Submitted:
15 April 2025
Posted:
16 April 2025
You are already at the latest version
Abstract
The increasing demand for sustainable, economical and environmentally friendly solutions has positioned microalgae as promising candidates in biotechnology, particularly in food, feed, nutraceutical, pharmaceutical, biofuel and bioremediation applications. This review explores the role of Nannochloropsis genus and other marine oleaginous microalgae in co-cultivation systems, highlighting their mechanisms of interaction with various microorganisms and their potential for various biotechnological purposes. Case studies of Nannochloropsis sp. co-cultures with other microalgae, bacteria and fungi are presented. The different types of associations are described as alternative strategies to enhance biomass productivity, lipid accumulation, and nutrient recycling. A key focus of this review is the potential of Nannochloropsis microalgae co-cultivation in food as it is part of the list of microalgae to be approved for consumption in the European Union, discussing their rich nutritional value, safety, and regulatory status. Additionally, the role of microalgae in the alternative protein sector is explored, with particular emphasis on their integration in cultivated meat products as nutrient suppliers and metabolic partners for animal cells. Despite their potential, several challenges such as scale-up, contamination risk and strain selection remain key obstacles to microalgal biotechnology widespread adoption. Future research should focus on optimizing microalgae-based co-cultures for food applications, addressing safety concerns, and further investigating their integration into functional foods and cellular agriculture products.
Keywords:
Nannochloropsis
; co-culture
; marine oleaginous microalgae
; biotechnology
; food
; bioactive compounds
1. Introduction
Microalgae constitute a wide group of photosynthetic organisms that have proven their value in biotechnology. Their simple cultivation requirements, rapid growth rates, high photon conversion efficiency and rich biochemical composition, have brought significant attention to their use in a myriad of applications. Among the vast diversity of microalgae, marine oleaginous species have gained increasing attention for their potential in biotechnology. These microorganisms are highly valued for their ability to thrive under varying salinity and temperature conditions while biosynthesizing a wide array of secondary metabolites with value in food, biofuel production and nutraceutical [1]. Their high productivity in vitamins, proteins, pigments, minerals and other bioactive compounds, combined with their rapid proliferation and simple growth requirements, make them promising candidates for food and nutraceutical applications [2].
In fact, microalgae are highly prized in food applications due to their rich protein content, which can vary from 6-70% of its dry weight (DW), with a well-balanced amino acid profile that includes the essential amino acids required for human nutrition [3]. Moreover, microalgae present high rentability due to their high protein yield per unit of biomass, making them an efficient and cost-effective alternative for large-scale protein production [3]. In addition to their high content, microalgae-derived proteins have also been associated with health benefits, including antioxidant, anti-inflammatory, and antiproliferative effects, further increasing their appeal in the development of functional foods [4]. That said, marine microalgae have been used as a valuable source of high-quality proteins offering a sustainable and nutrient-rich alternative to traditional protein sources.
Carotenoids of microalgae origin, such as β-carotene, astaxanthin, lycopene, fucoxanthin, among others, hold significant market value, due to their vibrant pigmentation and well-documented medicinal properties, making them highly relevant as natural colorants and functional ingredients in food formulations [2]. These compounds contribute to a healthy immune system and have been associated with the prevention of cardiovascular diseases, macular degeneration, and inflammatory conditions, as well as exhibiting antioxidant and antiproliferative activities [2].
In addition, marine microalgae represent a sustainable alternative to fish oils as a source of essential polyunsaturated fatty acids (PUFAs), particularly eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA) [1]. These omega-3 fatty acids (FAs) play a crucial role in cardiovascular health, inflammation regulation, and neurological function, with therapeutic applications in conditions such as Parkinson’s, Alzheimer’s, psoriasis, cancer, and rheumatoid arthritis [2]. While EPA serves as a precursor for bioactive lipids essential for cell membrane integrity, DHA is a major structural component of the brain, retina, and heart, underscoring its importance in human development and health [5]. Altogether, microalgae omega-3 and -6 FAs, are of extreme value in human and animal nutrition, being highly prized in food and feed application, especially given the incapacity of animals, including humans, of synthesizing these compounds naturally, emphasizing the need to obtain them from dietary sources [6].
As the demand for natural, sustainable and economically viable nutritional supplements continues to rise, optimizing microalgae bioprocesses for enhanced metabolite production remains a priority to ensure cost-effective and eco-friendly commercial applications. In this review, the role of microalgae-microorganisms co-culture is displayed in the context of maximizing production pathways that bring advantages to various biotechnological fields, with a focus on studies conducted on a particular marine microalgae species of great biotechnological interest. Table 1 summarizes the key bioactive compounds, lipid content, and applications of the species of marine oleaginous microalgae with increased biological interest.
From this list, Nannochloropsis spp. has stood out as a genus of particular interest due to its unique metabolic profile and broad biotechnological potential. Given the growing interest in co-cultivation as a strategy to develop more sustainable and efficient bioprocesses, along with the increasing focus on Nannochloropsis, it is important to consolidate existing research on the topic. Its rich nutritional composition, coupled with its adaptability as a versatile co-culture partner, has positioned Nannochloropsis as a promising candidate for novel bioprocessing strategies. A comprehensive understanding of past and ongoing studies will help clarify the principles and advantages of Nannochloropsis co-culture with other microorganisms, ultimately paving the way for unlocking its full biotechnological potential.
Nannochloropsis spp. is a marine oleaginous microalga that belongs to the Eustigmatophyceae class. The Nannochloropsis genus includes five well characterized species: N. oceanica, N. australis, N. granulata, N. oculata and N. limnetica (the only freshwater specie) [19]. Microchloropsis gaditana and Microchloropsis salina, initially classified as Nannochloropsis, were reclassified to the Microchloropsis genus due to significant genomic differences [20]. For simplification, this review will consider both Microchloropsis species as part of the Nannochloropsis group, as they share similar biochemical and nutritional properties.
Nannochloropsis species are single cells, oval-shaped, non-flagellated organisms with dimensions typically in the range of 2 to 8 µm. These microalgae are characterized by their thick, rigid cellulose-rich wall, composed mainly by glucose (~68%) and in minor concentrations (4 to 8%), by other monosaccharides such as rhamnose, mannose, ribose, xylose, fucose, and galactose [21]. Scholtz et al. (2014) [22] conducted a detailed study on the cell wall structure and composition of N. gaditana, revealing a two-layered cell wall. The inner layer is predominantly composed of cellulose (~75%) while the outer layer is rich in algaenan, a highly hydrophobic biopolymer of long-chain hydrocarbons [22]. The authors propose that this outer algaenan wall is a major contributor to the recalcitrance of N. gaditana cell walls, making them more resistant to enzymatic and chemical degradation [22]. Whilst this cell structure improves the organism’s ability to withstand harsh environmental conditions, it also presents a challenge in extracting intracellular compounds in industrial processes.
Unlike many other microalgae, Nannochloropsis cells are distinguished by their composition in chlorophyll a, but not chlorophyll b or c [7]. Apart from the freshwater microalgae N. limnetica, Nannochloropsis species thrive in marine environments, and are therefore cultured in artificial seawater-like environments. These species are grown under aerobic conditions, in the presence of a nitrogen source and light radiation to support their photosynthetic regime. Nonetheless, some can adopt heterotrophic or mixotrophic regimes when supplied with organic carbon sources. Commercial cultivation is often carried out in open raceway ponds or closed photobioreactors, under controlled optimal conditions to enhance productivity and metabolic performance [7].
The emerging focus on Nannochloropsis spp. is justified by their robust growth characteristics, ability to thrive in varying environmental conditions and feasibility to large-scale cultivation, make them ideal candidates for sustainable biomass production. Additionally, they have a notable biochemical composition, mainly regarding their rich lipid content, representing more than 20% of its DW, and up to 60% when cultured under stress conditions [23]. As oleaginous microalgae, Nannochloropsis sp. have a unique fatty acid profile, with the ability to synthetise a variety of saturated (SFAs), monounsaturated (MUFAs) and polyunsaturated fatty acids (PUFAs), the latter representing a significant share of the total fatty acids content [23]. In fact, it is the high PUFA content that makes these green microalgae particularly valuable, especially for omega-3 dietary supplements and biofuel production. Their ability to synthesize significant amounts of EPA, which can constitute 2% or more of their dry weight, further enhances their biotechnological and nutritional appeal for all the health benefits associated. Additionally, the protein and carbohydrate content represent around 25-30% and 30% DW, respectively, enriching the nutritional value of Nannochloropsis. Nevertheless, this data varies across strains, culture conditions, methodology and others. Moreover, the carotenoid profile is also noteworthy, namely for the production of violaxanthin and, in minor concentrations, astaxanthin, canthaxanthin, zeaxanthin and β-carotene, which also carry medicinal value, as previously stated [24]. As demonstrated, the nutritional value of Nannochloropsis is substantial and should be fully harnessed to enhance its applications in developing nutritious food, paving the way for more sustainable and health-promoting solutions. Recent advances on the biotechnology field have explored co-cultivation principles as strategies to enhance the sustainability, productivity and cost-efficiency of microalgal bioprocesses, representing an advantage over monocultures [25]. Co-cultivation refers to growing two or more different populations of cells or whole microorganisms in a shared and controlled environment. The goal is to mimic the natural habitat of cells and stimulate their natural biotic and abiotic interactions, to ultimately optimize their bioactivity [26]. This approach leverages several ecological principles that naturally occur, for instance, different species that require complementary resources, are able to co-exist together, with minimal competition and enhanced overall productivity [27]. The sustainable side of co-culture is directly related to the circular exchange of nutrients that occurs between autotrophic and heterotrophic species, mainly in recycling the nitrogen and carbon compounds, closing the cycles [28]. This way, the reciprocal exchange of nutrients between co-cultured species allows for a more efficient utilization of available resources, improving growth rates and metabolic output. Overall, there are several studies that show that co-cultivation experiments improve the production rates, growth profiles and may trigger new production pathways, which increases chemical diversity [29].
This review focus on the integration of marine oleaginous microalgae in co-culture systems with other microorganisms which holds of great interest in biotechnology with promising results in nutritional supplements, aquaculture, biodiesel production and wastewater treatment (WWT) [25]. Their ability to engage in productive interactions with other microorganisms opens new possibilities for enhancing biomass productivity, improving nutrient recycling, and creating more sustainable bioprocesses. Given these advantages, reviewing the potential of Nannochloropsis in co-cultivation systems is crucial for advancing its applications in biotechnology and ensure its full potential.
This review aims to explore the existing studies on the co-culture of marine oleaginous microalgae with other microorganisms, with particular focus on the Nannochloropsis species. It will uncover the mechanisms behind these interactions, identify the most common biotechnological applications and discuss the potential applications of Nannochloropsis co-cultures in the food industry. Finally, it will present the challenges and opportunities for future developments in marine oleaginous microalgae processes.
2. Materials and Methods
The Google Scholar and Clarivate Web of Science platforms were used to search for articles using the words “co-culture”, “marine microalgae”, “Nannochloropsis” or “oleaginous” as mandatory. Other words, such as “biotechnology”, “biofuel”, “wastewater”, “food”, “feed”, “nutraceuticals” or “cellular agriculture” were defined as optional. The search was limited to the years between 2001 and 2024. From the retrieved articles, a careful selection process was applied to prioritize original research over review papers whenever possible.
3. Mechanisms of Interaction in Microbial Consortia
Over time, organisms have developed diverse strategies to interact with each other to better adapt to their environment, tolerate adverse conditions and even resist to invasive species. These interactions can take different forms, depending on the benefits or disadvantages conferred to the organisms in association. That said, symbiosis, the interaction between two organisms of different species, can be of different types: mutualism, when both parties benefit from the interaction; commensalism, when only one specie is benefited while the other is unaffected; and parasitism, when one species gains advantage at the expense of the other [30].
These associations are frequent in microbial communities, including microalgae-microorganisms consortia. Compared to axenic cultures, co-cultivation is a strategy oftenly applied as it results in enhanced productivity, microbial resilience and metabolic efficiency, making polycultures extremely valuable in the bioprocessing field [25]. However, one of the main challenges in artificial polycultures is selecting the appropriate species, before promoting naturally occurring interactions for industrial purposes. A thorough study on the impact of both biotic and abiotic factors on a co-culture system is essential to ensure that the selected strains are benefit for industrial applications.
Robust microbial strains with high growth rates, strong tolerance to environmental fluctuations and valuable metabolic activities are essential for cost-efficient and scalable bioprocesses. When designing a co-culture system for biotechnological large-scale applications, strain selection must consider not only the inherent biological value of the organisms but also their compatibility with industrial settings. For instance, given Nannochloropsis is usually cultured under high salinity media, it is imperial to choose co-culture partners that exhibit halotolerance or halophilic traits. In fact, these characteristics are advantageous, not only because high salinity can suppress pathogenic species, mitigating contamination issues, but also because halotolerant microbes often offer metabolic versatility [31]. An example with industrial interest is bacteria Paracoccus spp., known for their flexible metabolism, ability to survive on diverse substrates, effective denitrification capacity, and overall resilience to environmental changes [32]. Such traits could help optimize nutrient cycling and enhance lipid production in a Nannochloropsis culture, with prospect for scaling up. Therefore, a detailed evaluation of the environmental adaptability of microorganisms is important in strain selection for co-cultivation purposes. Ultimately, the strategic selection of robust, high-performing strains lays the grounds for scalable, cost-effective, and sustainable microalgal bioprocesses.
Another relevant aspect to account for upon strain selection is the microbial mechanisms of interspecies interaction. Most of microalgal interactions with other species occur in aquatic habitats, through the exchange of metabolites or signalling molecules in the phycosphere – the rich microenvironment immediately surrounding a microalgal cell, crowded with various microorganisms [33]. The network of metabolites that is established in a microbial consortium can be categorized into two different mechanisms of communication based on the nature of the compounds involved, whether they are produced within cells (endometabolites) or secreted into the surrounding environment (exometabolites) [34].
3.1. Metabolite Exchange
One of the most fundamental interactions in microalgae-microorganism consortia is built on the exchange of nutrients. As photoautotrophic organisms, microalgae play a crucial role in the carbon cycle by fixing atmospheric carbon dioxide (CO2) and producing organic carbon substrates, essential micronutrients and oxygen (O2). These metabolites serve as energy sources for the heterotrophic microorganisms in association, which in return release CO2 and inorganic carbon byproducts back into the environment to be used by microalgae, establishing a mutualistic carbon-oxygen cycle [34].
A similar exchange occurs for nitrogen compounds. Some nitrogen-fixing bacteria (e.g., Azospirillum, Rhizobium) can convert the atmospheric nitrogen (N2) into bioavailable inorganic nitrogen, in the form of ammonium (NH4+) or nitrate (NO3-), ready to be utilized by microalgae [35]. The exchange of nutrients is also extended to sulphur compounds, derived from microalgal metabolism and posteriorly utilized by sulphur-metabolizing bacteria and fungi, establishing another cycle within microbial communities [35].
3.2. Chemical Interactions
Beyond nutrient exchange, microalgae can interact with the surrounding microorganisms through chemical signalling, which can trigger physiological responses and influence population dynamics by controlling gene expression. In microalgae, these interactions are mediated by allelochemical metabolites, a diverse group of signalling molecules that include pheromones, free fatty acids, alkaloids, aldehydes, peptides or extracellular metabolites, among others [36]. Such exchange of metabolites can have either a positive or negative effect on neighbouring communities: in some cases, microalgal allelochemicals inhibit the growth of competing species, disrupt biofilm formation or induce cell lysis, working as antimicrobial agents [36]. In others, byproducts of algal metabolism can induce an adaptative response, helping them and the surrounding organisms to better adapt to extreme environments or resist to stress conditions such as nutrient deprivation [37].
One of the most well-documented bacterial-chemical mechanisms of interaction, the quorum sensing (QS), has shown to influence microalgal behaviour, affecting nutrient acquisition, biofilm formation and self-motility [38]. In response, these photoautotrophic organisms have developed defence mechanisms, being able to produce QS-inhibitors that interfere with bacterial communications systems [38]. The production of indole-3-acetic acid (IAA) is a well-studied example of a chemical signalling mechanisms between microalgae and bacteria. When secreted into the extracellular space by Sulfitobacter species, the hormone IAA promotes microalgae growth and lipid production, which carries biotechnological value. However, at high concentrations, it becomes toxic for microalgae, favouring the bacteria, that take advantage of the nutrients from the lysed cells [39,40]. This example underlies the importance of carefully studying microbial interactions since a benefit relation can rapidly shift into a competitive or even parasitic association.
Despite these interrelations depending highly on the metabolome of species, abiotic factors also have a part in allelopathy behaviours. Nutrient limitation, low temperatures and high pH are known to stimulate the synthesis of allelochemicals, whereas high light irradiation, high temperatures, high nutrient bioavailability and low pH repress the secretion of signalling molecules [36].
3.3. Metabolomics of Nannochloropsis
In Nannochloropsis, the biosynthesis of triacylglycerol’s (TAGs) and EPA involves a series of enzymatic reactions that convert simple FAs precursors into complex lipid molecules [41]. Summarily, TAG synthesis primarily occurs through the acyl-CoA-dependent Kennedy pathway, which comprises sequential acylation steps of glycerol-3-phosphate into lysophosphatidic acid, phosphatidic acid, diacylglycerol and finally into TAG by action of various enzymes [42]. Under nitrogen-limited conditions, there is higher accumulation of TAGs, particularly C16 fatty acids, due to the overexpression of diacylglycerols acyltransferase genes [42]. Additionally, an acyl-CoA-independent pathway can also contribute to the TAG production, especially in CO2-rich environments, by utilizing photosynthetically assimilated carbon [43]. Regarding the EPA production, Nannochloropsis follows one metabolic route, ω-6 pathway, which varies from other EPA-producing microalgae, such as P. tricornutum that may employ two different pathways for EPA production [44]. In Nannochloropsis EPA biosynthesis occurs in the endoplasmic reticulum, where stearic acid (C18:0) is converted into EPA (C20:5) in various intermediate steps, through the action of fatty acid desaturates and a fatty acid elongase [43]. The newly synthesized EPA molecule is then transferred to the chloroplast membrane, a translocation process that constitutes a difficulty in cellular engineering approaches for increasing the EPA production [43]. This unique, single-route pathway for EPA production not only highlights the species’ metabolic specialization but also points to both opportunities and challenges for future metabolic engineering aimed at enhancing EPA yields in industrial applications [41].
A thorough understanding of the lipid metabolic pathways in Nannochloropsis is crucial for selecting appropriate co-cultivation strains and optimizing culture conditions to maximize bioactive lipid production. As previously discussed, multiple factors influence the lipid metabolism of microalgae, including the presence of other microorganisms in culture. For instance, TAG accumulation can be stimulated in the presence of denitrifying bacteria that would contribute to the depletion of nitrogen in the medium [45]. Conversely, the co-cultivation with photosynthetic microorganisms can promote competition for atmospheric carbon, potentially reducing Nannochloropsis‘ metabolic activity and therefore lowering lipid productivity [45]. On the contrary, anaerobic bacteria capable of recycling organic carbon produced by microalgae, re-introduce it into the system, increasing the availability of photosynthetic assimilated carbon and promoting lipid synthesis. Additionally, microalgae from the Nannochloropsis genera use B-vitamins as growth factors and health promoters, but lack the ability to synthesize them independently, needing an exogenous source [46]. Certain microorganisms can serve as a source of B-vitamins precursors, enhancing biomass growth and indirectly stimulating lipid accumulation by boosting the photosynthetic activity [46]. Hence, the presence of other microorganisms in co-culture systems can be a valuable asset for enhanced biomass and lipid production of Nannochloropsis, as various microbes can secrete signaling molecules, enzymes, or metabolites that directly impact the metabolic pathways.
Abiotic factors also play a fundamental role in microalgae metabolism and must be considered when designing co-culture systems. Nannochloropsis thrives at an optimal temperature of 25-29ºC, a pH of 7-10, salinity levels between 22-49 g/L, and in the presence of light as an obligate phototrophic species. Despite being a specie that can easily adapt to environmental fluctuations, stress conditions can have implications in the metabolic vias of Nannochloropsis. At lower temperatures (down to 9ºC) the production of unsaturated lipids is enhanced as a mechanism to maintain membrane fluidity, which results in higher EPA and PUFAs content, while higher temperatures (up to 31ºC) promote the synthesis of TAGs [47]. While Nannochloropsis sp. is considered to be a halophilic specie, tolerating a wide salinity range, salt concentration influences its growth and lipid profile [48]. Studies indicate that at lower salinity levels (25‰) EPA accumulation increases while total lipid content decreases [48]. The content in C16 is not significantly altered although it reaches higher levels in low salinity environments [48]. Light intensity and irradiation wavelength have also been reported to influence biomass growth and metabolites production in Nannochloropsis, with total lipid content increasing with light exposure along with the levels of TAGs. However, this condition resulted in reduced EPA synthesis due to alterations in chloroplast membrane, where EPA in allocated [49]. Moreover, the fatty acid profile is also responsive to variations of light wavelength, with blue and red light favoring the production of SFAs over white light [50].
All things considered, taking the influence of abiotic factors in Nannochloropsis cultures is fundamental when designing polyculture systems. The previously presented studies show that fluctuations from standard cultivation conditions have an impact in Nannochloropsis growth and bioactivity. Hence, it is important to, not only guarantee that the cultivation conditions are favorable for all involved species, ensuring optimum growth, but also account for the influence that culture partners may have on the environment settings. For instance, the high cell density of co-cultured partners can compromise light penetration, altering the culture conditions and influencing Nannochloropsis metabolic activity.
In fact, previous studies have demonstrated that co-cultivation strategies can impact the metabolism of microalgae through their mechanisms of interaction. Xu et al. (2018) [51] studied the co-culture of Chlamydomonas reinhardtii with Azotobacter chroococcumg, reporting that the co-cultivation strategy promoted the growth and lipid production of microalgae under nitrogen deprivation, by upregulating microalgae genes responsible for lipid metabolism [51]. In a different study, Leyva et al. (2015) [52] concluded that the presence of Azospirillum brasilense in Chlorella vulgaris cultures, promoted the activity of acetyl-CoA carboxylase and thereby enhanced lipid accumulation. Given that this enzyme is also part of Nannochloropsis lipid biosynthesis [7], co-culture strategies leveraging microbial interactions to enhance enzymatic activity could be explored while maintaining optimal cultivation conditions.
Further studies should focus on deepen the knowledge on the specific metabolic changes in Nannochloropsis when co-cultured with different microorganisms. Understanding the underlying regulatory mechanisms of lipid and biomass production will enable the selection of microbial partners that optimize microalgal productivity. Additionally, assessing how both biotic and abiotic factors impact co-culture systems will be essential for developing sustainable and efficient lipid production strategies.
The communication between microalgae and microorganisms is a highly complex and dynamic network of signalling molecules and metabolic exchanges, which can be either advantageous or harmful for the species involved. Hence, it is essential to conduct a strict study before inoculating two different species to assure that the desired goals are achieved. Understanding the microbial ways of communication, the mechanisms of microalgae when in the presence of other organisms and the competitiveness for resources, is vital when designing a synthetic co–culture of microorganisms.
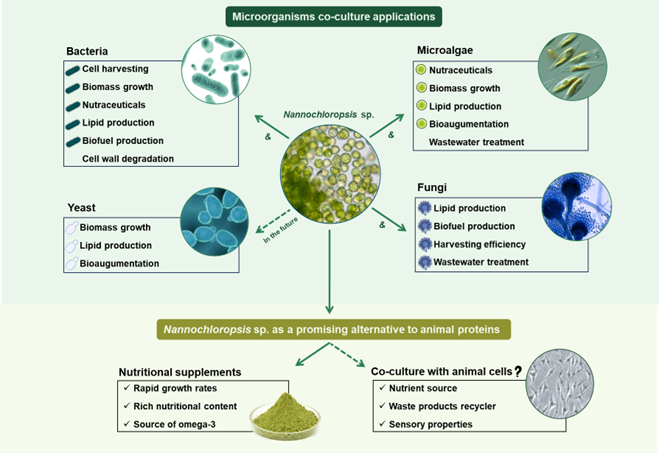
4. Co-Culture of Nannochloropsis with Microorganisms
The application of co-cultivation principles in bioprocessing has been greatly explored as the benefits of mixing cultures are being unraveled. The improvement of biomass and added-value compounds productivity, enhanced growth, efficient waste by-products removal or facility in biomass harvest, are the main advantageous traits of microalgae-microorganisms co-cultivation systems and will be detailed further in this article.
Table 2 summarizes the cases of co–culture of Nannochloropsis spp. with other various microorganism species found in literature.
4.1. Microalgae and Microalgae
The main characteristic of a co-culture system involving two microalgae species is the existing competition for the same limited resources. This can trigger mechanisms of adaptation that can ultimately lead to increased biomass growth, efficient nutrient uptake and improved lipid yields. Moreover, the complementary metabolic pathways of different species may enhance the overall production of valuable compounds, potentially leading to the discovery of new bioactive molecules with novel health-promoting properties [6]. Marine microalgae species have gain attention due to their ability to grow in saline water and with recycled nutrients, reducing the costs on freshwater. This feature makes them a cost-efficient and environmentally friendly alternative in biofuel and added-value compounds production [63].
Most cases of co-cultivation of Nannochloropsis sp. with other marine microalgae leverage on their capacity to biosynthesize compounds of interest. Maglie et al. (2021) [64] presented a comprehensive study on the co-cultivation of N. oculata with the marine microalga Tisochrysis lutea, demonstrating that while this co-cultivation strategy enhanced the production of valuable metabolites and optimized nutrient utilization, it also presented downsides. In fact, both species exhibited higher biomass growth in monocultures which the authors attributed to the initial cell density used, suggesting that the inoculum ratio is also an important factor to account for when designing artificial co-cultivation systems [64]. On the other hand, co-cultivation resulted in increased chlorophyll and carotenoid content, highlighting its potential in increasing second metabolites productivity [64]. Regarding the lipid profile there was a slight decrease in total fatty acid content, related once more, to the proportion of algae in the polyculture. Nonetheless, the combined system resulted in a richer omega-3 PUFAs content and a diversified profile, with N. oculata contributing to 17.4 mg/g DW composition in EPA, and T. lutea providing 13.9 mg/g DW of DHA [64]. Building on this, Thurn et al. (2022) [65], optimized the combined production of EPA and DHA by co-culturing two marine oleaginous species, T. lutea with M. salina, achieving a balanced 1:1 ratio of these essential FAs after 8 days, with a reported increase in DHA concentration of around 73% but no significant changes on EPA production improvement. The co-cultivation system enabled the simultaneous production of these omega-3 FAs though it did not enhance overall accumulation of EPA. Most of EPA-producer organisms are unable to synthesize DHA and vice-versa, making this strategy particularly attractive in obtaining a more diverse microalgal-lipid content, despite the need for further optimization [65]. The authors also registered a biomass increase of 31% due to optimal utilization of the light spectrum [65]. More recently, these authors expanded the approach to the green microalgae N. oceanica and M. salina (EPA producers), and the brown-microalgae I. galbana (DHA producer), achieving enhanced biomass production and richer omega-3 FA profiles likely due to the species capacity to absorb light at different wavelengths in the mixed culture [66]. While these studies highlight the potential of co-cultivation in producing two highly valuable bioactive compounds in a single system, they also underline important trade-offs that must be accounted for and further investigated to optimize mixed cultures bioprocesses.
Several studies have reinforced the potential of polycultures of marine oleaginous microalgae in biofuel production, where a high PUFAs content improves fuel properties and promotes carbon neutral processes with no additional carbon being released into the atmosphere [67]. Additionally, the enrichment in omega-3 FAs in combined microalgae cultures supports their application in the nutraceutical industry, particularly in the production of alternative fish-oil supplements and in the nutritional enrichment of aquaculture feed [6].
Beyond these applications, Nannochloropsis sp. cultivation with other microalgae species has also demonstrated increasing value in bioremediation. Davis et al. (2015) [54] explored the potential of using struvite, a phosphate mineral commonly recovered from wastewater, as a nutrient source for a combined marine culture of M. salina and P. tricornutum. The research findings showed that the nitrogen and phosphate content of struvite-based media was sufficient to support the cell culture in the same way as standard medium, with the advantage of increasing nutrient uptake and pigment production [54]. On a different approach, Sharma et al. (2020) [53] evaluated the bioremediation efficiency was achieved when culturing a microalgal consortia, of both fresh (Chlorella sp., Chlamydomonas reinhardtii, Scenedesmus bijugatus, and Scenedesmus dimorphus) and seawater (Nannochloropsis sp.) species in sewage wastewater. One of the artificial microbial consortia resulted in the removal of ~86% of total organic carbon, 87% of chemical oxygen and reduce the nitrate and phosphate compounds in ~94%, with efficient heavy metal removal [53]. This pollutant removal capacity of microalgae co-cultures can also be applied in aquaculture to treat marine wastewater, as it was studied by Davis et al. (2015) [68], when designing a co-cultivation system of marine P. tricornutum with freshwater Chlorella sp.. The synergistic interactions between both species resulted in a removal rate of total nitrogen of ~93% and of total phosphorous of ~96% [68]. P. tricornutum metabolites enhance the photosynthetic activity of Chlorella, which thereby increases the bioavailability of carbon sources, favourable for the heterotrophic regime of the seawater specie [68]. In the same work, the authors proved the efficacy of the proposed co-culture system outdoors in actual marine wastewater, bridging the gap for application in aquaculture ponds [68]. These results demonstrate the industrial applicability of microalgae-based co-cultivation systems for WWT, as they report high nutrient removal efficiency, particularly in reducing organic carbon, nitrogen, phosphorus, and heavy metals, aligning with the requirements for large-scale bioremediation practices.
Overall, these case studies highlight the potential of marine oleaginous microalgae co-culture in bioremediation, while holding increasing interest for biofuel production and dietary supplements. By improving nutrient recycling, enhancing biomass and secondary metabolites productivity, microalgae-microalgae systems offer a potentially more sustainable and cost-efficient approach.
4.2. Microalgae and Bacteria
The duality microalga-bacteria is a well-documented and naturally occurring interaction in aquatic environments. Bacteria thrive in the nutrient-rich phycosphere surrounding microalgal cells. The complex microbial consortia engage in all types of symbiotic interactions, from mutualistic nutrient cycling to QS regulation [6]. The mutualistic relations will be the focus of this chapter.
Mutualism between bacteria and microalgae is primarily based on nutrient exchange since microalgae work as a supplier of dissolved organic carbon necessary for bacterial growth. Simultaneously, bacteria decompose inorganic matter, providing a myriad of nutrients, vitamin precursors (vitamin B12), growth promoting hormones (IAA), CO2 and available nitrogen to microalgae [69].
The specific bacterial communities associated with microalgae can vary significantly depending on environmental factors, which influences their growth and biochemical composition in different ways. When characterizing the phycosphere of a Nannochloropsis sp. strain cultivated in outdoor photobioreactors, Lian et al. (2021) [33] identified eighteen bacterial strains. Among them, Maritalea porphyrae and Labrenzia aggegata were found to significantly enhance microalgae growth and chlorophyll production, whereas other bacteria, such as Flavonacteria, caused growth inhibition [33]. These results highlight the importance of thoroughly characterizing naturally occurring microalgae-bacterial interactions before designing a co-culture system, in order to select the bacterial strains that would contribute favourably to the process.
There are several reports on the co-culture of marine microalgae with bacteria for a variety of biotechnological purposes, including WWT, biofuel production, added-value metabolite synthesis, and contamination control in microalgae cultures and aquaculture ponds, and waste metabolites removal. Subasankari et al. (2020) [70] investigated the symbiotic relationship between N. oceanica and the halophilic bacterium Halomonas aquamarine. Their study revealed that bacterial co-cultivation enhanced microalgae biomass production, lipid accumulation and carotenoid synthesis. This improvement was attributed to the production of siderophores and vitamins by H. aquamarine, which function as algal growth factors, leading to a 14% increase in lipid content [70]. Similar beneficial effects have been observed in co-cultures involving other marine oleaginous microalgae, such as I. galbana and P. tricornutum with bacterial strains from the Marinobacter genus. These mixed cultures not only enhanced biomass productivity, but also increased lipid accumulation, with I. galbana yielding higher concentrations of DHA [71,72]. These findings underscore the potential of bacterial co-cultivation strategies to enhance the production of bioactive compounds with potentially significant health benefits.
One of the bottlenecks in large-scale algal biotechnology is biomass harvesting which often requires high energy-consuming techniques [73]. This cost-driver step is particularly relevant in biofuel production for the dewatering of cells [74]. To address this, researchers have explored bacterial co-cultivation as a strategy to enhance microalgal aggregation and reduce energy requirements during harvesting processes. Powell and Hill (2013) [59] exploited the role of Bacillus sp. bacteria in aggregation of N. oceanica cultures. Results showed that cell accumulation rapidly occurred in response to the bacterial secretion of extracellular polymeric substances (EPS), which promote cell adhesion and clustering [59]. Similarly, Tran et al. (2020) [75], identified bacterial isolates from the Rhodobacterales, Flavobacteriales and Sphingomonadales orders, as key players in promoting EPS production and consequent aggregation of N. oceanica, leading to improved biomass recovery. These findings support bacterial co-cultivation strategies as a cost-effective and sustainable approach for mitigating harvesting challenges.
In addition to aggregation, other major limitation in microalgal bioprocessing lies on the difficulty in degrading the stable and resistant cellulose-rich cell wall to gain access to the intracellular metabolites. As previously stated, Nannochloropsis species, in particularly, are known for their thick and rigid walls, which constitutes an obstacle for intracellular lipid extraction [19]. Generally, mechanical or chemical pre-treatment steps are employed to breach down or weaken cell walls, although, these methods have the downside of being costly and aggressive, with the possibility to cause degradation of the interest products. Co-culture can be applied has a more sustainable alternative as specific microbial strains might help overcome cell wall resistance, improving the bioavailability of intracellular compounds for food and biotechnological applications. Understanding the cell wall composition is crucial to select a co-cultivation partner that could hydrolyse the cell wall components without compromising the intracellular products. Muñoz et al. (2014) [60] studied the enzymatic capacity of bacteria to hydrolyse microalgal cell wall by co-culturing microalgae with several species of bacteria. Results showed that the most efficient strain in N. gaditana cell wall degradation was Raoultella ornithinolytica, whereas it proved to be ineffective towards Botryococcus braunii [60]. This disparity of results was proposed to be caused by the influence of external factors on cellulolytic bacterial metabolic performances, highlighting the need for a deeper understanding of interspecies relations [60]. This system promoted bacterial enzymatic cell wall degradation, enhancing lipid recovery without the need for expensive and energy-consuming pre-treatment processes [60]. The increase of the bioavailability of lipids represents, once more, an advantageous trait in the biofuel production although it could not be applied in the food sector as R. ornithinolytica is known to cause infections in humans [76]. While this case study presents the potential of microalgal-bacterial interactions as an alternative to costly pre-treatment processes, further research should prioritize identifying non-pathogenic species.
In conclusion, bacterial co-cultivation offers the opportunity to enhance Nannochloropsis sp. productivity, lipid accumulation and facilitate biomass harvesting and lipid extraction, being a promising and economical efficient approach for the scale-up of algal biotechnology.
4.3. Microalgae and Fungi
Microalgae-fungi associations are a naturally occurring symbiotic association that can improve biomass growth, lipid accumulation and WWT efficiency. The interest in the co–culture of those microorganisms focuses on the self–pelletization capacity of filamentous fungi, which enables them to aggregate with each other into fungal pellets [73]. When in association, the fungi will work as bio–flocculant agents entrapping the microalgae in their pellets, thereby facilitating the separation of biomass from liquid fraction [73]. In accordance with the previously identified bottlenecks of microalgal bioprocesses, this bio-flocculation property is of great significance in industrial applications, as it simplifies and cuts costs on cell harvesting. Beyond cell aggregation, microalgae and fungi co-cultures can enhance lipid production of TAGs and PUFAs, both of which are valuable for biofuel production and nutraceutical applications. Furthermore, among all microalgae-microbial associations, algae–fungi systems have demonstrated the highest efficiency in WWT due to the combined action of microalgal nutrient assimilation with fungal organic matter degradation [25].
Du et al. (2018) [77] explored the co-culture of N. oceanica with the oleaginous fungus Mortierella elongata, aiming for bio-flocculation and lipid yield improvement. The study implemented a cultivation strategy in which the microorganisms were grown separately, allowing M. elongata to develop its mycelial network before being added to the mixed culture. The results exhibited a significant enhancement in biomass aggregation, with the fungus specie efficiently capturing microalgal cells in its network and thereby facilitating cell separation and lipid extraction. Moreover, the co-culture led to an increase in lipid accumulation in both oleaginous species, with a significant rise in TAG and PUFAs content. Notably, M. elongata also enhanced the production of DHA, due to the high salinity of the medium. The study also applied genetic engineering techniques to further optimize the FA profile of N. oceanica [77]. A similar approach was explored by Wrede et al. (2014) [73] by co-cultivating the filamentous fungus Aspergillus fumigatus with various marine oleaginous microalgae species, (D. salina, N. oculata and T. chuii). The results confirmed that fungal-microalgal interaction improved bio-flocculation up to 90% after 24h of co-cultivation for almost all tested microalgal species. Fungal mycelia trapped microalgal cells within their filaments, simplifying biomass harvesting. In addition, the co-culture increased lipid yields and diverse content in TAGs and PUFAs [73]. Beyond biofuel production, this study has proved the efficacy of combined cultures of A. fumigatus with microalgae in WWT. The fungi successfully degraded organic matter while microalgae assimilated the resultant nitrogen and phosphorous, creating an efficient circular system with potential for large scale wastewater bioremediation [73].
The integration of microalgae and fungi represents a promising, scalable and sustainable strategy for addressing several challenges in biofuel production, microalgal harvesting and environmental remediation. This co-culture pairing holds the potential for several industrial applications, if the right strains and conditions are applied.
Microalgae and Yeast
Similar to other microbial interactions, the co-cultivation of microalgae and yeast has been highlighted in previous studies, demonstrating its biotechnological potential, particularly in enhancing lipid production[78]. This approach has been explored for applications in biofuel production, aquaculture feed, WWT and added-value compounds synthesis [79]. Microalgae and yeast form a complementary metabolic relationship with photosynthetic microalgae producing O2 and organic carbon sources that support yeast growth, while yeast produces CO2 and nutrients that stimulate microalgal metabolism [80]. This exchanges of gas and metabolites not only optimize biomass growth but also improve lipid accumulation, as yeasts can increase the bioavailability of key metabolites required for microalgae lipid biosynthesis [6]. Additionally, yeast simple cultivation requirements and ability to thrive on diverse feedstocks, allow for the reuse of industrial or agriculture waste streams, making these co-culture systems useful in applications associated to WWT [81,82]. Another characteristic of yeasts is their capacity to degrade heavy metals by bioaugmentation, making them valuable in environmental treatment of contaminated sites [83].
While co-cultivation strategies have been explored in freshwater microalgae-yeast systems, research on marine oleaginous microalgae, particularly for Nannochloropsis sp., remains limited. However, Cai et al. (2007) [84], analysed the growth and biochemical composition of a mixed culture of the marine microalgae I. galbana and the yeast Ambrosiozyma cicatricose, showing that when cultured together, both organisms reached higher growth rates and increased FAs production. Although, overall lipid concentrations did not significantly surpass those of monocultures, demonstrating that despite the potential of these systems, there is space to optimize their interactions [84].
Given the high lipid productivity of certain marine microalgae and the metabolic versatility and efficiency of yeast, further research into optimizing strain selection and culture conditions, could unlock new opportunities for scalable and cost-effective alternatives for biotechnological applications [79]. The integration of yeast in marine microalgal cultures presents an avenue of research with economical and sustainable potential and an opportunity for further scientific investigation.
4.4. Co-Culture of Other Microalgae with Microorganisms
The literature on microalgae-microorganisms consortia is extensive, particularly in biotechnological purposes. Several reviews have been published on microalgal co-cultivation strategies for biofuel production [85], bioremediation and WWT [86,87] and the synthesis of bioactive compounds [25,88]. Among the microalgae studied in polycultures, freshwater species Chlorella spp. and Scenedesmus obliquus stand out as some of the most common [89]. Chlorella spp. is particularly valued for its secondary metabolite production robust growth with diverse substrates, and compatibility with various microorganisms, while S. obliquus is recognized for its high lipid content and efficient nutrient uptake [89]. However, the co-cultivation of these and other microalgae species falls outside the scope of this review, which is primarily centered on Nannochloropsis spp.. Given its classification as a marine oleaginous microalga, it is relevant to compare its co-cultivation studies with those involving other microalgae from the same ecological and metabolic niche (presented in Table 1). Table 3 provides a summary of co-culture studies conducted with marine oleaginous species, beyond Nannochloropsis, offering a broader understanding on co-cultivation strategies in this specific group.
Despite the category of marine oleaginous species involving more species (Table 1), the ones presented in Table 3 were the ones with published articles on co-culture, found. In broad lines, Phaeodactylum tricornutum is a marine diatom widely recognized for its ability to synthesize significant amounts of EPA and fucoxanthin [9,10]. In co-culture systems, P. tricornutum has been paired with other microalgae or bacterial partners (e.g., Marinobacter sp.) to enhance lipid and biomass productivity and optimization of wastewater treatment [91,93]. In comparison with Nannochloropsis, P. tricornutum can be more sensitive to fluctuations in temperature and light variations, besides requiring complex and expensive harvesting steps in the downstream processing, which constitutes a challenge at industrial scale [105].
Isochrysis galbana, valuable for its high PUFAs content, containing both EPA and DHA, is usually applied in aquaculture feed and nutraceutical formulations [11,12]. Co-culture systems involving I. galbana and bacterial strains (Alteromonas sp., Labrenzia sp.) have shown improvements in biomass yield, lipid profile, and even pathogen inhibition [99]. However, I. galbana generally produces lower overall lipid yields compared to Nannochloropsis (Table 1).
Species from the Tetraselmis genera are known for their ability to adapt to various salinities levels besides their rentable lipid accumulation [13,14]. Co-culture studies often focus on the microalgae capacity to induce flocculation promoting the harvesting step in other microalgae cultures [100]. While Tetraselmis sp. demonstrates strong resilience and decent growth rates, its lipid profile tends to be less dominated by polyunsaturated fatty acids (PUFAs) than that of Nannochloropsis. Consequently, Nannochloropsis remains a more attractive option for industries seeking high-value PUFAs.
Finally, Dunaliella salina is best known for its high β-carotene content, making it a valuable asset in the nutraceutical and cosmetic sectors [15]. The few studies found on co-culture approaches, paired D. salina with bacterial strains (Halomonas sp.) aiming for enhanced biomass production and carotenoid yield while simultaneously addressing bioremediation challenges [104]. Although D. salina can accumulate lipids under stress, it is primarily cultivated for carotenoids rather than PUFAs, unlike Nannochloropsis.
While each marine oleaginous microalga has its unique advantages and added-value metabolites, Nannochloropsis excels in producing EPA-rich lipids, often exhibiting robust growth, high biomass and PUFAs yields and significant tolerance to salinity and temperature changes. This combination of resilience and high-value lipid production makes Nannochloropsis especially appealing for industrial applications in food, feed, biofuel and bioremediation sectors, with more published research on co-culture approaches than the other marine oleaginous microalgae. Consequently, Nannochloropsis remains a prime candidate for large-scale co-cultivation systems designed to optimize PUFA production while maintaining cost-effectiveness. Nonetheless, further exploration of the other valuable marine species presents an opportunity to uncover novel traits and applications that could enhance existing bioprocesses.
5.1. Microalgae in Food and Feed
In this review, as in previous reviews, microalgae consortia are presented has a valuable strategy to optimize biofuel production, WWT, bioremediation and added-value metabolites production. The ability to engineer symbiotic relationships between microalgae and microorganisms has led to scalable and cost-effective solutions for some of the major bottlenecks in microalgal biotechnology, including enhanced biomass harvesting, improved lipid productivity and efficient nutrient recycling [6]. However, the full potential of using these microorganisms in the food sector is still a topic to be explored. On the other hand, from a nutritional point of view, it has been stated that microalgae are a promising asset for food and feed purposes due to their high content in proteins, carbohydrates, lipids and carotenoids, holding great especially in representing a natural, sustainable and healthy alternative to other protein sources and in dietary supplements formulations.
In livestock, incorporating microalgal biomass in feed formulations has been shown to enhance animal growth, improve their immune response and increase their resistance towards bacterial and viral infections [106]. In aquaculture, microalgae serve as sustainable alternatives to traditional fishmeal (i.e., fish farming feed composed by smaller fishes or fish waste), contributing to release the pressure on wild fish stocks. Whether as an exclusive feed compound or as part of the regular feed, microalgae have been shown to improve the nutrient content, colour and overall health of aquaculture molluscs, crustaceans, shrimps and fish [107]. Moreover, terrestrial animal feeds supplemented with microalgae have demonstrated improvements in meat quality caused by the antioxidant properties and high content in PUFAs and pigments of microalgae [107].
In human consumption, the introduction of microalgae in human dietary is becoming more popular by the day, whether in the form of tablets, dried powder and omega-3 rich oils capsules, protein, carotenoids or other food supplements formulations. Moreover, as they can be easily incorporated in a variety of food products, their versatility has facilitated their consumption by their use as ingredients in common foods such as bread, snacks, dairy products and beverages. However, the value of microalgae in cooking, goes beyond their nutritious content, since they are also beneficial for their organoleptic, rheological and shelf-life properties [106]. As a viable alternative to conventional fish-oil supplements and animal protein, microalgae have the potential to help reduce the pressure on traditional food systems while still ensuring global food security.
The co-cultivation of microalgae with other microorganisms also presents interesting opportunities in the food and feed industry, particularly in enhancing nutritional quality while aiming for more sustainable processes. Combined cultures of microalgae are used in aquaculture systems to serve as rich nutritional feed, probiotics or to mitigate potential diseases and ensure overall seafood health, contributing to more sustainable aquaculture practices [108]. In the same way, these principles could be applied to human nutrition. The literature on microalgal polycultures designed for human consumption is still scarce. Most commercialized products combine multiple microalgal biomasses or their bioactive products in one nutrient-rich supplement [109]. Tyus (2016) [109] explored this idea by analysing the protein, carbohydrate and lipid production of microalgae co-cultures, so that it could be applied in nutraceuticals. The author concluded that the biochemical value of the co-culture system resulted in higher lipid, carbohydrates and protein profiles and monoculture biomass, highlighting the potential behind exploring microalgal mixed cultured in human supplements formulation. More studies on the matter should be performed to investigate the potential of microalgae-microalgae and microalgae-microbial systems in the development of functional foods, as it has been displayed throughout this work that it can be highly advantageous in the nutraceutical industry.
5.2. The Potential of Nannochloropsis in the Alternative Proteins’ World
The growing demand for sustainable, safe and animal-free protein sources has accelerated research in finding alternative sources of proteins, with microalgae emerging as promising alternatives to animal protein. Their rapid growth and high protein content, composed by all essential and non-essential amino acids have attracted significant attention for their use in plant-based food [110].
Nannochloropsis sp., in particular, has a high protein content of around 30% of its DW, with a balanced ratio between essential and non-essential amino acids, making it a viable protein source in food formulations and a sustainable alternative to animal proteins [111]. Moreover, due to its ability to rapidly accumulating omega-3 FAs, essential for human health, Nannochloropsis sp. hold great potential for food application as it can provide a rich nutritional value to microalgae-based products [24].
Beyond the simple use of Nannochloropsis biomass or bioactive compounds, the application of this marine species in food can be further enhanced resorting to co-cultivation principles. As it has been stated in the previous chapters, the synergistic interactions between Nannochloropsis and complementary microorganisms not only enhance biomass productivity and nutrient recycling but also stimulate the production of bioactive metabolites—such as antioxidants, vitamins, and flavor compounds [64]. These approaches can be used to improve the sustainability, the nutritional and the sensory qualities of microalgae food products. Moreover, the incorporation of co-culture systems in food technology can bring further advantages in overcoming Nannochloropsis-biotechnological hurdles, such as lower digestibility and limited bioavailability of intracellular compounds [60]. This represents a gap in scientific research on Nannochloropsis research and an opportunity for further investigation. Co-culture systems offer a more robust nutritional profile, reduced production costs, and improved sustainability, making them an attractive line of research for developing high-quality, functional plant-based foods and alternative protein sources.
The usage of microalgae can be also expanded to the novel field of cellular agriculture due to their nutritional benefits and suitability for co-culture. Innovative research is beginning to explore the co-cultivation of microalgae with animal cells to create integrated systems that could enhance the nutritional profile of cultured meat and seafood [112]. Alike the previously described microbial interactions, microalgae would provide the essential nutrients, vitamins and growth factors that support animal cell proliferation, while simultaneously recycling metabolic waste products such as CO2 and nitrogen and phosphorous compounds [112]. Besides nutritional enhancement, a co-culture system would also reduce the costs associated to cultured food and contribute to a closed-loop production with lower environmental impacts [113]. As a matter of fact, studies have shown that microalgae can successfully be integrated as part of the cultured meat production process in various ways: i) microalgae extracts can be employed as a nutrient source replacing the need to use Fetal Bovine Serum or basal medium [114], ii) recyclers of waste products in the spent culture medium [115], and iii) incorporated in scaffolds to promote thicker and healthier tissues by providing a constant flux of O2 and nutrients to the animal cells in co–culture [116].
In the same way, microalgae from the Nannochloropsis genus have the potential to be introduced in cultivated products, mainly in cultivated seafood production. Their high content in PUFAs would not only contribute to the nutritional enhancement in omega-3 FAs, especially EPA, but also potentially provide the fish flavour and smell to the cultivated product, approximating the cultivated seafood to conventional products [117].
Although no research on direct co-culture systems between marine oleaginous microalgae with animal cells was found, the promising synergy observed in microalgae-microbial systems provides a strong foundation for further exploration. Nannochloropsis sp. constitutes a versatile resource for the development of alternative food products, offering both the nutritional enrichment and process sustainability in emerging cellular agriculture technologies, by taking full advantage of its marine bioactive compounds.
5.3. Toxicology, Safety and Regulatory Aspects of Nannochloropsis sp.
The growing interest in Nannochloropsis sp. derives from its rich chemical composition, particularly in PUFAs, as it is a sustainable and vegetarian source of omega-3 FAs for both feed and food applications. Nonetheless, one of the setbacks related to the integration of microalgae biomass in food is in ensuring its safety for humans and animals. So far, no toxins produced by Nannochloropsis genus are known [19].
A significant setback reported regarding the use of Nannochloropsis in food and feed is related to its lower digestibility, primarily due to its thick cell walls [118]. To overcome this downside cell wall disruption strategies are required. Traditional physical or chemical pre-treatment methods can be costly and energy-consuming [60]. Co-culture strategies offer a more sustainable alternative, by utilizing bacterial or fungal pre-treatments to degrade Nannochloropsis cell wall [60]. To do so, it is important to understand the precise composition and structure of Nannochloropsis cell wall, to select optimal co-culture partners that could hydrolyse the highly resistant algaenan outer layer, while also aiming for non-pathogenic strains [22]. Further investigation should be done on the potential of opting for co-cultivation strategies as sustainable and cost-effective alternatives to improve bioavailability and digestibility of Nannochloropsis.
Additionally, there is a great concern related to the capacity of microalgae, and Nannochloropsis spp., of bioaccumulating contaminant substances intracellularly, such as heavy metals, polychlorinated biphenyls, and toxins present in the surrounding environment [19,119]. To date, there are no reports of allergic reactions directly attributed to Nannochloropsis sp., although its cultivation must be carefully managed under controlled and clean conditions, to minimize risks of contamination. In fact, the major legislation constrains are precisely associated to the safe use of microalgae in food, emphasising the necessity to thoroughly study the toxicity profile of microalgae–based products, before introducing them in human dietary [120].
Summarily, this microalga, by itself, does not constitute a toxic hazard for human consumption, animals or plants. However, more rigorous toxicological studies should be performed. A report published by the European Union (EU) Joint Research Centre classified Nannochloropsis as a safe source of EPA for food supplements in human dietary (with no production of toxins), providing that the quality control regulations are respected [121]. In the EU market, microalgae fall into the Novel Food Regulation No. 2015/2283, which has approved 20%, out of the 150 regularly consumed algae species [120].
While established species such as Arthrospira plattensis (commonly known as Spirulina), Chlorella, and Aphanizomenon flos-aquae are considered Not Novel Foods since they have been largely consumed priorly to 1997, more recently consumed species, such as Odontella aurita, Ulkenia sp., Tetraselmis chuii, Haematococcus pluvialis and Schizochytrium sp, have been approved and categorized as Novel Foods [122,123].
Nannochloropsis sp. is currently on the list of microalgae species To Be Approved for food and food supplements in the EU. As a source of valuable bioactive compounds such as EPA, lipids and arachidonic acid, this specie has drawn attention for its valuable use in food. The EU approval of microalgae species for human consumption is always evolving and does not necessarily accompany the worldwide legislation. In fact, some microalgae and microalgae-byproducts, despite being produced in Europe, are commercialized elsewhere [122]. In fact, there are already Nannochloropsis–based products being commercialized outside Europe: Qualitas Health trades liquid capsules rich in EPA oil secreted by N. oculata, and Optimally Organics produces a nutritional supplement composed by N. gaditana dried powder, laying the grounds for the development of future products [124].
Nannochloropsis sp. inclusion in the EU's Novel Foods list is progressing, with ongoing research and regulatory developments paving the way for its approval. The process is being facilitated as some products of Nannochloropsis species, such as N. gaditana or N. oculata go through the acceptance process, see approval and are commercialized in specific countries [118].
6. Current Challenges and Future Perspectives
The co-cultivation of marine oleaginous microalgae, particularly of the Nannochloropsis genus, with other microorganisms presents a promising approach for enhancing biomass production, nutrient recycling and high-valuable metabolites production, which supports their versatile applications in food, feed, nutraceuticals, biofuel, WWT and bioremediation.
However, several technical, research and scalability challenges must be addressed to take advantage of the full potential of co-culture systems. In fact, scalability has become one of the biggest challenges associated to microalgae-based co-cultures systems. Growth rate disparity, nutrient competition and variable environmental conditions can lead to the dominance of one species over the other, reducing the overall system efficiency [6]. Moreover, a thorough study on the strains utilized in the co-culture system must be performed priorly to ensure that there is a beneficial symbiotic relation that would generate a positive outcome, while opting for strains with potential for large scale cultivation. Additionally, contamination control also poses a significant challenge, being fundamental to maintain selective growth conditions without inhibiting beneficial interactions or cause safety concerns, especially in the production of functional foods [25].
In that sense, further research is needed to identify compatible microbial partners that can enhance the productivity and stability of marine microalgae co-cultures, ensuring that non-pathogenic microorganisms are utilized in food and nutraceutical bioprocessing. Advances in genetic engineering and synthetic biology offer potential solutions for improving compatibility, optimizing metabolic rates and biochemical synthesis. Investigating novel microbial consortia could further expand the range of beneficial interactions and applications. Furthermore, appropriate co-cultivation systems should be designed to better potentiate the simultaneous growth of both species by regulating nutrient availability, light exposure and ensuring overall ideal culture conditions that are essential for optimizing co-cultures performance [125].
Despite these challenges, the future of marine oleaginous microalgae co-cultivation is promising and far from being unravelled. The role of these photosynthetic organisms in microbial consortia is already advancing with several case studies supporting the advantages they bring to numerous biotechnological applications. The ability to enhance the nutritional profile and bioactive compounds in co-cultures makes them a valuable resource for the development of plant-based foods and dietary supplements with marine origin, and therefore research on the theme is highly encouraged. Co-culture approaches should be further investigated as sustainable alternatives to overcome some of the challenges related to Nannochloropsis-bioprocessing, such as low digestibility, harvesting inefficacy while enhancing added-value metabolites production.
Developing innovative co-cultivation strategies could unlock new ways to enhance the bioavailability, functionality, and sensory attributes of Nannochloropsis-based food products, ultimately facilitating its integration into the alternative protein market. The potential co-culture of these microalgae with animal cells, it is still at its early stages, constituting a new avenue of research and holding great potential for future applications in food and cellular agriculture.
7. Conclusions
In this review, it was demonstrated that Nannochloropsis sp., with its high nutritional value, adaptability, and simple cultivation requirements, holds great promise as a versatile co-cultivation partner for various biotechnological applications. Its potential to enhance biomass productivity, nutrient recycling, and bioactive compound production in polycultures, makes it a valuable candidate for industries ranging from food and nutraceuticals to biofuels and wastewater treatment. The value of marine bioactive compounds in human health was detailed as the role of these valuable microorganisms in human and animal dietary is of significant interest, pressing the need to explore novel strategies for introducing marine microalgae co-culture strategies in food technology. However, to fully unlock these opportunities, further research is required to overcome technical challenges such as optimizing species compatibility, balancing nutrient dynamics, and scaling up co-culture systems. Addressing these hurdles through innovative co-culture approaches will be crucial in paving the way for the sustainable and commercially viable application of Nannochloropsis in various sectors. Expanding research efforts in this field will not only improve the efficiency and feasibility of these systems but also contribute to the broader goal of developing more sustainable biotechnological solutions across various industries, particularly in food technology.
Author Contributions
Conceptualization, M.V.E.; writing—original draft preparation, M.V.E.; writing—review and editing, all authors; supervision, N.T.F. and F.C.F.; funding acquisition, N.T.F. and F.C.F. All authors have read and agreed to the published version of the manuscript.
Funding
Thanks for funding from Fundação para a Ciência e Tecnologia for PhD scholarships (2022.12900.BD and 2024.01989.BDANA), CleanFish project (2022.07677.PTDC), iBB (UIDB/04565/2020 and UIDP/04565/2020) and i4HB (LA/P/0140/2020). The Good Food Institute (GFI) supported this work with its competitive Grant Program through the “Algae2Fish” project. The work developed is on the scope of FEASTS (Project ID 101136749) funded by the European Commission funded in the call HORIZON-CL6-2023-FARM2FORK-01-13.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest, other than interests in the field including partnering with interested stakeholders and fostering start-up initiatives. F.C.F. regularly updates the disclosure of his collaborations at https://www.cienciavitae.pt/portal/4C18-FD61-0596, accessed on 13 March 2025.
Abbreviations
The following abbreviations are used in this manuscript:
| DHA | Docosahexaenoic Acid |
| DW | Dry Weight |
| EPA | Eicosapentaenoic Acid |
| EPS | Extracellular Polymeric Substances |
| EU | European Union |
| FA | Fatty Acid |
| IAA | Indole-3-acetic Acid |
| MUFA | Monounsaturated Fatty Acid |
| PUFA | Polyunsaturated Fatty Acid |
| SFA | Saturated Fatty Acid |
| TAG | Triacylglycerol |
| QS | Quorum Sensing |
| WWT | Wastewater Treatment |
References
- J. Wu et al. Bioactive substances and potentiality of marine microalgae. Food Sci Nutr 2021, 9, 5279–5292. [Google Scholar] [CrossRef] [PubMed]
- M. K. Anusree, K. M. K. Anusree, K. Manasa Leela, M. Sreehari, S. Raj, A. Sreenikethanam, and A. K. Bajhaiya. Marine microalgae: an emerging source of pharmaceuticals and bioactive compounds. New Horizons in Natural Compound Research, 2023. [Google Scholar] [CrossRef]
- S. M. Tibbetts, J. E. Milley, and S. P. Lall. Chemical composition and nutritional properties of freshwater and marine microalgal biomass cultured in photobioreactors. J Appl Phycol 2015, 27, 1109–1119. [Google Scholar] [CrossRef]
- J. C. Lee et al.. Marine algal natural products with anti-oxidative, anti-inflammatory, and anti-cancer properties. Cancer Cell Int 2013, 13, 1–7. [Google Scholar] [CrossRef]
- V. Mimouni et al.. The Potential of Microalgae for the Production of Bioactive Molecules of Pharmaceutical Interest. Curr Pharm Biotechnol 2012, 13, 2733–2750. [Google Scholar] [CrossRef]
- P. K. Das, J. Rani, S. Rawat, and S. Kumar. Microalgal Co-cultivation for Biofuel Production and Bioremediation: Current Status and Benefits. BioEnergy Research 2021, 15, 1–26. [Google Scholar] [CrossRef]
- Y. Ye, M. Liu, L. Yu, H. Sun, and J. Liu. Nannochloropsis as an Emerging Algal Chassis for Light-Driven Synthesis of Lipids and High-Value Products. Marine Drugs 2024, 22, 54. [Google Scholar] [CrossRef]
- Y. Ma, Z. Wang, C. Yu, Y. Yin, and G. Zhou. Evaluation of the potential of 9 Nannochloropsis strains for biodiesel production. Bioresour Technol 2014, 167, 503–509. [Google Scholar] [CrossRef]
- X. Wu, R. Ruan, Z. Du, and Y. Liu. Current Status and Prospects of Biodiesel Production from Microalgae. Energies 2012, 5, 2667–2682. [Google Scholar] [CrossRef]
- C. Celi, D. Fino, and F. Savorani. Phaeodactylum tricornutum as a source of value-added products: A review on recent developments in cultivation and extraction technologies. Bioresour Technol Rep 2022, 19, 101122. [Google Scholar] [CrossRef]
- Y. Chisti. Biodiesel from microalgae. Biotechnol Adv 2007, 25, 294–306. [Google Scholar] [CrossRef]
- N. Mishra. EXPLORING THE BIOLOGICALLY ACTIVE METABOLITES OF ISOCHRYSIS GALBANA IN PHARMACEUTICAL INTEREST: AN OVERVIEW. Int J Pharm Sci Res 2018, 2, 2162–2174. [Google Scholar] [CrossRef]
- W. K. Lee et al.. Year-Round Cultivation of Tetraselmis sp. for Essential Lipid Production in a Semi-Open Raceway System. Mar Drugs 2021, 19, 314. [Google Scholar] [CrossRef]
- S. Mehariya et al.. A comprehensive review on versatile microalga Tetraselmis: Potentials applications in wastewater remediation and bulk chemical production. J Environ Manage 2024, 365, 121520. [Google Scholar] [CrossRef] [PubMed]
- G. de S. Celente, T. M. Rizzetti, Y. Sui, and R. de C. de S. Schneider. Potential use of microalga Dunaliella salina for bioproducts with industrial relevance. Biomass Bioenergy 2022, 167, 106647. [Google Scholar] [CrossRef]
- G. Chi et al.. Production of polyunsaturated fatty acids by Schizochytrium (Aurantiochytrium) spp. Biotechnol Adv 2022, 55, 107897. [Google Scholar] [CrossRef]
- Singh, P. S. Nigam, and J. D. Murphy. Renewable fuels from algae: An answer to debatable land based fuels. Bioresour Technol 2011, 102, 10–16. [Google Scholar] [CrossRef]
- L. A. Meireles, A. C. Guedes, and F. X. Malcata. Lipid class composition of the microalga Pavlova lutheri: Eicosapentaenoic and docosahexaenoic acids. J Agric Food Chem 2003, 51, 2237–2241. [Google Scholar] [CrossRef]
- “The Biology of Nannochloropsis oceanica Suda & Miyashita (a microalga). Australian Government, 2019.
- M. W. Fawley, I. Jameson, and K. P. Fawley. The phylogeny of the genus Nannochloropsis (Monodopsidaceae, Eustigmatophyceae), with descriptions of N. australis sp. Nov. and Microchloropsis gen. Nov. Phycologia 2015, 54, 545–552. [Google Scholar] [CrossRef]
- M. R. Brown. The amino-acid and sugar composition of 16 species of microalgae used in mariculture. J Exp Mar Biol Ecol 1991, 145, 79–99. [Google Scholar] [CrossRef]
- M. J. Scholz et al.. Ultrastructure and composition of the Nannochloropsis gaditana cell wall. Eukaryot Cell 2014, 13, 1450–1464. [Google Scholar] [CrossRef]
- M. M. Rebolloso-Fuentes, A. Navarro-Pérez, F. García-Camacho, J. J. Ramos-Miras, and J. L. Guil-Guerrero. Biomass Nutrient Profiles of the Microalga Nannochloropsis. J Agric Food Chem 2001, 49, 2966–2972. [Google Scholar] [CrossRef]
- L. Zanella and F. Vianello. Microalgae of the genus Nannochloropsis: Chemical composition and functional implications for human nutrition. J Funct Foods 2020, 68, 103919. [Google Scholar] [CrossRef]
- G. Padmaperuma, R. V. Kapoore, D. J. Gilmour, and S. Vaidyanathan. Microbial consortia: a critical look at microalgae co-cultures for enhanced biomanufacturing. Crit Rev Biotechnol 2018, 38, 690–703. [Google Scholar] [CrossRef] [PubMed]
- L. Goers, P. Freemont, and K. M. Polizzi. Co-culture systems and technologies: taking synthetic biology to the next level. J R Soc Interface 2014, 11. [Google Scholar] [CrossRef]
- A. Corcoran and W. J. Boeing. Biodiversity Increases the Productivity and Stability of Phytoplankton Communities. PLoS One 2012, 7, e49397. [Google Scholar] [CrossRef]
- J. A. Christie-Oleza, D. Sousoni, M. Lloyd, J. Armengaud, and D. J. Scanlan. Nutrient recycling facilitates long-term stability of marine microbial phototroph-heterotroph interactions. Nat Microbiol 2017, 2, 17100. [Google Scholar] [CrossRef]
- D. M. Selegato and I. Castro-Gamboa. Enhancing chemical and biological diversity by co-cultivation. Front Microbiol 2023, 14, 1117559. [Google Scholar] [CrossRef]
- C. A. Santos and A. Reis. Microalgal symbiosis in biotechnology. Appl Microbiol Biotechnol 2014, 98, 5839–5846. [Google Scholar] [CrossRef]
- J. M. Uratani, R. Kumaraswamy, and J. Rodríguez. A systematic strain selection approach for halotolerant and halophilic bioprocess development: A review. Extremophiles 2014, 18, 629–639. [Google Scholar] [CrossRef]
- S. C. Baker, S. J. Ferguson, B. Ludwig, M. D. Page, O.-M. H. Richter, and R. J. M. van Spanning. Molecular genetics of the genus Paracoccus: metabolically versatile bacteria with bioenergetic flexibility. Microbiol Mol Biol Rev 1998, 62, 1046–1078. [Google Scholar] [CrossRef]
- J. Lian, P. Schimmel, S. Sanchez-Garcia, R. H. Wijffels, H. Smidt, and D. Sipkema. Different co-occurring bacteria enhance or decrease the growth of the microalga Nannochloropsis sp. CCAP211/78. Microb Biotechnol 2021, 14, 1159–1170. [Google Scholar] [CrossRef] [PubMed]
- G. Daly, V. Ghini, A. Adessi, M. Fondi, A. Buchan, and C. Viti. Towards a mechanistic understanding of microalgae–bacteria interactions: integration of metabolomic analysis and computational models. FEMS Microbiol Rev 2022, 46, 1–19. [Google Scholar] [CrossRef]
- J. L. Fuentes, I. Garbayo, M. Cuaresma, Z. Montero, M. González-Del-Valle, and C. Vílchez. Impact of Microalgae-Bacteria Interactions on the Production of Algal Biomass and Associated Compounds. Marine Drugs 2016, 14, 100. [Google Scholar] [CrossRef]
- S. Chaïb, J. C. A. Pistevos, C. Bertrand, and I. Bonnard. Allelopathy and allelochemicals from microalgae: An innovative source for bio-herbicidal compounds and biocontrol research. Algal Res 2021, 54, 102213. [Google Scholar] [CrossRef]
- K. Montgomery, J. C. Charlesworth, R. LeBard, P. T. Visscher, and B. P. Burns. Quorum Sensing in Extreme Environments. Life 2013, 3, 131–148. [Google Scholar] [CrossRef]
- J. Zhou, Y. Lyu, M. L. Richlen, D. M. Anderson, and Z. Cai. Quorum sensing is a language of chemical signals and plays an ecological role in algal-bacterial interactions. CRC Crit Rev Plant Sci 2016, 35, 81. [Google Scholar] [CrossRef]
- E. Segev et al.. Dynamic metabolic exchange governs a marine algal-bacterial interaction. Elife, 2016. [CrossRef]
- S. A. Amin et al.. Interaction and signalling between a cosmopolitan phytoplankton and associated bacteria. Nature 2015, 522, 98–101. [Google Scholar] [CrossRef]
- Y. Ye, M. Liu, L. Yu, H. Sun, and J. Liu. Nannochloropsis as an Emerging Algal Chassis for Light-Driven Synthesis of Lipids and High-Value Products. Mar Drugs 2024, 22, 54. [Google Scholar] [CrossRef]
- J. Li et al.. Choreography of Transcriptomes and Lipidomes of Nannochloropsis Reveals the Mechanisms of Oil Synthesis in Microalgae. Plant Cell 2014, 26, 1645–1665. [Google Scholar] [CrossRef]
- J. Yang et al.. PDAT regulates PE as transient carbon sink alternative to triacylglycerol in Nannochloropsis. Plant Physiol 2022, 189, 1345–1362. [Google Scholar] [CrossRef]
- W. A. Wan Razali, C. A. Evans, and J. Pandhal. Comparative Proteomics Reveals Evidence of Enhanced EPA Trafficking in a Mutant Strain of Nannochloropsis oculata. Front Bioeng Biotechnol 2022, 10, 838445. [Google Scholar] [CrossRef]
- M. M. Morris et al.. Bacterial community assembly, succession, and metabolic function during Nannochloropsis salina outdoor cultivation. 2021. Accessed: Mar. 29, 2025. Available online: https://www.osti. 1876.
- S. K, T. V, J. D, A. P, and R. K. Sarangi. Growth promoting studies on co-culturing Nannochloropsis oceanica with Halomonas aquamarina actively enhance the algal biomass and lipid production. Biocatal Agric Biotechnol 2020, 29, 101790. [Google Scholar] [CrossRef]
- N. Ferrer-Ledo, L. Stegemüller, M. Janssen, R. H. Wijffels, and M. J. Barbosa. Growth and fatty acid distribution over lipid classes in Nannochloropsis oceanica acclimated to different temperatures. Front Plant Sci 2023, 14, 1078998. [Google Scholar] [CrossRef]
- N. Gu, Q. Lin, G. Li, Y. Tan, L. Huang, and J. Lin. Effect of salinity on growth, biochemical composition, and lipid productivity of Nannochloropsis oculata CS 179. Eng Life Sci 2012, 12, 631–637. [Google Scholar] [CrossRef]
- Sukenik, Y. Carmeli, and T. Berner. REGULATION OF FATTY ACID COMPOSITION BY IRRADIANCE LEVEL IN THE EUSTIGMATOPHYTE NANNOCHLOROPSIS SP.1. J Phycol 1989, 25, 686–692. [Google Scholar] [CrossRef]
- W. Kim, M. G. Sung, K. Nam, M. Moon, J. H. Kwon, and J. W. Yang. Effect of monochromatic illumination on lipid accumulation of Nannochloropsis gaditana under continuous cultivation. Bioresour Technol 2014, 159, 30–35. [Google Scholar] [CrossRef]
- L. Xu, X. Cheng, and Q. Wang. Enhanced lipid production in Chlamydomonas reinhardtii by co-culturing with azotobacter chroococcum. Front Plant Sci 2018, 9, 338772. [Google Scholar] [CrossRef]
- L. A. Leyva, Y. Bashan, and L. E. de-Bashan. Activity of acetyl-CoA carboxylase is not directly linked to accumulation of lipids when Chlorella vulgaris is co-immobilised with Azospirillum brasilense in alginate under autotrophic and heterotrophic conditions. Ann Microbiol 2015, 65, 339–349. [Google Scholar] [CrossRef]
- J. Sharma et al.. Microalgal consortia for municipal wastewater treatment – Lipid augmentation and fatty acid profiling for biodiesel production. J Photochem Photobiol B 2020, 202, 111638. [Google Scholar] [CrossRef]
- R. W. Davis, A. J. Siccardi, N. D. Huysman, N. B. Wyatt, J. C. Hewson, and T. W. Lane. Growth of mono- and mixed cultures of Nannochloropsis salina and Phaeodactylum tricornutum on struvite as a nutrient source. Bioresour Technol 2015, 198, 577–585. [Google Scholar] [CrossRef]
- M. Maglie, C. Baldisserotto, A. Guerrini, A. Sabia, L. Ferroni, and S. Pancaldi. A co-cultivation process of Nannochloropsis oculata and Tisochrysis lutea induces morpho-physiological and biochemical variations potentially useful for biotechnological purposes. J Appl Phycol 2021, 33, 2817–2832. [Google Scholar] [CrossRef]
- L. Thurn, J. Schobel, and D. Weuster-Botz. Photoautotrophic Production of Docosahexaenoic Acid- and Eicosapentaenoic Acid-Enriched Biomass by Co-Culturing Golden-Brown and Green Microalgae. Fermentation 2024, 10, 220. [Google Scholar] [CrossRef]
- N. A. T. Tran, B. Tamburic, C. R. Evenhuis, and J. R. Seymour. Bacteria-mediated aggregation of the marine phytoplankton Thalassiosira weissflogii and Nannochloropsis oceanica. J Appl Phycol 2020, 32, 3735–3748. [Google Scholar] [CrossRef]
- A. Corcoran et al.. Scale-dependent enhancement of productivity and stability in xenic Nannochloropsis cultures. Algal Res 2022, 68, 102892. [Google Scholar] [CrossRef]
- R. J. Powell and R. T. Hill. Rapid Aggregation of Biofuel-Producing Algae by the Bacterium Bacillus sp. Strain RP1137. Appl Environ Microbiol 2013, 79, 6093–6101. [Google Scholar] [CrossRef]
- C. Muñoz, C. Hidalgo, M. Zapata, D. Jeison, C. Riquelme, and M. Rivas. Use of cellulolytic marine bacteria for enzymatic pretreatment in microalgal biogas production. Appl Environ Microbiol 2014, 80, 4199–4206. [Google Scholar] [CrossRef]
- Z. Y. Du et al.. Enhancing oil production and harvest by combining the marine alga Nannochloropsis oceanica and the oleaginous fungus Mortierella elongata. Biotechnol Biofuels 2018, 11, 1–16. [Google Scholar] [CrossRef]
- D. Wrede et al.. Co-Cultivation of Fungal and Microalgal Cells as an Efficient System for Harvesting Microalgal Cells, Lipid Production and Wastewater Treatment. PLoS One 2014, 9, e113497. [Google Scholar] [CrossRef]
- T. Ishika, N. R. Moheimani, and P. A. Bahri. Sustainable saline microalgae co-cultivation for biofuel production: A critical review. Renewable and Sustainable Energy Reviews 2017, 78, 356–368. [Google Scholar] [CrossRef]
- M. Maglie, C. Baldisserotto, A. Guerrini, A. Sabia, L. Ferroni, and S. Pancaldi. A co-cultivation process of Nannochloropsis oculata and Tisochrysis lutea induces morpho-physiological and biochemical variations potentially useful for biotechnological purposes. J Appl Phycol 2021, 33, 2817–2832. [Google Scholar] [CrossRef]
- L. Thurn, A. Stock, S. Gerwald, and D. Weuster-Botz. Simultaneous photoautotrophic production of DHA and EPA by Tisochrysis lutea and Microchloropsis salina in co-culture. Bioresour Bioprocess 2022, 9, 1–13. [Google Scholar] [CrossRef]
- L. Thurn, J. Schobel, and D. Weuster-Botz. Photoautotrophic Production of Docosahexaenoic Acid- and Eicosapentaenoic Acid-Enriched Biomass by Co-Culturing Golden-Brown and Green Microalgae. Fermentation 2024, 10, 220. [Google Scholar] [CrossRef]
- Ray, M. Nayak, and A. Ghosh. A review on co-culturing of microalgae: A greener strategy towards sustainable biofuels production. Science of The Total Environment 2022, 802, 149765. [Google Scholar] [CrossRef]
- W. Ding, X. Zhou, M. He, W. Jin, Y. Chen, and J. Sun. Pollutant removal and resource recovery of co-cultivated microalgae Chlorella sp. and Phaeodactylum tricornutum for marine aquaculture wastewater. Journal of Water Process Engineering 2024, 67, 106182. [Google Scholar] [CrossRef]
- M. Padri, N. Boontian, C. Piasai, and M. S. Tamzil. Construction of co-culture of microalgae with microorganisms for enhancing biomass production and wastewater treatment: a review. IOP Conf Ser Earth Environ Sci 2021, 623, 012024. [Google Scholar] [CrossRef]
- S. K, T. V, J. D, A. P, and R. K. Sarangi. Growth promoting studies on co-culturing Nannochloropsis oceanica with Halomonas aquamarina actively enhance the algal biomass and lipid production. Biocatal Agric Biotechnol 2020, 29, 101790. [Google Scholar] [CrossRef]
- Y. Y. Wang et al.. Co-cultivation of Isochrysis galbana and Marinobacter sp. can enhance algal growth and docosahexaenoic acid production. Aquaculture 2022, 556, 738248. [Google Scholar] [CrossRef]
- M. Chorazyczewski, I. S. Huang, H. Abdulla, X. Mayali, and P. V. Zimba. The Influence of Bacteria on the Growth, Lipid Production, and Extracellular Metabolite Accumulation by Phaeodactylum tricornutum (Bacillariophyceae). J Phycol 2021, 57, 931–940. [Google Scholar] [CrossRef]
- D. Wrede et al.. Co-Cultivation of Fungal and Microalgal Cells as an Efficient System for Harvesting Microalgal Cells, Lipid Production and Wastewater Treatment. PLoS One 2014, 9, e113497. [Google Scholar] [CrossRef]
- N. Uduman, Y. N. Uduman, Y. Qi, M. K. Danquah, G. M. Forde, and A. Hoadley. Dewatering of microalgal cultures: A major bottleneck to algae-based fuels. Journal of Renewable and Sustainable Energy, 2010; 1. [Google Scholar] [CrossRef]
- N. A. T. Tran, B. Tamburic, C. R. Evenhuis, and J. R. Seymour. Bacteria-mediated aggregation of the marine phytoplankton Thalassiosira weissflogii and Nannochloropsis oceanica. J Appl Phycol 2020, 32, 3735–3748. [Google Scholar] [CrossRef]
- R. Hajjar, G. Ambaraghassi, H. Sebajang, F. Schwenter, and S. H. Su. Raoultella ornithinolytica: Emergence and Resistance. Infect Drug Resist 2020, 13, 1091. [Google Scholar] [CrossRef] [PubMed]
- Z. Y. Du et al.. Enhancing oil production and harvest by combining the marine alga Nannochloropsis oceanica and the oleaginous fungus Mortierella elongata. Biotechnol Biofuels 2018, 11, 1–16. [Google Scholar] [CrossRef]
- Dias, J. Santos, A. Reis, and T. Lopes da Silva. Yeast and microalgal symbiotic cultures using low-cost substrates for lipid production. Bioresour Technol Rep 2019, 7, 100261. [Google Scholar] [CrossRef]
- N. Arora, A. Patel, J. Mehtani, P. A. Pruthi, V. Pruthi, and K. M. Poluri. Co-culturing of oleaginous microalgae and yeast: paradigm shift towards enhanced lipid productivity. Environmental Science and Pollution Research 2019, 26, 16952–16973. [Google Scholar] [CrossRef]
- R. K. Naidoo, Z. F. Simpson, J. R. Oosthuizen, and F. F. Bauer. Nutrient exchange of carbon and nitrogen promotes the formation of stable mutualisms between chlorella sorokinianaand saccharomyces cerevisiae under engineered synthetic growth conditions. Front Microbiol 2019, 10, 424255. [Google Scholar] [CrossRef]
- J. Ling, S. Nip, W. L. Cheok, R. A. de Toledo, and H. Shim. Lipid production by a mixed culture of oleaginous yeast and microalga from distillery and domestic mixed wastewater. Bioresour Technol 2014, 173, 132–139. [Google Scholar] [CrossRef]
- Dias, A. Reis, J. A. L. Santos, L. Gouveia, and T. Lopes da Silva. Primary brewery wastewater as feedstock for the yeast Rhodosporidium toruloides and the microalga Tetradesmus obliquus mixed cultures with lipid production. Process Biochemistry 2022, 113, 71–86. [Google Scholar] [CrossRef]
- W. Bahafid, N. Tahri Joutey, H. Sayel, I. Boularab, and N. El Ghachtouli. Bioaugmentation of chromium-polluted soil microcosms with Candida tropicalis diminishes phytoavailable chromium. J Appl Microbiol 2013, 115, 727–734. [Google Scholar] [CrossRef]
- S. Cai, C. Hu, and S. Du. Comparisons of Growth and Biochemical Composition between Mixed Culture of Alga and Yeast and Monocultures. J Biosci Bioeng 2007, 104, 391–397. [Google Scholar] [CrossRef]
- Azarpour, S. Zendehboudi, O. Mohammadzadeh, A. R. Rajabzadeh, and I. Chatzis. A review on microalgal biomass and biodiesel production through Co-cultivation strategy. Energy Convers Manag 2022, 267, 115757. [Google Scholar] [CrossRef]
- L. Leng et al.. Co-culture of fungi-microalgae consortium for wastewater treatment: A review. Bioresour Technol 2021, 330, 125008. [Google Scholar] [CrossRef]
- P. K. Das, J. Rani, S. Rawat, and S. Kumar. Microalgal Co-cultivation for Biofuel Production and Bioremediation: Current Status and Benefits. BioEnergy Research 2021, 15, 1–26. [Google Scholar] [CrossRef]
- Y. Tong, K. Honda, and C. J. C. Derek. A review on microalgal-bacterial co-culture: The multifaceted role of beneficial bacteria towards enhancement of microalgal metabolite production. Environ Res 2023, 228, 115872. [Google Scholar] [CrossRef] [PubMed]
- R. Naseema Rasheed et al.. Microalgal co-cultivation -recent methods, trends in omic-studies, applications, and future challenges. Front Bioeng Biotechnol 2023, 11, 1193424. [Google Scholar] [CrossRef]
- W. Ding, X. Zhou, M. He, W. Jin, Y. Chen, and J. Sun. Pollutant removal and resource recovery of co-cultivated microalgae Chlorella sp. and Phaeodactylum tricornutum for marine aquaculture wastewater. Journal of Water Process Engineering 2024, 67, 106182. [Google Scholar] [CrossRef]
- W. -W. Huang, B.-Z. Dong, Z.-P. Cai, and S.-S. Duan. Growth effects on mixed culture of Dunaliella salina and Phaeodactylum tricornutum under different inoculation densities and nitrogen concentrations. Afr J Biotechnol 2011, 10, 13164–13174. [Google Scholar] [CrossRef]
- N. L. Kadalag, P. R. Pawar, and G. Prakash. Co-cultivation of Phaeodactylum tricornutum and Aurantiochytrium limacinum for polyunsaturated omega-3 fatty acids production. Bioresour Technol 2022, 346, 126544. [Google Scholar] [CrossRef]
- M. Chorazyczewski, I. S. Huang, H. Abdulla, X. Mayali, and P. V. Zimba. The Influence of Bacteria on the Growth, Lipid Production, and Extracellular Metabolite Accumulation by Phaeodactylum tricornutum (Bacillariophyceae). J Phycol 2021, 57, 931–940. [Google Scholar] [CrossRef]
- Škufca, *!!! REPLACE !!!*; et al. . Interaction between Microalgae P. tricornutum and Bacteria Thalassospira sp. for Removal of Bisphenols from Conditioned Media. Int J Mol Sci 2022, 23, 8447. [Google Scholar] [CrossRef]
- T. T. Vuong, B. R. Kwon, J. I. Eom, B. K. Shin, and S. M. Kim. Interaction between marine bacterium Stappia sp. K01 and diatom Phaeodactylum tricornutum through extracellular fatty acids. J Appl Phycol 2020, 32, 71–82. [Google Scholar] [CrossRef]
- P. V. Phatarpekar, R. A. Sreepada, C. Pednekar, and C. T. Achuthankutty. A comparative study on growth performance and biochemical composition of mixed culture of Isochrysis galbana and Chaetoceros calcitrans with monocultures. Aquaculture 2000, 181, 141–155. [Google Scholar] [CrossRef]
- H. Wang et al.. Treatment of fishery wastewater by co-culture of Thalassiosira pseudonana with Isochrysis galbana and evaluation of their active components. Algal Res 2021, 60, 102498. [Google Scholar] [CrossRef]
- Y. Y. Wang et al.. Co-cultivation of Isochrysis galbana and Marinobacter sp. can enhance algal growth and docosahexaenoic acid production. Aquaculture 2022, 556, 738248. [Google Scholar] [CrossRef]
- S. V. Sandhya and K. K. Vijayan. Symbiotic association among marine microalgae and bacterial flora: a study with special reference to commercially important Isochrysis galbana culture. J Appl Phycol 2019, 31, 2259–2266. [Google Scholar] [CrossRef]
- M. Kawaroe, T. Prartono, A. Sunuddin, and D. Saputra. Marine Microalgae Tetraselmis suecica as Flocculant Agent of Bio-flocculation Method. Hayati 2016, 23, 62–66. [Google Scholar] [CrossRef]
- J. Park et al.. Phycospheric native bacteria Pelagibaca bermudensis and Stappia sp. Ameliorate biomass productivity of Tetraselmis striata (KCTC1432BP) in co-cultivation system through mutualistic interaction. Front Plant Sci 2017, 8, 244337. [Google Scholar] [CrossRef]
- J. Han, L. Zhang, S. Wang, G. Yang, L. Zhao, and K. Pan. Co-culturing bacteria and microalgae in organic carbon containing medium. Journal of Biological Research 2016, 23, 8. [Google Scholar] [CrossRef]
- N. Muradov et al.. Fungal-assisted algal flocculation: Application in wastewater treatment and biofuel production. Biotechnol Biofuels, 2015. [CrossRef]
- J. Zhang, B. Huang, and T. Tang. Effect of co-culture with Halomonas mongoliensis on Dunaliella salina growth and phenol degradation. Front Bioeng Biotechnol 2022, 10, 1072868. [Google Scholar] [CrossRef]
- T. Butler. The diatom Phaeodactylum tricornutum as a sustainable microalgal cell factory: towards a biorefinery approach. 2021.
- Kusmayadi, Y. K. Leong, H. W. Yen, C. Y. Huang, and J. S. Chang. Microalgae as sustainable food and feed sources for animals and humans – Biotechnological and environmental aspects. Chemosphere 2021, 271, 129800. [Google Scholar] [CrossRef]
- Dineshbabu, G. Goswami, R. Kumar, A. Sinha, and D. Das. Microalgae–nutritious, sustainable aqua- and animal feed source. J Funct Foods 2019, 62, 103545. [Google Scholar] [CrossRef]
- L. R. Martínez-Córdova, M. Emerenciano, A. Miranda-Baeza, and M. Martínez-Porchas. Microbial-based systems for aquaculture of fish and shrimp: an updated review. Rev Aquac 2015, 7, 131–148. [Google Scholar] [CrossRef]
- Tyus. Discovering Potential Protein, Carbohydrate, and Lipid Based Food Ingredients in a Co-Culture of Microalgae. Louisiana State University and Agricultural & Mechanical College. ProQuest Dissertation, s & Theses, 2019, Accessed: Feb. 08, 2025.
- J. Espinosa-Ramírez, A. C. Mondragón-Portocarrero, J. A. Rodríguez, J. M. Lorenzo, and E. M. Santos. Algae as a potential source of protein meat alternatives. Front Nutr 2023, 10, 1254300. [Google Scholar] [CrossRef] [PubMed]
- M. Kent, H. M. Welladsen, A. Mangott, and Y. Li. Nutritional Evaluation of Australian Microalgae as Potential Human Health Supplements. PLoS One 2015, 10, e0118985. [Google Scholar] [CrossRef]
- N. Rojas-Tavara and A. J. Donayre-Torres. Microalgae in lab-grown meat production. https://cjfs.agriculturejournals.cz/doi/10.17221/69/2023-CJFS.html CJFS 2023, 41, 406–418. [Google Scholar] [CrossRef]
- S. Hubalek, M. J. Post, and P. Moutsatsou. Towards resource-efficient and cost-efficient cultured meat. Curr Opin Food Sci 2022, 47, 100885. [Google Scholar] [CrossRef]
- Y. Okamoto, Y. Haraguchi, N. Sawamura, T. Asahi, and T. Shimizu. Mammalian cell cultivation using nutrients extracted from microalgae. Biotechnol Prog 2020, 36, e2941. [Google Scholar] [CrossRef]
- Y. Haraguchi and T. Shimizu. Microalgal culture in animal cell waste medium for sustainable ‘cultured food’ production. Arch Microbiol 2021, 203, 5525–5532. [Google Scholar] [CrossRef]
- Y. Haraguchi and T. Shimizu. Three-dimensional tissue fabrication system by co-culture of microalgae and animal cells for production of thicker and healthy cultured food. Biotechnol Lett 2021, 43, 1117–1129. [Google Scholar] [CrossRef]
- Babuskin Srinivasan, Krishnan K. R., Sivarajan Meenatchisundaram, and Sukumar Muthusamy. Functional foods enriched with marine microalga Nannochloropsis oculata as a source of ω-3 fatty acids. Food Technol Biotechnol 2014, 52, 292–299, Accessed online: https://wwwcabidigitallibraryorg/doi/full/105555/20143328456. [Google Scholar]
- L. Zanella and F. Vianello. Microalgae of the genus Nannochloropsis: Chemical composition and functional implications for human nutrition. J Funct Foods 2020, 68, 103919. [Google Scholar] [CrossRef]
- J. Ji, L. Shi, J. Ampofo, and Lord Abbey. Microalgae: Bioactive Composition, Health Benefits, Safety and Prospects as Potential High-Value Ingredients for the Functional Food Industry. Foods 2022, 11, 1744. [Google Scholar] [CrossRef]
- M. C. Mendes et al.. Algae as Food in Europe: An Overview of Species Diversity and Their Application†. Foods 2022, 11, 1871. [Google Scholar] [CrossRef]
- J. R. Centre et al., Microalgae-based products for the food and feed sector – An outlook for Europe. Publications Office, 2014. [CrossRef]
- “Algae as Novel Food in Europe. European Algae Biomass Association (EABA).
- “Food and Feed Information Portal Database | FIP.” Accessed: Aug. 05, 2024. Online.. Available: https://ec.europa.
- U. Al Hoqani, U. U. Al Hoqani, U. Al-Hoqani, R. Young, and S. Purton. Article in Perspectives in Phycology. 2016. [Google Scholar] [CrossRef]
- L. Goers, P. L. Goers, P. Freemont, and K. M. Polizzi. Co-culture systems and technologies: taking synthetic biology to the next level. J R Soc Interface, 2014; 96. [Google Scholar] [CrossRef]

Table 1.
Marine oleaginous microalgae species with biotechnological interest and their valuable bioactive compounds, total lipid content under optimal conditions and applications.
Table 1.
Marine oleaginous microalgae species with biotechnological interest and their valuable bioactive compounds, total lipid content under optimal conditions and applications.
| Marine microalgae Specie | Bioactive compounds |
Total Lipid content | Applications | Reference |
|---|---|---|---|---|
| Nannochloropsis spp. | PUFAs, EPA | 30% - 60% | Biofuel, nutraceuticals, feed and food | [7,8] |
|
Phaeodactylum tricornutum |
PUFAs, EPA, fucoxanthin | 20 - 30% | Nutraceuticals, pharmaceuticals, biofuel, cosmetics | [9,10] |
| Isochrysis galbana | PUFAs, DHA and EPA | 25 - 30 % | Aquaculture feed, nutraceuticals | [11,12] |
| Tetraselmis spp. | PUFAs | 12 - 15 % | Aquaculture feeds, biofuel and wastewater treatment | [13,14] |
| Dunaliella salina | PUFAs, Beta-carotene, lutein | 7 % | Biofuel, cosmetics, nutraceuticals and pharmaceuticals | [15] |
| Schizochytrium spp. | PUFAs, DHA | (up to 60% under stress) | DHA-rich oils, oral vaccines | [16,17] |
| Pavlova lutheri | PUFAs, DHA, EPA | 50 - 70 % | Aquaculture feed, biofuel | [18] |
Table 2.
Published literature on the co–culture of Nannochoropsis sp. with other microorganisms and respective application.
Table 2.
Published literature on the co–culture of Nannochoropsis sp. with other microorganisms and respective application.
| Co-culture type | Microalgae Specie | Co-culture Species | Application | Reference |
|---|---|---|---|---|
|
Microalgae- microalgae |
Nannochloropsis sp. | MAC11: Chlorella sp., Chlamydomonas reinhardtii, Scenedesmus bijugatus, and Oscillatoria MAC2: Chlorella sp., Kirchnella, Scenedesmus dimorphus, and Microcoleus* |
- Sewage wastewater treatment - Heavy metal removal by bioaugmentation |
[53] |
| M. salina |
Phaeodactylum tricornutum |
- Wastewater treatment | [54] | |
| N. oculata | Tisochrysis lutea | - Biomass growth and lipid production | [55] | |
| N. oceanica | Isochrysis galbana | - Biomass growth and lipid production | [56] | |
|
Microalgae- bacteria |
N. oceanica | Halomonas aquamarina | - Biomass growth and lipid production | [46] |
| N. oceanica | Bacterial isolates from the Rhodobacterales, Flavobacteriales and Sphingomonadale orders | - Facilitate biomass aggregation | [57] | |
| N. oceanica | Genera Algoriphagus, Oceanicaulis, and Marinobacter | - Enhance productivity and stability of cell cultures | [58] | |
| N. oceanica | Bacillus sp. | - Cell aggregation for biofuel production | [59] | |
| M. gaditana | Raoultella ornithinolytica | - Cell wall degradation for biofuel production | [60] | |
| Microalgae-fungi | N. oceanica | Mortierella elongata | - Biofuel production - Harvesting efficiency |
[61,62] |
| N. oculata | Aspergillus fumigatus | - Wastewater Treatment - Lipid production - Harvesting efficiency |
[62] |
1 MAC – Microalgae Consortia.
Table 3.
Published literature on the co–culture of various marine oleaginous microalgae species with other microorganisms and respective biotechnological application.
Table 3.
Published literature on the co–culture of various marine oleaginous microalgae species with other microorganisms and respective biotechnological application.
| Marine microalgae specie | Co-culture type | Co-culture partners | Application | Reference |
|---|---|---|---|---|
|
Phaeodactylum tricornutum |
Microalgae- microalgae |
Chlorella sp. | - Marine aquaculture wastewater treatment | [90] |
| Dunaliella salina | - Biomass growth, lipid and chlorophyll production | [91] | ||
| Aurantiochytrium limacinum | - EPA and DHA production | [92] | ||
| Microalgae-bacteria | Marinobacter sp. | - Biomass growth and lipid production | [93] | |
| Thalassospira sp. | - Bisphenols removal from media | [94] | ||
| Stappia sp. | - Biomass growth, lipid and carotenoid production | [95] | ||
|
Isochrysis galbana |
Microalgae- microalgae |
Chaetoceros calcitrans | - Added-value metabolites production | [96] |
| Microalgae-bacteria | Thalassiosira pseudonana | - Fishery wastewater treatment | [97] | |
| Marinobacter sp. | - Biomass growth and DHA production | [98] | ||
| Alteromonas sp. | - Biomass growth and metabolites production | [99] | ||
| Labrenzia sp. | - Biomass growth and metabolites production | [99] | ||
| Microalgae-yeast | Ambrosiozyma cicatricosa | - Biomass growth | [84] | |
| Tetraselmisspp. | Microalgae- microalgae |
T. lutea and Microchloropsis salina | - EPA and DHA production | [65] |
|
T. sueccia and Chlorella sp., Nannochloropsis sp. |
- Bio-flocculation for cell harvesting | [100] | ||
| Microalgae-bacteria |
T. striata and Pelagibaca bermudensis, Stappia sp. |
Biomass growth and lipid production | [101] | |
| T. chuii and Muricauda sp. | - Biomass growth | [102] | ||
| Microalgae-fungi |
T. suecica and Aspergillus fumigatus |
- Bio-flocculation for cell harvesting, biomass growth and lipid production | [103] | |
|
Dunaliella salina |
Microalgae-bacteria | Halomonas mongoliensis | - Bisphenols removal from wastewater | [104] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



