
Submitted:
14 April 2025
Posted:
15 April 2025
You are already at the latest version
Abstract
This study aims to investigate the endogenous gibberellin levels and related genes analysis of noxious invasive weed Heracleum sosnowskyi. Genome-wide identification, phylogenetic analysis, conserved motif analysis and gene structure characterization of GA-oxidases were performed. We analysed endogenous GAs levels and the expression of target HsGAoxs in response to GA3 within H. sosnowskyi developing ovaries. Twenty-seven HsGAoxs genes were identified, distributed across eleven chromosomes. Phylogenetic analysis classified proteins into the HsGA20ox, C19-HsGA2ox and HsGA3ox subfamilies, facilitating functional predictions. Among the thirteen HsGA2ox protein members, there were no C20-GA2ox subfamily that distinguish H. sosnowskyi from other model plant species. Analysis of gene structure and conserved motifs confirmed the phylogenetic grouping and suggested that the evolutionary pattern was maintained within these subfamilies. The observed increase in precursor and bioactive GA levels provides evidence that they play a crucial role in promoting fruit growth. Ovary phenotypes reflected the timing of peak gibberellin levels, specifically during the cell expansion period. Exogenous GA3 treatment promoted HsGA3ox1 expression within both the central and lateral regions of the umbel ovaries. Overall, the results show that GA levels are precisely regulated by multiple HsGAox genes for stable early fruit development, and that disturbances in this stability affect fruit development. This opens up the possibility of investigating the role of GA in H. sosnowskyi fruit formation and developing measures for invasion control.
Keywords:
Sosnowsky’s hogweed
; Apiaceae
; invasive plant
; gibberellic acid
; genome-wide analysis
; early fruit development
; 2-oxoglutarate-dependent dioxygenases
1. Introduction
Gibberellins (GAs) are a large group of plant hormones, encompassing 136 chemically similar compounds found in plants, fungi, and bacteria [1,2]. They belong to diterpene carboxylic acids with a tetracyclic ent-gibberellane skeleton, composed of 19–20 carbon atoms [3]. Biologically active GAs act as endogenous plant growth and development regulators, transmitting internal and environmental signals within the organism [1]. They stimulate cell elongation, cell division, regulate seed germination, control the transition between juvenile and generative plant phases, induce flowering, influence sex determination, and play a role in fruit setting and growth processes [4,5]. However, only a small group are biologically active: gibberellin A1 (GA1), gibberellin A3 (GA3), gibberellin A4 (GA4), and gibberellin A7 (GA7) [1]. Interestingly, GA1 and GA4 are the dominant forms in plant tissues [6,7,8]. GA3 and GA7 metabolism in plants is not a common process, and these GA species are present in trace amounts, however, microorganisms can metabolize GA20 to GA3, or GA7 directly from GA4, involving GA desaturases [9,10]. For plants, it is crucial to regulate GA levels precisely; however, the chemical structure specificity of 1,2-unsaturated GAs (GA3 and GA7) obstructs their inactivation in plants by 2β-hydroxylation, which results in their prolonged activity and may explain their low amounts in plants [2,10].
GA biosynthesis has been studied in many plant species, and it occurs in three stages according to subcellular localization, where involved enzymes act [4,11]. The first stage of GA biosynthesis takes place in plastids, where geranylgeranyl diphosphate is converted to ent-kaurene through an intermediate, catalysed by terpene synthases: ent-copalyl diphosphate synthase (CPS) and ent-kaurene synthase (KS) [12,13]. Subsequently, in the outer plastid membrane and endoplasmic reticulum, cytochrome P450 monooxygenases catalyse oxidation reactions, during which ent-kaurene is converted through four intermediates to GA12 [14,15]. The third stage of synthesis occurs in the cell cytoplasm, where the common precursor GA12 is oxidised by two distinct soluble 2-oxoglutarate-dependent dioxygenases (2-ODDs) – GA 20-oxidase (GA20ox) and GA 3-oxidase (GA3ox) – leading to the biologically active GA1 or GA4 [11,16,17]. To keep optimal levels of active GAs species within tissues, the inactivation process is essential for plant growth and development. The main mechanism involves two groups of 2-ODDs enzymes, differentiated by their substrate specificity: C19-GA 2-oxidase and C20-GA 2-oxidase, both of which utilise 2β-hydroxylation to catabolize precursors and biologically active GAs into inactive products [2,18,19,20].
The 2-ODD superfamily contains four enzyme subfamilies (GA20ox, GA3ox, C19-GA2ox and C20-GA2ox), involved in GA biosynthesis and catabolism, which are encoded by small multigene families and are conserved between multiple species [19,21,22,23]. Genes of GAoxs subfamilies were first isolated in model plant species: GA20ox in pumpkin [24], GA3ox in Arabidopsis thaliana [25], GA2ox in runner bean (Phaseolus coccineus) and in A. thaliana [26]. To date, GAoxs genes have been identified in many commercially important plants such as: cucumber [18], watermelon [27], tomato [28,29,30], maize [31], rice [32,33], peach [34], grapevine [35], wild cherry, wild strawberry [36], breadfruit [37] and others. With the identification of GAoxs enzymes and their genes, the mechanism of fruit-set became better understood, which opened opportunities to manipulate fruit-set and induce parthenocarpy [28,29,38,39].
H. sosnowskyi is a noxious invasive weed species enlisted into Invasive Alien Species of Union concern and national lists of invasive species in many EU countries [40,41]. H. sosnowskyi propagates only by seeds and after bearing them, eventually dies [42,43]. Interestingly, Pastinaca sativa (Apiaceae) is adapted to cope with some pests by producing seedless fruits. This strategy enables it to regulate outbreaks of herbivores [44]. Previous studies have indicated an exogenous GA3 effect in inducing seedlessness in H. sosnowskyi [45,46]. However, to date, analysis of endogenous GAs and related genes has not been available. With the recent article on the H. sosnowskyi genome, GAoxs and gene analysis have become accessible in this invasive species [47].
In this study, we identified 27 putative GAoxs enzymes in the H. sosnowskyi genome. Phylogenetic analysis clustered these proteins into three subgroups: GA20ox, GA3ox and C19-GA2ox. We identified the effects of exogenous GA3 on the expression of HsGA20ox1, HsGA3ox1 and HsGA2ox1, as well as on the endogenous GA profiles in gradually opening H. sosnowskyi flowers. These results open opportunities to further analyse the role of GAs in H. sosnowskyi fruit-set mechanism and develop invasion control strategies.
2. Results
2.1. Genome-Wide Identification and Analysis of GA-Oxidase Genes in Heracleum sosnowskyi
To identify GAox genes from GA20ox, GA2ox and GA3ox subfamilies in Heracleum sosnowskyi, the amino acid sequences of GA20ox, GA2ox and GA3ox from Arabidopsis thaliana and Oryza sativa were used as a reference in BLASTp program against the H. sosnowskyi genome. A total of 32 putative protein homologues were obtained. However, 5 protein candidates did not possess 2OG-Fe(II) oxygenase (PF03171) or non-haem dioxygenase N-terminal (PF14226) domains, as confirmed by InterPro Pfam (available online: http:// https://www.ebi.ac.uk/interpro) (Table S1). Thus, we concluded that H. sosnowskyi contains 27 GAox family members. We identified and renamed GA20ox, GA2ox and GA3ox genes in the H. sosnowskyi genome: 9 HsGA20ox1-9, 13 HsGA2ox1-13 and 5 HsGA3ox1-5 (Table 1).
The CDS lengths of the HsGA20ox, HsGA2ox and HsGA3ox genes varied from 978 (HsGA20ox7) to 1221 nt (HsGA20ox4), from 945 (HsGA2ox8) to 1665 nt (HsGA2ox2) and from 1020 (HsGA3ox5) to 1068 nt (HsGA3ox2), respectively. In HsGAox families, amino acid residue ranges slightly differed: HsGA20ox (325-406 aa), HsGA2ox (314-554 aa) and HsGA3ox (339-355 aa). The theoretical molecular weight ranged from 36.81 to 46.14 kDa (HsGA20ox), for HsGA2ox from 35.39 to 61.92 kDa and for HsGA3ox from 38.00 to 40.02 kDa (Table 1). Using Plant-mPLoc software, we identified the subcellular localization of the HsGAox proteins. Analysis revealed that all members from HsGA20ox, HsGA2ox and HsGA3ox are localized in the cytoplasm. Interestingly, only one member, HsGA2ox2 protein, displayed localization in both the cytoplasm and nucleus (Table 1).
To determine the exact genomic positions of HsGAox genes, chromosome localization was carried out. A total of 27 HsGAox genes were widely distributed across eleven H. sosnowskyi chromosomes (Figure 1). HsGA20ox gene subfamily members were identified to be distributed in chromosomes 1, 2, 3, 7 and 8. The vast majority of gene loci are localized in the lower arm of the chromosomes; however, HsGA20ox7 and HsGA20ox8 are situated in the upper arm and in the middle of chromosome 3, respectively. Thirteen HsGA2ox genes are scattered across six chromosomes. Eight HsGA2ox1, 2, 5, 6, 9, 10, 11, 12 loci are located in the lower arm of the chromosomes 2, 4, 5, 6, 10 and 11. The rest of the genes localized in the upper arm of the chromosomes 6, 10, and 11. The H. sosnowskyi GA3ox subfamily genes are scattered among chromosomes 5, 9, 10 and 11. HsGA3ox1, 4 and 5 are located in the lower arm of chromosomes 11 and 5, respectively. The HsGA3ox3 locus was located in the end of the upper arm of chromosome 9. Interestingly, chromosomes 1-6 are considered longer than others, but HsGAox genes are distributed across all chromosomes and do not show a dominant correlation to chromosome length.
2.2. Phylogenetic Analysis of the HsGAox Genes
To explore the phylogenetic relationship of HsGAox with homologues from other taxa, we involved two model species, Arabidopsis thaliana and Oryza sativa, in the analysis. A maximum-likelihood (ML) phylogenetic tree was built based on alignments of the complete protein sequences from 27 Heracleum sosnowskyi, 16 A. thaliana, and 21 O. sativa found in NCBI and Phytozome databases. Four subfamilies (GA20ox, C19-GA2ox, C20-GA2ox and GA3ox) were identified. However, H. sosnowskyi proteins clustered into 3 different subfamilies: GA20ox, C19-GA2ox and GA3ox. Interestingly, among thirteen HsGA2ox members, none belonged to the C20-GA2ox subfamily. Moreover, HsGA20ox1 and -2, HsGA20ox5 and -6 appear to be duplicate proteins. These results allow us to make speculations on the functions of the HsGA20ox, HsGA2ox, and HsGA3ox according to the phylogenetic classification mentioned above (Figure 2).
2.3. Conserved Motif and Gene Structure Analysis of the HsGAox Genes
To support the phylogenetic analysis, we predicted conserved motifs of the HsGAox protein family using the Multiple Em for Motif Elicitation (MEME) online tool. 10 conserved motifs for HsGA20ox, C19-HsGA2ox and HsGA3ox subfamily proteins were obtained and ranged from 21 to 50 amino acid residues (Figure 3B). The assessment of motifs 1–10 with InterPro Pfam revealed that motif 1 exhibits 2OG-Fe(II) oxygenase and motif 2 exhibits non-haem dioxygenase N-terminal domains. Except for HsGA3ox, all HsGA20oxs and HsGA2oxs were endowed with 8 motifs in the same pattern. In contrast, HsGA3oxs contained 7 motifs (HsGA3ox4 had 6 motifs), moreover, motif composition of the HsGA3oxs was distinct from HsGA20oxs and HsGA2oxs by the C-terminal end, where motif 6 or motif 8 was absent. In more detail, HsGA20oxs subfamily proteins are distinct from other HsGAoxs, containing a family specific motif 6 at N-terminal end of the sequence. Furthermore, motif 10 is unique for HsGA20ox and HsGA3ox subfamily members; in contrast, at the same location of HsGA2ox sequences, motif 5 is observed.
We analysed the intron and exon compositions, as well as dispositions in the gene structure of HsGAoxs (Figure 3C). Analysis revealed that the majority of HsGA20ox members had a consistent number of 3 exons and 2 introns, whereas HsGA20ox4 and -7 number of exons varied from 4 to 5, respectively. HsGA2ox subfamily genes contained 3-5 exons: 10 members (HsGA2ox1,3,5-12) had 3 exons, HsGA2ox4 – 4 exons and HsGA2ox2,13 – 5 exons. The HsGA3ox subfamily (HsGA3ox1-5) showed a relatively simple gene structure, consisting of 2 exons and 1 intron. In summary, the length of introns varied within three HsGAoxs gene subfamilies, contributing to the overall size differences among gene members in these subfamilies. To sum up, gene structure and motif analyses support the phylogenetic findings, indicating the conserved evolution of these gene subfamilies in all three species.
2.4. Analysis of HsGAoxs Expression in Response to GA3 Within Heracleum sosnowskyi Developing Ovaries
To evaluate the exogenous GA3 effect on the expression of GA biosynthesis enzyme genes, the Real-time quantitative PCR (qPCR) method was used to determine the quantity of gene transcripts in the terminal umbel ovaries, which were collected from the central and lateral parts at different stages of development. Specific primers for the putative HsGA20ox1, HsGA3ox1, and HsGA2ox1 genes were designed and used in the analysis. It was found that the HsGA3ox1 gene was most intensely expressed at 3 and 10 days after the start of the experiment, both in the central and lateral parts of the terminal umbel (Figure 4B). The quantity of HsGA20ox1 transcripts in the terminal umbel ovaries peaked 3 days after the start of the experiment, but the expression of this gene decreased after 10 days from the onset of flower spreading (Figure 4B). Due to the effect of GA3, the relative abundance of HsGA20ox1 gene transcripts decreased by almost half on the third day of the experiment, reaching 8.9 % in the central part (p = 0.04) and 8.7 % in the lateral part (p = 0.85). After 10 days, the expression of this gene was even weaker in both the central (2.4 %) and lateral (2.2 %) parts. Analysis of HsGA3ox1 gene expression revealed that 3 days after the GA3 treatment, the expression of this gene decreased from 41.5 % to 7.18 % (p = 0.8) in the central part of the umbel and from 45.3 % to 7.27 % (p = 0.72) in the lateral part. Ten days after the GA3 treatment, the relative abundance of HsGA3ox1 gene transcripts significantly increased, reaching 235 % (p < 0.001) in the central part and 168 % (p = 0.0019) in the lateral part of the umbel.
2.5. Changes in Endogenous GA levels in Heracleum sosnowskyi Ovaries After Treatment with Exogenous GA3
The dynamics of endogenous GAs were also analysed in another experiment in which we aimed to determine the distribution of metabolites of the C-13-hydroxylated GA biosynthesis pathway in the central and lateral parts of the terminal umbel after GA3 treatment (Figure 4C). The obtained results revealed that the level of biologically active GA1 was extremely low in both parts of the umbel at 0 and 3 days after application (DAA): 963 pg/g FW in the central part and 264 pg/g FW in the lateral part. However, at a later stage of ovary development (10 DAA), the hormone level in the central part of the umbel significantly increased to 16.3 ng/g FW (p = 0.011) and to 43.6 ng/g FW (p < 0.001) in the lateral part. Interestingly, the GA1 content in the ovaries of the central and lateral parts differed significantly, by almost a twofold difference (p < 0.001). The level of the precursor GA20 reached its peak in the central (72.47 ng/g FW) and lateral (46.64 ng/g FW) parts of the umbel during the 3 DAA period. However, during the 10 DAA period, the level of this metabolite significantly decreased to 4.37 ng/g FW (p < 0.001) and 2.87 ng/g FW (p = 0.005) in the ovaries of the central and lateral parts, respectively. The level of the GA20 catabolite (GA29) remained low throughout the experiment. The level of the GA1 catabolic form (GA8) increased in the central part (12.8 ng/g FW) and in the lateral part (7.3 ng/g FW) during the 3 DAA period, and then significantly decreased to 1.19 ng/g FW (p < 0.001) and 0.89 ng/g FW (p = 0.059) in each part of the umbel, respectively, during the later period (10 DAA). The highest levels of early hormone precursors GA44 and GA19 in the terminal ovaries were detected only at the 3 DAA developmental stage. Interestingly, in the early stage of ovary development, GA44 was found four times more abundant in the central part of ovary tissues (32.2 ng/g FW) than in the lateral part (7.35 ng/g FW), and the difference between them was significant (p < 0.001). After spraying the terminal umbels with exogenous GA3, the GA1 content increased insignificantly in the ovaries of the central part from 0.963 to 5.15 ng/g FW (p = 0.95), and significantly in the lateral part from 0.264 to 20.9 ng/g FW (p < 0.001) after 3 DAA. 10 days after application with GA3 solution, an uneven distribution of GA1 hormone levels was observed depending on the umbel area: significantly decreased in the central part to 388 pg/g FW (p < 0.001) and significantly decreased in the lateral part to 26.86 ng/g FW (p < 0.01). Due to GA3 treatment, the level of catabolite GA29 increased dramatically in the ovaries of the lateral part of the umbel: 6.66 ng/g FW (p < 0.001) and 6.28 ng/g FW (p < 0.01) in the 3 DAA and 10 DAA periods, respectively.
2.6. GA3 Impact on the Phenotype of Heracleum sosnowskyi Ovaries
Morphometric analysis of the samples revealed that the development of mericarps is not uniform in the central and lateral parts of the umbel: after 10 days, the mericarps in the central part of the umbel were 11 % shorter (U = 2000.5, p < 0.001), 10 % narrower (U = 2385, p < 0.01), and 9 % lighter (U = 2185.5, p < 0.001) than the mericarps from the lateral part of the umbel (Figure 4B). Three days after GA3 150 mg/l treatment, there was a statistically significant decrease in mericarp length (U = 1459, p < 0.001) and weight (U = 2004, p < 0.01) in the central part, and width (U = 2203.5, p < 0.001) in the lateral part. Ten days after application, there was a statistically significant decrease in mericarp length (21 %) (U = 1656, p < 0.001), width (30 %) (U = 1020, p < 0.001), and weight (59 %) (U = 1008.5, p < 0.001) in the central part; and a decrease in length (20 %) (U = 1337, p < 0.001), width (31 %) (U = 712.5, p < 0.001), and weight (41 %) (U = 703, p < 0.001) in the lateral part (Figure 4B).
3. Discussion
Gibberellins play an important role in the processes of fruit set, development, and ripening [48,49]. In horticulture, plants are often sprayed with bioactive GAs to obtain high-quality seedless fruits, but this can have a negative impact on the histological and morphological structure of the fruit [50,51,52,53]. The 2-ODD protein superfamily is the second-largest enzyme family in plants. Moreover, 2-ODDs have a crucial role in leading oxygenation or hydroxylation in various plant metabolic events [19]. Members of 2-ODDs have been identified in many model plant species: Arabidopsis thaliana, rice, cucumber, tomato and others [18,26,54,55,56,57,58,59]. However, information about 2-ODDs in invasive Heracleum sosnowskyi is very limited. A recent study by [47] made the genome of H. sosnowskyi available for genome-wide analysis. According to GAoxs protein sequences from A. thaliana and Oryza sativa [60], we identified and named 9 HsGA20ox1-9, 13 HsGA2ox1-13 and 5 HsGA3ox1-5 protein homologues from H. sosnowskyi (Table 1). All putative GAoxs contained 2OG-Fe(II) oxygenase (PF03171) and non-haem dioxygenase N-terminal (PF14226) domains, which are known to be specific for 2-ODDs [61,62]. In our study, we predicted the 3D protein structures of several HsGAoxs and identified that both previously named domains are located in the reaction crevice of the proteins (Figure 3A). This aligns with the previous models of suggested GAoxs in rice [63]. Interestingly, HsGA20ox1 and HsGA20ox2, seem to be duplicates, which have one amino acid substitution caused by single nucleotide polymorphism (Figure 3A). Gene duplication is generally seen as a major driver of genes developing different functions, a process steered by natural selection, which is important to consider in further HsGAox analysis [64,65]. Phylogenetic analysis suggested that HsGA20oxs belong to three distinct clades, similarly, HsGA3oxs seem to belong to two different paraphyletic groups. Interestingly, we have not found any members of C20-GA2oxs (Figure 2). However, the functional characterization using C20- and C19-GA substrates of HsGA2oxs might improve our classification. Previous studies indicate that GA2oxs are composed of three groups: C19-GA2ox-I, C19-GA2ox-II, and C20-GA2ox-I [11,58,66,67]. In our study, we observed that H. sosnowskyi C19-2ox members cluster together with O. sativa and A. thaliana members into two monophyletic groups: C19-GA2ox-I and C19-GA2ox-II (Figure 2). These findings suggest that C19-HsGA2oxs might have functional differences among the groups. Furthermore, it is known that different GAox genes display significant variations in their expression across different tissues [39,68,69,70,71]. Comparison of the phylogenetic tree with the gene structure and motif analysis, it is apparent that the most closely related members within subfamilies share similar gene structure and motif composition (Figure 2; Figure 3). Interestingly, HsGA20ox and HsGA2ox are found in greater numbers than HsGA3oxs, similarly to A. thaliana, O. sativa, Cucumis sativus, Salix matsudana [18,63,72,73]. To delve into the evolutionary relationships of GAox genes, we analysed motif composition and gene structure (Fig. 3). However, GA20ox and GA3ox subfamilies were distinct from GA2ox by the LPWKET motif, which is characteristic of GA20oxs and for some GA3oxs [39,63,72]. Whereas motif 9 was observed in all C19-HsGA2oxs, it is distinct from the rest of the 2-ODD subfamilies [63]. Gene structure revealed that all HsGA20oxs contain three exons and two introns except for HsGA20ox4 and -7, which are similar to the structure of the GA20ox subfamily in A. thaliana [58]. HsGA3oxs have a simple gene structure of one intron and two exons, consistent with those in A. thaliana, C. sativus and O. sativa [58,60]. Our findings reveal that closely related members within GAox subfamilies possess similar structures and motifs, implying shared functions.
A more detailed analysis of terminal umbel ovaries in the early stages of fruit development revealed a differential distribution of GA and metabolites between the central and lateral regions. This was particularly reflected in a significant increase in the levels of metabolites GA44 and GA20 in the control samples of the central umbel region at 0 and 3 days after application (DAA) (Figure 4D). This inter-regional distribution may be due to the complex umbel architecture, as the flowers in the lateral part of the umbel open first [74]. In addition, the literature indicates that GA is involved in the sex determination of male flowers [75,76,77], with high levels of GA precursors and the hormone GA4 accumulating in the floral parts of stamens [78]. As is known, male flowers in the umbels of some Apiaceae family representatives, such as Zizia aurea and Thaspium barbinode, are distributed centripetally [79]. However, the terminal umbel of Heracleum mantegazzianum is usually composed only of hermaphrodite flowers [74]. There is no literature data on the distribution of hermaphrodite and male flowers in the terminal umbel of H. sosnowskyi but based on our endogenous GA profiles (Figure 4D), it can be assumed that there should be more of them in the centre of the H. sosnowskyi umbel. Three days after exogenous GA3 treatment, the accumulation of the metabolite GA20 was significantly reduced in the ovaries of both parts of the umbel, but the level of the hormone GA1 in the ovary tissues increased (Figure 4D). Ten days after application, the synthesis of the hormone GA1 in the ovary tissues of the central part of the umbel was completely suppressed, but only partially in the lateral part of the umbel. The inhibitory effect of exogenous GA3 on GA biosynthesis can be identified by a significant decrease in the precursor GA20 and a marked increase in the amount of the catabolite GA29 in the ovaries of both parts of the umbel. However, it remains unclear why exogenous GA3 forms a GA1 gradient in different parts of the umbel (Figure 4D). It is known that an increase in the level of biologically active GA forms in the early stages of fruit development leads to the development of parthenocarpic fruit set [38,49,51,80,81]. It should be noted that in this experiment, flower pollination was not controlled, so the GA1 content at 10 DAA could have increased due to natural fertilization (Figure 4D).
Before analysing the abundance of GA biosynthesis gene (HsGA20ox1, HsGA3ox1, and HsGA2ox1) transcripts, these genes were cloned from H. sosnowskyi umbel tissues collected 10 days after application (DAA) and specific primers were designed for them. It was observed that in the early stage of fruit development (3 DAA), HsGA20ox1 was more highly expressed, but its expression decreased in later stages (10 DAA). On the other hand, the expression of the metabolic HsGA3ox1 gene was much more intense, with no differences detected between umbel regions. Other studies have indicated that the increase in GA20ox gene expression and the decrease in GA2ox are essential factors in regulating GA biosynthesis during fruit set [48,51,82,83]. It should also be noted that in other plant species (e.g., white clover, tomato, cultivated rice), more than one GA oxidase gene is involved in fruit set control [28,49,78,80,84]. However, our results suggest that intense fruit development is dependent on endogenous GA, which is supported by the obtained levels of endogenous GA (Figure 4E). Additionally, the results of phenotypic analysis of ovaries (Figure 4B) correspond to the peak of gibberellin during the cell expansion phase observed in the model plant tomato [85,86]. However, the effect of exogenous GA3 stimulated HsGA3ox1 expression in the central and lateral parts of the umbel ovaries, while the inhibition of HsGA2ox1 gene expression, as speculated, was not detected (Figure 4E). Furthermore, the obtained expression profile of the HsGA20ox1 gene at 10 DAA, both in the control and after GA3 treatment, suggests that fruit set had already occurred, as seen in some species such as pear, where after a similar treatment with exogenous biologically active GA, the expression of genes PbGA20ox1, PbGA20ox2, and PbGA20ox3 is suppressed [39]. Our results align with the suppressed expression of paralogous genes of the PbGA20ox (PbGA20ox1, PbGA20ox3) in pollinated pear ovaries [39].
4. Materials and Methods
4.1. Research Object and Growth Conditions
An invasive habitat of Heracleum sosnowskyi was selected for field trials, and an experimental field was established. The test site was located in an anthropogenic H. sosnowskyi habitat with an area of 6.48 ha in an abandoned area in Vilnius, Lithuania (54.739749°N, 25.258871°E WGS80). The experiment field was fenced with a “stop” line and marked with signs informing about the plant research being conducted. The studies were conducted on H. sosnowskyi plants during the flowering and fruit formation stages from 2021. Samples were collected from 3 plants. The terminal umbel was divided into central and lateral parts following the guidelines [87].
4.2. Application of Plants with GA3
Terminal umbels of Heracleum sosnowskyi were sprayed with the phytohormone gibberellic acid (GA3) (SERVA, Heidelberg, Germany) at a 150 mg/l concentration. Certain order umbels were sprayed twice. Distilled water was used as a control spray. Manual sprayers “Venus” (KWAZAR, Budy-Grzybek, Poland) were used for spraying. Inflorescences were sprayed from a distance of 15-20 cm, evenly applying 18.5 ml onto each umbel.
4.3. Sample Harvesting
Samples for morphological and anatomical analysis of Heracleum sosnowskyi flowers and mericarps were collected in August-September from terminal umbels. To study ovary development dynamics in terminal umbels, ovary samples were collected from the central and lateral parts of the terminal umbel at 0, 3 and 10 days after application (at the beginning of flowering). For biochemical and molecular studies, flowers of different developmental stages were collected, weighed, flash-frozen in liquid nitrogen and stored in a low-temperature freezer (Skadi Green line, ES) at -80 °C until analysis.
4.4. Bioinformatic Analysis of Heracleum sosnowskyi GAoxs Subfamilies
In order to perform a comprehensive phylogenomic analysis of the GAox subfamilies in Heracleum sosnowskyi, a prerequisite was the in silico extraction of all potential GAox homolog sequences from publicly available whole-genome sequence data. We extracted 16 Arabidopsis thaliana and 21 rice GAox sequences from NCBI Phytozome and Rice Genome Annotation Project databases. These sequences were grouped according to protein subfamilies and used as query sequences against H. sosnowskyi genome (NCBI: PRJNA928505) in BLASTp [88]. Overall, 32 amino acid homologues were obtained and revised. Candidates were accepted if they shared at least 40% identity and had an expected threshold e-value ≤1.0 × e-10. Subsequently, HsGAoxs protein candidates were submitted to InterPro Pfam database to identify the highly conserved and characteristic 2OG-FeII_Oxy (PF03171) and DIOX_N (PF14226) domains of the 2-ODDs superfamily [61,62]. Conserved motif analysis of HsGAoxs was performed using Multiple Em for Motif Elicitation (MEME) 5.5.7 software with default parameters [89]. The exon-intron gene structure of HsGAoxs genes was analysed using GSDS 2.0 online software [90]. Based on the CDS region range found in the NCBI database annotation of each HsGAox protein, we visualized gene locations on H. sosnowskyi chromosomes using MapGene2Chromosome v2 online tool [91]. In a subsequent analysis, the molecular weights of all HsGAox proteins were calculated using the Protein Molecular Weight tool from the bioinformatics.org website. Additionally, the Plant-mPLoc 2.0 software was applied to predict the subcellular localizations of HsGAox proteins [92]. Protein structures of HsGAoxs were predicted using Alphafold 3 [93], and the predicted model structure quality was assessed by the VoroMQA online tool using default parameters [94].
4.5. Multiple Sequence Alignment and Phylogenetic Analysis
Multiple sequence alignment of the GAoxs protein sequences from Heracleum sosnowskyi, Arabidopsis thaliana and Oryza sativa was conducted using MEGA X software through the application of MUSCLE (Multiple Sequence Comparison by Log-Expectation) algorithm with default parameters [95].
Optimal model fit for aligned protein sequences was generated, and a phylogenetic tree was constructed using Maximum Likelihood (ML) and Le-Gascuel model by MEGA 12 software [96]. The robustness of the phylogenetic inference was evaluated using 500 bootstrap replicates.
4.6. Extraction of Endogenous GAs
Endogenous GAs were extracted from fresh mericarp tissues that had been frozen in liquid nitrogen (N2) according to the specified methodology [22]. GA extraction consisted of three stages. The first stage entailed sample extraction with 80 % methanol (purity ≥ 99.95 %, Carl Roth GmbH + Co. KG, Karlsruhe, Germany), introduction of a deuterated 17,17-d2-GA standard mixture (internal standard) into the sample (purity ≥ 90 %, OlChemim, Olomouc, Czech Republic), and liquid-liquid extraction. The second stage involved solid-phase extraction using anion exchange “Bond Elut DEA” (Agilent, Santa Clara, USA) and reverse-phase “Sep-pak C18” (Waters, Wexford, Ireland) columns. The third stage was derivatization, during which the extracted samples were methylated with freshly prepared diazomethane according to De Boer and Backer [97] and trimethylsilylated with N-methyl-N-(trimethylsilyl)trifluoroacetamide (MSTFA) (purity ≥ 95 %, Macherey-Nagel, Düren, Germany). After extraction, the samples were evaporated under a stream of N2 gas and stored at –20 °C until analysis by GC-MS method.
4.7. Endogenous GA Analysis by Gas Chromatography-Mass Spectrometry
Qualitative analysis of endogenous Gas was performed by GC-MS using a “GC/MS-Q2010 PLUS” (Shimadzu, Kyoto, Japan) gas chromatography system coupled with single quadrupole “GC-MS-QP2010 ULTRA” (Shimadzu, Kyoto, Japan) mass spectrometer. 1 μl of the sample was manually injected into the injector of the gas chromatography system coupled to a non-polar “BPX5” column (30 m × 0.25 mm × 0.25 μm) (SGE Analytical Science, Kiln Farm Milton Keynes, United Kingdom). Helium was used as the carrier gas. The split ratio was 30:1 for sample introduction into the column. The initial column temperature was 60 °C and was immediately raised at a rate of 45 °C/min to 220 °C, followed by a gradual increase at 4 °C/min intervals up to 300 °C. The interface temperature was 240 °C. The analysis time for one sample was 25 min. Data were recorded in SIM (selected ion monitoring) mode after 5 min from injection. Target ions for GA qualitative analysis were used for the corresponding GAs and d2-GA: 300 and 302 (GA12), 239 and 241 (GA15), 314 and 316 (GA24), 270 and 272 (GA9), 284 and 286 (GA4), 506 and 508 (GA34), 284 and 286 (GA51), 207 and 209 (GA53), 207 and 209 (GA44), 374 and 376 (GA19), 418 and 420 (GA20), 506 and 508 (GA1), 594 and 596 (GA8), 506 and 508 (GA29). Compound identification was based on their retention times and the presence of additional characteristic ions in the obtained mass spectra, by comparing them with the known mass spectra of labelled and unlabelled GA methyl esters [98]. Endogenous GA levels were calculated based on peak areas, with corrections for the abundance of natural isotopes in the samples and the presence of unlabelled GAs in the internal GA standards [99].
4.8. Identification of GA Biosynthesis Gene Fragments
To amplify putative Heracleum sosnowskyi GA 20-oxidase (GA20ox), GA 3-oxidase (GA3ox) and GA 2-oxidase (GA2ox) gene fragments, a PCR based method was applied [22,100]. Based on the conserved amino acid sequences and nucleotide sequences of the GA20ox and GA3ox enzyme subfamilies in wild carrot (Daucus carota) (Apiaceae), degenerate primers were designed to amplify these genes. For GA2ox, a degenerated primer pair from cucumber (Cucumis sativus) (Cucurbitacea) was used. cDNA molecules were synthesized from frozen fresh mericarp samples of H. sosnowskyi. The resulting cDNA was used as a template in a PCR reaction, which was performed using the “Phusion High-Fidelity PCR Master Mix” (Thermo Scientific, Vilnius, Lithuania) kit, according to the manufacturer’s protocol. PCR products were analysed by electrophoresis on a 2 % agarose gel, stained with “Midori Green Advance” (Nippon Genetics, Duren, Germany) dye and visualized with a “FastGeneR GelPic LED Box” device (Nippon Genetics, Duren, Germany). The expected size of the DcGA20ox PCR product was 813 base pairs (bp), the expected size of the DcGA3ox product was 497 bp, and the expected size of the CsGA2ox product was 300 bp. PCR product bands were cut from the agarose gel and samples were purified using the “Gene JETTM PCR Purification Kit” according to the manufacturer’s instructions (Thermo Scientific, Vilnius, Lithuania). Samples were prepared for sequencing according to the recommendations of “Microsynth SEQLAB” (Germany). Sequencing results allowed for the identification of regions in the H. sosnowskyi genome based on which specific primer pairs were designed. Using “NCBI PrimerBLAST” and “Primer3” software, forward (FW) and reverse (REV) primers were generated for the putative H. sosnowskyi HsGA20ox1, HsGA3ox1, HsGA2ox1 genes (Table S1).
4.9. Real-Time Quantitative PCR
Real-time quantitative PCR (qPCR) was performed to assess the transcript levels of late gibberellin biosynthesis enzyme (GA20ox, GA3ox, GA2ox) genes in Heracleum sosnowskyi ovary samples. Specific primers were used in the qPCR analysis to evaluate the expression of the genes encoding these proteins (Table S1). 1 µl of cDNA diluted 5 times with double distilled water was used as the template for the qPCR reaction. The total reaction mixture volume was 15 µl. Reactions were performed in an “AZURE CIELOTM Real-time PCR System” (Azure Biosystems, Dublin, USA) thermocycler using the two-step cycle protocol of the “Maxima SYBR Green noROX qPCR Master Mix (2X)” (Thermo Scientific, Vilnius, Lithuania) kit. Cycling conditions consisted of one cycle at 95 °C for 10 min and 40 cycles at 95 °C for 15 s followed by 60 °C for 1 min. The cucumber (Cucumis sativus) β-actin gene (AB010922) was used as the internal standard. The relative abundance of each gene was calculated as the average of two technical replicates using the “Real-time PCR Miner” bioinformatics software and expressed as a percentage [101]. Technical replicates with a Ct difference ≥ 0.5 from the mean were discarded. All experiments were repeated twice with three biological replicates.
4.10. Statistical Analysis
Using descriptive statistical methods, the obtained results were presented as arithmetic means with standard errors (mean ± SE). The normal distribution of data was assessed using the Shapiro-Wilk test. If the assumption of data normality was met, parametric methods were employed, and the homoscedasticity of the data was assessed using Levene’s test with Brown-Forsythe correction. If the data were not normally distributed, statistically significant differences between group medians were assessed using the Kruskal-Wallis H-test. For pairwise comparisons between experimental groups, the Mann-Whitney U test or Dunn’s z test was applied. Statistically significant differences were indicated when p < 0.05. Data statistical analysis and graphical representation were performed using “PAST 4.16” and “R” version 4.0.2 in “RStudio” version 1.3.1093 with “ggplot2” package [102,103].
5. Conclusions
A total of 27 HsGAox genes were widely distributed across eleven Heracleum sosnowskyi chromosomes. Phylogenetic classification allows us to make speculations about the functions of the HsGA20ox HsGA2ox, and HsGA3ox. The analysed H. sosnowskyi proteins clustered into three different subfamilies. Among thirteen HsGA2ox members, none belonged to the C20-GA2ox subfamily, contrary to the two subfamilies of GA2oxs in other plant species. Gene structure and motif analyses support the phylogenetic findings, indicating the conserved evolution of these gene subfamilies across three species. Intense fruit development is dependent on endogenous GA, as supported by the obtained levels of endogenous GA. The phenotypic analysis of ovaries corresponds to the peak of gibberellin during the cell expansion phase. However, the effect of exogenous GA3 stimulated HsGA3ox1 expression in the central and lateral parts of the umbel ovaries. Overall, these results confirm that GA homeostasis is a tightly regulated, complex system including multiple HsGAox genes. It maintains a stable hormone level in the early stages of fruit development. A high or low level of the hormone in a particular umbel part can disrupt the development of the fruit. These results open opportunities to further analyse the role of GAs in H. sosnowskyi fruit-set mechanism and develop invasion control strategies.
In this study, 27 HsGAox genes were identified in H. sosnowskyi, distributed across eleven chromosomes. Phylogenetic analysis revealed three distinct HsGAox subfamilies, indicating the functional roles of HsGA20ox, HsGA2ox, and HsGA3ox. The HsGA2ox group, which is distinct from other plant species and lacks C20-GA2ox. The conserved motif and gene structure patterns across species support the phylogenetic classification. It was found that endogenous gibberellin (GA) levels are crucial for robust fruit development, and ovary phenotypes coincide with the highest GA levels during cell expansion. External GA3 application upregulated HsGA3ox1 in the ovary regions of the umbels. Complex GA homeostasis, regulated by multiple HsGAox genes, is crucial for stable early fruit development; GA imbalances in the umbel parts negatively affect fruit formation. These findings provide a basis for investigating the role of GA in H. sosnowskyi fruit formation and developing strategies to control the invasive spread.
Author Contributions
Conceptualization, T.Ž. and S.J.; methodology, J.B., D.L. and T.Ž.; software, J.B., D.L., S.R. and T.Ž; formal analysis, S.R. and S.J.; investigation, D.L., J.B. and T.Ž.; data curation, S.J and T.Ž.; writing–original draft preparation, T.Ž.; writing—review and editing, D.L., S.R., J.B. and S.J.; visualization, D.L., S.R. and T.Ž; supervision, S.J. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Nature Research Centre R&D II programme.
Data Availability Statement
The data supporting the reported results can be found in the archive of scientific reports of the Nature Research Centre.
Acknowledgments
We thank Nijolė Bareikienė, engineer of the Laboratory of Plant Physiology of the Nature Research Center, for the support and help provided.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Sponsel, V.M. Signal Achievements in Gibberellin Research: The Second Half-century. In Annual Plant Reviews, Volume 49: The Gibberellins; Wiley Online Library, 2016; pp. 1–36. [Google Scholar]
- Hedden, P. The Current Status of Research on Gibberellin Biosynthesis. Plant Cell Physiol 2020, 61, 1832–1849. [Google Scholar] [CrossRef] [PubMed]
- Hedden, P.; Thomas, S.G. Gibberellin Biosynthesis and Its Regulation. Biochemical Journal 2012, 444, 11–25. [Google Scholar] [CrossRef] [PubMed]
- Hedden, P. Gibberellins. In Encyclopedia of Applied Plant Sciences; Elsevier Inc., 2016; Vol. 1, pp. 411–420. ISBN 9780123948083. [Google Scholar]
- Hedden; Thomas. In The Gibberellins; 2016; Vol. 1, ISBN 9788578110796.
- Lange, T.; Hedden, P.; Graebe, J.E. Gibberellin Biosynthesis in Cell-Free Extracts from Developing Cucurbita maxima Embryos and the Identification of New Endogenous Gibberellins. Planta 1993, 189, 350–358. [Google Scholar] [CrossRef]
- MacMillan, J. Occurrence of Gibberellins in Vascular Plants, Fungi, and Bacteria. J Plant Growth Regul 2001, 20, 387–442. [Google Scholar] [CrossRef]
- Curaba, J.; Moritz, T.; Blervaque, R.; Parcy, F.; Raz, V.; Herzog, M.; Vachon, G. AtGA3ox2, a Key Gene Responsible for Bioactive Gibberellin Biosynthesis, Is Regulated during Embryogenesis by Leafy Cotyledon2 and FUSCA3 in Arabidopsis. Plant Physiol 2004, 136, 3660–3669. [Google Scholar] [CrossRef] [PubMed]
- Ward, D.A.; MacMillan, J.; Gong, F.; Phillips, A.L.; Hedden, P. Gibberellin 3-Oxidases in Developing Embryos of the Southern Wild Cucumber, Marah macrocarpus. Phytochemistry 2010, 71, 2010–2018. [Google Scholar] [CrossRef]
- Bhattacharya, A.; Kourmpetli, S.; Ward, D.A.; Thomas, S.G.; Gong, F.; Powers, S.J.; Carrera, E.; Taylor, B.; Gonzalez, F.N. de C.; Tudzynski, B.; et al. Characterization of the Fungal Gibberellin Desaturase as a 2-Oxoglutarate-Dependent Dioxygenase and Its Utilization for Enhancing Plant Growth. Plant Physiol 2012, 160, 837–845. [Google Scholar] [CrossRef]
- Yamaguchi, S. Gibberellin Metabolism and Its Regulation. Annu Rev Plant Biol 2008, 59, 225–251. [Google Scholar] [CrossRef]
- Aach, H.; Bode, H.; Robinson, D.G.; Graebe, J.E. Ent-Kaurene Synthase Is Located in Proplastids of Meristematic Shoot Tissues. Planta 1997, 211–219. [Google Scholar] [CrossRef]
- Sun, T.P.; Gubler, F. Molecular Mechanism of Gibberellin Signalling in Plants. Annu Rev Plant Biol 2004, 55, 197–223. [Google Scholar] [CrossRef]
- Helliwell, C.A.; Sullivan, J.A.; Mould, R.M.; Gray, J.C.; James Peacock, W.; Dennis, E.S. A Plastid Envelope Location of Arabidopsis Ent-Kaurene Oxidase Links the Plastid and Endoplasmic Reticulum Steps of the Gibberellin Biosynthesis Pathway. Plant Journal 2001, 28, 201–208. [Google Scholar] [CrossRef]
- Nelson, D.R.; Schuler, M.A.; Paquette, S.M.; Werck-Reichhart, D.; Bak, S. Comparative Genomics of Rice and Arabidopsis. Analysis of 727 Cytochrome P450 Genes and Pseudogenes from a Monocot and a Dicot. Plant Physiol 2004, 135, 756–772. [Google Scholar] [CrossRef]
- Pimenta Lange, M.J.; Lange, T. Gibberellin Biosynthesis and the Regulation of Plant Development. Plant Biol 2006, 8, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Sponsel, V.M.; Hedden, P. Gibberellin Biosynthesis and Inactivation. In Plant Hormones; Springer, 2010; pp. 63–94. [Google Scholar]
- Pimenta Lange, M.J.; Liebrandt, A.; Arnold, L.; Chmielewska, S.M.; Felsberger, A.; Freier, E.; Heuer, M.; Zur, D.; Lange, T. Functional Characterization of Gibberellin Oxidases from Cucumber, Cucumis sativus L. Phytochemistry 2013, 90, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Kawai, Y.; Ono, E.; Mizutani, M. Evolution and Diversity of the 2-Oxoglutarate-Dependent Dioxygenase Superfamily in Plants. Plant Journal 2014, 78, 328–343. [Google Scholar] [CrossRef]
- Pearce, S.; Huttly, A.K.; Prosser, I.M.; Li, Y.D.; Vaughan, S.P.; Gallova, B.; Patil, A.; Coghill, J.A.; Dubcovsky, J.; Hedden, P.; et al. Heterologous Expression and Transcript Analysis of Gibberellin Biosynthetic Genes of Grasses Reveals Novel Functionality in the GA3ox Family. BMC Plant Biol 2015, 15. [Google Scholar] [CrossRef] [PubMed]
- Rebers, M.; Kaneta, T.; Kawaide, H.; Yamaguchi, S.; Yang, Y.Y.; Imai, R.; Sekimoto, H.; Kamiya, Y. Regulation of Gibberellin Biosynthesis Genes during Flower and Early Fruit Development of Tomato. Plant Journal 1999, 17, 241–250. [Google Scholar] [CrossRef]
- Lange, T.; Kappler, J.; Fischer, A.; Frisse, A.; Padeffke, T.; Schmidtke, S.; Lange, M.J.P. Gibberellin Biosynthesis in Developing Pumpkin Seedlings. Plant Physiol 2005, 139, 213–223. [Google Scholar] [CrossRef]
- Sakamoto, T.; Miura, K.; Itoh, H.; Tatsumi, T.; Ueguchi-Tanaka, M.; Ishiyama, K.; Kobayashi, M.; Agrawal, G.K.; Takeda, S.; Abe, K.; et al. An Overview of Gibberellin Metabolism Enzyme Genes and Their Related Mutants in Rice. Plant Physiol 2004, 134, 1642–1653. [Google Scholar] [CrossRef]
- Lange, T. Purification and Partial Amino-Acid Sequence of Gibberellin 20-Oxidase from Cucurbita Maxima L. Endosperm. 1994; Vol. 195. [Google Scholar]
- Chiang, H.-H.; Hwang, L.; Goodman, H.M. Lsolation of the Arabidopsis GA4 Locus; American Society of Plant Physiologists, 1995; Vol. 7. [Google Scholar]
- Thomas, S.G.; Phillips, A.L.; Hedden, P. Molecular Cloning and Functional Expression of Gibberellin 2-Oxidases, Multifunctional Enzymes Involved in Gibberellin Deactivation. Plant Biol 1999, 96, 4698–4703. [Google Scholar] [CrossRef]
- Kang, H.-G.; Jun, S.-H.; Kim, J.; Kawaide, H.; Kamiya, Y.; An, G. Cloning and Molecular Analyses of a Gibberellin 20-Oxidase Gene Expressed Specifically in Developing Seeds of Watermelon. Plant Physiol 1999, 121, 373–382. [Google Scholar] [CrossRef]
- García-Hurtado, N.; Carrera, E.; Ruiz-Rivero, O.; López-Gresa, M.P.; Hedden, P.; Gong, F.; García-Martínez, J.L. The Characterization of Transgenic Tomato Overexpressing Gibberellin 20-Oxidase Reveals Induction of Parthenocarpic Fruit Growth, Higher Yield, and Alteration of the Gibberellin Biosynthetic Pathway. J Exp Bot 2012, 63, 5803–5813. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Bello, L.; Moritz, T.; López-Díaz, I. Silencing C19-GA 2-Oxidases Induces Parthenocarpic Development and Inhibits Lateral Branching in Tomato Plants. J Exp Bot 2015, 66, 5897–5910. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Wang, X.; Zhang, L.; Lin, S.; Liu, D.; Wang, Q.; Cai, S.; El-Tanbouly, R.; Gan, L.; Wu, H.; et al. Identification and Characterization of Tomato Gibberellin 2-Oxidases (GA2oxs) and Effects of Fruit-Specific SlGA2ox1 Overexpression on Fruit and Seed Growth and Development. Hortic Res 2016, 3. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Shan, X.; Jiang, Z.; Zhao, L.; Jin, F. Genome-Wide Identification and Expression Analysis of the GA2ox Gene Family in Maize (Zea mays L.) under Various Abiotic Stress Conditions. Plant Physiology and Biochemistry 2021, 166, 621–633. [Google Scholar] [CrossRef]
- Toyomasu, T.; Kawaide, H.; Sekimoto, H.; von Numers, C.; Phillips, A.L.; Hedden, P.; Kamiya Toyomasu, Y. ; Numers, von Frontier Research Pro-Gram, The Institute of Physical and Chemical Research. 1997.
- Lo, S.-F.; Yang, S.-Y.; Chen, K.-T.; Hsing, Y.-I.; Zeevaart, J.A.D.; Chen, L.-J.; Yu, S.-M. A Novel Class of Gibberellin 2-Oxidases Control Semidwarfism, Tillering, and Root Development in Rice. Plant Cell 2008, 20, 2603–2618. [Google Scholar] [CrossRef]
- Cheng, J.; Ma, J.; Zheng, X.; Lv, H.; Zhang, M.; Tan, B.; Ye, X.; Wang, W.; Zhang, L.; Li, Z. Functional Analysis of the Gibberellin 2-Oxidase Gene Family in Peach. Front Plant Sci 2021, 12, 619158. [Google Scholar] [CrossRef]
- Giacomelli, L.; Rota-Stabelli, O.; Masuero, D.; Acheampong, A.K.; Moretto, M.; Caputi, L.; Vrhovsek, U.; Moser, C. Gibberellin Metabolism in Vitis Vinifera L. During Bloom and Fruit-Set: Functional Characterization and Evolution of Grapevine Gibberellin Oxidases. J Exp Bot 2013, 64, 4403–4419. [Google Scholar] [CrossRef]
- Sabir, I.A.; Manzoor, M.A.; Shah, I.H.; Abbas, F.; Liu, X.; Fiaz, S.; Shah, A.N.; Jiu, S.; Wang, J.; Abdullah, M.; et al. Evolutionary and Integrative Analysis of Gibberellin-Dioxygenase Gene Family and Their Expression Profile in Three Rosaceae Genomes (F. Vesca, P. Mume, and P. Avium) Under Phytohormone Stress. Front Plant Sci 2022, 13. [Google Scholar] [CrossRef]
- Zhou, Y.; Underhill, S.J.R. Breadfruit (Artocarpus Altilis) Gibberellin 2-Oxidase Genes in Stem Elongation and Abiotic Stress Response. Plant Physiology and Biochemistry 2016, 98, 81–88. [Google Scholar] [CrossRef]
- Fos, M.; Nuez, F.; Garcı́a-Martı́nez, J.L. The Gene Pat-2, Which Induces Natural Parthenocarpy, Alters the Gibberellin Content in Unpollinated Tomato Ovaries. Plant Physiol 2000, 122, 471–480. [Google Scholar] [CrossRef]
- Wang, H.; Wu, T.; Liu, J.; Cong, L.; Zhu, Y.; Zhai, R.; Yang, C.; Wang, Z.; Ma, F.; Xu, L. PbGA20ox2 Regulates Fruit Set and Induces Parthenocarpy by Enhancing GA4 Content. Front Plant Sci 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Gederaas, L.; Loennechen Moen, T.; Skjelseth, S.; Larsen, L.K. Alien Species in Norway - with the Norwegian Black List 2012; Norwegian Biodiversity Infomation Centre (NBIC): Trondheim, 2012; ISBN 9788292838372. [Google Scholar]
- Gudžinskas, Z.; Kazlauskas, M.; Pilate, D.; Balalaikins, M.; Pilats, M.; Šaulys, A.; Šaulienė, I.; Šukienė, L. Invasive Organisms the Border Region of Lithuania and Latvia; BMK Press: Vilnius, 2014. [Google Scholar]
- Tkachenko, K.G. Peculiarities and Seed Productivity in Some Heracleum Species Grown in Leningrad Area. Rastitelnye Resursy 1989, 1, 52–61. [Google Scholar]
- Gudžinskas, Z.; Žalneravičius, E. Seedling Dynamics and Population Structure of Invasive Heracleum Sosnowskyi (Apiaceae) in Lithuania. Ann Bot Fenn 2018. [Google Scholar] [CrossRef]
- Zangerl, A.R.; Berenbaum, M.R.; Nitao, J.K. Parthenocarpic Fruits in Wild Parsnip: Decoy Defence against a Specialist Herbivore. Evol Ecol 1991, 5, 136–145. [Google Scholar] [CrossRef]
- Koryznienė, D.; Jurkonienė, S.; Žalnierius, T.; Gavelienė, V.; Jankovska-Bortkevič, E.; Bareikienė, N.; Būda, V. Heracleum Sosnowskyi Seed Development under the Effect of Exogenous Application of GA3. PeerJ 2019, 2019, 1–14. [Google Scholar] [CrossRef]
- Žalnierius, T.; Šveikauskas, V.; Aphalo, P.J.; Gavelienė, V.; Būda, V.; Jurkonienė, S. Gibberellic Acid (GA3) Applied to Flowering Heracleum Sosnowskyi Decreases Seed Viability Even If Seed Development Is Not Inhibited. Plants 2022, 11, 314. [Google Scholar] [CrossRef]
- Schelkunov, M.I.; Shtratnikova, V.Y.; Klepikova, A. V; Makarenko, M.S.; Omelchenko, D.O.; Novikova, L.A.; Obukhova, E.N.; Bogdanov, V.P.; Penin, A.A.; Logacheva, M.D. The Genome of the Toxic Invasive Species Heracleum Sosnowskyi Carries an Increased Number of Genes despite Absence of Recent Whole-genome Duplications. The Plant Journal 2024, 117, 449–463. [Google Scholar] [CrossRef]
- Rieu, I.; Eriksson, S.; Powers, S.J.; Gong, F.; Griffiths, J.; Woolley, L.; Benlloch, R.; Nilsson, O.; Thomas, S.G.; Hedden, P. Genetic Analysis Reveals That C19-GA 2-Oxidation Is a Major Gibberellin Inactivation Pathway in Arabidopsis. Plant Cell 2008, 20, 2420–2436. [Google Scholar] [CrossRef]
- Dorcey, E.; Urbez, C.; Blázquez, M.A.; Carbonell, J.; Perez-Amador, M.A. Fertilization-Dependent Auxin Response in Ovules Triggers Fruit Development through the Modulation of Gibberellin Metabolism in Arabidopsis. Plant Journal 2009, 58, 318–332. [Google Scholar] [CrossRef]
- Vivian-Smith, A.; Koltunow, A.M. Genetic Analysis of Growth-Regulator-Induced Parthenocarpy in Arabidopsis. Plant Physiol 1999, 121, 437–451. [Google Scholar] [CrossRef] [PubMed]
- Serrani, J.C.; Fos, M.; Atarés, A.; García-Martínez, J.L. Effect of Gibberellin and Auxin on Parthenocarpic Fruit Growth Induction in the Cv Micro-Tom of Tomato. J Plant Growth Regul 2007, 26, 211–221. [Google Scholar] [CrossRef]
- Tiwari, A.; Offringa, R.; Heuvelink, E. Auxin-Induced Fruit Set in Capsicum Annuum L. Requires Downstream Gibberellin Biosynthesis. J Plant Growth Regul 2012, 31, 570–578. [Google Scholar] [CrossRef]
- Asahira, T.; Nitsch, J.P. Tubérisation in Vitro: Ullucus Tuberosus et Dioscorea. Bulletin de la Societe Botanique de France 1968, 115, 345–352. [Google Scholar] [CrossRef]
- Shohat, H.; Cheriker, H.; Kilambi, H.V.; Illouz Eliaz, N.; Blum, S.; Amsellem, Z.; Tarkowská, D.; Aharoni, A.; Eshed, Y.; Weiss, D. Inhibition of Gibberellin Accumulation by Water Deficiency Promotes Fast and Long-term ‘Drought Avoidance’Responses in Tomato. New Phytologist 2021, 232, 1985–1998. [Google Scholar] [CrossRef]
- Toyomasu, T.; Kawaide, H.; Sekimoto, H.; von Numers, C.; Phillips, A.L.; Hedden, P.; Kamiya Toyomasu, Y. ; Numers, von Frontier Research Pro-Gram, The Institute of Physical and Chemical Research. 1997.
- Sakamoto, T.; Miura, K.; Itoh, H.; Tatsumi, T.; Ueguchi-Tanaka, M.; Ishiyama, K.; Kobayashi, M.; Agrawal, G.K.; Takeda, S.; Abe, K.; et al. An Overview of Gibberellin Metabolism Enzyme Genes and Their Related Mutants in Rice. Plant Physiol 2004, 134, 1642–1653. [Google Scholar] [CrossRef] [PubMed]
- Itoh, H.; Ueguchi-Tanaka, M.; Sentoku, N.; Kitano, H.; Matsuoka, M.; Kobayashi, M. Cloning and Functional Analysis of Two Gibberellin 3-Hydroxylase Genes That Are Differently Expressed during the Growth of Rice.
- Han, F.; Zhu, B. Evolutionary Analysis of Three Gibberellin Oxidase Genesin Rice, Arabidopsis, and Soybean. Gene 2011, 473, 23–35. [Google Scholar] [CrossRef]
- Phillips, A.L.; Ward, D.A.; Uknes, S.; Appleford, N.E.J.; Lange, T.; Huttly, A.K.; Gaskin, P.; Graebe, J.E.; Hedden, P. Isolation and Expression of Three Gibberellin 20-Oxidase CDNA Clones from Arabidopsis. Plant Physiol 1995, 108, 1049–1057. [Google Scholar] [CrossRef]
- Sun, H.; Pang, B.; Yan, J.; Wang, T.; Wang, L.; Chen, C.; Li, Q.; Ren, Z. Comprehensive Analysis of Cucumber Gibberellin Oxidase Family Genes and Functional Characterization of CsGA20ox1 in Root Development in Arabidopsis. Int J Mol Sci 2018, 19. [Google Scholar] [CrossRef]
- Hedden, P.; Phillips, A.L. Gibberellin Metabolism: New Insights Revealed by the Genes. Trends Plant Sci 2000, 5, 523–530. [Google Scholar] [CrossRef]
- Aravind, L.; Koonin, E. V The DNA-Repair Protein AlkB, EGL-9, and Leprecan Define New Families of 2-Oxoglutarate-and Iron-Dependent Dioxygenases. 2001. [Google Scholar]
- Huang, Y.; Wang, X.; Ge, S.; Rao, G.Y. Divergence and Adaptive Evolution of the Gibberellin Oxidase Genes in Plants Genome Evolution and Evolutionary Systems Biology. BMC Evol Biol 2015, 15. [Google Scholar] [CrossRef] [PubMed]
- Lynch, M.; Conery, J.S. The Evolutionary Fate and Consequences of Duplicate Genes. Science (1979) 2000, 290, 1151–1155. [Google Scholar] [CrossRef] [PubMed]
- Sémon, M.; Wolfe, K.H. Consequences of Genome Duplication. Curr Opin Genet Dev 2007, 17, 505–512. [Google Scholar] [CrossRef]
- Chen, S.; Wang, X.-J.; Tan, G.-F.; Zhou, W.-Q.; Wang, G.-L. Gibberellin and the Plant Growth Retardant Paclobutrazol Altered Fruit Shape and Ripening in Tomato. Protoplasma 2020, 257, 853–861. [Google Scholar] [CrossRef]
- Schomburg, F.M.; Bizzell, C.M.; Lee, D.J.; Zeevaart, J.A.D.; Amasino, R.M. Overexpression of a Novel Class of Gibberellin 2-Oxidases Decreases Gibberellin Levels and Creates Dwarf Plants. Plant Cell 2003, 15, 151–163. [Google Scholar] [CrossRef]
- Zhou, Y.; Underhill, S.J.R. Breadfruit (Artocarpus altilis) Gibberellin 2-Oxidase Genes in Stem Elongation and Abiotic Stress Response. Plant Physiology and Biochemistry 2016, 98, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Mitchum, M.G.; Yamaguchi, S.; Hanada, A.; Kuwahara, A.; Yoshioka, Y.; Kato, T.; Tabata, S.; Kamiya, Y.; Sun, T.P. Distinct and Overlapping Roles of Two Gibberellin 3-Oxidases in Arabidopsis Development. Plant Journal 2006, 45, 804–818. [Google Scholar] [CrossRef]
- Hu, J.; Mitchum, M.G.; Barnaby, N.; Ayele, B.T.; Ogawa, M.; Nam, E.; Lai, W.-C.; Hanada, A.; Alonso, J.M.; Ecker, J.R.; et al. Potential Sites of Bioactive Gibberellin Production during Reproductive Growth in Arabidopsis. Plant Cell 2008, 20, 320–336. [Google Scholar] [CrossRef]
- Lo, S.-F.; Yang, S.-Y.; Chen, K.-T.; Hsing, Y.-I.; Zeevaart, J.A.D.; Chen, L.-J.; Yu, S.-M. A Novel Class of Gibberellin 2-Oxidases Control Semidwarfism, Tillering, and Root Development in Rice. Plant Cell 2008, 20, 2603–2618. [Google Scholar] [CrossRef]
- He, H.; Yamamuro, C. Interplays between Auxin and GA Signaling Coordinate Early Fruit Development. Hortic Res 2022, 9. [Google Scholar] [CrossRef]
- Wei, H.; Chen, J.; Zhang, X.; Lu, Z.; Liu, G.; Lian, B.; Yu, C.; Chen, Y.; Zhong, F.; Zhang, J. Characterization, Expression Pattern, and Function Analysis of Gibberellin Oxidases in Salix matsudana. Int J Biol Macromol 2024, 266, 131095. [Google Scholar] [CrossRef]
- Perglova, I.; Pergl, J.; Pysek, P.; Pyšek, P. Reproductive Ecology of Heracleum mantegazzianum. In Ecology and management of giant hogweed; CAB International: Wallingford, 2007; pp. 55–73. ISBN 1845932064. [Google Scholar]
- Koornneef, M.; Van der Veen, J.H. Induction and Analysis of Gibberellin Sensitive Mutants in Arabidopsis thaliana (L.) Heynh. Theoretical and Applied genetics 1980, 58, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Goto, N.; Pharis, R.P. Role of Gibberellins in the Development of Floral Organs of the Gibberellin-Deficient Mutant, Ga1-1, of Arabidopsis thaliana. Canadian Journal of Botany 1999, 77, 944–954. [Google Scholar] [CrossRef]
- Pimenta Lange, M.J.; Knop, N.; Lange, T. Stamen-Derived Bioactive Gibberellin Is Essential for Male Flower Development of Cucurbita maxima L. J Exp Bot 2012, 63, 2681–2691. [Google Scholar] [CrossRef] [PubMed]
- Hirano, K.; Nakajima, M.; Asano, K.; Nishiyama, T.; Sakakibara, H.; Kojima, M.; Katoh, E.; Xiang, H.; Tanahashi, T.; Hasebe, M. The GID1-Mediated Gibberellin Perception Mechanism Is Conserved in the Lycophyte Selaginella moellendorffii but Not in the Bryophyte Physcomitrella patens. Plant Cell 2007, 19, 3058–3079. [Google Scholar] [CrossRef]
- Bell, C.R. Breeding Systems and Floral Biology of the Umbelliferae; or, Evidence for Specialization in Unspecialized Flowers. In The biology and chemistry of the Umbelliferae; Academic Press, 1971; pp. 93–107. [Google Scholar]
- Serrani, J.C.; Carrera, E.; Ruiz-Rivero, O.; Gallego-Giraldo, L.; Peres, L.E.P.; García-Martínez, J.L. Inhibition of Auxin Transport from the Ovary or from the Apical Shoot Induces Parthenocarpic Fruit-Set in Tomato Mediated by Gibberellins. Plant Physiol 2010, 153, 851–862. [Google Scholar] [CrossRef]
- Alabadí, D.; Blázquez, M.A.; Carbonell, J.; Ferrándiz, C.; Pérez-Amador, M.A. Instructive Roles for Hormones in Plant Development. International Journal of Developmental Biology 2009, 53, 1597–1608. [Google Scholar] [CrossRef]
- Olimpieri, I.; Siligato, F.; Caccia, R.; Soressi, G.P.; Mazzucato, A.; Mariotti, L.; Ceccarelli, N. Tomato Fruit Set Driven by Pollination or by the Parthenocarpic Fruit Allele Are Mediated by Transcriptionally Regulated Gibberellin Biosynthesis. Planta 2007, 226, 877–888. [Google Scholar] [CrossRef]
- Serrani, J.C.; Ruiz-Rivero, O.; Fos, M.; García-Martínez, J.L. Auxin-Induced Fruit-Set in Tomato Is Mediated in Part by Gibberellins. Plant Journal 2008, 56, 922–934. [Google Scholar] [CrossRef]
- Cong, L.; Yue, R.; Wang, H.; Liu, J.; Zhai, R.; Yang, J.; Wu, M.; Si, M.; Zhang, H.; Yang, C.; et al. 2,4-D-Induced Parthenocarpy in Pear Is Mediated by Enhancement of GA4 Biosynthesis. Physiol Plant 2018. [CrossRef]
- Gillaspy, G.; Ben-David, H.; Gruissem, W. Fruits: A Developmental Perspective. Plant Cell 1993, 5, 1439–1451. [Google Scholar] [CrossRef] [PubMed]
- McAtee, P.; Karim, S.; Schaffer, R.; David, K. A Dynamic Interplay between Phytohormones Is Required for Fruit Development, Maturation, and Ripening. Front Plant Sci 2013, 4, 1–7. [Google Scholar] [CrossRef]
- Moravcová, L.; Perglová, I.; Pyšek, P.; Jarošík, V.; Pergl, J. Effects of Fruit Position on Fruit Mass and Seed Germination in the Alien Species Heracleum mantegazzianum (Apiaceae) and the Implications for Its Invasion. Acta Oecologica 2005, 28, 1–10. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A New Generation of Protein Database Search Programs; Oxford University Press, 1997; Vol. 25. [Google Scholar]
- Bailey, T.L.; Elkan, C. The Value of Prior Knowledge in Discovering Motifs with MEME. In Proceedings of the Ismb; 1995; Vol. 3, pp. 21–29. [Google Scholar]
- Hu, B.; Jin, J.; Guo, A.-Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An Upgraded Gene Feature Visualization Server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [PubMed]
- Chao, J.-T.; Kong, Y.-Z.; Wang, Q.; Sun, Y.-H.; Gong, D.-P.; Lv, J.; Liu, G.-S. MapGene2Chrom, a Tool to Draw Gene Physical Map Based on Perl and SVG Languages. Yi Chuan 2015, 37, 91–97. [Google Scholar]
- Chou, K.-C.; Shen, H.-B. Plant-MPLoc: A Top-Down Strategy to Augment the Power for Predicting Plant Protein Subcellular Localization. PLoS One 2010, 5, e11335. [Google Scholar] [CrossRef]
- Abramson, J.; Adler, J.; Dunger, J.; Evans, R.; Green, T.; Pritzel, A.; Ronneberger, O.; Willmore, L.; Ballard, A.J.; Bambrick, J.; et al. Accurate Structure Prediction of Biomolecular Interactions with AlphaFold 3. Nature 2024, 630, 493–500. [Google Scholar] [CrossRef]
- Olechnovič, K.; Venclovas, Č. VoroMQA: Assessment of Protein Structure Quality Using Interatomic Contact Areas. Proteins: Structure, Function, and Bioinformatics 2017, 85, 1131–1145. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple Sequence Alignment with High Accuracy and High Throughput. Nucleic Acids Res 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Suleski, M.; Sanderford, M.; Sharma, S.; Tamura, K. MEGA12: Molecular Evolutionary Genetic Analysis Version 12 for Adaptive and Green Computing. Mol Biol Evol 2024, 41, msae263. [Google Scholar] [CrossRef]
- De Boer, T.J.; Backer, H.J. A New Method for the Preparation of Diazomethane. Recueil des Travaux Chimiques des Pays-Bas 1954, 73, 229–234. [Google Scholar] [CrossRef]
- Gaskin, P.; MacMillan, J. GC-MS of the Gibberellins and Related Compounds. Methodology and a library of spectra. University of Bristol (Cantock’s Enterprises), Bristol, England, UK 1991.
- Hasegawa, H.; Shinohara, Y.; Hashimoto, T.; Matsuda, R.; Hayashi, Y. Prediction of Measurement Uncertainty in Isotope Dilution Gas Chromatography/Mass Spectrometry. J Chromatogr A 2006, 1136, 226–230. [Google Scholar] [CrossRef] [PubMed]
- Frisse, A.; Pimenta, M.J.; Lange, T. Expression Studies of Gibberellin Oxidases in Developing Pumpkin Seeds. Plant Physiol 2003, 131, 1220–1227. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Fernald, R.D. Comprehensive Algorithm for Quantitative Real-Time Polymerase Chain Reaction. Journal of computational biology 2005, 12, 1047–1064. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. Past: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontologia electronica 2001, 4, 1. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing. 2021.
Figure 1.
Distribution of HsGAox genes on Heracleum sosnowskyi chromosomes. The scale refers to the lengths of the chromosomes.
Figure 1.
Distribution of HsGAox genes on Heracleum sosnowskyi chromosomes. The scale refers to the lengths of the chromosomes.
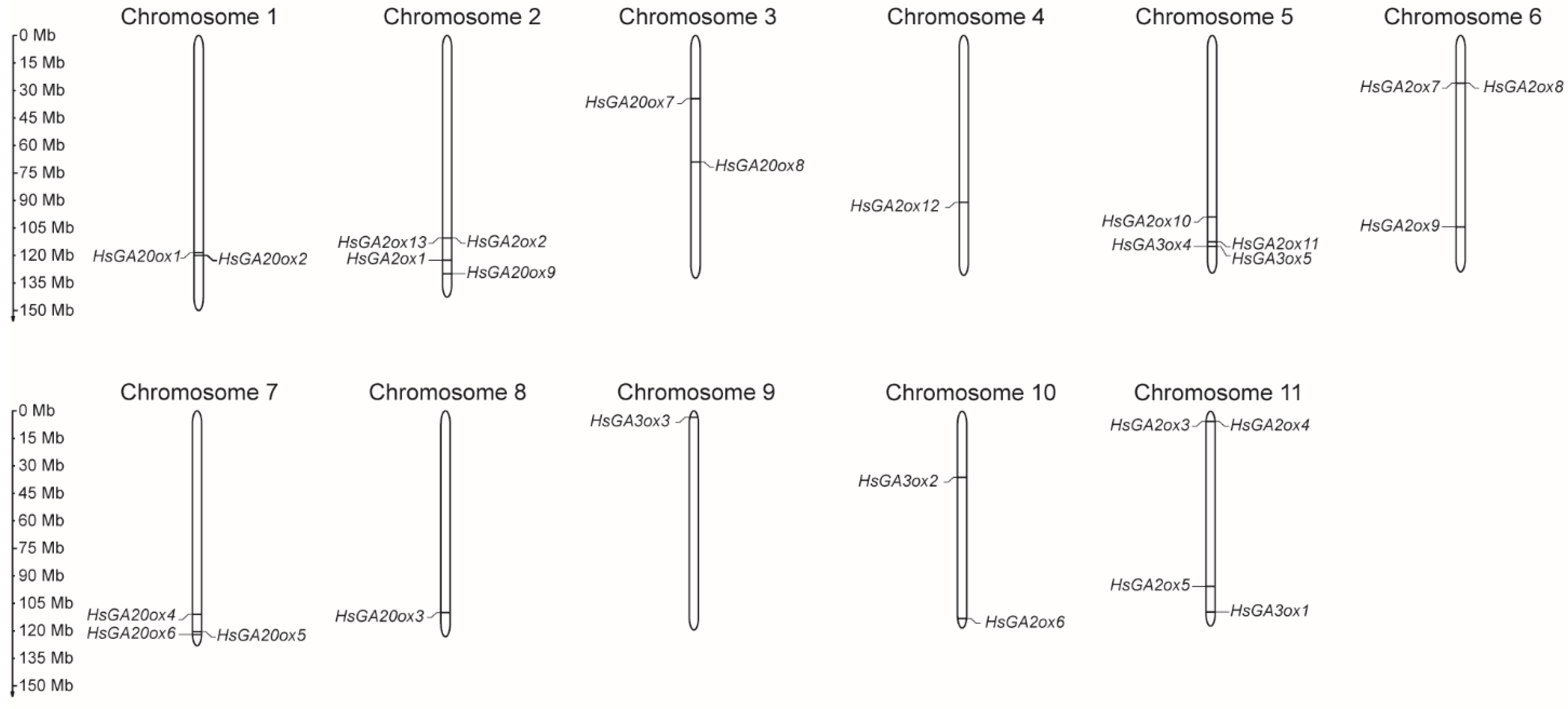
Figure 2.
The phylogenetic relationship analysis of GAox proteins among Heracleum sosnowskyi, Arabidopsis thaliana and Oryza sativa. The tree was constructed according to the ML method using MEGA 12.0 software with 1000 bootstrap replicates. The GAox subfamilies and analysed species were labelled with different colours. Scale bar refers to a phylogenetic distance of amino acid substitutions per site.
Figure 2.
The phylogenetic relationship analysis of GAox proteins among Heracleum sosnowskyi, Arabidopsis thaliana and Oryza sativa. The tree was constructed according to the ML method using MEGA 12.0 software with 1000 bootstrap replicates. The GAox subfamilies and analysed species were labelled with different colours. Scale bar refers to a phylogenetic distance of amino acid substitutions per site.
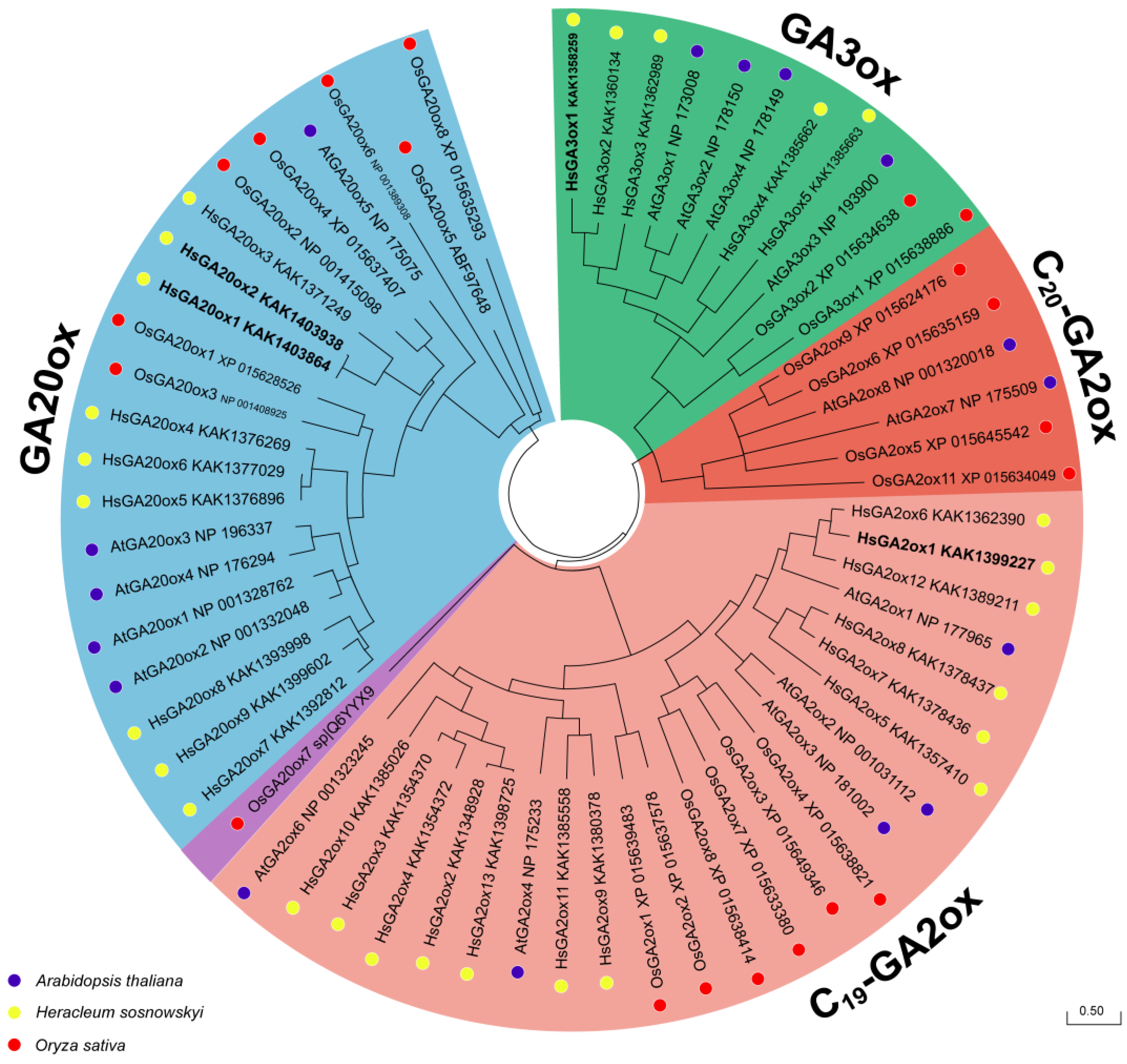
Figure 3.
Representation of conserved protein domains, motifs and gene structure patterns of Heracleum sosnowskyi GAox. (A) Generated models of putative HsGA20ox1, HsGA20ox2, HsGA2ox1 and HsGA3ox1 proteins. Non-haem dioxygenase N-terminal and 2OG-Fe(II) oxygenase domains are highlighted by dark blue and light blue, respectively. (B) Schematic view of HsGAox protein sequences and 10 conserved motifs distributed among them. (C) The gene structure map of HsGAox members.
Figure 3.
Representation of conserved protein domains, motifs and gene structure patterns of Heracleum sosnowskyi GAox. (A) Generated models of putative HsGA20ox1, HsGA20ox2, HsGA2ox1 and HsGA3ox1 proteins. Non-haem dioxygenase N-terminal and 2OG-Fe(II) oxygenase domains are highlighted by dark blue and light blue, respectively. (B) Schematic view of HsGAox protein sequences and 10 conserved motifs distributed among them. (C) The gene structure map of HsGAox members.
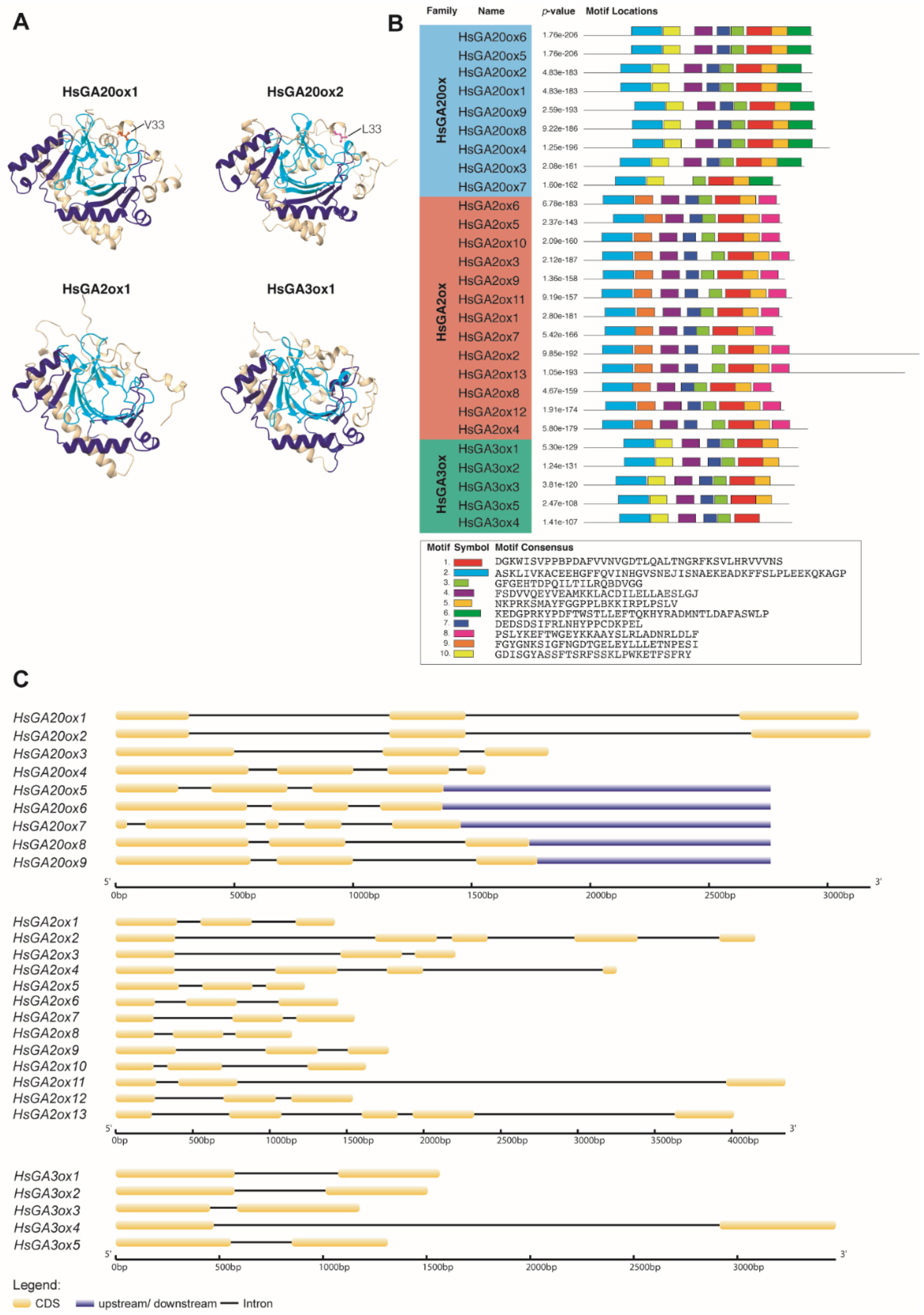
Figure 4.
(A) Schematic picture of Heracleum sosnowskyi terminal inflorescence. The effect of 150 mg/l gibberellic acid (GA3) on phenotype (B), morphometry (C), endogenous gibberellin (GA) content (D) and GA biosynthesis gene expression (E) in H. sosnowskyi ovary tissues from the central and lateral parts of terminal inflorescence. DAA – days after application GA3. Error bars represent the standard error of the mean. Asterisks in the same colour columns indicate statistically significant differences (* – p < 0.01; ** – p < 0.001). Scale bar represents 1 mm.
Figure 4.
(A) Schematic picture of Heracleum sosnowskyi terminal inflorescence. The effect of 150 mg/l gibberellic acid (GA3) on phenotype (B), morphometry (C), endogenous gibberellin (GA) content (D) and GA biosynthesis gene expression (E) in H. sosnowskyi ovary tissues from the central and lateral parts of terminal inflorescence. DAA – days after application GA3. Error bars represent the standard error of the mean. Asterisks in the same colour columns indicate statistically significant differences (* – p < 0.01; ** – p < 0.001). Scale bar represents 1 mm.
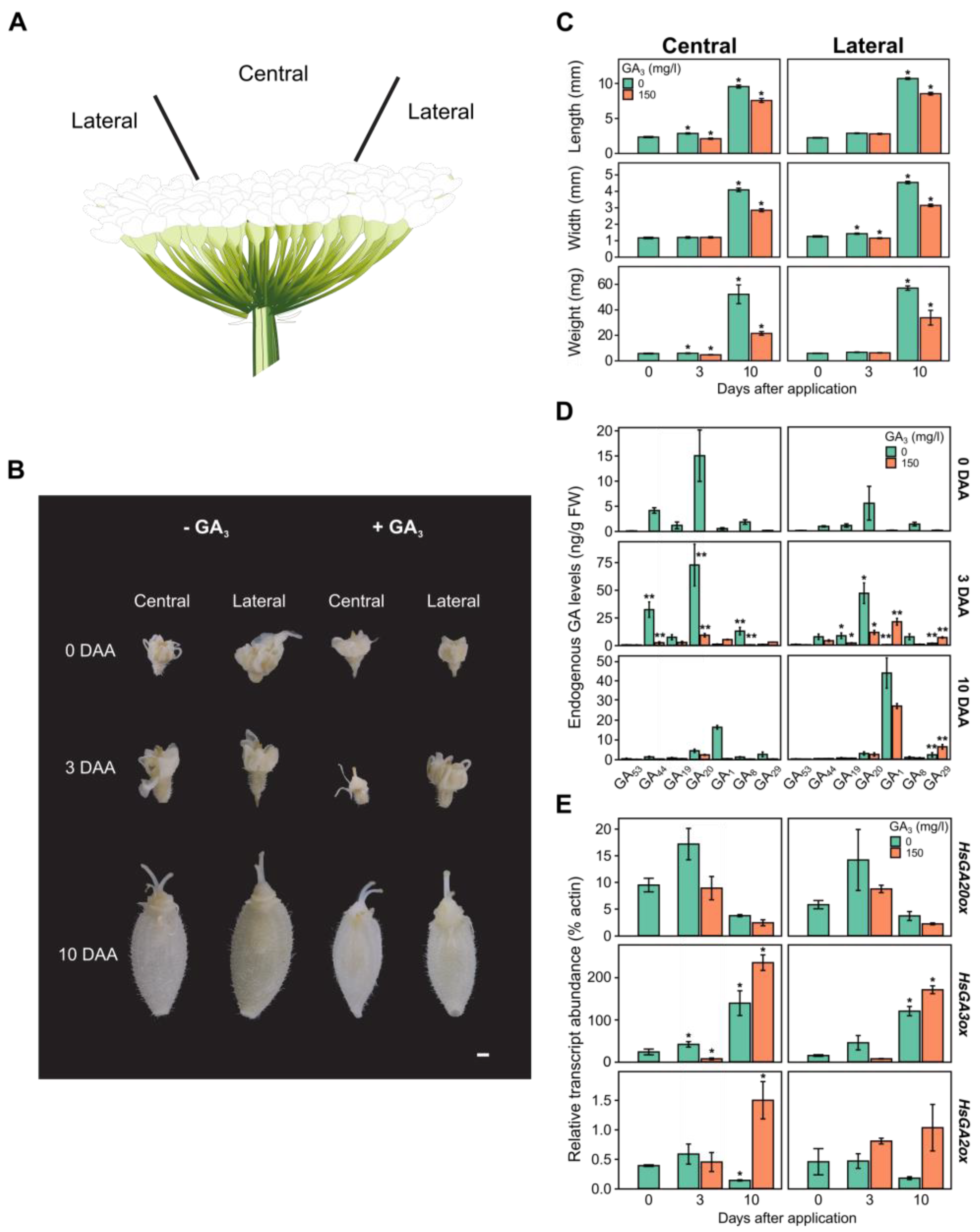

Table 1.
Characterization of the HsGAox superfamily in Heracleum sosnowskyi.
| Protein Name | Accession no. | Gene Name | CDS (nt) | Length (aa) | MW (kDa) | Domain Location | Type | Subcellular Localization |
|---|---|---|---|---|---|---|---|---|
| HsGA20ox1 | KAK1403938 | HsGA20ox1 | 1134 | 377 | 42.88 | 44-144a 209-306b |
GA20ox | Cytoplasm |
| HsGA20ox2 | KAK1403864 | HsGA20ox2 | 1134 | 377 | 42.89 | 44-144a 209-306b |
GA20ox | Cytoplasm |
| HsGA20ox3 | KAK1371249 | HsGA20ox3 | 1095 | 364 | 41.33 | 43-144a 209-306b |
GA20ox | Cytoplasm |
| HsGA20ox4 | KAK1376269 | HsGA20ox4 | 1221 | 406 | 46.14 | 63-169a 227-325b |
GA20ox | Cytoplasm |
| HsGA20ox5 | KAK1376896 | HsGA20ox5 | 1140 | 379 | 42.96 | 61-167a 225-323b |
GA20ox | Cytoplasm |
| HsGA20ox6 | KAK1377029 | HsGA20ox6 | 1140 | 379 | 43.02 | 61-167a 225-323b |
GA20ox | Cytoplasm |
| HsGA20ox7 | KAK1392812 | HsGA20ox7 | 978 | 325 | 36.81 | 62-164a 227-325b |
GA20ox | Cytoplasm |
| HsGA20ox8 | KAK1393998 | HsGA20ox8 | 1152 | 383 | 43.73 | 34-135a 181-260b |
GA20ox | Cytoplasm |
| HsGA20ox9 | KAK1399602 | HsGA20ox9 | 1149 | 382 | 43.38 | 66-170a 230-328b |
GA20ox | Cytoplasm |
| HsGA2ox1 | KAK1399227 | HsGA2ox1 | 987 | 328 | 36.87 | 26-115a 176-273b |
C19-GA2ox | Cytoplasm |
| HsGA2ox2 | KAK1348928 | HsGA2ox2 | 1665 | 554 | 61.92 | 20-79a 172-291b |
C19-GA2ox | Cytoplasm Nucleus |
| HsGA2ox3 | KAK1354370 | HsGA2ox3 | 1047 | 348 | 38.86 | 20-88a 173-291b |
C19-GA2ox | Cytoplasm |
| HsGA2ox4 | KAK1354372 | HsGA2ox4 | 1113 | 370 | 41.05 | 20-88a 173-292b |
C19-GA2ox | Cytoplasm |
| HsGA2ox5 | KAK1357410 | HsGA2ox5 | 987 | 328 | 36.77 | 40-96a 180-275b |
C19-GA2ox | Cytoplasm |
| HsGA2ox6 | KAK1362390 | HsGA2ox6 | 975 | 342 | 36.31 | 23-110a 173-269b |
C19-GA2ox | Cytoplasm |
| HsGA2ox7 | KAK1378436 | HsGA2ox7 | 954 | 317 | 35.81 | 26-111a 170-265b |
C19-GA2ox | Cytoplasm |
| HsGA2ox8 | KAK1378437 | HsGA2ox8 | 945 | 314 | 35.39 | 22-123a 166-261b |
C19-GA2ox | Cytoplasm |
| HsGA2ox9 | KAK1380378 | HsGA2ox9 | 999 | 332 | 37.21 | 21-80a 174-275b |
C19-GA2ox | Cytoplasm |
| HsGA2ox10 | KAK1385026 | HsGA2ox10 | 981 | 326 | 36.65 | 19-88a 169-276b |
C19-GA2ox | Cytoplasm |
| HsGA2ox11 | KAK1385558 | HsGA2ox11 | 1035 | 344 | 38.19 | 19-84a 172-288b |
C19-GA2ox | Cytoplasm |
| HsGA2ox12 | KAK1389211 | HsGA2ox12 | 996 | 331 | 37.05 | 27-118a 177-276b |
C19-GA2ox | Cytoplasm |
| HsGA2ox13 | KAK1398725 | HsGA2ox13 | 1593 | 530 | 59.09 | 20-79a 172-291b |
C19-GA2ox | Cytoplasm |
| HsGA3ox1 | KAK1358259 | HsGA3ox1 | 1065 | 354 | 39.98 | 56-158a 211-308b |
GA3ox | Cytoplasm |
| HsGA3ox2 | KAK1360134 | HsGA3ox2 | 1068 | 355 | 40.02 | 57-159a 210-309b |
GA3ox | Cytoplasm |
| HsGA3ox3 | KAK1362989 | HsGA3ox3 | 1047 | 348 | 39.05 | 46-145a 196-295b |
GA3ox | Cytoplasm |
| HsGA3ox4 | KAK1385662 | HsGA3ox4 | 1035 | 344 | 38.59 | 48-141a 201-297b |
GA3ox | Cytoplasm |
| HsGA3ox5 | KAK1385663 | HsGA3ox5 | 1020 | 339 | 38.00 | 50-152a 206-302b |
GA3ox | Cytoplasm |
aa: amino acid; CDS: coding sequence; MW: the theoretical molecular weight of proteins; nt: nucleotide;a non-haem dioxygenase N-terminal domain;b 2OG-Fe(II) oxygenase domain.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



