
Submitted:
14 April 2025
Posted:
15 April 2025
You are already at the latest version
Abstract
Heparan-sulfate (HS) and chondroitin-sulfate (CS) proteoglycans (PGs) are essential regulators of many biological processes including cell differentiation, signalisation and proliferation. PGs interact, mainly via their glycosaminoglycan (GAG) chains, with a large number of ligands including growth factors, enzymes and extracellular matrix components, thereby modulating their biological activities. HSPGs and CSPGs share a common tetrasaccharide linker region, which undergoes modifications particularly phosphorylation of the xylose residue by the kinase Fam20B. Here, we demonstrated that Fam20B gain-of-function decreased, in a dose dependent manner, the synthesis of both CS- and HS-attached PGs. In addition, we showed that blockage of GAG chain synthesis by Fam20B was suppressed by mutation of aspartic acid residues D289 and D309 of the catalytic domain. Interestingly, we bring evidence that, in contrast to Fam20B, expression of the 2-phosphoxylose phosphatase XYLP increased, in a dose dependent manner, GAG chain synthesis and rescue the blockage of GAG chains synthesis induced by Fam20B. In line with previous reports, we found that Fam20B loss-of-function reduced GAG chain synthesis. Finaly, we found that Fam20B inhibits proliferation and migration of glioblastoma cells, thus revealing the critical role of GAG chains of PGs in glioblastoma cell tumorigenesis. This study revealed that both gain- and loss-of-function of Fam20B led to decreased GAG chain synthesis, therefore suggesting that a balance between phosphorylation and dephosphorylation of the xylose by Fam20B and XYLP, respectively is probably an essential factor of the regulation of the rate of PG synthesis.
Keywords:
Fam20B
; proteoglycans
; glycosaminoglycan synthesis
; glioblastoma
1. Introduction
Proteoglycans (PGs) are complex macromolecules that are found on cell surface and in the extracellular matrix [1]. They are formed by the covalent attachment of one or more glycosaminoglycan (GAG) chains onto the core protein. PG-GAG chains interact with a large number of ligands including growth factors and morphogens and their receptors as well as extracellular matrix structural molecules where they regulate activity and ligand stability [2,3]. PGs regulate several biological processes including cell-cell and cell-matrix interactions, development, signaling, proliferation, differentiation and various pathological conditions such as inflammation, tumor progression and viral and bacterial infection, [4,5,6,7]. Most commonly occurring PGs are chondroitin-sulphate PGs (CSPGs) and heparan-sulphate PGs (HSPGs). These PGs share a common tetrasaccharide linker region (GlcAβ1-3Galβ1-3Galβ1-4Xyl-O-Ser) for the attachment of the GAG chain onto the PG core protein. The first step in the synthesis of the tetrasaccharide primer is the addition of xylose on specific serine residues of the core protein. This initial and limiting step in GAG synthesis is catalyzed by xylosyltransferase I and II [8,9,10]. Subsequently, two galactose residues are attached to xylose by β1,4-Galactosyltransferase 7 and β1,3-Galactosyltransferase 6 (β4GalT7 and β3GalT6), and one glucuronic acid by β1,3-Glucuronosyltransferase (GlcAT-I), therefore completing the synthesis of the terasaccharide primer. HS- and CS-GAG chains of PGs are built up on this linkage tetrasaccharide region by the alternate addition of N-acetylhexosamine and glucuronic acid residues [11]. The specificity of interaction between ligands and GAG chains is influenced by the fine structure of the GAG chain, primarily its sulfation pattern [12]. The tetrasaccharide primer also undergoes various modifications including sulfation of galactose residues and phosphorylation of xylose [13,14].
Fam20 (family of sequence similarity 20) contains three members Fam20A, Fam20B and Fam20C [15]. Their function has recently been established [16,17]. The substrates for Fam20A are unknown; however, Fam20A is important for the enamel biomineralization and tooth eruption [17,18]. Mutations in Fam20A are associated with various tooth disorders named human amelogenesis imperfecta and gingival hyperplasia syndrome [19], and in some cases the renal calcification are also involved along with tooth disorders such as in Enamel Renal Syndrome [18,20]. Fam20C is a Golgi casein kinase and is mainly expressed in the biomineralized tissues. Fam20C phosphorylates casein and the Small Integrin-Binding Ligand, N-linked Glycoproteins (SIBLINGs) which are critical for biomineralization. Mutations in Fam20C are associated with Raine syndrome, characterized by a lethal osteosclerotic bone dysplasia [16,21,22]. Recently, Fam20C from the C. elegans (ceFam20C) have been crystallized [23]. The 3D structure revealed the presence of protein kinase-like fold with five disulfide and two Asparagine residues for N-linked glycosylation. The ceFam20C also contains a DFG (Asp-Phe-Gly) variant motif, in which the aspartate residue Asp387 co-ordinates the divalent cation required for catalysis and a catalytic segment that includes the putative catalytic aspartate residue, Asp366. The Asp387 and Asp366 residues are conserved in all the Fam20 family [23].
Fam20B is a kinase that phosphorylates the xylose in the tetrasaccharide linkage region of PGs [24,25]. Genetic studies showed that loss-of-function mutations in Fam20B in zebra fish decreases the amount of cellular GAG chains and cause cartilage and skeleton defects [26]. Fam20B knockout in mice results in embryonic lethality and embryos showed multisystem organ hypoplasia and delayed development [17]. On the other hand, it has been shown that gain-of-function of XYLP, the 2-phosphoxylose phosphatase that de-phosphorylates xylose in the tetrasaccharide linker region, increases the amounts of GAGs in Hela cells and inversely, loss-of-function of XYLP decreases GAGs synthesis [27].
The aim of the present study was to investigate the potential regulatory function of FAM20B in the synthesis of PG. Here, we demonstrate that Fam20B gain-of-function inhibits the synthesis of PGs and showed that both CS/DS- and HS-PGs are affected. Interestingly, we found that Fam20B inhibition of PG synthesis is dose-dependent and is rescued by the phosphatase XYLP. Furthermore, based on the crystal structure of ceFam20C, we showed that aspartic acid residues D289 and D309 are essentials for Fam20B suppression of the synthesis of CS and HS-attached PGs.
2. Materials and Methods
2.1. Cell Lines and Culture Conditions
The Chinese hamster ovary cells (CHO-K1), CHO galactosyltransferase I deficient cell line pgsB-618, human embryonic kidney cells HEK293, human brain tumor T98G, human lung adenocarcinoma cells A549 and human primary skin fibroblasts were cultured in DMEM (4.5 mg/ml glucose) or DMEM-F12 medium supplemented with 2 mM glutamin, 100 IU/ml penicillin, 100 µg/ml streptomycin and 10% fetal bovine serum (FBS, Dutscher Brumath, France) at 37°C with humidified atmosphere in a 5% CO2.
2.2. Vector Constructions and Cell Transfection
Fam20B was amplified from human placenta cDNA library (Takara Bio) by PCR using5’GAATTCCACCATGAAGCTAAAGCAGCGAGTCGTG3’ (forward) including EcoRI site and 5’GGATCCTTACAAGTGTGAGAGAGCCATCCT3’ (reverse) primers including BamHI site, using Advantage GC 2 Polymerase Mix (Takara Bio). XYLP was cloned by PCR using 5’GAATTCCACCATGCT TTTCCGCAACCGCTTC 3’ (forward) including EcoRI site and 5’CTCGAGTTAGAATCCTTCCCTGTGACATGC 3’ (reverse) primers including XhoI site. The amplified Fam20B and XYLP products were ligated into pCR2.1-TOPO vector (Invitrogen). Flag-XYLP and HA-XYLP cDNAs were generated by PCR. The coding region for Fam20B was excised and ligated into the pCMV vector (Stratagene, Valencia, CA) by double digestion with EcoRI and BamHI to generate pCMV-Fam20B. The coding region for Flag-XYLP and HA-XYLP were excised and ligated into the pCMV vector by double digestion with EcoRI and XhoI to generate pCMV-Flag-XYLP and pCMV-HA-XYLP. Decorin and HA-syndecan 4 cDNAs were generated by PCR and cloned into EcoRI and BamHI or EcoRI and XhoI sites of pCMV empty vector to generate pCMV-DCN and pCMV-HA-SDC4, respectively.
For transfection, cells were seeded in 6-well culture plate until 80% confluency and transfected with 250 ng of either pCMV-DCN or pCMV-HA-SDC4 in combination with 1 µg pCMV-Fam20B, pCMV-HA-XYLP, pCMV-Flag-XYLP, pCMV-Fam20BD289A, pCMV-Fam20BD309A or pCMV-empty vector using lipofectamine 2000 transfection reagent (Invitrogen, Carlsbad, CA) according to manufacturer’s instructions. Expression of decorin and of syndecan 4 was analyzed at 48 h post-transfection by Western blotting.
2.3. Fam20B Knockdown
Sense 5’CACCGTATAGCCGAGACCATGTGG3’ and antisense 5’AA ACCCACA TGGTCTCGGCTATAC3’ oligonucleotides containing 20 bp sequence (underlined) targeting Fam20B exon 3 and cohesive ends (bold) with the vector were annealed and ligated into BbsI sites of pUC57-attbU6 sgRNA vector which express gRNA using avian-derived U6 promoter. Cells were transfected with empty pUc57-attbU6 (control), pUc57 attbU6/Fam20BgRNA in association with pspCas9 plasmid expressing Cas9. To facilitate screening, cells were co-transfected with pSVneo plasmid which express the neomycin resistance. Positive cells were screened using resistance to neomycin and gene mutation was determined by sequence analysis. The Fam20B genomic region targeted was amplified by PCR using specific primers. Sequencing results indicated that FAM20B gene harboured frameshift mutation mediated by deletion of two nucleotides at the sites targeted by gRNA, creating premature stop codon that prevents the translation of full length FAM20B. All analyzed sequences from the transfected cell clones exhibited the same deletions in FAM20B genomic region.
2.4. Site-Directed Mutagenesis
Site directed mutagenesis of the residues Asp289 to Ala289 and Asp309 to Ala309 in Fam20B were performed using the QuikChange XLII (Agilent, CA, USA) according to the recommendations of the manufacturer. pCMV-Fam20B expression vector was used as template. Sense and antisense oligonucleotides introducing the desired mutations were for D289A: 5’CTGATTGGCAATGCTGCCCGCATCACTAT GAG3’ (forward) and 5’CTCATAGTGATGGCGGGCAGCATTGCCAATCAG3’ (reverse) and for D309A: 5’TGCTCATCCTTCTTGCTAATGCCAAAAGCTTTGG3’ (forward) and 5’C CAAAGCTTTTGGCATTAGCAAGAAGGATGAGC A3’ (reverse). Full length mutated cDNA Fam20BD289A and Fam20BD309A were checked by DNA sequencing.
2.5. N-Glycosylation Analysis
Cells were grown in 6-well culture plate at 80% confluency then transfected with pCMV-Fam20B expression vector. At 24 h post-transfection the cells were lysed in the HEPES buffer and protein concentration was measured by the Bradford method [28]. Twenty µg of protein was digested with PNGase F (New England Biolabs), which cleaves asparagine-linked (N-linked) oligosaccharides, according to the manufacturer’s instructions.
2.6. Western Blotting
Total protein from cells was extracted using RIPA buffer (150 mM NaCl, 50 mM Tris-HCl, pH 7.5, 1% deoxycholate, 0.1% SDS, 1% Triton X-100) supplemented with protease and phosphatase inhibitors (Roche Diagnostics, Indianapolis, IN, USA). Cell lysates were sonicated on ice and protein concentration of the samples was determined by the Bradford method. Proteins from culture medium and from cell lysates (50 μg/lane) were separated on 10% SDS-PAGE gels, transferred to a PVDF membrane (Bio-RAD), and subsequently blocked in PBS-Tween 20 containing 5% nonfat milk or 5% BSA. Membranes were then incubated overnight with primary antibodies directed against Fam20B (Cat# HPA 007409, 1/1000, Sigma-Aldrich), decorin (Cat# MAB143, 1:1000, R&D Systems), HA (Cat# 901501, 1:10000, BioLegend), M2-Flag (Cat# F1804, Sigma-Aldrich) or βactin (Cat#3700, 1/2000, CST) followed by incubation with horseradish peroxidase-conjugated secondary antibodies (Cat# 7074, 1:2000, CST or Cat# 7076, 1:2000 CST). Antibodies were diluted in 5% BSA/0.01% tween 20 in PBS. The blots were then developed using Clarity Western ECL substrate (BIO-RAD, Hercules, CA) according to the instructions of the manufacturer.
2.7. Metabolic Labelling of GAG Chains
Metabolic labelling of PG-GAG chains was carried out using 35S-sulfate incorporation method as described by De Vries et al, (1986) [29]. Briefly, cells were grown in 6-well culture plate and transfected with pCMV-Fam20B, pCMV-Fam20BD289A, pCMV-Fam20BD309A, or pCMV empty vector. Cells were then radiolabeled with 10 µCi/ml of 35S-sulfate (Perkin Elmer) in sulfate free media containing 2% dialyzed FBS. After overnight incubation, the culture medium was collected, digested with papain (1 mg/ml) and 35S-labeled GAGs were precipitated by cetylpyridinium chloride (CPC) as described by Bronson et al., (1987) [30]. The CPC precipitated radiolabeled GAGs were separated by SDS-PAGE on a 4-12% Nu-PAGE gel. The gel was dried and exposed to autoradiography film. To measure the rate of sulfate incorporation into GAG chains of PGs, HEK293 transfected with pCMV empty vector or pCMV-Fam20B were radiolabeled with 10 µCi/ml of [35S]-sulfate for 6 h then, conditioned culture medium was collected and digested with papain (1mg/ml). [35S]-labeled GAG chains were precipitated by CPC dissolved in solvable and mixed in scintillation fluid (Perkin Elmer, MA, USA). The radioactivity associated with GAGs was measured by liquid scintillation counting (Packard, Rungis, France).
2.8. Immunofluorescence Analysis
The CHO-K1 cells were grown on glass coverslips and transfected with Fam20B expression vector pCMV-Fam20B or with pCMV empty vector (control). At 36 h after transfection, cells were fixed with 3% (w/v) paraformaldehyde in PBS for 20 min and were permeabilized by treatment with 0.1% (w/v) Triton X-100/PBS solution for 4 min. After extensive washing in 0.2% (w/v) fish skin gelatin in PBS, cells were then incubated with primary antibodies anti-Fam20B (Cat# HPA 007409, 1/100 Sigma-Aldrich) and anti-HS (Cat# 370255-1, 1:100, AMSBIO) for 20 min. Cells were washed several times in 0.2% (w/v) fish skin gelatin in PBS and incubated with secondary antibodies coupled with Alexa Fluor 555 and Alexa Fluor 488 (Cat# A-21428 or Cat# A-11017, Molecular Probes) for 20 min. Cells were washed with PBS and nuclei were stained with Hoechst/PBS solutions then coverslips were mounted with Moviol (National Diagnostics, U.K.) containing 1% propylgallate (Sigma-Aldrich). Digital images were captured with an inverted microscope Lieca DMI3000 B (Leica Microsystems, Germany).
2.9. Data Analysis and Statistical Procedures
Each experiment was repeated at least three times independently. Quantitative data were expressed as mean ± S.D. Statistical analysis was performed with an unpaired two-tailed Student's t-test, and effects were considered statistically significant at *P<0.05. One representative immunoblot of three independent experiments was shown in results.
3. Results
3.1. Fam20B Gain-of-Function Reduces the Synthesis PG-GAG Chains
It has been shown that loss of Fam20B impairs the synthesis of PGs leading to premature termination of PG-GAG chain elongation [25]. To gain more insight in the role of Fam20B in the synthesis of PG-GAG chains, we sought to determine whether Fam20B gain-of-function affects the synthesis of PG-GAG chains. To this end, we expressed Fam20B in HEK293 cells and analyzed the impact on the synthesis of PGs. Western-blot analysis showed that cells transfected with the expression vector pCMV-Fam20B efficiently expressed a polypeptide of about 46 kDa corresponding to Fam20B protein, which was absent in cells transfected with pCMV empty vector (control) (Figure 1A). Fam20B contains potential N-glycosylation sites. To test whether Fam20B is N-glycosylated, proteins from HEK293 cells expressing Fam20B were treated with the enzyme N-glycosidase F (PNGase) and analyzed by Western blot. Treatment with PNGase produced a decrease of about 2 kDa in the apparent molecular mass of Fam20B polypeptide (Figure 1B), indicating that Fam20B is sensitive to PNGase and therefore is N-glycosylated. We next analyzed the effect of Fam20B gain-of-function on the synthesis of GAG chains using metabolic incorporation of radiolabelled [35S]-sulfate into GAG chains of PGs. Interestingly, the level of PG synthesis was decreased by 75% in cells transfected with Fam20B expression vector compared to control cells transfected with empty vector (Figure 1C). This was confirmed by SDS-PAGE analysis of radiollabeled GAG chains of PGs which shows strong reduction (90%) in overall GAG chains produced in Fam20B expressing cells compared to control (Figure 1D,E). These results indicated that Fam20B gain-of-function strongly reduced the synthesis of PG-GAG chains.
3.2. Fam20B Gain-of-Function Reduces the Synthesis of Both CS- and HS-GAG Chains
Given the fact that Fam20B phophorylates the xylose residue of the tetrasaccharide linker common to both CS and HS, we hypothesized that Fam20B may impact the synthesis of both HSPGs and CSPGs. To examine this hypothesis, we used decorin and syndecan 4 as reporter for the synthesis of CSPGs and HSPGs, respectively. HEK293 cells were transfected with pCMV-Fam20B expression vector or pCMV empty vector (control) together with either pCMV-DCN vector expressing decorin or pCMV-HA-SDC4 vector expressing HA-syndecan 4 with an N-terminal tagged HA epitope. The expression of decorin and syndecan 4 was analyzed by Western blot. Transfection of HEK293 cells with pCMV-DCN vectors resulted, as expected, in the secretion in the medium of high amount of CS/DS-attached decorin that migrate as an elongated smear during gel electrophoresis due to heterogeneity of attached GAG chains, however when decorin was co-expressed with Fam20B, the amount of CS/DS-attached decorin in the medium was strongly reduced (Figure 2A). To further confirm that Fam20B reduces the synthesis of GAG-attached decorin, we took advantage of CHO pgs-B618 cells deficient for β4GalT7 enzyme. These cells initiate GAG synthesis by catalyzing the transfer of xylose residues on the PG core protein, but they are not able to catalyse the second step which consist in the addition of galactose on the xylose residue due to lack of β4GalT7, therefore precluding the formation of completed tetrasaccharide primer and subsequent elongation of PG-GAG chain. Expression of decorin in CHO pgs-B618 cells resulted in the synthesis in the culture medium of high amount of GAG chain-free decorin (Figure 2B). As expected, co-expression of decorin with β4GalT7 restored the synthesis of GAG-attached decorin. However, when decorin and β4GalT7 were expressed along with Fam20B the synthesis of GAG-attached decorin was dramatically reduced (Figure 2B), indicating that Fam20B gain-of-function blocks GAG chains formation.
Next, we examine whether the effect of Fam20B on the synthesis of GAG-attached decorin is dose-dependent. To this end, HEK293 cells were transfected with decorin expression vector along with increasing concentrations of Fam20B expression vector. Remarkably, as shown in Figure 2C the synthesis of GAG-attached decorin decreases with increased expression of Fam20B. Indeed, the amount of decorin expressed dropped by 50%, 70% and 95% when cells were transfected with 100 ng, 250 ng and 500 ng of Fam20B expression vector, respectively (Figure 2C,D). This data clearly indicated that inhibition of the synthesis of GAG-attached decorin by Fam20B is dose-dependent. Interestingly, analysis of intracellular decorin in cell lysates of control and Fam20B expressing cells clearly revealed the presence of high amount of GAG chain-free decorin inside the cells expressing Fam20B which increases with increasing expression of Fam20B (Figure 2E). The amount of intracellular GAG chain-free decorin was increased by about 10%, 30% and 40% in cells transfected with 100 ng, 250 ng and 500 ng of Fam20B expression vector, respectively compared to control (Figure 2E,F). These results indicate that Fam20B gain-of-function led to accumulation of GAG chain-free decorin inside the cells which is not prone for secretion due probably to the blockage in the elongation of CS/DS GAG chain. To determine whether this process is cell type specific, we studied the effect of Fam20B expression on the synthesis of decorin in human primary skin fibroblasts and lung fibroblast cell line A549 which naturally produce and secrete high amount of CS/DS-attached decorin. As expected, analysis of decorin from conditioned medium of human primary skin fibroblast cells showed high amount of CS/DS-attached decorin, as revealed by immunobloting using anti-decorin antibodies (Figure 2G), however transfection with Fam20B expression vector led to a strong decrease (90%) in CS/DS-attached decorin in the medium (Figure 2G,H). Similar results were obtained in lung fibroblast cells A549. Transfection with Fam20B expression vector reduced the amount of GAG-attached decorin secreted in the medium by about 80%, compared to control (Figure 2I,J). Altogether, these results clearly showed that the effect of Fam20B on the synthesis of CS/DS-attached decorin is dose-dependent and is not cell-type specific.
We next tested whether Fam20B produced similar inhibitory effect on the synthesis of HS-attached PGs, as observed for CSPG. For these purposes, we used syndecan 4 as HSPG reporter. HEK293 cells were transfected with HA-syndecan 4 expression vector and pCMV-empty vector (control) or with HA-syndecan 4 and Fam20B expression vectors. Transfection of cells with HA-syndecan 4 vector resulted in high expression of HS-attached syndecan 4 that migrates as a smear in SDS-PAGE (Figure 3A). Interestingly, co-expression with Fam20B led to remarkable reduction (80%) in the synthesis of HS-attached syndecan 4 (Figure 3A,B), indicating that Fam20B gain-of-function dramatically decreases the synthesis of HS-attached syndecan 4. To determine whether reduction in the synthesis of HS-attached syndecan 4 is Fam20B dose-dependent, HEK293 cells were transfected with HA-syndecan 4 expression vector along with increasing concentrations of Fam20B expression vector. The results showed that the amount of HS-attached syndecan 4 dropped by about 20%, 50% and 80% when cells were transfected with 100 ng, 250 ng and 500 ng of Fam20B expression vector, respectively (Figure 3C,D). In contrast, the amount of HS-free syndecan 4 in the cells was increasing by about 10% to 25% with increased expression of Fam20B (Figure 3C,E). Altogether, these results demonstrated that gain-of-function of Fam20B reduced the synthesis of PGs of both type, CS and HS, and led to accumulation of GAG-free PG core protein inside the cells.
Cell surface HS-attached PGs are important for many vital cell-signaling processes [4,5,6]. To examine the effect of Fam20B gain-of-function on cell surface HSPGs, we carried out indirect immunofluorescence analysis of cell surface HSPGs in CHO-K1 cells transfected with Fam20B expression vector or with pCMV vector (control), using anti-HS monoclonal antibody 10E4, which is commonly used to detect HS chains of PGs [31,32,33]. Prominent staining of the cell membrane HSPGs was observed in CHO-K1 cells transfected with empty vector (control), whereas no or low HSPG staining could be observed in cell membrane of cells transfected with Fam20B expression vector (Figure 3F). When recombinant cells were probed with anti-Fam20B antibodies, efficient expression of Fam20B was revealed, whereas cells transfected with pCMV (control) showed no staining (Figure 3F). Noteworthy, when cells express Fam20B, no or very low staining for cell surface HSPG was observed (Figure 3F, Merge), indicating that Fam20B gain-of-function impaired the synthesis of cell surface HS-GAG chains.
3.3. XYLP Rescues the Synthesis of GAG-Attached PGs Blockage Induced by Fam20B
It has been shown that XYLP is the 2-phosphoxylose phosphatase that de-phosphorylates xylose in the tetrasaccharide linker region of PGs [27]. We hypothesize that Fam20B gain-of-function may result in sustained phosphorylation of xylose residue which may lead to blockage in the elongation of CS and HS GAG chains, therefore expression of XYLP may rescue the elongation process. We first analyzed the effect of XYLP on the synthesis of decorin in HEK293 cells. Cells were transfected with pCMV-DCN and empty pCMV (control) vector or with pCMV-DCN and increased concentrations of pCMV-XYLP expression vector. As shown in Figure 4A, expression of XYLP increased, in a dose-dependent manner, the amount of GAG-attached decorin. Indeed, the amount of CS/DS free decorin present in the medium was gradually decreasing as the concentration of XYLP increases (Figure 4A). This strongly suggest that XYLP gain-of-function enhanced the synthesis of GAG-attached decorin.
To determine whether expression of XYLP may rescue Fam20B-induced inhibition of GAG-attached decorin, HEK293 cells were transfected with decorin and Fam20B expression vector or with decorin and Fam20B along with XYLP expression vector. As expected, GAG-attached decorin is efficiently produced in cells transfected with decorin expression vector, whereas its expression was reduced in cells expressing Fam20B (Figure 4B). Remarkably, expression of XYLP rescued the synthesis of GAG-attached decorin leading to the synthesis of high molecular mass CS/DS attached decorin, therefore overcoming the inhibitory effect of Fam20B on the synthesis of CS/DS attached decorin (Figure 4B). Similar results were obtained for syndecan 4. Transfection of HEK293 cells with HA-syndecan 4 expression vector resulted in the synthesis of high amount of GAG-attached syndecan 4, however co-expression with Fam20B dramatically reduced the synthesis of HS-attached syndecan 4 (Figure 4C). Interestingly, expression of XYLP rescued Fam20B-induced inhibition of the synthesis of HS-attached syndecan 4 (Figure 4C). Altogether, these results demonstrate that Fam20B gain-of-function blocks the synthesis of GAG-attached PGs that can be rescued by XYLP.
3.4. Aspartic Acid Residues in Catalytic Domain and DFG Motif Are Essential for Fam20B Activity
Based on the structure/function studies and 3D structure analysis of C. elegans Fam20C (ceFam20C), it has been shown that the aspartic residue of the 387DHG389 sequence, a variant of the DFG motif, present in canonical protein kinases which co-ordinates Mn2+ cation and the putative catalytic aspartic residue present in the catalytic segment 366DRHHYE371 are essential for ceFam20C activity [23]. The corresponding aspartic residues are present in the human Fam20B and are Asp309 and Asp289, respectively. We therefore explored the effect of the mutation of these residues on human Fam20B activity. To this end, Asp309 to alanine (D309A) and Asp289 to alanine (D289A) mutants of Fam20B were engineered and expressed in CHO-K1 cells. Western blot analysis showed that Fam20BD309A and Fam20BD289A mutants were expressed in a similar amount as wild-type Fam20B protein (Figure 5A). To determine whether the D309A and D289A mutations impair Fam20B activity, decorin and syndecan 4 were co-expressed with either wild-type Fam20B or with the mutants Fam20BD309A and Fam20BD289A, respectively and GAG-attached decorin and syndecan 4 were analyzed by immunoblot. In contrast to wild-type Fam20B, co-expression with the mutants Fam20BD309A and Fam20BD289A did not suppress the synthesis of either CS/DS-attached decorin (Figure 5B) nor that of HS-attached syndecan 4 (Figure 5C). Indeed, high amount of CS/DS-containing decorin and HS-attached syndecan 4 was produced in the presence of Fam20BD309A and Fam20BD289A, respectively indicating that mutation of Asp309 and Asp289 to alanine abolished the ability of Fam20B to suppress PG-GAG synthesis and therefore impair the activity of Fam20B. To further confirm this finding, wild-type Fam20B and the mutants Fam20BD289A and Fam20BD309A were transfected into HEK293 cells and neosynthesized PG-GAG chains were radiolabelled using 35S-sulfate incorporation. SDS-PAGE analysis of radiolabeled GAG chains produced by Fam20BD289A and Fam20BD309A expressing cells showed similar pattern as in the control (Figure 5D), whereas cell expressing wild-type Fam20B presented strong reduction (90%) in overall GAG chains produced (Figure 5D,E).
We next analyzed the effect of Fam20B mutations on the synthesis of decorin in human lung cancer cell line A549 which abundantly produced this PG in the medium. Immunoblot analysis of decorin from culture supernatants of lung fibroblast cells expressing Fam20BD289A and Fam20BD309A mutants showed high amount of CS/DS-attached decorin as in control cells (Figure 6F). In contrast, the amount of CS/DS-attached decorin produced by cells expressing wild-type Fam20B was decreased by about 75% (Figure 5F,G). These results showed that Fam20BD289A and Fam20BD309A mutants did not suppress the synthesis of CS/DS-attached decorin in lung fibroblasts as wild-type Fam20B does. Thus, these results clearly demonstrated that expression of an altered form of Fam20B did not alter the synthesis of GAG-attached PGs, therefore confirming that Fam20B gain-of-function is a negative regulator of PG synthesis.
3.5. Knockout of Fam20B Reduced the Synthesis of Both CS and HSPGs
We next analyzed the effect of the loss of Fam20B in HEK293 cells on the synthesis of PG-GAG chains by generating Fam20B-knockout HEK293 cells using CRISPR/Cas9 technique. Fam20B-knockout clone harbouring deletion mutation in exon 3 leading to premature stop codon was selected (Figure 6A) and transfected with vectors expressing either decorin or HA-syndecan 4. As shown by Western blot, transfection of control and Fam20B-knockout cells with decorin expression vectors resulted in the synthesis of high amount of CS/DS-attached decorin (Figure 6B). However, the amount of GAG-attached decorin produced in Fam20B-knockout HEK293 cells were reduced by about 75%, compared to control cells (Figure 6B,C). Similar results were observed for the HSPG syndecan 4. Expression of HS-attached stndecan 4 was dramatically reduced in Fam20B-knockout cells, compared to control cells. (Figure 6D,E). These results showed that absence of Fam20B is associated with decreased synthesis of GAG-attached PGs. Altogether, these data showed that both loss- and gain-of-function of Fam20B downregulate the synthesis of GAG-attached PGs arguing for a strict dependence of PG synthesis on Fam20B expression levels and therefore suggest that modulation of the expression of Fam20B regulates the rate of PG synthesis.
3.6. Fam20B Inhibits Proliferation and Migration of Glioblastoma Cells
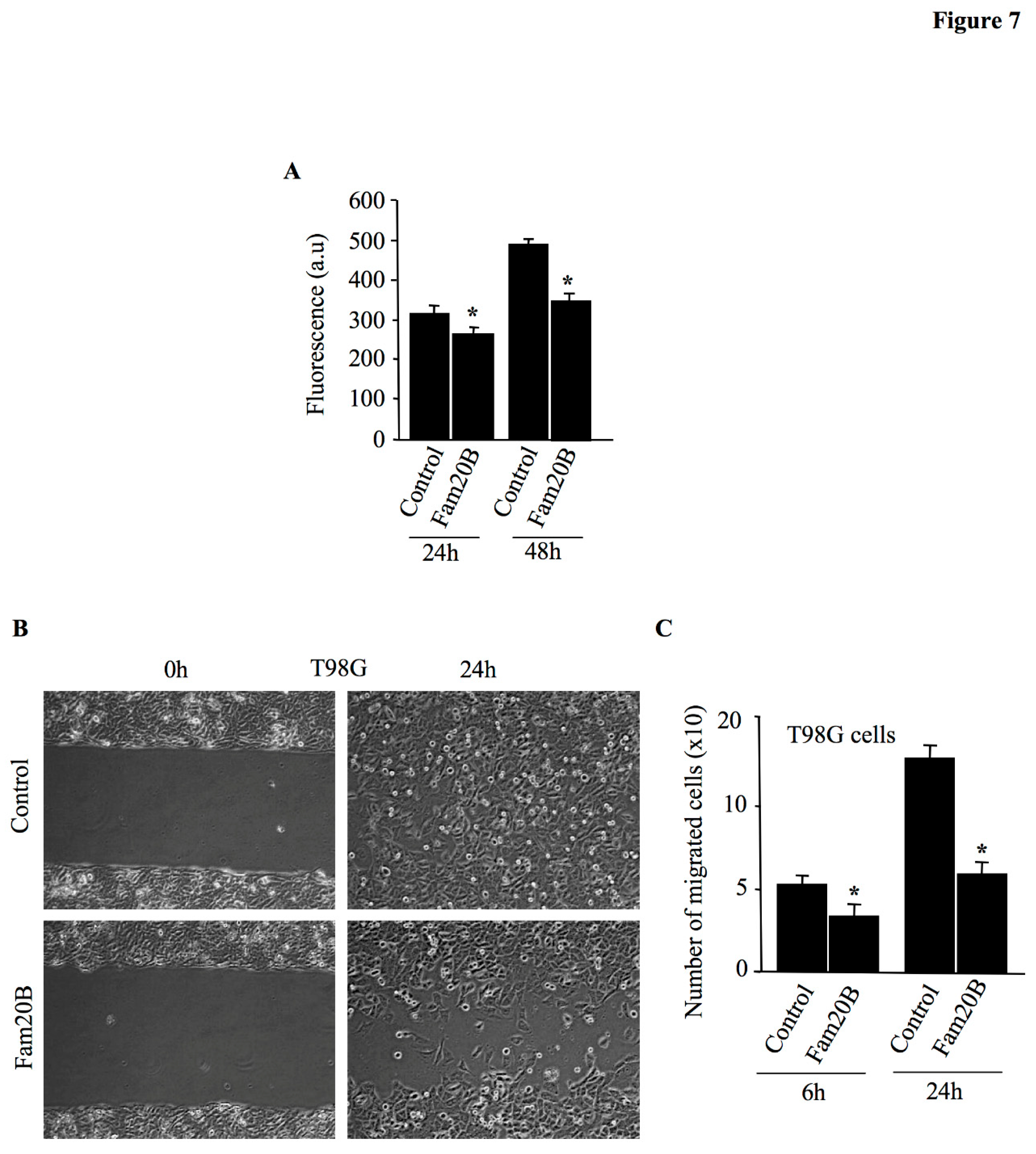
Because PGs play an important role in cell migration and proliferation, we hypothesized that Fam20B-dependent inhibition of PG-GAG synthesis may affect these processes. To test this hypothesis, Fam20B was expressed in brain tumor glioblastoma cell line T98G. Cell proliferation was measured at 24 h and 48 h after transfection. As shown in Figure 7A, glioblastoma cells transfected with Fam20B presented a reduction in cell proliferation of about 14% at 24 h and 28% at 48 h, compared to control. We next analyzed the ability of Fam20B-transfected cells to migrate using scratch wound-healing experiments. Interestingly, T98G cells transfected with Fam20B expression vector were impaired in their ability to migrate when compared to control cells (Figure 7B,C). The migration assays reveal a slower rate of cell migration by the Fam20B-transfected cells compared with control cells. Indeed, expression of Fam20B in glioblastoma cells reduced the number of cells that migrate in scratch area by about 36% at 6h and 53% at 24 h, compared to control (Figure 7B,C). These results support that Fam20B-dependant inhibition of PG-GAG synthesis impairs cell proliferation and migration, therefore highlighting the critical role of GAG chains of PGs in glioblastoma cell tumorigenesis.
4. Discussion
In this study we showed that Fam20B gain-of-function induced a strong reduction in overall PG-GAG chains produced in the cell. By using decorin and syndecan 4 as reporter proteins for CS and HS-attached PGs, we showed that Fam20B gain-of-function negatively regulates the synthesis of both CS- and HS-attached PGs. This has been further confirmed using human skin and lung fibroblasts expressing high amount of endogenous decorin in the medium. Fam20B gain-of-function blocks the synthesis of decorin in both skin and lung fibroblast cells. Similar results were obtained for endogenous HSPGs. By using anti-HS 10E4 antibody which is commonly used to detect HS chains of PGs [31,32,33], we showed that Fam20B gain-of-function strongly attenuates the expression of cell surface HSPGs. Remarkably, we showed that Fam20B gain-of-function reduced the synthesis of both CS/DS-attached decorin and HS-attached syndecan 4 in a dose dependent manner and induces intracellular accumulation of GAG-free decorin and syndecan 4. Therefore, suggesting that lack of GAG elongation prevents PGs from being secreted. Interestingly, reduced secretion in culture medium of decorin and syndecan 4 in Fam20B expressing cells was accompanied with increased amounts of intracellular non elongated decorin and syndecan 4 that are probably not competent for secretion and accumulates in the cells. Altogether, these data clearly demonstrate that Fam20B gain-of-function negatively regulates the synthesis of PGs of both types CS and HS. As the synthesis of CS and HS GAGs share a common tetrasaccharide primer, Fam20B gain-of-function affects the synthesis of the primer. Indeed, blocking of any step in the synthesis of this primer will obviously impact the synthesis of both CS and HS.
Of note, it has been reported that stable expression of Fam20B in Hela cells increased the amounts of GAGs with particularly augmented short CS chains [24]. On the other hand, it has been shown that loss-of-function of XYLP, the 2-phosphoxylose phosphatase that de-phosphorylates xylose of PG primer region, decreased HS and CS synthesis and inversely, gain-of-function increased their synthesis [27]. Using transient expression of Fam20B in several cell lines as well as primary fibroblast cells we found that Fam20B gain-of-function reduced the synthesis of GAG-attached PGs in all the cells tested. The discrepancy may be due to the selection of stable clones or to an unknown factor that has to be identified. In the other hand, in agreement with our study, it has been shown the knockout of Fam20B in bone osteosarcoma cells U2OS led to reduction in the synthesis of GAGs [25]. This study also showed that GalTII present higher activity towards phosphorylated Gal-Xylose, compared to its unphosphorylated counterpart, thus loss of Fam20B causes impaired GalTII activity resulting in the formation of incomplete linkage tetrasaccharides capped with sialic acid and cannot be elongated [25].
During the physiological conditions there is a balance between the phosorylation and dephosphorylation of the xylose by Fam20B and XYLP, respectively. Therefore, gain- or loss-of-function of either Fam20B or XYLP may disturb this balance thus affect the rate of GAG synthesis. How Fam20B inhibits the synthesis of GAG chains is unknown, however one can hypothesize that Fam20B gain-of-function may lead to sustained phosphorylation of xylose residue on the core protein, therefore competing with XYLP phosphatase that de-phosphorylates xylose of PG linker region, a process that may be required prior to subsequent GAG elongation. Noteworthy, a working model for the role of Fam20B in the synthesis of the tetrasaccharide linkage and subsequent GAG elongation suggests that xylose phosphorylation by Fam20B occurs predominantly after addition of the first galactosyl residue to the growing linkage region and that the removal of the xylose 2-phosphate moiety by the xylose 2-phosphatase, XYLP occurs following completion of the tetrasaccharide linkage to allow GAG elongation [34]. If the phosphate is not removed by XYLP, EXTL2 transfers the GlcNAc residue via an α1,4-linkage on the phosphorylated tetrasaccharide, leading to synthesis of phosphorylated pentasaccharide structure unable to serve as primer for CS/HS polymerization [27]. Whereas it has been reported that phosphorylated tetrasaccharide is a preferred substrate for ChGn-1 enzyme that transfers a GalNAc residue to the phosphorylated tetrasaccharide in the protein linkage region of CS.
Interestingly, it has been shown that phosphorylation of xylose before addition of the first galactosyl residue occurs. Indeed, analysis of [32P]-labelled O-linked glycan chains and stubs attached to intracellular decorin, revealed that 52% of Xyl-decorin was phosphorylated, therefore indicating that phosphorylation of xylose occurs before addition of galactose residue on Xyl-decorin and continues when the Gal residues are added and is complete at the trisaccharide stage [34,35]. In a previous study, we have reported that β4GalT7 efficiently catalyzes the transfer of galactose residue onto the non-phosphorylated xyloside analog, whereas the phosphorylated xylose analog at position 2-O was not substrate, suggesting that the 2-O phosphorylation precludes the transfer of the galactose on xylose [13]. In line with this, Siegbahn et al., showed that most modifications of position 2 in xylose, rendered analogs less prone to galactosylation by β4GalT7 [36] and showed that xylose analogs carrying modification on the 2-O position were not able to prime GAG synthesis in human cell lines [37]. These data indicate that the hydroxyl in position 2 might act as a hydrogen bond acceptor. The crystal structure of the β4GalT7 has been published recently and revealed that the Tyr177, Tyr179, Trp207, and Leu209 are important for the hydrophobic binding of the xylose and that the Asp211 forms a strong hydrogen bond with the OH present in the 4th position of the acceptor xylose. Whereas, the Asp212 interacts via a hydrogen bond with the OH present at 2nd position of the xylose [38]. This, suggested that phosphorylation of xylose before addition of Gal residue by Fam20B may lead to premature termination of the tetrasaccharide primer synthesis and subsequently to unelongated GAGs. This may constitute a mechanism to counteract excessive synthesis of elongated GAG chains.
All the members of the Fam20 family contains a conserved C-terminal domain including the catalytic domain DRHHYE and a DFG motif which is crucial for the metal ion binding. It has been shown that mutation of the aspartic acid residues in the catalytic segment and in the DFG motif of Fam20C abolished the enzyme activity [21]. These aspartic acid residues are conserved in all members of the Fam20 family and correspond to Asp289 and Asp309 of the human Fam20B. Mutation of either of the two aspartic residues to alanine abolished the ability of Fam20B to block the synthesis of both CS- and HS-attached PGs, suggesting that mutation of Asp289 and Asp309 impaired Fam20B activity, consistent with the putative role of theses residues in catalysis and in the binding of the divalent cation Mn2+, respectively.
On the other hand, analysis of biological process including cell proliferation and migration indicated that both cell proliferation and migration were impaired in Fam20B-transfected cells. Expression of Fam20B in glioblastoma cells led to a significant reduction in proliferation and in the ability of cells to migrate in scratch-wound healing assays. The importance of PGs and their GAG chains in cell proliferation and migration is well established [39] and numerous PGs have been implicated in tumor growth and metastasis [40]. Therefore, Fam20B-dependent inhibition of PG-GAG synthesis could contribute to the cell proliferation and migration defects observed in glioblastoma cells.
Author Contributions
M.O., L.B and I.S. conceived and designed the experiments. L.B. and I.S. conducted the experiments. M.O., L.B. and I.S. analyzed the results. M.O. and L.B. wrote the paper. All authors reviewed the manuscript.
Funding
This work was supported by grant from Région Lorraine.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data available on request.
Conflicts of interest: The authors declare no conflicts of interest.
References
- Perrimon, N.; Bernfield, M. Specificities of heparan sulphate proteoglycans in developmental processes. Nature 2000, 404, 725–728. [Google Scholar] [CrossRef] [PubMed]
- Knudson, C.B.; Nofal, G.A.; Pamintuan, L.; Aguiar, D.J. The chondrocyte pericellular matrix: a model for hyaluronan-mediated cell-matrix interactions. Biochem Soc Trans. 1999, 27, 142–147. [Google Scholar] [CrossRef]
- Melrose, J.; Shu, C.; Whitelock, J.M.; Lord, M.S. The cartilage extracellular matrix as a transient developmental scaffold for growth plate maturation. Matrix Biol. 2016, 52-54, 363–383. [Google Scholar] [CrossRef]
- Couchman, J.R. Transmembrane signaling proteoglycans. Annu Rev Cell Dev Biol. 2010, 26, 89–114. [Google Scholar] [CrossRef]
- Manon-Jensen, T.; Itoh, Y.; Couchman, J.R. Proteoglycans in health and disease: The multiple roles of syndecan shedding. FEBS Journal. 2010, 277, 3876–3889. [Google Scholar] [CrossRef]
- Li, L.; Ly, M.; Linhardt, R.J. Proteoglycan sequence. Molecular BioSystems. 2012, 8, 1613–1625. [Google Scholar] [CrossRef] [PubMed]
- Gesslbauer, B.; Theuer, M.; Schweiger, D.; Adage, T.; Kungl, A.J. New targets for glycosaminoglycans and glycosaminoglycans as novel targets. Expert Rev Proteomics. 2013, 10, 77–95. [Google Scholar] [CrossRef] [PubMed]
- Götting, C.; Kuhn, J.; Kleesiek, K. Human xylosyltransferases in health and disease. Cellular and Molecular Life Sciences. 2007, 64, 1498–1517. [Google Scholar] [CrossRef]
- Wilson, I.B.H. The never-ending story of peptide O-xylosyltransferase. Cellular and Molecular Life Sciences CMLS. 2004, 61, 794–809. [Google Scholar] [CrossRef]
- Venkatesan, N.; Barré, L.; Magdalou, J.; Mainard, D.; Netter, P.; Fournel-Gigleux, S.; Ouzzine, M. Modulation of xylosyltransferase i expression provides a mechanism regulating glycosaminoglycan chain synthesis during cartilage destruction and repair. FASEB Journal. 2009, 23, 813–822. [Google Scholar] [CrossRef]
- Breton, C.; Fournel-Gigleux, S.; Palcic, M.M. Recent structures, evolution and mechanisms of glycosyltransferases. Current Opinion in Structural Biology. 2012, 22, 540–549. [Google Scholar] [CrossRef] [PubMed]
- Habuchi, H.; Habuchi, O.; Kimata, K. Sulfation pattern in glycosaminoglycan: does it have a code? Glycoconj J. 2004, 21, 47–52. [Google Scholar] [CrossRef]
- Gulberti, S.; Lattard, V.; Fondeur, M.; Jacquinet, J.C.; Mulliert, G.; Netter, P.; Magdalou, J.; Ouzzine, M.; Fournel-Gigleux, S. Phosphorylation and sulfation of oligosaccharide substrates critically influence the activity of human β1,4-galactosyltransferase 7 (GalT-I) and β1,3-glucuronosyltransferase I (GlcAT-I) involved in the biosynthesis of the glycosaminoglycan-protein linkage region of proteoglycans. Journal of Biological Chemistry. 2005, 280, 1417–1425. [Google Scholar]
- Gulberti, S.; Lattard, V.; Fondeur, M.; Jacquinet, J.C.; Mulliert, G.; Netter, P.; Magdalou, J.; Ouzzine, M.; Fournel-Gigleux, S. Modifications of the glycosaminoglycan-linkage region of proteoglycans: phosphorylation and sulfation determine the activity of the human beta1,4-galactosyltransferase 7 and beta1,3-glucuronosyltransferase I. TheScientificWorldJournal [electronic resource]. 2005, 5, 510–514. [Google Scholar] [CrossRef] [PubMed]
- Nalbant, D.; Youn, H.; Nalbant, S.I.; Sharma, S.; Cobos, E.; Beale, E.G.; Du, Y.; Williams, S.C. FAM20: an evolutionarily conserved family of secreted proteins expressed in hematopoietic cells. BMC genomics. 2005, 6, 11. [Google Scholar] [CrossRef]
- Tagliabracci, V.S.; Xiao, J.; Dixon, J.E. Phosphorylation of substrates destined for secretion by the Fam20 kinases. Biochemical Society Transactions. 2013, 41, 1061–1065. [Google Scholar] [CrossRef] [PubMed]
- Vogel, P.; Hansen, G.; Read, R.; Vance, R.; Thiel, M.; Liu, J.; Wronski, T.; Smith, D.; Jeter-Jones, S.; Brommage, R. Amelogenesis imperfecta and other biomineralization defects in Fam20a and Fam20c null mice. Veterinary Pathology Online. 2012, 49, 998–1017. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.H.; Seymen, F.; Lee, K.E.; Lee, S.K.; Kweon, Y.S.; Kim, K.J.; Jung, S.E.; Song, S.J.; Yildirim, M.; Bayram, M. Novel FAM20A mutations in hypoplastic amelogenesis imperfecta. Human mutation. 2012, 33, 91–94. [Google Scholar] [CrossRef]
- O'Sullivan, J.; Bitu, C.C.; Daly, S.B.; Urquhart, J.E.; Barron, M.J.; Bhaskar, S.S.; Martelli-Junior, H.; dos Santos Neto, P.E.; Mansilla, M.A.; Murray, J.C.; et al. Whole-Exome sequencing identifies FAM20A mutations as a cause of amelogenesis imperfecta and gingival hyperplasia syndrome. Am J Hum Genet. 2011, 88, 616–620. [Google Scholar] [CrossRef]
- Jaureguiberry, G.; Parry, D.; Quentric, M.; Himmerkus, N.; Koike, T.; Poulter, J.; Klootwijk, E.; Robinette, S.; Howie, A.; Patel, V. Nephrocalcinosis (Enamel Renal Syndrome) Caused by Autosomal Recessive FAM20A Mutations. Nephron Physiology. 2013, 122, 1–6. [Google Scholar] [CrossRef]
- Tagliabracci, V.S.; Engel, J.L.; Wen, J.; Wiley, S.E.; Worby, C.A.; Kinch, L.N.; Xiao, J.; Grishin, N.V.; Dixon, J.E. Secreted Kinase Phosphorylates Extracellular Proteins That Regulate Biomineralization. Science. 2012, 336, 1150–1153. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, H.O.; Xu, A.; Ogura, E.; Manning, G.; Irvine, K.D. The Raine Syndrome Protein FAM20C Is a Golgi Kinase That Phosphorylates Bio-Mineralization Proteins. PLoS ONE. 2012, 7, e42988. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Tagliabracci, V.S.; Wen, J.; Kim, S.-A.; Dixon, J.E. Crystal structure of the Golgi casein kinase. Proceedings of the National Academy of Sciences. 2013. [Google Scholar] [CrossRef] [PubMed]
- Koike, T.; Izumikawa, T.; Tamura, J.I.; Kitagawa, H. FAM20B is a kinase that phosphorylates xylose in the glycosaminoglycan–protein linkage region. Biochemical Journal. 2009, 421, 157–162. [Google Scholar] [CrossRef]
- Wen, J.; Xiao, J.; Rahdar, M.; Choudhury, B.P.; Cui, J.; Taylor, G.S.; Esko, J.D.; Dixon, J.E. Xylose phosphorylation functions as a molecular switch to regulate proteoglycan biosynthesis. Proc Natl Acad Sci U S A. 2014, 111, 15723–15728. [Google Scholar] [CrossRef]
- Eames, B.F.; Yan, Y.-L.; Swartz, M.E.; Levic, D.S.; Knapik, E.W.; Postlethwait, J.H.; Kimmel, C.B. Mutations in fam20b and xylt1 Reveal That Cartilage Matrix Controls Timing of Endochondral Ossification by Inhibiting Chondrocyte Maturation. PLoS Genet. 2011, 7, e1002246. [Google Scholar] [CrossRef]
- Koike, T.; Izumikawa, T.; Sato, B.; Kitagawa, H. Identification of Phosphatase That Dephosphorylates Xylose in the Glycosaminoglycan-Protein Linkage Region of Proteoglycans. Journal of Biological Chemistry. 2014, 289, 6695–6708. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical biochemistry. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- De Vries, B.; Van den Berg, W.; Vitters, E.; Van de Putte, L. Quantitation of glycosaminoglycan metabolism in anatomically intact articular cartilage of the mouse patella: in vitro and in vivo studies with 35S-sulfate, 3H-glucosamine, and 3H-acetate. Rheumatology international. 1986, 6, 273–281. [Google Scholar] [CrossRef]
- Bronson, R.E.; Bertolami, C.N.; Siebert, E.P. Modulation of fibroblast growth and glycosaminoglycan synthesis by interleukin-1. Coll Relat Res. 1987, 7, 323–332. [Google Scholar] [CrossRef]
- Mani, K.; Cheng, F.; Sandgren, S.; Van Den Born, J.; Havsmark, B.; Ding, K.; Fransson, L.-Å. The heparan sulfate–specific epitope 10E4 is NO-sensitive and partly inaccessible in glypican-1. Glycobiology. 2004, 14, 599–607. [Google Scholar] [CrossRef] [PubMed]
- David, G.; Bai, X.M.; Van Der Schueren, B.; Cassiman, J.-J.; Van Den Berghe, H. Developmental changes in heparan sulfate expression: in situ detection with mAbs. The Journal of Cell Biology. 1992, 119, 961–975. [Google Scholar] [CrossRef] [PubMed]
- Leteux, C.; Chai, W.; Nagai, K.; Herbert, C.G.; Lawson, A.M.; Feizi, T. 10E4 antigen of scrapie lesions contains an unusual nonsulfated heparan motif. Journal of Biological Chemistry. 2001, 276, 12539–12545. [Google Scholar] [CrossRef]
- Moses, J.; Oldberg, A.; Cheng, F.; Fransson, L.A. Biosynthesis of the proteoglycan decorin--transient 2-phosphorylation of xylose during formation of the trisaccharide linkage region. Eur J Biochem. 1997, 248, 521–526. [Google Scholar] [CrossRef]
- Moses, J.; Oldberg, A.; Fransson, L.A. Initiation of galactosaminoglycan biosynthesis. Separate galactosylation and dephosphorylation pathways for phosphoxylosylated decorin protein and exogenous xyloside. Eur J Biochem. 1999, 260, 879–884. [Google Scholar] [CrossRef] [PubMed]
- Siegbahn, A.; Manner, S.; Persson, A.; Tykesson, E.; Holmqvist, K.; Ochocinska, A.; Ronnols, J.; Sundin, A.; Mani, K.; Westergren-Thorsson, G.; et al. Rules for priming and inhibition of glycosaminoglycan biosynthesis; probing the [small beta]4GalT7 active site. Chemical Science. 2014, 5, 3501–3508. [Google Scholar] [CrossRef]
- Siegbahn, A.; Aili, U.; Ochocinska, A.; Olofsson, M.; Ronnols, J.; Mani, K.; Widmalm, G.; Ellervik, U. Synthesis, conformation and biology of naphthoxylosides. Bioorg Med Chem. 2011, 19, 4114–4126. [Google Scholar] [CrossRef]
- Tsutsui, Y.; Ramakrishnan, B.; Qasba, P.K. Crystal Structures of β-1,4-Galactosyltransferase 7 Enzyme Reveal Conformational Changes and Substrate Binding. Journal of Biological Chemistry. 2013, 288, 31963–31970. [Google Scholar] [CrossRef]
- Mythreye, K.; Blobe, G.C. Proteoglycan signaling co-receptors: roles in cell adhesion, migration and invasion. Cell Signal. 2009, 21, 1548–1558. [Google Scholar] [CrossRef]
- Afratis, N.; Gialeli, C.; Nikitovic, D.; Tsegenidis, T.; Karousou, E.; Theocharis, A.D.; Pavao, M.S.; Tzanakakis, G.N.; Karamanos, N.K. Glycosaminoglycans: key players in cancer cell biology and treatment. FEBS J. 2012, 279, 1177–1197. [Google Scholar] [CrossRef]
Figure 1.
Fam20B blocks the synthesis of PG-GAG chains. HEK293 cells were transfected with pCMV-Fam20B or pCMV empty vector (control) and (A) Fam20B was detected by Western blot using anti-Fam20B antibodies. β-actin was used as loading control. (B) Western blot analysis of the sensitivity of Fam20B to digestion with PNGase F. (C) Analysis of the level of PG-GAG chain synthesis in HEK293 cells transfected with either Fam20B expression vector or empty plasmid (control) using 35S-sulfate incorporation method. (D) SDS-PAGE analysis of radiolabeled GAG chains produced in culture medium of HEK293 cells transfected with either Fam20B expression vector or empty plasmid (control) using 35S-sulfate incorporation method. The amounts of 35S-sulfate GAGs were normalized to DNA and relative to control. (E) The bar graph represents the quantification of GAG chains on the autoradiography of the SDS-PAGE. Data were presented as mean ± SD of three separate experiments. Statistical significance was evaluated using Student’s t test (*, P<0.05).
Figure 1.
Fam20B blocks the synthesis of PG-GAG chains. HEK293 cells were transfected with pCMV-Fam20B or pCMV empty vector (control) and (A) Fam20B was detected by Western blot using anti-Fam20B antibodies. β-actin was used as loading control. (B) Western blot analysis of the sensitivity of Fam20B to digestion with PNGase F. (C) Analysis of the level of PG-GAG chain synthesis in HEK293 cells transfected with either Fam20B expression vector or empty plasmid (control) using 35S-sulfate incorporation method. (D) SDS-PAGE analysis of radiolabeled GAG chains produced in culture medium of HEK293 cells transfected with either Fam20B expression vector or empty plasmid (control) using 35S-sulfate incorporation method. The amounts of 35S-sulfate GAGs were normalized to DNA and relative to control. (E) The bar graph represents the quantification of GAG chains on the autoradiography of the SDS-PAGE. Data were presented as mean ± SD of three separate experiments. Statistical significance was evaluated using Student’s t test (*, P<0.05).
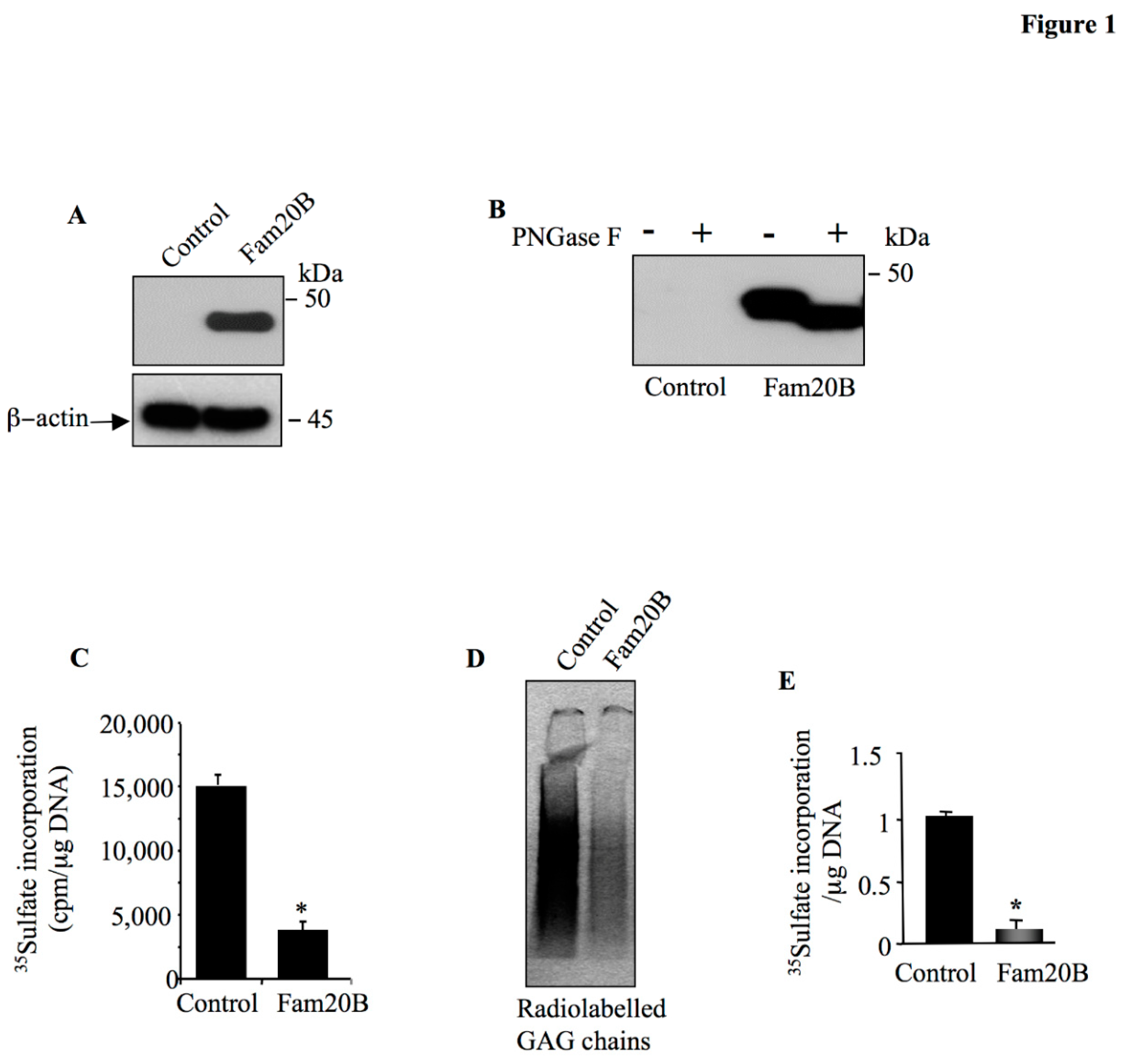
Figure 2.
Fam20B suppression of GAG chain synthesis is dose-dependent. (A) HEK293 cells were co-transfected with pCMV-DCN along with pCMV-Fam20B or empty pCMV vector (control) and the expression of DCN was analyzed in medium by Western blot using anti-DCN antibodies. (B) CHO PgsB-618 cells were co-transfected with pCMV-DCN along with empty pCMV (control), pCMV-Fam20B, pCMV-β4GalT7 or pCMV-Fam20B and pCMV-β4GalT7. DCN produced in the medium was analyzed by Western blot using anti-DCN antibodies. (C) Western blot analysis of Fam20B and DCN produced in the medium HEK293 cells transfected with pCMV-DCN and increased concentrations of Fam20B. (D) The bar graph represents the quantification of DCN in the Western blot. (E) Detection of intracellular DCN in cells transfected with pCMV-DCN and increased concentrations of Fam20B. β-actin was used as loading control. (F) The bar graph represents the quantification of DCN in the Western blot. (G) Western blot analysis of secreted DCN in the medium of human primary skin fibroblasts transfected with pCMV-Fam20B or empty vector (control). (H) The bar graph represents the quantification of DCN in the Western blot. (I) Detection of secreted DCN in the medium of A549 cells transfected with pCMV-Fam20B or empty vector (control). (J) The bar graph represents the quantification of DCN in the Western blot. Data were presented as mean ± SD of three separate experiments. Statistical significance was evaluated using Student’s t test (*, P<0.05).
Figure 2.
Fam20B suppression of GAG chain synthesis is dose-dependent. (A) HEK293 cells were co-transfected with pCMV-DCN along with pCMV-Fam20B or empty pCMV vector (control) and the expression of DCN was analyzed in medium by Western blot using anti-DCN antibodies. (B) CHO PgsB-618 cells were co-transfected with pCMV-DCN along with empty pCMV (control), pCMV-Fam20B, pCMV-β4GalT7 or pCMV-Fam20B and pCMV-β4GalT7. DCN produced in the medium was analyzed by Western blot using anti-DCN antibodies. (C) Western blot analysis of Fam20B and DCN produced in the medium HEK293 cells transfected with pCMV-DCN and increased concentrations of Fam20B. (D) The bar graph represents the quantification of DCN in the Western blot. (E) Detection of intracellular DCN in cells transfected with pCMV-DCN and increased concentrations of Fam20B. β-actin was used as loading control. (F) The bar graph represents the quantification of DCN in the Western blot. (G) Western blot analysis of secreted DCN in the medium of human primary skin fibroblasts transfected with pCMV-Fam20B or empty vector (control). (H) The bar graph represents the quantification of DCN in the Western blot. (I) Detection of secreted DCN in the medium of A549 cells transfected with pCMV-Fam20B or empty vector (control). (J) The bar graph represents the quantification of DCN in the Western blot. Data were presented as mean ± SD of three separate experiments. Statistical significance was evaluated using Student’s t test (*, P<0.05).
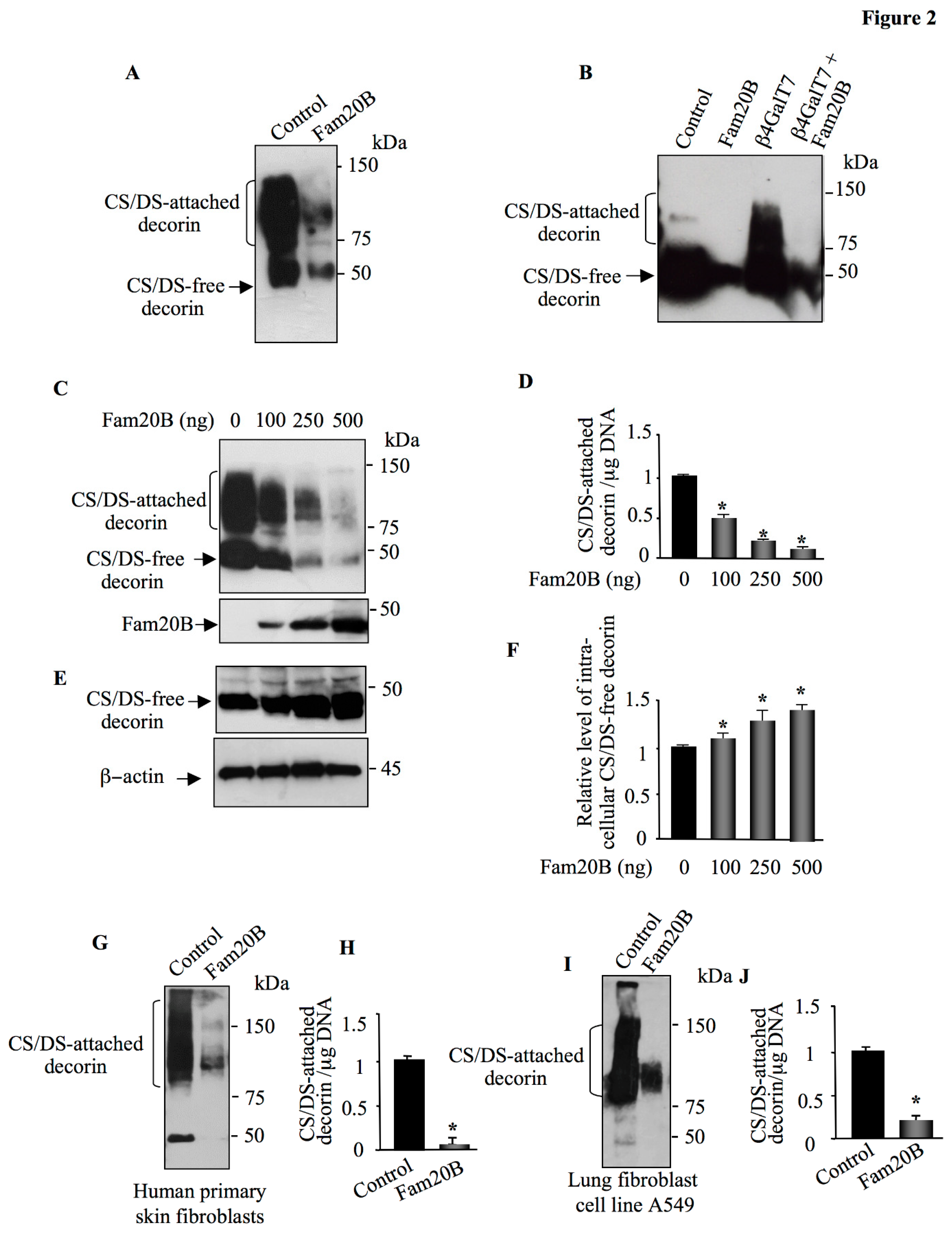
Figure 3.
Fam20B blocks the synthesis of HSPGs. (A) HEK293 cells were co-transfected with pCMV-HA-SDC4 and either pCMV-Fam20B or empty pCMV (control) and SDC4 was analyzed in cell lysate by Western blot using anti-HA antibodies. β-actin was used as loading control. (B) The bar graph represents the quantification of the Western blots. (C) Western blot analysis of SDC4 and of Fam20B in cell lysate of HEK293 cells transfected with increased concentrations of Fam20B. β-actin was used as loading control. (D) The bar graph represents the quantification of HS-attached SDC4 in the Western blot. (E) The bar graph represents the quantification of HS-free SDC4 in the Western blot. (F) CHO-K1 cells were transfected with pCMV-Fam20B or empty pCMV (control) vector and expression of cell surface HSPGs was examined by immunofluorescence using anti-heparan sulfate antibody 10E4 (green). The expression of Fam20B was analyzed using anti-Flag antibody (red). Nuclei were stained (blue) using Hoechst/PBS solution. Data were presented as mean ± SD of three separate experiments. Statistical significance was evaluated using Student’s t test (*, P<0.05).
Figure 3.
Fam20B blocks the synthesis of HSPGs. (A) HEK293 cells were co-transfected with pCMV-HA-SDC4 and either pCMV-Fam20B or empty pCMV (control) and SDC4 was analyzed in cell lysate by Western blot using anti-HA antibodies. β-actin was used as loading control. (B) The bar graph represents the quantification of the Western blots. (C) Western blot analysis of SDC4 and of Fam20B in cell lysate of HEK293 cells transfected with increased concentrations of Fam20B. β-actin was used as loading control. (D) The bar graph represents the quantification of HS-attached SDC4 in the Western blot. (E) The bar graph represents the quantification of HS-free SDC4 in the Western blot. (F) CHO-K1 cells were transfected with pCMV-Fam20B or empty pCMV (control) vector and expression of cell surface HSPGs was examined by immunofluorescence using anti-heparan sulfate antibody 10E4 (green). The expression of Fam20B was analyzed using anti-Flag antibody (red). Nuclei were stained (blue) using Hoechst/PBS solution. Data were presented as mean ± SD of three separate experiments. Statistical significance was evaluated using Student’s t test (*, P<0.05).
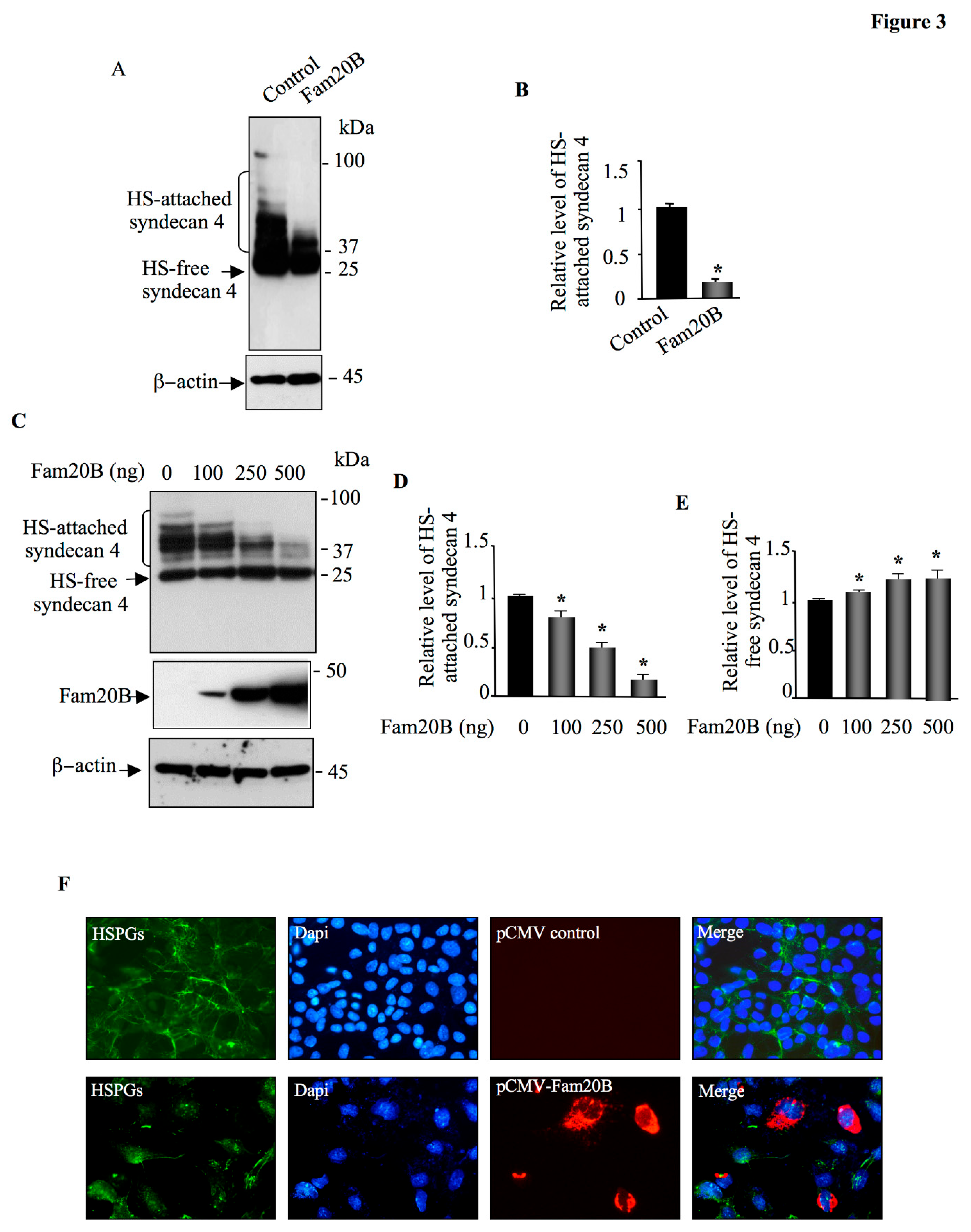
Figure 4.
XYLP rescue the synthesis of GAG-chains produced by Fam20B. (A) HEK293 Cells were transfected with pCMV-DCN together with increased concentrations of pCMV-HA-XYLP. Expression of DCN and XYLP was analyzed by Western blot using anti-DCN and anti-HA antibodies, respectively. β-actin was used as loading control. (B) HEK293 cells were transfected with pCMV-DCN (control), pCMV-DCN and pCMV-Fam20B or pCMV-DCN and pCMV-Fam20B together with pCMV-HA-XYLP. DCN, FAM20B and XYLP were analyzed by Western blot using anti-DCN, anti-Fam20B and anti-HA antibodies, respectively. β-actin was used as loading control. (C) HEK293 cells were transfected with pCMV-HA-SDC4, pCMV-HA-SDC4 and pCMV-Fam20B or pCMV-HA-SDC4 and pCMV-Fam20B together with pCMV-Flag-XYLP. SDC4, FAM20B and XYLP were analyzed in cell lysate by Western blot using anti-HA, anti-Fam20B and anti-Flag antibodies, respectively. β-actin was used as loading control. Data were presented as mean ± SD of three separate experiments.
Figure 4.
XYLP rescue the synthesis of GAG-chains produced by Fam20B. (A) HEK293 Cells were transfected with pCMV-DCN together with increased concentrations of pCMV-HA-XYLP. Expression of DCN and XYLP was analyzed by Western blot using anti-DCN and anti-HA antibodies, respectively. β-actin was used as loading control. (B) HEK293 cells were transfected with pCMV-DCN (control), pCMV-DCN and pCMV-Fam20B or pCMV-DCN and pCMV-Fam20B together with pCMV-HA-XYLP. DCN, FAM20B and XYLP were analyzed by Western blot using anti-DCN, anti-Fam20B and anti-HA antibodies, respectively. β-actin was used as loading control. (C) HEK293 cells were transfected with pCMV-HA-SDC4, pCMV-HA-SDC4 and pCMV-Fam20B or pCMV-HA-SDC4 and pCMV-Fam20B together with pCMV-Flag-XYLP. SDC4, FAM20B and XYLP were analyzed in cell lysate by Western blot using anti-HA, anti-Fam20B and anti-Flag antibodies, respectively. β-actin was used as loading control. Data were presented as mean ± SD of three separate experiments.
Figure 5.
The aspartic acid residues Asp289 and Asp309 are essential for Fam20B function. (A) Detection of wild-type Fam20B and mutants in HEK293 cells transfected with pCMV-Fam20B, pCMV-Fam20BD289A, pCMV-Fam20BD309A or pCMV empty vector (control) using anti-FAM20B antibodies. β-actin was used as loading control. (B) HEK293 cells were co-transfected with pCMV-DCN along with pCMV-Fam20B, pCMV-Fam20BD289A, pCMV-Fam20BD309A or pCMV empty vector (control) and DCN produced in the medium was analyzed by Western blot using anti-DCN antibodies. (C) HEK293 cells were co-transfected with pCMV-HA-SDC4 along with pCMV-Fam20B, pCMV-Fam20BD289A, pCMV-Fam20BD309A or pCMV empty vector (control) and SDC4 was analyzed by Western blot using anti-HA antibodies. β-actin was used as loading control. (D) PG-GAG chains produced in the medium of HEK293 cells transfected with pCMV-Fam20B, pCMV-Fam20BD289A, pCMV-Fam20BD309A or pCMV empty vector (control) were metabolically labelled by 35S-sulfate incorporation and isolated by CPC precipitation then separated by SDS-PAGE and revealed by autoradiography. (E) The bar graph represents the quantification of 35S-sullfate radiolabelled GAG chains of the autoradiography. (F) A549 cells were transfected with pCMV-Fam20B, pCMV-Fam20BD289A, pCMV-Fam20BD309A or pCMV empty vector (control) and DCN secreted by the cells in the medium was analyzed by Western blot using anti-DCN antibodies. (G) The bar graph represents the quantification of the Western blot. Data were presented as mean ± SD of three separate experiments. Statistical significance was evaluated using Student’s t test (*, P<0.05).
Figure 5.
The aspartic acid residues Asp289 and Asp309 are essential for Fam20B function. (A) Detection of wild-type Fam20B and mutants in HEK293 cells transfected with pCMV-Fam20B, pCMV-Fam20BD289A, pCMV-Fam20BD309A or pCMV empty vector (control) using anti-FAM20B antibodies. β-actin was used as loading control. (B) HEK293 cells were co-transfected with pCMV-DCN along with pCMV-Fam20B, pCMV-Fam20BD289A, pCMV-Fam20BD309A or pCMV empty vector (control) and DCN produced in the medium was analyzed by Western blot using anti-DCN antibodies. (C) HEK293 cells were co-transfected with pCMV-HA-SDC4 along with pCMV-Fam20B, pCMV-Fam20BD289A, pCMV-Fam20BD309A or pCMV empty vector (control) and SDC4 was analyzed by Western blot using anti-HA antibodies. β-actin was used as loading control. (D) PG-GAG chains produced in the medium of HEK293 cells transfected with pCMV-Fam20B, pCMV-Fam20BD289A, pCMV-Fam20BD309A or pCMV empty vector (control) were metabolically labelled by 35S-sulfate incorporation and isolated by CPC precipitation then separated by SDS-PAGE and revealed by autoradiography. (E) The bar graph represents the quantification of 35S-sullfate radiolabelled GAG chains of the autoradiography. (F) A549 cells were transfected with pCMV-Fam20B, pCMV-Fam20BD289A, pCMV-Fam20BD309A or pCMV empty vector (control) and DCN secreted by the cells in the medium was analyzed by Western blot using anti-DCN antibodies. (G) The bar graph represents the quantification of the Western blot. Data were presented as mean ± SD of three separate experiments. Statistical significance was evaluated using Student’s t test (*, P<0.05).
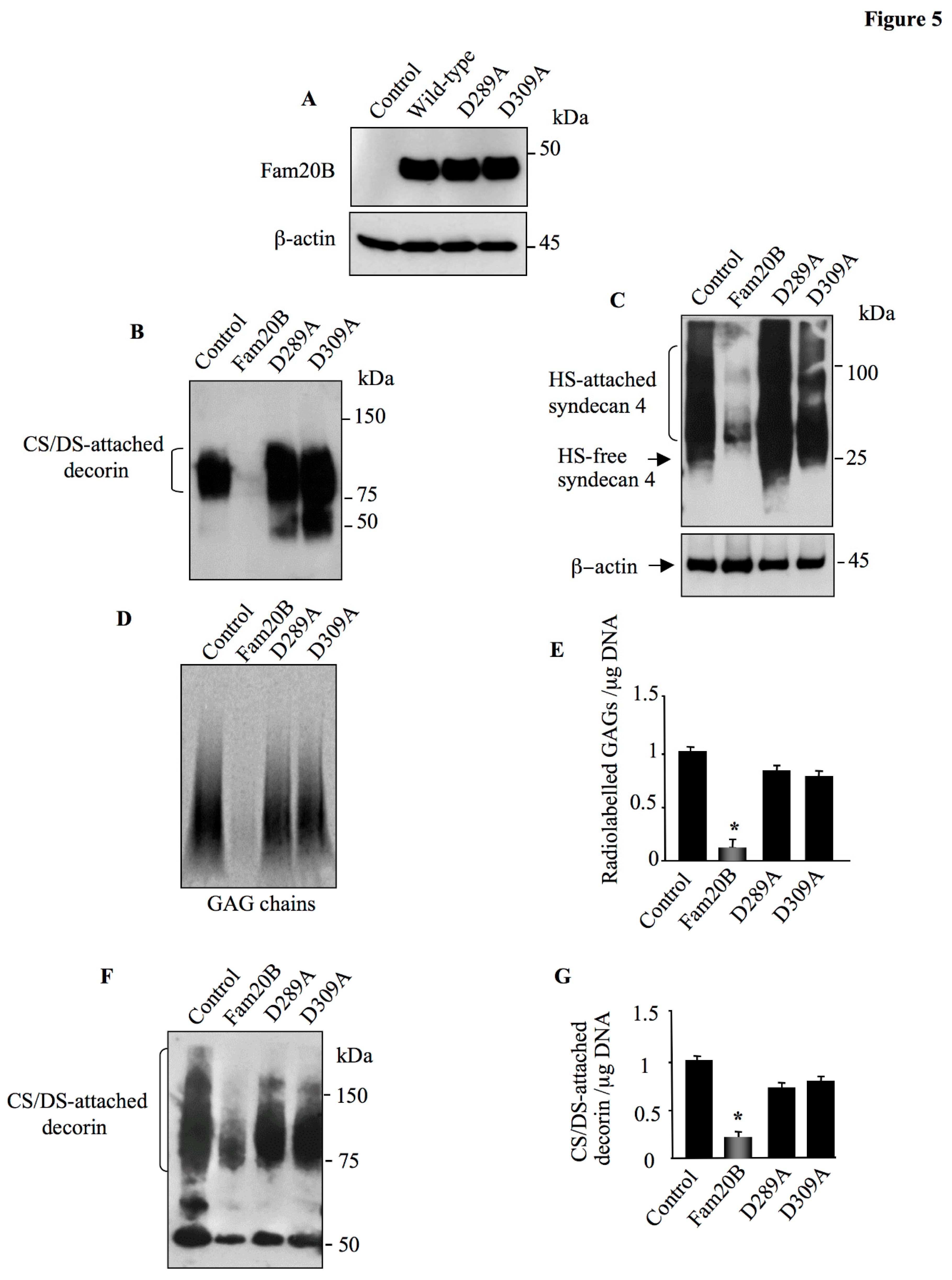
Figure 6.
CRISPR/Cas9 knockdown of Fam20B. (A) Alignment of Fam20B targeted sequence from wild-type and mutant HEK293 cells. (B). Detection of DCN in the culture medium of control and Fam20B-knockout HEK293 cells transfected with pCMV-DCN. (C) The bar graph represents the quantification of the Western blot. (D) Western blot analysis of SDC4 in cell lysate of HEK293 cells (control) and Fam20B-knockout HEK293 cells transfected with pCMV-HA-SDC4. β-actin was used as loading control. (E) The bar graph represents the quantification of the Western blot. Data were presented as mean ± SD of three separate experiments. Statistical significance was evaluated using Student’s t test (*, P<0.05).
Figure 6.
CRISPR/Cas9 knockdown of Fam20B. (A) Alignment of Fam20B targeted sequence from wild-type and mutant HEK293 cells. (B). Detection of DCN in the culture medium of control and Fam20B-knockout HEK293 cells transfected with pCMV-DCN. (C) The bar graph represents the quantification of the Western blot. (D) Western blot analysis of SDC4 in cell lysate of HEK293 cells (control) and Fam20B-knockout HEK293 cells transfected with pCMV-HA-SDC4. β-actin was used as loading control. (E) The bar graph represents the quantification of the Western blot. Data were presented as mean ± SD of three separate experiments. Statistical significance was evaluated using Student’s t test (*, P<0.05).
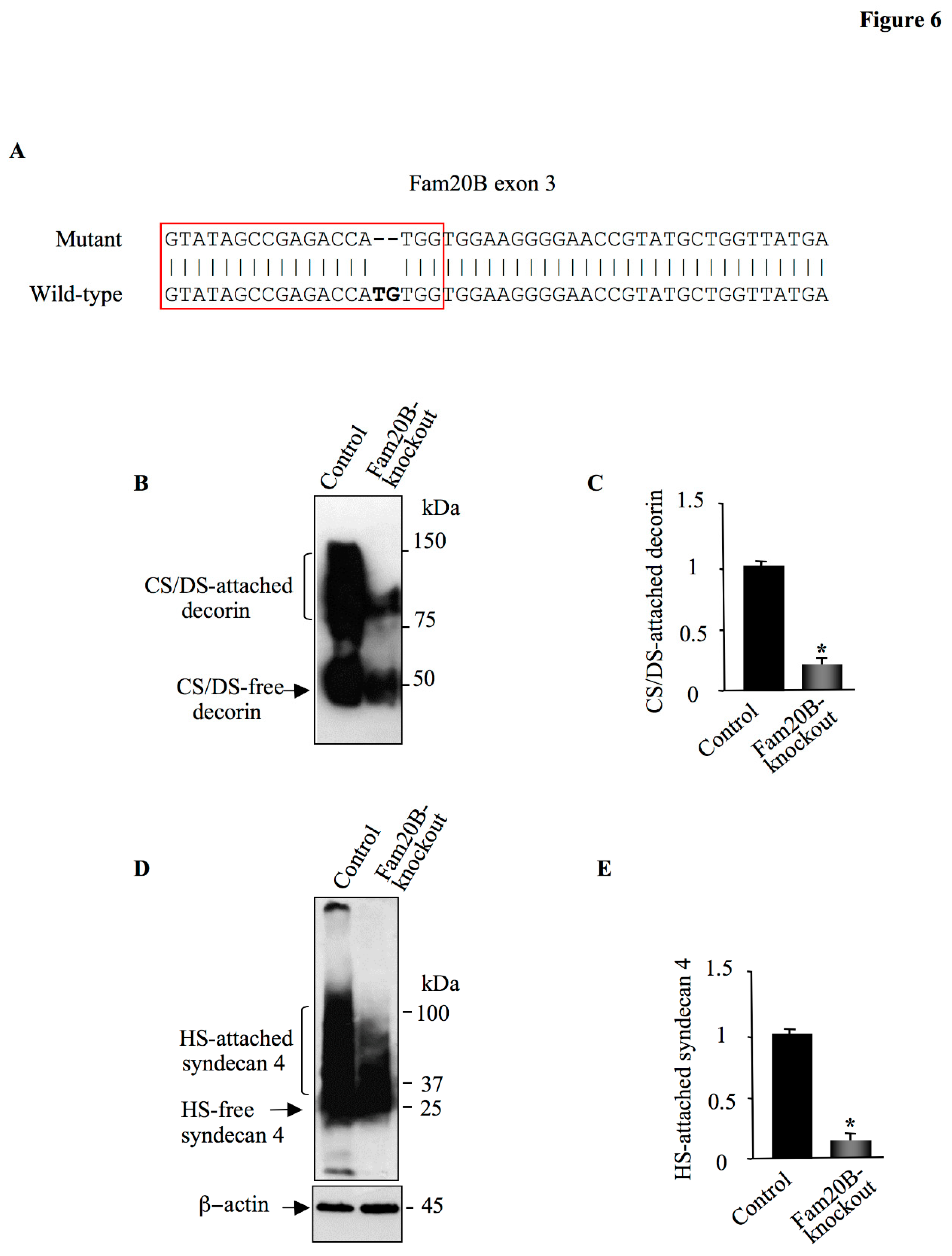
Figure 7.
Fam20B reduces the proliferation and migration of glioblastoma cells T98G. (A) Glioblastoma cells T98G were plated in 96 well plates and transfected with pCMV-Fam20B or empty pCMV vector (control) and cell proliferation was measured by CyQUANT NF Cell Proliferation Assay Kit at 24 h and 48 h after transfection. (B) Migration of glioblastoma cell line T98G transfected with pCMV-Fam20B or pCMV empty vector (control) was assessed using scratch wound-healing assay as described in Materials and Methods section and cell migration was photographed at 0 and 24h after scratch by phase-contrast microscopy. (C) The bar graph depicting the number of cells that migrate in the scratch area after 6 h and 24 h. Data were presented as mean ± SD of three separate experiments with three replicates each. Statistical significance was evaluated using Student’s t test (*, P<0.05).
Figure 7.
Fam20B reduces the proliferation and migration of glioblastoma cells T98G. (A) Glioblastoma cells T98G were plated in 96 well plates and transfected with pCMV-Fam20B or empty pCMV vector (control) and cell proliferation was measured by CyQUANT NF Cell Proliferation Assay Kit at 24 h and 48 h after transfection. (B) Migration of glioblastoma cell line T98G transfected with pCMV-Fam20B or pCMV empty vector (control) was assessed using scratch wound-healing assay as described in Materials and Methods section and cell migration was photographed at 0 and 24h after scratch by phase-contrast microscopy. (C) The bar graph depicting the number of cells that migrate in the scratch area after 6 h and 24 h. Data were presented as mean ± SD of three separate experiments with three replicates each. Statistical significance was evaluated using Student’s t test (*, P<0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



