
Submitted:
05 April 2025
Posted:
09 April 2025
You are already at the latest version
Abstract
Nuisance and harmful algal blooms can negatively impact freshwater, estuarine, and coastal marine systems globally, and pose serious risks to water quality, human and ecosystem health, and food production. Algae can produce toxic compounds, directly interfere with aquaculture species through (e.g.) production of foam or mucilage and can result in hypoxic conditions when the bloom senesces. Application of US Environmental Protection Agency (USEPA) registered algaecides can be effective, scalable, and inexpensive, but there is growing interest in plant or bacterial derived compounds that do not require the use of chemicals such as hydrogen peroxide or copper. The algaecide C7X1 is a plant based organic algaecide that proves effective against a wide variety of algae, including harmful algal species such as Microcystis, Heterosigma, and Pseudo-nitzschia. Performance is comparable to other USEPA-registered algaecides, with minimal toxin extracellular toxin release and a potential lifetime of weeks in treated waters. The mode of action is inhibition of photosynthesis, suggesting that direct off-target impacts to zooplankton and other organisms would be minimal.
Keywords:
harmful algal blooms
; treatment
; Microcystis
; microcystin
; Pseudo-nitzschia
; Chattonella
; Heterosigma akashiwo
1. Introduction
Harmful algal blooms are ubiquitous in freshwater, brackish, and marine environments [1,2,3,4]. HABs can include microalgae, cyanobacteria, and macroalgae, and can include both low and high biomass events, toxic algae (such as Pseudo-nitzschia which produces domoic acid [5], or Microcystis which produces microcystins [6]), and fish-killing algae (such as Heterosigma akashiwo, exact mechanisms unknown [7]), amongst many others. Freshwater HAB toxins are finding their way into marine environments and contaminating seafood with poorly understood consequences [8,9,10,11]. Blooms of fish and shellfish killing HABs are occurring in many regions and are especially threatening to aquaculture. While long-term preventive mitigation and management efforts (i.e., nutrient reduction) are optimal [12,13], the problem is formidable, leading to both costly and long-term (years to decades) solutions to address these issues. Blooms threatening public and environmental health may be best mitigated through short-term solutions including immediate control, such as the use of USEPA-registered chemical algaecides (c.f. [14]). However, use of commercially available algaecides can be problematic. Some use copper as an inhibitor [15], many others use oxidants such as hydrogen peroxide [16], and there are numerous regions that restrict application of these products, in part due to the long-lasting legacy of copper-based algaecides and pesticides in aquatic systems [17]. For many other potential algaecides, development and testing can be hampered or even blocked because of permitting requirements under the US Federal Insecticide, Fungicide, and Rodenticide Act (FIFRA) and the National Pollutant Discharge Elimination System (NPDES), leading to exploration of algaecides that are based on natural products and therefore exempt from FIFRA requirements.
So-called natural algaecides have garnered considerable attention given the potential for reduced impacts on non-target organisms, and can exhibit high efficiency, selectivity, and are often considered to be more ecosystem-friendly [18]. The majority of these compounds are derived from bacteria [18,19,20] but there is also rich literature on plant-based compounds, including for application specifically to HAB organisms [21,22,23,24,25,26]. However, a recent meta-analysis of various treatments used in freshwater systems identified only four effective chemicals: copper sulfate, hydrogen peroxide, peracetic acid, and simazine, while the bacterial, physical, or plant-based treatments did not significantly improve water quality [27]. Thus despite testing numerous natural compounds, no single natural algaecide has emerged as a potential treatment that is as cost-effective, scalable, and efficient as traditional treatments despite the desire to find an environmentally friendly alternative to traditional algaecides.
Anjon AG is a research and development company that specializes in organic pesticides for agriculture. The majority of their agricultural products are registration exempt under Sec. 25(b) and 40 CFR 152.25(f) of FIFRA. Based on this experience, the company has formulated an algaecide, C7X1, that uses a variety of FIFRA-exempt ingredients (Table 1) rather than a specific compound. This contribution provides benchtop-scale assessment of C7X1 as a potential natural algaecide which meets the requirements (i.e. effectiveness, scalability, durability, ease of application, streamlined permitting processes) for water resource managers considering short-term mitigation of algal blooms [14].
2. Results
2.1. Exposure Concentrations for Inhibition of Photosynthesis
The algal cultures and field samples were tested using a range of C7X1 concentrations to assess efficacy in suppressing photosynthesis. The concentration of algaecide resulting in 50% inhibition of maximal Fv/Fm (IC50 values) and the experimental treatment where FvFm reached a value of < 0.05 (complete shutdown of photosynthesis) were comparable across algae in benchtop testing, with IC50 values of approximately 400-14,000 ppm C7X1 and complete shutdown of photosynthesis within 24 hours ranging from 4,000-20,000 ppm (Table 2). Testing on water obtained from Pinto Lake, CA provided comparable results. Cultures exposed to 7,000 ppm C7X1 for 24 hours exhibited a decline in chlorophyll ranging from 9.5-85% (Figure 1) and noticeable deformation and loss of pigmentation in some of the cultures (Figure 2).
Pinto Lake water from 28 August 2024 was serially diluted to assess whether there is a biomass effect with the algaecide, and to assess time of exposure to the algaecide. Figure 2 shows a timeseries after treatment with 1000 ppm C7X1. There were significant differences with both group (amount of biomass) and time, as well as the interaction of group and time (ANOVA with Tukey Honestly Significant Difference post-hoc test, p<0.001).
Figure 3 shows an M. aeruginosa culture treated with C7X1 at 0 (control), 625, 1250, 2,500, 5,000, 6,250, and 12,500 ppm and measured sequentially at 0, 1, 22, 25, 48, and 52 hours. The IC50 values steadily decrease with time, ranging from 7,341 ppm (0 hours) to 3,466 ppm (48 hours) ppm C7X1. There also appeared to be a threshold response with a rapid decrease in Fv/Fm above 2,500 ppm. Based on these results, subsequent tests typically used 24-hour exposure for IC50 values (Table 2) for intercomparison of results. Note that at time 0 (immediately after adding C7X1) Fv/Fm dropped to <0.05 at concentrations of 6,250 ppm and higher for Pinto Lake water, demonstrating that C7X1 is capable of nearly instantaneous suppression of photosynthesis at sufficiently high concentrations.
2.2. Stability of the Algaecide
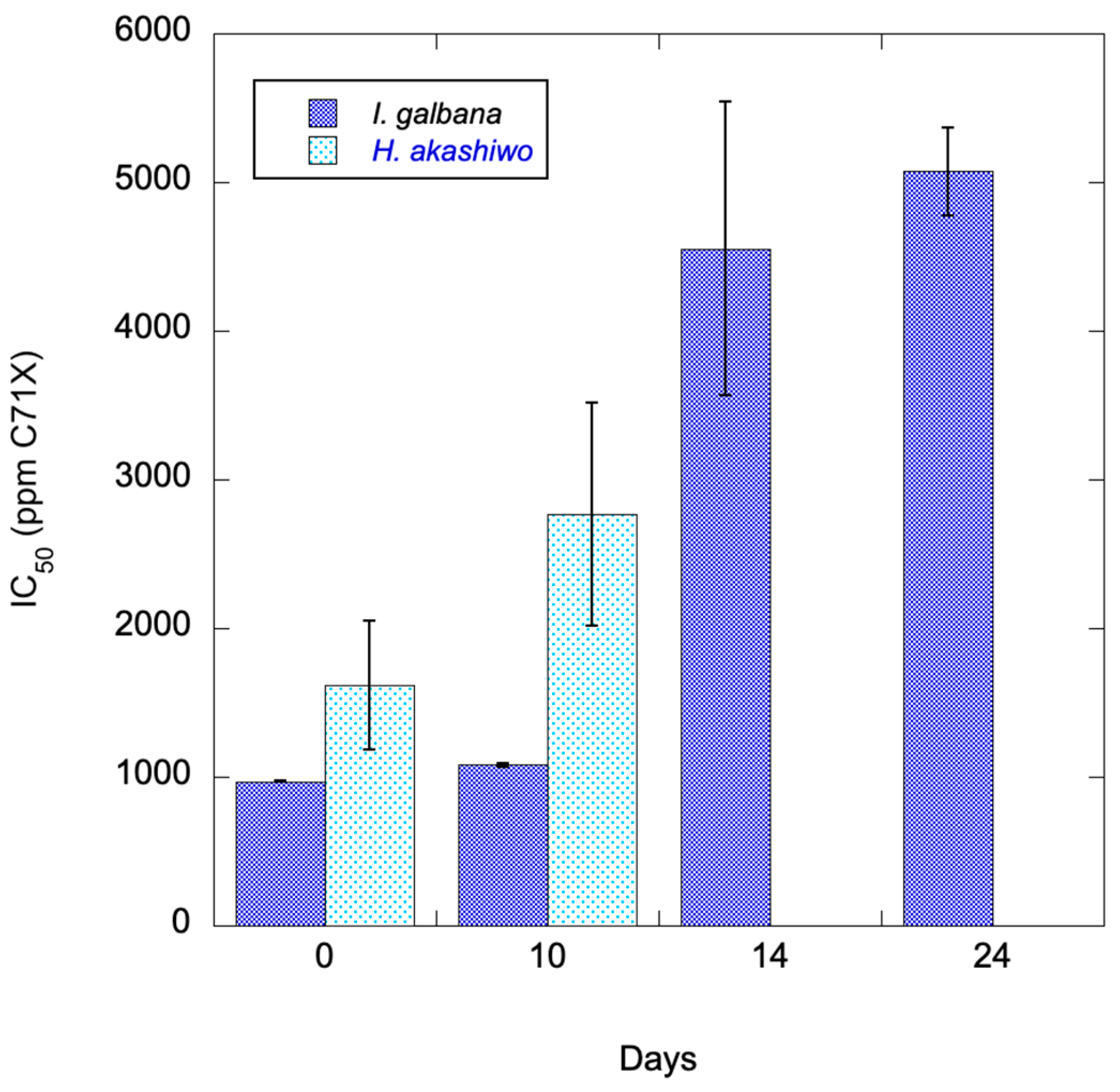
As per manufacturer recommendations, C7X1 was stored at room temperature. Repeated experiments over six months with the same algal strains showed no significant difference (ANOVA, p>0.5) with time. To assess whether it would degrade under simulated environmental conditions, C7X1 was diluted 1:10 with filtered seawater and placed in a UV-transparent quartz glass tube. The tube was left outside for 24 days, and the C7X1 was used for IC50 curves using Isochrysis galbana. At each treatment time, laboratory-stored C7X1 was used as a control. Figure 3 shows the gradual degradation of efficacy, resulting in increasingly higher IC50 values. A similar but truncated experiment was performed using C7X1 diluted 1:10 with 0.2 µm filtered Pinto Lake water, and stored in a quartz tube under ambient conditions. H. akashiwo was treated with the control and UV-treated algaecide at 0, 3, and 10 days. At 3 days there was no difference between UV and non-UV C7X1. At 10 days, the IC50 increased from 1,618 ppm to 2,769 ppm or about double, compared to I. galbana which increased about 10% at the same timepoint.
Figure 4.
IC50 for I. galbana and H. akashiwo exposed to C7X1 that was exposed to natural sunlight in UV-transparent quartz tubes. Error bars are the standard deviation of biological and technical replicates.
Figure 4.
IC50 for I. galbana and H. akashiwo exposed to C7X1 that was exposed to natural sunlight in UV-transparent quartz tubes. Error bars are the standard deviation of biological and technical replicates.
2.3. Vertical Column Experiments
A typical deployment scenario for use of algaecide is to spray it onto the surface of a water column. To assess how C7X1 would perform in that scenario (at lab scale), 0.5 m, 1 L vertical Plexiglas columns were filled with either Pinto Lake water collected 18 August 2024 and 28 September 2024, or H. akashiwo culture, which was chosen as it is a strong vertical migrator.
2.3.1. Pinto Lake, 18 August 2024
For the first experiment, the water was mixed, dispensed into four columns, and allowed to equilibrate for 24 hours in an environmental chamber. Chlorophyll concentration at the time of sampling was 45.4 µg L-1. The IC50 for the whole water was 3,179 ppm (standard error=370). At the start of the experiment, each column had separated into a surface layer, diffuse cells throughout the column, and bottom layer of cells (Figure 5). One column was not treated (Control), the other three were treated at a concentration of the equivalent of 5,000 ppm for the full column volume. Column 2 was mixed before spraying the C7X1 at the top of the column, Column 3 was treated and then mixed, dispersing the C7X1 through the full volume, and Column 4 was sprayed at the surface with no mixing. Initial Fv/Fm was 0.348 (+/- 0.03), consistent with a moderately healthy community dominated by cyanobacteria.
The columns were sampled after 1.5 hours and resampled after 24 and 48 hours at the top, 10 cm, 20 cm, immediately above the biomass at the bottom of the column, and from within the bottom layer (Figure 3). Samples were analyzed for Fv/Fm and rETR curves. At the termination of the experiment, the Control column was treated with 5,000 ppm C7X1 and mixed, replicating Column 3. Columns 2 and 4 were maintained for a total of 7 days in an environmental chamber to assess recovery.
At 1.5 hours the Control Fv/Fm was 0.357, or healthy for a cyanobacterial bloom. The treated columns exhibited depressed Fv/Fm at the surface, Fv/Fm < 0.1, and ETR curves were not plotted since there was essentially no fluorescence signal. Column 3 exhibited no fluorescence (Fv/Fm <0.05) and no additional sampling was conducted. The Control column (replicating Column 3 at the end of the experiment) also showed no variable fluorescence after treating and mixing the column.
Variable fluorescence was negligible in the surface of the treated columns at 24 hours. Photosynthetic performance is plotted as rETR curves in Figure 6 for sampling at 48 hours. Maximum rETR was reduced by about half throughout the column, with progressively more impairment approaching the surface. The mixed and treated column (Column 2) was more uniform, with least impairment in the bottom layer of flocculent material. Column 3, which was not mixed, exhibited a stronger vertical gradient of impairment. This is potentially due to the C7X1 being more evenly distributed in Column 2 as mixed cells sank out, while the C7X1 was presumably more gradually mixed down the column as buoyant cells sank, since the C7X1 was less dense than the Pinto Lake water. At 72 hours all the treated columns were clear of cells except for the bottom flocculant material. The columns remained clear through day 7, when the experiment was terminated.
2.3.2. Pinto Lake, 28 September 2024
The second column experiment was set up in an identical fashion except the treated and mixed column was omitted. Initial Fv/Fm was 0.457 (+/- 0.01). Chlorophyll concentration at the time of sampling was 80.2 µg L-1. The IC50 was 8,832 (SE=2,932). Columns were again treated with 5,000 ppm C7X1. However, except for the Control, all columns immediately cleared of cells, and variable fluorescence dropped to <0.05 through the full vertical extent. The experiment was therefore terminated without additional rETR curves, and the columns were maintained to determine whether there would be recovery. None but the control exhibited positive Fv/Fm values at 7 days. Figure 4D provides the rETR data for the initial sampling before treatment, and at 24 hours for the Control at 24 hours. When the unmixed column was sprayed with C7X1, it was noted that the upper 10 cm immediately developed a blue tint consistent with cell lysis and release of phycocyanin. Given the rapidity of the response and the much lower maximum rETR compared to August, it is likely that the bloom in Pinto Lake was senescing and therefore more susceptible to the algaecide, compared to the August experiment.
2.3.3. Heterosigma Akashiwo Cultures
A third set of column experiments were conducted with H. akashiwo strain EBL 71. Stock cultures were diluted into 2 L cell-free media at a final concentration of 39.4 µg L-1 chlorophyll and dispensed into four columns, which were held for 24 hours in an environmental chamber. Columns were Control (replicates) and treated (replicates) with surface spray and no mixing. Based on IC50 values for H. akashiwo, treatment was reduced to 1,000 ppm C7X1. Initial Fv/Fm of the stock culture indicated that the cells were healthy (Fv/Fm=0.54 +/- 0.02) but below optimal values of ~0.65. At the start of the experiment cells had migrated to the upper third of the columns. Photosynthetic response as rETR curves were measured at the beginning (before treatment) of the experiment and at 24 hours.
The rETR curves (Figure 7) were similar between replicates so data were pooled. At 24 hours, the treated columns exhibited greatly reduced photosynthesis compared to the control columns with fairly uniform reductions (except for one treated column sampled at 20 cm, or the middle of the column, which was even more suppressed). At 48 hours, the control columns were still healthy, but the treated columns were cleared, with all cells on the bottom and no photosynthetic activity.
2.3.4. Pinto Lake Large Volume Columns
The last set of experiments were scaled up to 1.5 m, 20 L volume Plexiglas columns that were maintained in an environmental chamber. The initial chlorophyll was 5.49 µg L-1, and the experiment was designed to mimic pre-bloom conditions. BG11 media at 0.1% of stock concentration (17.6 µM N, 0.23 µM P) was added to all columns. replicate treatment columns were sprayed with the equivalent of 1000 ppm for the 20 L volume at the top of the column. Initial Fv/Fm was 0.432, or healthy for a mixed assemblage with cyanobacteria. At one week, the control column Fv/Fm was still healthy at 0.447, while the treated column was reduced to 0.136 at 20 cm depth. At two weeks, Fv/Fm increased in the control to 0.509 while the treatment increased to 0.247. At one month the experiment was terminated, with Fv/Fm of 0.498 and 0.225 for the control and treated columns. The control columns had a pronounced surface accumulation of algae, dominated by Volvox colonies. The treated columns had a surface layer of senescent and unhealthy Volvox and the water column was dominated by detritus. Final chlorophyll concentrations in the control and treated columns, sampled at 10 cm to avoid the surface mats, was 2.25 (+/- 1.48) and 27.5 (+/- 26.1) µg L-1 chlorophyll.
2.4. Toxin Release
For the M. aeruginosa and P. multiseries cultures, samples from the IC50 experiments were sampled for total and dissolved (released) toxin concentrations for microcystins and domoic acid, respectively. Data for the P. multiseries experiment are presented as percent dissolved toxin release. The control (no C7X1 treatment) samples had 22.0% (+/- 1.5%) dissolved toxin and 885.0 µg L-1 (+/- 55.2 µg L-1) total toxin. Replicates were analyzed after exposure for 24 hours to 500, 1,000, 2,000, 4,000, 5,000, and 10,000 ppm C7X1. Results are presented in Figure 8A. There was no significant difference to the control up to 1,000 ppm C7X1, and 58.9% (+/- 6.9%) release at the highest dose of 10,000 ppm.
For M. aeruginosa, there was negligible dissolve microcystin in the control (0.1%) with 33.2 µg L-1 total microcystin. Replicates were analyzed after exposure for 24 hours to 5,000, 10,000, 20,000, and 25,000 ppm C7X1. Results are presented in Figure 8B. Samples were also analyzed from the column experiment on 28 September 2024 using Pinto Lake water. The control had 23.7 µg L-1 total microcystins, with 16.6% in the dissolved phase. After 24 hours exposure to 5,000 ppm C7X1, dissolved toxins increased to 60.9% and was the same concentration at the top and bottom of the columns. As noted above, there appeared to be release of phycocyanin in the treated columns, consistent with cell lysis after treatment with C7X1 despite the relatively high Fv/Fm value for the control of 0.457 (+/- 0.01).
3. Discussion
3.1. Exposure Concentrations for Inhibition of Photosynthesis
The C7X1 algaecide exhibited a range of IC50 values and lethal doses depending on the algal strain (Table 2). An effective algaecide must be concentrated enough that application to large waterbodies is reasonable. For comparison, Kinley-Baird et al. [14] reported minimum effective exposure concentrations for a broad range of commercially available algaecides as mg L-1 of active ingredient. C7X1 is 20.1% active ingredients (Table 1) with a density of 925 mg L-1, so 1000 ppm (v/v) would be approximately equivalent to 0.19 mg L-1 active ingredients. While the density difference between C7X1 and either fresh or saltwater is enough to form a layer, it is miscible in both and was observed to gradually mix/diffuse downward.
Kinley-Baird et al. [14] assessed dose based on decline in cell densities and chlorophyll a rather than Fv/Fm, but the values reported ranged from 0.51-1.01 mg L-1 for the copper-based algaecides, 11.4 mg L-1 for the peroxide-based algaecides, and 0.139 mg L-1 for endothall acid. Zhou et al. [28] tested Diuron and ethyl 2-methylacetoacetate (EMA) on Microcystis, and identified significant declines in cell biomass at 48 and 72 hours with exposure to 0.75 mg L-1 Diuron and 500 mg L-1 EMA, respectively. C7X1 IC50 values ranged from 0.076-2.57 mg L-1, so within the range of effective dose for commercially available treatments.
An advantage of C7X1 compared to many other compounds is that it is FIFRA-exempt and will not contribute to legacy contamination compared to copper-based treatments. For that reason, there has been considerable interest in identifying plant-based algaecidal compounds. Zhao et al. [23] identified the plant-based alkaloids nefirine and nuciferine at concentrations of 0.25-4.5 mg L-1 to be effective against Microcystis, with a half-maximal effective concentration (EC50) of 0.52 mg L-1. Other natural products have also been considered including mixtures of compounds from Chinese traditional medicines [21]. A combination of compounds from golden thread and areca seed was most effective, at a dose of 0.048% (w/v), or 480 mg L-1. For C7X1, the comparable concentration to achieve a similar reduction, noting that each study used different metrics for impairment, would be 0.19 mg L-1. C7X1 thus provides comparable results to other plant-based compounds, as indicated by reduced photosynthetic performance at similar concentrations with significant reductions in chlorophyll at 24 hours (Figure 1).
3.2. Mode of Action for Inhibition
Pulsed Amplitude Fluorometry has been proposed as a convenient and sensitive metric for evaluating algaecide performance [28] and has been widely used to assess a broad range of compounds as algaecides [29,30,31,32]. Variable fluorescence is generally considered to be an indicator of overall photosynthetic performance, with lower values representing a response to environmental stressors. In general, a decrease in Fv/Fm represents reduced photosynthetic capacity. All the algae showed a decrease in Fv/Fm with exposure to C7X1, consistent with a negative response to the active ingredients that directly impacts photosynthetic performance. Examining Figure 5 and Figure 6, exposure to C7X1 decreased the initial slope (α) as well as an increase in photo-inhibition (β). At the same time, rETRmax decreased compared to the controls. Reduction in rETRmax can result from lowering electron transport chain and Calvin cycle activities [33], suggesting that the algaecide impaired photosystem II (PSII). The simultaneous decline in α and rETRmax and increase in β are all consistent with general impairment of the photosystem, likely at multiple points in the photosynthetic process.
Visual examination of cells by microscopy (Figure 2) showed distortion of shape and bleaching or a reduction in pigments, but cells remained intact. Reduction in chlorophyll over 24 hours suggests that pigments were being destroyed or catabolized within the cell and is consistent with the changes in α and β. All of these results are consistent with C7X1 damaging the photosynthetic apparatus and likely resulting in cellular oxidative stress [34], similar to other plant-derived natural products tested as algaecides [23].
3.3. Extent of Toxin Release After Exposure to C7X1
Minimizing algal toxin release is a potential priority in management actions where blooms are near drinking water intake sites or there is potential for use of contaminated water in agricultural applications [35,36,37]. Much less is known about the potential threat of dissolved domoic acid in marine and estuarine environments, but studies show that it can be ubiquitous in estuarine and coastal marine environments and is likely related to low-level contamination of bivalves [9,38].
While human and environmental health risks are more clearly associated with total toxin load rather than dissolved toxin [14], the ability to clear the water column, as shown in the vertical column experiments, and sequester biomass with minimal release of toxins is preferential for an effective algaecide [39]. Benchtop testing with M. aeruginosa and P. multiseries demonstrates that there is low to moderate toxin release at concentrations of C7X1 at the IC50 treatment level, and that very high concentrations are required to approach full release (Figure 8).
Column experiments in August and September resulted in flocculation of the cells and sequestration of toxin to the bottom of the column in the first set of experiments (August), but considerable release with uniform distribution of dissolved microcystins in the second set of experiments (September). While this is likely related to the overall health of the assemblages in August and September, more testing at mesocosm or contained field trial scales is warranted to determine whether C7X1 can both clear the water column and sequester cells and toxins at depth to develop management strategies that both visibly mitigate algal blooms and protect the public from inadvertent exposure to dissolved toxins [39].
3.4. From Bench Scale to Environmental Testing
Laboratory results using cultures and Pinto Lake water at up to 20 L volumes suggest that C7X1 is a promising algaecide that is comparable to both commercially available and compound-specific formulations. If C7X1 were to move from bench scale testing to environmental application, these results provide some guidelines for use and further testing. The column experiments demonstrate that C7X1 will penetrate vertically without mixing, making the algaecide amenable to surface spray application. The efficacy of C7X1 is both biomass-dependent (Figure 3) and organism dependent (Table 2), such that some basic knowledge of the system where it is applied would be beneficial. The 20 L column experiment, while not exposed to UV, did demonstrate the ability to suppress bloom conditions after one month, with an approximately 10-fold reduction in surface biomass relative to the control and no change in community composition, but with an increase in detritus and senescent algae. C7X1 has a reasonably long decay time when tested with UV exposure and is shelf-stable when not exposed to UV, suggesting that repeat application would be required on timescales of weeks. While off-target effects were not addressed in this study, qualitative examination of the plankton community from the Pinto Lake experiments suggest that at doses near the IC50 range, zooplankton activity and abundance were not reduced.
4. Conclusions
The overall objective of this study was to provide preliminary data on the efficacy of the algaecide C7X1 with bench scale testing, prior to demonstration-scale and full application of the algaecide. The C7X1 formulation is effective against a wide range of algae, results in minimal toxin release, and at bench scale, is appropriate at high concentrations for immediate suppression of blooms, or at lower concentrations for longer-term suppression of algae.
The frequency and intensity of harmful algal blooms is increasing both within the United States and globally, increasingly requiring management strategies that include active mitigation [4]. C7X1 provides a “green” alternative to other USEPA-registered algaecides and could potentially be used in waterbodies with streamlined permitting since all ingredients are exempt under FIFRA. While many other algaecide studies have focused on freshwater HABs such as Microcystis, the applicability of C7X1 to freshwater, estuarine, and marine HABs highlights the potential for broad application if logistical and management challenges that emerge at scale can be overcome.
5. Materials and Methods
5.1. Algal Strains and Field Sampling
Testing of C7X1 was performed using both monospecific algal cultures (not axenic; Table 2) and water collected from Pinto Lake, CA. Pinto Lake is a shallow natural lake located 8.3 km inland from Monterey Bay, and is well-characterized through an ongoing ~weekly time-series occupied from 2009-present [9,40]. Whole water was collected 18 August 2024, 28 September 2024, and 28 January 2025. See Appendix A for additional environmental data.
For each collection, surface (<1m) water was collected in a 20L polycarbonate carboy and transported to the lab where it was maintained at ~100 µmol photons m-2 s-1 light (12:12 light:dark) and 15°C in an environmental chamber. Samples were inspected visually using a Leica MZ 12.5 dissecting scope equipped with dark field, and community composition was quantified using a relative abundance scale: Rare (R): <1%; Present (P): 1 - <10%; Common (C): 10 - <25%; Abundant (A): 25-<50%. The assemblages were Dolichospermum (A) and Ceratium (A) for July, Microcystis (A) with low levels (P) of Dolichospermum and Aphanizomenon in August, Microcystis (A) with moderate levels (C) of Dolichospermum and low levels (P) of Aphanizomenon and Ceratium in September, and Aphanizomenon, Microcystis (C) and Ceratium (R) with bacteria and detritus making up the rest of the assemblage. Chlorophyll levels were 16.3, 59.6, 78.8, and 4.5 µg L-1 respectively.
Cultures used in this study included Microcystis aeruginosa, Gloeocapsa alpicola, Aphanizomenon flos-aquae, Heterosigma akashiwo, Pseudo-nitzschia multiseries, Isochrysis galbana, and Chattonella spp. (Table 3). Freshwater strains were maintained at room temperature (~20°C) and ambient light on either Bold 3N or BG-11 obtained from University of Texas Culture Culture Collection. Estuarine species were maintained on Guillard’s f/2 or L1 media (32 PSU for all but H. akashiwo, which was at 24 PSU) at 15°C or 20°C with 150 µmol photons m-2 s-1 light and 12:12 light:dark in environmental chambers. All experiments were conducted with algae in exponential or early stationary growth phase. A subset of cultures were documented with and without exposure to C7X1 at doses sufficient to inhibit photosynthesis using a Zeiss Axio Imager A.1 microscope equipped with a Teledyne Lumenera color camera.
5.2. Experimental Treatments
Algal biomass was asssessed using relative fluorescence units (RFU) on a Turner Designs 10AU fluorometer, with RFU converted to equivalent chlorophyll using calibrations between RFU and acetone-extracted chlorophyll measurements [41]. Algal samples were diluted to approximately 50 µg L-1 chlorophyll with 0.2 µm filtered base water (media or Pinto Lake water) except for one test with Pinto Lake water collected 18 August 2024 where the water was serially diluted 59.6 µg L-1 to 29.8, 14.9, and 7.5 µg L-1 to determine if there was a biomass effect with exposure to algaecide. The C7X1 algaecide was provided by Anjon AG. For exposure to algaecide, 5 mL water was subsampled into 10 mL borosilicate glass tubes and C7X1 was added at varying concentrations, with all concentrations reported as ppm, or µL L-1 (v/v). The primary metric for assessment was change in variable fluorescence (Fv/Fm) after 24 hour exposure, but a subset of experiments were sampled at both shorter (hours) and longer (days) timepoints to assess the temporal impact of the algaecide. Treatments were conducted in replicate or triplicate and control samples (no algaecide) were included for each experiment. Algaecide was added with or without 10-fold dilution using base water so that no more than 5% of the 5 mL test volume was added for each treatment. Treated samples were maintained under the same conditions as the Pinto Lake water or cultures for up to 7 days. The cultures were also exposed to 7,000 ppm C7X1 for 24 hours and then analyzed for extracted chlorophyll content to determine if the algaecide resulted in decreased chlorophyll.
For Pinto Lake water, additional experiments were conducted in 0.5 m (~1 L) and 1.5 m (~20 L) columns to assess the ability of C7X1 to penetrate a vertical column and suppress surface aggregation. Pinto Lake water was added to the columns, allowed to equilibrate for 24 hours in an enviromental chamber, and then exposed to C7X1 using a spray bottle to simulate typical deployment conditions where the algaecide is sprayed onto the surface of a bloom. One set of experiments (0.5 m colums) also tested the effect of adding the C7X1 at the surface and then immediately mixing the full column, which essentially replicates the 5 mL benchtop testing at scale. The 0.5 m columns were kept in environmental chambers for up to five days of testing while the 1.5 m columns, which used Pinto Lake water collected 28 January 2025, were maintained for 4 weeks. The columns were sampled at varying depths by inserting a glass tube sealed at the top end so that water could be collected from specific depths without disturbing the vertical structure of the column. Sampling took place by removing and replacing the seal at the top to collect a small volume of water at depth. Replicates (control and treatment) were conducted sequentially for logistical reasons.
5.3. Assessment of Variable Fluorescence and Electron Transport Rates
The primary metric for assessing the efficacy of C7X1 was through measurement of variable fluorescence (Fv/Fm) and relative Electron Transport Rates (rETR), which provide a photosynthesis vs. irradiance curve for a given sample. Discrete samples were analyzed using a Heinz-Walz WATER-PAM [42]. The instrument was blanked with 0.2 μm filtered base water. Discrete samples were dark-adapted for 30 min, and then the gain was adjusted on the WATER-PAM. Subsequent samples used the same gain setting, with three technical replicates per sample. The first PAM reading provides variable fluorescence, while subsequent measurements are subjected to increasing irradiance to calculate the relative Electron Transport Rate (rETR) response curve, functionally equivalent to a photosynthesis vs. irradiance (PvsE) curve. The data were fit to the 4-parameter curve proposed by Platt et al. [43]. The curve fit provides rETRmax (μmol photons m−2 s−1), or the maximum electron transport rate, α, the initial slope of the curve (μmol electrons m− 2 s− 1/μmol photons m− 2 s− 1), Ek (μmol photons m−2 s−1), the irradiance at half-saturation, and β (μmol electrons m− 2 s− 1/μmol photons m− 2 s− 1, a photo-inhibition term.
5.4. Inhibitory Concentration Curves and Statistics
The Fv/Fm data were used to calculate the half-maximal inhibitory concentration (IC50) at 24 hours, and the dose at which photosynthesis was completely inhibited (Fv/Fm < 0.05). IC50 values were determined in Kaleidagraph v. 5.01 (Synergy Software) using a modified dose-response curve:
where x=concentration of C7X1, y=the Fv/Fm value, a=the minimum response value, b=the range of transition (ymax – ymin), c=IC50, and d=the slope of the fit. Error estimates are the standard error of the model fit for each variable. All fits had r2 values > 0.9.
y=a+b/(1+(x/c)^d)
Eukaryotic algae generally exhibit ymax value of ~0.65 (healthy) to 0 (no photosynthetic response). Cyanobacteria typically exhibit lower ymax values (~0.3-0.4) for healthy cells when using 660 nm red excitation light on the WATER-PAM [44]. Repeated IC50 values at multiple time points were used to assess degradation of C7X1 (loss of efficacy) after exposure to ambient outdoor environmental conditions in a sealed quartz tube for up to 24 days, allowing penetration of UV light into the stock solution and natural oscillations in irradiance and temperature. Experiments were conducted January 2025 in Santa Cruz, CA, USA.
Comparison of differences between groups were conducted in Kaleidagraph using one-way and two-way ANOVA, with Tukey Honestly Significant Difference post hoc test. Significance was set at a value of 0.05.
5.5. Toxin Testing
For the Microcystis and Pseudo-nitzschia cultures, samples from the IC50 experiments were analyzed at 24 hours for release of toxins, while the 0.5 m column experiments with Pinto Lake water were tested at 24 hours from the surface and at the bottom of the columns for control and treatment columns. Approximately 1 mL of whole water was collected for total (cellular and dissolved) toxins, and a second 1 mL sample was syringe filtered under low pressure using a 13 mm PTFE syringe filter. The filtrate was analyzed for dissolved toxin. Domoic acid and microcystin LR, LA, YR, and RR were analyzed on an Agilent 6150 LC/MS with JetSpray Technology and Select Ion Monitoring using an external calibration curve with pure standards obtained from NRC Canada [9]. Since the primary interest for this analysis was release of toxins upon exposure to C7X1, concentrations are reported as percent dissolved toxin relative to the control samples.
Funding
This research received no external funding.
Data Availability Statement
The original contributions presented in this study are included in the article. Further inquiries can be directed to the corresponding author.
Acknowledgments
The C7X1 algicide was provided as an in-kind donation by Robert Breeze, Anjon AG. Kendra Negrey provided technical support and field sampling for this project, and Eleanor Kudela participated in laboratory data collection and experimental setup.
Conflicts of Interest
The author declares no conflicts of interest.
Appendix A
Appendix A.1. Field sampling
Water was collected from Pinto Lake, CA, USA (36.95° N, 121.77° W) as part of this analysis. Pinto Lake is a shallow spring-fed natural lake 8.3 km inland from Monterey Bay and covers 37 surface hectares. Approximately weekly sampling is conducted from the boat dock on the south end of the lake. Relevant environmental data bracketing the collection dates are provided in Table A1.

Table A1.
Environmental conditions at Pinto Lake. Community Composition uses a relative abundance index: Rare (R): <1%; Present (P): 1 - <10%; Common (C): 10 - <25%; Abundant (A): 25-<50%.
Table A1.
Environmental conditions at Pinto Lake. Community Composition uses a relative abundance index: Rare (R): <1%; Present (P): 1 - <10%; Common (C): 10 - <25%; Abundant (A): 25-<50%.
| Date | Temperature (°C) | Chlorophyll (µg L-1) | Community Composition |
|---|---|---|---|
| 7/31/24 | 24.3 | 900.6 | A - Dolichospermum, R - Microcystis, Aphanizomenon, Straustrum, Cymbella |
| 8/6/24 | 24.7 | 546.9 | A - Microcystis, R - Aphanizomenon, Dolichospermum, Straustrum, diatom (Leptocylindrus-like) |
| 8/13/24 | 24 | 55.8 | A - Microcystis, R - Aphanizomenon, Dolichospermum, Straustrum, Ceratium |
| 8/20/24 | 24.3 | 59.6 | A - Microcystis, R - Aphanizomenon, Dolichospermum, Oscillatoria, Ceratium |
| 8/27/24 | 23.7 | 45.4 | A - Microcystis, R - Aphanizomenon, Dolichospermum, Straustrum |
| 9/3/24 | 23.7 | 78.3 | A - Microcystis, P - Dolichospermum, R - Aphanizomenon, Ceratium |
| 9/17/24 | 22.1 | 77.7 | A - Microcystis, C - Dolichospermum, R - Aphanizomenon, Ceratium |
| 9/24/24 | 22.3 | 85.9 | A - Microcystis, P - Dolichospermum, R - Aphanizomenon, Pediastrum |
| 10/1/24 | 21.9 | 74.9 | A - Microcystis, R - Dolichospermum, Pediastrum |
| 10/15/24 | 21.4 | 120.4 | A - Microcystis, R - Dolichospermum, Oscillatoria, Ceratium, diatom (Leptocylindrus-like) |
| 10/22/24 | 18.9 | 957.1 | A - Microcystis, R - Dolichospermum, Aphanizomenon, Oscillatoria, flagellate |
| 11/5/24 | 16.9 | 2446.3 | A - Microcystis, R - Aphanizomenon, Ceratium |
| 11/12/24 | 15.9 | 188.2 | A - Microcystis |
| 11/25/24 | 14.4 | 174.0 | A - Microcystis, R - Ceratium |
| 12/3/24 | 12.8 | 1340.3 | A - Microcystis, R - Aphanizomenon |
| 12/10/24 | 11.7 | 372.4 | A - Microcystis, R - Aphanizomenon |
| 12/17/24 | 12.2 | 1360.4 | A - Microcystis, R - Aphanizomenon |
| 1/7/25 | 12 | 12.6 | C - Aphanizomenon, Microcystis, R - Ceratium, flagellate |
| 1/14/25 | 10.6 | 6.6 | C - Aphanizomenon, Microcystis, R - Ceratium, flagellate, tons of sparkles/bacteria |
| 1/21/25 | 10.3 | 5.5 | C - Aphanizomenon, Microcystis, R - Ceratium |
| 2/5/25 | 12.9 | 3.6 | A - Microcystis, P - Aphanizomenon, R - flagellate |
| 2/11/25 | 11.3 | 2.1 | A - Microcystis, R - flagellate |
| 2/18/25 | 12.5 | 1.7 | A - Microcystis, R - Ceratium |
| 2/25/25 | 14.8 | 2.1 | C - Microcystis, Ceratium, Coelastrum, tons of sparkles/bacteria |
| 3/4/25 | 14.3 | 2.8 | C - Microcystis, Ceratium, diatom (Leptocylindrus-like), tons of sparkles/bacteria, |
| 3/18/25 | 14.5 | 6.7 | A - Microcystis, R - Ceratium |
| 3/25/25 | 18.5 | 6.9 | C - Microcystis, Aphanizomenon, flagellate, R - Ceratium, Coelastrum, diatom (Leptocylindrus-like) |
References
- Feng, L.; Wang, Y.; Hou, X.; Qin, B.; Kutser, T.; Qu, F.; Chen, N.; Paerl, H.W.; Zheng, C. Harmful Algal Blooms in Inland Waters. Nat Rev Earth Environ 2024, 5, 631–644. [Google Scholar] [CrossRef]
- Anderson, D.M.; Fensin, E.; Gobler, C.J.; Hoeglund, A.E.; Hubbard, K.A.; Kulis, D.M.; Landsberg, J.H.; Lefebvre, K.A.; Provoost, P.; Richlen, M.L.; et al. Marine Harmful Algal Blooms (HABs) in the United States: History, Current Status and Future Trends. Harmful Algae 2021, 102, 101975. [Google Scholar] [CrossRef]
- Enevoldsen, H.O.; Isensee, K.; Lee, Y.J. State of the Ocean Report 2024. UNESCO-IOC, 2024. [Google Scholar]
- Woods Hole Oceanographic Institution; Anderson, D.M.; Backer, L.C.; Bouma-Gregson, K.; Bowers, H.A.; Bricelj, V.M.; D’Anglada, L.; Deeds, J.; Dortch, Q.; Doucette, G.J.; et al. Harmful Algal Research & Response: A National Environmental Science Strategy (HARRNESS), 2024-2034; Woods Hole Oceanographic Institution, 2024. [Google Scholar]
- Trainer, V.L.; Bates, S.S.; Lundholm, N.; Thessen, A.E.; Cochlan, W.P.; Adams, N.G.; Trick, C.G. Pseudo-Nitzschia Physiological Ecology, Phylogeny, Toxicity, Monitoring and Impacts on Ecosystem Health. Harmful Algae 2012, 14, 271–300. [Google Scholar] [CrossRef]
- Harke, M.J.; Steffen, M.M.; Gobler, C.J.; Otten, T.G.; Wilhelm, S.W.; Wood, S.A.; Paerl, H.W. A Review of the Global Ecology, Genomics, and Biogeography of the Toxic Cyanobacterium, Microcystis Spp. Harmful Algae 2016, 54, 4–20. [Google Scholar] [CrossRef] [PubMed]
- Mehdizadeh Allaf, M. Heterosigma Akashiwo, a Fish-Killing Flagellate. Microbiology Research 2023, 14, 132–147. [Google Scholar] [CrossRef]
- Ibelings, B.W.; Chorus, I. Accumulation of Cyanobacterial Toxins in Freshwater “Seafood” and Its Consequences for Public Health: A Review. Environmental Pollution 2007, 150, 177–192. [Google Scholar] [CrossRef]
- Peacock, M.B.; Gibble, C.M.; Senn, D.B.; Cloern, J.E.; Kudela, R.M. Blurred Lines: Multiple Freshwater and Marine Algal Toxins at the Land-Sea Interface of San Francisco Bay, California. Harmful Algae 2018, 73, 138–147. [Google Scholar] [CrossRef]
- Miller, M.A.; Kudela, R.M.; Mekebri, A.; Crane, D.; Oates, S.C.; Tinker, M.T.; Staedler, M.; Miller, W.A.; Toy-Choutka, S.; Dominik, C.; et al. Evidence for a Novel Marine Harmful Algal Bloom: Cyanotoxin (Microcystin) Transfer from Land to Sea Otters. PLoS ONE 2010, 5, e12576. [Google Scholar] [CrossRef]
- Gibble, C.M.; Peacock, M.B.; Kudela, R.M. Evidence of Freshwater Algal Toxins in Marine Shellfish: Implications for Human and Aquatic Health. Harmful Algae 2016, 59, 59–66. [Google Scholar] [CrossRef]
- Glibert, P.M.; Al-Azri, A.; Icarus Allen, J.; Bouwman, A.F.; Beusen, A.H.W.; Burford, M.A.; Harrison, P.J.; Zhou, M. Key Questions and Recent Research Advances on Harmful Algal Blooms in Relation to Nutrients and Eutrophication. In Global Ecology and Oceanography of Harmful Algal Blooms; Glibert, P.M., Berdalet, E., Burford, M.A., Pitcher, G.C., Zhou, M., Eds.; Ecological Studies; Springer International Publishing: Cham, 2018; Vol. 232, pp. 229–259. ISBN 978-3-319-70068-7. [Google Scholar]
- Paerl, H.W.; Otten, T.G.; Kudela, R. Mitigating the Expansion of Harmful Algal Blooms Across the Freshwater-to-Marine Continuum. Environ. Sci. Technol. 2018, 52, 5519–5529. [Google Scholar] [CrossRef]
- Kinley-Baird, C.; Calomeni, A.; Berthold, D.E.; Lefler, F.W.; Barbosa, M.; Rodgers, J.H.; Laughinghouse, H.D. Laboratory-Scale Evaluation of Algaecide Effectiveness for Control of Microcystin-Producing Cyanobacteria from Lake Okeechobee, Florida (USA). Ecotoxicology and Environmental Safety 2021, 207, 111233. [Google Scholar] [CrossRef] [PubMed]
- Hennessey, A.V.; McDonald, M.B.; Johnson, P.P.; Gladfelter, M.F.; Merrill, K.L.; Tenison, S.E.; Ganegoda, S.S.; Hoang, T.C.; Torbert, H.A.; Beck, B.H.; et al. Evaluating the Tolerance of Harmful Algal Bloom Communities to Copper. Environmental Pollution 2025, 368, 125691. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Andrés, J.; Romero-Martínez, L.; Seoane, S.; Acevedo-Merino, A.; Moreno-Garrido, I.; Nebot, E. Evaluation of Algaecide Effectiveness of Five Different Oxidants Applied on Harmful Phytoplankton. Journal of Hazardous Materials 2023, 452, 131279. [Google Scholar] [CrossRef] [PubMed]
- Kiaune, L.; Singhasemanon, N. Pesticidal Copper (I) Oxide: Environmental Fate and Aquatic Toxicity. In Reviews of Environmental Contamination and Toxicology Volume 213; Whitacre,, D.M., Ed.; Reviews of Environmental Contamination and Toxicology; Springer New York: New York, NY, 2011; Vol. 213, pp. 1–26. ISBN 978-1-4419-9859-0. [Google Scholar]
- Li, H.; Xing, R.; Ji, X.; Liu, Y.; Chu, X.; Gu, J.; Wang, S.; Wang, G.; Zhao, S.; Cao, X. Natural Algicidal Compounds: Strategies for Controlling Harmful Algae and Application. Plant Physiology and Biochemistry 2024, 215, 108981. [Google Scholar] [CrossRef]
- Coyne, K.J.; Wang, Y.; Johnson, G. Algicidal Bacteria: A Review of Current Knowledge and Applications to Control Harmful Algal Blooms. Front. Microbiol. 2022, 13, 871177. [Google Scholar] [CrossRef]
- Yang, S.; Williams, S.J.; Courtney, M.; Burchill, L. Warfare under the Waves: A Review of Bacteria-Derived Algaecidal Natural Products. Nat. Prod. Rep. 2025, 10.1039.D4NP00038B. [Google Scholar] [CrossRef]
- Zhou, L.H.; Zheng, T.L.; Wang, X.; Ye, J.L.; Tian, Y.; Hong, H.S. Effect of Five Chinese Traditional Medicines on the Biological Activity of a Red-Tide Causing Alga—Alexandrium Tamarense. Harmful Algae 2007, 6, 354–360. [Google Scholar] [CrossRef]
- Liu, F.; He, Z.-B.; Li, H.-Y.; Liu, J.-S.; Yang, W.-D. Inhibition of Five Natural Products from Chinese Herbs on the Growth of Chattonella Marina. Environ Sci Pollut Res 2016, 23, 17793–17800. [Google Scholar] [CrossRef]
- Zhao, G.; Hong, Y.; Li, L.; Zhang, H.; Xu, R.; Hao, Y. Selection and Characterization of Plant-Derived Alkaloids with Strong Antialgal Inhibition: Growth Inhibition Selectivity and Inhibitory Mechanism. Harmful Algae 2022, 117, 102272. [Google Scholar] [CrossRef]
- Zhu, X.; Dao, G.; Tao, Y.; Zhan, X.; Hu, H. A Review on Control of Harmful Algal Blooms by Plant-Derived Allelochemicals. Journal of Hazardous Materials 2021, 401, 123403. [Google Scholar] [CrossRef]
- Lan, Y.; Chen, Q.; Gou, T.; Sun, K.; Zhang, J.; Sun, D.; Duan, S. Algicidal Activity of Cyperus Rotundus Aqueous Extracts Reflected by Photosynthetic Efficiency and Cell Integrity of Harmful Algae Phaeocystis Globosa. Water 2020, 12, 3256. [Google Scholar] [CrossRef]
- Hu, J.; Kokoette, E.; Xu, C.; Huang, S.; Tang, T.; Zhang, Y.; Liu, M.; Huang, Y.; Yu, S.; Zhu, J.; et al. Natural Algaecide Sphingosines Identified in Hybrid Straw Decomposition Driven by White-Rot Fungi. Advanced Science 2023, 10, 2300569. [Google Scholar] [CrossRef] [PubMed]
- Anantapantula, S.S.; Wilson, A.E. Most Treatments to Control Freshwater Algal Blooms Are Not Effective: Meta-Analysis of Field Experiments. Water Research 2023, 243, 120342. [Google Scholar] [CrossRef]
- Zhou, S.; Shao, Y.; Gao, N.; Deng, Y.; Qiao, J.; Ou, H.; Deng, J. Effects of Different Algaecides on the Photosynthetic Capacity, Cell Integrity and Microcystin-LR Release of Microcystis Aeruginosa. Science of The Total Environment 2013, 463–464, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Hossain, I.; Mays, B.; Hanhart, S.L.; Hubble, J.; Azizihariri, P.; McLean, T.I.; Pierce, R.; Lovko, V.; John, V.T. An Effective Algaecide for the Targeted Destruction of Karenia Brevis. Harmful Algae 2024, 138, 102707. [Google Scholar] [CrossRef]
- Guo, P.; Liu, Y.; Liu, C. Effects of Chitosan, Gallic Acid, and Algicide on the Physiological and Biochemical Properties of Microcystis Flos-Aquae. Environ Sci Pollut Res 2015, 22, 13514–13521. [Google Scholar] [CrossRef]
- Mehdizadeh Allaf, M.; Erratt, K.J.; Peerhossaini, H. Comparative Assessment of Algaecide Performance on Freshwater Phytoplankton: Understanding Differential Sensitivities to Frame Cyanobacteria Management. Water Research 2023, 234, 119811. [Google Scholar] [CrossRef]
- Lürling, M.; Meng, D.; Faassen, E. Effects of Hydrogen Peroxide and Ultrasound on Biomass Reduction and Toxin Release in the Cyanobacterium, Microcystis Aeruginosa. Toxins 2014, 6, 3260–3280. [Google Scholar] [CrossRef]
- Ralph, P.J.; Gademann, R. Rapid Light Curves: A Powerful Tool to Assess Photosynthetic Activity. Aquatic Botany 2005, 82, 222–237. [Google Scholar] [CrossRef]
- Nishiyama, Y. Oxidative Stress Inhibits the Repair of Photodamage to the Photosynthetic Machinery. The EMBO Journal 2001, 20, 5587–5594. [Google Scholar] [CrossRef]
- Bittencourt-Oliveira, M.D.C.; Cordeiro-Araújo, M.K.; Chia, M.A.; Arruda-Neto, J.D.D.T.; Oliveira, Ê.T.D.; Santos, F.D. Lettuce Irrigated with Contaminated Water: Photosynthetic Effects, Antioxidative Response and Bioaccumulation of Microcystin Congeners. Ecotoxicology and Environmental Safety 2016, 128, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Cordeiro-Araújo, M.K.; Chia, M.A.; Arruda-Neto, J.D.D.T.; Tornisielo, V.L.; Vilca, F.Z.; Bittencourt-Oliveira, M.D.C. Microcystin-LR Bioaccumulation and Depuration Kinetics in Lettuce and Arugula: Human Health Risk Assessment. Science of The Total Environment 2016, 566–567, 1379–1386. [Google Scholar] [CrossRef] [PubMed]
- Melaram, R.; Newton, A.R.; Chafin, J. Microcystin Contamination and Toxicity: Implications for Agriculture and Public Health. Toxins 2022, 14, 350. [Google Scholar] [CrossRef] [PubMed]
- Trapp, A.; Hayashi, K.; Fiechter, J.; Kudela, R.M. What Happens in the Shadows - Influence of Seasonal and Non-Seasonal Dynamics on Domoic Acid Monitoring in the Monterey Bay Upwelling Shadow. Harmful Algae 2023, 129, 102522. [Google Scholar] [CrossRef]
- Greenfield, D.I.; Duquette, A.; Goodson, A.; Keppler, C.J.; Williams, S.H.; Brock, L.M.; Stackley, K.D.; White, D.; Wilde, S.B. The Effects of Three Chemical Algaecides on Cell Numbers and Toxin Content of the Cyanobacteria Microcystis Aeruginosa and Anabaenopsis Sp. Environmental Management 2014, 54, 1110–1120. [Google Scholar] [CrossRef]
- Kudela, R.M. Characterization and Deployment of Solid Phase Adsorption Toxin Tracking (SPATT) Resin for Monitoring of Microcystins in Fresh and Saltwater. Harmful Algae 2011, 11, 117–125. [Google Scholar] [CrossRef]
- Welschmeyer, N.A. Fluorometric Analysis of Chlorophyll a in the Presence of Chlorophyll b and Pheopigments. Limnology & Oceanography 1994, 39, 1985–1992. [Google Scholar] [CrossRef]
- Schreiber, U. Pulse-Amplitude-Modulation (PAM) Fluorometry and Saturation Pulse Method: An Overview. Papageorgiou, G.C., Govindjee, Eds.; In Chlorophyll a Fluorescence; Advances in Photosynthesis and Respiration; Springer Netherlands: Dordrecht, 2004; Vol. 19, pp. 279–319. ISBN 978-1-4020-3217-2. [Google Scholar]
- Platt, T, T.; Gallegos, C.; Harrison, W. Photoinhibition of Photosynthesis in Natural Assemblages of Marine Phytoplankton. Journal of Marine Research 1980, 38, 687–701.
- Santabarbara, S.; Villafiorita Monteleone, F.; Remelli, W.; Rizzo, F.; Menin, B.; Casazza, A.P. Comparative Excitation-emission Dependence of the FV / FM Ratio in Model Green Algae and Cyanobacterial Strains. Physiologia Plantarum 2019, 166, 351–364. [Google Scholar] [CrossRef]
Figure 1.
Percent decrease in chlorophyll after exposure to C7X1 at 7,000 ppm for 24 hours. Error bars are standard deviations from biological replicates.
Figure 1.
Percent decrease in chlorophyll after exposure to C7X1 at 7,000 ppm for 24 hours. Error bars are standard deviations from biological replicates.
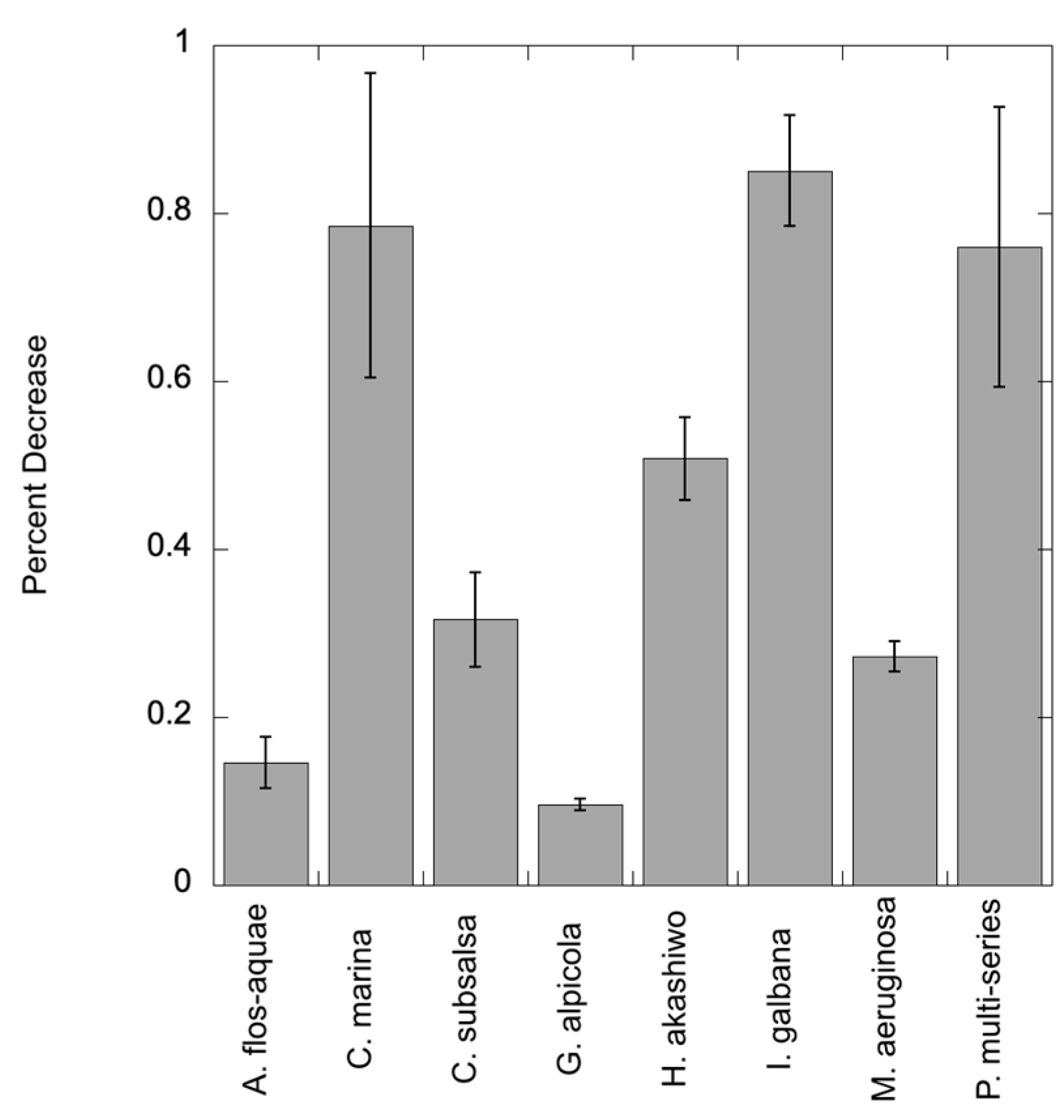
Figure 2.
Micrographs of representative algae exposed to 7,000 ppm C7X1 for 24 hours. A: M. aeruginosa control; B: H. akashiwo control; C: C. marina control. D: M. aeruginosa treated; E: H. akashiwo treated; F: C. marina treated. Each pair of micrographs were imaged at the same magnification and illumination.
Figure 2.
Micrographs of representative algae exposed to 7,000 ppm C7X1 for 24 hours. A: M. aeruginosa control; B: H. akashiwo control; C: C. marina control. D: M. aeruginosa treated; E: H. akashiwo treated; F: C. marina treated. Each pair of micrographs were imaged at the same magnification and illumination.
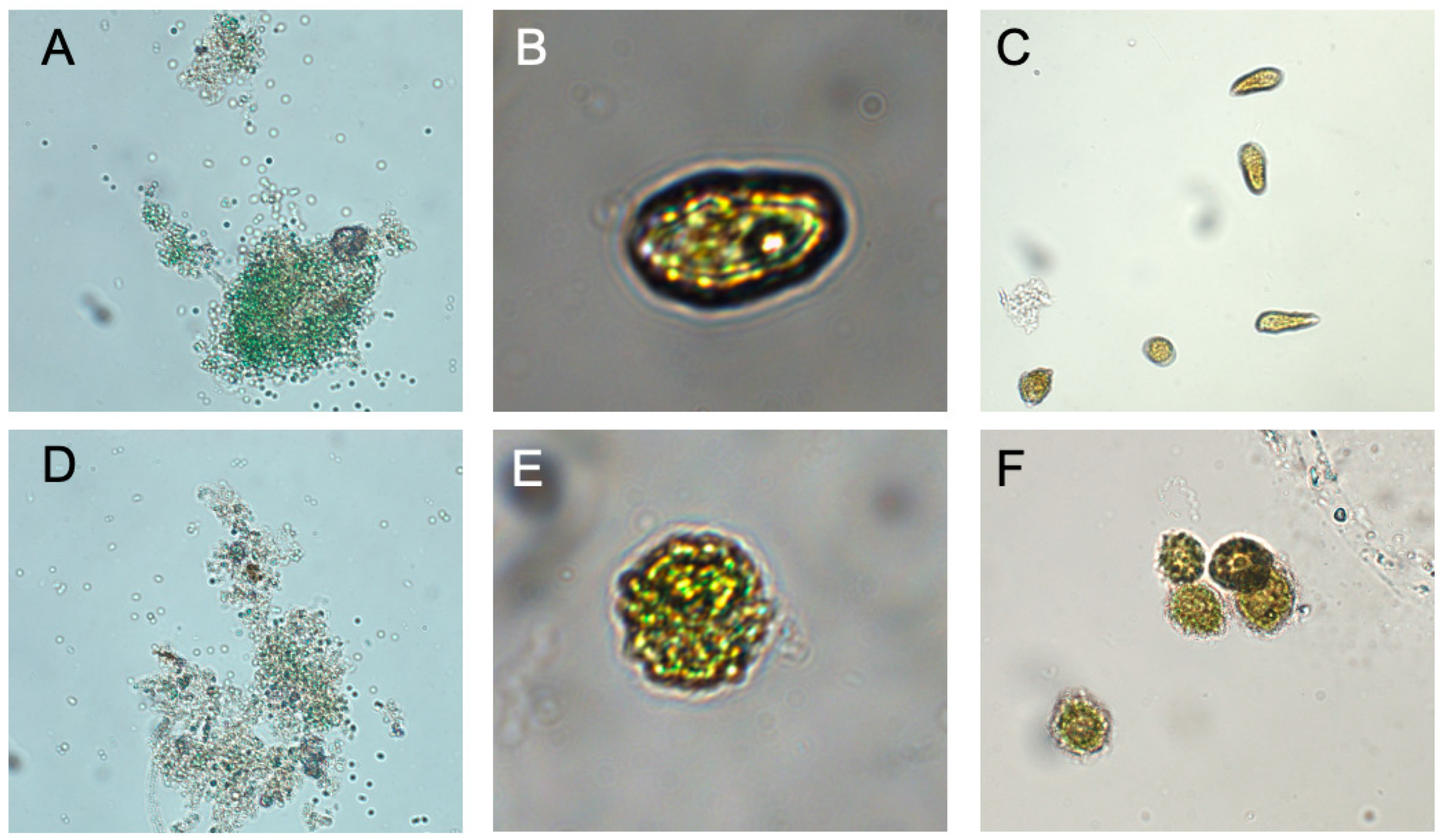
Figure 3.
Variable fluorescence (Fv/Fm) from Pinto Lake that was serially diluted and exposed to 1000 ppm C7X1, with sampling conducted from 0 to 72 hours. Error bars are the standard deviation of biological and technical replicates.
Figure 3.
Variable fluorescence (Fv/Fm) from Pinto Lake that was serially diluted and exposed to 1000 ppm C7X1, with sampling conducted from 0 to 72 hours. Error bars are the standard deviation of biological and technical replicates.
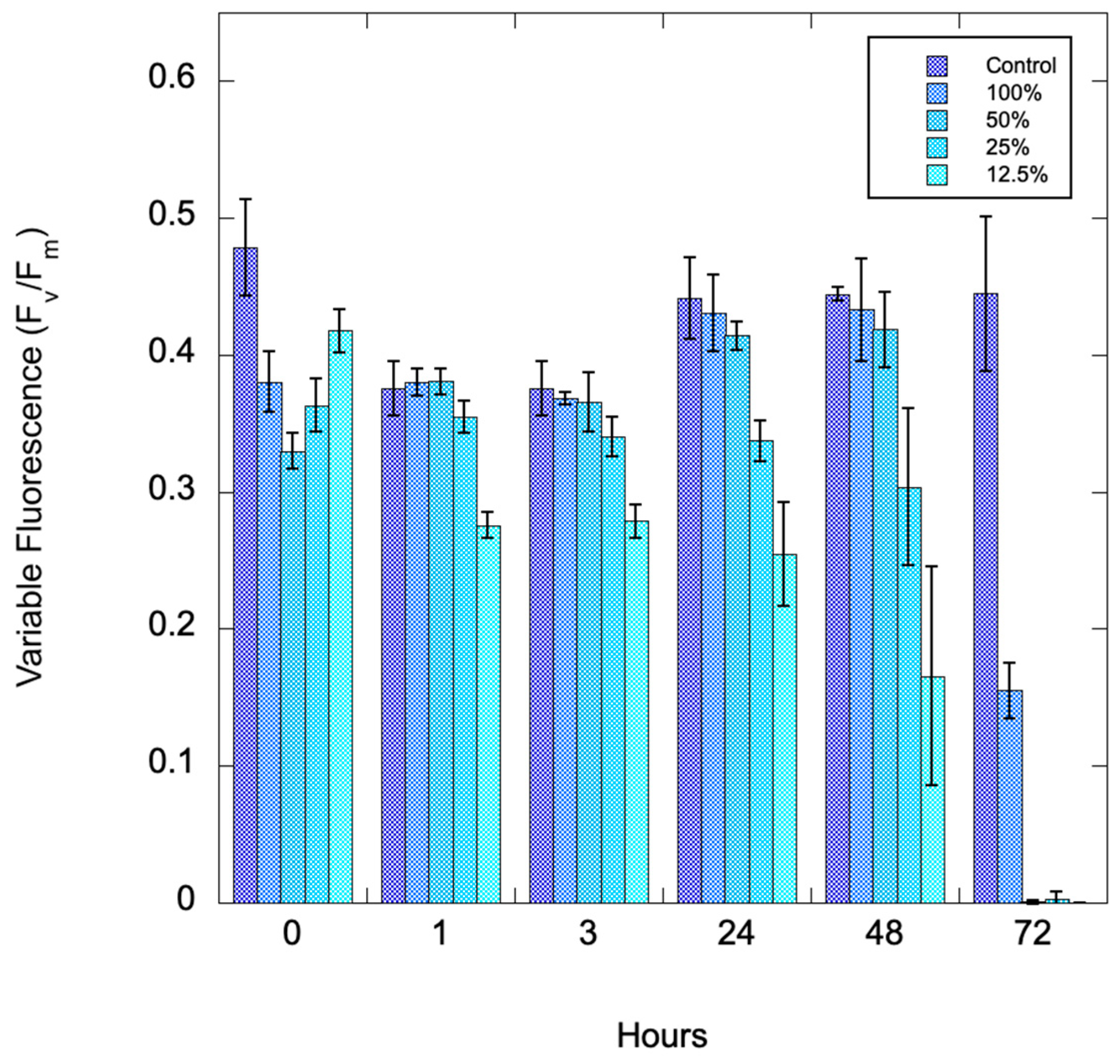
Figure 5.
Vertical column experiments with Pinto Lake water at 48 hours after treatment. Left: Control; Middle: treated and mixed; Right: treated at the surface only. Annotations on the left indicate depths at which samples were collected.
Figure 5.
Vertical column experiments with Pinto Lake water at 48 hours after treatment. Left: Control; Middle: treated and mixed; Right: treated at the surface only. Annotations on the left indicate depths at which samples were collected.

Figure 6.
Relative Electron Transport Rate (rETR) curves for the columns prior to treatment (A), Column 2 (C2) which was mixed and then treated, and (C) Column 4 (C4), which was sprayed at the top with no mixing. Depth sampling for C2 and C4 are as indicated in Figure 4. Error bars are standard deviations for biological and technical replicates.
Figure 6.
Relative Electron Transport Rate (rETR) curves for the columns prior to treatment (A), Column 2 (C2) which was mixed and then treated, and (C) Column 4 (C4), which was sprayed at the top with no mixing. Depth sampling for C2 and C4 are as indicated in Figure 4. Error bars are standard deviations for biological and technical replicates.
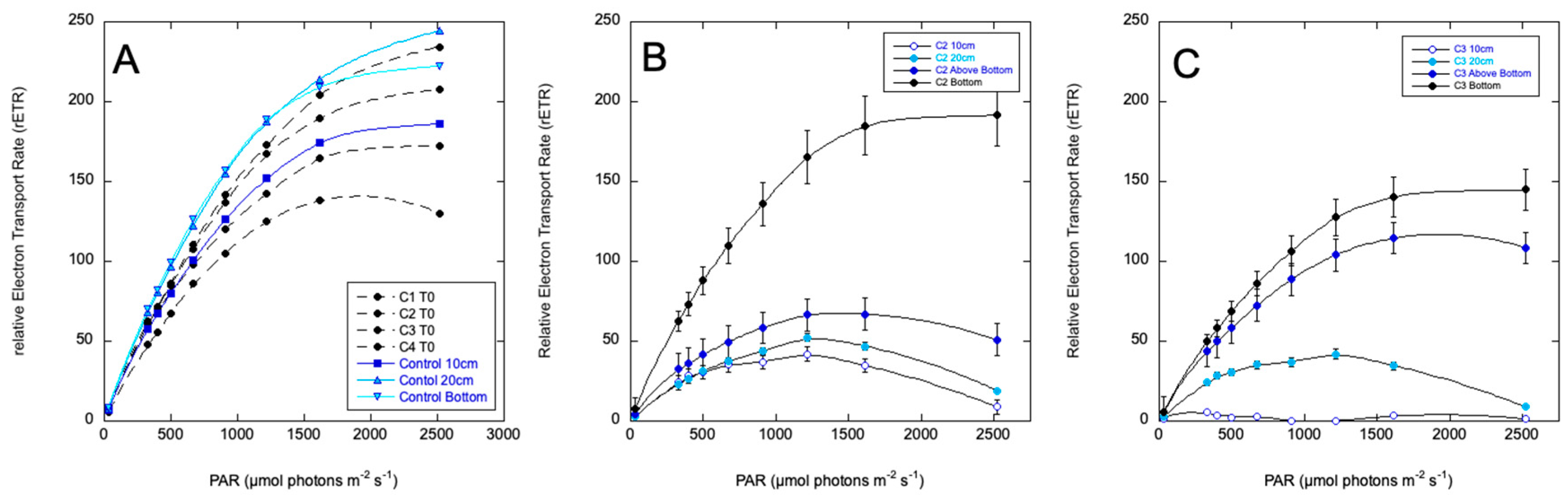
Figure 7.
Relative Electron Transport Rate (rETR) curves for the H. akashiwo columns at the initiation of the experiment (T0) and at 24 hours. Error bars are standard deviations for biological and technical replicates.
Figure 7.
Relative Electron Transport Rate (rETR) curves for the H. akashiwo columns at the initiation of the experiment (T0) and at 24 hours. Error bars are standard deviations for biological and technical replicates.
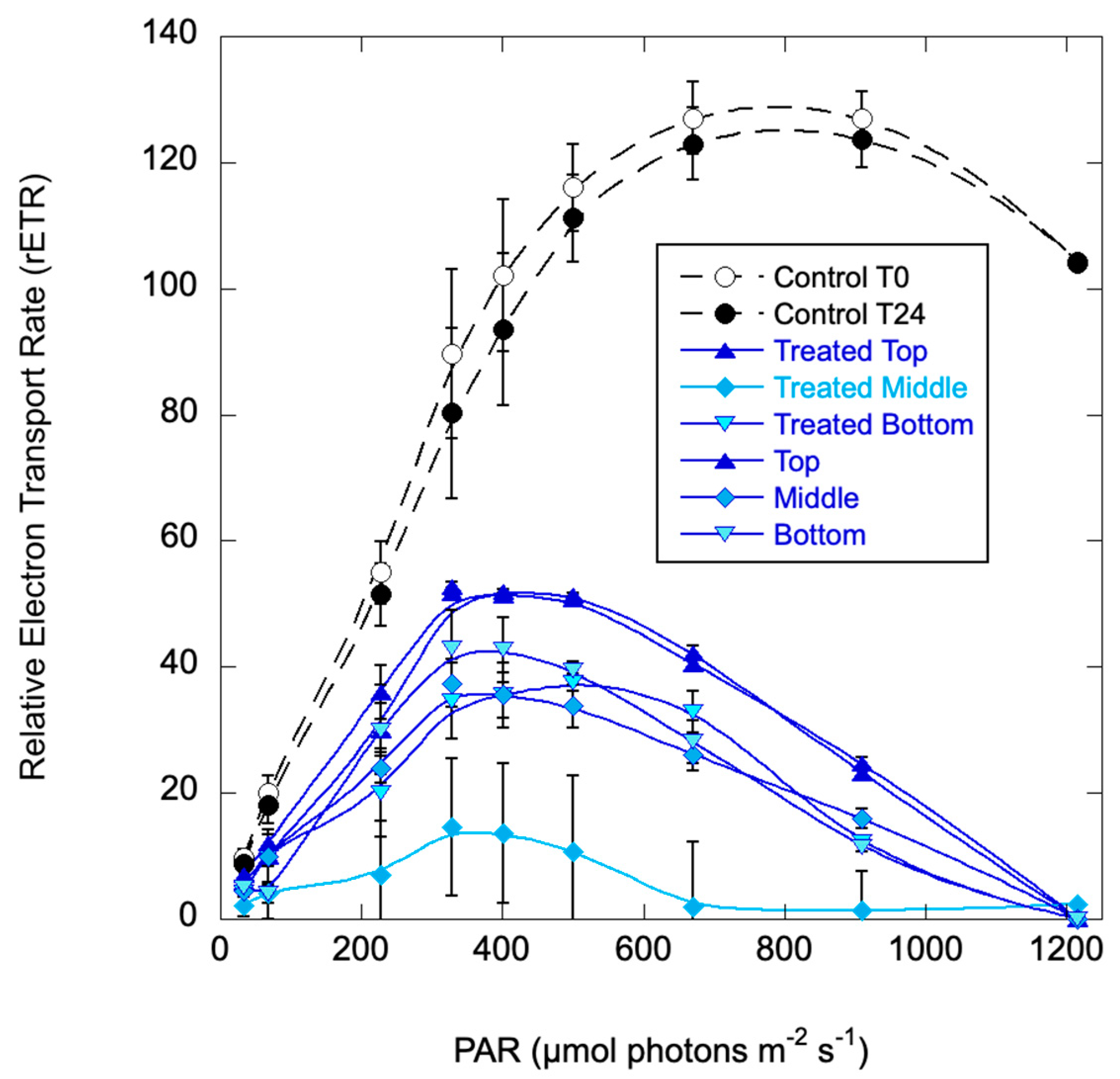
Figure 8.
Percent toxin release from P. multiseries (A), M. aeruginosa (B, light blue) and the column experiment from 28 September 2024 using Pinto Lake water (B, dark blue). Error bars are standard deviation from biological and technical replicates. .
Figure 8.
Percent toxin release from P. multiseries (A), M. aeruginosa (B, light blue) and the column experiment from 28 September 2024 using Pinto Lake water (B, dark blue). Error bars are standard deviation from biological and technical replicates. .
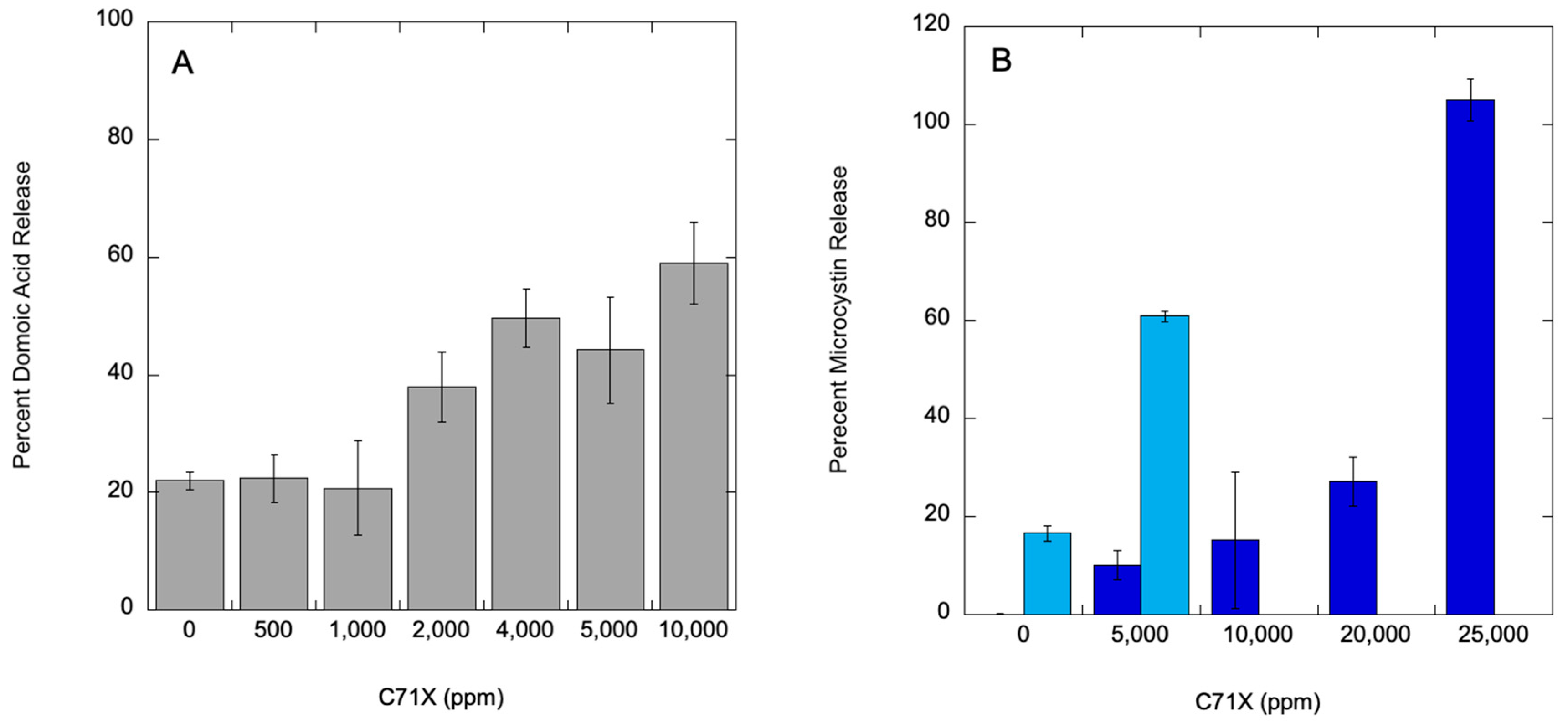
Table 1.
Ingredients in the Anjon AG C7X1 algaecide. (A) indicates active ingredients, (I) indicates inert ingredients.
Table 1.
Ingredients in the Anjon AG C7X1 algaecide. (A) indicates active ingredients, (I) indicates inert ingredients.
| Ingredient Name | CAS | Percent (%) |
|---|---|---|
| Peppermint (A) | N/A | 0.1 |
| Lemongrass Oil (A) | 8007-02-1 | 1.0 |
| Cinnamon (A) | N/A | 2.0 |
| Cottonseed Oil (A) | 8001-29-4 | 2.0 |
| Geranium Oil (A) | 8000-46-2 | 2.0 |
| Thyme (A) | N/A | 2.0 |
| Rosemary (A) | N/A | 3.0 |
| Clove (A) | N/A | 4.0 |
| Garlic (A) | N/A | 4.0 |
| Total Active Ingredients: | 20.1 | |
|
Citrus Pectin (I) |
9000-69-5 |
|
| Citrus Peel Extract (I) | 94226-47-4 | |
| Guar Gum (I) | 9000-30-0 | |
| Isopropyl Alcohol (I) | 67-63-0 | |
| Water (I) | N/A | |
| Xanthan Gum (I) | 11138-66-2 | |
| Total Inert Ingredients: | 79.1 |
Table 2.
The concentration at which Fv/Fm is inhibited by 50% (IC50) with C7X1 using Equation 1. (concentrations are ppm), and the dose at which Fv/Fm was <0.05 after 24 hours.
Table 2.
The concentration at which Fv/Fm is inhibited by 50% (IC50) with C7X1 using Equation 1. (concentrations are ppm), and the dose at which Fv/Fm was <0.05 after 24 hours.
| Algal Strain | IC50 (standard error) | Fv/Fm < 0.05 |
|---|---|---|
| Anabaena flos-aquae | 13,247 (6,962) | 20,000 |
| Gloeocapsa alpicola | 2,015 (292) | 10,000 |
| Microcystis aeruginosa | 976 (96) | 6,000 |
| Chattonella marina | 393 (135) | 10,000 |
| Chattonella subsalsa | 713 (140) | 10,000 |
| Heterosigma akashiwo | 1,618 (835) | 5,000 |
| Isochrysis galbana | 5,314 (563) | 10,000 |
| Pseudo-nitzschia multiseries | 4,073 (748) | 10,000 |
Table 3.
Algal cultures used for testing. UTEX is the University of Texas Austin culture collection, NCMA is the National Center for Marine Algae and Biota, EBL is the Environmental Biological Laboratory at Moss Landing Marine Labs, San Jose State University.
Table 3.
Algal cultures used for testing. UTEX is the University of Texas Austin culture collection, NCMA is the National Center for Marine Algae and Biota, EBL is the Environmental Biological Laboratory at Moss Landing Marine Labs, San Jose State University.
| Algal Strain | Strain/ID | Source |
|---|---|---|
| Anabaena flos-aquae | B 1444 | UTEX |
| Gloeocapsa alpicola | 589 | UTEX |
| Microcystis aeruginosa | LB 2385 | UTEX |
| Chattonella marina | CCMP 2049 | NCMA |
| Chattonella subsalsa | CCMP 2821 | NCMA |
| Heterosigma akashiwo | EB L71 | EBL |
| Isochrysis galbana | CCMP 1323 | NCMA |
| Pseudo-nitzschia multiseries | EBL 123 | EBL |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



