Submitted:
03 April 2025
Posted:
08 April 2025
You are already at the latest version
Abstract
The global proliferation of plastic waste presents an urgent environmental challenge, exacerbated by the limitations of current recycling and disposal technologies. In re-sponse, biocatalytic degradation using plastic-degrading enzymes has emerged as a promising solution for sustainable plastic waste management. This review provides a comprehensive overview of recent advances in the discovery, characterization, and engineering of enzymes capable of depolymerizing synthetic plastics. Enzymes such as PETases, cutinases, lipases, laccases, and peroxidases are examined in terms of their catalytic mechanisms, substrate specificity, and microbial origins. Special attention is given to developments in protein engineering, including the creation of highly efficient variants like FAST-PETase, as well as the integration of enzymes into immobilized sys-tems and synthetic microbial consortia. The review also explores novel sources of plas-tic-degrading activity, including insect gut microbiomes and environmental meta-genomes, highlighting their potential as genetic reservoirs for future biotechnological applications. Finally, the practical applications of these enzymes in enzymatic recy-cling, bioremediation and industrial processes are discussed, while identifying current limitations and future directions. The convergence of enzyme discovery, synthetic bi-ology, and circular economy principles may ultimately lead to scalable and sustainable strategies for mitigating plastic pollution.
Keywords:
plastic-degrading enzymes
; enzymatic recycling
; bioremediation
1. Introduction
Plastics have been used in almost every sectors of modern world, including packaging, construction, automotive, electronics, agriculture and healthcare [1]. They became ubiquitous commodities, due to their desirable properties such as low cost, light weight and high durability [2]. Synthetic polymers like polyethylene (PE), polypropylene (PP), polyvinyl chloride (PVC), polystyrene (PS), polyethylene terephthalate (PET) and polyurethane (PU) are engineered to meet high standards in performance that natural materials barely match [3]. However, the same properties that confer utility also result in environmental persistence. Most synthetic plastics are highly recalcitrant to natural degradation processes, leading to long-term accumulation in landfills, waterways and ecosystems [4].
Global production of plastics was over 400 million tons in 2020 and is expected to be doubled in 2050 unless major policy and technological interventions are implemented [5]. Alarmingly, only a small proportion of plastics (approximately 9% in the United States) are recycled through current waste management systems [6]. The remainder is either incinerated, contributing to air pollution and greenhouse gas emissions, or discarded in landfills where it may leach harmful additives into soil and groundwater [6]. A significant portion also ends up in terrestrial and marine environments, breaking down into microplastics that pervade water bodies, sediments and even the food we consume [7]. The persistence and ubiquity of microplastics have sparked growing concern among scientists and policymakers due to their potential toxicity and capacity to bioaccumulate in food chains [8].
Traditional strategies in plastic waste management have proven to be inadequate in addressing the scale and complexity of plastic pollution. Mechanical recycling, while, typically works best with clean and single-type plastic waste, and often produces lower-quality recycled materials due to degradation and contamination [9]. Chemical recycling methods such as pyrolysis or solvolysis can depolymerize plastics into monomers or fuels [9]. But they typically require high energy input, specialized infrastructure and often produce toxic by-products [9]. Incineration, though effective at volume reduction, releases harmful emissions and is not considered a sustainable solution [10]. Landfilling, the most common disposal method globally, does not eliminate plastic waste but merely displaces the environmental burden [11].
Given these challenges, there is increasing interest in developing biological approaches to manage plastic waste that are both effective and environmentally benign [12]. In particular, enzyme-mediated depolymerization offers a promising route for breaking down synthetic polymers under mild conditions [13]. Enzymes are naturally occurring biocatalysts capable of accelerating chemical reactions with remarkable specificity and efficiency. They are expressed in a wide range of microorganisms, including bacteria and fungi, many of which have evolved mechanisms to degrade natural polymers such as cellulose, lignin, and cutin [14]. The recent discoveries in microbial enzymes that can also degrade synthetic polymers have opened new avenues for sustainable treatment of plastic waste [15,16].
A landmark discovery in this field was made in 2016 when researchers identified Ideonella sakaiensis, a bacterium capable of using PET as its primary carbon source [17]. This organism secretes PETase, an enzyme that hydrolyzes PET into its constituent monomers, terephthalic acid (TPA) and ethylene glycol (EG), under ambient conditions [17]. This breakthrough demonstrated the feasibility of biological PET degradation and stimulated a surge of research aimed at discovering and optimizing enzymes with similar capabilities [3,12,15,16]. Since then, several classes of plastic-degrading enzymes have been identified, each with distinct substrate preferences and catalytic mechanisms [3,4,12,15,16,18]. These include various polyester hydrolases such as PETase, cutinases, and lipases. They also include polyurethane-degrading enzymes and oxidative enzymes like laccases and peroxidases. Additionally, alkane hydroxylases initiate the breakdown of long-chain hydrocarbons found in polyolefins like PE and PP [18,19].
The mechanisms by which these enzymes function vary depending on the chemical structure of the target plastic [3,18,20]. Enzymes that act on PET and PU typically hydrolyze ester or urethane bonds, while those targeting polyolefins rely on oxidative processes to introduce functional groups that destabilize the carbon backbone [15,18,20]. The effectiveness of enzymatic degradation depends on several factors. These include the crystallinity of plastic, molecular weight and the presence of additives or contaminants [20]. Environmental conditions such as temperature and pH also play a crucial role [20].
In recent years, there has been remarkable progress in improving plastic-degrading enzymes [21,22]. Advances have come through protein engineering, directed evolution and machine learning-based design [21,22]. For example, engineered PETase variants have been developed with better thermal stability and catalytic efficiency [20,21,23,24]. These improvements allow rapid depolymerization of post-consumer PET at higher temperatures [20,21,23,24]. Other studies have focused on building multi-enzyme systems or fusion proteins that replicate the natural metabolic pathways of plastic-degrading microorganisms [16,19]. Additionally, enzyme immobilization techniques, in which enzymes are fixed onto solid supports, have been employed to improve enzyme reusability and operational stability in industrial applications [20,21].
These advancements are beginning to translate into real-world applications [25]. Enzymatic recycling is emerging as a promising alternative to traditional methods [25]. Companies and research consortia are developing pilot-scale processes for enzymatic PET breakdown and repolymerization [25]. Enzyme-based solutions are also being investigated for the treatment of plastic-contaminated environments, including soil and marine ecosystems, as well as for the degradation of microplastics in wastewater treatment facilities [26]. Despite significant technical and economic challenges, enzyme-mediated plastic degradation is rapidly emerging as a critical component of the global strategy to combat plastic pollution [3].
This review provides a comprehensive overview of current knowledge on plastic-degrading enzymes. It covers the types of enzymes involved, their biochemical properties and mechanisms of action, microbial sources and the specific plastic substrates they target. Furthermore, it also highlights recent breakthroughs in enzyme discovery and engineering. In addition, it discusses practical applications and limitations, and explores future directions for research and development. By synthesizing key findings from across the literature, this work aims to shed light on the potential of enzymatic technologies to support a circular plastic economy and contribute meaningfully to environmental sustainability.
2. Plastic–Degrading Enzymes
2.1. Classification of Plastic-Degrading Enzymes
Plastic-degrading enzymes can be broadly categorized based on the types of chemical bonds they act upon and the nature of the plastic substrates they target [20]. Primarily, these enzymes fall into two main categories; hydrolytic enzymes, which cleave hydrolysable bonds like esters or amides, and oxidative enzymes, which degrade inert carbon-carbon backbones through oxidation processes [20]. (see Table 1 for an overview)
2.1.1. Hydrolytic Enzymes
Hydrolytic enzymes target plastics that contain ester or urethane bonds [15,27]. As a result, plastics like PET and certain types of polyurethanes are susceptible to enzymatic hydrolysis [20,27]. Key enzyme classes in this category include;
2.1.2. Oxidative Enzymes
Oxidative enzymes are sourced from lignin-degrading fungi [20,30]. They initiate the degradation of plastics lacking hydrolysable bonds, such as polyolefins (PE, PP), PS, and PVC [20,30];
2.1.3. Specialized and Auxiliary Enzymes
Some enzymes defy simple categorization but play crucial roles in specific degradation pathways:
This classification and the accompanying examples provide a foundation for understanding enzyme-mediated plastic degradation mechanisms. They also help to guide the selection or engineering of enzymes for specific plastic substrates. The following sections examine the biochemical properties and catalytic mechanisms of each major enzyme class in greater detail.
2.2. Mechanisms of Plastic-Degrading Enzymes
2.2.1. PETases and Polyester Hydrolases
PETase, first identified in the bacterium I. sakaiensis in 2016, specifically hydrolyzes PET, a common polyester found in bottles and textiles [17,20]. PETase specifically cleaves ester bonds in PET to yield MHET primarily, alongside minor quantities of TPA [17,20]. The enzyme MHETase, also produced by I. sakaiensis, subsequently hydrolyzes MHET into TPA and EG [17,20]. Structurally, PETases belong to the α/β-hydrolase fold family [34]. They utilize a catalytic serine residue to perform a nucleophilic attack on ester bonds, forming an intermediate complex that is subsequently hydrolyzed into monomers [20,34]. PETases possess an open active-site cleft that facilitates binding to polymer chains [34]. Additionally, they feature specialized surface-binding loops that enhance accessibility to the substrate [34].
Figure 1.
Schematic illustration of enzyme-mediated plastic degradations.
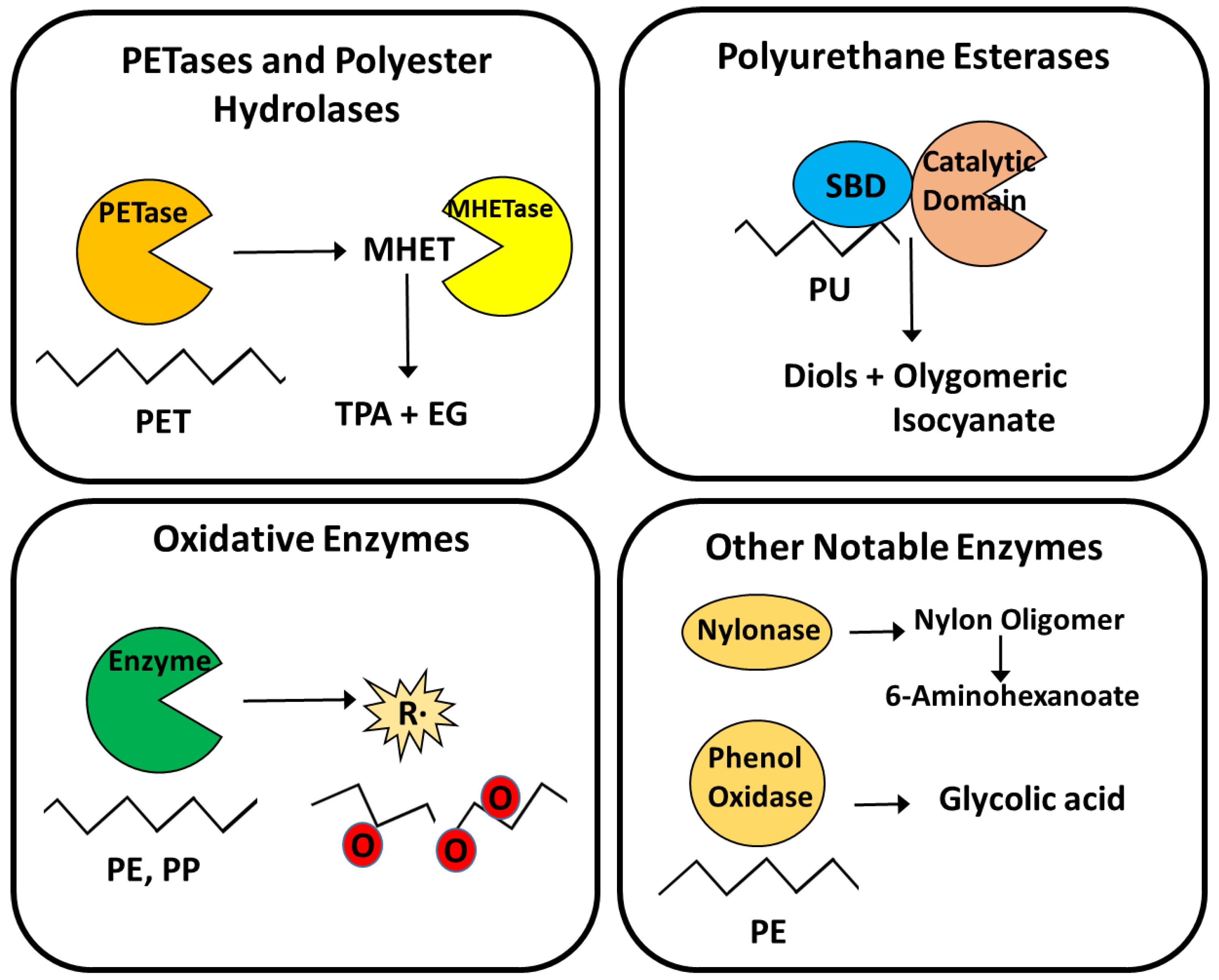
Cutinases from organisms such as Fusarium solani and Thermobifida share the catalytic triad mechanism (Ser-His-Asp) with PETases [20,35,36]. They possess an open and shallow active site architecture that is well suited for interacting with polymer substrates [20]. Because of this open active site architecture, cutinases exhibit broad substrate specificity toward synthetic polymers [37]. Thus, they can also depolymerize polycaprolactone (PCL), polylactic acid (PLA), polybutylene succinate (PBS) and polyester-type PU that possess ester linkages [27,38,39,40,41].
Lipases, particularly those derived from Candida antarctica, can hydrolyze aliphatic polyesters such as PCL and PLA, breaking down the ester bonds within their polymer backbones [20,42]. These enzymes also operate through a classical serine hydrolase mechanism [20,42]. They are particularly effective when acting on polymers in an amorphous state or when the substrate is exposed to elevated temperatures [20,42]. Higher temperatures enhance the mobility of polymer chains, thereby improving enzyme accessibility and catalytic efficiency [20].
2.2.2. Polyurethane Esterases
Enzymes secreted by bacterial genera such as Pseudomonas and Comamonas are commonly referred to as polyurethane esterases or polyurethanases [43,44]. These enzymes feature a dual-domain architecture, consisting of a hydrophobic surface-binding domain (SBD) and a catalytic domain [45]. Initially, the SBD facilitates enzyme adhesion to the polymer surface [45]. Subsequently, the catalytic domain hydrolyzes urethane ester linkages, releasing smaller fragments such as diols and oligomeric polyisocyanates [45].
Proteases derived from fungal species such as Aspergillus tubingensis and bacterial species like Bacillus subtilis play a key role in degrading PU [27,46]. These enzymes specifically target peptide-like urethane and urea bonds within the polymer [27,46,47]. Rather than acting alone, these proteases often work in synergy with esterases, which hydrolyze the ester linkages present in PU [20,47]. This combined enzymatic activity enhances overall degradation efficiency [20,47]. As a result, the polymer is broken down into smaller, more accessible intermediates that can be further metabolized or chemically processed [20,47].
2.2.3. Laccases, Peroxidases and Oxidative Enzymes
Oxidative enzymes, predominantly laccases and peroxidases derived from white-rot fungi such as Trametes versicolor and Phanerochaete chrysosporium [48,49]. These enzymes initiate the degradation of polymers that lack hydrolysable bonds, including polyolefins like PE and PP, as well as PS and PVC [48,49]. The enzymes catalyze radical-mediated oxidation reactions, abstracting electrons from polymer backbones to form reactive radical intermediates [27,48,49]. These radicals then introduce oxygen-containing functional groups such as hydroxyl and carbonyl groups into the polymer chains [27,48,49]. This oxidative functionalization reduces polymer chain stability and promotes subsequent biodegradation by secondary microbial enzymes or spontaneous chemical degradation [27,48,49,50]. For instance, manganese peroxidase can effectively degrade PE and facilitate PVC dechlorination by breaking carbon–carbon bonds and releasing hydrochloric acid (HCl), aiding microbial assimilation [48,49,50].
PS, characterized by aromatic side groups, presents unique challenges [51]. Oxidative enzymes such as laccases can generate radicals and initiate chain oxidation [27]. Notably, an esterase from the fungus Lantinus tigrinus directly depolymerizes PS, forming non-toxic intermediates [52]. So, the enzyme demonstrated rare enzymatic activity specifically targeting PS [52]. Additionally, laccase-mediated Fenton-like reactions have been explored to generate hydroxyl radicals that significantly enhance the oxidative degradation of PS and other recalcitrant plastics [53].
2.2.4. Other Notable Enzymes
In addition to the major groups above, there are specialized enzymes and pathways worth noting. Nylon-degrading enzymes such as nylonases (e.g., 6-aminohexanoate-dimer hydrolase) evolved in certain Flavobacterium and Pseudomonas strains to break down nylon oligomers – a famous case of microbes adapting to a synthetic polymer waste [54]. While nylon oligomers are not high-molecular-weight plastics, this example illustrates how enzyme evolution can create entirely new activities toward man-made polymer [54].
Another example is the metabolism of styrene (the monomer of PS) by bacteria like Pseudomonas putida [32,55]. P. putida can’t directly depolymerize PS, but it can consume styrene after pyrolysis or partial degradation [55,56]. It uses a styrene monooxygenase and downstream enzymes to convert styrene into valuable products like polyhydroxyalkanoate (PHA) or other intermediates [57]. This approach, combining partial thermal degradation with microbial enzymatic conversion, is a form of hybrid degradation for plastics like PS [57].
Alkane hydroxylases deserve mention for polyolefins. These bacterial enzymes normally oxidize long-chain alkanes (like those in petroleum); some studies suggest they can slowly oxidize PE chains, since PE is essentially a very long alkane [58,59,60,61]. Strains of Rhodococcus and Pseudomonas have been found to induce alkane hydroxylase and other oxidases when grown on PE films, leading to formation of carbonyl groups on PE [58,59,61]. Although the activity is low, this indicates that common hydrocarbon-degrading enzymes can play a role in plastic degradation if the organism can access the polymer surface [58,59,61]. Often, physical or chemical pretreatment, such as ultraviolet (UV) exposure to create initial cracks and oxidized bonds in PE, is needed to enable such enzymes to attack the polymer [4,62,63].
Finally, recent research has uncovered insect-derived enzymes capable of plastic depolymerization [64]. In 2022, scientists found that the saliva of waxworms (Galleria mellonella larvae) contains two phenol oxidase enzymes that rapidly oxidize and depolymerize PE at room temperature [64]. These enzymes are believed to directly catalyze the oxidation of PE backbone [64,65]. So, they can bypass the typical requirement for high-temperature or UV pretreatment [64]. This reaction breaks the long polymer chains into glycolic acid and other short-chain products within hours [64]. This discovery represents the first known case of enzymes that can efficiently degrade PE under mild conditions [64]. It also introduces a new category of plastic-degrading enzymes derived from insect biochemistry, distinct from those previously identified in microbial systems [64].
In summary, plastic-degrading enzymes utilize a range of catalytic mechanisms, from hydrolysis of ester and urethane bonds to radical-mediated oxidation of carbon–carbon backbones. These diverse biochemical strategies enable the breakdown of both hydrolysable and non-hydrolysable plastics. Hydrolytic enzymes, such as PETases and cutinases, effectively depolymerize polyesters like PET, while oxidative enzymes like laccases and peroxidases initiate the degradation of more inert polymers such as PE, PP and PS. The versatility of these enzymatic approaches highlights their promise as environmentally friendly tools for addressing plastic waste.
3. Microbial Sources of Plastic-Degrading Enzymes
A diverse array of microorganisms, including bacteria, fungi, and symbiotic communities, produce enzymes capable of degrading plastics. This section details some key microbial sources and highlights their abilities to target various plastic types (see Table 2 for an overview).
3.1. Bacteria
Several bacteria isolated from environments rich in plastic waste, such as soil, compost, and marine ecosystems, have demonstrated the ability to degrade plastics. One of the most notable examples is I. sakaiensis [17]. It can degrade low-crystallinity PET in near completion by secreting PETase and MHETase [17]. Many thermophilic actinomycete bacteria, including Thermobifida fusca and Thermomonospora species, secrete cutinases and esterases that can break ester bonds in PET and PU [37,67].
Pseudomonas is a genus particularly rich in degradation capabilities: different Pseudomonas species can utilize PU, PS, and even partially oxidized PE as carbon sources [78]. For instance, P. chlororaphis and P. fluorescens produce extracellular polyurethane esterases (PueA, PueB) that break down polyester-PU [78,79]. Additionally, newly discovered strains like Pseudomonas sp. TDA1 have demonstrated the capability to utilize PU foam as their sole carbon source [43]. Another species, P. putida is known for metabolizing styrene (from PS) and even converting PET-derived terephthalate into valuable bioproducts [80,81,82].
Other notable bacterial genera include Bacillus, known for degrading PU and pre-oxidized PE, and Rhodococcus, which has demonstrated slow mineralization of PS and PE films over extended incubation periods [61,82,83,84,85]. In marine settings, Alcanivorax and Kocuria species have been reported to colonize and degrade floating plastic debris, likely relying on their alkane-degrading enzymes to attack PE [86,87,88].
Overall, bacterial plastic degraders are often found in environments loaded with plastic waste, suggesting adaptive evolution of their enzymatic repertoire.
3.2. Fungi
Fungi, especially saprophytic molds and lignin-degrading fungi, are prolific producers of relevant to plastic biodegradation (laccases, peroxidases, cutinases, etc.) [18]. Ascomycete fungi such as Aspergillus, Penicillium, Fusarium, and Cladosporium have been repeatedly isolated from plastic-contaminated sites and shown to cause degradation of plastics [27]. Aspergillus niger and Penicillium simplicissimum, for example, have demonstrated the ability to colonize and embrittle PE films after pretreatment, likely through secretion of oxidative enzymes and cutinase-like esterases [89]. F. solani produces the classic cutinase enzyme and can depolymerize PET and PU coatings [20]. Basidiomycete fungi, known for their lignin-degrading systems, also play a major role [27]. White-rot fungi like Trametes versicolor, Phanerochaete chrysosporium, and Pleurotus ostreatus are frequently cited for their ability to degrade recalcitrant plastics such as PE and PVC by virtue of their laccases and peroxidases [27]. For PS, fungi have been less studied, but Lantinus species have shown some PS breakdown via esterase activity [90]. Fungal enzymes often have high stability and are secreted in large amounts, making fungi attractive for enzyme production in plastic waste treatment. Notably, fungi can sometimes directly penetrate plastics with their hyphae, aiding in mechanical disintegration in combination with enzymatic action [91].
3.3. Symbiotic and Other Sources
Symbiotic microbial communities within insect guts present another exciting avenue for plastic biodegradation. The gut microbes of waxworms and mealworms enable these larvae to digest plastics like PE and PS, respectively. Waxworms (G. mellonella larvae) secrete enzymes in their saliva and gut capable of oxidizing PE effectively under mild conditions [64]. In mealworms (Tenebrio molitor larvae), gut bacteria including Exiguobacterium and Yokenella have been implicated in degrading PS foam, as the worms can subsist on a PS diet and excrete degraded fragments [92,93,94].
Additionally, ruminants (like cows) have also been suggested as a source: recent studies found that the microflora in cow rumen fluid can degrade PET films, possibly due to a mix of cutinases or lipases present from their plant-based diet digestion [95,96].
Compost and soil consortia (mixed communities) sometimes outperform pure cultures, as different organisms can attack different components of a plastic simultaneously [97]. For example, a mixed fungal consortium of Candida (yeast) and Trichoderma was shown to more effectively break down UV-treated PE than either alone, likely combining Candida’s oxidative metabolism with Trichoderma’s enzymatic arsenal [97]. Metagenomic surveys have indeed found that environments chronically exposed to plastic (e.g. landfill soils) harbor a higher diversity of plastic-degrading enzymes, indicating microbial communities collectively adapt to exploit plastic as a nutrient source [98].
In summary, microbial communities from diverse environments, including soil, marine ecosystems, insect guts and ruminant digestive tracts, serve as both a genetic reservoir of potential plastic-degrading enzymes and active agents for plastic biodegradation. These microbes not only secrete effective depolymerizing enzymes but also metabolize the resulting breakdown products, demonstrating their utility as both tools and templates for sustainable plastic waste treatment.
4. Recent Advances in Plastic-Degrading Enzymes
Over the past decade, research into plastic-degrading enzymes has significantly accelerated, driven by technological advancements in genomics, protein engineering, and synthetic biology [20,24,99]. These efforts have resulted in the discovery of novel enzymes, enhanced catalytic properties of existing enzymes, and innovative deployment strategies for scalable plastic biodegradation [100,101] (see Table 3 for an overview).
4.1. Discovery of New Enzymes and Pathways
Scientists are leveraging genomics and environmental screening to find novel plastic-degrading enzymes [98,102]. A 2021 global study scanned over 200 million DNA sequences from environmental samples and identified 30,000 enzyme homologs with the potential to degrade 10 different plastics [102,103]. The presence and abundance of these enzymes were found to correlate with local plastic pollution levels, suggesting an evolutionary response by microbial communities to exploit plastics as carbon sources [103]. Notably, many of the identified sequences represented entirely new proteins unrelated to known hydrolases, presenting a rich reservoir for future enzyme characterization and development [103].
Among specific discoveries, the phenol oxidases found in the saliva of waxworms (G. mellonella) in 2022 marked a breakthrough as the first enzymes capable of rapidly depolymerizing PE without the need for pretreatment [64]. Similarly, in 2020, researchers isolated a Pseudomonas strain from a waste site that can degrade and even grow on PU, utilizing it as a sole carbon source [104]. They identified enzymes from the strain that break PU into absorbable metabolites [104]. Each new enzyme expands the range of plastics that might be biologically tackled, such as recently discovered PET hydrolases in leaf compost, marine environments, and even in the cow rumen microbiome [95,105].
4.2. Protein Engineering for Enhanced Enzyme Performance
Perhaps the most dramatic progress has come from engineering enzymes like PETases to be faster and more robust. In 2018, scientists solved the crystal structure of I. sakaiensis PETase and noted it was similar to cutinases, providing clues for improvement [106]. By 2020, a team engineered the “double mutant” PETase (inspired by cutinase active-site features), which showed a substantial increase in PET degradation rate [107]. Around the same time, French company Carbios and academic collaborators evolved an enhanced cutinase (from leaf compost bacteria) that achieved 90% depolymerization of PET in just 10 hours [66,101]. This enzyme was 10,000 times more efficient than the wild-type at breaking PET bonds, enabling the complete recycling of PET bottles into new plastic of equal quality [66,101]. Following this, in 2020 researchers at University of Portsmouth linked PETase and MHETase together, creating a “super-enzyme” that could digest PET roughly 6 times faster than PETase alone [108]. Although too large for expression by a single microbe, this chimeric enzyme demonstrated the potential of multi-enzyme systems to accelerate depolymerization [108].
The latest breakthrough came in 2022 with the development of FAST-PETase (Functional, Active, Stable, and Tolerant PETase) [109]. Using machine-learning algorithms to guide mutations, researchers introduced 5 mutations into PETase, improving its thermostability and activity across various pH ranges [109]. FAST-PETase operates effectively at temperatures up to 50 °C (near the glass transition of PET) and remains active in various conditions [109]. Impressively, FAST-PETase degraded post-consumer PET products from 51 different sources within one week [109]. The enzyme could even digest the amorphous portions of a commercial water bottle without any preprocessing [109]. This robustness allowed the team to demonstrate a closed-loop PET recycling [109]. PET trash was enzymatically depolymerized to monomers, purified, and re-polymerized into new PET plastic, completing a full recycle with the help of the engineered enzyme [109]. FAST-PETase represents a significant step toward practical enzymatic recycling at scale, and its design showcases the power of AI-driven protein engineering [110].
Beyond PET, efforts are underway to engineer enzymes for other plastics. For instance, protein engineering of leaf-branch compost cutinase (LCC) has yielded variants capable of degrading both PET and polyethylene furanoate (PEF), a bio-based polyester [111]. Additionally, studies have demonstrated that cutinases can hydrolyze polyamide 6,6 (PA 6,6), indicating potential for broader application in polyamide degradation [112]. These advancements highlight the versatility of engineered cutinases in addressing various plastic waste challenges.
Enzyme engineering has also targeted thermostability, since plastic depolymerization is often more efficient at higher temperatures where the polymer is softer [100]. Strategies like introducing disulfide bonds or stabilizing mutations have extended enzyme working ranges to 70–75 °C (as with some engineered cutinases), which greatly accelerates PET degradation due to increased chain mobility [66]. Similarly, directed evolution of Aspergillus laccase has produced variants that are more tolerant of the harsh conditions needed to oxidize PE surfaces [113].
4.3. Enzyme Immobilization and Reactor Systems
Another advancement is in how enzymes are deployed. Immobilization of PETases and lipases on solid supports or within biofilms improves enzyme reusability, operational stability, and resistance to denaturation [114]. For instance, immobilizing PETase onto nanostructured cobalt phosphate allowed for high enzyme loading with minimal mass transfer limitations, leading to improved stability and reusability [114]. Similarly, lipases immobilized on magnetic nanoparticles via polydopamine coating exhibited enhanced pH and thermal stability compared to their free counterparts [115]. These immobilized systems are employed in packed-bed and fluidized-bed reactors designed to continuously process plastic waste [116]. There have been pilot demonstrations of enzymatic recycling: for instance, Carbios announced a demonstration plant where tons of PET waste are enzymatically converted to monomers for repolymerization [101]. Immobilization also confers increased thermostability, allowing enzymes to function at higher temperatures and degrade more crystalline plastics [117]. This is particularly important for recycling commercial-grade plastic products that exhibit greater resistance to enzymatic attack [16].
4.4. Metabolic Engineering and Synthetic Biology
Instead of using isolated enzymes, some efforts focus on engineering whole-cell systems due to recent advancements in synthetic biology and metabolic engineering [118]. For example, E. coli or Pseudomonas have been genetically modified to secrete PETases and take up the monomers, effectively creating an “engineered microbial recycler” [119]. One study engineered P. putida to consume terephthalic acid (from PET) and funnel it into the production of biopolymer PHA, thus coupling plastic degradation with bio-upcycling [120].
More advanced strategies involve constructing synthetic consortia wherein one microbial species secretes oxidative enzymes to initiate plastic degradation, while another species carries out depolymerization and downstream bioconversion [121]. Such systems represent a step toward integrated biorefineries for plastic waste, combining biodegradation, resource recovery, and sustainable chemical production in a single platform [121].
5. Applications in Recycling, Bioremediation and Industry
The practical deployment of plastic-degrading enzymes is a rapidly evolving field, with potential applications across recycling, environmental cleanup, and industrial operations [122]. These strategies aim to address the limitations of traditional plastic waste management by enabling selective, sustainable, and scalable alternatives [122].
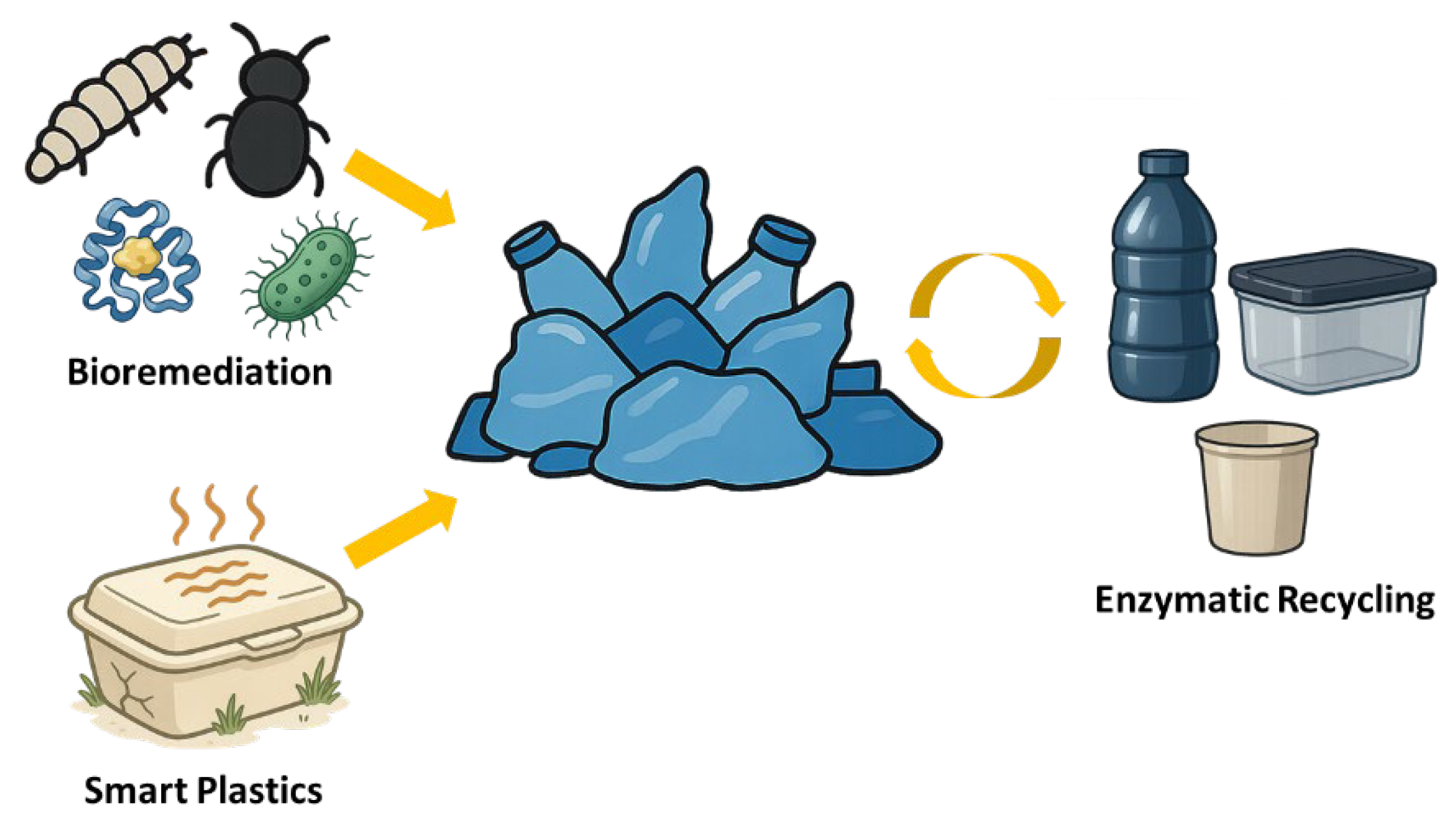
Figure 2.
Schimatic Illustration about Applications of Plastic-Degrading Enzymes.
5.1. Enzymatic Recycling
Enzymatic recycling offers a pathway to circular plastic economy by depolymerizing plastic wastes into their original monomers, which can then be purified and repolymerized into new plastics [123]. PET serves as the leading example. Companies such as Carbios have developed industrial-scale enzymatic recycling plants that utilize PETases and cutinases to break down PET from post-consumer waste—such as bottles and textiles—into TPA and EG [25]. These monomers can then be repolymerized into virgin-grade PET without loss in material quality [25]. Unlike mechanical recycling, which degrades polymer quality, enzymatic recycling yields monomers of equal purity to petrochemical sources [25]. The closed-loop PET recycling demonstrated with FAST-PETase exemplifies this approach [124]. Enzymatic recycling is also being explored for PU, converting them into polyols and other components that can serve as raw material for new polymer synthesis [125].
Addressing mixed plastic waste remains a challenge [100]. One solution involves designing enzyme cocktails tailored to degrade different plastic types either sequentially or simultaneously [100,126]. Another strategy is upcycling, where enzymes convert plastic into chemical intermediates that can be transformed into value-added products [126]. For example, Pseudomonas species can metabolize PET monomers into β-ketoadipate, a precursor for high-performance polymers and biodegradable plastics [32]. These strategies enhance the economic appeal of biocatalytic plastic processing, as well as promote a plastic circular economy.
5.2. Bioremediation and Environmental Cleanup
Using microbes or enzymes to degrade plastics in situ (in the environment) is a more challenging application but one with great appeal [11,46]. Laboratory studies have shown that adding plastic-degrading fungi or bacteria to soil can accelerate the degradation of plastic films, especially if combined with pretreatments like plowing the soil with oxidative chemicals [127]. For marine bioremediation, researchers are investigating bioaugmentation with marine bacteria that have natural plastic-degrading abilities, or deploying enzyme-coated scaffolds that could capture microplastics and degrade them [12]. For example, enzyme-producing bacterial consortia have been tested in composting facilities to degrade compostable plastics like PLA and polybutylene succinate faster than normal [128]. However, for non-compostable plastics like PE and PP, direct bioremediation is slow without pretreatment [129].
Innovative approaches include the use of insects as biodegradation vectors. Waxworms can ingest PE, initiating oxidation and partial breakdown through enzymes in their saliva and digestive tracts [64]. Mealworms, on the other hand, can consume PS foam, aided by gut microbiota such as Exiguobacterium and Yokenella [92,93,94]. While promising, these biotic strategies require careful ecological evaluation before field application.
5.3. Industrial and Commercial Uses
Enzymes are increasingly considered for integration into industrial workflows to assist with plastic processing and waste management. One concept under development involves embedding enzymes or their precursors into plastic materials, creating smart plastics that degrade upon exposure to specific environmental triggers (e.g., heat, moisture, or pH changes) [130,131]. For instance, one study describes the development of a PLA-based plastic with embedded, engineered PLA hydrolase that fully disintegrates under home-compost conditions within 20-24 weeks [130]. These enzyme-activated systems could allow for post-use degradation without compromising product durability during use [131].
Enzymes are also being investigated for use in textile and packaging industries. Cutinases, for example, could be employed to clean PET-based industrial waste (e.g., textile off-cuts or film trimmings) by depolymerizing them directly on-site [132]. In wastewater treatment facilities, enzymatic degradation of microplastic particles using membrane-immobilized enzymes offers a promising avenue for reducing microplastic load in effluents [133]. Nonetheless, this kind of technology is currently underappreciated due to a lack of information and proof, necessitating extensive large-scale study for adoption [133].
While many of these applications remain at the research or pilot stage, they illustrate the potential of enzymatic technologies to provide flexible, efficient, and environmentally friendly solutions across multiple sectors affected by plastic pollution.
5. Current Limitations and Future Prospects
Despite significant progress in the discovery, engineering and application of plastic-degrading enzymes, several limitations hinder their large-scale implementation. A key challenge lies in the relatively low degradation efficiency of many enzymes, especially against common plastics such as PE, PP and high-crystallinity PET [20]. These materials are inherently resistant due to their dense molecular structures and lack of hydrolyzable functional groups [20]. Enzymatic breakdown of these plastics is often slow, requiring weeks to months for measurable weight loss under laboratory conditions—far from sufficient for industrial application [20].
Crystalline regions in polymers are largely inaccessible to enzymes, necessitating pretreatments such as heat, UV exposure, or mechanical milling to enhance surface reactivity [15]. However, these additional steps add complexity and cost. Enzyme performance is also limited by narrow optimal conditions [20]. Most enzymes function within specific temperature and pH ranges and are sensitive to additives commonly found in plastic waste, such as dyes and plasticizers [20]. Enhancing enzyme robustness—particularly thermostability and chemical resistance—remains a major objective of protein engineering [101].
Another significant hurdle is substrate specificity. Most known enzymes act on a narrow range of polymers or chemical bonds, meaning that mixtures of different plastics require enzyme cocktails or sequential processing steps [20]. Designing effective reactors and optimizing these multienzyme processes pose considerable technical challenges [101]. Furthermore, achieving complete mineralization of plastics (conversion to CO₂, water, or biomass) is still rare, especially for polyolefins and PS. Partial degradation or fragmentation into microplastics—without full assimilation—can exacerbate environmental issues rather than solve them [20].
From an economic standpoint, enzyme production costs must compete with low-cost virgin plastics. Although fermentation technologies from industries like detergents offer scalable enzyme production, commercial viability will depend on improving enzyme efficiency and reducing downstream processing costs [20]. Regulatory barriers also exist, particularly for environmental release of engineered enzymes or microorganisms [16]. Safety, ecological impact and the potential for horizontal gene transfer are key concerns that must be addressed through thorough risk assessment [88].
Despite these limitations, future prospects for enzymatic degradation of plastics are promising. Protein engineering, including machine-learning-guided design and directed evolution, is accelerating the development of faster and more robust enzymes, as demonstrated by recent PETase variants like FAST-PETase [23,24,110]. These approaches are now being applied to other enzyme families, potentially expanding the suite of biocatalysts capable of degrading various plastic types [20,130].
The pipeline of new enzyme discovery is also growing [98]. Metagenomic surveys continue to uncover novel enzyme candidates from environments heavily contaminated with plastic waste [97,98]. These discoveries offer the potential for building synthetic microbial consortia specifically tailored to different polymer types[98]. For example, a consortium might include an oxidase-secreting microbe to initiate breakdown of polyolefins, followed by hydrolase-producing microbes to complete depolymerization and others to metabolize resulting monomers [20].
Another promising strategy lies in chemo-enzymatic hybrid systems [131]. These combine mild chemical or physical pretreatments (e.g., plasma, UV, green oxidants) to soften or oxidize the plastic surface, followed by enzymatic action to depolymerize and assimilate the material [20]. Such combinations may offer a practical balance between cost, speed and completeness of degradation, particularly for highly recalcitrant plastics.
On the materials side, there is increasing interest in designing future plastics to be more amenable to enzymatic recycling [123,130]. This includes incorporating cleavable linkers or biodegradable segments into otherwise conventional polymers. Tailored enzymes could then recognize and efficiently cleave these weak points, enabling rapid and targeted degradation.
In summary, plastic-degrading enzymes have progressed from niche biological curiosities to credible agents for recycling and bioremediation. While scaling and broadening their applicability remain key challenges, advances in microbiology, bioengineering, and materials science are converging to unlock their full potential. With continued interdisciplinary effort, enzymatic technologies may soon provide scalable, sustainable, and circular solutions for managing global plastic waste.
6. Conclusion
The ongoing plastic pollution crisis demands transformative solutions, and enzyme-based plastic degradation offers a compelling avenue grounded in nature’s own catalytic machinery. Through years of research, enzymes such as PETase, cutinase, and laccase have emerged as potent tools for targeting a wide array of synthetic polymers. Advances in genomics, metagenomics, and protein engineering have vastly expanded the known repertoire of plastic-degrading enzymes and enhanced their catalytic capabilities under industrially relevant conditions.
Applications in enzymatic recycling, environmental bioremediation, and industrial plastic management are rapidly transitioning from conceptual frameworks to tangible solutions. From the closed-loop recycling of PET using engineered enzymes like FAST-PETase to the potential deployment of enzyme-functionalized filters in wastewater systems, these biotechnologies are redefining how plastic waste can be handled at the molecular level.
However, significant challenges persist. Current enzymes exhibit limited activity against highly crystalline or chemically inert plastics, leading to the need for pretreatment, which could be another limit as cost of process and complexity. Also the stability of enzymes and cost-effective production of them remains a bottleneck. Mixed plastic waste streams further complicate processing, demanding multifaceted enzymatic systems or innovative reactor designs. Moreover, concerns around incomplete degradation and the environmental deployment of engineered biological agents must be addressed through careful risk assessment and policy development.
Despite these limitations, the field continues to progress at an accelerated pace. Indeed, the number of landmark studies in this field has been growing explosively since 2020. Emerging directions include the development of synthetic microbial consortia, chemo-enzymatic hybrid strategies, and the creation of enzyme-sensitive materials designed with end-of-life degradation in mind. Integrating enzymatic solutions with broader systems of circular economy, sustainable manufacturing and environmental stewardship holds immense promise.
In conclusion, the convergence of enzyme discovery, bioengineering, and industrial application is shaping a new frontier in sustainable plastic management. Continued interdisciplinary collaboration and innovation will be essential to scale these technologies and expand their impact. As the field matures, enzymatic plastic degradation could become a cornerstone of global strategies to mitigate plastic pollution and foster a more sustainable material future.
Funding
This research received no external funding.
Conflicts of Interest
The author declares no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| PET | polyethylene terephthalate |
| PU | polyurethane |
| PE | polyethylene |
| PP | polypropylene |
| PVC | polyvinyl chloride |
| PS | polystyrene |
| PCL | polycaprolactone |
| PLA | polylactic acid |
| PHA | polyhydroxyalkanoate |
References
- Khan, S.; Iqbal, A. Organic polymers revolution: Applications and formation strategies, and future perspectives. J. Polym. Sci. Eng 2023, 6, 3125. [Google Scholar] [CrossRef]
- Stanley, J.; Culliton, D.; Jovani-Sancho, A.-J.; Neves, A.C. The Journey of Plastics: Historical Development, Environmental Challenges, and the Emergence of Bioplastics for Single-Use Products. Eng 2025, 6, 17. [Google Scholar] [CrossRef]
- Mohanan, N.; Montazer, Z.; Sharma, P.K.; Levin, D.B. Microbial and Enzymatic Degradation of Synthetic Plastics. Front Microbiol 2020, 11, 580709. [Google Scholar] [CrossRef] [PubMed]
- Wei, R.; Zimmermann, W. Microbial enzymes for the recycling of recalcitrant petroleum-based plastics: how far are we? Microbial biotechnology 2017, 10, 1308–1322. [Google Scholar] [CrossRef]
- Dokl, M.; Copot, A.; Krajnc, D.; Van Fan, Y.; Vujanović, A.; Aviso, K.B.; Tan, R.R.; Kravanja, Z.; Čuček, L. Global projections of plastic use, end-of-life fate and potential changes in consumption, reduction, recycling and replacement with bioplastics to 2050. Sustainable Production and Consumption 2024, 51, 498–518. [Google Scholar] [CrossRef]
- Kibria, M.G.; Masuk, N.I.; Safayet, R.; Nguyen, H.Q.; Mourshed, M. Plastic waste: challenges and opportunities to mitigate pollution and effective management. International Journal of Environmental Research 2023, 17, 20. [Google Scholar] [CrossRef]
- Duis, K.; Coors, A. Microplastics in the aquatic and terrestrial environment: sources (with a specific focus on personal care products), fate and effects. Environmental Sciences Europe 2016, 28, 2. [Google Scholar] [CrossRef]
- Emenike, E.C.; Okorie, C.J.; Ojeyemi, T.; Egbemhenghe, A.; Iwuozor, K.O.; Saliu, O.D.; Okoro, H.K.; Adeniyi, A.G. From oceans to dinner plates: The impact of microplastics on human health. Heliyon 2023, 9. [Google Scholar] [CrossRef]
- Jeswani, H.; Krüger, C.; Russ, M.; Horlacher, M.; Antony, F.; Hann, S.; Azapagic, A. Life cycle environmental impacts of chemical recycling via pyrolysis of mixed plastic waste in comparison with mechanical recycling and energy recovery. Sci. Total Environ. 2021, 769, 144483. [Google Scholar] [CrossRef]
- Fayshal, M.A. Current practices of plastic waste management, environmental impacts, and potential alternatives for reducing pollution and improving management. Heliyon 2024, 10. [Google Scholar] [CrossRef]
- Wojnowska-Baryła, I.; Bernat, K.; Zaborowska, M. Plastic waste degradation in landfill conditions: the problem with microplastics, and their direct and indirect environmental effects. Int. J. Env. Res. Public Health 2022, 19, 13223. [Google Scholar] [CrossRef] [PubMed]
- Cai, Z.; Li, M.; Zhu, Z.; Wang, X.; Huang, Y.; Li, T.; Gong, H.; Yan, M. Biological degradation of plastics and microplastics: a recent perspective on associated mechanisms and influencing factors. Microorganisms 2023, 11, 1661. [Google Scholar] [CrossRef]
- Wu, Y.; Hu, Q.; Che, Y.; Niu, Z. Opportunities and challenges for plastic depolymerization by biomimetic catalysis. Chemical Science 2024. [CrossRef] [PubMed]
- Hahn, S.; Hennecke, D. What can we learn from biodegradation of natural polymers for regulation? Environmental Sciences Europe 2023, 35, 50. [Google Scholar] [CrossRef]
- Roy, A.; Chakraborty, S. A comprehensive review on sustainable approach for microbial degradation of plastic and it’s challenges. Sustainable Chemistry for the Environment 2024, 100153. [Google Scholar] [CrossRef]
- Dhali, S.L.; Parida, D.; Kumar, B.; Bala, K. Recent trends in microbial and enzymatic plastic degradation: a solution for plastic pollution predicaments. Biotechnology for Sustainable Materials 2024, 1, 11. [Google Scholar] [CrossRef]
- Yoshida, S.; Hiraga, K.; Takehana, T.; Taniguchi, I.; Yamaji, H.; Maeda, Y.; Toyohara, K.; Miyamoto, K.; Kimura, Y.; Oda, K. A bacterium that degrades and assimilates poly (ethylene terephthalate). Science 2016, 351, 1196–1199. [Google Scholar] [CrossRef]
- Temporiti, M.E.E.; Nicola, L.; Nielsen, E.; Tosi, S. Fungal enzymes involved in plastics biodegradation. Microorganisms 2022, 10, 1180. [Google Scholar] [CrossRef]
- Retnadhas, S.; Ducat, D.C.; Hegg, E.L. Nature-Inspired Strategies for Sustainable Degradation of Synthetic Plastics. JACS Au 2024, 4, 3323–3339. [Google Scholar] [CrossRef]
- Suresh, V.; Shams, R.; Dash, K.K.; Shaikh, A.M.; Béla, K. Comprehensive Review on Enzymatic Polymer Degradation: A Sustainable Solution for Plastics. Journal of Agriculture and Food Research 2025, 101788. [Google Scholar] [CrossRef]
- Yao, J.; Liu, Y.; Gu, Z.; Zhang, L.; Guo, Z. Deconstructing PET: Advances in enzyme engineering for sustainable plastic degradation. Chem. Eng. J. 2024, 154183. [Google Scholar] [CrossRef]
- Joho, Y.; Vongsouthi, V.; Gomez, C.; Larsen, J.S.; Ardevol, A.; Jackson, C.J. Improving plastic degrading enzymes via directed evolution. Protein Engineering, Design and Selection 2024, gzae009. [Google Scholar] [CrossRef] [PubMed]
- Hong, H.; Ki, D.; Seo, H.; Park, J.; Jang, J.; Kim, K.-J. Discovery and rational engineering of PET hydrolase with both mesophilic and thermophilic PET hydrolase properties. Nat. Commun. 2023, 14, 4556. [Google Scholar] [CrossRef] [PubMed]
- Stevensen, J.; Janatunaim, R.Z.; Ratnaputri, A.H.; Aldafa, S.H.; Rudjito, R.R.; Saputro, D.H.; Suhandono, S.; Putri, R.M.; Aditama, R.; Fibriani, A. Thermostability and Activity Improvements of PETase from Ideonella sakaiensis. ACS Omega 2025. [Google Scholar] [CrossRef]
- Oda, K.; Wlodawer, A. Development of enzyme-based approaches for recycling PET on an industrial scale. Biochemistry 2024, 63, 369–401. [Google Scholar] [CrossRef]
- Bhattacharjee, U.; Baruah, K.N.; Shah, M.P. Exploring sustainable strategies for mitigating microplastic contamination in soil, water, and the food chain: A comprehensive analysis. Environmental Chemistry and Ecotoxicology 2025. [Google Scholar] [CrossRef]
- Okal, E.J.; Heng, G.; Magige, E.A.; Khan, S.; Wu, S.; Ge, Z.; Zhang, T.; Mortimer, P.E.; Xu, J. Insights into the mechanisms involved in the fungal degradation of plastics. Ecotoxicol. Environ. Saf. 2023, 262, 115202. [Google Scholar] [CrossRef]
- Gricajeva, A.; Nadda, A.K.; Gudiukaite, R. Insights into polyester plastic biodegradation by carboxyl ester hydrolases. J. Chem. Technol. Biotechnol. 2022, 97, 359–380. [Google Scholar] [CrossRef]
- Salas, R.; Villa, R.; Cano, S.; Nieto, S.; García-Verdugo, E.; Lozano, P. Biocatalytic hydrolysis of di-urethane model compounds in ionic liquid reaction media. Catal. Today 2024, 430, 114516. [Google Scholar] [CrossRef]
- Mester, T.; Tien, M. Oxidation mechanism of ligninolytic enzymes involved in the degradation of environmental pollutants. Int. Biodeterior. Biodegrad. 2000, 46, 51–59. [Google Scholar] [CrossRef]
- Kumar, A.; Chandra, R. Ligninolytic enzymes and its mechanisms for degradation of lignocellulosic waste in environment. Heliyon 2020, 6. [Google Scholar] [CrossRef] [PubMed]
- Tiso, T.; Winter, B.; Wei, R.; Hee, J.; de Witt, J.; Wierckx, N.; Quicker, P.; Bornscheuer, U.T.; Bardow, A.; Nogales, J. The metabolic potential of plastics as biotechnological carbon sources–review and targets for the future. Metab. Eng. 2022, 71, 77–98. [Google Scholar] [CrossRef]
- Mudondo, J.; Lee, H.-S.; Jeong, Y.; Kim, T.H.; Kim, S.; Sung, B.H.; Park, S.-H.; Park, K.; Cha, H.G.; Yeon, Y.J. Recent advances in the chemobiological upcycling of polyethylene terephthalate (PET) into value-added chemicals. J. Microbiol. Biotechnol. 2022, 33, 1. [Google Scholar] [CrossRef]
- Liu, C.; Shi, C.; Zhu, S.; Wei, R.; Yin, C.-C. Structural and functional characterization of polyethylene terephthalate hydrolase from Ideonella sakaiensis. Biochem. Biophys. Res. Commun. 2019, 508, 289–294. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, M.V.D.; Calandrini, G.; da Costa, C.H.S.; da Silva de Souza, C.G.; Alves, C.N.; Silva, J.R.A.; Lima, A.H.; Lameira, J. Evaluating cutinase from Fusarium oxysporum as a biocatalyst for the degradation of nine synthetic polymer. Sci. Rep. 2025, 15, 2887. [Google Scholar] [CrossRef] [PubMed]
- Jbilou, F.; Dole, P.; Degraeve, P.; Ladavière, C.; Joly, C. A green method for polybutylene succinate recycling: Depolymerization catalyzed by lipase B from Candida antarctica during reactive extrusion. Eur. Polym. J. 2015, 68, 207–215. [Google Scholar] [CrossRef]
- Khairul Anuar, N.F.S.; Huyop, F.; Ur-Rehman, G.; Abdullah, F.; Normi, Y.M.; Sabullah, M.K.; Abdul Wahab, R. An Overview into Polyethylene Terephthalate (PET) Hydrolases and Efforts in Tailoring Enzymes for Improved Plastic Degradation. International Journal of Molecular Sciences 2022, 23, 12644. [Google Scholar] [CrossRef]
- Hu, X.; Gao, Z.; Wang, Z.; Su, T.; Yang, L.; Li, P. Enzymatic degradation of poly(butylene succinate) by cutinase cloned from Fusarium solani. Polym. Degradation Stab. 2016, 134, 211–219. [Google Scholar] [CrossRef]
- Ravi, J.; Ponnuraj, K.; Ragunathan, P. Enzymatic biodegradation of Poly(ε-Caprolactone) (PCL) by a thermostable cutinase from a mesophilic bacteria Mycobacterium marinum. Sci. Total Environ. 2025, 972, 179066. [Google Scholar] [CrossRef]
- Hu, T.; Wang, Y.; Ma, L.; Wang, Z.; Tong, H. Biodegradation of polybutylene succinate by an extracellular esterase from Pseudomonas mendocina. Int. Biodeterior. Biodegrad. 2024, 195, 105910. [Google Scholar] [CrossRef]
- Shalem, A.; Yehezkeli, O.; Fishman, A. Enzymatic degradation of polylactic acid (PLA). Appl. Microbiol. Biotechnol. 2024, 108, 413. [Google Scholar] [CrossRef] [PubMed]
- Blázquez-Sánchez, P.; Engelberger, F.; Cifuentes-Anticevic, J.; Sonnendecker, C.; Griñén, A.; Reyes, J.; Díez, B.; Guixé, V.; Richter, P.K.; Zimmermann, W.; et al. Antarctic Polyester Hydrolases Degrade Aliphatic and Aromatic Polyesters at Moderate Temperatures. Applied and Environmental Microbiology 2022, 88, e01842-21. [Google Scholar] [CrossRef] [PubMed]
- Puiggené, Ò.; Espinosa, M.J.C.; Schlosser, D.; Thies, S.; Jehmlich, N.; Kappelmeyer, U.; Schreiber, S.; Wibberg, D.; Kalinowski, J.; Harms, H.; Heipieper, H.J.; et al. Extracellular degradation of a polyurethane oligomer involving outer membrane vesicles and further insights on the degradation of 2,4-diaminotoluene in Pseudomonas capeferrum TDA1. Sci. Rep. 2022, 12, 2666. [Google Scholar] [CrossRef]
- Akutsu, Y.; Nakajima-Kambe, T.; Nomura, N.; Nakahara, T. Purification and Properties of a Polyester Polyurethane-Degrading Enzyme from Comamonas acidovorans TB-35. Appl. Environ. Microbiol. 1998, 64, 62–67. [Google Scholar] [CrossRef]
- Nakajima-Kambe, T.; Shigeno-Akutsu, Y.; Nomura, N.; Onuma, F.; Nakahara, T. Microbial degradation of polyurethane, polyester polyurethanes and polyether polyurethanes. Appl. Microbiol. Biotechnol. 1999, 51, 134–140. [Google Scholar] [CrossRef]
- Yang, X.G.; Wen, P.P.; Yang, Y.F.; Jia, P.P.; Li, W.G.; Pei, D.S. Plastic biodegradation by in vitro environmental microorganisms and in vivo gut microorganisms of insects. Front Microbiol 2022, 13, 1001750. [Google Scholar]
- Solanki, P.; Putatunda, C.; Kumar, A.; Bhatia, R.; Walia, A. Microbial proteases: ubiquitous enzymes with innumerable uses. 3 Biotech 2021, 11, 428. [Google Scholar] [CrossRef]
- Bautista-Zamudio, P.A.; Flórez-Restrepo, M.A.; López-Legarda, X.; Monroy-Giraldo, L.C.; Segura-Sánchez, F. Biodegradation of plastics by white-rot fungi: A review. Sci. Total Environ. 2023, 901, 165950. [Google Scholar] [CrossRef]
- Srikanth, M.; Sandeep, T.; Sucharitha, K.; Godi, S. Biodegradation of plastic polymers by fungi: a brief review. Bioresources and bioprocessing 2022, 9, 42. [Google Scholar] [CrossRef]
- Ehara, K.; Iiyoshi, Y.; Tsutsumi, Y.; Nishida, T. Polyethylene degradation by manganese peroxidase in the absence of hydrogen peroxide. J. Wood Sci. 2000, 46, 180–183. [Google Scholar] [CrossRef]
- Zhang, Y.; Pedersen, J.N.; Eser, B.E.; Guo, Z. Biodegradation of polyethylene and polystyrene: From microbial deterioration to enzyme discovery. Biotechnol. Adv. 2022, 60, 107991. [Google Scholar] [CrossRef] [PubMed]
- Tahir, L.; Ishtiaq Ali, M.; Zia, M.; Atiq, N.; Hasan, F.; Ahmed, S. Production and characterization of esterase in Lantinus tigrinus for degradation of polystyrene. Polish J. Microbiol. 2013, 62, 101–108. [Google Scholar] [CrossRef]
- Niu, Q.; Meng, Q.; Yang, H.; Wang, Y.; Li, X.; Li, G.; Li, Q. Humification process and mechanisms investigated by Fenton-like reaction and laccase functional expression during composting. Bioresour. Technol. 2021, 341, 125906. [Google Scholar] [CrossRef] [PubMed]
- Negoro, S. Biodegradation of nylon oligomers. Appl. Microbiol. Biotechnol. 2000, 54, 461–466. [Google Scholar] [CrossRef]
- Lv, S.; Li, Y.; Zhao, S.; Shao, Z. Biodegradation of Typical Plastics: From Microbial Diversity to Metabolic Mechanisms. International Journal of Molecular Sciences 2024, 25, 593. [Google Scholar] [CrossRef]
- Warhurst, A.; Fewson, C. Microbial metabolism and biotransformation of styrene. J. Appl. Bacteriol. 1994, 77, 597–606. [Google Scholar] [CrossRef]
- Johnston, B.; Adamus, G.; Ekere, A.I.; Kowalczuk, M.; Tchuenbou-Magaia, F.; Radecka, I. Bioconversion of Plastic Waste Based on Mass Full Carbon Backbone Polymeric Materials to Value-Added Polyhydroxyalkanoates (PHAs). Bioengineering 2022, 9. [Google Scholar] [CrossRef]
- Jeon, H.J.; Kim, M.N. Functional analysis of alkane hydroxylase system derived from Pseudomonas aeruginosa E7 for low molecular weight polyethylene biodegradation. Int. Biodeterior. Biodegrad. 2015, 103, 141–146. [Google Scholar] [CrossRef]
- Rong, Z.; Ding, Z.-H.; Wu, Y.-H.; Xu, X.-W. Degradation of low-density polyethylene by the bacterium Rhodococcus sp. C-2 isolated from seawater. Sci. Total Environ. 2024, 907, 167993. [Google Scholar] [CrossRef]
- Jin, J.; Arciszewski, J.; Auclair, K.; Jia, Z. Enzymatic polyethylene biorecycling: Confronting challenges and shaping the future. J. Hazard. Mater. 2023, 460, 132449. [Google Scholar] [CrossRef]
- Tao, X.; Ouyang, H.; Zhou, A.; Wang, D.; Matlock, H.; Morgan, J.S.; Ren, A.T.; Mu, D.; Pan, C.; Zhu, X.; et al. Polyethylene Degradation by a Rhodococcous Strain Isolated from Naturally Weathered Plastic Waste Enrichment. Environ. Sci. Technol. 2023, 57, 13901–13911. [Google Scholar] [CrossRef] [PubMed]
- Restrepo-Flórez, J.-M.; Bassi, A.; Thompson, M.R. Microbial degradation and deterioration of polyethylene – A review. Int. Biodeterior. Biodegrad. 2014, 88, 83–90. [Google Scholar] [CrossRef]
- Amobonye, A.; Bhagwat, P.; Singh, S.; Pillai, S. Plastic biodegradation: Frontline microbes and their enzymes. Sci. Total Environ. 2021, 759, 143536. [Google Scholar] [CrossRef] [PubMed]
- Sanluis-Verdes, A.; Colomer-Vidal, P.; Rodriguez-Ventura, F.; Bello-Villarino, M.; Spinola-Amilibia, M.; Ruiz-Lopez, E.; Illanes-Vicioso, R.; Castroviejo, P.; Aiese Cigliano, R.; Montoya, M.; et al. Wax worm saliva and the enzymes therein are the key to polyethylene degradation by Galleria mellonella. Nat. Commun. 2022, 13, 5568. [Google Scholar] [CrossRef]
- Spínola-Amilibia, M.; Illanes-Vicioso, R.; Ruiz-López, E.; Colomer-Vidal, P.; Rodriguez-Ventura, F.; Peces Pérez, R.; Arias, C.F.; Torroba, T.; Solà, M.; Arias-Palomo, E.; et al. Plastic degradation by insect hexamerins: Near-atomic resolution structures of the polyethylene-degrading proteins from the wax worm saliva. Science Advances 2023, 9, eadi6813. [Google Scholar] [CrossRef]
- Tournier, V.; Topham, C.M.; Gilles, A.; David, B.; Folgoas, C.; Moya-Leclair, E.; Kamionka, E.; Desrousseaux, M.L.; Texier, H.; Gavalda, S.; et al. An engineered PET depolymerase to break down and recycle plastic bottles. Nature 2020, 580, 216–219. [Google Scholar] [CrossRef]
- Sabari V L, D.; Rajmohan, G.; S B, R.; S, S.; Nagasubramanian, K.; G, S.K.; Venkatachalam, P. Improving the binding affinity of plastic degrading cutinase with polyethylene terephthalate (PET) and polyurethane (PU); an in-silico study. Heliyon 2025, 11. [Google Scholar] [CrossRef]
- Roberts, C.; Edwards, S.; Vague, M.; León-Zayas, R.; Scheffer, H.; Chan, G.; Swartz Natasja, A.; Mellies Jay, L. Environmental Consortium Containing Pseudomonas and Bacillus Species Synergistically Degrades Polyethylene Terephthalate Plastic. mSphere 2020, 5. [Google Scholar] [CrossRef]
- Muringayil Joseph, T.; Azat, S.; Ahmadi, Z.; Moini Jazani, O.; Esmaeili, A.; Kianfar, E.; Haponiuk, J.; Thomas, S. Polyethylene terephthalate (PET) recycling: A review. Case Studies in Chemical and Environmental Engineering 2024, 9, 100673. [Google Scholar] [CrossRef]
- Li, S.; Liu, L.; Garreau, H.; Vert, M. Lipase-Catalyzed Biodegradation of Poly(ε-caprolactone) Blended with Various Polylactide-Based Polymers. Biomacromolecules 2003, 4, 372–377. [Google Scholar] [CrossRef]
- Howard, G.T. Biodegradation of polyurethane: a review. Int. Biodeterior. Biodegrad. 2002, 49, 245–252. [Google Scholar] [CrossRef]
- Dang, T.C.H.; Nguyen, D.T.; Thai, H.; Nguyen, T.C.; Hien Tran, T.T.; Le, V.H.; Nguyen, V.H.; Tran, X.B.; Thao Pham, T.P.; Nguyen, T.G.; et al. Plastic degradation by thermophilic Bacillus sp. BCBT21 isolated from composting agricultural residual in Vietnam. Advances in Natural Sciences: Nanoscience and Nanotechnology 2018, 9, 015014. [Google Scholar] [CrossRef]
- Santo, M.; Weitsman, R.; Sivan, A. The role of the copper-binding enzyme – laccase – in the biodegradation of polyethylene by the actinomycete Rhodococcus ruber. Int. Biodeterior. Biodegrad. 2013, 84, 204–210. [Google Scholar] [CrossRef]
- Iiyoshi, Y.; Tsutsumi, Y.; Nishida, T. Polyethylene degradation by lignin-degrading fungi and manganese peroxidase. J. Wood Sci. 1998, 44, 222–229. [Google Scholar] [CrossRef]
- Ru, J.; Huo, Y.; Yang, Y. Microbial Degradation and Valorization of Plastic Wastes. Front Microbiol 2020, 11. [Google Scholar] [CrossRef]
- Kim, H.R.; Lee, H.M.; Yu, H.C.; Jeon, E.; Lee, S.; Li, J.; Kim, D.H. Biodegradation of Polystyrene by Pseudomonas sp. Isolated from the Gut of Superworms (Larvae of Zophobas atratus). Environ. Sci. Technol. 2020, 54, 6987–6996. [Google Scholar] [CrossRef]
- Negoro, S.; Taniguchi, T.; Kanaoka, M.; Kimura, H.; Okada, H. Plasmid-determined enzymatic degradation of nylon oligomers. J. Bacteriol. 1983, 155, 22–31. [Google Scholar] [CrossRef]
- Stern, R.V.; Howard, G.T. The polyester polyurethanase gene (pueA) from Pseudomonas chlororaphis encodes a lipase. FEMS Microbiol. Lett. 2000, 185, 163–168. [Google Scholar] [CrossRef]
- Howard, G.T.; Mackie, R.I.; Cann, I.K.; Ohene-Adjei, S.; Aboudehen, K.S.; Duos, B.G.; Childers, G.W. Effect of insertional mutations in the pueA and pueB genes encoding two polyurethanases in Pseudomonas chlororaphis contained within a gene cluster. J. Appl. Microbiol. 2007, 103, 2074–2083. [Google Scholar] [CrossRef]
- Werner, A.Z.; Avina, Y.-S. C.; Johnsen, J.; Bratti, F.; Alt, H.M.; Mohamed, E.T.; Clare, R.; Mand, T.D.; Guss, A.M.; Feist, A.M.; et al. Adaptive laboratory evolution and genetic engineering improved terephthalate utilization in Pseudomonas putida KT2440. Metab. Eng. 2025, 88, 196–205. [Google Scholar] [CrossRef]
- Bao, T.; Qian, Y.; Xin, Y.; Collins, J.J.; Lu, T. Engineering microbial division of labor for plastic upcycling. Nat. Commun. 2023, 14, 5712. [Google Scholar] [CrossRef] [PubMed]
- Sivan, A.; Szanto, M.; Pavlov, V. Biofilm development of the polyethylene-degrading bacterium Rhodococcus ruber. Appl. Microbiol. Biotechnol. 2006, 72, 346–352. [Google Scholar] [CrossRef] [PubMed]
- Yao, Z.; Seong, H.J.; Jang, Y.-S. Degradation of low density polyethylene by Bacillus species. Applied Biological Chemistry 2022, 65, 84. [Google Scholar] [CrossRef]
- Yun, S.-D.; Lee, C.O.; Kim, H.-W.; An, S.J.; Kim, S.; Seo, M.-J.; Park, C.; Yun, C.-H.; Chi, W.S.; Yeom, S.-J. Exploring a New Biocatalyst from Bacillus thuringiensis JNU01 for Polyethylene Biodegradation. Environmental Science & Technology Letters 2023, 10, 485–492. [Google Scholar]
- Shah, Z.; Krumholz, L.; Aktas, D.F.; Hasan, F.; Khattak, M.; Shah, A.A. Degradation of polyester polyurethane by a newly isolated soil bacterium, Bacillus subtilis strain MZA-75. Biodegradation 2013, 24, 865–877. [Google Scholar] [CrossRef]
- Dong, C.; Wang, Z.; Shao, Z. Degradation from hydrocarbons to synthetic plastics: the roles and biotechnological potential of the versatile Alcanivorax in the marine blue circular economy. Blue Biotechnology 2024, 1, 14. [Google Scholar] [CrossRef]
- Zadjelovic, V.; Erni-Cassola, G.; Obrador-Viel, T.; Lester, D.; Eley, Y.; Gibson, M.I.; Dorador, C.; Golyshin, P.N.; Black, S.; Wellington, E.M.H.; et al. A mechanistic understanding of polyethylene biodegradation by the marine bacterium Alcanivorax. J. Hazard. Mater. 2022, 436, 129278. [Google Scholar] [CrossRef]
- Zhai, X.; Zhang, X.H.; Yu, M. Microbial colonization and degradation of marine microplastics in the plastisphere: A review. Front Microbiol 2023, 14, 1127308. [Google Scholar] [CrossRef]
- Khatua, S.; Simal-Gandara, J.; Acharya, K. Myco-remediation of plastic pollution: current knowledge and future prospects. Biodegradation 2024, 35, 249–279. [Google Scholar] [CrossRef]
- Tahir, L.; Ishtiaq Ali, M.; Zia, M.; Atiq, N.; Hasan, F.; Ahmed, S. Production and characterization of esterase in Lantinus tigrinus for degradation of polystyrene. Pol. J. Microbiol. 2013, 62, 101–108. [Google Scholar] [CrossRef]
- Černoša, A.; Cortizas, A.M.; Traoré, M.; Podlogar, M.; Danevčič, T.; Gunde-Cimerman, N.; Gostinčar, C. A screening method for plastic-degrading fungi. Heliyon 2024, 10, e31130. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Yang, J.; Wu, W.M.; Zhao, J.; Song, Y.; Gao, L.; Yang, R.; Jiang, L. Biodegradation and Mineralization of Polystyrene by Plastic-Eating Mealworms: Part 2. Role of Gut Microorganisms. Environ. Sci. Technol. 2015, 49, 12087–12093. [Google Scholar] [CrossRef] [PubMed]
- Park, J.-W.; Kim, M.; Kim, S.-Y.; Bae, J.; Kim, T.-J. Biodegradation of polystyrene by intestinal symbiotic bacteria isolated from mealworms, the larvae of Tenebrio molitor. Heliyon 2023, 9, e17352. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Su, T.; Zhao, J.; Wang, Z. Biodegradation of Polystyrene by Tenebrio molitor, Galleria mellonella, and Zophobas atratus Larvae and Comparison of Their Degradation Effects. Polymers 2021, 13, 3539. [Google Scholar] [CrossRef]
- Quartinello, F.; Kremser, K.; Schoen, H.; Tesei, D.; Ploszczanski, L.; Nagler, M.; Podmirseg, S.M.; Insam, H.; Piñar, G.; Sterflingler, K.; et al. Together Is Better: The Rumen Microbial Community as Biological Toolbox for Degradation of Synthetic Polyesters. Frontiers in Bioengineering and Biotechnology 2021, 9. [Google Scholar] [CrossRef]
- Benavides Fernández, C.D.; Guzmán Castillo, M.P.; Quijano Pérez, S.A.; Carvajal Rodríguez, L.V. Microbial degradation of polyethylene terephthalate: a systematic review. SN Applied Sciences 2022, 4, 263. [Google Scholar] [CrossRef]
- Lin, X.; Zhang, R.; Cui, F.-Q.; Hong, W.; Yang, S.; Ju, F.; Xi, C.; Sun, X.; Song, L. Natural-selected plastics biodegradation species and enzymes in landfills. PNAS Nexus 2025, 4. [Google Scholar] [CrossRef]
- Jahanshahi, D.A.; Barzani, M.R.R.; Bahram, M.; Ariaeenejad, S.; Kavousi, K. Metagenomic exploration and computational prediction of novel enzymes for polyethylene terephthalate degradation. Ecotoxicol. Environ. Saf. 2025, 289, 117640. [Google Scholar] [CrossRef]
- Viljakainen, V.R.; Hug, L.A. New approaches for the characterization of plastic-associated microbial communities and the discovery of plastic-degrading microorganisms and enzymes. Comput Struct Biotechnol J 2021, 19, 6191–6200. [Google Scholar] [CrossRef]
- Barclay, A.; Acharya, K.R. Engineering Plastic Eating Enzymes Using Structural Biology. Biomolecules 2023, 13. [Google Scholar] [CrossRef]
- Liu, Y.-J.; Zhou, J.; Li, Y.; Yan, X.; Xu, A.; Zhou, X.; Liu, W.; Xu, Y.; Su, T.; Wang, S.; et al. State-of-the-art advances in biotechnology for polyethylene terephthalate bio-depolymerization. Green Carbon 2025. [Google Scholar] [CrossRef]
- Wani, A.K.; Akhtar, N.; Naqash, N.; Rahayu, F.; Djajadi, D.; Chopra, C.; Singh, R.; Mulla, S.I.; Sher, F.; Américo-Pinheiro, J.H.P. Discovering untapped microbial communities through metagenomics for microplastic remediation: recent advances, challenges, and way forward. Environ. Sci. Pollut. Res. Int. 2023, 30, 81450–81473. [Google Scholar] [CrossRef] [PubMed]
- Zrimec, J.; Kokina, M.; Jonasson, S.; Zorrilla, F.; Zelezniak, A. Plastic-Degrading Potential across the Global Microbiome Correlates with Recent Pollution Trends. mBio 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Cao, K.; Liu, H.; Wang, Y.; Zhang, B.; Han, H.; Cui, Z.; Cao, H. Discovery of a polyester polyurethane-degrading bacterium from a coastal mudflat and identification of its degrading enzyme. J. Hazard. Mater. 2025, 483, 136659. [Google Scholar] [CrossRef] [PubMed]
- Danso, D.; Schmeisser, C.; Chow, J.; Zimmermann, W.; Wei, R.; Leggewie, C.; Li, X.; Hazen, T.; Streit, W. New Insights into the Function and Global Distribution of Polyethylene Terephthalate (PET)-Degrading Bacteria and Enzymes in Marine and Terrestrial Metagenomes. Applied and Environmental Microbiology 2018, 84, e02773-17. [Google Scholar] [CrossRef]
- Joo, S.; Cho, I.J.; Seo, H.; Son, H.F.; Sagong, H.-Y.; Shin, T.J.; Choi, S.Y.; Lee, S.Y.; Kim, K.-J. Structural insight into molecular mechanism of poly(ethylene terephthalate) degradation. Nat. Commun. 2018, 9, 382. [Google Scholar] [CrossRef]
- Austin, H.P.; Allen, M.D.; Donohoe, B.S.; Rorrer, N.A.; Kearns, F.L.; Silveira, R.L.; Pollard, B.C.; Dominick, G.; Duman, R.; El Omari, K.; et al. Characterization and engineering of a plastic-degrading aromatic polyesterase. Proc Natl Acad Sci U S A 2018, 115, E4350–E4357. [Google Scholar] [CrossRef]
- Knott, B.C.; Erickson, E.; Allen, M.D.; Gado, J.E.; Graham, R.; Kearns, F.L.; Pardo, I.; Topuzlu, E.; Anderson, J.J.; Austin, H.P.; et al. Characterization and engineering of a two-enzyme system for plastics depolymerization. Proc Natl Acad Sci U S A 2020, 117, 25476–25485. [Google Scholar] [CrossRef]
- Lu, H.; Diaz, D.J.; Czarnecki, N.J.; Zhu, C.; Kim, W.; Shroff, R.; Acosta, D.J.; Alexander, B.R.; Cole, H.O.; Zhang, Y.; et al. Machine learning-aided engineering of hydrolases for PET depolymerization. Nature 2022, 604, 662–667. [Google Scholar] [CrossRef]
- Stockinger, P.; Niederhauser, C.; Farnaud, S.; Buller, R. Computational analysis reveals temperature-induced stabilization of FAST-PETase. Comput. Struct. Biotechnol. J. 2025, 27, 969–977. [Google Scholar] [CrossRef]
- Furukawa, M.; Kawakami, N.; Tomizawa, A.; Miyamoto, K. Efficient Degradation of Poly(ethylene terephthalate) with Thermobifida fusca Cutinase Exhibiting Improved Catalytic Activity Generated using Mutagenesis and Additive-based Approaches. Sci. Rep. 2019, 9, 16038. [Google Scholar] [CrossRef] [PubMed]
- Araújo, R.; Silva, C.; O’Neill, A.; Micaelo, N.; Guebitz, G.; Soares, C.M.; Casal, M.; Cavaco-Paulo, A. Tailoring cutinase activity towards polyethylene terephthalate and polyamide 6,6 fibers. J. Biotechnol. 2007, 128, 849–857. [Google Scholar] [CrossRef] [PubMed]
- Camarero, S.; Pardo, I.; Cañas, A.I.; Molina, P.; Record, E.; Martínez, A.T.; Martínez, M.J.; Alcalde, M. Engineering platforms for directed evolution of Laccase from Pycnoporus cinnabarinus. Appl. Environ. Microbiol. 2012, 78, 1370–1384. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Samak, N.A.; Hao, X.; Chen, Z.; Yang, G.; Zhao, X.; Mu, T.; Yang, M.; Xing, J. Nano-immobilization of PETase enzyme for enhanced polyethylene terephthalate biodegradation. Biochem. Eng. J. 2021, 176, 108205. [Google Scholar] [CrossRef]
- Ren, Y.; Rivera, J.G.; He, L.; Kulkarni, H.; Lee, D.K.; Messersmith, P.B. Facile, high efficiency immobilization of lipase enzyme on magnetic iron oxide nanoparticles via a biomimetic coating. BMC Biotechnol. 2011, 11, 63. [Google Scholar] [CrossRef]
- Lu, D.; Wu, J.; Jin, S.; Wu, Q.; Deng, L.; Wang, F.; Nie, K. The enhancement of waste PET particles enzymatic degradation with a rotating packed bed reactor. Journal of Cleaner Production 2024, 434, 140088. [Google Scholar] [CrossRef]
- Sheldon, R.A.; van Pelt, S. Enzyme immobilisation in biocatalysis: why, what and how. Chem. Soc. Rev. 2013, 42, 6223–6235. [Google Scholar] [CrossRef]
- Qi, X.; Yan, W.; Cao, Z.; Ding, M.; Yuan, Y. Current Advances in the Biodegradation and Bioconversion of Polyethylene Terephthalate. Microorganisms 2021, 10. [Google Scholar] [CrossRef]
- Brandenberg, O.F.; Schubert, O.T.; Kruglyak, L. Towards synthetic PETtrophy: Engineering Pseudomonas putida for concurrent polyethylene terephthalate (PET) monomer metabolism and PET hydrolase expression. Microbial Cell Factories 2022, 21, 119. [Google Scholar] [CrossRef]
- Dissanayake, L.; Jayakody, L.N. Engineering Microbes to Bio-Upcycle Polyethylene Terephthalate. Frontiers in Bioengineering and Biotechnology 2021, 9. [Google Scholar] [CrossRef]
- Qi, X.; Ma, Y.; Chang, H.; Li, B.; Ding, M.; Yuan, Y. Evaluation of PET Degradation Using Artificial Microbial Consortia. Front Microbiol 2021, 12, 778828. [Google Scholar] [CrossRef] [PubMed]
- Alaghemandi, M. Sustainable Solutions Through Innovative Plastic Waste Recycling Technologies. Sustainability 2024, 16, 10401. [Google Scholar] [CrossRef]
- Highmoore, J.F.; Kariyawasam, L.S.; Trenor, S.R.; Yang, Y. Design of depolymerizable polymers toward a circular economy. Green Chem. 2024, 26, 2384–2420. [Google Scholar] [CrossRef]
- García-Meseguer, R.; Ortí, E.; Tuñón, I.; Ruiz-Pernía, J.J.; Aragó, J. Insights into the Enhancement of the Poly(ethylene terephthalate) Degradation by FAST-PETase from Computational Modeling. J. Am. Chem. Soc. 2023, 145, 19243–19255. [Google Scholar] [CrossRef]
- Zarezadeh, E.; Tangestani, M.; Jafari, A.J. A systematic review of methodologies and solutions for recycling polyurethane foams to safeguard the environment. Heliyon 2024, 10, e40724. [Google Scholar] [CrossRef]
- Ballerstedt, H.; Tiso, T.; Wierckx, N.; Wei, R.; Averous, L.; Bornscheuer, U.; O’Connor, K.; Floehr, T.; Jupke, A.; Klankermayer, J. MIXed plastics biodegradation and UPcycling using microbial communities: EU Horizon 2020 project MIX-UP started January 2020. Environmental Sciences Europe 2021, 33, 99. [Google Scholar] [CrossRef]
- Xiong, Z.; Zhang, Y.; Chen, X.; Sha, A.; Xiao, W.; Luo, Y.; Han, J.; Li, Q. Soil Microplastic Pollution and Microbial Breeding Techniques for Green Degradation: A Review. Microorganisms 2024, 12. [Google Scholar] [CrossRef]
- Mistry, A.N.; Kachenchart, B.; Wongthanaroj, A.; Somwangthanaroj, A.; Luepromchai, E. Rapid biodegradation of high molecular weight semi-crystalline polylactic acid at ambient temperature via enzymatic and alkaline hydrolysis by a defined bacterial consortium. Polym. Degradation Stab. 2022, 202, 110051. [Google Scholar] [CrossRef]
- Idris, S.N.; Amelia, T.S.M.; Bhubalan, K.; Lazim, A.M.M.; Zakwan, N.A.M.A.; Jamaluddin, M.I.; Santhanam, R.; Amirul, A.-A. A.; Vigneswari, S.; Ramakrishna, S. The degradation of single-use plastics and commercially viable bioplastics in the environment: A review. Environ. Res. 2023, 231, 115988. [Google Scholar] [CrossRef]
- Guicherd, M.; Ben Khaled, M.; Guéroult, M.; Nomme, J.; Dalibey, M.; Grimaud, F.; Alvarez, P.; Kamionka, E.; Gavalda, S.; Noël, M.; et al. An engineered enzyme embedded into PLA to make self-biodegradable plastic. Nature 2024, 631, 884–890. [Google Scholar] [CrossRef]
- Sun, S. Enzyme-Embedded Biodegradable Plastic for Sustainable Applications: Advances, Challenges, and Perspectives. ACS Applied Bio Materials 2025, 8, 1785–1796. [Google Scholar] [CrossRef] [PubMed]
- Kawai, F.; Furushima, Y.; Mochizuki, N.; Muraki, N.; Yamashita, M.; Iida, A.; Mamoto, R.; Tosha, T.; Iizuka, R.; Kitajima, S. Efficient depolymerization of polyethylene terephthalate (PET) and polyethylene furanoate by engineered PET hydrolase Cut190. AMB Express 2022, 12, 134. [Google Scholar] [CrossRef] [PubMed]
- Mandal, M.; Roy, A.; Popek, R.; Sarkar, A. Micro- and nano- plastic degradation by bacterial enzymes: A solution to ‘White Pollution’. The Microbe 2024, 3, 100072. [Google Scholar] [CrossRef]

Table 1.
Summary of Major Enzyme Classes in Plastic Biodegradation.
| Main Category | Enzyme Class | Plastic Targets |
|---|---|---|
| Hydrolytic Enzymes | Polyester Hydrolases | PET, biodegradable polyesters |
| Urethane Hydrolases Proteases/Ureases |
PU PU |
|
| Oxidative Enzymes | Laccases/Peroxidases | PE, PP, PS, PVC |
| Specialized and Auxiliary Enzymes | Styrene Monooxygenase | PS |
| MHETase | PET1 |
Note: 1 In combination with PETase.
Table 2.
Enzyme Classes, Target Plastic Substrates, and Representative Microbial Sources.
| Enzyme or Enzyme Class | Primary Plastic Substrate(s) | Source Microorganism(s) |
|---|---|---|
| PETase (PET hydrolase) | PET | I. sakaiensis (bacterium) [17] |
| MHETase (TPA hydrolase) | MHET | I. sakaiensis [17] |
| Cutinases (esterases) | PET; polyesters (e.g., PCL, PBS); cutin | Thermobifida spp. (actinomycetes) [20,66,67] F. solani (fungus) [3,20,68] |
| Lipases (esterases) | Aliphatic polyesters (PCL, PLA); minor action on PET |
C. antarctica (yeast) [36,69]; Pseudomonas spp. (bacteria) [70] |
| Polyurethane esterases | Polyester-based PU | Pseudomonas chlororaphis, Pseudomonas fluorescens (bacteria) [43,71] |
| PU ether hydrolase (e.g., PudA) | Polyether-PU | Pseudomonas sp. (membrane-bound with SBD) [43,71] |
| Proteases | PU (urethane/urea bonds) |
B. subtilis (bacterium) [47,72]; A. tubingensis (fungus) [47] |
| Urease | PU (carbamate/urea linkages) |
Penicillium sp. (fungus) [73]; Bacillus sp. (bacterium) [73] |
| Laccase (multicopper oxidase) | PE, PP, PVC (oxidative cleavage) |
T. versicolor (fungus) [73]; Rhodococcus ruber (bacterium) [73] |
| Manganese/Lignin Peroxidases | PE, PVC (oxidative cleavage) | P. chrysosporium (fungus) [74] |
| Alkane hydroxylase | PE (initiates alkane oxidation) |
R. ruber [62]; Pseudomonas aeruginosa [62] |
| Esterase (PS-depolymerase) | PS (polystyrene) | L. tigrinus (fungus) [75] |
| Phenol oxidases | PE (oxidative depolymerization) | Waxworm (G. mellonella) saliva enzymes [64] |
| Styrene monooxygenase | Styrene (from PS) | P. putida (bacterium) [76] |
| Nylon oligomer hydrolase | Nylon-6 oligomers | Flavobacterium sp. KI72 (bacterium) [77] |
Table 3.
Recent Advances in Plastic-Degrading Enzymes.
| Category | Key Advances | References |
|---|---|---|
| Discovery of New Enzymes |
Metagenomic surveys identified 30,000 enzyme homologs | [98,102,103] |
| PE-degrading waxworm enzymes discovered | [64] | |
| Novel PU-degrading Pseudomonas isolated. | [104] | |
| Protein Engineering | Structure-based design led to double-mutant PETase | [106,107] |
| LCC evolved to degrade PET in 10 hours | [66,101] | |
| Creation of PETase-MHETase “super-enzyme” for enhanced rate | [108] | |
| FAST-PETase developed using AI for enhanced stability and activity | [109,110] | |
| Engineered cutinases target PET, PEF, and polyamide | [111,112] | |
| Enzyme Immobilization | PETase immobilized on solid supports improves stability and reusability | [114] |
| lipases immobilized on nanoparticles improves stability | [115] | |
| Packed/fluidized-bed reactors for continuous waste treatment | [101,116] | |
| Pilot-scale enzymatic recycling demonstrated by Carbios. | [16,117] | |
| Synthetic Biology | Whole-cell systems to express PETase and metabolize monomers | [118,119,120] |
| Synthetic consortia for cooperative degradation and upcycling | [121] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



