Submitted:
08 April 2025
Posted:
08 April 2025
You are already at the latest version
Abstract
In the marine red alga Pyropia yezoensis, filamentous phases of the life cycle, e.g., the conchocelis (sporophyte) and conchosporangium (conchosporophyte), proliferate by tip growth. In this study, we investigated the possible involvement of phosphoinositide turnover and actin polymerization in the spontaneous initiation and tip growth of new branches in conchocelis cells. Treatment with LY294002 and U73122, specific inhibitors of phosphoinositide-phosphate 3-kinase and phospholipase C, respectively, reduced side-branch formation and inhibited elongation of branches. In addition, two inhibitors of the actin cytoskeleton, cytochalasin B (CCB) and latrunculin B (LAT-B), had similar effects on tip growth. However, CCB did not alter the branching rate of single-celled conchocelis, whereas LAT-B did. Since CCB and LAT-B affect actin polymerization by different mechanisms, this result suggests differences in the contributions of actin polymerization to branch initiation versus tip growth. These findings demonstrate the critical and diverse functional roles played by phosphoinositide turnover and actin polymerization in the regulation of the initiation and maintenance of tip growth in the conchocelis phase of P. yezoensis.
Keywords:
tip growth
; branch formation
; stem cell
; actin polymerization
; phosphoinositide-phosphate 3-kinase
; phospholipase C
; conchocelis
; Pyropia yezoensis
1. Introduction
Tip growth is a type of cellular growth occurring in filamentous bodies and tissues such as fungal hyphae, angiosperm pollen tubes and root hairs, moss protonemata, green algal rhizoids, and thali of filamentous brown algae [1-3]. The mechanisms regulating tip growth have been analyzed extensively in the tip-growing cells of angiosperms, namely, root hairs and pollen tubes that are non-dividing single cells with elongation occurring at the tip, and they involve ion fluxes, asymmetric distributions of F-actin and phosphoinositides, trafficking of membrane and cell wall materials, and production of reactive oxygen species and plant hormones [4-13]. Similar observations have been made for tip growth in hyphae and protonemata of filamentous multicellular organisms that grow via cell division restricted at the tip stem cells [1,2,4]. In single-cell and hyphae types of tip growth, modulations of ion fluxes, actin polymerization, phosphoinositide turnover, and membrane trafficking commonly occur at the apex of the growing cells, thereby establishing a restricted point of growth that polarizes growth direction and leads to tip growth [12,14,15].
The marine red alga Pyropia yezoensis (Bangiales, Rhodophyta) is a representative species for nori aquaculture in Asian countries such as Japan, South Korea, and China [16]. Despite being cultivated in industrial large-scale aquaculture systems at the sea surface, P. yezoensis is relatively poorly studied; thus, the mechanisms regulating its growth, life cycle, development, and environmental stress responses are mostly unknown. However, we have recently determined that tip growth is used for proliferation during filamentous phases of the P. yezoensis life cycle, e.g., the conchocelis (sporophyte) and the conchosporangium (conchosporophyte) [17-20]. In these phases, elongation and division occurs only in the apical cell of the filament, and each cell division produces two different cell types: a new apical cell at the tip of the filament and a neighboring differentiated non-dividing cell [17-20]. Similar hyphae-type tip growth was also observed in the gametophytic protonema of the moss Physcomitrium patens, where the tip cell is now recognized as a stem cell that divides to produce a copy of itself and a differentiated non-dividing cell [21]. Thus, tip cells of the conchocelis and conchosporangium of P. yezoensis also are thought to be stem cells. Therefore, elucidation of mechanisms regulating production and maintenance of the apical stem cell is fundamental for understanding tip growth as a growth mode of the two filamentous phases in the P. yezoensis life cycle.
In addition to polarized tip growth in the conchocelis and conchosporangia, directional migration of asexual spores from leafy gametophytes is polarity dependent in P. yezoensis [22-25]. Establishment of polarity along the anterior–posterior axis is regulated by phosphoinositide-phosphate 3-kinase (PI3K) and phospholipase C (PLC). PI3K phosphorylates a hydroxyl group (-OH group) at the 3rd position of the inositol ring of phosphoinositide (PtdIns), phosphoinositide 4-phosphate, phosphoinositide 5-phosphate, and phosphoinositide 4,5-bisphosphate [PtdIns(4,5)P2] to generate phosphoinositide 3-phosphate, phosphoinositide 3,4-bisphosphate, phosphoinositide 3,5-bisphosphate, and phosphoinositide 3,4,5-trisphosphate, respectively. PLC hydrolyzes PtdIns(4,5)P2 to produce two second messengers, inositol 1,4,5-trisphosphate and diacylglycerol [26,27]. Phosphoinositide turnover mediated by these enzymes is critical for the directional migration of asexual spores of P. yezoensis. In addition, F-actin is distributed asymmetrically to the forward side of migrating cells [22,24,25]. These findings led us to hypothesize that PI3K, PLC, and F-actin are involved in polarized tip growth in P. yezoensis.
We previously developed a novel experimental system using single-celled conchocelis for the study of tip growth in P. yezoensis [20]. This system enables us to identify factors involved in the generation of a branch initial from differentiated non-dividing cells, tip growth of the branch initial, and establishment and maintenance of the stem cell by cell elongation and cell division [20]. In this study, we addressed whether phosphoinositide turnover and actin polymerization are critical for tip growth in single-celled conchocelis from P. yezoensis. Our findings provide novel insights into the regulation of hyphae-type tip growth.
2. Results
2.1. Effects of PI3K and PLC Inhibitors
We first examined the possible involvement of PI3K and PLC in the generation of branch initials and their subsequent tip growth using chemical inhibitors of these enzymes. Single-celled conchocelis cells were treated with 0, 5, or 10 μM LY294002, a specific inhibitor of PI3K, or LY303511, its inactive analog, for 3 days. The branching rate and branch length were lower in the presence of LY294002 in a dose-dependent manner (Figure S1), with strong inhibition at 5 μM; however, LY303511 had no significant effect, even at 10 μM (Fig. 1). Similarly, a 3-day treatment of single-celled conchocelis with U73122, a specific inhibitor of PLC, inhibited both branching rate and branch length in a dose-dependent manner (Figure S2), whereas these effects were not observed after using U73433, an inactive analog of U73122 (Fig. 2). Thus, we concluded that PI3K and PLC activities are important for generation and tip growth of branch initials from differentiated conchocelis cells.
Figure 1.
Effects of a PI3K inhibitor on tip growth. (A–C) Photographs of single conchocelis cells treated with 0.5% DMSO as a control (A), 10 μM LY294002 (B), or its inactive analog 10 μM LY303511 (C) for 3 days. Arrows indicate newly generated branches. Bars: 25 μm. (D, E) Branching rate (D) and branch length (E) following treatment of single conchocelis cells with 0.5% DMSO (control), 10 μM LY294002, or 10 μM LY303511 for 3 days. Center line, median line; box limits, interquartile range with upper and lower quartiles; points, data; whiskers, range with maximum and minimum values; crosses, mean value. Lowercase letters denote significant differences in branching rate (D) and branch length (E) based on three independent experiments (n = 3) as determined by the Tukey-Kramer test (p < 0.05) for each set of treatments.
Figure 1.
Effects of a PI3K inhibitor on tip growth. (A–C) Photographs of single conchocelis cells treated with 0.5% DMSO as a control (A), 10 μM LY294002 (B), or its inactive analog 10 μM LY303511 (C) for 3 days. Arrows indicate newly generated branches. Bars: 25 μm. (D, E) Branching rate (D) and branch length (E) following treatment of single conchocelis cells with 0.5% DMSO (control), 10 μM LY294002, or 10 μM LY303511 for 3 days. Center line, median line; box limits, interquartile range with upper and lower quartiles; points, data; whiskers, range with maximum and minimum values; crosses, mean value. Lowercase letters denote significant differences in branching rate (D) and branch length (E) based on three independent experiments (n = 3) as determined by the Tukey-Kramer test (p < 0.05) for each set of treatments.
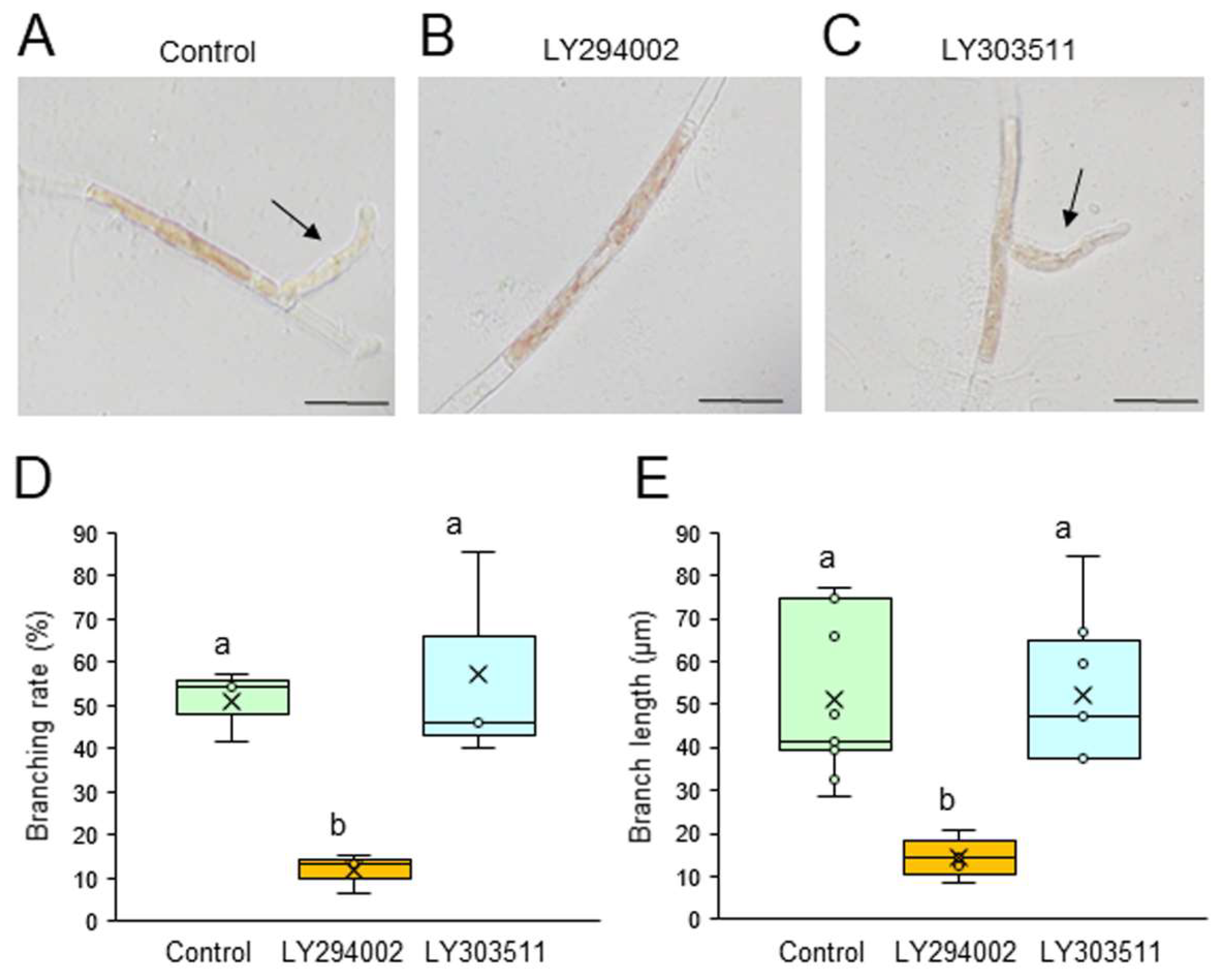
Figure 2.
Effects of a PLC inhibitor on tip growth. (A–C) Photographs of single conchocelis cells treated with 0.5% DMSO as a control (A), 0.1 μM U73122 (B), or its inactive analog 0.1 μM U73433 (C) for 3 days. Arrows indicate newly generated branches. Bars: 25 μm. (D, E) Branching rate (D) and branch length (E) following treatment of single conchocelis cells with 0.5% DMSO (control), 0.1 μM U73122, or 0.1 μM U73433 for 3 days. Center line, median line; box limits, interquartile range with upper and lower quartiles; points, data; whiskers, range with maximum and minimum values; crosses, mean value. Lowercase letters denote significant differences in branching rate (D) and branch length (E) based on three independent experiments (n = 3) as determined by the Tukey-Kramer test (p < 0.05) for each set of treatments.
Figure 2.
Effects of a PLC inhibitor on tip growth. (A–C) Photographs of single conchocelis cells treated with 0.5% DMSO as a control (A), 0.1 μM U73122 (B), or its inactive analog 0.1 μM U73433 (C) for 3 days. Arrows indicate newly generated branches. Bars: 25 μm. (D, E) Branching rate (D) and branch length (E) following treatment of single conchocelis cells with 0.5% DMSO (control), 0.1 μM U73122, or 0.1 μM U73433 for 3 days. Center line, median line; box limits, interquartile range with upper and lower quartiles; points, data; whiskers, range with maximum and minimum values; crosses, mean value. Lowercase letters denote significant differences in branching rate (D) and branch length (E) based on three independent experiments (n = 3) as determined by the Tukey-Kramer test (p < 0.05) for each set of treatments.
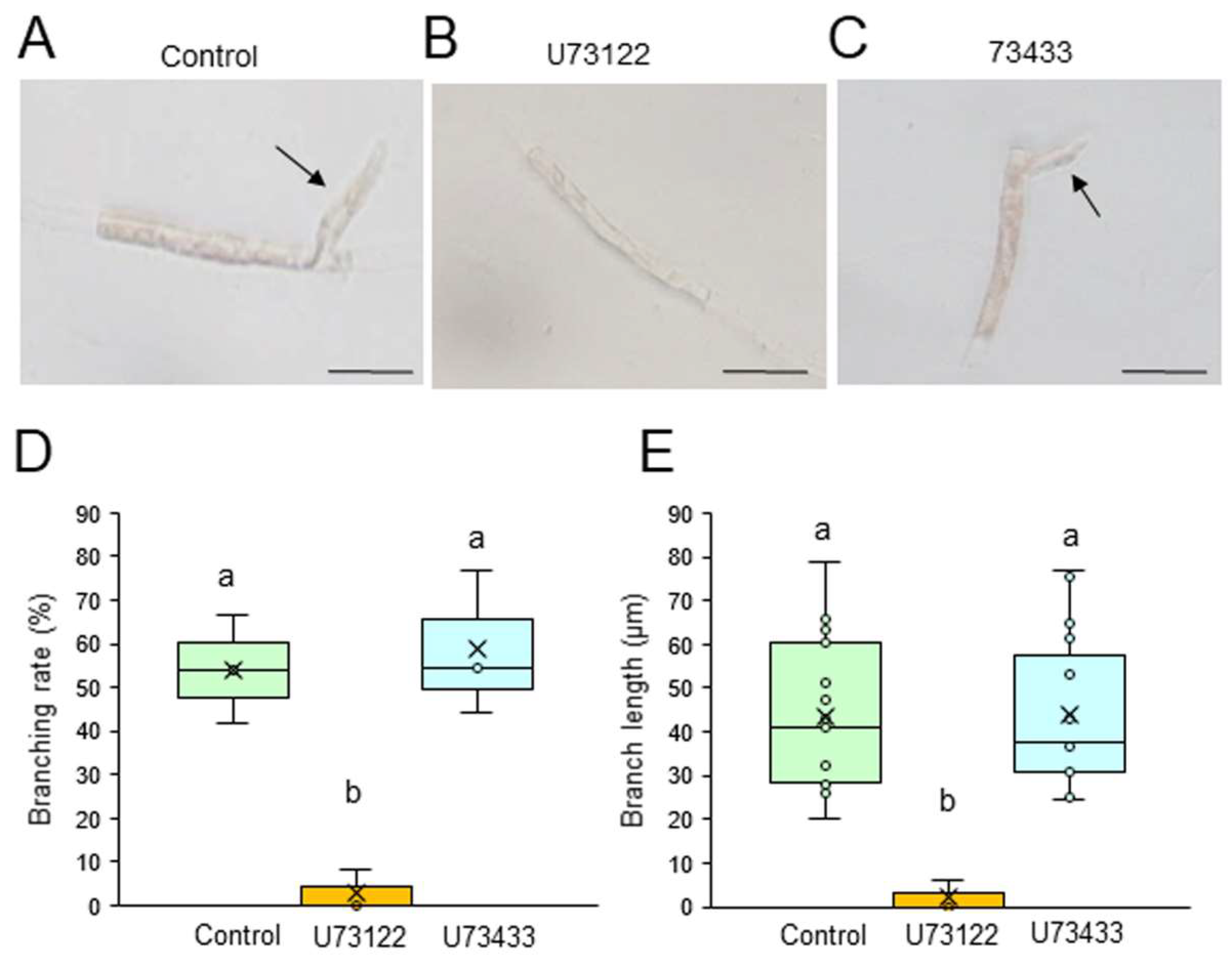
2.2. Effects of Actin Polymerization Inhibitors
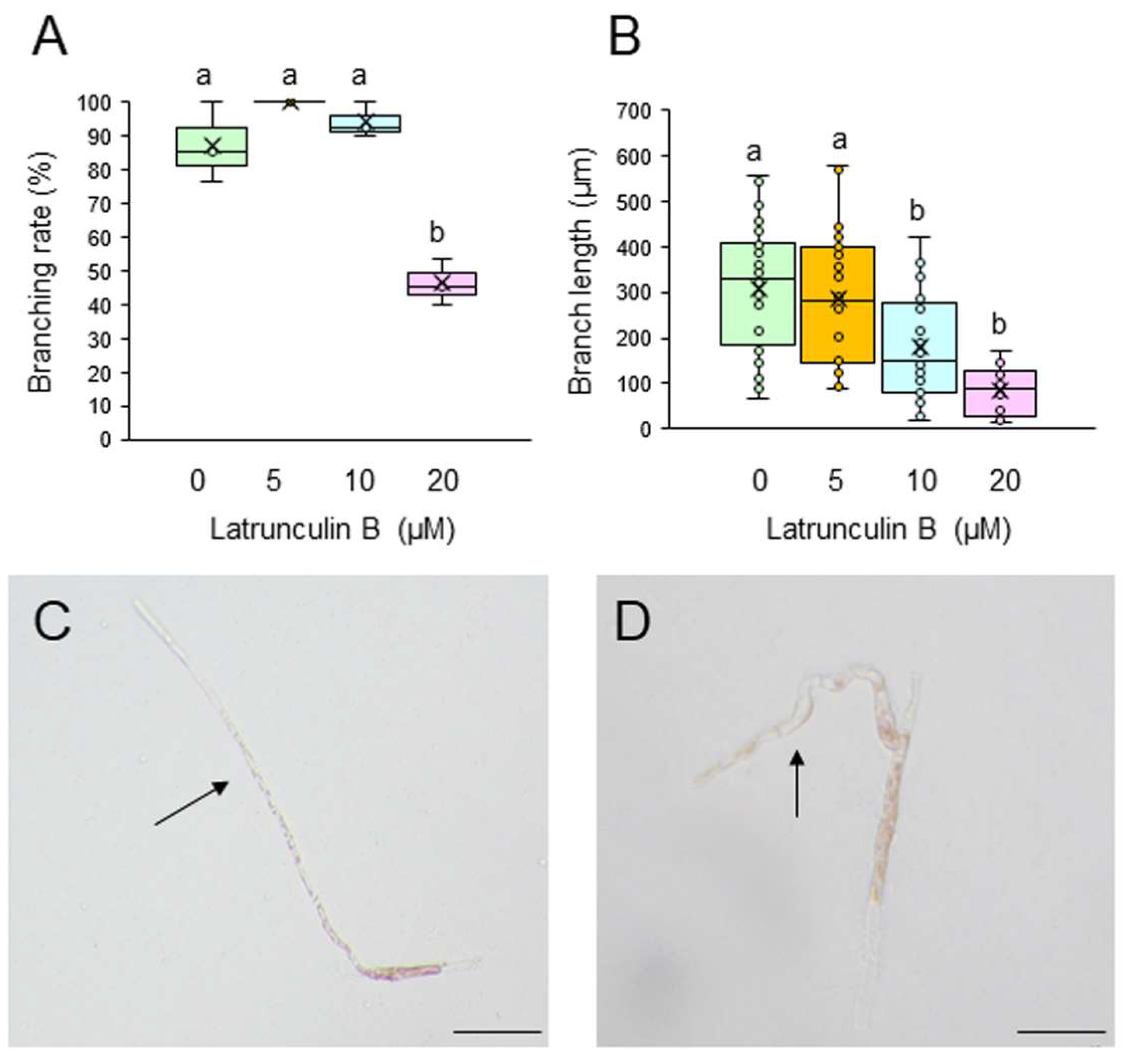
To test whether F-actin is involved in the generation and tip growth of branch initials, single-celled conchocelis was treated with different concentrations of the actin cytoskeleton inhibitors cytochalasin B (CCB) and latrunculin B (LAT-B). Incubation of single conchocelis cells with 10, 20, or 30 μM CCB for 7 days did not affect branching rate (Figure 3A). By contrast, the same treatments resulted in a shorter branch length (Figure 3B), and the branches displayed a wavy shape (compare Figure 3C and 3D). When the cells were incubated with 5, 10, or 20 μM LAT-B for 3 or 7 days, branching rate and branch length were inhibited in a dose-dependent manner (Figure 4A and 4B) and, as observed with the CCB treatments, branches were wavy (compare Figure 4C and 4D). When LAT-B was removed by washing the cells with seawater, we observed recovery of both branching rate and branch growth (Figure S3). Therefore, although the effects of the two inhibitors differed, our experimental results indicate the involvement of F-actin in generation of branch initials, branch tip growth, and establishment of stem cells in conchocelis cells.
3. Discussion
This study demonstrated that PI3K and PLC activities and actin polymerization have critical roles in the generation and tip growth of branch initials in single-celled conchocelis of P. yezoensis. These findings are consistent with the mechanisms of tip growth in the pollen tubes and root hairs of terrestrial plants, where more detailed analyses have been performed, including pharmacological inhibition of PI3K and PLC [28,29], overexpression and knockdown of genes encoding PLCs and PIP5Ks [28.30-36], and visualization of the subcellular location of phosphoinositide, PLC, and F-actin [28,37-41]. P. yezoensis lacks a reverse-genetic experimental system that could provide direct evidence for the functions of genes and the subcellular localization of gene products, which hinders functional analyses of PI3K, PLC, and actin polymerization [42-45].
Nevertheless, our findings provide novel information about the regulation of tip growth in algae.
Our observations of generation and tip growth of branch initials in single-celled conchocelis from P. yezoensis indicate that PI3K and PLC activities are involved in both processes , 2, S1, and S2). Thus, we infer that phosphoinositide turnover activates signal transduction pathways at the branching point and apex of the stem cells and that this is necessary for establishment and tip growth of branches in differentiated conchocelis cells. The cytoskeleton inhibitor CCB inhibited the growth of branches but not their initiation (Figure 3), whereas LAT-B inhibited both initiation and growth of branches and S3). In fact, tip growth in single-celled conchocelis can be divided into four sequential events: generation of the branch initial on a differentiated conchocelis cell, elongation of the branch initial, initial cell division to establish the apical stem cell, and ongoing branch elongation based on division of the tip stem cells. Since it is well known that CCB and LAT-B inhibit F-actin-based G-actin polymerization and access of G-actin to F-actin, respectively, we speculate that the generation of the branch initial involves relocalizing pre-existing F-actin at the branch position in differentiated non-dividing conchocelis cells and does not require additional actin polymerization. By contrast, establishment of the apical stem cell by the first cell division in the new branch and the tip growth of the stem cell require new production and/or further accumulation of F-actin. Accordingly, we propose that the regulatory mechanisms for establishment of the branch initial and tip growth of branches differ from those of hyphae-type tip growth, although F-actin, PI3K, and PLC are involved in both processes.
Despite the different effects of CCB and LAT-B on tip growth, treatment of single-celled conchocelis with either inhibitor resulted in abnormal growth of branches and a wavy shape (Figure 3D and 4D). A similar effect was observed after treatment of conchocelis cells with PI3K and PLC inhibitors, although the effect was very weak, probably because of the short treatment time (see Figure 2C). Thus, we conclude that the determination of the growth direction is also important for normal tip growth; this and the required elongation and division of stem cells in the branch initials are all regulated by both phosphoinositide turnover and actin polymerization.
Previously, we demonstrated the presence of auxin in conchocelis cells of P. yezoensis as part of a comprehensive analysis of plant hormones in Bangiales [46,47]. Furthermore, the functional involvement of auxin in tip growth in both conchocelis and conchosporangium is supported by chemical biology evidence [20], consistent with the involvement of auxin in tip growth of the brown alga Ectocarpus siliculosus [3], which also produces auxin [48]. In terrestrial plants, auxin signal transduction plays important roles in tip growth [10,49,50,51,52,53,54,55,56]. In addition, it is well known that the auxin receptor transport inhibitor response 1 (TIR1) and transcription factors like Auxin/Indole-3-Acetic Acid (Aux/IAA) and auxin response factor (ARF) are main components of the auxin signal transduction [49,57,58]. However, P. yezoensis has no orthologs of genes encoding TIR1, Aux/IAA, and ARF, key faactors in auxin signaling in terrestrial plants [46,47]. This suggests that red algae may have a novel pathway for auxin signal transduction. Thus, identification of factors involved in auxin sensing and signal transduction is one of the important issues for elucidating the regulatory mechanisms of tip growth in P. yezoensis.
In conclusion, our study demonstrated that the activities of PI3K and PLC and the polymerization of actin are critically involved in tip growth in conchocelis cells of P. yezoensis. In addition, we have already demonstrated the critical involvement of auxin in tip growth of these cells [20]. At present, however, the relationships among auxin, actin polymerization, and phosphoinositide turnover in regulation of tip growth are unknown. In the future, it will be valuable to determine whether acceleration of actin polymerization and activation of PI3K and PLC are auxin dependent. To this end, developing methods for F-actin visualization in living cells and reverse-genetic techniques for analyzing gene functions in P. yezoensis is essential. These efforts could provide novel insights into the mechanisms regulating the generation of branch initials, establishment of the stem cell, and tip growth of branches in conchocelis cells of P. yezoensis.
4. Materials and Methods
4.1. Algal Materials and Culture Conditions
The filamentous conchocelis of P. yezoensis (strain U-51) was maintained in sterilized artificial seawater according to Li et al. [59], with weekly changes of the seawater. Briefly, the culture conditions were 60 μmol photons m−2 s−1 light with a long-day photoperiod (14 h light/10 h dark) at 15°C, with aeration with sterilized air filtered through a 0.22-μm filter (Whatman, Maidstone, UK).
4.2. Preparation of Single-Celled Conchocelis
As described by Taya et al. [20], aggregates of filamentous conchocelis were chopped using a razor blade, and single conchocelis cells were collected by filtration of fragmented conchocelis through a 10-µm nylon mesh to remove large pieces. The filtrate was subsequently incubated in 9-cm dishes [Asunoru dish, 90 mm (diameter) × 20 mm (height), As One Corporation, Osaka, Japan] containing 30 ml seawater at 15°C for 10 min. Non-branched single conchocelis cells were identified using an Olympus IX73 light microscope equipped with an Olympus DP22 camera, picked up using a micropipette, and transferred to 96-well plates (each cell/well containing 200 µl artificial seawater).
4.3. Treatment of Single-Celled Conchocelis with Inhibitors of PI3K and PLC
Cells were incubated in 200 µl artificial seawater containing 5 or 10 µM LY294002 (Cayman Chemical, Ann Arbor, Michigan, USA) or LY303511 (Bio-Techne, Minneapolis, Minnesota, USA), which are active and inactive versions of PI3K-specific inhibitors, respectively, or containing 5 or 10 µM U73122 (Cayman Chemical, Ann Arbor, Michigan, USA) or U73433 (Cayman Chemical, Ann Arbor, Michigan, USA), which are active and inactive versions of PLC-specific inhibitors, respectively. Single-celled conchocelis was cultured statically in wells under the conditions described above except for aeration. Control treatments included 1% DMSO (FUJIFILM Wako Pure Chemical Corporation, Osaka, Japan) without the chemical inhibitors. The effects of the chemical inhibitors were evaluated after incubation for 3 days at 15°C by calculating branching rate (number of cells producing a branch as a percentage of the total number of cells observed) and measuring the length of the branches, both of which were performed by direct observation or by photographing the cells using an Olympus IX73 light microscope equipped with an Olympus DP22 camera.
4.4. Treatment of Single-Celled Conchocelis with Inhibitors of Actin Polymerization
Cells were incubated in 200 µl artificial seawater containing 10, 20, or 30 µM cytochalasin B (CCB; FUJIFILM Wako Pure Chemical Corporation, Osaka, Japan) or 5, 10, or 20 µM latrunculin B (LAT-B; Adipogen Life Sciences, San Diego, California, USA), under the same conditions used for the PI3K and PLC inhibitor experiments. Control treatments included 1% DMSO (FUJIFILM Wako Pure Chemical Corporation, Osaka, Japan) without the chemical inhibitors. After incubation for 7 days with CCB or LAT-B at 15°C, the effects of these actin polymerization inhibitors were evaluated as above.
4.5. Statistical Analysis
Mean values ± SD were calculated from triplicate experiments. Statistically significant differences in the interactions between the duration of incubation with the various chemicals and the regeneration rate of the branch initials or the length of branches were determined by one-way ANOVA with the Tukey-Kramer test (p < 0.05). Significant differences for each set of treatments were determined using a cutoff value of p < 0.05.
Supplementary Materials
The following supporting information can be downloaded at: Preprints.org, Figure S1: Effects of LY294002 on tip growth; Figure S2: Effects of U793122 on tip growth; Figure S3: Recovery from the effects of latrunculin B (LAT-B) on tip growth after removing the inhibitor.
Author Contributions
R.I. collected and analyzed the data and generated figures. K.M. conceived and designed the study, generated figures, and wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was not supported by any research fund.
Data Availability Statement
Data are contained within this article or Supplementary Materials.
Acknowledgments
We are grateful to the Marine Resources Research Center of Aichi Fisheries Research Institute for kindly providing P. yezoensis strain U51. We also thank Kensuke Taya, Lin Li, Ho Viet Khoa, Shoko Kitano, Haruka Makiyama, Makiha Fujita, Tatsunori Okitsu, and Yuta Yamazaki for their technical assistance.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Rounds, C.M.; Bezanilla, M. Growth mechanisms in tip-growing plant cells. Annu. Rev. Plant Biol. 2013, 64, 243–265. [Google Scholar] [CrossRef] [PubMed]
- Riquelme, M.; Aguirre, J.; Bartnicki-García, S.; Braus, G.H.; Feldbrügge, M.; Fleig, U.; Hansberg, W.; Herrera-Estrella, A.; Kämper, J.; Kück, U.; Mouriño-Pérez, R.R.; Takeshita, N.; Fischer, R. Fungal morphogenesis, from the polarized growth of hyphae to complex reproduction and infection structures. Microbiol. Mol. Biol. Rev. 2018, 82, e00068–17. [Google Scholar] [CrossRef] [PubMed]
- Rabillé, H.; Billoud, B.; Tesson, B.; Le Panse, S.; Rolland, É.; Charrier, B. The brown algal mode of tip growth: Keeping stress under control. PLoS Biol. 2019, 17, e2005258. [Google Scholar] [CrossRef]
- Geitmann, A.; Emons, A.M. The cytoskeleton in plant and fungal cell tip growth. J. Microsc. 2000, 198(Pt 3) Pt 3, 218–245. [Google Scholar] [CrossRef]
- Campanoni, P.; Blatt, M.R. Membrane trafficking and polar growth in root hairs and pollen tubes. J. Exp. Bot. 2007, 58, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Ischebeck, T.; Seiler, S.; Heilmann, I. At the poles across kingdoms: phosphoinositides and polar tip growth. Protoplasma 2010, 240, 13–31. [Google Scholar] [CrossRef]
- Pei, W.; Du, F.; Zhang, Y.; He, T.; Ren, H. Control of the actin cytoskeleton in root hair development. Plant Sci. 2012, 187, 10–18. [Google Scholar] [CrossRef]
- Guan, Y.; Guo, J.; Li, H.; Yang, Z. Signaling in pollen tube growth: crosstalk, feedback, and missing links. Mol. Plant 2013, 6, 1053–1064. [Google Scholar] [CrossRef]
- Malhó, R.; Serrazina, S.; Saavedra, L.; Dias, F.V.; Ul-Rehman, R. Ion and lipid signaling in apical growth-a dynamic machinery responding to extracellular cues. Front. Plant Sci. 2015, 6, 816. [Google Scholar] [CrossRef]
- Velasquez, S.M.; Barbez, E.; Kleine-Vehn, J.; Estevez, J.M. Auxin and cellular elongation. Plant Physiol. 2016, 170, 1206–1215. [Google Scholar] [CrossRef]
- Zhang, S.; Huang, L.; Yan, A.; Liu, Y.; Liu, B.; Yu, C.; Zhang, A.; Schiefelbein, J.; Gan, Y. Multiple phytohormones promote root hair elongation by regulating a similar set of genes in the root epidermis in Arabidopsis. J. Exp. Bot. 2016, 67, 6363–6372. [Google Scholar] [CrossRef] [PubMed]
- Bibeau, J.P.; Galotto, G.; Wu, M.; Tüzel, E.; Vidali, L. Quantitative cell biology of tip growth in moss. Plant Mol Biol. 2021, 107, 227–244. [Google Scholar] [CrossRef]
- Oubohssaine, M.; Hnini, M.; Rabeh, K. Phospholipid signaling in plant growth and development: Insights, biotechnological implications and future directions. J. Plant Physiol. 2025, 307, 154454. [Google Scholar] [CrossRef]
- Lee, Y.J.; Yang, Z. Tip growth: signaling in the apical dome. Curr. Opin. Plant Biol. 2008, 11, 662–671. [Google Scholar] [CrossRef] [PubMed]
- Scholz, P.; Anstatt, J.; Krawczyk, H.E.; Ischebeck, T. Signalling pinpointed to the tip: the complex regulatory network that allows pollen tube growth. Plants 2020, 9, 1098. [Google Scholar] [CrossRef] [PubMed]
- Blouin, N.A.; Brodie, J.A.; Grossman, A.C.; Xu, P.; Brawley, S.H. Porphyra: a marine crop shaped by stress. Trends Plant Sci. 2011, 16, 29–37. [Google Scholar] [CrossRef]
- Takahashi, M.; Mikami, K. Phototropism in the marine red macroalga Pyropia yezoensis. Am. J. Plant Sci. 2016, 7, 2412–2428. [Google Scholar] [CrossRef]
- Mikami, K.; Li, C.; Irie, R.; Hama, Y. A unique life cycle transition in the red seaweed Pyropia yezoensis depends on apospory. Commun. Biol. 2019, 2, 299. [Google Scholar] [CrossRef]
- Hiwatashi, Y.; Shimada, M.; Mikami, K.; Takada, N. Establishment of a live-imaging analysis for polarized growth of conchocelis in the multicellular red alga Neopyropia yezoensis. Front. Plant Sci. 2022, 12, 716011. [Google Scholar] [CrossRef]
- Taya, K.; Takeuchi, S.; Takahashi, M.; Hayashi, K.I.; Mikami, K. Auxin regulates apical stem cell regeneration and tip growth in the marine red alga Neopyropia yezoensis. Cells 2022, 11, 2652. [Google Scholar] [CrossRef]
- Kofuji, R.; Hasebe, M. Eight types of stem cells in the life cycle of the moss Physcomitrella patens. Curr. Opin. Plant Biol. 2014, 17, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Saga, N.; Mikami, K. Phosphatidylinositol 3-kinase activity and asymmetrical accumulation of F-actin are necessary for establishment of cell polarity in the early development of monospores from the marine red alga Porphyra yezoensis. J. Exp. Bot. 2008, 59, 3575–3586. [Google Scholar] [CrossRef]
- Li, L.; Saga, N.; Mikami, K. Effects of cell wall synthesis on cell polarity in the red alga Porphyra yezoensis. Plant Signal. Behav. 2008, 3, 1126–1128. [Google Scholar] [CrossRef]
- Li, L.; Saga, N.; Mikami, K. Ca2+ influx and phosphoinositide signalling are essential for the establishment and maintenance of cell polarity in monospores from the red alga Porphyra yezoensis. J. Exp. Bot. 2009, 60, 3477–3489. [Google Scholar] [CrossRef] [PubMed]
- Mikami, K.; Li, L.; Takahashi, M.; Saga, N. Photosynthesis-dependent Ca2+ influx and functional diversity between phoss pholipases in the formation of cell polarity in migrating cells of red algae. Plant Signal. Behav. 2009, 4, 911–913. [Google Scholar] [CrossRef]
- Heilmann, I. Plant phosphoinositide signaling - dynamics on demand. Biochim. Biophys. Acta 2016, 1861(Pt B), 1345–1351. [Google Scholar] [CrossRef]
- Fang, Y.; Jiang, J.; Ding, H.; Li, X.; Xie, X. Phospholipase C: Diverse functions in plant biotic stress resistance and fungal pathogenicity. Mol. Plant Pathol. 2023, 24, 1192–1202. [Google Scholar] [CrossRef] [PubMed]
- Helling, D.; Possart, A.; Cottier, S.; Klahre, U.; Kost, B. Pollen tube tip growth depends on plasma membrane polarization mediated by tobacco PLC3 activity and endocytic membrane recycling. Plant Cell 2006, 18, 3519–3534. [Google Scholar] [CrossRef]
- Lee, Y.; Bak, G.; Choi, Y.; Chuang, W.I.; Cho, H.T.; Lee, Y. Roles of phosphatidylinositol 3-kinase in root hair growth. Plant Physiol. 2008, 147, 624–635. [Google Scholar] [CrossRef]
- Kusano, H.; Testerink, C.; Vermeer, J.E.; Tsuge, T.; Shimada, H.; Oka, A.; Munnik, T.; Aoyama, T. The Arabidopsis phosphatidylinositol phosphate 5-kinase PIP5K3 is a key regulator of root hair tip growth. Plant Cell 2008, 20, 367–380. [Google Scholar] [CrossRef]
- Stenzel, I.; Ischebeck,T. ; König, S.; Hołubowska, A.; Sporysz, M.; Hause, B.; Heilmann, I. The type B phosphatidylinositol-4-phosphate 5-kinase 3 is essential for root hair formation in Arabidopsis thaliana. Plant Cell 2008, 20, 124–141. [Google Scholar] [CrossRef] [PubMed]
- Saavedra, L.; Balbi, V.; Lerche, J.; Mikami, K.; Heilmann, I.; Sommarin, M. PIPKs are essential for rhizoid elongation and caulonemal cell development in the moss Physcomitrella patens. Plant J. 2011, 67, 635–647. [Google Scholar] [CrossRef] [PubMed]
- Saavedra, L.; Mikami, K.; Malhó, R.; Sommarin, M. : PIP kinases and their role in plant tip growing cells. Plant Signal. Behav. 2012, 7, 1302–1305. [Google Scholar] [CrossRef] [PubMed]
- Kuo, H.F.; Chang, T.Y.; Chiang, S.F.; Wang, W.D.; Charng, Y.Y.; Chiou, T.J. Arabidopsis inositol pentakisphosphate 2-kinase, AtIPK1, is required for growth and modulates phosphate homeostasis at the transcriptional level. Plant J. 2014, 80, 503–515. [Google Scholar] [CrossRef]
- Wada, Y.; Kusano, H.; Tsuge, T.; Aoyama, T. Phosphatidylinositol phosphate 5-kinase genes respond to phosphate deficiency for root hair elongation in Arabidopsis thaliana. Plant J. 2015, 81, 426–437. [Google Scholar] [CrossRef]
- Zhang, Q.; van Wijk, R.; Zarza, X.; Shahbaz, M.; van Hooren, M.; Guardia, A.; Scuffi, D.; García-Mata, C.; Van den Ende, W.; Hoffmann-Benning, S.; Haring, M.A.; Laxalt, A.M.; Munnik, T. Knock-down of Arabidopsis PLC5 reduces primary root growth and secondary root formation whereas overexpression improves drought tolerance and causes stunted root hair growth. Plant Cell Physiol. 2018, 59, 2004–2019. [Google Scholar] [CrossRef]
- Kost, B.; Lemichez, E.; Spielhofer, P.; Hong, Y.; Tolias, K.; Carpenter, C.; Chua, N.H. Rac homologues and compartmentalized phosphatidylinositol 4,5-bisphosphate act in a common pathway to regulate polar pollen tube growth. J. Cell Biol. 1999, 145, 317–330. [Google Scholar] [CrossRef]
- Chen, C.Y.; Wong, E.I.; Vidali, L.; Estavillo, A.; Hepler, P.K.; Wu, H.M.; Cheung, A.Y. The regulation of actin organization by actin-depolymerizing factor in elongating pollen tubes. Plant Cell 2002, 14, 2175–2190. [Google Scholar] [CrossRef]
- Dowd, P.E.; Coursol, S.; Skirpan, A.L.; Kao, T.H.; Gilroy, S. Petunia phospholipase c1 is involved in pollen tube growth. Plant Cell 2006, 18, 1438–1453. [Google Scholar] [CrossRef]
- Cheung, A.Y.; Duan, Q.H.; Costa, S.S.; de Graaf, B.H.; Di Stilio, V.S.; Feijo, J.; Wu, H.M. The dynamic pollen tube cytoskeleton: live cell studies using actin-binding and microtubule-binding reporter proteins. Mol. Plant 2008, 1, 686–702. [Google Scholar] [CrossRef]
- Hempel, F.; Stenzel, I.; Heilmann, M.; Krishnamoorthy, P.; Menzel, W.; Golbik, R.; Helm, S.; Dobritzsch, D.; Baginsky, S.; Lee, J.; Hoehenwarter, W.; Heilmann, I. MAPKs influence pollen tube growth by controlling the formation of phosphatidylinositol 4,5-bisphosphate in an apical plasma membrane domain. Plant Cell 2017, 29, 3030–3050. [Google Scholar] [CrossRef]
- Mikami, K.; Hirata, R.; Takahashi, M.; Uji, T.; Saga, N. Transient transformation of red algal cells: Breakthrough toward genetic transformation of marine crop Porphyra species. In Genetic Transformation; Alvarez, M., ed.; InTech Open Access Publisher, Rijeka, Croatia, 2011, pp 241-258.
- Mikami, K. Current advances in seaweed transformation. In An Integrated View of the Molecular Recognition and Toxinology - From Analytical Procedures to Biomedical Applications; Baptista, G.R., ed., InTech Open Access Publisher, Rijeka, Croatia, 2013, pp 323-347.
- Mikami, K. A technical breakthrough close at hand: Feasible approaches toward establishing a gene-targeting genetic transformation system in seaweeds. Front. Plant Sci. 2014, 5, 498. [Google Scholar] [CrossRef] [PubMed]
- Mikami, K. Recent developments in nuclear reverse-genetic manipulations that advance seaweed biology in the genomic era. J. Aquat. Res. Mar. Sci. 2018, 2018, 39–42. [Google Scholar]
- Mikami, K.; Mori, I.C.; Matsuura, T.; Ikeda, Y.; Kojima, M.; Sakakibara, H.; Hirayama, T. Comprehensive quantification and genome survey reveal the presence of novel phytohormone action modes in red seaweeds. J. Appl. Phycol. 2016, 28, 2539–2548. [Google Scholar] [CrossRef]
- Mori, I.C.; Ikeda, Y.; Matsuura, T.; Hirayama, T.; Mikami, K. Phytohormones in red seaweeds: a technical review of methods for analysis and a consideration of genomic data. Bot. Mar. 2017, 60, 153–170. [Google Scholar] [CrossRef]
- Le Bail, A.; Billoud, B.; Kowalczyk, N.; Kowalczyk, M.; Gicquel, M.; Le Panse, S.; Stewart, S.; Scornet, D.; Cock, J.M.; Ljung, K.; Charrier, B. Auxin metabolism and function in the multicellular brown alga Ectocarpus siliculosus. Plant Physiol. 2010, 153, 128–144. [Google Scholar] [CrossRef]
- Leyser, O. Auxin signaling. Plant Physiol. 2018, 176, 465–479. [Google Scholar] [CrossRef]
- Shibata, M.; Sugimoto, K. A gene regulatory network for root hair development. J. Plant Res. 2019, 132, 301–309. [Google Scholar] [CrossRef]
- Li, M.; Zhu, Y.; Li, S.; Zhang, W.; Yin, C.; Lin, Y. Regulation of phytohormones on the growth and development of plant root hair. Front. Plant Sci. 2022, 13, 865302. [Google Scholar] [CrossRef]
- Li, E.; Zhang, Y.L. : Qin, Z.; Xu, M.; Qiao, Q.; Li, S.; Li, SW.; Zhang, Y. Signaling network controlling ROP-mediated tip growth in Arabidopsis and beyond. Plant Commun. 2023, 4, 100451. [Google Scholar] [CrossRef]
- Bittisnich, D.J.; Williamson, R.E. Tip-localised H+-fluxes and the applicability of the acid-growth hypothesis to tip-growing cells: Control of chloronemal extension in Funaria hygrometrica by auxin and light. Planta 1989, 178, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.Z.; Lin, Y.; Zhang, X.L.; Pang, D. W; Zha,o J. IAA stimulates pollen tube growth and mediates the modification of its wall composition and structure in Torenia fournieri. J. Exp. Bot. 2008, 59, 2529–2543. [Google Scholar] [CrossRef]
- Bhosale, R.; Giri, J.; Pandey, B.K.; Giehl, R.F.H.; Hartmann, A.; Traini, R.; Truskina, J.; Leftley, N.; Hanlon, M.; Swarup, K.; Rashed, A.; Voß, U.; Alonso, J.; Stepanova, A.; Yun, J.; Ljung, K.; Brown, K.M.; Lynch, J.P.; Dolan, L.; Vernoux, T.; Bishopp, A.; Wells, D.; von Wirén, N.; Bennett, M.J.; Swarup, R. A mechanistic framework for auxin dependent Arabidopsis root hair elongation to low external phosphate. Nat Commun. 2018, 9, 1409. [Google Scholar] [CrossRef] [PubMed]
- Schoenaers, S.; Balcerowicz, D.; Breen, G.; Hill, K.; Zdanio, M.; Mouille, G.; Holman, T.J.; Oh, J.; Wilson, M.H.; Nikonorova, N.; Vu, L.D.; De Smet, I.; Swarup, R.; De Vos, W.H.; Pintelon, I.; Adriaensen, D.; Grierson, C.; Bennett, M.J.; Vissenberg, K. The auxin-regulated CrRLK1L kinase ERULUS controls cell wall composition during root hair tip growth. Curr. Biol. 2018, 28, 722–732.e6. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, K. The interaction and integration of auxin signaling components. Plant Cell Physiol. 2012, 53, 965–975. [Google Scholar] [CrossRef]
- Fukui, K.; Hayashi, K.I. Manipulation and sensing of auxin metabolism, transport and signaling. Plant Cell Physiol. 2018, 59, 1500–1510. [Google Scholar] [CrossRef]
- Li, C.; Ariga, I.; Mikami, K. Difference in nitrogen starvation-inducible expression patterns among phylogenetically diverse ammonium transporter genes in the red seaweed Pyropia yezoensis. Am. J. Plant Sci. 2019, 10, 1325–1349. [Google Scholar] [CrossRef]
Figure 3.
Effects of the actin polymerization inhibitor cytochalasin B (CCB) on tip growth. (A, B) Branching rate (A) and branch length (B) following treatment of single conchocelis cells with 0.5% DMSO (control) or 10, 20, or 30 μM CCB for 7 days. Center line, median line; box limits, interquartile range with upper and lower quartiles; points, data; whiskers, range with maximum and minimum values; crosses, mean value. Lowercase letters denote significant differences in branching rate (A) and branch length (B) based on three independent experiments (n = 3) as determined by the Tukey-Kramer test (p < 0.05) for each set of treatments. (C, D) Photographs of single-celled conchocelis treated with 0.5% DMSO (C) or 20 μM CCB (D) for 7 days. Arrows indicate newly generated branches. Bars: 100 μm for (C) and 50 μm (D).
Figure 3.
Effects of the actin polymerization inhibitor cytochalasin B (CCB) on tip growth. (A, B) Branching rate (A) and branch length (B) following treatment of single conchocelis cells with 0.5% DMSO (control) or 10, 20, or 30 μM CCB for 7 days. Center line, median line; box limits, interquartile range with upper and lower quartiles; points, data; whiskers, range with maximum and minimum values; crosses, mean value. Lowercase letters denote significant differences in branching rate (A) and branch length (B) based on three independent experiments (n = 3) as determined by the Tukey-Kramer test (p < 0.05) for each set of treatments. (C, D) Photographs of single-celled conchocelis treated with 0.5% DMSO (C) or 20 μM CCB (D) for 7 days. Arrows indicate newly generated branches. Bars: 100 μm for (C) and 50 μm (D).
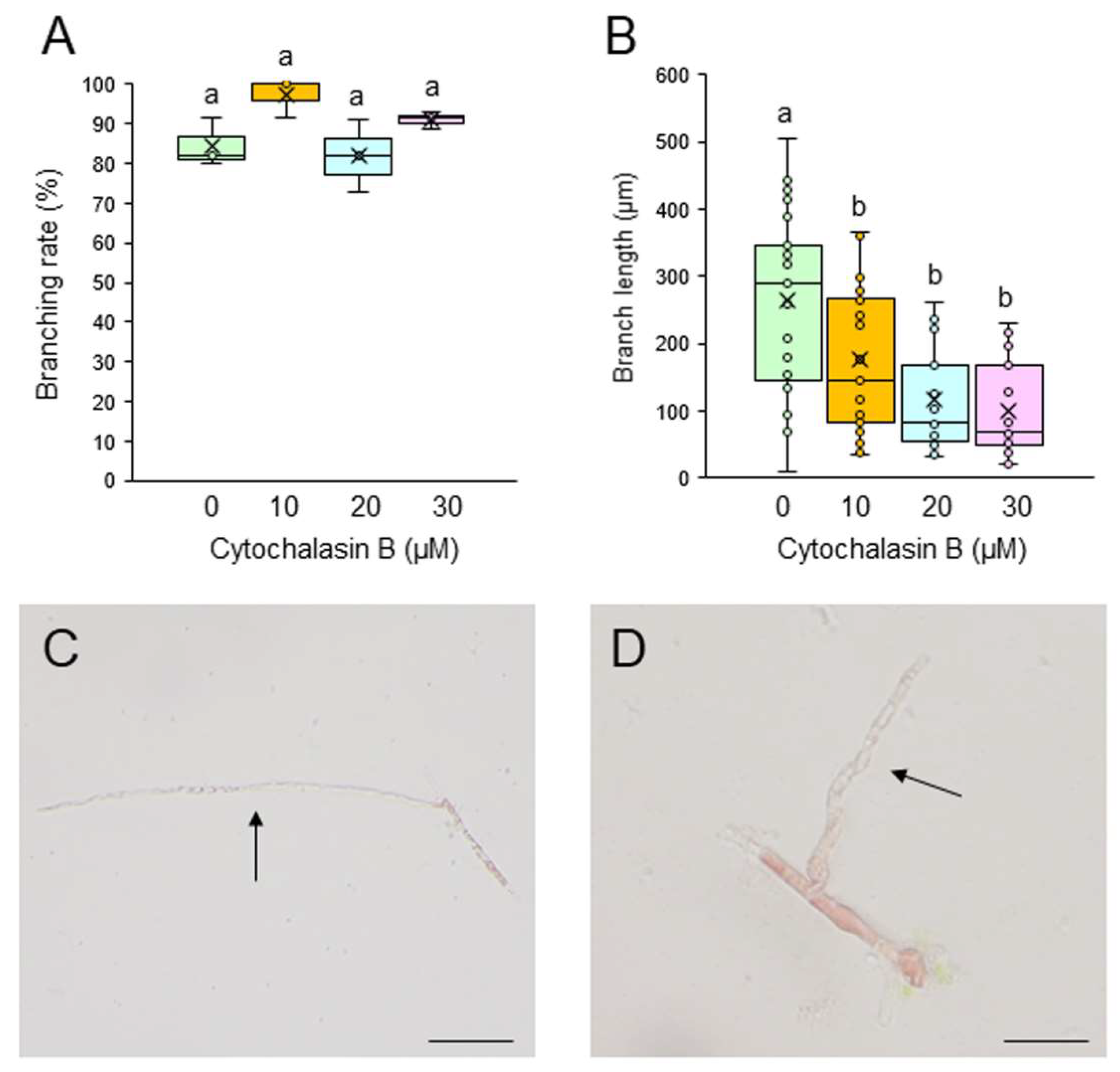
Figure 4.
Effects of the actin polymerization inhibitor latrunculin B (LAT-B) on tip growth. (A, B) Branching rate (A) and branch length (B) following treatment of single-celled conchocelis with 0.5% DMSO (control) or 5, 10, or 20 μM LAT-B for 7 days. Center line, median line; box limits, interquartile range with upper and lower quartiles; points, data; whiskers, range with maximum and minimum values; crosses, mean value. Lowercase letters denote significant differences in branching rate (A) and branch length (B) based on three independent experiments (n = 3) as determined by the Tukey-Kramer test (p < 0.05) for each set of treatments. (C, D) Photographs of single-celled conchocelis treated with 0.5% DMSO (C) or 20 μM LAT-B (D) for 7 days. Arrows indicate newly generated branches. Bars: 100 μm (C) and 50 μm (D).
Figure 4.
Effects of the actin polymerization inhibitor latrunculin B (LAT-B) on tip growth. (A, B) Branching rate (A) and branch length (B) following treatment of single-celled conchocelis with 0.5% DMSO (control) or 5, 10, or 20 μM LAT-B for 7 days. Center line, median line; box limits, interquartile range with upper and lower quartiles; points, data; whiskers, range with maximum and minimum values; crosses, mean value. Lowercase letters denote significant differences in branching rate (A) and branch length (B) based on three independent experiments (n = 3) as determined by the Tukey-Kramer test (p < 0.05) for each set of treatments. (C, D) Photographs of single-celled conchocelis treated with 0.5% DMSO (C) or 20 μM LAT-B (D) for 7 days. Arrows indicate newly generated branches. Bars: 100 μm (C) and 50 μm (D).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



