Submitted:
06 April 2025
Posted:
07 April 2025
You are already at the latest version
Abstract
The rise of multidrug-resistant (MDR) Pseudomonas aeruginosa remains an unresolved and substantial challenge to public health, which highlights an urgent need for newer therapeutic strategies. Despite the availability of innumerable antibiotics that effectively eliminate bacterial infections, their unregulated consumption and overexploitation has promoted the development of multidrug resistance by inducing selection pressure. As the world progresses into the post-antibiotic era, antivirulence therapies that exploit a ‘disarm-don’t kill approach’ are gaining momentum as a promising alternative to existing antimicrobial regimens. In view of extensive research being conducted to explore alternate intervention strategies against P. aeruginosa, this review augments scientific literature on repurposing of Food and Drug Administration (FDA)-approved drugs as antivirulence agents, focusing on their ability to disarm quorum sensing (QS), suppress virulence factor production, and disrupt biofilm formation. Drugs from various categories, including but not limited to antifungals, antidiabetics, antihypertensives, antiparasitics, NSAIDs, and antibiotics have been reported to override QS circuitry and QS-regulated virulence pathways in P. aeruginosa through in vitro and in vivo studies. Further, pre-clinical studies with FDA-approved drugs have been substantiated by in silico analysis predicting strong binding affinities to key QS receptors of P. aeruginosa such as LasR, RhlR, and PqsR, underscoring their potential mechanisms of action. Besides, with the well-documented safety profiles, pharmacokinetics, and clinical efficacy of the existing drugs, this repurposing approach streamlines the drug development process, minimizes costs, and accelerates the transition to clinical application. This review underscores the transformative potential of drug repurposing as a cost-effective and sustainable solution to the escalating antimicrobial resistance crisis and advocates for further research to optimize and clinically validate these promising antivirulence therapies.
Keywords:
Pseudomonas aeruginosa
; Drug repurposing
; Antivirulence therapy
; Multidrug resistance
; FDA-approved drugs
; Quorum quenching & Virulence attenuation
; Biofilm inhibition
1. Introduction
Pseudomonas aeruginosa, an aerobic-tolerant Gram-negative bacterium, has been recognized as the most notorious pathogen with a genome size ranging between 5.5-7 Mbp [1,2,3]. This versatile bacterium is ubiquitously distributed in natural habitats like soil, sewage, aqueous environments and in the mammalian gut [4]. In addition to being part of the normal human microflora, it is highly embroiled in infections contracted from both hospital and community settings. Being a prevalent microorganism in hospital-acquired infections, it accounts for 11-13.8% of all the characterized microbiological isolates among nosocomial infections [5]. In patients with cystic fibrosis, chronic obstructive pulmonary disease, or those undergoing immunosuppressive therapies, P. aeruginosa is responsible for inflicting severe respiratory infections, resulting in substantial reduction in the quality of life along with increased in mortality ranging between 18% to 61% [6]. Furthermore, the pathogen displays broad host range and wide tissue tropism, causing a myriad of grave infections such as gastrointestinal diseases, burn wound infections, keratitis, conjunctivitis, endophthalmitis, dacryocystitis, corneal ulcers, and osteomyelitis [7,8]. One of the most alarming yet fascinating aspects of P. aeruginosa is its magnificent adaptability and resilience against a wide range of antibiotics, which has led to its emergence as a promising nosocomial pathogen and inclusion within the ESAKPE pathogens [2,9]. The bacterium displays intrinsic resistance which can be attributed to mechanisms/factors such as low outer membrane permeability, presence of efflux pumps, and production of β-lactamases [10,11]. Additionally, it can also assimilate resistance genes via horizontal gene transfer, thereby exacerbating the complexity of therapeutic interventions [12,13]. With its ability to persist in hospital environments, pseudomonal isolates frequently exhibit a strong resistome, i.e., they harbor several mechanisms that contribute towards multidrug resistance and enhance bacterial survival. Moreover, the Centre for Disease Control and Prevention (CDC) has regarded MDR-P. aeruginosa as a notable menace over the past decade, reporting at least 32,600 cases with 2,700 deaths, incurring annual healthcare expenses of approximately $767 million globally [14,15]. Besides, MDR strains of P. aeruginosa have been implicated in a plethora of life-threatening infections, including malignancies, severe lower respiratory tract infections, cystic fibrosis, ventilator-associated pneumonia, and bacteraemia [13,16].
With MDR pathogens expanding at an undefined rate, the drug development pipeline is also dwindling. In this context, 15 out of 18 major global pharmaceutical corporations have abandoned antimicrobial drug discovery initiatives in the past decade [17]. The landscape of drug development is grappling with significant challenges arising from protracted timelines, escalating costs, and the intricate complexities associated with conducting clinical trials [18]. Furthermore, conventional drug development paradigms are characterized by a substantial failure rate of around 45%, which is largely attributable to concerns encompassing safety as well as toxicity [19,20]. Consequently, this switches the emphasis of a researcher from the challenges of drug development towards drug repurposing, also referred to as drug repositioning, by exploring and leveraging the safety and efficacy information of existing drugs. Since these drugs have already received approval from the Food and Drug Administration (FDA), they facilitate a more rapid response to the urgent medical needs by offering alternate therapeutic options for diseases having limited effective treatments [21,22,23]. As repurposed drugs bypass the initial 6–7 years typically needed for drug development and move directly into pre-clinical and clinical testing stages, the drug repurposing approach substantially saves manpower, precious time, and financial investment required to bring a drug to market compared to the de novo development of a novel pharmaceutical [24]. A comparison between the de novo drug development process and drug repurposing approach has been depicted in Figure 1. Various pre-approved drugs with anticancer, antifungal, antiparasitic, antihypertensive, antiviral, antipsychotic, and antidiabetic properties are aggressively repurposed for harnessing their newly-elucidated antimicrobial as well as antivirulence prospects [25]. Interestingly, approximately 30% of all drugs and biologics that are authorized by FDA are repositioned pharmaceuticals [20]. Recent statistics indicate that the drug repurposing market was valued at approximately USD 28.9 billion in 2023. Moreover, recent projections suggest that the pharmaceutics market (repurposing) will grow at a compound annual growth rate of 4.7% between 2024 to 2034, reaching around $47.8 billion by 2034 [26]. This growth has been predicted with the increasing prevalence of chronic and rare diseases, advancements in computational biology and artificial intelligence, and the need for cost-effective drug development strategies. North America currently dominates the market, accounting for 46.3% of the revenue in 2023, with a market size of $14.8 billion. As these trends continue, the drug repurposing market is expected to grow, offering great opportunities for pharmaceutical companies and healthcare providers to meet the global medical needs more effectively [27].
An alternative strategy to combat antimicrobial resistance (AMR) is through disarmament of bacterial virulence (antivirulence) by silencing signalling pathways, instead of posing a survival challenge and killing it (antibacterial) [28,29]. Antivirulence drugs (drugs showing attenuation of virulence), hence, reduce the risk of inducing selection pressure which is often witnessed when antimicrobial drugs are employed [30,31]. There are multiple ways by which the virulence of P. aeruginosa can be silenced, including iron chelation, lectin inhibitors, biofilm disruption, and quorum quenching (QQ) [32]. Among these methods, QQ is a unique approach for employing antivirulence therapy. P. aeruginosa regulates its virulence pathways through an intricate cell density-dependent mechanism called quorum sensing (QS), which further relies on chemical signal molecules called autoinducers (AIs), that help in establishing both intraspecies and interspecies communication between bacteria [33,34,35]. Coordinated bacterial responses prompted by the QS mechanisms induce the expression of diverse virulence factors that would otherwise be produced at a very small magnitude by individual cells. Therefore, drugs with QQ prospects can possibly be explored for antivirulence stratagems to combat hard-to-treat infections caused by MDR-P. aeruginosa [36]. Thus, the discovery of "antivirulence" potential in pre-approved drugs presents an exciting opportunity in pharmaceutical sciences.
2. Targeting the QS Pathways: A Promising Antivirulence Approach Against P. aeruginosa
The QS circuitry of P. aeruginosa is the most complicated yet extensively studied among all the QS systems known till date. The pathogen shows various integrated signalling pathways which work intricately in a hierarchal manner to augment bacterial virulence, thereby contributing to pathogenesis of P. aeruginosa. Approximately 10% of all pseudomonal genes are directly or indirectly regulated by the QS systems [37,38]. P. aeruginosa harbours 3 distinct and widely recognized QS pathways, comprising the Las-, Rhl-, and Pseudomonas quinolone signal (Pqs)-based system. The LasR/LasI and RhlR/RhlI QS systems regulate the signal transduction via N-3-oxo dodecanoyl-L-homoserine lactone (3-oxo-C12-HSL) and N-butyryl-L-homoserine lactone (C4-HSL) Ais, respectively [7]. The Las system comprises two key components, LasR and LasI, which regulate transcription and act as synthase proteins. LasI synthesizes 3-oxo-C12-HSL, which activates the cytoplasmic receptor, LasR, which further helps regulate the expression of genes associated with biofilm formation, hemolysins, proteases, elastases and exotoxin A production [39]. Similarly, RhlI, a synthase enzyme responsible for the production of C4-HSL, binds to its corresponding receptor RhlR and activates the production of virulence genes that produce pyocyanin, hydrogen cyanide, siderophores, elastases, alkaline protease, and regulates bacterial motility [40]. On the other hand, the Pqs system is chemically distinct and operates through a non-AHL-based QS molecule, 2-heptyl-3-hydroxy-4-quinolone (PQS), that is synthesized from the pqs operon (pqsABCDE) and is transported via outer membrane vesicles. It regulates the expression of genes involved in biofilm formation, swimming motility, and production of pyocyanin, proteases, elastases, rhamnolipids, pyochelin and pyoverdine (siderophores) that aid immune evasion and trigger lysis of neutrophils, macrophages, and other host cells [7]. The interlinked expression of these 3 QS pathways plays a quintessential role in driving pseudomonal virulence, fitness, and persistence [41].
To promote antivirulence response in P. aeruginosa, the QS pathways can be targeted by various approaches. The first strategy comprises enzyme-mediated degradation of QS signal (AHLs) by three known classes of enzymes namely: lactonases, acylases, and oxidoreductases [42]. Lactonases are metalloproteins that hydrolyse the ester bond within the homoserine lactone ring, resulting in the formation of the corresponding AHL molecule [43]. Acylases, on the other hand, irreversibly break down AHL molecules by hydrolysing the acyl-amide bond that links the acyl tail to the lactone ring [44]. This reaction releases a fatty acid chain and a homoserine lactone moiety as separate components [45]. In contrast, oxidoreductases modify AHLs it into an inactive form by oxidizing or reducing the acyl side chain [46]. They are the least abundant and least studied among the identified AHL-targeting enzymes. Another antivirulence strategy works by inhibiting the biosynthesis of QS molecules by directly obstructing AHL and PQS production in P. aeruginosa [47]. AHL production can be suppressed by targeting AHL synthases, interfering with substrate biosynthesis, or directly inhibiting synthase enzymes, as seen with fabI inhibition in P. aeruginosa. Similarly, inhibitors of PQS production, such as methyl anthranilate and farnesol, have been shown to modulate virulence in P. aeruginosa by lowering QS molecule biosynthesis [48,49]. The third strategy employs inhibition of QS signal transduction by exploiting analogues of native QS molecules or by designing small-molecule receptor inhibitors (called antagonists), which disrupt molecular interactions between the QS receptor(s) and their respective natural ligands (QS molecules) [47]. For example, PqsR antagonists, such as HHQ analogues with strong electron-withdrawing groups, competitively inhibit PQS signalling and attenuate bacterial virulence associated with the Pqs system [50].
As an emerging alternative to antimicrobial therapies, drugs with QQ prospects have been acclaimed for their potent antivirulence potential, disrupting QS mechanisms, and rendering the pathogen avirulent. Since QS is not involved primarily in survival of P. aeruginosa, but functions an auxiliary facet in modulating bacterial virulence, its abrogation does not directly extend any selection pressure over the pathogen. Interestingly, any drug that displays QQ potential attenuates the QS circuitry at a sub-inhibitory concentrations (sub-MICs) that do not pose a survival threat to the pathogen [47,51]. This antivirulence strategy, also designated as a ‘disarm-don’t kill approach’, therefore lowers the risk of developing anthropogenic resistance in P. aeruginosa, a phenomenon commonly observed with conventional antimicrobial drugs [52]. Hence, this review article is an attempt towards collating scientific literature focused on FDA-approved drugs that have been shown to effectively function as antivirulence drugs against P. aeruginosa.
3. Repositioning FDA-Approved Drugs Against P. aeruginosa: Evidences from Pre-Clinical Studies
Owing to widespread drug resistance in P. aeruginosa and the gradual downfall in drug development across pharmaceutical giants, there has been a leaning interest towards antivirulence therapies by repurposing FDA-approved drugs. By leveraging the established safety and efficacy profiles of existing drugs, this strategy allows for a faster and more cost-effective transition to new therapeutic applications [53]. These drugs have demonstrated efficacy in attenuating QS pathways, biofilm formation, and the production of host-damaging virulence factors, thereby mitigating bacterial pathogenicity in vitro and in vivo. With the rise of AMR in bacterial pathogens, antibiotics are increasingly being viewed as ineffective therapies, making drug repurposing for antimicrobial purposes less beneficial. Instead, drug repositioning for antivirulence strategies is gaining attention, as it targets pathogen virulence without directly affecting its survival, potentially reducing selective pressure. Various screening studies are now focusing on repurposing drugs for antivirulence applications. In this context, the upcoming section provides a comprehensive overview of repurposed antivirulence drugs specifically targeting P. aeruginosa, including different drug classes comprising antifungal, antidiabetic, antiparasitic, antibiotics, and nonsteroidal anti-inflammatory drugs, etc., detailing their mode of action and therapeutic efficacy through pre-clinical investigations. By emphasizing the antivirulence roles of drugs, this discussion underscores a transformative shift in the approach to treating MDR pathogens, hence, emphasizing the urgency of developing alternative solutions to address the growing AMR crisis.
3.1. Antifungal Drugs
5-Fluorocytosine (FC), a fluorinated cytosine analogue, was initially developed in 1957 for its potential as an anti-tumour agent but was later found to be ineffective against tumours. It was subsequently tested for its efficacy against candidiasis and cryptococcosis, where it demonstrated significant antifungal activity [54]. Since then, it has been extensively utilized for treating infections caused by Candida spp. and Cryptococcus neoformans in humans [55]. FC was the first ever antifungal drug to have been examined for its anti-QS activity against P. aeruginosa [56]. The study confirmed antivirulence potential of FC by inhibiting pyoverdine production by downregulating pvdS transcription. Moreover, administration of FC (30 mg/kg body weight) completely protected C57BL/6 mice infected intratracheally with P. aeruginosa (106 CFU). In a subsequent study, di Bonaventura et al. (2022) highlighted the antibiofilm effectiveness of ciclopirox against P. aeruginosa, which was originally used to treat superficial fungal and yeast infections, including seborrheic dermatitis, vaginal candidiasis and onychomycosis [58]. Besides the antifouling ability, ciclopirox was also found to downregulate the expression of the mexC but could not suppress the transcription of virulence genes such as aprA and toxA [57]. Recently an imidazole antifungal drug, miconazole, was scrutinized for its ability to suppress pseudomonal virulence [59]. Exposure to miconazole significantly reduced QS-regulated hemolysin, pyocyanin, and rhamnolipid production along with suppressing protease activity by 59%, 47-49%, 42-47%, and 36-40%, respectively, which was accompanied by biofilm inhibition up to ~ 45-48% [60]. Miconazole also downregulated the expression of various QS regulators: rhlR, pqsR, lasI, and lasR, which was further substantiated by molecular docking analysis confirming strong interactions between the antifungal agent and QS receptors. Furthermore, the effect of miconazole was evaluated in three-week-old healthy female albino mice that were injected intraperitoneally with P. aeruginosa PAO1. Consequently, miconazole-treated mice showed 100% survival. Henceforth, FC, ciclopirox and miconazole demonstrate significant promise as antifungal agents also harbouring antivirulence and antifouling potential against P. aeruginosa (Table 1), highlighting their ability to mitigate bacterial infection in animal models.
3.2. Antihypertensive Drugs
Till now, three antihypertensives have been elucidated for their antivirulence potential against P. aeruginosa, namely, glyceryl trinitrate (GTN), guanfacine, and propranolol. The first study in this regard surfaced when Abbas and Shaldam (2017) showed that GTN significantly reduced the synthesis of QS molecules and bacterial virulence in P. aeruginosa by impeding the production of pyocyanin (75%) and protease (78.5%), conjointly eradicating biofilm formation by 66.6% [61]. Interestingly, molecular docking showed profound interaction between GTN and ligand-binding pockets of LasR and RhlR with binding energies of -93.47 and -77.23, respectively. Similarly, guanfacine has also been shown to diminish biofilm development and pyocyanin production in P. aeruginosa [62]. More recently, sub-inhibitory concentrations of propranolol reportedly attenuated biofilm formation, interfering with motility phenotypes and inhibiting virulence factors (protease, hemolysin and pyocyanin production) in P. aeruginosa. [63]. Moreover, this drug was also proven to curtail the expression of QS-associated genes with in silico studies revealing highly competitive binding towards the LasR receptor. Furthermore, intraperitoneal administration of propranolol protected albino mice from P. aeruginosa-induced peritonitis, revealing its protective efficacy in vivo. Therefore, the aforementioned drugs (Table 2) can be exploited as potent antivirulence agents against P. aeruginosa, providing a strong foundation for their application as anti-infective drugs.
3.3. Antiparasitic Drugs
Niclosamide, a drug for treating tapeworm infections in humans, received approval from the US FDA in 1982 and is also listed among the World Health Organization's essential medicines [64]. In this context, a comprehensive study has highlighted the QQ potential of niclosamide, indicating its ability to quench AHL biosynthesis, thereby, inhibiting biofilm formation, swarming motility, and production of virulence factors like pyocyanin, elastase, protease and rhamnolipids in P. aeruginosa [65]. The authors also validated the antivirulence potential and protective effects of niclosamide against pseudomonal infection induced in Gardnerella mellonella (insect infection model). Another study by Yuan et al. (2022b) affirmed the dimetridazole-mediated attenuation of pseudomonal virulence in vitro as well as in vivo [66]. Results demonstrated that dimetridazole remarkably silenced the transcription of QS regulatory genes (lasR, rhlR, and pqsR), accompanied by the phenotypic impediment of QS-controlled proteases, pyocyanin, and biofilm formation. Moreover, the drug was shown to improve the survival of Caenorhabditis elegans and female C57BL/6 mice, effectively providing protection against pseudomonal infection in vivo. A recent in vitro study probed albendazole for its effect on bacterial virulence and biofilm formation in P. aeruginosa [67]. In the presence of albendazole (1/8 MIC), the production of hallmark virulence factors, including hemolysin, alginate, protease, rhamnolipids, elastase and pyocyanin were significantly inhibited by 52.6%, 64.1%, 67.3%, 54.5%, 50.5% and 60.8%, respectively. This reduction was driven by the suppressed expression of lasI, lasR, rhlI, rhlR, pqsA and pqsR, leading to a marked decrease in AHL production in albendazole-treated cultures. In silico analysis further revealed strong associations between albendazole and QS receptors of P. aeruginosa. Besides, albendazole was effective in impeding the swimming, swarming, and twitching motility phenotypes by 35.65%, 29.96%, and 34.26%, further delaying biofilm formation and eliciting an anti-fouling response. In a subsequent study, the authors highlighted the anti-QS potential and antivirulence prospects of ivermectin by showcasing its ability to significantly attenuate pyocyanin, hemolysin and pyochelin production [68]. Results also demonstrated ivermectin’s anti-proteolytic activity with a notable reduction of total bacterial proteases by 24.34%. Moreover, molecular docking predicted strong interactions between ivermectin and PqsR receptor of P. aeruginosa with a binding energy of -11.6 kcal/mol. Overall, the potency of various antiparasitics as QQ drugs against P. aeruginosa opens a new horizon towards exploring antivirulence therapies (Table 3).
3.3. Antidiabetic Drugs
Gliptins and metformin are among the most frequently prescribed FDA-approved antidiabetic medications that are extensively used in clinical practice. Besides the hypoglycaemic activity of metformin, it has also been shown to exhibit potent antivirulence activity by abrogating QS mechanisms in P. aeruginosa [69]. At sub-inhibitory concentrations, metformin significantly inhibited pyocyanin, hemolysin, protease, and elastase production, which coincided with reduced biofilm formation (68%), and retarded motility phenotypes [69]. Moreover, in silico analysis predicted molecular interactions between QS receptors (LasR/RhlR) and metformin, stipulating its potential antivirulence properties through these competitive interactions. In a subsequent study, the authors scrutinized the QQ prospects of sitagliptin against P. aeruginosa, illustrating its aptness in curtailing the bacterial virulence analogously to metformin [70]. Additionally, qRT-PCR studies indicated that expression of lasI, lasR, rhlI, rhlR, pqsA, and pqsR was downregulated by 41.19%, 38.79%, 41.35%, 32.44%, 55.15% and 75.21% respectively, in the presence of sitagliptin at sub-lethal doses [70]. Interestingly, in vivo experimentation by Hegazy et al. (2020) revealed that all albino mice infected with P. aeruginosa intraperitoneally and later treated with sitagliptin showed 100% survival [71]. Furthermore, metformin nanoemulsions have been developed that efficiently attenuate QS circuitry and biofilm formation (48.7-68.6%) in P. aeruginosa [72]. The nanoformulations significantly suppressed swarming motility (88.8-94.2 %) and showed a marked reduction in pyocyanin production, protease activity, and pqsA gene expression compared to metformin alone. Besides, metformin nanoemulsions protected mice from pseudomonal infection in vivo. A recent in vitro study conducted with metformin demonstrated notable QQ proficiency through reduced AHL biosynthesis in P. aeruginosa, subsequently attenuating bacterial virulence and biofilm formation in standard strains (PAO1/PA14) [73]. Moreover, a combination of vildagliptin and metformin was shown to hinder biofilm formation, pyocyanin production, motility phenotypes along with production of virulent extracellular enzymes in P. aeruginosa at significantly higher rates [74]. Interestingly, the drug combination at sub-lethal doses diminished P. aeruginosa-induced infection in mice by modulating bacterial virulence. Therefore, the scientific evidences presented in this section strongly validate the repositioning of antidiabetic drugs for their antivirulence properties (Table 4).
3.4. Nonsteroidal Anti-Inflammatory Drugs
Beyond the conventional role of analgesics in pain relief and modulation of inflammation, these are also being extensively examined for their antivirulence properties to treat pseudomonal infections. El-Mowafy et al. pioneered the first investigation on aspirin, revealing its potential QQ activity in vitro [75]. Aspirin reportedly attenuated bacterial virulence by inhibiting the production of elastase, total proteases, and pyocyanin, which was accompanied by reduced expression of lasI, lasR, rhlI, rhlR, pqsA and pqsR genes by 38%, 72%, 69%, 72%, 74%, and 43%, respectively. The downstream effects resulted in impediment of motility phenotypes and biofilm formation. These findings were also supported by in silico analysis that revealed strong binding affinity of aspirin towards LasR receptor. Subsequently, another study unveiled that tenoxicam at 1/4 MIC functions as a potent inhibitor of QS-regulated virulence in P. aeruginosa by attenuating pyoverdine, rhamnolipids, pyocyanin, elastase, proteases, and hemolysins production at phenotypic level [76]. The authors confirmed these findings by studying survival rates in P. aeruginosa-induced peritonitis mice model, wherein intraperitoneal administration of tenoxicam improved survival by ~80%. On similar lines, paracetamol at sub-inhibitory concentrations has been shown to mitigate antifouling response and migration-arrest (swarming) capability against P. aeruginosa by 64% and 57%, respectively [77]. Interestingly, diclofenac and its nanoformulation, diclofenac-loaded PLGA nanoparticles, have been investigated for their antivirulence prospects [78]. P. aeruginosa cultures treated with nanoparticle formulation showed significantly lowered hemolytic activity (32–88%), twitching motility, and biofilm formation, which coincided with reduced expression of lasI and lasR by 0.51- and 0.75-folds, respectively. Researchers have also highlighted the QQ potential of naproxen, as evidenced by the suppression of QS genes (lasI/rhlI) coupled with reduction in virulence factors such as bacterial protease, hemolysin, pyocyanin, biofilm, and motility phenotypes [79]. Additionally, molecular docking simulations uncovered naproxen's high affinity towards QS regulatory proteins. Most recently, aceclofenac has been scrutinized for its anti-QS potential and role in disarming pseudomonal virulence [68]. Results indicated that aceclofenac was capable of curtailing pyocyanin, protease, hemolysin and pyochelin production in P. aeruginosa PAO1. Besides, the findings were substantiated by in silico analysis which revealed strong molecular interactions between aceclofenac and QS receptors: LasR, RhlR and PqsR receptors with binding energies of -8.8, -8.5 and -7.7 kcal/mol, respectively. Since NSAIDs demonstrate strong antivirulence prospects, they can potentially serve as alternate drugs for targeting P. aeruginosa. However, the lack of in vivo studies becomes a critical research gap that needs to be addressed prior to their usage. A brief overview of repurposed NSAIDs demonstrating antivirulence properties have been enlisted in Table 5.
3.5. Antibiotics
While antibiotics have long been exploited for their broad-spectrum antibacterial potential, emerging research highlights their dual nature extending killing effects at higher concentrations while exhibiting antivirulence properties at sub-inhibitory concentrations [32]. Till date, several antibiotics have been used for this purpose, with erythromycin being the first one in this regard [80]. At sub-MICs, erythromycin effectively abolished the QS-regulated proteolytic and hemolytic nature of P. aeruginosa, thereby significantly reducing its virulence in mice. Later, Skindersoe et al. (2008) evaluated the effect of azithromycin, ceftazidime, and ciprofloxacin on QS in P. aeruginosa [81]. These drugs abrogated elastase, protease, and hemolysin production, extending their QQ ability. Another investigation affirmed the azithromycin-mediated attenuation of QS circuitry by restraining swimming, swarming and twitching motilities, biofilm establishment and QS signal production in vitro in P. aeruginosa PAO1 [82]. Doxycycline, a widely used tetracycline antibiotic, has been scrutinized by Husain and Ahmad (2013) for its antivirulence attributes targeting P. aeruginosa [83]. The outcomes demonstrated that sub-MICs of doxycycline abrogated elastase (67.2%), pyocyanin (69.1%), and protease (65%) production, impeding swarming motility (74%), and preventing biofilm formation in PAO1. Despite metronidazole’s primary role in targeting anaerobic bacteria, a study by Abbas (2015) has revealed its potential to influence virulence mechanisms in P. aeruginosa [84]. Production of pyocyanin, pyoverdine, protease and hemolysin was notably reduced in the presence of metronidazole supplemented at sub-MICs. Additionally, treatment with metronidazole resulted in a significantly compromised swimming and twitching motilities along with reduced biofilm formation. Further, Husain et al. (2016) assessed the use of ceftazidime as an antivirulence agent for treating pseudomonal infections, demonstrating its ability to reduce swarming motility with a sharp decrease in protease, elastase, and pyocyanin production [85]. Reduction in pyocyanin levels was affirmed by the suppression of PQS-activated transcription in P. aeruginosa. Moreover, PAO1-infected C. elegans displayed improved survival rate upon treatment with ceftazidime. Molecular docking also indicated that ceftazidime exhibits a strong affinity for LasR and RhlR ligand-binding domains. Other ꞵ-lactam antibiotics, cefepime & imipenem, have also been examined wherein the authors showcased the attenuating ability of the drugs against elastase, total proteases, hemolysin and pyocyanin production in P. aeruginosa [86]. Another antibiotic, clofoctol has been scrutinized for its role in abolishing pyocyanin production, swarming motility and biofilm formation in PAO1 strain [17]. Furthermore, real time PCR analysis showed downregulation of Pqs-controlled virulence genes. Secnidazole is another antibiotic with a mechanism of action similar to metronidazole, and its antivirulence efficacy has been characterized by the mitigation of key QS genes, including lasI, lasR, rhlI, rhlR, pqsA, and pqsR, which prevents P. aeruginosa-inflicted mortality in mice [87]. On similar lines, another study revealed the pathogen-disarming effect of nitrofurazone which significantly suppressed Pqs-dependent virulence traits in PAO1, including pyocyanin and rhamnolipids production, and swarming motility, and was also able to cease biofilm formation [88]. Ceftriaxone also exhibited potent QQ ability against PAO1 both in vitro and in vivo [89]. This antibiotic notably disrupted motility phenotypes, reduced pyocyanin production, extended antifouling properties, and enhanced the survival of C. elegans infected with PAO1. Additionally, in silico analysis predicted the high binding affinity for the ligand-binding pockets of QS receptors (LasR/PqsR). Recent findings by Naga et al. (2021) attributed the QS impeding activity of cefoperazone against P. aeruginosa, highlighting the subduction of QS-related virulence factors and downregulating the expression of lasI and rhlI genes, hence being a promising candidate for repurposing [90]. Nitrofurantoin, an antibiotic used for treating urinary tract infections, has recently been investigated for its ability to harbor anti-QS potential, thereby reducing pyocyanin, pyochelin, hemolysin, and total protease production [68]. Interestingly, nitrofurantoin also demonstrated strong molecular interaction with the QS receptors (LasR, RhlR, & PqsR) with binding energy -8.5, -7.6 and -6.7 kcal/mol, respectively. Collectively, these findings underscore the anti-QS and antivirulence potential of various antibiotics against P. aeruginosa PAO1 (Table 6). By combining standard in vitro and in silico methods along in vivo investigations, these antibiotics can be repurposed beyond their antimicrobial potential to curb the pseudomonal virulence.
In addition to the aforementioned drugs, several other chemical compounds demonstrating significant antivirulence properties against P. aeruginosa have also been reported in the scientific literature. A comprehensive list of such drugs along with their mechanisms of action and specific effects on virulence determinants, has been summarized in Table 7. Based on the existing literature, Figure 2 depicts the currently known molecular targets and bacterial pathways that are disrupted by various FDA-approved drugs for potentiating anti-QS and antivirulence responses against P. aeruginosa.
4. Future Perspectives
Despite the propitious outcomes of repurposed drugs as antivirulence agents, several challenges and opportunities remain to be addressed to ensure their successful clinical translation. There is a need for systematic screening pipelines to identify antivirulence activity in existing drugs by exploiting high-throughput screening technologies, coupled with computational methods like in silico modelling and machine learning. These approaches should focus on identifying compounds that have minimal impact on the host microbiota and pose a lower risk of fostering antimicrobial resistance. The translation of preclinical findings to clinical settings remains a critical hurdle. Rigorous validation through in vivo models, followed by well-designed clinical trials is essential to establish safety and efficacy. Collaboration between academic researchers, pharmaceutical companies, and regulatory bodies can facilitate a smoother transition. Additionally, optimizing drug delivery through innovative formulations such as nanoparticles, liposomes, or hydrogels can foster stability and bioavailability, ensuring targeted action towards infection sites. Combination therapies offer potential for amplifying therapeutic outcomes. Synergistic combination of antivirulence drugs with conventional antibiotics could simultaneously compromise bacterial defences while eradicating the infection and reducing the likelihood of resistance development. Furthermore, the concepts of antivirulence and drug revitalizing should also be applied to other MDR pathogens, expanding the potential of this approach to address a wide range of infectious diseases. Integrating artificial intelligence and machine learning could accelerate the discovery of new antivirulence candidates and help in predicting drug-target interactions and further streamline clinical trial designs. By addressing these challenges and harnessing technological advancements, antivirulence therapies have the potential to offer sustainable solutions to combat MDR pathogens proficiently.
5. Conclusions
P. aeruginosa poses an escalating threat globally which necessitates newer approaches to address its expanding resistome without further promoting antimicrobial resistance unlike conventional antibiotics. Antivirulence therapies present a groundbreaking alternative by targeting the mechanisms of pathogenicity without compromising bacterial survival. By prioritizing the "disarm-don’t kill" philosophy, these approaches offer a hopeful avenue to counteract this notorious pathogen while minimizing the risk of developing antimicrobial resistance. Drug repositioning has gained attention as a practical and efficient strategy for identifying antivirulence agents. As highlighted in this review, diverse classes of repurposed drugs, ranging from antifungals to antidiabetics, have shown significant potential to disrupt bacterial communication and pathogenicity, which have been validated by in vitro, in vivo, and in silico experimentation. This paradigm shift towards repurposed drugs underscores their potential as sustainable and effective toolkit for combating pseudomonal infections. In summary, antivirulence therapies and drug repurposing together hold the potential to reshape the landscape of MDR infection management, and emphasizes the promise of repurposed drugs as perpetual, cost-effective, and efficient alternatives for addressing P. aeruginosa infections. These therapies offer a fresh perspective to confront this versatile pathogen while reducing the likelihood of resistance development, paving the way for a new era in the management of infectious diseases.
Abbreviations
AcF: Aceclofenac, AHL: Acyl homoserine lactone, AHL’s: Acyl homoserine lactones, AI: Artificial intelligence, AMR: Antimicrobial resistance, AtS: Atorvastatin, AZM: Azithromycin, CDC: Centre for Disease Control and Prevention, CFT: Ceftazidime, COPD: Chronic obstructive pulmonary disease, CPR: Ciprofloxacin, CT: Ceftriaxone, FC: Flurocytosine, FDA: Food and Drug Administration, GTN: Glyceryl trinitrate, HHQ: 2-heptyl-4-(1H)-quinolone, HSL: Homoserine lactones, IvM: Ivermectin, LvC: Levocetrizine, MDR: Multidrug resistance, MIC: Minimum inhibitory concentration, NP: Nanoparticle, NSAIDs: Non-steroidal anti-inflammatory drugs, NT: Nitrofurantoin, PCR: Polymerase chain reaction, PLGA: Poly lactic-co-glycolic acid, PQS: Pseudomonas quinolone signal, QQ: Quorum quenching, QS: Quorum sensing, QSSM: Quorum sensing signalling molecules, UTI: Urinary tract infection.
Author Contributions
BS: Data curation, Formal Analysis, Methodology, Writing – original draft. JC: Conceptualization, Supervision, Data curation, Formal Analysis, Writing – review & editing. LK: Data curation, Formal Analysis, SR: Formal Analysis. KH: Supervision, Formal Analysis, Writing – review & editing.
Funding
The authors did not receive support from any organization for the submitted work.
Data availability
All the datasets generated and analyzed during the current study have all been cited in this
manuscript.
Acknowledgements
JC and LK would like to acknowledge the Indian Council of Medical Research (ICMR), Govt. of India, New Delhi, for providing Senior Research Fellowship (SRF).
Conflict of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Driscoll JA, Brody SL, Kollef MH. The Epidemiology, Pathogenesis and Treatment of Pseudomonas aeruginosa Infections. Drugs. 2007;67:351-68. [CrossRef]
- Pang Z, Raudonis R, Glick BR, Lin T-J, Cheng Z. Antibiotic resistance in Pseudomonas aeruginosa: mechanisms and alternative therapeutic strategies. Biotechnology Advances. 2019;37:177-92. [CrossRef]
- Stover CK, Pham XQ, Erwin AL, Mizoguchi SD, Warrener P, Hickey MJ, et al. Complete genome sequence of Pseudomonas aeruginosa PAO1, an opportunistic pathogen. Nature. 2000;406:959-64. [CrossRef]
- Sathe N, Beech P, Croft L, Suphioglu C, Kapat A, Athan E. Pseudomonas aeruginosa: Infections and novel approaches to treatment “Knowing the enemy” the threat of Pseudomonas aeruginosa and exploring novel approaches to treatment. Infectious Medicine. 2023;2:178-94. [CrossRef]
- Lizioli A, Privitera G, Alliata E, Antonietta Banfi EM, Boselli L, Panceri ML, et al. Prevalence of nosocomial infections in Italy: result from the Lombardy survey in 2000. Journal of Hospital Infection. 2003;54:141-8. [CrossRef]
- Kang CI, Kim SH, Kim HB, Park SW, Choe YJ, Oh Md, et al. Pseudomonas aeruginosaBacteremia: Risk Factors for Mortality and Influence of Delayed Receipt of Effective Antimicrobial Therapy on Clinical Outcome. Clinical Infectious Diseases. 2003;37:745-51. [CrossRef]
- Chadha J, Harjai K, Chhibber S. Revisiting the virulence hallmarks of Pseudomonas aeruginosa: a chronicle through the perspective of quorum sensing. Environmental Microbiology. 2022;24:2630-56. [CrossRef]
- El Zowalaty ME, Al Thani AA, Webster TJ, El Zowalaty AE, Schweizer HP, Nasrallah GK, et al. Pseudomonas Aeruginosa: Arsenal of Resistance Mechanisms, Decades of Changing Resistance Profiles, and Future Antimicrobial Therapies. Future Microbiology. 2015;10:1683-706. [CrossRef]
- Nguyen L, Garcia J, Gruenberg K, MacDougall C. Multidrug-Resistant Pseudomonas Infections: Hard to Treat, But Hope on the Horizon? Current Infectious Disease Reports. 2018;20. [CrossRef]
- C Reygaert W. An overview of the antimicrobial resistance mechanisms of bacteria. AIMS Microbiology. 2018;4:482-501. [CrossRef]
- Strateva T, Yordanov D. Pseudomonas aeruginosa – a phenomenon of bacterial resistance. Journal of Medical Microbiology. 2009;58:1133-48. [CrossRef]
- Patel JB, Richter SS. Mechanisms of Resistance to Antibacterial Agents. Manual of Clinical Microbiology2015. p. 1212-45. [CrossRef]
- Aggarwal M, Patra A, Awasthi I, George A, Gagneja S, Gupta V, et al. Drug repurposing against antibiotic resistant bacterial pathogens. European Journal of Medicinal Chemistry. 2024;279. [CrossRef]
- Kunz Coyne AJ, El Ghali A, Holger D, Rebold N, Rybak MJ. Therapeutic Strategies for Emerging Multidrug-Resistant Pseudomonas aeruginosa. Infectious Diseases and Therapy. 2022;11:661-82. [CrossRef]
- Burrows LL. The Therapeutic Pipeline for Pseudomonas aeruginosa Infections. ACS Infectious Diseases. 2018;4:1041-7. [CrossRef]
- Botelho J, Grosso F, Peixe L. Antibiotic resistance in Pseudomonas aeruginosa – Mechanisms, epidemiology and evolution. Drug Resistance Updates. 2019;44. [CrossRef]
- D'Angelo F, Baldelli V, Halliday N, Pantalone P, Polticelli F, Fiscarelli E, et al. Identification of FDA-Approved Drugs as Antivirulence Agents Targeting the pqs Quorum-Sensing System of Pseudomonas aeruginosa. Antimicrobial Agents and Chemotherapy. 2018;62. [CrossRef]
- Chong CR, Sullivan DJ. New uses for old drugs. Nature. 2007;448:645-6. [CrossRef]
- Xue H, Li J, Xie H, Wang Y. Review of Drug Repositioning Approaches and Resources. International Journal of Biological Sciences. 2018;14:1232-44. [CrossRef]
- Rudrapal M, J. Khairnar S, G. Jadhav A. Drug Repurposing (DR): An Emerging Approach in Drug Discovery. Drug Repurposing - Hypothesis, Molecular Aspects and Therapeutic Applications. 2020.
- Delavan B, Roberts R, Huang R, Bao W, Tong W, Liu Z. Computational drug repositioning for rare diseases in the era of precision medicine. Drug Discovery Today. 2018;23:382-94. [CrossRef]
- Parvathaneni V, Kulkarni NS, Muth A, Gupta V. Drug repurposing: a promising tool to accelerate the drug discovery process. Drug Discovery Today. 2019;24:2076-85. [CrossRef]
- Law GL, Tisoncik-Go J, Korth MJ, Katze MG. Drug repurposing: a better approach for infectious disease drug discovery? Current Opinion in Immunology. 2013;25:588-92. [CrossRef]
- Padhy BM, Gupta YK. Drug repositioning: Re-investigating existing drugs for new therapeutic indications. Journal of Postgraduate Medicine. 2011;57. [CrossRef]
- Walker D, Rampioni G, Visca P, Leoni L, Imperi F. Drug repurposing for antivirulence therapy against opportunistic bacterial pathogens. Emerging Topics in Life Sciences. 2017;1:13-22. [CrossRef]
- Newswire P. Drug Repurposing Market Projected to Reach USD 47.8 Billion by 2034 with a CAGR of 4.7% - Transparency Market Research. https://www.prnewswire.com/news-releases/drug-repurposing-market-projected-to-reach-usd-47-8-billion-by-2034-with-a-cagr-of-4-7---transparency-market-research-302199223.html, 2024 (accessed 5 April 2025).
- Presswire E. Drug Repurposing Market Forecasted To Hit US$ 51.8 Billion By 2033 Biovista, Excelra, Fios Genomics, Novartis AG. https://www.einpresswire.com/article/780644699/drug-repurposing-market-forecasted-to-hit-us-51-8-billion-by-2033-biovista-excelra-fios-genomics-novartis-ag, 2025 (accessed 5 April 2025).
- Lee Y. Targeting virulence for antimicrobial chemotherapy. Current Opinion in Pharmacology. 2003;3:513-9. [CrossRef]
- Marra A. Can virulence factors be viable antibacterial targets? Expert Review of Anti-infective Therapy. 2014;2:61-72. [CrossRef]
- Rasko DA, Sperandio V. Anti-virulence strategies to combat bacteria-mediated disease. Nature Reviews Drug Discovery. 2010;9:117-28. [CrossRef]
- Cegelski L, Marshall GR, Eldridge GR, Hultgren SJ. The biology and future prospects of antivirulence therapies. Nature Reviews Microbiology. 2008;6:17-27. [CrossRef]
- Mudgil U, Khullar L, Chadha J, Prerna, Harjai K. Beyond antibiotics: Emerging antivirulence strategies to combat Pseudomonas aeruginosa in cystic fibrosis. Microbial Pathogenesis. 2024;193. [CrossRef]
- Pena RT, Blasco L, Ambroa A, González-Pedrajo B, Fernández-García L, López M, et al. Relationship Between Quorum Sensing and Secretion Systems. Frontiers in Microbiology. 2019;10. [CrossRef]
- Nealson KH, Platt T, Hastings JW. Cellular Control of the Synthesis and Activity of the Bacterial Luminescent System. Journal of Bacteriology. 1970;104:313-22. [CrossRef]
- Hawver LA, Jung SA, Ng W-L, Shen A. Specificity and complexity in bacterial quorum-sensing systems. FEMS Microbiology Reviews. 2016;40:738-52. [CrossRef]
- Pérez-Pérez M, Jorge P, Pérez Rodríguez G, Pereira MO, Lourenço A. Quorum sensing inhibition in Pseudomonas aeruginosa biofilms: new insights through network mining. Biofouling. 2017;33:128-42. [CrossRef]
- Williams P, Cámara M. Quorum sensing and environmental adaptation in Pseudomonas aeruginosa: a tale of regulatory networks and multifunctional signal molecules. Current Opinion in Microbiology. 2009;12:182-91. [CrossRef]
- Schuster M, Peter Greenberg E. A network of networks: Quorum-sensing gene regulation in Pseudomonas aeruginosa. International Journal of Medical Microbiology. 2006;296:73-81. [CrossRef]
- Rutherford ST, Bassler BL. Bacterial Quorum Sensing: Its Role in Virulence and Possibilities for Its Control. Cold Spring Harbor Perspectives in Medicine. 2012;2:a012427-a. [CrossRef]
- Papenfort K, Bassler BL. Quorum sensing signal–response systems in Gram-negative bacteria. Nature Reviews Microbiology. 2016;14:576-88. [CrossRef]
- Lee J, Zhang L. The hierarchy quorum sensing network in Pseudomonas aeruginosa. Protein & Cell. 2014;6:26-41. [CrossRef]
- Dong Y-H, Xu J-L, Li X-Z, Zhang L-H. AiiA, an enzyme that inactivates the acylhomoserine lactone quorum-sensing signal and attenuates the virulence of Erwinia carotovora. Proceedings of the National Academy of Sciences. 2000;97:3526-31. [CrossRef]
- Dong Y-H, Wang L-H, Xu J-L, Zhang H-B, Zhang X-F, Zhang L-H. Quenching quorum-sensing-dependent bacterial infection by an N-acyl homoserine lactonase. Nature. 2001;411:813-7. [CrossRef]
- Leadbetter JR, Greenberg EP. Metabolism of Acyl-Homoserine Lactone Quorum-Sensing Signals by Variovorax paradoxus. Journal of Bacteriology. 2000;182:6921-6. [CrossRef]
- Lin YH, Xu JL, Hu J, Wang LH, Ong SL, Leadbetter JR, et al. Acyl-homoserine lactone acylase from Ralstonia strain XJ12B represents a novel and potent class of quorum-quenching enzymes. Molecular Microbiology. 2003;47:849-60. [CrossRef]
- Uroz S, Chhabra SR, Cámara M, Williams P, Oger P, Dessaux Y. N-Acylhomoserine lactone quorum-sensing molecules are modified and degraded by Rhodococcus erythropolis W2 by both amidolytic and novel oxidoreductase activities. Microbiology. 2005;151:3313-22. [CrossRef]
- Chadha J, Harjai K, Chhibber S. Repurposing phytochemicals as anti-virulent agents to attenuate quorum sensing-regulated virulence factors and biofilm formation in Pseudomonas aeruginosa. Microbial Biotechnology. 2022;15:1695-718. [CrossRef]
- Calfee MW, Coleman JP, Pesci EC. Interference with Pseudomonas quinolone signal synthesis inhibits virulence factor expression by Pseudomonas aeruginosa. Proceedings of the National Academy of Sciences. 2001;98:11633-7. [CrossRef]
- Cugini C, Calfee MW, Farrow JM, Morales DK, Pesci EC, Hogan DA. Farnesol, a common sesquiterpene, inhibits PQS production in Pseudomonas aeruginosa. Molecular Microbiology. 2007;65:896-906. [CrossRef]
- Lu C, Kirsch B, Zimmer C, de Jong Johannes C, Henn C, Maurer Christine K, et al. Discovery of Antagonists of PqsR, a Key Player in 2-Alkyl-4-quinolone-Dependent Quorum Sensing in Pseudomonas aeruginosa. Chemistry & Biology. 2012;19:381-90. [CrossRef]
- Kalia VC, Patel SKS, Kang YC, Lee J-K. Quorum sensing inhibitors as antipathogens: biotechnological applications. Biotechnology Advances. 2019;37:68-90. [CrossRef]
- Rasmussen TB, Givskov M. Quorum sensing inhibitors: a bargain of effects. Microbiology. 2006;152:895-904. [CrossRef]
- Kulkarni VS, Alagarsamy V, Solomon VR, Jose PA, Murugesan S. Drug Repurposing: An Effective Tool in Modern Drug Discovery. Russian Journal of Bioorganic Chemistry. 2023;49:157-66. [CrossRef]
- Titsworth E, Grunberg E. Chemotherapeutic Activity of 5-Fluorocytosine and Amphotericin B Against Candida albicans in Mice. Antimicrobial Agents and Chemotherapy. 1973;4:306-8. [CrossRef]
- Vermes A. Flucytosine: a review of its pharmacology, clinical indications, pharmacokinetics, toxicity and drug interactions. Journal of Antimicrobial Chemotherapy. 2000;46:171-9. [CrossRef]
- Imperi F, Massai F, Facchini M, Frangipani E, Visaggio D, Leoni L, et al. Repurposing the antimycotic drug flucytosine for suppression of Pseudomonas aeruginosa pathogenicity. Proceedings of the National Academy of Sciences. 2013;110:7458-63. [CrossRef]
- Di Bonaventura G, Lupetti V, De Fabritiis S, Piccirilli A, Porreca A, Di Nicola M, et al. Giving Drugs a Second Chance: Antibacterial and Antibiofilm Effects of Ciclopirox and Ribavirin against Cystic Fibrosis Pseudomonas aeruginosa Strains. International Journal of Molecular Sciences. 2022;23. [CrossRef]
- Gupta AK, Plott T. Ciclopirox: a broad-spectrum antifungal with antibacterial and anti-inflammatory properties. International Journal of Dermatology. 2004;43:3-8. [CrossRef]
- Heel RC, Brogden RN, Pakes GE, Speight TM, Avery GS. Miconazole. Drugs. 1980;19:7-30. [CrossRef]
- Gad AI, El-Ganiny AM, Eissa AG, Noureldin NA, Nazeih SI. Miconazole and phenothiazine hinder the quorum sensing regulated virulence in Pseudomonas aeruginosa. The Journal of Antibiotics. 2024;77:454-65. [CrossRef]
- Abbas HA, Shaldam MA. Glyceryl trinitrate is a novel inhibitor of quorum sensing in Pseudomonas aeruginosa. African Health Sciences. 2017;16. [CrossRef]
- Okada BK, Li A, Seyedsayamdost MR. Identification of the Hypertension Drug Guanfacine as an Antivirulence Agent in Pseudomonas aeruginosa. ChemBioChem. 2019;20:2005-11. [CrossRef]
- Alotaibi HF, Alotaibi H, Darwish KM, Khafagy E-S, Abu Lila AS, Ali MAM, et al. The Anti-Virulence Activities of the Antihypertensive Drug Propranolol in Light of Its Anti-Quorum Sensing Effects against Pseudomonas aeruginosa and Serratia marcescens. Biomedicines. 2023;11. [CrossRef]
- Chen W, Mook RA, Premont RT, Wang J. Niclosamide: Beyond an antihelminthic drug. Cellular Signalling. 2018;41:89-96. [CrossRef]
- Imperi F, Massai F, Ramachandran Pillai C, Longo F, Zennaro E, Rampioni G, et al. New Life for an Old Drug: the Anthelmintic Drug Niclosamide Inhibits Pseudomonas aeruginosa Quorum Sensing. Antimicrobial Agents and Chemotherapy. 2013;57:996-1005. [CrossRef]
- Yuan Y, Yang X, Zeng Q, Li H, Fu R, Du L, et al. Repurposing Dimetridazole and Ribavirin to disarm Pseudomonas aeruginosa virulence by targeting the quorum sensing system. Frontiers in Microbiology. 2022;13. [CrossRef]
- Chadha J, Khullar L, Gulati P, Chhibber S, Harjai K. Repurposing albendazole as a potent inhibitor of quorum sensing-regulated virulence factors in Pseudomonas aeruginosa: Novel prospects of a classical drug. Microbial Pathogenesis. 2024;186. [CrossRef]
- Chadha J, Mudgil U, Khullar L, Ahuja P, Harjai K. Revitalizing common drugs for antibacterial, quorum quenching, and antivirulence potential against Pseudomonas aeruginosa: in vitro and in silico insights. 3 Biotech. 2024;14. [CrossRef]
- Abbas HA, Elsherbini AM, Shaldam MA. Repurposing metformin as a quorum sensing inhibitor in Pseudomonas aeruginosa. African Health Sciences. 2017;17. [CrossRef]
- Abbas HA, Shaldam MA, Eldamasi D. Curtailing Quorum Sensing in Pseudomonas aeruginosa by Sitagliptin. Current Microbiology. 2020;77:1051-60. [CrossRef]
- Hegazy WAH, Khayat MT, Ibrahim TS, Nassar MS, Bakhrebah MA, Abdulaal WH, et al. Repurposing Anti-diabetic Drugs to Cripple Quorum Sensing in Pseudomonas aeruginosa. Microorganisms. 2020;8. [CrossRef]
- Gomaa SE, Shaker GH, Mosallam FM, Abbas HA. Knocking down Pseudomonas aeruginosa virulence by oral hypoglycemic metformin nano emulsion. World Journal of Microbiology and Biotechnology. 2022;38. [CrossRef]
- Chadha J, Khullar L, Gulati P, Chhibber S, Harjai K. Anti-virulence prospects of Metformin against Pseudomonas aeruginosa: A new dimension to a multifaceted drug. Microbial Pathogenesis. 2023;183. [CrossRef]
- Khayat MT, Abbas HA, Ibrahim TS, Elbaramawi SS, Khayyat AN, Alharbi M, et al. Synergistic Benefits: Exploring the Anti-Virulence Effects of Metformin/Vildagliptin Antidiabetic Combination against Pseudomonas aeruginosa via Controlling Quorum Sensing Systems. Biomedicines. 2023;11. [CrossRef]
- El-Mowafy SA, Abd El Galil KH, El-Messery SM, Shaaban MI. Aspirin is an efficient inhibitor of quorum sensing, virulence and toxins in Pseudomonas aeruginosa. Microbial Pathogenesis. 2014;74:25-32. [CrossRef]
- Askoura M, Saleh M, Abbas H. An innovative role for tenoxicam as a quorum sensing inhibitor in Pseudomonas aeruginosa. Archives of Microbiology. 2019;202:555-65. [CrossRef]
- Seleem NM, Atallah H, Abd El Latif HK, Shaldam MA, El-Ganiny AM. Could the analgesic drugs, paracetamol and indomethacin, function as quorum sensing inhibitors? Microbial Pathogenesis. 2021;158. [CrossRef]
- Rostamnejad D, Esnaashari F, Zahmatkesh H, Rasti B, Zamani H. Diclofenac-loaded PLGA nanoparticles downregulate LasI/R quorum sensing genes in pathogenic P. aeruginosa isolates. Archives of Microbiology. 2024;206. [CrossRef]
- Esnaashari F, Rostamnejad D, Zahmatkesh H, Zamani H. In vitro and in silico assessment of anti-quorum sensing activity of Naproxen against Pseudomonas aeruginosa. World Journal of Microbiology and Biotechnology. 2023;39. [CrossRef]
- Sofer D, Gilboa-Garber N, Belz A, Garber NC. ‘Subinhibitory’ Erythromycin Represses Production of Pseudomonas aeruginosa Lectins, Autoinducer and Virulence Factors. Chemotherapy. 1999;45:335-41. [CrossRef]
- Skindersoe ME, Alhede M, Phipps R, Yang L, Jensen PO, Rasmussen TB, et al. Effects of Antibiotics on Quorum Sensing in Pseudomonas aeruginosa. Antimicrobial Agents and Chemotherapy. 2008;52:3648-63. [CrossRef]
- Bala A, Kumar R, Harjai K. Inhibition of quorum sensing in Pseudomonas aeruginosa by azithromycin and its effectiveness in urinary tract infections. Journal of Medical Microbiology. 2011;60:300-6. [CrossRef]
- Husain FM, Ahmad I. Doxycycline interferes with quorum sensing-mediated virulence factors and biofilm formation in Gram-negative bacteria. World Journal of Microbiology and Biotechnology. 2013;29:949-57. [CrossRef]
- Abbas HA. Inhibition of Virulence ofPseudomonas aeruginosa: A Novel Role of Metronidazole Against Aerobic Bacteria. Research Journal of Pharmacy and Technology. 2015;8. [CrossRef]
- Husain FM, Ahmad I, Baig MH, Khan MS, Khan MS, Hassan I, et al. Broad-spectrum inhibition of AHL-regulated virulence factors and biofilms by sub-inhibitory concentrations of ceftazidime. RSC Advances. 2016;6:27952-62. [CrossRef]
- El-Mowafy SA, Abd El Galil KH, Habib E-SE, Shaaban MI. Quorum sensing inhibitory activity of sub-inhibitory concentrations of β-lactams. African Health Sciences. 2017;17. [CrossRef]
- Saleh MM, Abbas HA, Askoura MM. Repositioning secnidazole as a novel virulence factors attenuating agent in Pseudomonas aeruginosa. Microbial Pathogenesis. 2019;127:31-8. [CrossRef]
- Baldelli V, D’Angelo F, Pavoncello V, Fiscarelli EV, Visca P, Rampioni G, et al. Identification of FDA-approved antivirulence drugs targeting the Pseudomonas aeruginosa quorum sensing effector protein PqsE. Virulence. 2020;11:652-68. [CrossRef]
- Kumar L, Brenner N, Brice J, Klein-Seetharaman J, Sarkar SK. Cephalosporins Interfere With Quorum Sensing and Improve the Ability of Caenorhabditis elegans to Survive Pseudomonas aeruginosa Infection. Frontiers in Microbiology. 2021;12. [CrossRef]
- Naga NG, El-Badan DE, Rateb HS, Ghanem KM, Shaaban MI. Quorum Sensing Inhibiting Activity of Cefoperazone and Its Metallic Derivatives on Pseudomonas aeruginosa. Frontiers in Cellular and Infection Microbiology. 2021;11. [CrossRef]
- Bandara MBK, Zhu H, Sankaridurg PR, Willcox MDP. Salicylic Acid Reduces the Production of Several Potential Virulence Factors of Pseudomonas aeruginosa Associated with Microbial Keratitis. Investigative Opthalmology & Visual Science. 2006;47. [CrossRef]
- Yuan M, Chua SL, Liu Y, Drautz-Moses DI, Yam JKH, Aung TT, et al. Repurposing the anticancer drug cisplatin with the aim of developing novel Pseudomonas aeruginosa infection control agents. Beilstein Journal of Organic Chemistry. 2018;14:3059-69. [CrossRef]
- Gerner E, Almqvist S, Werthén M, Trobos M. Sodium salicylate interferes with quorum-sensing-regulated virulence in chronic wound isolates of Pseudomonas aeruginosa in simulated wound fluid. Journal of Medical Microbiology. 2020;69:767-80. [CrossRef]
- Saqr AA, Aldawsari MF, Khafagy E-S, Shaldam MA, Hegazy WAH, Abbas HA. A Novel Use of Allopurinol as A Quorum-Sensing Inhibitor in Pseudomonas aeruginosa. Antibiotics. 2021;10. [CrossRef]
- Soltani S, Fazly Bazzaz BS, Hadizadeh F, Roodbari F, Soheili V. New Insight into Vitamins E and K1 as Anti-Quorum-Sensing Agents against Pseudomonas aeruginosa. Antimicrobial Agents and Chemotherapy. 2021;65. [CrossRef]
- Al-Rabia MW, Asfour HZ, Alhakamy NA, Bazuhair MA, Ibrahim TS, Abbas HA, et al. Cilostazol is a promising anti-pseudomonal virulence drug by disruption of quorum sensing. AMB Express. 2024;14. [CrossRef]
Figure 1.
Comparison of de novo Drug Discovery and Drug Repurposing Approaches. De novo drug discovery follows a lengthy process from target identification to clinical trials and regulatory approval. In contrast, drug repurposing accelerates development by identifying existing drugs and bypassing Phase I trials, thereby expediting regulatory review and market repositioning. This highlights the efficiency of drug repurposing in accelerating drug availability for clinical use.
Figure 1.
Comparison of de novo Drug Discovery and Drug Repurposing Approaches. De novo drug discovery follows a lengthy process from target identification to clinical trials and regulatory approval. In contrast, drug repurposing accelerates development by identifying existing drugs and bypassing Phase I trials, thereby expediting regulatory review and market repositioning. This highlights the efficiency of drug repurposing in accelerating drug availability for clinical use.
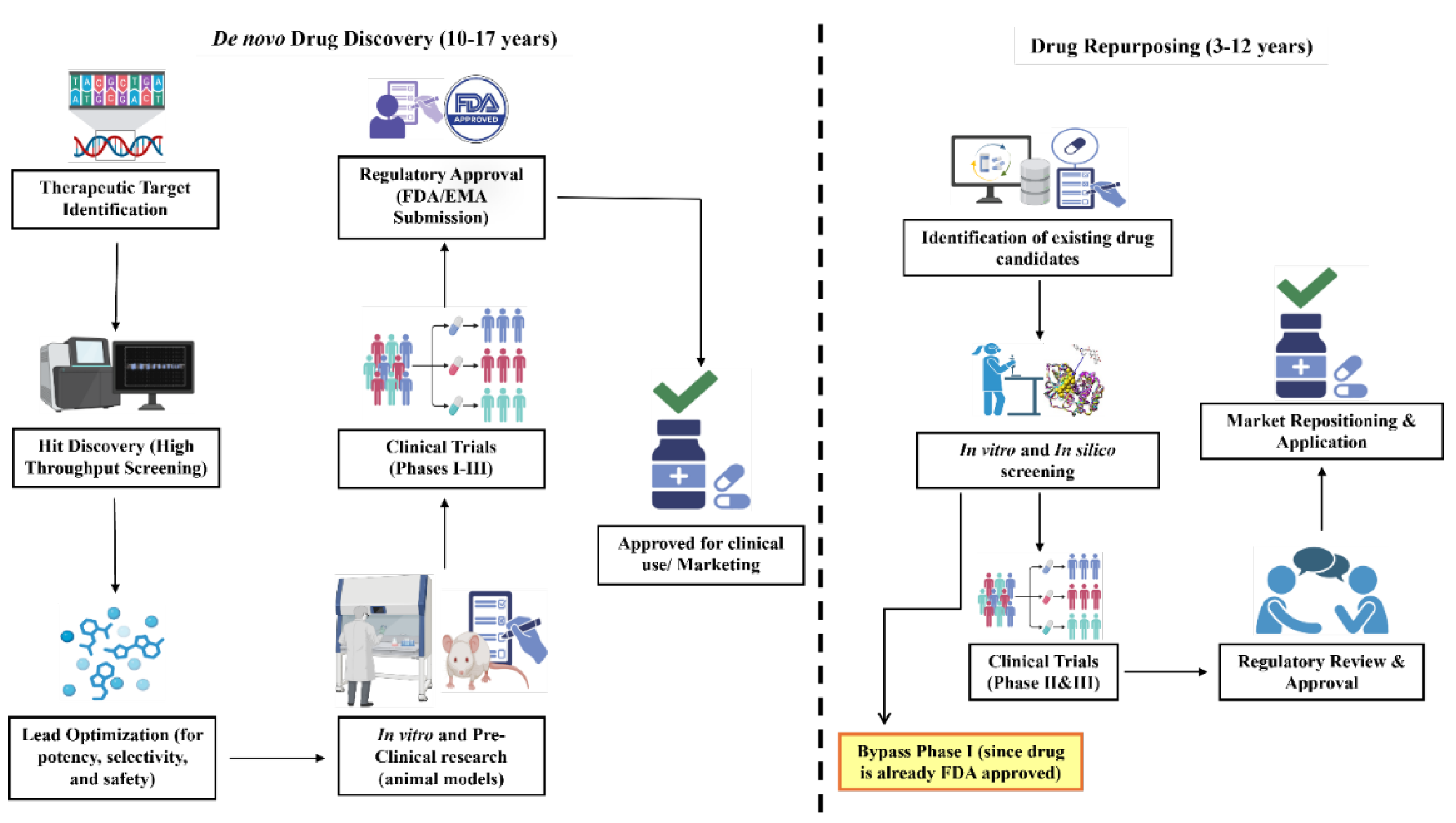
Figure 2.
Mechanistic overview of repurposed drugs targeting P. aeruginosa virulence pathways, including (1) Inhibition of AHL biosynthesis (Albendazole, Metformin, Cefepime, Imipenem); (2) Bacterial motility inhibition (Propranolol, Niclosamide, Albendazole, Metformin, Sitagliptin, Aspirin, Diclofenac, Paracetamol, Naproxen, Azithromycin, Ceftazidime, Doxycycline, Metronidazole, Secnidazole, Clofoctol, Ceftriaxone, Nitrofurazone, Salicylic acid, Allopurinol, Cilostazol); (3) Attenuation of virulence factor production (Flurocytosine, Miconazole, Glyceryl trinitrate, Propranolol, Guanfacine, Niclosamide, Ivermectin, Albendazole, Metformin, Sitagliptin, Dimetridazole, Aspirin, Tenoxicam, Diclofenac, Naproxen, Aceclofenac, Erythromycin, Azithromycin, Ceftazidime, Ciprofloxacin, Doxycycline, Metronidazole, Cefepime, Imipenem, Secnidazole, Clofoctol, Nitrofurantoin, Ceftriaxone, Cefoperazone, Nitrofurazone, Salicylic acid, Sodium salicylate, Vitamin E &K, Allopurinol, Ribavirin, Phenothiazine, Fexofenadine, Cilostazol, Levocetrizine, Atorvastatin); (4) Inhibition of biofilm formation (Ciclopirox, Miconazole, Glyceryl trinitrate, Guanfacine, Niclosamide, Albendazole, Dimetridazole, Metformin, Sitagliptin, Aspirin, Diclofenac, Paracetamol, Naproxen, Azithromycin, Doxycycline, Metronidazole, Secnidazole, Clofoctol, Ceftriaxone, Cisplatin, Nitrofurazone, Sodium salicylate, Vitamin E & K, Allopurinol, Ribavirin, Phenothiazine, Cilostazol); (5) QS receptor binding (Miconazole, Glyceryl trinitrate, Propranolol, Ivermectin, Albendazole, Metformin, Sitagliptin, Aspirin, Naproxen, Aceclofenac, Ceftazidime, Nitrofurantoin, Ceftriaxone, Phenothiazine, Fexofenadine, Levocetrizine, Atorvastatin); and (6) Inhibition of downregulation of virulence genes (Flurocytosine, Ciclopirox, Albendazole, Sitagliptin, Aspirin, Dimetridazole, Diclofenac, Naproxen, Ceftazidime, Secnidazole, Clofoctol, Cefoperazone, Sodium salicylate, Ribavirin, Cilostazol).
Figure 2.
Mechanistic overview of repurposed drugs targeting P. aeruginosa virulence pathways, including (1) Inhibition of AHL biosynthesis (Albendazole, Metformin, Cefepime, Imipenem); (2) Bacterial motility inhibition (Propranolol, Niclosamide, Albendazole, Metformin, Sitagliptin, Aspirin, Diclofenac, Paracetamol, Naproxen, Azithromycin, Ceftazidime, Doxycycline, Metronidazole, Secnidazole, Clofoctol, Ceftriaxone, Nitrofurazone, Salicylic acid, Allopurinol, Cilostazol); (3) Attenuation of virulence factor production (Flurocytosine, Miconazole, Glyceryl trinitrate, Propranolol, Guanfacine, Niclosamide, Ivermectin, Albendazole, Metformin, Sitagliptin, Dimetridazole, Aspirin, Tenoxicam, Diclofenac, Naproxen, Aceclofenac, Erythromycin, Azithromycin, Ceftazidime, Ciprofloxacin, Doxycycline, Metronidazole, Cefepime, Imipenem, Secnidazole, Clofoctol, Nitrofurantoin, Ceftriaxone, Cefoperazone, Nitrofurazone, Salicylic acid, Sodium salicylate, Vitamin E &K, Allopurinol, Ribavirin, Phenothiazine, Fexofenadine, Cilostazol, Levocetrizine, Atorvastatin); (4) Inhibition of biofilm formation (Ciclopirox, Miconazole, Glyceryl trinitrate, Guanfacine, Niclosamide, Albendazole, Dimetridazole, Metformin, Sitagliptin, Aspirin, Diclofenac, Paracetamol, Naproxen, Azithromycin, Doxycycline, Metronidazole, Secnidazole, Clofoctol, Ceftriaxone, Cisplatin, Nitrofurazone, Sodium salicylate, Vitamin E & K, Allopurinol, Ribavirin, Phenothiazine, Cilostazol); (5) QS receptor binding (Miconazole, Glyceryl trinitrate, Propranolol, Ivermectin, Albendazole, Metformin, Sitagliptin, Aspirin, Naproxen, Aceclofenac, Ceftazidime, Nitrofurantoin, Ceftriaxone, Phenothiazine, Fexofenadine, Levocetrizine, Atorvastatin); and (6) Inhibition of downregulation of virulence genes (Flurocytosine, Ciclopirox, Albendazole, Sitagliptin, Aspirin, Dimetridazole, Diclofenac, Naproxen, Ceftazidime, Secnidazole, Clofoctol, Cefoperazone, Sodium salicylate, Ribavirin, Cilostazol).
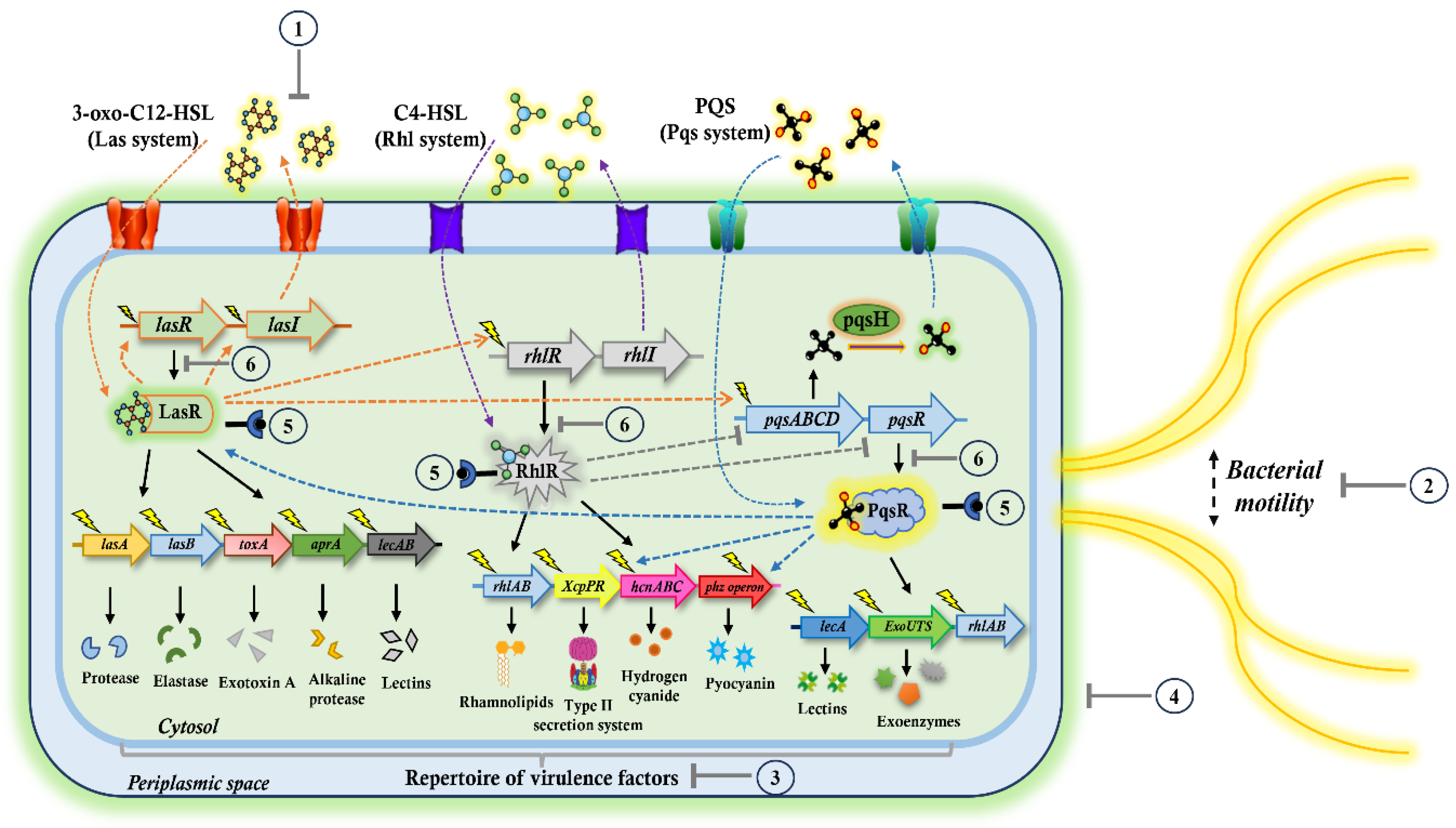

Table 1.
Repurposed antifungal drugs against P. aeruginosa showing antivirulence potential.
| Drug | Repurposed use | References | ||
|---|---|---|---|---|
| In vitro findings | In vivo findings | In silico findings | ||
| 5-Fluorocytosine | Reduction in pyoverdine production (~4 folds) by downregulating pvdS transcription | Protected mice from fatal effects of P. aeruginosa | - | [56] |
| Ciclopirox | Biofilm reduction and mexC gene downregulation | - | - | [57] |
| Miconazole | Curtails pyocyanin (47–49%), hemolysin (59%), rhamnolipid (42–47%) and protease production (36–40%) along with biofilm inhibition (45–48%) | Rescued mice from PAO1 infection | Suggests strong binding between miconazole and LasR, RhlR, and PqsR proteins with binding energies −9.069, −6.613, −6.485 kcal/mol, respectively | [60] |
Table 2.
Repurposed antihypertensive drugs against P. aeruginosa showing antivirulence potential.
| Drug | Repurposed use | References | ||
|---|---|---|---|---|
| In vitro findings | In vivo findings | In silico findings | ||
| Glyceryl trinitrate (GTN) | Eradicates biofilm (67%); impedes pyocyanin (75%) and protease (79%) production | - | Showed interaction between GTN and LasR (-93.47 Kcal/mol) and RhlR (-77.23 kcal/mol) | [61] |
| Guanfacine | Inhibited biofilm formation and pyocyanin (~1.5 folds) production | - | - | [62] |
| Propranolol | Reduced production of virulence factors (protease, hemolysin and pyocyanin production), motility phenotypes and biofilm formation (~2.5 folds) | Showed protection in mice from pseudomonal infections | Revealed high binding capacity of propranolol with LasR | [63] |
Table 3.
Repurposed antiparasitic drugs against P. aeruginosa showing antivirulence potential.
| Drug | Repurposed use | References | ||
|---|---|---|---|---|
| In vitro findings | In vivo findings | In silico findings | ||
| Niclosamide | Hampered biofilm formation (2-folds), swarming motility and production of pyocyanin (85-90%), elastase, protease and rhamnolipids (~25%) | Reduced virulence in G. mellonella insect model | - | [65] |
| Dimetridazole | Silenced transcription of virulence genes: lasR, rhlR, and pqsR (~2-6 folds) along with reducing protease, pyocyanin production and biofilm formation | Rescued both C. elegans and mice from pseudomonal infection | - | [66] |
| Albendazole | Attenuates hemolysin (33%), alginate (37%), protease, rhamnolipids (29%), elastase and pyocyanin (47%) production; restrained motility phenotypes; suppressed expression of lasI, lasR, rhlI, rhlR, pqsA and pqsR genes; showed antifouling response against PAO1 | - | Exhibited strong association with LasR, RhlR and PqsR receptors with binding energy of -8.8, -6.5 and -6.3 kcal/mol, respectively | [67] |
| Ivermectin | Diminished production of pyocyanin (29%), hemolysin (69%), pyochelin (58%) and protease (24%) | - | Showed high binding affinity with PqsR (-11.6 kcal/mol) | [68] |
Table 4.
Repurposed antidiabetic drugs against P. aeruginosa showing antivirulence potential.
| Drug | Repurposed use | References | ||
|---|---|---|---|---|
| In vitro findings | In vivo findings | In silico findings | ||
| Metformin | Inhibition of pyocyanin, hemolysin, protease and elastase activity; anti fouling potential (~67%); restrained swimming and twitching motilities | - | Revealed significant interaction with LasR (-6.4 kcal/mol) and RhlR (-6.0 kcal/mol) receptor of P. aeruginosa | [69, 73] |
| Sitagliptin | Attenuates pyocyanin, hemolysin (~92%), protease and elastase production; inhibited swimming, swarming, twitching motilities; and biofilm formation; downregulated expression of lasI, lasR, rhlI, rhlR, pqsA and pqsR genes | - | Showed strong association with LasR quorum sensing receptor of P. aeruginosa | [70] |
| Sitagliptin | Suppressed expression of virulence genes; inhibited P. aeruginosa’s virulence enzymes, pyocyanin production, motility phenotypes and biofilm production (~55%) | Rescued mice form P. aeruginosa infection, showing 100% survival | Showed multiple binding interactions with QS receptors | [71] |
| Metformin nano emulsions (MET-NEs) | Supressed swarming motility (89–94%); reduction in pyocyanin (60–80%) production, protease activity (78–99%), and pqsA gene expression compared to metformin alone | Showed protective activity against pseudomonal infections | - | [72] |
| Combination of Vildagliptin & Metformin | Diminish biofilm formation, bacterial motility, and the production of virulent extracellular enzymes and pyocyanin pigment (~30%); downregulated expression of QS-encoding genes | Reduced P. aeruginosa infection in mice | Revealed strong affinity with LasR, QscR, and PqsR receptor of P. aeruginosa | [74] |
Table 5.
Repurposed NSAIDs against P. aeruginosa showing antivirulence potential.
| Drug | Repurposed use | References | ||
|---|---|---|---|---|
| In vitro findings | In vivo findings | In silico findings | ||
| Aspirin | Reported significant inhibition of elastase, total protease and pyocyanin production; antifouling activity; restrained motilities; suppressed expression of virulence genes (lasI, lasR, rhlI, rhlR, pqsA and pqsR) | - | Revealed strong cohesion between aspirin and LasR receptor with S score of -12.02 | [75] |
| Tenoxicam | Attenuates pyoverdine (7%), rhamnolipids (27%), pyocyanin (29%), elastase, proteases (34%), and hemolysin production | P. aeruginosa-infected mice showed 80% survival | - | [76] |
| Paracetamol | Antifouling activity (~67%) ; reduced swarming motility (~58%) | - | - | [77] |
| Diclofenac loaded PLGA (Poly (lactic-co-glycolic acid)) nanoparticles (NPs) | Downregulation in lasI (0.28-0.57-fold) and lasR (0.07-0.39-fold) gene expression; anti-hemolytic activity; supressed biofilm formation (9–27%) and twitching motility | - | - | [78] |
| Naproxen | Curtails bacterial protease (~25), hemolysin, pyocyanin, biofilm (48- 63%), and motility; restrained lasI and rhlI gene expression | - | Showed high affinity towards P. aeruginosa’s QS-receptors | [79] |
| Aceclofenac (AcF) | Represses pyocyanin (16%), protease (20%), hemolysin (55%) and pyochelin (37%) production | - | Demonstrated strong associations with LasR, RhlR and PqsR receptor of P. aeruginosa having binding energies –8.8, –8.5, –7.7 kcal/mol |
[68] |
Table 6.
Repurposed antibiotics against P. aeruginosa showing antivirulence potential.
| Drug | Repurposed use | References | ||
|---|---|---|---|---|
| In vitro findings | In vivo findings | In silico findings | ||
| Erythromycin | Anti-proteolytic and anti-hemolytic activity | Reduced virulence in mice upon pseudomonal infection | - | [80] |
| Azithromycin (AZM) | Diminishes production of elastase, protease, and hemolysin; curtails motility phenotypes (swimming, swarming and twitching) and biofilm formation (~66%) | - | - | [81, 82] |
| Ceftazidime (CFT) | Attenuates elastase (63%), protease (56%), hemolysin (58-72%) and pyocyanin (61%) production; restrains swarming motility (82%); downregulated PQS-activated transcription | Showed improved virulence in P. aeruginosa-infected C. elegans | Reveals high binding affinity for RhlR and LasR domains (–7.54 and –7.31 kcal/mol) of P. aeruginosa | [81, 85, 86] |
| Ciprofloxacin (CPR) | Reduced the production of elastase (~50%), protease (~34%), and hemolysin | - | - | [81] |
| Doxycycline | Abrogated elastase (67%), pyocyanin (69%), and protease (65%) production; suppresses swarming motility (74%) and development of biofilm in PAO1 | - | - | [83] |
| Metronidazole | Represses production of pyocyanin (44%), pyoverdine (83%), protease (60%) and hemolysin; inhibits swimming and twitching motility; antifouling activity (87%) | - | - | [84] |
| Cefepime | Inhibits hemolysin (69-83%), elastase (61-70%) , total protease (51-61%) and pyocyanin (63-73%) production | - | - | [86] |
| Imipenem | Anti-proteolytic activity (50-62%); attenuates elastase (52-66%), pyocyanin (51-57%) and hemolysin (55-69%) production | - | - | [86] |
| Secnidazole | Suppression of QS-related genes (lasI, lasR, rhlI, rhlR, pqsA, and pqsR); Inhibits QS-related virulence factors; curtails swimming and twitching motility; diminishes biofilm formation | Reduction in mortality in P. aeruginosa-infected mice | - | [87] |
| Clofoctol | Abolishes pyocyanin production, swarming motility and biofilm formation; downregulation of QS-controlled genes | - | - | [17] |
| Nitrofurazone | Ceases pyocyanin, rhamnolipids production, and swarming motility; antifouling activity | - | - | [88] |
| Ceftriaxone (CT) | Disrupted motility phenotypes, pyocyanin production (41%), biofilm formation | Enhanced survival of C. elegans infected with PAO1 | Demonstrates high binding affinity towards LasR (-6.6 kcal/mol) and PqsR (-6.7 kcal/mol) receptors | [89] |
| Cefoperazone | Diminishes expression of lasI (77%) and rhlI (44%) genes; represses QS-related virulence factors production | - | - | [90] |
| Nitrofurantoin (NT) | Abrogated pyochelin (33%), pyocyanin (82%,), hemolysin (77%) and total protease (18%) production | - | Showed strong interaction with LasR, RhlR and PqsR receptor having binding energies –8.5, –7.6 and –6.7 kcal/mol, respectively | [68] |
Table 7.
Repurposed drugs as anti-virulence potential against P. aeruginosa.
| Drug | Therapeutic purpose | Repurposed use/Antivirulence potential | References |
|---|---|---|---|
| Salicylic Acid | Analgesic and Anti-inflammatory agent | Curtails twitching motility and production of protease (37%) | [91] |
| Cisplatin | Anticancer drug | Eradication of in vitro and in vivo biofilms (~ 99%) | [92] |
| Sodium salicylate | Analgesic, Antipyretic, Anti-inflammatory | Downregulation of expression of QS-related genes along with reduced virulence factors production (pyocyanin, siderophore production and biofilm formation (59 %)) | [93] |
| Allopurinol | Treatment of gout and used as eye drops (0.4%) | Diminishes QS-controlled virulence factors, antifouling potential (61%), inhibits motility phenotypes (swimming: 92%; twitching: 87% & swarming: 85%) | [94] |
| Vitamin E and K1 | Antioxidant and Blood clotting | Antibiofilm activity (Vitamin E: 37%, Vitamin K1: 63%); significant inhibition of pyocyanin production (75% & 60%), pyoverdine production (61% & 60%), and protease activity (87% & 43%) | [95] |
| Ribavirin | Antiviral drug | Disarming the QS-controlled proteases, pyocyanin and biofilm formation along with suppressing regulatory genes lasR, rhlR, and pqsR | [66] |
| Phenothiazine | Antipsychotic drug | Attenuating virulence factors production such as hemolysin, protease, rhamnolipid activity, pyocyanin production and biofilm formation; higher binding affinity to LasR, RhlR, and PqsR QS-proteins; reduced pathogenesis in vivo | [60] |
| Cilostazol | Antiplatelet and a vasodilator drug | Anti-proteolytic and antifouling activity, inhibition of swarming motility, diminished pyocyanin production; downregulation of QS-gene regulation; protection of mice against pathological changes in liver, spleen and kidney tissues | [96] |
| Fexofenadine (FeX) | Antihistamine drug | Diminished the pyochelin, pyocyanin (71%), hemolysin (81%) and total protease production in PAO1; and strong association between FeX and PqsR receptor | [68] |
| Levocetrizine (LvC) | Antihistamine drug | Inhibited the phenotypic virulence by reducing hemolysin, protease, pyocyanin and pyoschelin production; acts as ligand for LasR (–7.5 kcal/mol) receptor of PAO1 | [68] |
| Atorvastatin (AtS) | Anti-cholesterol drug | Anti-proteolytic activity, curtailed pyocyanin (77.24%), pyochelin (70%) and hemolysin (77.1%) production; high binding affinity towards PqsR receptor | [68] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



