
Submitted:
02 April 2025
Posted:
03 April 2025
You are already at the latest version
Abstract
Bats exhibit a unique repertoire of vocal behaviors, with many species employing echolocation to actively sense their environment, while using communication calls for social interactions. This review explores the neural circuits underlying these behaviors, from brainstem and midbrain networks that generate and control vocalizations, to higher-order brain regions, such as the frontal cortex, which may contribute to modulation of vocal behaviors. While much is known about brainstem mechanisms for echolocation, less is understood about the neural control of communication calls and the integration of these systems. Recent findings highlight the interplay between auditory, motor, and spatial processing networks in shaping bat vocalizations, with evidence for both shared and distinct neural pathways for echolocation and communication. We propose a framework for vocal production circuits in bats, synthesizing findings from diverse species and experimental techniques, and identify key questions to guide future research. This review underscores the importance of bats as models for studying vocal production and how they can provide insights into the evolution and adaptation of neural mechanisms across species.
Keywords:
vocal production
; bats
; neural activity
; neuroethology
Introduction
Bats are highly vocal animals, and the majority of species in this megadiverse order (comprising over 1,400 species) use their vocalizations for two main and quite distinct purposes. Firstly, many bats produce echolocation calls to actively sense their environment by processing the returning echoes from their surroundings. Secondly, bats also emit diverse social communication calls (Figure 1), with some species producing complex songs (Jahelková et al., 2008; Smotherman et al., 2016) and showing evidence for vocal learning (Elie et al., 2024; Knörnschild et al., 2012).
In general, bats use an astounding variety of ecological niches and exploit nearly all possible food sources in nature (such as insects, pollen, fruit, nectar, and blood (Denzinger et al., 2016)). The design of ultrasonic echolocation pulses has come to vary widely between bat species, reflecting the fine-tuning of each species' sensory and physical traits to its specific environment. The frequency composition, length, timing, and structure of calls all differ across each species’ orientation signals. While some bat species from the same family may exhibit remarkably different vocal adaptations in echolocation, such as members of the Vespertilionidae, other distantly related species have evolved strikingly similar echolocation calls. Most bats emit short (1-3 ms), downward frequency-modulated (FM) sweeps (Jones & Teeling, 2006), such as those in the genera Myotis or Carollia (Figure 1E). Several species of bats emit orientation signals that feature a prominent “constant frequency” (CF) component, flanked on one or both sides by a short FM component (together, FM-CF echolocation), and a longer overall call (e.g. up to 70 ms, Figure 1A). Only a handful of species are known to use FM-CF echolocation pulses, including horseshoe bats (Rhinolophus spp.), hipposiderid bats (Hipposideros spp.), and the mustached bat (Pteronotus parnellii), species which originate in different suborders (Jones & Teeling, 2006), making this echolocation strategy an example of convergent evolution.
Bats that perform FM-CF echolocation must solve different problems than bats that employ FM echolocation pulses. Bats with FM echolocation pulses rely more strongly on time information conveyed by returning echoes, and therefore rapidly and flexible adapt the pulse emission rate as they approach objects (as exemplified, e.g. in the terminal buzz of Eptesicus fuscus (Moss & Surlykke, 2001)). As these echolocation pulses are easily degraded by overlapping sounds, FM bats tend to shorten pulse duration during approaches and in cluttered environments (Jones & Teeling, 2006; Moss et al., 2006). Meanwhile, FM-CF echolocators use longer calls that are more tolerant to overlap, since the CF component of emitted pulses and returning echoes will have distinct, non-overlapping frequencies (this strategy will be discussed in detail below. For a review, see Hiryu et al., 2016). The long CF component is also a feature which aids in predation, as movement from prey insects is detected as perturbations in the CF component of returning echoes (Schnitzler & Denzinger, 2011). Importantly, these different echolocation design strategies may each entail specialized neuroanatomical circuits to produce them and evaluate the resulting echoes.
While bats are famous for their echolocation behavior, they also communicate using a large repertoire of vocalizations which are each deployed in specific social contexts (such as distress or during territorial combat, Knörnschild et al., 2014), with some species showing highly specialized behaviors such as singing (Smotherman et al., 2016), vocal imitation of adult calls by pups (Knörnschild et al., 2010), and infant-directed vocalizations (Fernandez & Knörnschild, 2020). Bats are one of the few mammals in which evidence of vocal learning has been found (Knörnschild, 2014; Lattenkamp & Vernes, 2018; Vernes & Wilkinson, 2019), and juveniles of the Greater sac-winged bat (Saccopteryx bilineata) go through a “babbling” phase, where they attempt and slowly learn to produce adult-like syllables (Fernandez et al., 2021). Meanwhile, adults may emit social “contact” calls (Carter et al., 2009), and social group membership may be signalled through a vocal signature, analogous to human dialects (Esser & Schubert, 1998; Knörnschild et al., 2012).
The brain circuitry and neuronal activity underlying vocal behavior has been the subject of many studies over the past years. While a large body of literature has focused on the role of brainstem networks in echolocation calls, much less is known about neural control of communication calls, and in what way these brain circuits overlap or diverge. Furthermore, while recent studies have explored the role of higher-order brain regions, such as the frontal cortex, our understanding of their specific contributions in vocal production remains limited.
In this review, we aim to bring together diverse findings from in vivo electrophysiological experiments, as well as pharmacological and electrical manipulations, which together seek to describe, perturb and identify the neural underpinnings of executive motor control of vocal production. Taking a bottom-up approach, we will discuss investigations in brain regions from brainstem circuits to cortical contributions, highlight functional interactions with auditory and spatial processing, and propose a tentative pathway for vocal production in bats. The brain areas implicated in vocal production and control in bats are summarized in Figure 2, and they will be discussed in detail throughout this review. We emphasize that our current knowledge of bat vocalization is derived from diverse species models, techniques, and occasionally conflicting empirical findings, making it challenging to create a unified overview of the field. Our aim is to chart progress to date and highlight unresolved questions that can guide future studies of vocal production behavior in bats and other animal models.
Vocal Control in Brainstem and Midbrain Circuits
I. Laryngeal Output and Its Control in the Medulla
Nearly all echolocating bats use the larynx to produce echolocation calls. The primary exceptions are bats within the genus Rousettus, which perform tongue clicks to emit broad-spectrum sonar pulses (Figure 1C).
Sounds used for the detection and localization of objects must have certain properties: they must have short wavelengths (i.e. be high in pitch), in order for the sound wave to reflect off the surface of small objects (Hartridge, 1920), and be short in duration, to permit the computation of object location in space, particularly for proximate objects.
The bat larynx exhibits several evolutionary adaptations to meet the demands of echolocation, such as ossified cricoid and thyroid cartilages and exceptionally fast laryngeal muscles (Carter, 2020; Elemans et al., 2011). These specializations make it possible to produce high-frequency echolocation pulses with call durations and inter-pulse intervals in the millisecond or even sub-millisecond range, while still allowing for the production of communication calls in lower frequencies and on longer time scales (Jones & Teeling, 2006). Some bat species, such as Daubenton’s bats (Myotis daubentonii) of the vespertilionid family, can expand their frequency range to up to 7 octaves, producing the lowest frequency portion by recruiting ventricular folds, and achieving exceptionally high frequencies through the use of specifically adapted vocal membranes (Håkansson et al., 2022).
In the mammalian vocal system, laryngeal muscles are innervated by motor neurons that are in turn controlled by structures within the brainstem. The nucleus ambiguus (NA) within the medulla has attracted special attention in the search for the neural basis of vocal production, as it presents the architecture required for vocal motor control: it projects to motor neurons that, in addition to the laryngeal muscles, also control supralaryngeal articulator muscles and expiration (Jürgens, 2009; Nieder & Mooney, 2020), it is innervated by the periaqueductal gray (PAG, see below) and receives projections directly from the motor cortex in at least two distantly-related bat species, Carollia perspicillata (Nevue et al., 2023) and Rousettus aegyptiacus (Wirthlin et al., 2024).
Electrophysiological recordings in the NA of the Rufous horseshoe bat (Rhinolophus rouxii) revealed neuronal spike patterns time-locked to onsets or offsets of their FM-CF echolocation calls. NA neuron firing started tens of milliseconds before vocalization onset and abruptly stopped a few milliseconds before the end of the calls, and firing rates were positively correlated with the emitted call frequency (Rübsamen & Betz, 1986). It is an open question whether NA neurons track call durations or other properties of echolocation calls (such as frequency) in bat species with shorter FM echolocation signal designs as well as in FM-CF echolocators.
II. The Periaqueductal Gray and Pontine Nuclei
Upstream of the NA (Figure 2), neurons in the bat PAG play a key role in the control and ‘gating’ of innate vocalizations, as has been widely demonstrated in rodents, cats, and primates (Jürgens, 2009; Nieder & Mooney, 2020; Tschida et al., 2019). Several studies investigating different bat species have demonstrated that stimulation in the PAG elicits vocalizations (see Figure 3 for an overview).
When Suga et al. (1973) electrically stimulated the lateral PAG in species of phyllostomids, noctilionids and vespertilionids – among them the Big brown bat (Eptesicus fuscus) –, all bats uniquely produced echolocation calls which were, in all cases, similar to their species-specific echolocation signal. In contrast, when Valentine et al. (2002) electrically stimulated the lateral PAG of E. fuscus, they only observed the production of communication calls. However, both echolocation and a suite of communication calls were observed following electrical stimulation to the lateral and ventro-lateral areas of the caudal PAG in the Pale spear-nosed bat (Phyllostomus discolor, Fenzl & Schuller, 2002).
Taken together, these results may suggest that these two vocalization regimes in bats are controlled by distinct regions within the vocally-selective portions of the PAG, as has been demonstrated in primates (Jürgens 1994) and cats (Subramanian et al. 2021). However, as it is difficult to compare the precise regions targeted across these studies and especially challenging to do so between species, it is also possible that the bat PAG features spatial overlap of call type-specific neural populations, as evidenced in mice (Tschida et al., 2019), or even that there are subtle differences between bat species with regards to the functional design of the PAG.
The PAG communicates extensively with neighboring structures within the dorsal pontine tegmentum, such as the paralemniscal area (PLA, Schuller et al., 1997). When neuronal activity in this brain region is blocked pharmacologically, stimulation in the PAG can no longer elicit echolocation calls, but PAG-induced communication calls are still observed (in P. discolor, Fenzl & Schuller, 2005). By comparison, pharmacological or electrical stimulation of the PLA alone leads only to the production of echolocation calls, but never communication calls in rhinolophid and phyllostomid bats (Fenzl & Schuller, 2002; Schuller & Radtke-Schuller, 1990).
Neurons in the PLA exhibit varied response properties during echolocation production. Distinct populations of vocally-activated neurons exhibit complex firing patterns that are unique to instances when the bats (R. rouxii) vocalize, as opposed to simply hearing playback of their own vocalizations (see below, Metzner, 1989, 1993).
Caudal to the PLA is the parabrachial complex, a group of nuclei mainly serving as a relay of cardiovascular, gastrointestinal and respiratory information between the medulla and higher structures (Chamberlin, 2004). As bats vocalize during the expiration phase of a breath (Suthers et al., 1972), it follows that the vocal pathway is tightly linked with respiratory control.
When Smotherman et al. (2006) enhanced GABAergic (i.e. inhibitory, see Glossary) synaptic activity in the parabrachial complex of the greater horseshoe bat (Rhinolophus ferrumequinum), they not only observed longer respirations but also prolonged echolocation call durations. In contrast, when GABAergic synaptic activity was blocked, the breaths became shorter and the animals were no longer able to accommodate two echolocation calls in one expiration, a typical behavior in this animal.
Together with the above, these findings indicate that the midbrain and brainstem nuclei perform a suite of complex operations that range from the ‘switching on and off’ of vocalization to fine-tuned control of complex vocal behaviors that crucially depend on sensory feedback. PAG-generated signals for echolocation depend on intact neuronal activity patterns in the PLA to achieve the corresponding motor action in the larynx, while PAG signals for communication calls may act through alternative pathways. Meanwhile, pontine structures medial to the lateral lemniscus are a site of convergence for multiple functions, serving the top-down vocal-motor output circuitry and bottom-up auditory processing functions in parallel.
The audiovocal interface: auditory and motor circuits
I. Control of vocalization is feedback-dependent: Examples from Doppler-shift compensating bats
The proper control of vocalization requires tight integration between sensory processing, premotor preparation, and motor execution. When hearing is disrupted, as in deaf populations, it becomes difficult or impossible to control vocal pitch and loudness (Monsen et al., 1979). A clear and elegant example illustrating the importance of the ascending auditory pathway in vocal production is the Doppler shift compensation behavior. In 1968, Schnitzler discovered that R. ferrumequinum emits FM-CF calls with a particular frequency while at rest, but will lower the pitch of the CF component during flight by 150-200 Hz, to compensate for flight-induced Doppler shift of the returning echo (Schnitzler, 1968). The purpose of this compensation is to ensure that the echo returns to the bat at approximately the same frequency to which the auditory system is most sensitive (Neuweiler, 1980; Suga et al., 1976). This adjustment of the echolocation frequency was named Doppler shift compensation and has been observed in many species with FM-CF echolocation, such as rhinolophids (Smotherman & Metzner, 2003), hipposiderids (Schuller, 1980), and mormoopids (Gaioni et al., 1990).
Appropriate adjustment of the constant frequency call naturally requires auditory feedback needed to compute the degree of pitch compensation. How then, do different auditory regions contribute to this vocal behavior?
Metzner demonstrated that the paralemniscal area in rhinolophid bats features different populations of vocally active neurons. Some neurons show suppression during vocalization, while another subpopulation of neurons ramp up activity in the pre-vocal phase, and/or maintain high firing rates for the duration of the vocalization. Interestingly, these vocally-active neurons exhibited echo-delay sensitivity, firing when artificial echoes were played back to the animal shortly (5-18 ms) after echolocation onset, but not when echoes were played following playback of a call (Metzner, 1989, 1993).
The vocal-related inhibition seen here, which lasts for the duration of the vocalization, may be driven by inhibitory inputs from nuclei of the lateral lemniscus, which is the first station along the ascending auditory pathway which exhibits attenuated responses to self-generated sounds (Suga & Schlegel, 1972; Suga & Shimozawa, 1974). We note that this ‘corollary discharge’ mechanism may still originate upstream of the inferior colliculus, from higher-order vocal control regions (Crapse & Sommer, 2008). This inhibition is critical in the case of the echolocating bat. While other animals that communicate vocally must also deal with self-stimulation, bats risk becoming desensitized to returning echoes if auditory-responsive neurons are already engaged in responding to their own vocalizations.
In subsequent studies, systematic perturbation of the paralemniscus-adjacent parabrachial nucleus using pharmacological agents targeting GABAergic and glutamatergic neuronal populations showed that local inhibitory activity was crucial both for maintaining the constant frequency at the preferred frequency at rest, and for performing the correct pitch compensation. When GABA antagonists or glutamate agonists were injected into the region, the resting frequency rose, and the bats could not lower the CF component in response to playback which mimicked the Doppler shift induced by natural flight. Conversely, when GABA agonists or glutamate antagonists were applied, the resting frequency lowered, and the bats over-compensated for the Doppler shift, lowering CFs below levels observed in typical conditions (Smotherman et al., 2003).
In these manipulations, FM-CF echolocation calls were shifted in pitch as a whole. Only when the investigators then injected the GABA antagonist into the principal sensory nucleus of the trigeminal region, ventral and caudal to the parabrachial nucleus, was the frequency, amplitude, and temporal structure of the call itself perturbed (Smotherman et al., 2003).
Together, these results indicate that the pontine structures ventral to the PAG and the inferior colliculus and medial to the lateral lemniscal fibers perform specialized functions important for (1) computing necessary vocal adjustments following sensory input, (2) inhibiting population responses to self-generated sounds while maintaining high sensitivity for returning echoes, and (3) maintaining the frequency and temporal structure of vocalizations even as they are being produced.
II. Auditory cortex involvement in sensory feedback and production
At the level of the cerebral cortex, the auditory cortex (AC) of bats is highly specialized to accommodate the demands of echolocation. In all bats that have been studied using functional neuroanatomical and electrophysiological methods, the auditory cortex is hypertrophied, exhibiting a large area dedicated to the processing of high-frequency sounds (O’Neill, 1995). Similarly to their subcortical counterparts, these populations are recruited during vocalization.
In Doppler-shifting bats, such as P. parnellii, the tonotopic plane of the primary auditory cortex is not uniformly responsive to sounds across all frequencies, but features a large region dedicated to the frequencies surrounding the preferred frequency for echoes which Doppler-shifting aims to return to the bat. This exquisite sensitivity to a particular frequency originates in the inner ear (“auditory fovea”) and is observable across many stations along the auditory pathway, at the level of the basilar membrane (Bruns & Schmieszek, 1980; Kössl & Vater, 1985), the inferior colliculus (Neuweiler, 1980; Rübsamen et al., 1988; Schuller & Pollak, 1979), and the auditory cortex (Schnitzler & Denzinger, 2011).
AC neurons tuned to echolocation frequencies also exhibit sensitivity to echoes arriving at particular delays relative to vocal onset, in a phenomenon called “delay tuning” (O’Neill & Suga, 1979; Suga & O’Neill, 1979). In many species, delay tuned neurons appear to be organized in the spatial arrangement in the AC by their preferred delay duration, resulting in a “chronotopy” (Hagemann et al., 2010), analogous to tonotopy of primary auditory areas. This chronotopic arrangement of delay sensitive neurons is observable even in juvenile bats (Kössl et al., 2012).
The sensitivity of the AC to temporal information may be exploited to adjust vocal timing. Following lesioning of the AC, Yuma myotis bats (Myotis yumanensis) failed to adjust their call rate when flying towards obstacles (Suga, 1969).
In the foregoing, we have discussed how neural spiking activity correlates with vocalization and the adjustment of vocal parameters. However, local field potentials (LFP, see Glossary) in the AC may serve as an index of vocal-related neuronal activity, or even additionally present a functional contribution to vocal production. In Seba’s short-tailed bat (Carollia perspicillata), oscillations in the beta range [12-30 Hz] before the onset of vocalization are predictive of whether an echolocation or a communication call is about to be emitted (García-Rosales et al., 2022).
In addition to its central role in sound processing, and the plausible involvement in vocal adjustment described above, the AC may serve to prepare the auditory network for subsequent processing of echoes through its reciprocal connections with other forebrain regions (see below).
Cerebellar Contributions to Vocal Production
One of the best described functions of cerebellum is its role in movement control and regulation, which it executes both through inputs to brainstem motor circuits and through projections to the motor cortex that run via the thalamus, as described in primates (Jürgens, 2002; Tanaka et al., 2021). Its function in vocal control has rarely been investigated to date, but abnormal vocalizations after cerebellar lesions have been described in macaques (Macaca fuscata), and cerebellar disorders in humans can lead to deficits in speech production (Ackermann et al., 2007; Gemba et al., 1995).
In bats, the involvement of the cerebellum in vocalizations was demonstrated many years ago by (Horikawa & Suga, 1986) through cerebellar lesions in P. parnellii, which resulted in an increased variation of the CF component in FM-CF echolocation calls. A recent study from our research group on C. perspicillata revealed that cerebellar firing rates and LFP oscillation patterns differ depending on whether the bat produces an echolocation or a communication call (Hariharan et al., 2024) (see Figure 1 for example calls in both bat species). Spike patterns were predictive of the call type, even half a second before the onset of the vocalization, indicative of premotor activity.
Active sensing and social representations in spatial processing circuits
I. Echolocation fine-tuning and read-out in the superior colliculus
The mammalian superior colliculus (SC) has been implicated in spatial processing and orientation behaviors by integrating sensory information across different modalities, especially from the visual and acoustic domain (Hoffmann et al., 2016; Hoy et al., 2019; Krauzlis et al., 2013). Neurons in the bat SC show clear delay tuning, thereby encoding target distance and creating an egocentric map of the environment (Hoffmann et al., 2016; Kothari et al., 2018; Valentine & Moss, 1997). As the perception of spatial features is dependent upon echolocation call emissions, it seems the bat SC has extended its function from controlling eye or head movements to the initiation and shaping of echolocation calls: When Valentine et al. (2002) electrically stimulated the SC in E. fuscus, they not only observed head and pinna movements, but could elicit stereotypical echolocation calls. These calls occurred after a significantly longer latency than vocalizations induced by stimulation of the PAG, indicative of a longer pathway to motor neurons.
The production of echolocation calls through SC stimulation had already been shown in rhinolophid bats by Schuller & Radtke-Schuller (1990), who found that the amplitude level of the elicited call increased with increasing stimulation current, a feature they did not observe when eliciting calls from any other brainstem area. This finding points to an active role of the SC in shaping and adapting vocalizations depending on the current spatial conditions.
This function was further confirmed when Sinha & Moss (2007) and later also Wohlgemuth, Kothari, et al. (2018) recorded neuronal activity in the SC in E. fuscus while the bat was tracking a moving insect. Vespertilionid bats modify their FM call duration and interval depending on the distance of the target they pursue. When the target was further away, which was associated with a longer echolocation call, neurons in the SC fired 20 to 30 milliseconds before the onset of the call. As the target moved closer and the calls and call intervals became shorter, the first spike of this pre-vocal bout shifted closer in time to the call, up to 5 milliseconds before the onset of super short calls during the terminal buzz. Another set of spike bursts were consistently observed at 3 milliseconds before every echolocation call, independently of the subsequent call duration or call interval. Thus, on the one hand, the SC prospectively encodes echolocation call parameters, a signal that could be sent downstream to control motor output, and on the other hand, it signals the initiation of every echolocation call irrespective of its temporal characteristics, a potentially useful signal for priming the network for an incoming echo.
A follow-up study in E. fuscus could show that delay tuning of SC neurons is also influenced by echolocation properties (Kothari et al., 2018): While the bats were freely flying, exploring an environment with multiple obstacles, and naturally increased their call rate as they approached certain objects, neurons in the SC shifted their preferred echo delay from longer to shorter time periods and exhibited sharper tuning to these delays, obtaining a higher spatial resolution at close range. Sharpening in delay tuning with increasing echolocation rate may however be independent of active vocal production as it was observed in the AC and inferior colliculus (IC) to playback of call-echo sequences vs. single call-echo pairs (Beetz et al., 2016; Macías et al., 2018). High echolocation call rates may additionally be accompanied by an increase in the power of gamma oscillations [40-140 Hz] in the SC (Kothari et al., 2018), a rhythmic activity pattern that is known to emerge when groups of neurons engage in attention tasks (Fries, 2009), but which could also stem from sequences of evoked potentials triggered by echolocation calls produced at high rates.
There is also evidence for some functional mapping in the SC: Wohlgemuth, Kothari, et al. (2018) found that neurons in dorsal SC layers of E. fuscus are more active after an echolocation call, putatively processing sensory information, and neurons in ventral SC layers show pre-motor coding as they are primarily active a few milliseconds before call onset.
Taken together, these findings implicate the midbrain SC as a powerful network where spatial information is not only analyzed, but where sampling of the environment may be optimized through dynamic adaptation of echolocation behavior, according to the animal’s current position relative to targets and its need for finer spatial or temporal resolution.
II. Potential Contributions of the Inferior Colliculus
The IC, which is exposed on the dorsal brain surface in many bat species (van Tussenbroek et al. 2024), is not usually considered a spatial processing site across mammals, but rather as a critical station along the auditory pathway. However, in the special case of bats, IC neurons may indeed read out spatial information from echolocation calls, e.g. through delay tuning (Dear and Suga 1995; Macías et al. 2012). Although lesioning the IC does not seem to impair the production of echolocation calls (Suga 1969), this brain region may still play a role in controlling echolocation call parameters. When Diebold et al. (2024) transiently inactivated the IC in E. fuscus during free flight, bats not only performed more poorly in navigating through an obstacle, but also altered several echolocation call parameters. Specifically, they showed increased call rates, extended call durations and broader frequency bandwidths. This could either mean that bats require a functioning IC to control and adapt these aspects of echolocation calls, or that the behavioral changes reflect a compensatory mechanism to a deficit in processing spatial features of the environment.
Furthermore, the IC could play a role in social interaction and communication. When Salles et al. (2024) recorded IC spike rates of E. fuscus during social encounters, they found increased activity during communication calls associated with aggressive interaction, but also with calls produced during mating. However, it is not known whether spike rates differ for self-produced vocalizations versus calls of conspecifics.
III. Processing of Spatial and Social Information in the Hippocampus
Beyond the midbrain, other regions in the mammalian brain are also highly involved in spatial processing and therefore could, in the special case of bats, play a role in vocalizations, among them the hippocampus. As first described in rodents (O’Keefe & Dostrovsky, 1971) and later shown for many other mammals (Ekstrom et al., 2003; Ono et al., 1993; Ulanovsky & Moss, 2007) and birds (Payne et al., 2021), neurons in the hippocampus form place fields, becoming active whenever the animal is in a specific location within an environment.
That place cells are informed by echolocation calls, was shown by Ulanovsky & Moss (2011) in E. fuscus, when they recorded hippocampal activity while the bats explored an arena. Immediately after an echolocation call, the place field of the animal’s current location was clear and well localized. But as several hundreds of milliseconds passed after the call, the spatial code became less selective, resulting in larger and more diffuse place fields. The same group could also show a few years later that in periods with high call rates, hippocampal place fields are smaller, resulting in a higher resolution at close range (Wohlgemuth, Yu, et al., 2018). A similar feature had already been described for the egocentric code in the SC (Kothari et al., 2018). In primates and bats, hippocampal cells also code for the location to which the animal attends through visual gaze or a directional echolocation pulse (Rolls & O’Mara, 1995; Ulanovsky & Moss, 2011).
Further experiments indicate that hippocampal oscillatory activity is modulated by echolocation calls. Theta rhythms [~5-10 Hz], which are highly prominent in rodents during active exploration (Hasselmo, 2005), appear in bats only intermittently in bouts (Yartsev et al., 2011) and become more frequent with rising echolocation call rates (Ulanovsky & Moss, 2007).
Until now, research in the hippocampus has focused on the role of echolocation calls on spatial representations, but hippocampal activity during communication calls has not been investigated. Recent studies in the Egyptian fruit bat (Rousettus aegyptiacus) however suggest that hippocampal neurons also encode aspects of social interaction, such as the movement of a conspecific (Omer et al., 2018), the identity of other bats in a group setting (Forli & Yartsev, 2023), or even the identity of human experimenters (Snyder et al. 2024). Relatedly, in rodents the hippocampal subregion CA2 is known for its role in social interaction (Hitti & Siegelbaum, 2014; Oliva et al., 2020). This raises the question whether hippocampal activity is also modulated by the perception or production of social communication calls.
Forebrain Networks in Vocalization
I. Basal Ganglia and Dopaminergic Neurons
While midbrain circuits clearly play a central role in vocal production, several cortical structures and connected subcortical regions in the forebrain fulfill critical functions. Apart from the cortex, which will be discussed below, studies have investigated for example the caudate nucleus (CN) in the dorsal striatum, which, as part of the basal ganglia (BG), is a critical component in the motor system. When Schwartz & Smotherman (2011) investigated the expression of the immediate early gene C-Fos, a marker for neuronal activation, in the Mexican free-tailed bat (Tadarida brasiliensis), they found a clear increase in expression in the CN after periods of extensive vocalization, but not when the bats had been listening only to playback calls. Additionally, oscillatory LFP activity in the CN was linked to the type of call produced in C. perspicillata (Weineck et al., 2020). Specifically beta oscillations [12-30 Hz] were predictive of the call type, exhibiting higher power before the onset of echolocation calls compared to communication calls. Simultaneously, the CN showed call-type-dependent synchronized activity with an auditory-selective area in the frontal cortex, the frontal auditory field (FAF, see below). This synchrony was expressed as theta coherence, a measure of phase relationship and consistency between two structures (see Glossary), which was strongest before and during communication calls, but was reduced before the onset of echolocation calls.
In a previous study, Tressler et al. (2011) had demonstrated a causal role of dopaminergic neurons, which are an integral part of the BG, in vocal production. After systemic injection of a dopamine-selective neurotoxin T. brasiliensis showed clear changes in their echolocation calls as amplitude, duration and frequency bandwidth decreased. Surprisingly, similar effects were observed after the injection of a dopamine receptor agonist, which points to a nonlinear relationship between dopamine levels and vocal production.
II. Production of Emotive Vocalizations via the Amygdala
While the BG most likely play a role in fine motor control of all vocalizations, communication calls in particular may feature arousing and affective elements. An area highly involved in emotion and motivation across species is the amygdala (Janak & Tye, 2015). In several bat species, it has been shown that amygdala neurons respond differentially to communication calls of conspecifics, depending on their emotional significance (P. parnellii: Naumann & Kanwal, 2011; E. fuscus: Gadziola et al., 2016). Ma & Kanwal (2014) probed the role of the amygdala in vocal production by electrically or pharmacologically stimulating the basolateral or central nuclei in P. parnellii. After stimulation, bats elicited both echolocation and communication calls. While there were no clear region-specific boundaries regarding the call types, some topography was observed across the rostrocaudal axis, with more echolocation calls elicited after stimulation in caudal regions and more communication calls produced after rostral stimulations. The bats emitted several different types of communication calls, with the majority of evoked social vocalizations resembling calls emitted during aggressive encounters (see Figure 1B right). This is in accordance with the previously described function in aggression and fear of the rodent amygdala (Adhikari et al., 2015; Hong et al., 2014), but also highlights its complex role in various aspects of emotional behavior. Interestingly, vocalizations were emitted after a considerable latency of more than 200 milliseconds, with longer latencies on average for echolocation calls. This points to a long signaling pathway for vocal-motor execution that could potentially run through the cortex since the amygdala shares reciprocal connections with several cortical areas (Figure 2), for example the anterior cingulate cortex (ACC, Fillinger et al., 2017, 2018; Fitzpatrick et al., 1998).
III. Neuronal Activity Preceding Vocal Onset in Frontal Cortical Areas
In primates, the ACC has long been implicated in the voluntary control of vocalizations (Aitken, 1981; Hage, 2010). In bats, we find several indications that neuronal activity in the ACC is involved in vocal production: Electrical stimulation in the ACC of P. parnellii could elicit either echolocation or communication calls, depending on the area of stimulation. More anterior sites led to echolocation calls, whereas stimulation in posterior areas rather produced communication calls (Gooler & O’Neill, 1987). The calls were observed with a considerable latency of approximately 100 milliseconds after stimulation, indicative of a longer signaling pathway to the motor neurons which potentially runs through the PAG, as postulated in primates (Nieder & Mooney, 2020). In the ACC of T. brasiliensis, Schwartz & Smotherman (2011) identified increased expression of C-Fos after extended periods of vocalization, indicative of high neuronal firing. The expression of FoxP2, a gene implicated in the development and learning of language, has also been demonstrated in the ACC of multiple bat species of the rhinolophid, phyllostomid and pteropodid families (Rodenas-Cuadrado et al., 2017; Yin et al., 2017).
An increasing body of work has also implicated frontal cortical areas beyond the ACC in vocal production. A recent study on R. aegyptiacus, for example, recorded neuronal firing rates in dorsal parts of the frontal cortex and identified cells that were selectively active either to self-produced communication calls or to the vocalizations of conspecifics, with very little overlap between the populations (Figure 4, Rose et al., 2021). A subsequent study investigated the motor cortex of R. aegyptiacus (Wirthlin et al., 2024), and recorded neurons specifically in an area, where previously tongue, jaw and nose movements in response to electric stimulation had been reported (Halley et al., 2022), and which they termed as the orofacial motor cortex. In humans, damage to the orofacial motor cortex can strongly affect the ability to speak (Jürgens, 2002; Nieder & Mooney, 2020), and in the Alston’s singing mouse (Scotinomys teguina), a rodent from Central American rainforests that engages in elaborate territorial counter-singing and turn-taking behavior, inactivation of the orofacial motor cortex affects singing response to a conspecific’s playback (Okobi et al., 2019). In R. aegyptiacus, Wirthlin et al. (2024) found that neurons in this region fire during or even before self-produced spontaneous vocalizations, but are less active to the auditory stimuli of other calling bats. Interestingly, the motor cortex seems to be able to bypass the vocal motor pathway through the PAG and can target the NA in the medulla directly, as demonstrated through neural tracing in this study on R. aegyptiacus, but also in C. perspicillata (Nevue et al., 2023).
Multiple studies, including from our research group, have focused their investigations on the frontal auditory field (FAF), an area in the frontal cortex located at the anterior sulcus, which responds to pure tones and complex sounds, as demonstrated for P. parnellii (Kanwal et al., 2000; Kobler et al., 1987), C. perspicillata (Eiermann & Esser, 2000; López-Jury et al., 2019) and T. brasiliensis (Macias et al., 2022). This brain region is reciprocally connected to the AC (Figure 2) (Kobler et al., 1987; Vernes et al., 2022), but also receives direct input from the suprageniculate nucleus in the thalamus, as part of a fast, non-canonical and non-lemniscal auditory pathway in which the IC and AC are bypassed (Casseday et al., 1989; Kobler et al., 1987). As shown for P. parnellii, the FAF also sends direct projections to the deep layers of the SC and therefore targets a critical region in the vocal production network (Kobler et al., 1987), as touched upon above.
During vocal production in C. perspicillata, oscillatory LFP activity in the FAF can be highly predictive of the type of vocalization the animal is about to emit: oscillatory power in the delta range [1-4 Hz] and in the gamma range [30-120 Hz] is higher before the onset of an echolocation call, than before the onset of a communication call (Figure 4, García-Rosales et al., 2022; Weineck et al., 2020). Additionally, the FAF also synchronizes its activity with other cortical regions in a call-type dependent manner. Simultaneous neuronal recordings in FAF and AC could show that up until the onset of any vocalization, gamma and delta rhythms in the FAF are predictive of oscillations in the AC, indicative of a top-down information flow from FAF to AC. But at the moment an echolocation call is emitted, this directionality reverses as now delta oscillations in the AC lead those oscillations in the FAF. This reversal does not, however, take place after a communication call is produced, in which case delta waves in the FAF continue to predict AC activity (García-Rosales et al., 2022). These results are highly suggestive that auditory and frontal areas operate in concert to direct vocal production and auditory feedback. After an echolocation call, the FAF requires input from the AC, which may transmit critical auditory information on echo delays (Hechavarría et al., 2013; Hechavarría & Kössl, 2014) among other sensory cues.
Together, these studies indicate frontal cortical areas are candidates for executive vocal control, as neural populations in these regions presage the structure and type of vocalizations before they are uttered. But despite the number of experiments that have been performed over recent years, a comparison across frontal cortical regions and across bat species is difficult because the terminology and the homology to brain regions in other mammalian species, and even between bat species, has not been consistently defined. The first study that described the FAF also identified a direct projection coming from the mediodorsal nucleus of the thalamus (Kobler et al., 1987). This projection is often used as a criterion, although not undisputed, to define the prefrontal cortex in rodents and primates (Fuster, 2015; Rose & Woolsey, 1949; Uylings et al., 2003). The prefrontal cortex is believed to select and guide adaptive behavior based on the current context (Dalley et al., 2004; Euston et al., 2013; Fuster, 2015), a function which could extend to aspects of vocal control in bats. But to what extent the FAF is part of a potential bat prefrontal cortex and what other cortical areas can be included in this definition, has not yet been explored. Relatedly, it is unclear whether previous studies investigating, nominally, the FAF and the ACC were in fact studying entirely distinct or overlapping regions.
A similar ambiguity surrounds the anatomical delimitation of the bat motor and premotor cortices. Some progress has been made in R. aegyptiacus, where motor functions were mapped in motor and somatosensory cortices through electrical microstimulation (Halley et al., 2022). However, this mapping may not easily generalize to other bat species which employ laryngeal echolocation, as R. aegyptiacus performs tongue-based echolocation which does not engage the larynx.
Concluding Remarks
I. Are echolocation and Communication Calls Controlled Through the Same Vocal Pathways?
In this review, we have aimed to bring together diverse reports illustrating how neural activity in bats gives rise to vocal production behavior (for an overview, see Figure 3 and Supplementary Table 1). Two major points come into relief when considering this collection of findings as a whole. First, the general blueprint for motor preparatory and vocal production activity is generally conserved across taxa. The “usual suspects” have emerged as key nodes in the motor output system (premotor brainstem nuclei, the midbrain PAG, lemniscal and collicular structures, basal ganglia, and auditory and frontal cortices) both when we compare species within the bat clade and when looking beyond to other vocal species of primates (including humans) and rodents.
However, quite distinct from most other mammals, bats not only use their vocalization for social interactions, but also to navigate through and perceive their environment. The presented evidence makes it clear how intimately vocal production and sensory processing are intertwined. In possibly all vertebrates that generate sound, and even in invertebrates such as crickets (Poulet & Hedwig, 2002), neural mechanisms have evolved to monitor and inhibit responses to self-generated sound. In the bat, this issue is elevated to a matter of central importance, since echoes may often return while the bat is still producing an echolocation call. Additionally, information gleaned from these echoes is needed to rapidly and dynamically adjust the following echolocation, all on extremely short timescales.
In addition to these challenges, the brain must support both modes of vocalization, echolocation as well as communication. How this is achieved is still one of the pressing questions in this field. Since echolocation is a means of navigation, and communication calls are used to mediate social relationships, one possibility is that these vocal regimes engage at least partially distinct sets of neural populations. In this review, we have explored several brain regions that seem to be selective for echolocation, at least according to our current knowledge. Among them are the SC and some pontine nuclei, such as the PLA. Other areas, including the PAG, the amygdala and the ACC, show evidence of some regional segregation for echolocation and communication circuits (for a review see Fenzl & Schuller, 2007). But also within the same region, echolocation and communication calls may be represented and controlled in overlapping neuronal populations through distinct activity and connectivity patterns. This may make it possible to dynamically fine-tune and adapt vocalizations according to the current context and behavioral state. One prime candidate for such a hub is the frontal cortex, which in bats has been investigated under the designations FAF and motor cortex. The dense interaction network of the frontal cortex with multiple other cortical and subcortical regions puts it in an ideal position for voluntary control of vocalization through integration of sensory feedback and modulation of motor patterns, selecting an adequate behavioral response for the current context. It is worth noting here that echolocation calls may additionally serve a social purpose and communicate personal information about the emitter (Kohles et al., 2020), making the context and purpose of a vocalization more fluent than originally thought.
Currently, we can only hypothesize about how the system evolved to support both the production of echolocation and communication calls. Although not conclusive, most current evidence suggests that laryngeal echolocation evolved once in a species basal to all bats approximately 85 to 65 million years ago (Liu et al., 2022; Teeling et al., 2012; Wang et al., 2017), and it is possible that this common ancestor already possessed a well developed vocal circuit for communication. Neural networks for echolocation may have evolved to use already existing vocal pathways, and over time, some additional brain regions could have been recruited to address the specialized processing requirements of echolocation calls. Alternatively, communication circuits could have evolved in parallel with echolocation, driven by the requirements of social interaction in large group settings. Comparing vocal production circuits in bats and other close mammalian relatives could give insight into this question.
II. Limitations and Future Directions
What has become evident up until this point, is that the subject which has captured the attention of most researchers in studying the bat brain is the echolocation behavior. While this is indeed the most salient and unusual feature of these flying mammals, considerably less attention has been paid to the neural basis of social communication. With the exception of a handful of studies (such as Fenzl & Schuller, 2005; García-Rosales et al., 2022; Ma & Kanwal, 2014; Weineck et al., 2020), few have undertaken a systematic investigation into how vocal production circuits underlying echolocation and communication differ at the neuronal or circuit level. We acknowledge however that it is challenging to measure reliable neural activity during intraspecific communication in naturalistic settings, though it has been achieved in the larger Rousettus bats, and even more so to compare it to such a different behavior as biosonar navigation in a compelling manner.
One limitation of electrophysiological recording techniques, the most commonly used in this field, is that the spatial extent of any given measurement is quite small, while complex sensory and motor functions are well-understood to be achieved through the coordinated activity of many areas of the brain at once. We should therefore remain open to the possibility that some activity patterns observed in a particular region may not reflect the contribution of that region itself so much as a signal which is inherited from a different structure, upstream of the region under observation and hidden from the experimenter’s field of view. This cautionary measure may be especially pertinent when studying multifaceted behavior such as the Doppler shift compensation or social communication.
Throughout the review, we have highlighted the many bat species in which vocalizations and associated neuronal activity patterns have been investigated. These species stem from multiple families and often differ largely in their habitat, guild and social behavior, which is evidenced by their diversity in the design of echolocation calls adapted according to their ecological needs. In contrast to the more common approach in neuroscience that focuses on one or two species as model organisms for an entire clade, this diversity in bat research opens the door to investigations and comparisons of neuronal networks with regard to their evolution, their ecological niche, and how one affects the other. To date, only a handful of studies on vocal production are similar enough to allow for such a comparison across species. But from research in the auditory pathway we know that neuronal organization in the brain can differ greatly depending on the complex interplay of the species' behavioral adaptation and its position on the evolutionary tree. Examples of this, as discussed above, are the ‘auditory fovea’ of CF bats and the chronotopic organization of delay-tuned neurons in the AC (Kössl et al., 2014). As not only echolocation but also communication calls vary greatly across species, we can expect a similar diversity in neural circuits specialized for vocal production. Identifying these differences and mapping them to a species’ adaptation and phylogenetic origin can play a crucial part in understanding the evolution of neuronal mechanisms that underlie the diverse behaviors across animals.
Glossary
GABAergic neurons - Neurons that release gamma-aminobutyric acid (GABA), which acts as an inhibitory neurotransmitter in the mature mammalian brain. By inhibiting excitatory neurons in the brain, they can gate signal flow and contribute to rhythmic brain oscillations.
Dopaminergic neurons - Neurons releasing the neurotransmitter dopamine. In the central nervous system, they are mostly located in the substantia nigra and the ventral tegmental area and play a critical role in movement control, reward and motivation.
LFP - Local field potential, measures the aggregate extracellular potentials of populations of neurons and can be used to measure rhythmic activity in a brain region, such as gamma or theta oscillations (named after their oscillatory frequency range).
Coherence - A measure of synchrony that relies on the phase relationship and consistency of oscillations.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
SSB and JCH conceived the central focus of the review. SSB and AK reviewed the literature and wrote the manuscript. SSB, AK, JCH and FGR revised and edited the manuscript.
Acknowledgements
This work was funded by the DFG priority program, project numbers 520617944 and 520223571, and by the DFG Heisenberg program, project number 525183217. We thank Jagmeet Kanwal for providing examples of Pteronotus parnellii vocalizations, and Michael Yartsev for providing an example of cortical recordings during vocalizations. We also wish to thank Adele Costalunga for graphical support.
References
- Ackermann, H., Mathiak, K., & Riecker, A. (2007). The contribution of the cerebellum to speech production and speech perception: clinical and functional imaging data. Cerebellum (London, England), 6(3), 202–213. [CrossRef]
- Adhikari, A., Lerner, T. N., Finkelstein, J., Pak, S., Jennings, J. H., Davidson, T. J., Ferenczi, E., Gunaydin, L. A., Mirzabekov, J. J., Ye, L., Kim, S.-Y., Lei, A., & Deisseroth, K. (2015). Basomedial amygdala mediates top-down control of anxiety and fear. Nature, 527(7577), 179–185. [CrossRef]
- Aitken, P. G. (1981). Cortical control of conditioned and spontaneous vocal behavior in rhesus monkeys. Brain and Language, 13(1), 171–184. [CrossRef]
- Beetz, M. J., Hechavarría, J. C., & Kössl, M. (2016). Temporal tuning in the bat auditory cortex is sharper when studied with natural echolocation sequences. Scientific Reports, 6, 29102. [CrossRef]
- Bruns, V., & Schmieszek, E. (1980). Cochlear innervation in the greater horseshoe bat: demonstration of an acoustic fovea. Hearing Research, 3(1), 27–43. [CrossRef]
- Carter, G. G., Fenton, M. B., & Faure, P. A. (2009). White-winged vampire bats (Diaemus youngi) exchange contact calls. Canadian Journal of Zoology, 87(7), 604–608. [CrossRef]
- Carter, R. T. (2020). Reinforcement of the larynx and trachea in echolocating and non-echolocating bats. Journal of Anatomy, 237(3), 495–503. [CrossRef]
- Casseday, J. H., Kobler, J. B., Isbey, S. F., & Covey, E. (1989). Central acoustic tract in an echolocating bat: an extralemniscal auditory pathway to the thalamus. The Journal of Comparative Neurology, 287(2), 247–259. [CrossRef]
- Chamberlin, N. L. (2004). Functional organization of the parabrachial complex and intertrigeminal region in the control of breathing. Respiratory Physiology & Neurobiology, 143(2-3), 115–125. [CrossRef]
- Crapse, T. B., & Sommer, M. A. (2008). Corollary discharge across the animal kingdom. Nature Reviews. Neuroscience, 9(8), 587–600. [CrossRef]
- Dalley, J. W., Cardinal, R. N., & Robbins, T. W. (2004). Prefrontal executive and cognitive functions in rodents: neural and neurochemical substrates. Neuroscience and Biobehavioral Reviews, 28(7), 771–784. [CrossRef]
- Dear, S. P., & Suga, N. (1995). Delay-tuned neurons in the midbrain of the big brown bat. Journal of Neurophysiology, 73(3), 1084–1100. [CrossRef]
- Denzinger, A., Kalko, E. K. V., Tschapka, M., Grinnell, A. D., & Schnitzler, H.-U. (2016). Guild Structure and Niche Differentiation in Echolocating Bats. In Bat Bioacoustics (pp. 141–166). Springer New York. [CrossRef]
- Diebold, C. A., Lawlor, J., Allen, K., Capshaw, G., Humphrey, M. G., Cintron-De Leon, D., Kuchibhotla, K. V., & Moss, C. F. (2024). Rapid sensorimotor adaptation to auditory midbrain silencing in free-flying bats. Current Biology: CB. (21). [CrossRef]
- Eiermann, A., & Esser, K. (2000). Auditory responses from the frontal cortex in the short-tailed fruit bat Carollia perspicillata. Neuroreport, 11(2), 421–425. [CrossRef]
- Ekstrom, A. D., Kahana, M. J., Caplan, J. B., Fields, T. A., Isham, E. A., Newman, E. L., & Fried, I. (2003). Cellular networks underlying human spatial navigation. Nature, 425(6954), 184–187. [CrossRef]
- Elemans, C. P. H., Mead, A. F., Jakobsen, L., & Ratcliffe, J. M. (2011). Superfast muscles set maximum call rate in echolocating bats. Science, 333(6051), 1885–1888. [CrossRef]
- Elie, J. E., Muroy, S. E., Genzel, D., Na, T., Beyer, L. A., Swiderski, D. L., Raphael, Y., & Yartsev, M. M. (2024). Role of auditory feedback for vocal production learning in the Egyptian fruit bat. Current Biology: CB, 34(17), 4062–4070.e7. [CrossRef]
- Esser, K.-H., & Schubert, J. (1998). Vocal Dialects in the Lesser Spear-Nosed Bat Phyllostomus discolor. The Science of Nature, 85(7), 347–349. [CrossRef]
- Euston, D. R., Gruber, A. J., & Mcnaughton, B. L. (2013). The role of medial prefrontal cortex in memory and decision making. Neuron, 76(6), 1057–1070. [CrossRef]
- Fenzl, T., & Schuller, G. (2002). Periaqueductal gray and the region of the paralemniscal area have different functions in the control of vocalization in the neotropical bat, Phyllostomus discolor. The European Journal of Neuroscience, 16(10), 1974–1986. (9). [CrossRef]
- Fenzl, T., & Schuller, G. (2005). Echolocation calls and communication calls are controlled differentially in the brainstem of the bat Phyllostomus discolor. BMC Biology, 3, 17. (1). [CrossRef]
- Fenzl, T., & Schuller, G. (2007). Dissimilarities in the vocal control over communication and echolocation calls in bats. Behavioural Brain Research, 182(2), 173–179. [CrossRef]
- Fernandez, A. A., Burchardt, L. S., Nagy, M., & Knörnschild, M. (2021). Babbling in a vocal learning bat resembles human infant babbling. Science, 373(6557), 923–926. [CrossRef]
- Fernandez, A. A., & Knörnschild, M. (2020). Pup Directed Vocalizations of Adult Females and Males in a Vocal Learning Bat. Frontiers in Ecology and Evolution, 8. [CrossRef]
- Fillinger, C., Yalcin, I., Barrot, M., & Veinante, P. (2017). Afferents to anterior cingulate areas 24a and 24b and midcingulate areas 24a’ and 24b' in the mouse. Brain Structure & Function, 222(3), 1509–1532. [CrossRef]
- Fillinger, C., Yalcin, I., Barrot, M., & Veinante, P. (2018). Efferents of anterior cingulate areas 24a and 24b and midcingulate areas 24a’ and 24b' in the mouse. Brain Structure & Function, 223(4), 1747–1778. [CrossRef]
- Fitzpatrick, D. C., Olsen, J. F., & Suga, N. (1998). Connections among functional areas in the mustached bat auditory cortex. Journal of Comparative Neurology, 391(3), 366–396. [CrossRef]
- Forli, A., & Yartsev, M. M. (2023). Hippocampal representation during collective spatial behaviour in bats. Nature, 621(7980), 796–803. [CrossRef]
- Fries, P. (2009). Neuronal gamma-band synchronization as a fundamental process in cortical computation. Annual Review of Neuroscience, 32(1), 209–224. [CrossRef]
- Fuster, J. M. (2015). The Prefrontal Cortex (Fifth Edit, pp. 1–444). Academic Press. [CrossRef]
- Gadziola, M. A., Shanbhag, S. J., & Wenstrup, J. J. (2016). Two distinct representations of social vocalizations in the basolateral amygdala. Journal of Neurophysiology, 115(2), 868–886. [CrossRef]
- Gaioni, S. J., Riquimaroux, H., & Suga, N. (1990). Biosonar behavior of mustached bats swung on a pendulum prior to cortical ablation. Journal of Neurophysiology, 64(6), 1801–1817. [CrossRef]
- García-Rosales, F., López-Jury, L., González-Palomares, E., Wetekam, J., Cabral-Calderín, Y., Kiai, A., Kössl, M., & Hechavarría, J. C. (2022). Echolocation-related reversal of information flow in a cortical vocalization network. Nature Communications, 13(1), 3642. (13). [CrossRef]
- Gemba, H., Miki, N., & Sasaki, K. (1995). Cortical field potentials preceding vocalization and influences of cerebellar hemispherectomy upon them in monkeys. Brain Research, 697(1-2), 143–151. [CrossRef]
- Gooler, D. M., & O’Neill, W. E. (1987). Topographic representation of vocal frequency demonstrated by microstimulation of anterior cingulate cortex in the echolocating bat, Pteronotus parnelli parnelli. Journal of Comparative Physiology. A, Sensory, Neural, and Behavioral Physiology, 161(2), 283–294. (16). [CrossRef]
- Hagemann, C., Esser, K.-H., & Kössl, M. (2010). Chronotopically organized target-distance map in the auditory cortex of the short-tailed fruit bat. Journal of Neurophysiology, 103(1), 322–333. [CrossRef]
- Hage, S. R. (2010). Neuronal networks involved in the generation of vocalization. In S. M. Brudzynski (Ed.), Handbook of Behavioral Neuroscience (Vol. 19, pp. 339–349). Elsevier. [CrossRef]
- Håkansson, J., Mikkelsen, C., Jakobsen, L., & Elemans, C. P. H. (2022). Bats expand their vocal range by recruiting different laryngeal structures for echolocation and social communication. PLoS Biology, 20(11), e3001881. [CrossRef]
- Halley, A. C., Baldwin, M. K. L., Cooke, D. F., Englund, M., Pineda, C. R., Schmid, T., Yartsev, M. M., & Krubitzer, L. (2022). Coevolution of motor cortex and behavioral specializations associated with flight and echolocation in bats. Current Biology: CB, 32(13), 2935–2941.e3. [CrossRef]
- Hariharan, S., Palomares, E. G., Babl, S. S., López-Jury, L., & Hechavarria, J. C. (2024). Cerebellar activity predicts vocalization in fruit bats. Current Biology: CB, 0(0). (11). [CrossRef]
- Hartridge, H. (1920). The avoidance of objects by bats in their flight. The Journal of Physiology, 54(1-2), 54–57. [CrossRef]
- Hasselmo, M. E. (2005). What is the function of hippocampal theta rhythm? - Linking bahavioral data to phasic properties of field potential and unit recording data. Hippocampus, 15(7), 936–949. [CrossRef]
- Hechavarría, J. C., & Kössl, M. (2014). Footprints of inhibition in the response of cortical delay-tuned neurons of bats. Journal of Neurophysiology, 111(8), 1703–1716. [CrossRef]
- Hechavarría, J. C., Macías, S., Vater, M., Voss, C., Mora, E. C., & Kössl, M. (2013). Blurry topography for precise target-distance computations in the auditory cortex of echolocating bats. Nature Communications, 4(1), 2587. [CrossRef]
- Hiryu, S., Mora, E. C., & Riquimaroux, H. (2016). Behavioral and physiological bases for Doppler shift compensation by echolocating bats. In Bat Bioacoustics (pp. 239–263). Springer New York. [CrossRef]
- Hitti, F. L., & Siegelbaum, S. A. (2014). The hippocampal CA2 region is essential for social memory. Nature, 508(7494), 88–92. [CrossRef]
- Hoffmann, S., Vega-Zuniga, T., Greiter, W., Krabichler, Q., Bley, A., Matthes, M., Zimmer, C., Firzlaff, U., & Luksch, H. (2016). Congruent representation of visual and acoustic space in the superior colliculus of the echolocating bat Phyllostomus discolor. The European Journal of Neuroscience, 44(9), 2685–2697. [CrossRef]
- Hong, W., Kim, D.-W., & Anderson, D. J. (2014). Antagonistic control of social versus repetitive self-grooming behaviors by separable amygdala neuronal subsets. Cell, 158(6), 1348–1361. [CrossRef]
- Horikawa, J., & Suga, N. (1986). Biosonar signals and cerebellar auditory neurons of the mustached bat. Journal of Neurophysiology, 55(6), 1247–1267. (14). [CrossRef]
- Hoy, J. L., Bishop, H. I., & Niell, C. M. (2019). Defined Cell Types in Superior Colliculus Make Distinct Contributions to Prey Capture Behavior in the Mouse. Current Biology: CB, 29(23), 4130–4138.e5. [CrossRef]
- Jahelková, H., Horáček, I., & Bartonička, T. (2008). The advertisement song of Pipistrellus nathusii (Chiroptera, Vespertilionidae): a complex message containing acoustic signatures of individuals. [CrossRef]
- Janak, P. H., & Tye, K. M. (2015). From circuits to behaviour in the amygdala. Nature, 517(7534), 284–292. [CrossRef]
- Jones, G., & Teeling, E. C. (2006). The evolution of echolocation in bats. Trends in Ecology & Evolution, 21(3), 149–156. [CrossRef]
- Jürgens, U. (2002). Neural pathways underlying vocal control. Neuroscience and Biobehavioral Reviews, 26(2), 235–258. [CrossRef]
- Jürgens, U. (2009). The neural control of vocalization in mammals: a review. Journal of Voice: Official Journal of the Voice Foundation, 23(1), 1–10. [CrossRef]
- Kanwal, J. S., Gordon, M., Peng, J. P., & Heinz-Esser, K. (2000). Auditory responses from the frontal cortex in the mustached bat, Pteronotus parnellii. Neuroreport, 11(2), 367–372. [CrossRef]
- Knörnschild, M. (2014). Vocal production learning in bats. Current Opinion in Neurobiology, 28, 80–85. [CrossRef]
- Knörnschild, M., Feifel, M., & Kalko, E. K. V. (2014). Male courtship displays and vocal communication in the polygynous bat Carollia perspicillata. Behaviour, 151(6), 781–798. [CrossRef]
- Knörnschild, M., Nagy, M., Metz, M., Mayer, F., & von Helversen, O. (2010). Complex vocal imitation during ontogeny in a bat. Biology Letters, 6(2), 156–159. [CrossRef]
- Knörnschild, M., Nagy, M., Metz, M., Mayer, F., & von Helversen, O. (2012). Learned vocal group signatures in the polygynous bat Saccopteryx bilineata. Animal Behaviour, 84(4), 761–769. [CrossRef]
- Kobler, J. B., Isbey, S. F., & Casseday, J. H. (1987). Auditory pathways to the frontal cortex of the mustache bat, Pteronotus parnellii. Science (New York, N.Y.), 236(4803), 824–826. [CrossRef]
- Kohles, J. E., Carter, G. G., Page, R. A., & Dechmann, D. K. N. (2020). Socially foraging bats discriminate between group members based on search-phase echolocation calls. Behavioral Ecology: Official Journal of the International Society for Behavioral Ecology. [CrossRef]
- Kössl, M., Hechavarria, J. C., Voss, C., Macias, S., Mora, E. C., & Vater, M. (2014). Neural maps for target range in the auditory cortex of echolocating bats. Current Opinion in Neurobiology, 24(1), 68–75. [CrossRef]
- Kössl, M., & Vater, M. (1985). The cochlear frequency map of the mustache bat, Pteronotus parnellii. Journal of Comparative Physiology. A, Sensory, Neural, and Behavioral Physiology, 157(5), 687–697. [CrossRef]
- Kössl, M., Voss, C., Mora, E. C., Macias, S., Foeller, E., & Vater, M. (2012). Auditory cortex of newborn bats is prewired for echolocation. Nature Communications, 3(1), 773. [CrossRef]
- Kothari, N. B., Wohlgemuth, M. J., & Moss, C. F. (2018). Dynamic representation of 3D auditory space in the midbrain of the free-flying echolocating bat. eLife, 7. (20). [CrossRef]
- Krauzlis, R. J., Lovejoy, L. P., & Zénon, A. (2013). Superior colliculus and visual spatial attention. Annual Review of Neuroscience, 36(Volume 36, 2013), 165–182. [CrossRef]
- Lattenkamp, E. Z., & Vernes, S. C. (2018). Vocal learning: a language-relevant trait in need of a broad cross-species approach. Current Opinion in Behavioral Sciences, 21, 209–215. [CrossRef]
- Liu, Z., Chen, P., Xu, D.-M., Qi, F.-Y., Guo, Y.-T., Liu, Q., Bai, J., Zhou, X., & Shi, P. (2022). Molecular convergence and transgenic evidence suggest a single origin of laryngeal echolocation in bats. iScience, 25(4), 104114. [CrossRef]
- López-Jury, L., Mannel, A., García-Rosales, F., & Hechavarria, J. C. (2019). Modified synaptic dynamics predict neural activity patterns in an auditory field within the frontal cortex. The European Journal of Neuroscience, 51(4), 1011–1025. [CrossRef]
- Macias, S., Bakshi, K., Troyer, T., & Smotherman, M. (2022). The prefrontal cortex of the Mexican free-tailed bat is more selective to communication calls than primary auditory cortex. Journal of Neurophysiology, 128(3), 634–648. [CrossRef]
- Macías, S., Luo, J., & Moss, C. F. (2018). Natural echolocation sequences evoke echo-delay selectivity in the auditory midbrain of the FM bat, Eptesicus fuscus. Journal of Neurophysiology, 120(3), 1323–1339. [CrossRef]
- Macías, S., Mora, E. C., Hechavarría, J. C., & Kössl, M. (2012). Properties of echo delay-tuning receptive fields in the inferior colliculus of the mustached bat. Hearing Research, 286(1-2), 1–8. [CrossRef]
- Ma, J., & Kanwal, J. S. (2014). Stimulation of the basal and central amygdala in the mustached bat triggers echolocation and agonistic vocalizations within multimodal output. Frontiers in Physiology, 5, 55. (15). [CrossRef]
- Metzner, W. (1989). A possible neuronal basis for Doppler-shift compensation in echo-locating horseshoe bats. Nature, 341(6242), 529–532. [CrossRef]
- Metzner, W. (1993). An audio-vocal interface in echolocating horseshoe bats. The Journal of Neuroscience: The Official Journal of the Society for Neuroscience, 13(5), 1899–1915. (4). [CrossRef]
- Monsen, R. B., Engebretson, A. M., & Vemula, N. R. (1979). Some effects of deafness on the generation of voice. The Journal of the Acoustical Society of America, 66(6), 1680–1690. [CrossRef]
- Moss, C. F., Bohn, K., Gilkenson, H., & Surlykke, A. (2006). Active listening for spatial orientation in a complex auditory scene. PLoS Biology, 4(4), e79. [CrossRef]
- Moss, C. F., & Surlykke, A. (2001). Auditory scene analysis by echolocation in bats. The Journal of the Acoustical Society of America, 110(4), 2207–2226. [CrossRef]
- Naumann, R. T., & Kanwal, J. S. (2011). Basolateral amygdala responds robustly to social calls: spiking characteristics of single unit activity. Journal of Neurophysiology, 105(5), 2389–2404. [CrossRef]
- Neuweiler, G. (1980). Auditory processing of echoes: Peripheral processing. In Animal Sonar Systems (pp. 519–548). Springer US. [CrossRef]
- Nevue, A. A., Mello, C. V., & Portfors, C. V. (2023). Bats possess the anatomical substrate for a laryngeal motor cortex. bioRxiv.org: The Preprint Server for Biology. [CrossRef]
- Nieder, A., & Mooney, R. (2020). The neurobiology of innate, volitional and learned vocalizations in mammals and birds. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences, 375(1789), 20190054. [CrossRef]
- O’Keefe, J., & Dostrovsky, J. (1971). The hippocampus as a spatial map. Preliminary evidence from unit activity in the freely-moving rat. Brain Research, 34, 171–175. https://www.ncbi.nlm.nih.gov/pubmed/5124915.
- Okobi, D. E., Jr, Banerjee, A., Matheson, A. M. M., Phelps, S. M., & Long, M. A. (2019). Motor cortical control of vocal interaction in neotropical singing mice. Science (New York, N.Y.), 363(6430), 983–988. [CrossRef]
- Oliva, A., Fernández-Ruiz, A., Leroy, F., & Siegelbaum, S. A. (2020). Hippocampal CA2 sharp-wave ripples reactivate and promote social memory. Nature, 587(7833), 264–269. [CrossRef]
- Omer, D. B., Maimon, S. R., Las, L., & Ulanovsky, N. (2018). Social place-cells in the bat hippocampus. Science (New York, N.Y.), 359(6372), 218–224. [CrossRef]
- O’Neill, W. E. (1995). The Bat Auditory Cortex. In Hearing by Bats (pp. 416–480). Springer New York. [CrossRef]
- O’Neill, W. E., & Suga, N. (1979). Target range-sensitive neurons in the auditory cortex of the mustache bat. Science (New York, N.Y.), 203(4375), 69–73. [CrossRef]
- Ono, T., Nakamura, K., Nishijo, H., & Eifuku, S. (1993). Monkey hippocampal neurons related to spatial and nonspatial functions. Journal of Neurophysiology, 70(4), 1516–1529. [CrossRef]
- Payne, H. L., Lynch, G. F., & Aronov, D. (2021). Neural representations of space in the hippocampus of a food-caching bird. Science, 373(6552), 343–348. [CrossRef]
- Poulet, J. F. A., & Hedwig, B. (2002). A corollary discharge maintains auditory sensitivity during sound production. Nature, 418(6900), 872–876. [CrossRef]
- Prat, Y., Taub, M., Pratt, E., & Yovel, Y. (2017). An annotated dataset of Egyptian fruit bat vocalizations across varying contexts and during vocal ontogeny. Figshare. [CrossRef]
- Rodenas-Cuadrado, P. M., Mengede, J., Baas, L., Devanna, P., Schmid, T. A., Yartsev, M., Firzlaff, U., & Vernes, S. C. (2017). Mapping the distribution of language related genes FoxP1, FoxP2, and CntnaP2 in the brains of vocal learning bat species. The Journal of Comparative Neurology, 526(8), 1235–1266. [CrossRef]
- Rolls, E. T., & O’Mara, S. M. (1995). View-responsive neurons in the primate hippocampal complex. Hippocampus, 5(5), 409–424. [CrossRef]
- Rose, J. E., & Woolsey, C. N. (1949). Organization of the mammalian thalamus and its relationships to the cerebral cortex. Electroencephalography and Clinical Neurophysiology, 1(1-4), 391–404. [CrossRef]
- Rose, M. C., Styr, B., Schmid, T. A., Elie, J. E., & Yartsev, M. M. (2021). Cortical representation of group social communication in bats. Science (New York, N.Y.), 374(6566), eaba9584. (1). [CrossRef]
- Rübsamen, R., & Betz, M. (1986). Control of echolocation pulses by neurons of the nucleus ambiguus in the rufous horseshoe bat, Rhinolophus rouxi. Journal of Comparative Physiology. A, Sensory, Neural, and Behavioral Physiology, 159(5), 675–687. (3). [CrossRef]
- Rübsamen, R., Neuweiler, G., & Sripathi, K. (1988). Comparative collicular tonotopy in two bat species adapted to movement detection,Hipposideros speoris andMegaderma lyra. Journal of Comparative Physiology. A, Neuroethology, Sensory, Neural, and Behavioral Physiology, 163(2), 271–285. [CrossRef]
- Salles, A., Loscalzo, E., Montoya, J., Mendoza, R., Boergens, K. M., & Moss, C. F. (2024). Auditory processing of communication calls in interacting bats. iScience, 27(6), 109872. [CrossRef]
- Schnitzler, H.-U. (1968). Die Ultraschall-Ortungslaute der Hufeisen-Fledermäuse (Chiroptera-Rhinolophidae) in verschiedenen Orientierungssituationen. Journal of comparative physiology. A, Neuroethology, sensory, neural, and behavioral physiology, 57(4), 376–408. [CrossRef]
- Schnitzler, H.-U., & Denzinger, A. (2011). Auditory fovea and Doppler shift compensation: adaptations for flutter detection in echolocating bats using CF-FM signals. Journal of Comparative Physiology. A, Neuroethology, Sensory, Neural, and Behavioral Physiology, 197(5), 541–559. [CrossRef]
- Schuller, G., Fischer, S., & Schweizer, H. (1997). Significance of the paralemniscal tegmental area for audio-motor control in the moustached bat,Pteronotus p. Parnellii: The afferent and efferent connections of the paralemniscal area. The European Journal of Neuroscience, 9(2), 342–355. [CrossRef]
- Schuller, G., & Pollak, G. (1979). Disproportionate frequency representation in the inferior colliculus of doppler-compensating Greater Horseshoe bats: Evidence for an acoustic fovea. Journal of Comparative Physiology. A, Neuroethology, Sensory, Neural, and Behavioral Physiology, 132(1), 47–54. [CrossRef]
- Schuller, G., & Radtke-Schuller, S. (1990). Neural control of vocalization in bats: mapping of brainstem areas with electrical microstimulation eliciting species-specific echolocation calls in the rufous horseshoe bat. Experimental Brain Research, 79(1), 192–206. (7). [CrossRef]
- Schwartz, C. P., & Smotherman, M. S. (2011). Mapping vocalization-related immediate early gene expression in echolocating bats. Behavioural Brain Research, 224(2), 358–368. (24). [CrossRef]
- Sinha, S. R., & Moss, C. F. (2007). Vocal premotor activity in the superior colliculus. The Journal of Neuroscience: The Official Journal of the Society for Neuroscience, 27(1), 98–110. (18). [CrossRef]
- Smotherman, M., Knörnschild, M., Smarsh, G., & Bohn, K. (2016). The origins and diversity of bat songs. Journal of Comparative Physiology. A, Neuroethology, Sensory, Neural, and Behavioral Physiology, 202(8), 535–554. [CrossRef]
- Smotherman, M., Kobayasi, K., Ma, J., Zhang, S., & Metzner, W. (2006). A mechanism for vocal-respiratory coupling in the mammalian parabrachial nucleus. The Journal of Neuroscience: The Official Journal of the Society for Neuroscience, 26(18), 4860–4869. (6). [CrossRef]
- Smotherman, M., & Metzner, W. (2003). Fine control of call frequency by horseshoe bats. Journal of Comparative Physiology. A, Neuroethology, Sensory, Neural, and Behavioral Physiology, 189(6), 435–446. (5). [CrossRef]
- Smotherman, M., Zhang, S., & Metzner, W. (2003). A neural basis for auditory feedback control of vocal pitch. The Journal of Neuroscience: The Official Journal of the Society for Neuroscience, 23(4), 1464–1477. [CrossRef]
- Snyder, M. C., Qi, K. K., & Yartsev, M. M. (2024). Neural representation of human experimenters in the bat hippocampus. Nature Neuroscience, 1–5. [CrossRef]
- Suga, N. (1969a). Echo-location and evoked potentials of bats after ablation of inferior colliculus. The Journal of Physiology, 203(3), 707–728. [CrossRef]
- Suga, N. (1969b). Echo-location of bats after ablation of auditory cortex. The Journal of Physiology, 203(3), 729–739. (23). [CrossRef]
- Suga, N., Neuweiler, G., & Müller, J. (1976). Peripheral auditory tuning for fine frequency analysis by the CF-FM bat,Rhinolophus ferrumequinum: IV. Properties of peripheral auditory neurons. Journal of Comparative Physiology. A, Neuroethology, Sensory, Neural, and Behavioral Physiology, 106(1), 111–125. [CrossRef]
- Suga, N., & O’Neill, W. E. (1979). Neural axis representing target range in the auditory cortex of the mustache bat. Science (New York, N.Y.), 206(4416), 351–353. [CrossRef]
- Suga, N., & Schlegel, P. (1972). Neural attenuation of responses to emitted sounds in echolocating bats. Science (New York, N.Y.), 177(4043), 82–84. [CrossRef]
- Suga, N., Schlegel, P., Shimozawa, T., & Simmons, J. (1973). Orientation sounds evoked from echolocating bats by electrical stimulation of the brain. The Journal of the Acoustical Society of America, 54(3), 793–797. (8). [CrossRef]
- Suga, N., & Shimozawa, T. (1974). Site of neural attenuation of responses to self-vocalized sounds in echolocating bats. Science (New York, N.Y.), 183(130), 1211–1213. [CrossRef]
- Suthers, R. A., Thomas, S. P., & Suthers, B. J. (1972). Respiration, wing-beat and ultrasonic pulse emission in an echo-locating bat. The Journal of Experimental Biology, 56(1), 37–48. [CrossRef]
- Tanaka, M., Kunimatsu, J., Suzuki, T. W., Kameda, M., Ohmae, S., Uematsu, A., & Takeya, R. (2021). Roles of the cerebellum in motor preparation and prediction of timing. Neuroscience, 462, 220–234. [CrossRef]
- Teeling, E. C., Dool, S., & Springer, M. S. (2012). Phylogenies, fossils and functional genes: the evolution of echolocation in bats. In G. F. Gunnell & N. B. Simmons (Eds.), Evolutionary History of Bats (pp. 1–22). Cambridge University Press. [CrossRef]
- Tressler, J., Schwartz, C., Wellman, P., Hughes, S., & Smotherman, M. (2011). Regulation of bat echolocation pulse acoustics by striatal dopamine. The Journal of Experimental Biology, 214(Pt 19), 3238–3247. (25). [CrossRef]
- Tschida, K., Michael, V., Takatoh, J., Han, B.-X., Zhao, S., Sakurai, K., Mooney, R., & Wang, F. (2019). A specialized neural circuit gates social vocalizations in the mouse. Neuron, 103(3), 459–472.e4. [CrossRef]
- Ulanovsky, N., & Moss, C. F. (2007). Hippocampal cellular and network activity in freely moving echolocating bats. Nature Neuroscience, 10(2), 224–233. (22). [CrossRef]
- Ulanovsky, N., & Moss, C. F. (2011). Dynamics of hippocampal spatial representation in echolocating bats. Hippocampus, 21(2), 150–161. [CrossRef]
- Uylings, H. B. M., Groenewegen, H. J., & Kolb, B. (2003). Do rats have a prefrontal cortex? Behavioural Brain Research, 146(1-2), 3–17. [CrossRef]
- Valentine, D. E., & Moss, C. F. (1997). Spatially selective auditory responses in the superior colliculus of the echolocating bat. The Journal of Neuroscience: The Official Journal of the Society for Neuroscience, 17(5), 1720–1733. [CrossRef]
- Valentine, D. E., Sinha, S. R., & Moss, C. F. (2002). Orienting responses and vocalizations produced by microstimulation in the superior colliculus of the echolocating bat, Eptesicus fuscus. Journal of Comparative Physiology. A, Neuroethology, Sensory, Neural, and Behavioral Physiology, 188(2), 89–108. (17). [CrossRef]
- van Tussenbroek, I. A., Knörnschild, M., Nagy, M., ten Cate, C. J., & Vernes, S. C. (2024). Morphological diversity in the brains of 12 neotropical bat species. Acta Chiropterologica, 25(2). [CrossRef]
- Vernes, S. C., Devanna, P., Hörpel, S. G., Alvarez van Tussenbroek, I., Firzlaff, U., Hagoort, P., Hiller, M., Hoeksema, N., Hughes, G. M., Lavrichenko, K., Mengede, J., Morales, A. E., & Wiesmann, M. (2022). The pale spear-nosed bat: A neuromolecular and transgenic model for vocal learning. Annals of the New York Academy of Sciences, 1517(1), 125–142. [CrossRef]
- Vernes, S. C., & Wilkinson, G. S. (2019). Behaviour, biology and evolution of vocal learning in bats. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences, 375(1789), 20190061. [CrossRef]
- Wang, Z., Zhu, T., Xue, H., Fang, N., Zhang, J., Zhang, L., Pang, J., Teeling, E. C., & Zhang, S. (2017). Prenatal development supports a single origin of laryngeal echolocation in bats. Nature Ecology & Evolution, 1(2), 21. [CrossRef]
- Weineck, K., García-Rosales, F., & Hechavarría, J. C. (2020). Neural oscillations in the fronto-striatal network predict vocal output in bats. PLoS Biology, 18(3), e3000658. (12). [CrossRef]
- Wirthlin, M. E., Schmid, T. A., Elie, J. E., Zhang, X., Kowalczyk, A., Redlich, R., Shvareva, V. A., Rakuljic, A., Ji, M. B., Bhat, N. S., Kaplow, I. M., Schäffer, D. E., Lawler, A. J., Wang, A. Z., Phan, B. N., Annaldasula, S., Brown, A. R., Lu, T., Lim, B. K., … Zhang, X. (2024). Vocal learning-associated convergent evolution in mammalian proteins and regulatory elements. Science, 383(6690), eabn3263. (2). [CrossRef]
- Wohlgemuth, M. J., Kothari, N. B., & Moss, C. F. (2018). Functional organization and dynamic activity in the superior colliculus of the echolocating bat, Eptesicus fuscus. The Journal of Neuroscience: The Official Journal of the Society for Neuroscience, 38(1), 245–256. (19). [CrossRef]
- Wohlgemuth, M. J., Yu, C., & Moss, C. F. (2018). 3D hippocampal place field dynamics in free-flying echolocating bats. Frontiers in Cellular Neuroscience, 12, 270. [CrossRef]
- Yartsev, M. M., Witter, M. P., & Ulanovsky, N. (2011). Grid cells without theta oscillations in the entorhinal cortex of bats. Nature, 479(7371), 103–107. [CrossRef]
- Yin, J.-X., Ruan, Y.-N., Liu, J.-L., Zhang, S.-Y., & Racey, P. (2017). FoxP2 expression in an echolocating bat (Rhinolophus ferrumequinum): Functional implications. Zeitschrift Für Saugetierkunde [Mammalian Biology], 85(1), 24–29. [CrossRef]
Figure 1.
Examples of echolocation and communication calls of three bat species from different families. (A-B) Pteronotus parnellii. (A) Oscillogram (top) and spectrogram (bottom) of an echolocation call with CF and FM components and (B) a selection of communication calls, namely sHFM (left), dRFM (middle) and rBNB (right). Examples reproduced with permission from Jagmeet Kanwal, Georgetown University. (C-D) Rousettus aegyptiacus. (C) Oscillogram (top) and spectrogram (bottom) of echolocation pulse produced with tongue click and (D) a selection of communication calls. Examples reproduced from publicly available dataset provided by Prat et al. (2017). (E-F) Carollia perspicillata. (E) Oscillogram (top) and spectrogram (bottom) of echolocation call with FM component and (F) a selection of communication calls recorded from the laboratory-housed colony. .
Figure 1.
Examples of echolocation and communication calls of three bat species from different families. (A-B) Pteronotus parnellii. (A) Oscillogram (top) and spectrogram (bottom) of an echolocation call with CF and FM components and (B) a selection of communication calls, namely sHFM (left), dRFM (middle) and rBNB (right). Examples reproduced with permission from Jagmeet Kanwal, Georgetown University. (C-D) Rousettus aegyptiacus. (C) Oscillogram (top) and spectrogram (bottom) of echolocation pulse produced with tongue click and (D) a selection of communication calls. Examples reproduced from publicly available dataset provided by Prat et al. (2017). (E-F) Carollia perspicillata. (E) Oscillogram (top) and spectrogram (bottom) of echolocation call with FM component and (F) a selection of communication calls recorded from the laboratory-housed colony. .
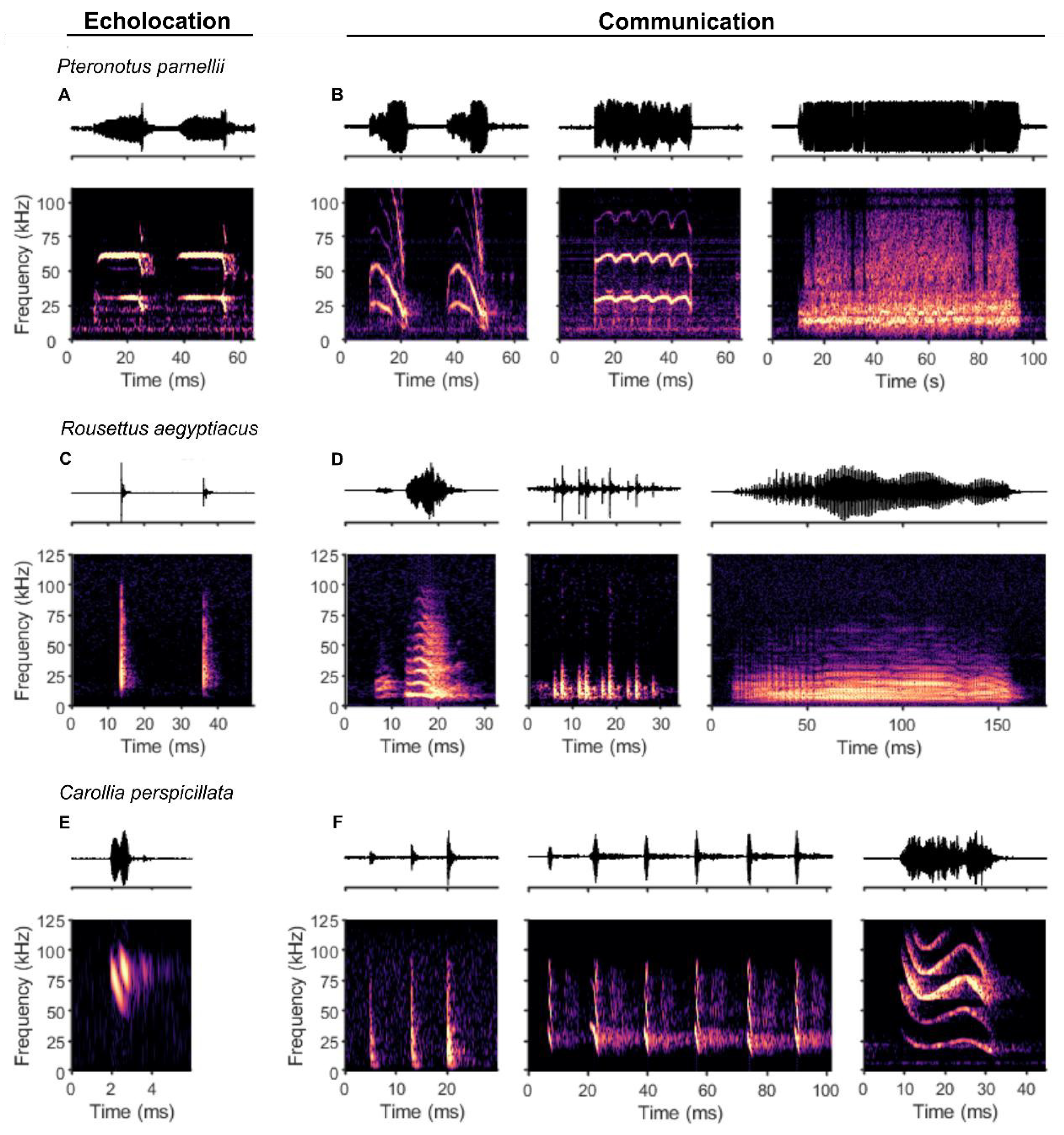
Figure 2.
Schematic illustration of important brain structures in vocal production and their anatomical connections. Solid lines indicate direct projections, dashed lines indicate indirect connections via further brain regions. Abbreviations: AC - auditory cortex, AMY - amygdala, BG - basal ganglia, CEREB - cerebellum, FC - frontal cortex, IC - inferior colliculus, NA - nucleus ambiguus, PAG - periaqueductal gray, PB - parabrachial complex, PLA - paralemniscal area, RF - reticular formation, SC - superior colliculus.
Figure 2.
Schematic illustration of important brain structures in vocal production and their anatomical connections. Solid lines indicate direct projections, dashed lines indicate indirect connections via further brain regions. Abbreviations: AC - auditory cortex, AMY - amygdala, BG - basal ganglia, CEREB - cerebellum, FC - frontal cortex, IC - inferior colliculus, NA - nucleus ambiguus, PAG - periaqueductal gray, PB - parabrachial complex, PLA - paralemniscal area, RF - reticular formation, SC - superior colliculus.
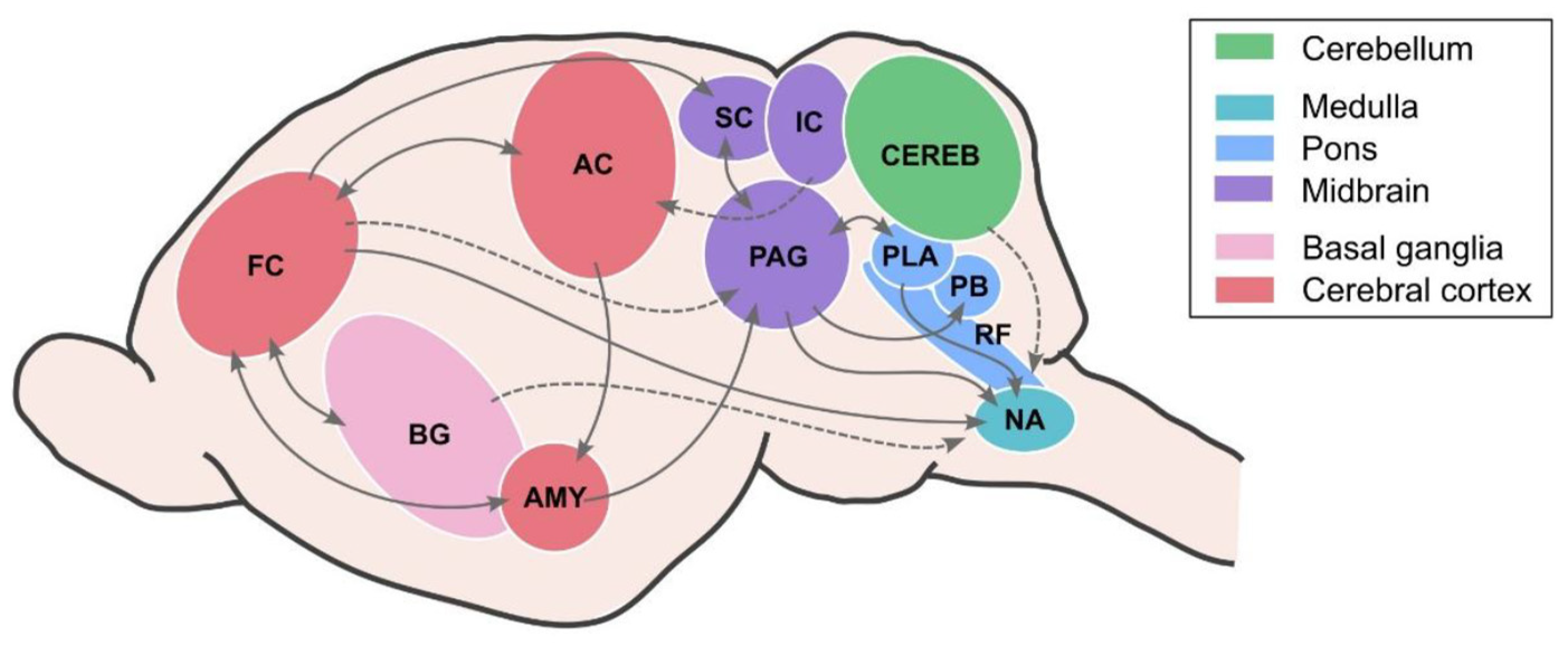
Figure 3.
Research on the neuronal network of vocal production mapped to the bat cladogram, showing the investigated species, the studied brain region and a summary of the main findings for each family. Echolocation and communication calls are abbreviated echo and comm, respectively. Superscript numbers indicate corresponding references and are marked in the reference list.
Figure 3.
Research on the neuronal network of vocal production mapped to the bat cladogram, showing the investigated species, the studied brain region and a summary of the main findings for each family. Echolocation and communication calls are abbreviated echo and comm, respectively. Superscript numbers indicate corresponding references and are marked in the reference list.
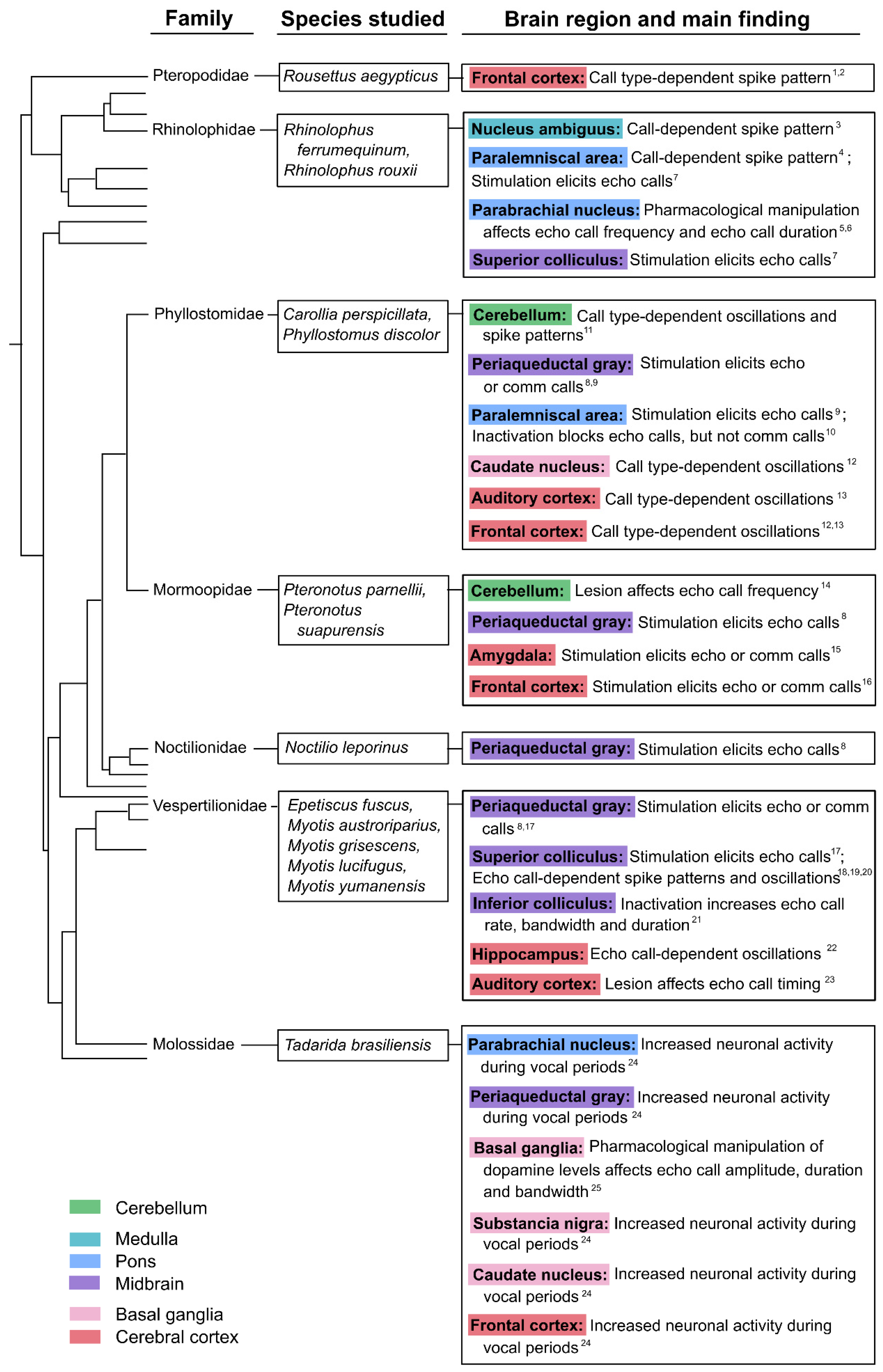
Figure 4.
Neuronal correlates of vocalization in the frontal cortex from two different bat species. (A-D) Exemplary anatomical and electrophysiology data from Rousettus aegyptiacus. (A-B) Location of tetrode recording in the frontal cortex (yellow circle in (A), dashed box and inset in (B)). (C) Single unit activity during self-produced vocalizations (green) or calls of conspecifics (yellow) during group interaction: call-aligned raster plots (top), average firing rate (middle) of two example neurons, and z-scored firing rates (bottom). (D) Average proportions of responsive neurons to different stimuli or vocal events across bats. Figure reproduced from Rose et al. (2021) with permission from AAAS. (E-H) Exemplary anatomical and electrophysiological data from Carollia perspicillata. (E-F) Location of laminar probe recording in the FAF (yellow circle in (E), dashed box and inset in (F)). (G) Exemplary LFP traces aligned to a single echolocation call (blue) and a single social communication call (red) across cortical depth. (H) Mean change in the power spectral density in representative LFP bands during the pre-vocal period (0.5-0 s) of an echolocation (blue) or a communication call (red) compared with baseline (steady state) activity. Figure reproduced with permission from García-Rosales et al. (2022).
Figure 4.
Neuronal correlates of vocalization in the frontal cortex from two different bat species. (A-D) Exemplary anatomical and electrophysiology data from Rousettus aegyptiacus. (A-B) Location of tetrode recording in the frontal cortex (yellow circle in (A), dashed box and inset in (B)). (C) Single unit activity during self-produced vocalizations (green) or calls of conspecifics (yellow) during group interaction: call-aligned raster plots (top), average firing rate (middle) of two example neurons, and z-scored firing rates (bottom). (D) Average proportions of responsive neurons to different stimuli or vocal events across bats. Figure reproduced from Rose et al. (2021) with permission from AAAS. (E-H) Exemplary anatomical and electrophysiological data from Carollia perspicillata. (E-F) Location of laminar probe recording in the FAF (yellow circle in (E), dashed box and inset in (F)). (G) Exemplary LFP traces aligned to a single echolocation call (blue) and a single social communication call (red) across cortical depth. (H) Mean change in the power spectral density in representative LFP bands during the pre-vocal period (0.5-0 s) of an echolocation (blue) or a communication call (red) compared with baseline (steady state) activity. Figure reproduced with permission from García-Rosales et al. (2022).
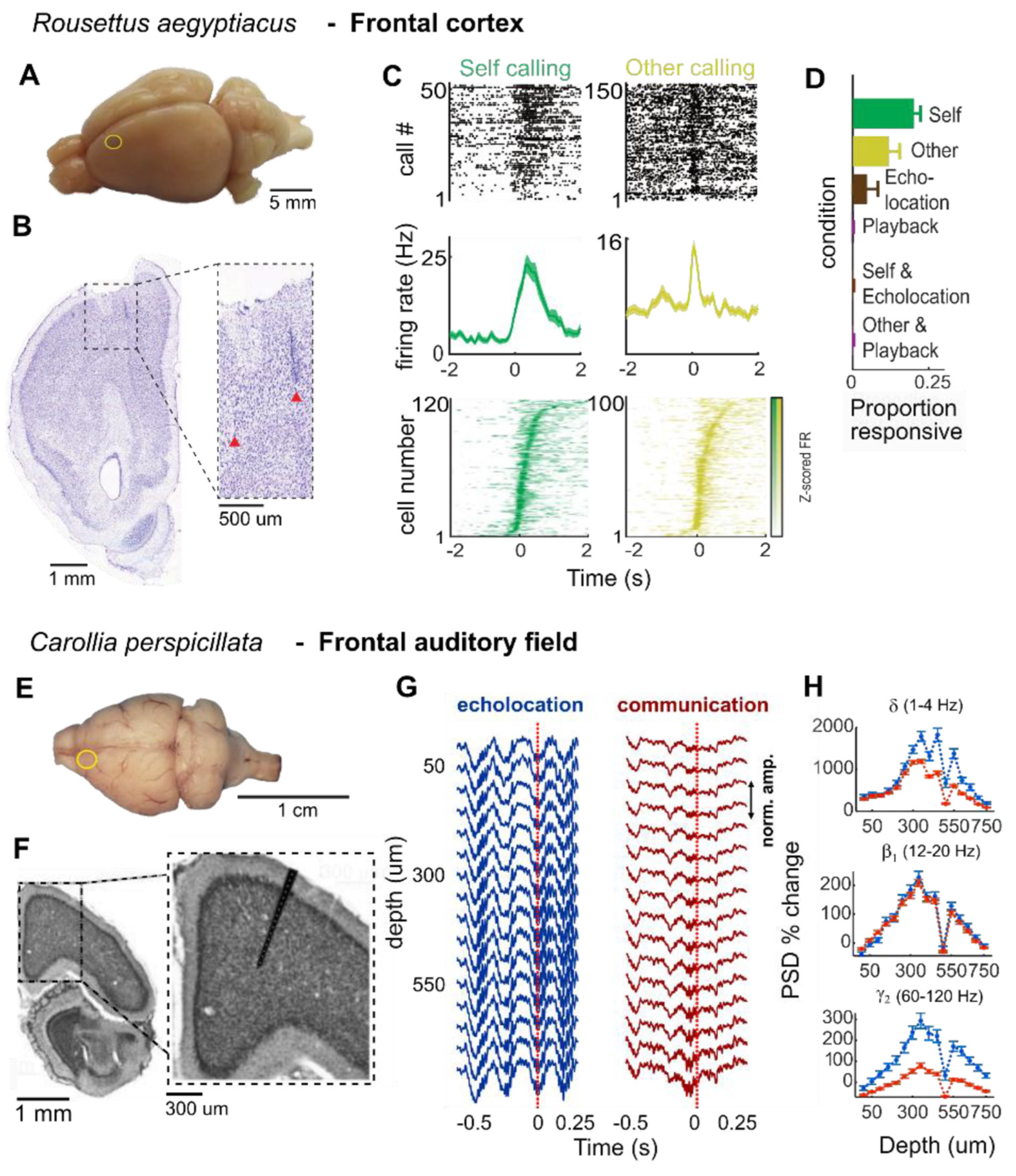
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



