
Submitted:
02 April 2025
Posted:
02 April 2025
You are already at the latest version
Abstract
Helicobacter pylori is a motile bacterial pathogen that causes severe gastric diseases globally. H. pylori motility and chemotaxis are key colonization factors. Motility and chemotaxis are studied in many microbes, including H. pylori, using soft agar assays. In these assays, bacteria are inoculated into low-percentage agar and expand in a motility and chemotaxis-dependent manner. H. pylori similarly expand in soft agar but if a plate is inoculated at multiple points, the expanded H. pylori colonies do not merge and leave gaps. The basis of these gaps was unknown. We report here that gap formation was not affected by media components such as nutrient and agar concentrations, nor did it require chemotaxis but did rely on quorum sensing. To broaden our understanding of this H. pylori property, an H. pylori Tn7 transposon library was screened for mutants that lost gap formation. Fourteen mutants were obtained, and the transposon sites mapped to genes encoding outer membrane proteins, cysteine rich proteins, phosphatidyl glycerophosphate synthase, an endorestriction nuclease and several hypothetical proteins. Our results suggest that H. pylori may use specific proteins to avoid contact with other H. pylori, a behavior that may relate to previous observations that different H. pylori strains do not mix populations in stomach glands.
Keywords:
Helicobacter pylori
; soft agar
; chemotaxis
; motility
; clonal competition
; quorum sensing
; outer membrane proteins
; hypothetical proteins
1. Introduction
Bacterial motility is a widespread trait found in over 50% of bacterial species and has been shown to confer fitness advantages in diverse settings [1]. Bacterial motility is often studied using bacterial media with a low percentage of agar, so-called soft or semi-solid agar plates [2,3]. After inoculation, bacteria migrate outward from the initial point, creating an expanded colony whose formation depends on growth, motility abilities, and chemotaxis responses [2,4,5]. Soft agar migration rates are influenced both by bacterial properties, such as chemotaxis, motility and growth, as well as media physical properties, including agar concentration[6] and nutrient availability [4,5].
In soft agar motility assays, bacteria migrate outwards from the point of inoculation resulting in colony expansion. Often these assays are done with several inoculations per plate, and in this case, the expanding colonies can potentially run into each other. When this happens, bacterial species show varying responses. In some cases, colonies merge or cross over each other, while others produce gaps with low bacterial numbers [5,7]. Our initial studies that frame this work showed that H. pylori colonies form visible gaps in motility assays, so this work focusses on them. Three prominent theories have been proposed to explain soft agar bacterial colony gaps: (1) gaps form because the migrating bacteria deplete nutrients in front of the colony, resulting in a poor ability to grow in the region between the colonies [5]; (2) gaps form because the migrating colonies induce mechanical stress in the agar that in turn creates physical barriers, e.g. thickened agar, that cannot be crossed by a competing colony[6]; and (3) gaps form because the colonies produce inhibitory molecules that arrest the growth of competing colonies [7].
In some bacterial species, soft agar colony gaps are formed only between non-clonal strains. Proteus mirabilus, for example, forms gaps only between non-clonal strains [8]. This response has been attributed to a genetic determinant mechanism to distinguish and recognize non-kin cells [9]. Clonal discrimination behaviors have been identified in various growth conditions including and outside of soft agar for Bacillus subtilis [5,7], Pseudoalteromonas haloplanktis [5], Salmonella typhimurium [5], Escherichia coli [5,7], Paenibacillus dendritiofrmis [7], Pseudomonas putida [6],Vibrio cholerae [10] and H. pylori [11]. Altogether, it’s clear that numerous reasons can underly gap formation in soft agar.
Helicobacter pylori is a motile microbial pathogen that is frequently studied in soft agar motility assays [4,12,13]. H. pylori colonizes the gastric tissues of more than half of the world’s population, causing chronic infections that can lead to gastritis, ulcers and gastric cancers [14,15,16,17]. H. pylori relies on motility and chemotaxis for host colonization [12,18,19]. H. pylori has lophotrichous sheathed flagella that confer strong motility in viscous environments, and a single chemotaxis system comprised of four chemoreceptors [20,21]. It uses flagellar motility to migrate in rich media with soft agar concentrations below 0.6% [22].
At the start of this work, we noticed that H. pylori soft agar colonies form gaps when they encounter each other. Given the myriad reasons for gap formation, we sought to understand the basis in H. pylori. We examined various parameters that might contribute to gap formation including agar concentration, media composition, and bacterial genetic determinants. Our results support that agar and nutrient concentrations influence the rate of H. pylori colony migration, as observed previously [4,22], but these variables did not alter gap formation. By exploring quorum sensing mutants and screening a transposon library, we found that gap formation is influenced by genetic factors. Our combined results suggest that the H. pylori soft agar gaps may be formed based on cell-cell recognition, a new idea that paves the way to understand this phenomenon.
2. Materials and Methods
Bacterial strains and growth conditions
For this work, the following strains were used: H. pylori wild type G27 [23], G27 pTM115 expressing GFP (KO446) [21], 26695 [24], PMSS1 [25], and SS1 [26]. The following mutants were used, all in the G27 or mG27 background: ∆chePep KO1332 [27], ∆cheV2 KO1278[28], ∆cheV3 KO1279[29], ∆tlpA KO740 [30], ∆tlpB KO583[31], ∆tlpC KO568[30], ∆tlpD (KO#1006)[32] and ∆luxS (KO1791). For mutant isolation, a G27-based transposon library was used [33]. H. pylori strains were grown on solid media consisting of Columbia Horse Blood Agar (CHBA) containing 5% (vol/vol) defibrinated horse blood (Hemostat Labs), 0.2% w/v-β cyclodextrin, 10 µg of vancomycin per ml, 5 µg of cefsulodin per ml, 2.5 U of polymyxin B per ml, 5 µg of trimethoprim per ml, 8 µg of amphotericin B per ml (all chemicals from Thermo Fisher or Gold Biotech). For growth in liquid media, Brucella Broth (BB) (Thermo Fisher) plus 10% (vol/vol) heat-inactivated fetal bovine serum (HI-FBS) was used (BB10). For antibiotic resistance marker selection, bacterial media were supplemented with 25µg of chloramphenicol (Cm) per mL, or 15 µg of kanamycin (Km) per mL. For long term storage, strains were frozen at -80°C in brain heart infusion media supplemented with 10% HI-FBS, 1% (wt/vol) β-cyclodextrin, 25% glycerol, and 5% dimethyl sulfoxide. All H. pylori cultures were grown at 37°C under microaerobic conditions of 5% O2, 10% CO2, and 85% N2.
Soft Agar Motility Assays
Soft agar motility assays were conducted with plates that were prepared using Brucella broth (BB) media with 2.5% HI-FBS and solidified with 0.35% (w/v) bacto agar (Fisher Scientific) for standard soft agar plates, or different agar or BB amounts when examining these variables. Soft agar plates were inoculated by touching bacterial growth from CHBA plates using a sterile micropipette tip and inserting into the soft agar. Plates were incubated in microaerobic conditions at 37°C for 6-10 days. Distances between the inoculation point and the colony edges were measured using a ruler. Soft agar plates were visualized using Microtek Scan Maker i900 or Biorad Chemidoc MP Imaging System with White Epi Illumination setting on the UV Transilluminator
Isolation of Tn-7 mutants without clear gap formation phenotype
To isolate mutants that did not form soft agar gaps, a G27 transposon pool of mutants were cultured on CHBA for 3 days in microaerobic conditions. Soft agar plates were inoculated at two points 1 cm apart from each other. After incubating the plates for six days, the expanded colonies formed were partitioned to three zones: the non-competing front, the competing front and the merge region, as shown in Figure QA. Bacteria from each region were collected by poking sterile pipette tips into the agar at that region, serially diluted, and re-plated on Cm supplemented CHBA plates to obtain single colonies. Twenty single colonies were collected from each of the three zones. These individual isolates were then plated again on standard soft agar to test behavior. Isolates that exhibited loss of gap formation were termed “merging isolates” and were stored at -80C for further analysis.
Identification of transposon insertion sites
To identify the location of transposon insertion sites in the merging isolates, genomic DNA was extracted using the Wizard Genomic Prep Kit (Promega). Nested PCR was conducted on the extracted DNA. For the first round of PCR, amplification was carried out with a random primer with a constant tail region plus a transposon-specific primer, called CAT Tn7N and CAT Tn7S listed in Table A1[33]. For the second round of PCR, amplification was carried out with a primer specific to the transposon plus a primer complementary to the constant tail region of the random primer, primers CAT Tn7 N2 and CAT Tn7 S2. Therefore, the final PCR products contain a portion of the transposon and the surrounding genomic information. These PCR products were then sent for DNA sequencing (Azenta) and analyzed by mapping to the G27 genome using NCBI Nucleotide Blast.
Genomic Transformations
To retransform the transposon mutants into a clean background, genomic DNA was isolated from the alpA::Tn-7 using the Wizard Genomic Prep Kit (Promega). H. pylori G27 WT was grown on CHBA for three days and then collected into 200 uL of BB10, such that the OD600 range was between 2-5, following the protocol in Hu et al. 2023. 100ng of genomic DNA was added to the bacterial suspension and incubated at room temperature for 5 minutes. The bacterial suspension was plated on CHBA for 24 hours to allow recovery before being re-suspended in 300uL of brucella and plated on CHBA plates supplemented with chloramphenicol. After 3-5 days of growth, single colonies were harvested, re-colony purified, and then either stored at -80C or used for DNA extraction.
Microscopy
To examine bacteria present in soft agar assays, samples were collected from the gaps by a pipet tip and visualized by phase contrast microscopy at 200X, 400X and 1000X magnification, using a Nikon ECLIPSE E600 microscope with a Hamamatsu C7472-95 digital camera.
Growth Curves
H. pylori strains were cultured overnight (shaking) in BB10 for 12-14 hours. Overnight cultures were back-diluted to OD600 of 0.1 using Brucella Broth supplemented with 2.5% HI-FBS to match the components in the soft agar assays. Growth assays were completed with CLARIOstarPlus Microplate Reader (BMG LABTECH) under shaking, microaerobic conditions of 5% O2, 10% CO2, and 85% N2 at 37°C. Four biological replicates per strain were conducted and OD600 was measured hourly for 20-25 technical replicates for each time point over the course of 24 hours.
Clear Gap Materials Assessment
To assess whether the gaps had growth inhibitory compounds, gap material was extracted from between G27 WT colonies after 8-days of soft agar incubation by inserting a pipet tip and sucking up 200µl of the clear gap from 5-6 positions spanning across the entire length of the competing fronts. Multiple plates were used to accumulate 1.5 mL. To this, 1.5 mL of the strain to be tested at a back diluted OD600 of 0.1 in fresh BB10 was added and incubated in microaerophilic conditions with shaking for 8 hours. Treated strains included G27 GFP+, which is kanamycin resistant. Control cultures were treated with either 1.5 mL of uninoculated agar or with BB10 only. Aliquots from treated cultures were serially diluted and plated on CHBA with either Kan or Cm; this step ensured that only that test strains would grow. CHBA plates were incubated for 3 days and counted for colony forming units.
3. Results
H. pylori ceases soft agar migration and forms a gap when two colonies meet
This project was initially started from the observation that H. pylori on long-incubated soft agar plates migrated from the inoculation points and saturated all the available media but produced a pattern of clear gaps in the soft agar that separated colonies (Figure 1A). This result suggested that colonies from different inoculation points failed to intersect or merge. We next characterized the soft agar behavior in more detail using a wild-type H. pylori G27 strain that expresses GFP from the pTM115 plasmid [21]. H. pylori GFP+ was inoculated on a standard H. pylori soft agar plate at two points, one centimeter apart from each other. The plates were incubated and analyzed over the course of six days, measuring the distances migrated away from (non-competitive) or toward the other colony (competitive). The bacteria displayed little migration at days 1-2, as noted previously [22], but displayed visible outward migration from day 3 on (Figure 1B i-iii). During days 1-2, the distance migrated by the competitive and non-competitive front from the inoculation points were similar (Figure 1C). After 3 days, the two fronts started to show different behavior that was statistically significant, with the competitive bacterial front migrating slower than the non-competitive front (Figure 1C). Over days 3-6, the non-competitive bacterial fronts continued migrating and the colonies developed an asymmetric shape (Figure 1B) and the difference in distances traveled by both fronts became increasing significant by the final day of the assay. We confirmed this behavior in other H. pylori strains including 26695, PMSS1, and SS1, observing that they too formed gaps (not shown). These results show that H. pylori colonies respond to each other even when at a distance and slow their migration as they approach each other.
Formation of colony gaps is chemotaxis independent
Soft agar migration depends on chemotaxis and motility. We therefore explored how bacterial chemotaxis affected gap formation. H. pylori G27 mutants with partial chemotaxis defects (∆chePep, ∆cheV2, ∆cheV3) were used for this analysis because they maintain partial ability to migrate in soft agar [27,28,29]. These strains still formed gaps (Figure 2A). Similarly, mutants with single chemoreceptor mutations (∆tlpA, ∆tlpB, ∆tlpC, ∆tlpD) retain soft agar migration [30,31,32] and still produced gaps (Figure 2B). These results suggest that chemotaxis does not affect gap formation between H. pylori soft agar colonies.
H. pylori colony gap formation is independent of nutrient level and agar percentages
We next explored whether media parameters influenced gap formation. One previously hypothesis reason for gap formation is the nutrient-based explanation, which theorizes that gaps form as nutrients decrease because there are insufficient nutrients to support growth [5]. This theory predicts that lower nutrients would result in increased gap sizes. We therefore varied the concentration of brucella broth in the media. Decreasing the brucella broth below normal (0.5X or 0.75X) or above (1.5x) changed migration rate, as reported before [4,34], but had no effect on gap formation (Figure 3A). These results suggest that nutrient availability does not affect H. pylori gap formation.
Another variable known to affect gap formation are physical properties, such as agar percentage [6]. We therefore modified the percent of agar in the soft agar plates. Gaps were readily apparent at the competitive bacterial fronts at agar concentrations from 0.3% to 0.45% (Figure 3B). At the lowest percent, 0.25%, there was only a minimal gap (Figure 3B). This result suggested that gaps require a minimal agar percentage for either formation or stability. At higher agar percent, the colonies had low migration but still retained gaps (Figure 3B). These results indicate that agar concentration may affect gap stability but does not clearly alter gap formation.
Soft agar colony gaps contain low densities of H. pylori cells with spiral morphology.
The gaps formed between colonies lack the normal opaqueness associated with bacterial growth, so we next examined whether these regions contained bacteria and whether these bacteria were alive. We first attempted to image the bacteria directly in the agar, by pouring thin, soft agar onto microscope slides with the petri dish as done previously [22], but these agar slides were too optically dense to see bacteria. So, samples were instead collected directly from the soft agar in either the gap or colony, and imaged using phase contrast microscopy (Figure 4). Low magnification images showed that there was bacterial density in the gaps, albeit lower than at the non-competitive front in agreement with the macroscopic soft agar plate appearance (Figure 4A, B). At 1000X, cells in the gap could be seen that retained a spiral morphology (Figure 4Aiii). In contrast, cells within the middle of the colony showed non-spiral shaped cells in aggregates (Figure 4Biii). Direct plating of gap material showed H. pylori growth (Figure 4C). These results suggest that the gaps contain a low density of H. pylori cells that are spiral in morphology, suggesting they are potentially viable.
Assessing inhibitory compounds in gap regions.
One possibility is that the gap region contains compounds that inhibit the accumulation of bacteria. This possibility was tested by exposing an experimental strain to material collected from gaps. Specifically, gaps were formed by wild-type H. pylori strain G27 (kanamycin-sensitive) for 8 days, and then material was collected from the gaps by pipetting. This material was then used to treat an H. pylori G27 strain that was kanamycin-resistant (H. pylori G27 pTM115). We then plated the treated samples on kanamycin CHBA, so that only the experimental strain would grow, comparing treated samples to those that were either left untreated or treated with uninoculated agar samples (experimental design shown in Supplemental Figure 1). After treatment, the samples were serially diluted and plated (Figure 5A-C). Treatment with gap material resulted in an ~ 10-fold decrease in CFU counts compared to no treatment (Figure 5D), but control experiments showed that agar alone was slightly inhibitory. When comparing the two agar treatments to each other (± H. pylori) there was lower growth but this decrease did not achieve statistical significance. These results suggest that there may be some inhibitory activity in the gap region, but it is not substantial or toxic.
H. pylori gap formation has a genetic basis.
We next analyzed whether there might be genetic determinants that affected gap formation. One H. pylori property that influences group behavior is quorum sensing via the auto-inducer 2 (AI-2) quorum sensing system [35]. To test whether quorum sensing was involved in gap formation, we analyzed the behavior of H. pylori lacking the AI-2 producing enzyme, luxS. H. pylori ∆luxS mutants did not form gaps (Figure 6B), suggesting gap formation depended on bacterial properties including quorum sensing. To identify other genetic loci that controlled gap formation, we screened an H. pylori G27 mini Tn7 transposon mutant pool [33]. As done above, we incubated two inoculums of this pool one centimeter apart for six days, and samples were taken from the gap region to enrich for mutants that had lost the ability to form gaps. As controls, isolates were also collected from within the colony, at both the non-competing and competing fronts as defined in Figure 1A. These samples were single colony purified and retested for gap formation. Isolates from within the colony, from either the non-competing or competing fronts, retained the ability to form gaps (Figure 6A) but isolates from the gap region lost this ability, creating colonies that merged (Figure 6A). To characterize whether the “merge” (loss-of-gap) phenotype was associated with the Tn7-targetted locus, we isolated genomic DNA from these merge isolates and used this DNA to transform a fresh G27 WT strain to chloramphenicol resistance. These recreated strains also lost gap formation, suggesting the Tn7-mutated locus conferred the merge phenotype (Figure 6B). Analysis of the gap region using microscopy revealed that the merging Tn7 mutants had a high number of bacteria in the gap zone compared with G27 WT (not shown). These results suggested that the transposons had interrupted genes that were required for gap formation.
To identify the mutant loci, 14 single colony Tn7 mutants with the merge phenotype were isolated from the G27 Tn7 pool and nested PCR was used to identify the disrupted genes. The disrupted genes included those coding for the AlpA and AlpB outer membrane proteins, two cysteine rich proteins, the HpyAIV type II restriction enzyme, phosphatidyl glycerophosphate synthase, and four hypothetical proteins (Table 1). ). These results suggest that H. pylori soft agar colony gap formation has a genetic basis, with multiple genes implicated.
To gain insight into the underlying reason the Tn mutants merged, we analyzed various aspects of their behavior These mutants retained merging behavior under varying nutrient and agar concentrations (Supplemental Figure 2). We observed that several of these mutants had elevated migration rates (Supplemental Figure 3). We evaluated their growth rates, but found this was not altered (Supplemental Figure 4). To explore whether fast migration rate could have potentially contributed to the loss of the gaps, e.g. perhaps by allowing the mutant strains to overcome physical resistance of the soft agar, we examined a known mutant with a documented elevated migration rate, H. pylori G27 ∆pilO [13]. This mutant, however, retained a normal ability to produce gaps between colonies, suggesting fast migration may facilitate but is not sufficient to cause merging (Supplemental Figure 3).
4. Discussion
In this work, we present findings that H. pylori migrating colonies do not merge and instead form low-density bacterial regions that appear as gaps, a phenomenon observed in other microbes. The H. pylori gaps are independent of chemotaxis, nutrient levels, or agar concentrations. They also do not have substantial soluble anti-microbial properties. Instead, our data suggest these gaps form as a consequence of H. pylori physiological properties, as evidenced by the finding of H. pylori mutants that lose the ability to form gaps. This work may have implications for the observed behavior of H. pylori in vivo, in which one infection blocks colonization by a second H. pylori strain [11,19,21].
Previous work has shown that soft agar colonies of some bacterial species merge while others form gaps and avoid direct colony-to-colony contact [5]. In some species, the phenomenon of merging or gap formation between competing bacterial colonies has been correlated with nutrient availability [5,7] or agar percentage [5,6]. Studies with S. typhimurium linked “slow down” of migration speed to excess nutrients, that caused a decrease in bacterial front velocity and the creation of gaps, with cells documented to switch to a less motile state [5]. B. subtilis colonies, in contrast, merge under low nutrient conditions, where migration speeds were faster than under nutrient replete conditions [7]. Gap formation in H. pylori, however, is not affected by nutrient amounts. Specifically, elevating or lowering brucella broth concentrations (Figure 3) or manipulating the concentration of FBS (not shown) had no effect on gap formation. In line with this finding, partial chemotaxis mutants still formed gaps in soft agar (Figure 2). Altogether, these results suggest that H. pylori gap formation is largely independent of media composition and chemotactic response.
We also explored whether the gaps between colonies might contain a potential secretion that decreases bacterial density. We collected gap material and treated fresh H. pylori, but found that the gaps slightly decreased bacterial numbers, but this decrease was not statistically significant (Figure 4). This finding suggested that gap formation is not due to cell growth inhibition via secretion of toxic compounds.
To gain ideas for how gap formation occurs in H. pylori, we examined quorum sensing mutants and isolated transposon mutants that loose gap formation. Quorum-sensing defective luxS mutants were the first we observed to lose gap formation (Figure 6). This finding strongly supported the idea that gap formation is influenced by bacterial properties, in this case, production of AI-2. Consistent with this idea, our transposon mutant screen identified 10 transposon insertions that lost gap formation. These disrupted loci mapped to a variety of cellular processes, including outermembrane proteins, cysteine-rich proteins, metabolic proteins, restriction enzymes and hypothetical proteins. All these proteins play important roles in H. pylori biology.
AlpAB are two outer membrane proteins characterized as adhesins that facilitate adherence between H. pylori and host-cell laminin, and also contribute to outer membrane vesicles in H. pylori biofilms [36,37,38,39,40]. The alpA-alpB operon was previously found to play a role in colonization in gerbils [38], mice [36], guinea pigs [41] and human epithelial cells [40,42] and triggers a host immune response [36,43]. AlpA and AlpB contain 518 amino acids, and are highly similar [36], dependent on each other functionally [36] and expressed in similar amounts in 200 clinical H. pylori isolates from human gastric biopsies [37]. Based on the locations of the transposon insertions, the region upstream the alpAB operon and alpA have been disrupted, indicating the merge phenotypes we’ve observed may be due to loss of at least alpA and maybe alpB.
Another transposon disrupted the gene for the type II restriction endonuclease HpyAIV. In the H. pylori strain G27, HpyAIV was recently identified as a gene that is upregulated transcriptionally by 2.59 fold in biofilm cells relative to planktonic cells [44]. Another disrupted gene, HPG27_1061, has not been characterized in G27 but its homolog was found to be a secreted protein dependent on environmental salt concentrations [45,46]. There are limited studies on the hypothetical proteins, cysteine-rich proteins, and the phosphatidyl glycerophosphate synthase, in H. pylori identified in this study suggesting we have much to learn about the roles of these proteins in general.
An interesting gene that was required for gap formation was luxS, a gene that facilitates a cell density based signaling mechanism via the production of Auto Inducer 2 (AI-2), a molecule used to regulate population growth amongst many bacterial species [47]. In H. pylori, AI-2 quorum sensing has been shown to affect expression of many genes, some involved in motility [48,49], hypothetical proteins, endorestriction nucleases and outer membrane proteins [50]. Thus, the role of luxS in gap formation could be due to any of several processes. Orthologs of some of the other identified genes, HPG27_147 and HPG27_412, have been shown to be impacted by LuxS in other H. pylori strains [50]. These genes encode a possible secreted beta-lactamase and a phosphatidyl glycerophosphate synthase. Unlike many bacteria that use AI-2 as an attractant, H. pylori perceive this molecule as a chemorepellent, moving away from its source using the TlpB chemoreceptors [49,51]. Although H. pylori possess a negative chemotactic response to AI-2, gap formation still occurred in tlpB mutants, suggesting that chemotactic responses are not a causative factor for gaps. AI-2 was recently proposed to reduce H. pylori adhesion to AGS cells by suppressing the expression of outer membrane proteins including AlpA and AlpB [52] and suppression of adhesion in H. pylori biofilms leads to the direct inhibition of growth [51]. One possibility is that under high cell density, these adhesins that are regulated by quorum sensing may facilitate binding to specific substrates or structures within the soft agar, indicating that spatial segregation may be maintained through specific molecular mechanisms. Though virtually all H. pylori strains contain genes encoding LuxS, AlpA, and AlpBtheir contribution to spatial organization between bacterial populations and potential binding to remains to be fully elucidated.
Altogether, our results suggest that H. pylori forms gaps on soft agar due to bacterial-intrinsic properties. We favor a model in which quorum sensing and other H. pylori genes, e.g. alpAB, create a physiological state that leads to gaps, which may involve a combination of changes to adhesive properties, lowering of growth or possibly even inhibiting growth of the encroaching colony. One possibility is that H. pylori detects the encroaching colony as “foreign” and has the ability to inhibit these cells. Although clonal populations are genetically identical, recent research in other pathogenic bacteria with isogenic strains has highlighted the role of sociomicrobiology in biofilms [10] and in vivo [35,53]. For example, isogenic P. aeruginosa strains can outcompete each other in murine lungs [53] and S. pneumoniae engages in fratricide to outcompete isogenic strains in murine nasal cavities [35]. Multiple studies have documented competition dynamics between H. pylori inoculums that affect gland occupation in vivo [21] [11], however kin selection and competition mechanisms have not been well characterized in H. pylori. Our experiments allude to a possible a non-secreted spatial regulation of competing H. pylori populations. Further understanding H. pylori social distancing mechanisms could reveal mechanisms that regulate H. pylori populations in the host and help us better understand previously documented gland occupation dynamics in vivo.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. The following supporting information can be downloaded at: www.mdpi.com/xxx/s1, Table S1. Primers used to locate Tn-7 insertion in merging mutants from Salama et al., 2004. Supplemental Figure 1. Clear gap treatment growth curve experiment schematic. Supplemental Figure 2. Merge mutant phenotypes are not affected by Soft Agar Nutrient and Agar Components. Supplemental Figure 3. Faster migration rates are not sufficient for gap formation. Supplemental Figure 4. Merge Tn-7 Mutants Planktonic growth in Liquid BB media with 2.5% FBS is similar to G27 WT.
Author Contributions
Conceptualization, YE and SK; methodology, YE, SK, and SH; investigation, YE, SK and SH; writing—original draft preparation, YE and KMO.; writing—review and editing, YE, SK, SH and KMO; supervision, SK and KMO; funding acquisition, KMO. All authors have read and agreed to the published version of the manuscript. Funding: The described project was supported by National Institutes of Health National Institute of Allergy and Infectious Disease (NIAID) grants number R01 AI116946 and R01 AI164682 (to K.M.O.). The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication.
Acknowledgments
The authors are grateful to all who contributed their guidance, support, and patience throughout this work. In times of uncertainty, collaboration and scientific inquiry remain beacons of progress, reminding us that knowledge, shared and pursued together, can illuminate even the most challenging paths forward.
References
- Wuichet, K.; Zhulin, I.B. Origins and Diversification of a Complex Signal Transduction System in Prokaryotes. Sci. Signal. 2010, 3. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, A.J.; Berg, H.C. Migration of Bacteria in Semisolid Agar. Proc. Natl. Acad. Sci. U.S.A. 1989, 86, 6973–6977. [Google Scholar] [CrossRef] [PubMed]
- Croze, O.A.; Ferguson, G.P.; Cates, M.E.; Poon, W.C.K. Migration of Chemotactic Bacteria in Soft Agar: Role of Gel Concentration. Biophysical Journal 2011, 101, 525–534. [Google Scholar] [CrossRef] [PubMed]
- Sanders, L.; Andermann, T.M.; Ottemann, K.M. A Supplemented Soft Agar Chemotaxis Assay Demonstrates the Helicobacter pylori Chemotactic Response to Zinc and Nickel. Microbiology 2013, 159, 46–57. [Google Scholar] [CrossRef]
- Sekowska, A.; Masson, J.-B.; Celani, A.; Danchin, A.; Vergassola, M. Repulsion and Metabolic Switches in the Collective Behavior of Bacterial Colonies. Biophysical Journal 2009, 97, 688–698. [Google Scholar] [CrossRef]
- Espeso, D.R.; Martínez-García, E.; De Lorenzo, V.; Goñi-Moreno, Á. Physical Forces Shape Group Identity of Swimming Pseudomonas putida Cells. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef]
- Strassmann, J.E.; Gilbert, O.M.; Queller, D.C. Kin Discrimination and Cooperation in Microbes. Annu. Rev. Microbiol. 2011, 65, 349–367. [Google Scholar] [CrossRef]
- Tipping, M.J.; Gibbs, K.A. Peer Pressure from a Proteus mirabilis Self-Recognition System Controls Participation in Cooperative Swarm Motility. PLoS Pathog 2019, 15, e1007885. [Google Scholar] [CrossRef]
- Budding, A.E.; Ingham, C.J.; Bitter, W.; Vandenbroucke-Grauls, C.M.; Schneeberger, P.M. The Dienes Phenomenon: Competition and Territoriality in Swarming Proteus mirabilis. J Bacteriol 2009, 191, 3892–3900. [Google Scholar] [CrossRef]
- Nadell, C.D.; Drescher, K.; Wingreen, N.S.; Bassler, B.L. Extracellular Matrix Structure Governs Invasion Resistance in Bacterial Biofilms. The ISME Journal 2015, 9, 1700–1709. [Google Scholar] [CrossRef]
- Fung, C.; Tan, S.; Nakajima, M.; Skoog, E.C.; Camarillo-Guerrero, L.F.; Klein, J.A.; Lawley, T.D.; Solnick, J.V.; Fukami, T.; Amieva, M.R. High-Resolution Mapping Reveals That Microniches in the Gastric Glands Control Helicobacter pylori Colonization of the Stomach. PLoS Biol 2019, 17, e3000231. [Google Scholar] [CrossRef] [PubMed]
- Foynes, S.; Dorrell, N.; Ward, S.J.; Stabler, R.A.; McColm, A.A.; Rycroft, A.N.; Wren, B.W. Helicobacter pylori Possesses Two CheY Response Regulators and a Histidine Kinase Sensor, CheA, Which Are Essential for Chemotaxis and Colonization of the Gastric Mucosa. Infect Immun 2000, 68, 2016–2023. [Google Scholar] [CrossRef]
- Sagoo, J.; Abedrabbo, S.; Liu, X.; Ottemann, K.M. Helicobacter pylori cheV1 Mutants Recover Semisolid Agar Migration Due to Loss of a Previously Uncharacterized Type IV Filament Membrane Alignment Complex Homolog. J Bacteriol 2024, 206, e00406–23. [Google Scholar] [CrossRef] [PubMed]
- Malfertheiner, P.; Camargo, M.C.; El-Omar, E.; Liou, J.-M.; Peek, R.; Schulz, C.; Smith, S.I.; Suerbaum, S. Helicobacter pylori Infection. Nat Rev Dis Primers 2023, 9, 19. [Google Scholar] [CrossRef] [PubMed]
- Uemura, N.; Yamaguchi, S.; Schlemper, R.J. Helicobacter pylori Infection and the Development of Gastric Cancer. The New England Journal of Medicine 2001. [Google Scholar] [CrossRef]
- Polk, D.B.; Peek, R.M. Helicobacter pylori: Gastric Cancer and Beyond. Nat Rev Cancer 2010, 10, 403–414. [Google Scholar] [CrossRef]
- Hu, Q.; Zhang, Y.; Zhang, X.; Fu, K. Gastric Mucosa-Associated Lymphoid Tissue Lymphoma and Helicobacter pylori Infection: A Review of Current Diagnosis and Management. Biomark Res 2016, 4, 15. [Google Scholar] [CrossRef]
- Ottemann, K.M.; Lowenthal, A.C. Helicobacter pylori Uses Motility for Initial Colonization and To Attain Robust Infection. Infect Immun 2002, 70, 1984–1990. [Google Scholar] [CrossRef]
- Terry, K.; Williams, S.M.; Connolly, L.; Ottemann, K.M. Chemotaxis Plays Multiple Roles during Helicobacter pylori Animal Infection. Infect Immun 2005, 73, 803–811. [Google Scholar] [CrossRef]
- Johnson, K.S.; Ottemann, K.M. Colonization, Localization, and Inflammation: The Roles of H. pylori Chemotaxis in Vivo. Current Opinion in Microbiology 2018, 41, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Keilberg, D.; Zavros, Y.; Shepherd, B.; Salama, N.R.; Ottemann, K.M. Spatial and Temporal Shifts in Bacterial Biogeography and Gland Occupation during the Development of a Chronic Infection. mBio 2016, 7, e01705–16. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Roujeinikova, A.; Ottemann, K.M. FliL Functions in Diverse Microbes to Negatively Modulate Motor Output via Its N-Terminal Region. mBio 2023, 14, e00283–23. [Google Scholar] [CrossRef] [PubMed]
- Censini, S.; Lange, C.; Xiang, Z.; Crabtree, J.E.; Ghiara, P.; Borodovsky, M.; Rappuoli, R.; Covacci, A. Cag, a Pathogenicity Island of Helicobacter pylori, Encodes Type I-Specific and Disease-Associated Virulence Factors. Proc. Natl. Acad. Sci. U.S.A. 1996, 93, 14648–14653. [Google Scholar] [CrossRef] [PubMed]
- Tomb, J.-F.; White, O.; Kerlavage, A.R.; Clayton, R.A.; Sutton, G.G.; Fleischmann, R.D.; Ketchum, K.A.; Klenk, H.P.; Gill, S.; Dougherty, B.A.; et al. The Complete Genome Sequence of the Gastric Pathogen Helicobacter pylori. Nature 1997, 388, 539–547. [Google Scholar] [CrossRef]
- Arnold, I.C.; Lee, J.Y.; Amieva, M.R.; Roers, A.; Flavell, R.A.; Sparwasser, T.; Müller, A. Tolerance Rather Than Immunity Protects From Helicobacter pylori–Induced Gastric Preneoplasia. Gastroenterology 2011, 140, 199–209.e8. [Google Scholar] [CrossRef]
- Lee, A.; O’Rourke, J.; De Ungria, M.; Robertson, B.; Daskalopoulos, G.; Dixon, M. A Standardized Mouse Model of Helicobacter pylori Infection: Introducing the Sydney Strain. Gastroenterology 1997, 112, 1386–1397. [Google Scholar] [CrossRef]
- Howitt, M.R.; Lee, J.Y.; Lertsethtakarn, P.; Vogelmann, R.; Joubert, L.-M.; Ottemann, K.M.; Amieva, M.R. ChePep Controls Helicobacter pylori Infection of the Gastric Glands and Chemotaxis in the Epsilonproteobacteria. mBio 2011, 2, e00098–11. [Google Scholar] [CrossRef]
- Pittman, M.S.; Goodwin, M.; Kelly, D.J. Chemotaxis in the Human Gastric Pathogen Helicobacter pylori: Different Roles for CheW and the Three CheV Paralogues, and Evidence for CheV2 Phosphorylation. Microbiology 2001, 147, 2493–2504. [Google Scholar] [CrossRef]
- Lowenthal, A.C.; Simon, C.; Fair, A.S.; Mehmood, K.; Terry, K.; Anastasia, S.; Ottemann, K.M. A Fixed-Time Diffusion Analysis Method Determines That the Three Chev Genes of Helicobacter pylori Differentially Affect Motility. Microbiology 2009, 155, 1181–1191. [Google Scholar] [CrossRef]
- Andermann, T.M.; Chen, Y.-T.; Ottemann, K.M. Two Predicted Chemoreceptors of Helicobacter pylori Promote Stomach Infection. Infect Immun 2002, 70, 5877–5881. [Google Scholar] [CrossRef]
- McGee, D.J.; Langford, M.L.; Watson, E.L.; Carter, J.E.; Chen, Y.-T.; Ottemann, K.M. Colonization and Inflammation Deficiencies in Mongolian Gerbils Infected by Helicobacter pylori Chemotaxis Mutants. Infect Immun 2005, 73, 1820–1827. [Google Scholar] [CrossRef] [PubMed]
- Collins, K.D.; Andermann, T.M.; Draper, J.; Sanders, L.; Williams, S.M.; Araghi, C.; Ottemann, K.M. The Helicobacter pylori CZB Cytoplasmic Chemoreceptor TlpD Forms an Autonomous Polar Chemotaxis Signaling Complex That Mediates a Tactic Response to Oxidative Stress. J Bacteriol 2016, 198, 1563–1575. [Google Scholar] [CrossRef]
- Salama, N.R.; Shepherd, B.; Falkow, S. Global Transposon Mutagenesis and Essential Gene Analysis of Helicobacter pylori. J Bacteriol 2004, 186, 7926–7935. [Google Scholar] [CrossRef]
- DeLoney-Marino, C.R.; Wolfe, A.J.; Visick, K.L. Chemoattraction of Vibrio fischeri to Serine, Nucleosides, and N -Acetylneuraminic Acid, a Component of Squid Light-Organ Mucus. Appl Environ Microbiol 2003, 69, 7527–7530. [Google Scholar] [CrossRef]
- Shen, P.; Lees, J.A.; Bee, G.C.W.; Brown, S.P.; Weiser, J.N. Pneumococcal Quorum Sensing Drives an Asymmetric Owner–Intruder Competitive Strategy during Carriage via the Competence Regulon. Nat Microbiol 2018, 4, 198–208. [Google Scholar] [CrossRef]
- Odenbreit, S.; Faller, G.; Haas, R. Role of the AlpAB Proteins and Lipopolysaccharide in Adhesion of Helicobacter pylori to Human Gastric Tissue. International Journal of Medical Microbiology 2002, 292, 247–256. [Google Scholar] [CrossRef]
- Odenbreit, S.; Swoboda, K.; Barwig, I.; Ruhl, S.; Borén, T.; Koletzko, S.; Haas, R. Outer Membrane Protein Expression Profile in Helicobacter pylori Clinical Isolates. Infect Immun 2009, 77, 3782–3790. [Google Scholar] [CrossRef]
- Senkovich, O.A.; Yin, J.; Ekshyyan, V.; Conant, C.; Traylor, J.; Adegboyega, P.; McGee, D.J.; Rhoads, R.E.; Slepenkov, S.; Testerman, T.L. Helicobacter pylori AlpA and AlpB Bind Host Laminin and Influence Gastric Inflammation in Gerbils. Infect Immun 2011, 79, 3106–3116. [Google Scholar] [CrossRef]
- Yonezawa, H.; Osaki, T.; Woo, T.; Kurata, S.; Zaman, C.; Hojo, F.; Hanawa, T.; Kato, S.; Kamiya, S. Analysis of Outer Membrane Vesicle Protein Involved in Biofilm Formation of Helicobacter pylori. Anaerobe 2011, 17, 388–390. [Google Scholar] [CrossRef]
- Yonezawa, H.; Osaki, T.; Fukutomi, T.; Hanawa, T.; Kurata, S.; Zaman, C.; Hojo, F.; Kamiya, S. Diversification of the AlpB Outer Membrane Protein of Helicobacter pylori Affects Biofilm Formation and Cellular Adhesion. J Bacteriol 2017, 199. [Google Scholar] [CrossRef]
- De Jonge, R.; Durrani, Z.; Rijpkema, S.G.; Kuipers, E.J.; Van Vliet, A.H.M.; Kusters, J.G. Role of the Helicobacter pylori Outer-Membrane Proteins AlpA and AlpB in Colonization of the Guinea Pig Stomach. Journal of Medical Microbiology 2004, 53, 375–379. [Google Scholar] [CrossRef] [PubMed]
- Odenbreit, S.; Till, M.; Hofreuter, D.; Faller, G.; Haas, R. Genetic and Functional Characterization of the alpAB Gene Locus Essential for the Adhesion of Helicobacter pylori to Human Gastric Tissue. Molecular Microbiology 1999, 31, 1537–1548. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Wu, J.Y.; Beswick, E.J.; Ohno, T.; Odenbreit, S.; Haas, R.; Reyes, V.E.; Kita, M.; Graham, D.Y.; Yamaoka, Y. Functional and Intracellular Signaling Differences Associated with the Helicobacter pylori AlpAB Adhesin from Western and East Asian Strains. Journal of Biological Chemistry 2007, 282, 6242–6254. [Google Scholar] [CrossRef]
- Hathroubi, S.; Hu, S.; Ottemann, K.M. Genetic Requirements and Transcriptomics of Helicobacter pylori Biofilm Formation on Abiotic and Biotic Surfaces. npj Biofilms Microbiomes 2020, 6, 56. [Google Scholar] [CrossRef]
- Voss, B.J.; Loh, J.T.; Hill, S.; Rose, K.L.; McDonald, W.H.; Cover, T.L. Alteration of the Helicobacter pylori Membrane Proteome in Response to Changes in Environmental Salt Concentration. Proteomics Clinical Apps 2015, 9, 1021–1034. [Google Scholar] [CrossRef]
- Caston, R.R.; Loh, J.T.; Voss, B.J.; McDonald, W.H.; Scholz, M.B.; McClain, M.S.; Cover, T.L. Effect of Environmental Salt Concentration on the Helicobacter pylori Exoproteome. Journal of Proteomics 2019, 202, 103374. [Google Scholar] [CrossRef]
- Joyce, E.A.; Bassler, B.L.; Wright, A. Evidence for a Signaling System in Helicobacter pylori : Detection of a luxS -Encoded Autoinducer. J Bacteriol 2000, 182, 3638–3643. [Google Scholar] [CrossRef]
- Rader, B.A.; Campagna, S.R.; Semmelhack, M.F.; Bassler, B.L.; Guillemin, K. The Quorum-Sensing Molecule Autoinducer 2 Regulates Motility and Flagellar Morphogenesis in Helicobacter pylori. J Bacteriol 2007, 189, 6109–6117. [Google Scholar] [CrossRef]
- Rader, B.A.; Wreden, C.; Hicks, K.G.; Sweeney, E.G.; Ottemann, K.M.; Guillemin, K. Helicobacter pylori Perceives the Quorum-Sensing Molecule AI-2 as a Chemorepellent via the Chemoreceptor TlpB. Microbiology 2011, 157, 2445–2455. [Google Scholar] [CrossRef]
- Yang, H.; Huang, X.; Zhang, X.; Zhang, X.; Xu, X.; She, F.; Wen, Y. AI-2 Induces Urease Expression Through Downregulation of Orphan Response Regulator HP1021 in Helicobacter pylori. Front. Med. 2022, 9, 790994. [Google Scholar] [CrossRef]
- Anderson, J.K.; Huang, J.Y.; Wreden, C.; Sweeney, E.G.; Goers, J.; Remington, S.J.; Guillemin, K. Chemorepulsion from the Quorum Signal Autoinducer-2 Promotes Helicobacter pylori Biofilm Dispersal. mBio 2015, 6, e00379–15. [Google Scholar] [CrossRef] [PubMed]
- Helicobacter - 2021 - Wen - AI-2 Represses CagA Expression and Bacterial Adhesion Attenuating the Helicobacter.Pdf.
- Czechowska, K.; McKeithen-Mead, S.; Al Moussawi, K.; Kazmierczak, B.I. Cheating by Type 3 Secretion System-Negative Pseudomonas aeruginosa during Pulmonary Infection. Proc. Natl. Acad. Sci. U.S.A. 2014, 111, 7801–7806. [Google Scholar] [CrossRef]
Figure 1.
H. pylori soft agar inoculations produce gaps between adjacent colonies .H. pylori strain G27 pTM115 (GFP+) was inoculated in standard H. pylori soft agar media, incubated under microaerobic conditions, and imaged at the indicated times. (A) At 7 days of incubation, soft agar inoculations have three zones: non-competing front, competing front and gap. (B) The plate shown in panel A was imaged days one-six. (C) The distances migrated by the competing (C) and non-competing (NC) fronts from the inoculation point were measured daily over six days for 4 biological replicates. Shown are averages with error bars representing SEM. Biological replicates were compared statistically using paired T-test analysis with significance indicated by *(p-value<0.05), **(p-value<0.01) and *** (p-value<0.001). .
Figure 1.
H. pylori soft agar inoculations produce gaps between adjacent colonies .H. pylori strain G27 pTM115 (GFP+) was inoculated in standard H. pylori soft agar media, incubated under microaerobic conditions, and imaged at the indicated times. (A) At 7 days of incubation, soft agar inoculations have three zones: non-competing front, competing front and gap. (B) The plate shown in panel A was imaged days one-six. (C) The distances migrated by the competing (C) and non-competing (NC) fronts from the inoculation point were measured daily over six days for 4 biological replicates. Shown are averages with error bars representing SEM. Biological replicates were compared statistically using paired T-test analysis with significance indicated by *(p-value<0.05), **(p-value<0.01) and *** (p-value<0.001). .
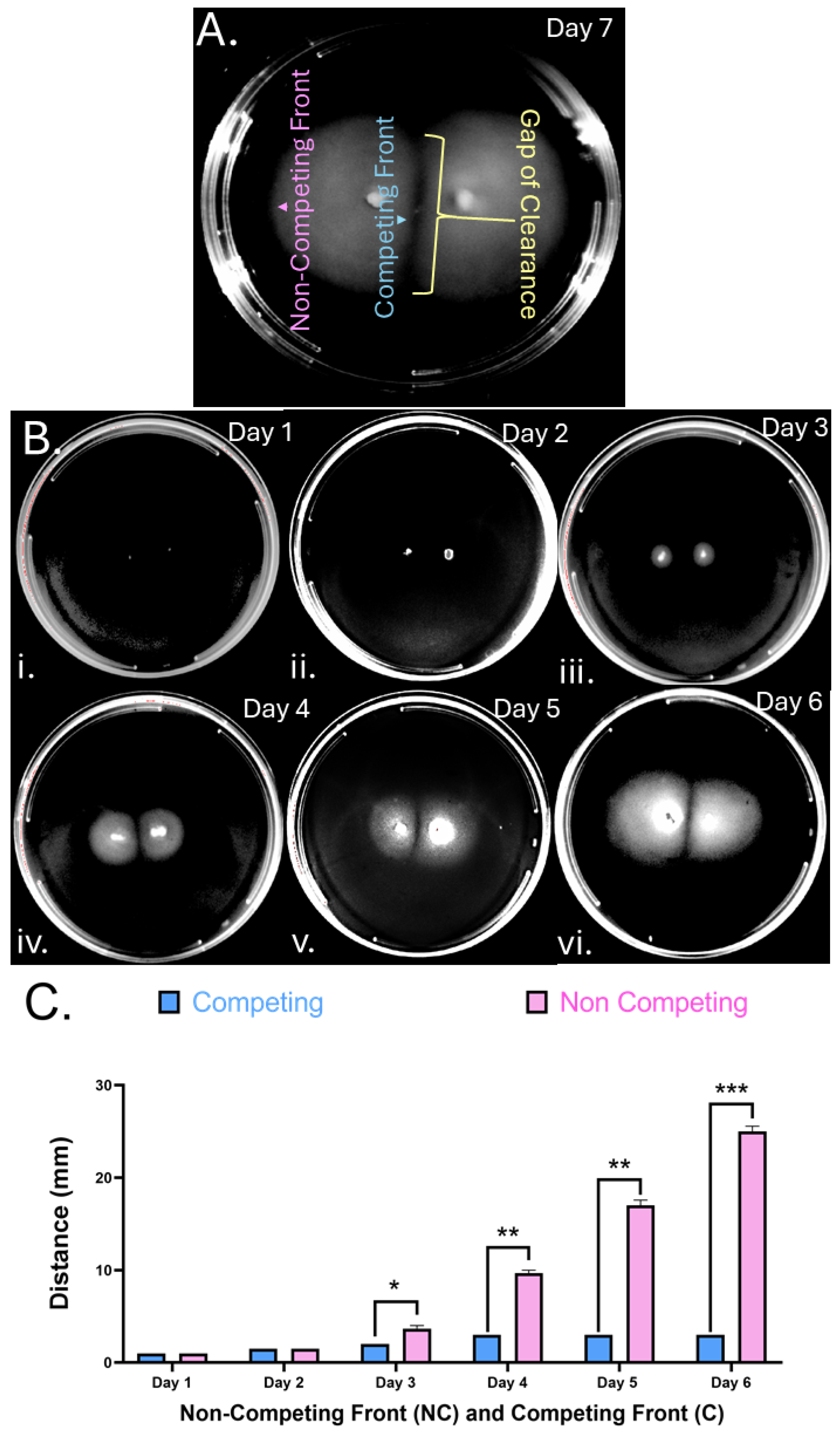
Figure 2.
H. pylori soft agar colony gaps are independent of chemotaxis. Partial chemotaxis and chemoreceptor mutants in strain G27 were inoculated in soft agar and imaged after 9 days (panel A) or 6 days (panel B). (A) Chemotaxis signaling mutants lacking (i) ∆chePep, (ii) ∆cheV2, (iii) ∆cheV3. (B) Chemotaxis receptor mutants (i) ∆tlpA, (ii) ∆tlpB, (iii) ∆tlpC, (iv) ∆tlpD. Images are representative of four biological replicates per mutant.
Figure 2.
H. pylori soft agar colony gaps are independent of chemotaxis. Partial chemotaxis and chemoreceptor mutants in strain G27 were inoculated in soft agar and imaged after 9 days (panel A) or 6 days (panel B). (A) Chemotaxis signaling mutants lacking (i) ∆chePep, (ii) ∆cheV2, (iii) ∆cheV3. (B) Chemotaxis receptor mutants (i) ∆tlpA, (ii) ∆tlpB, (iii) ∆tlpC, (iv) ∆tlpD. Images are representative of four biological replicates per mutant.
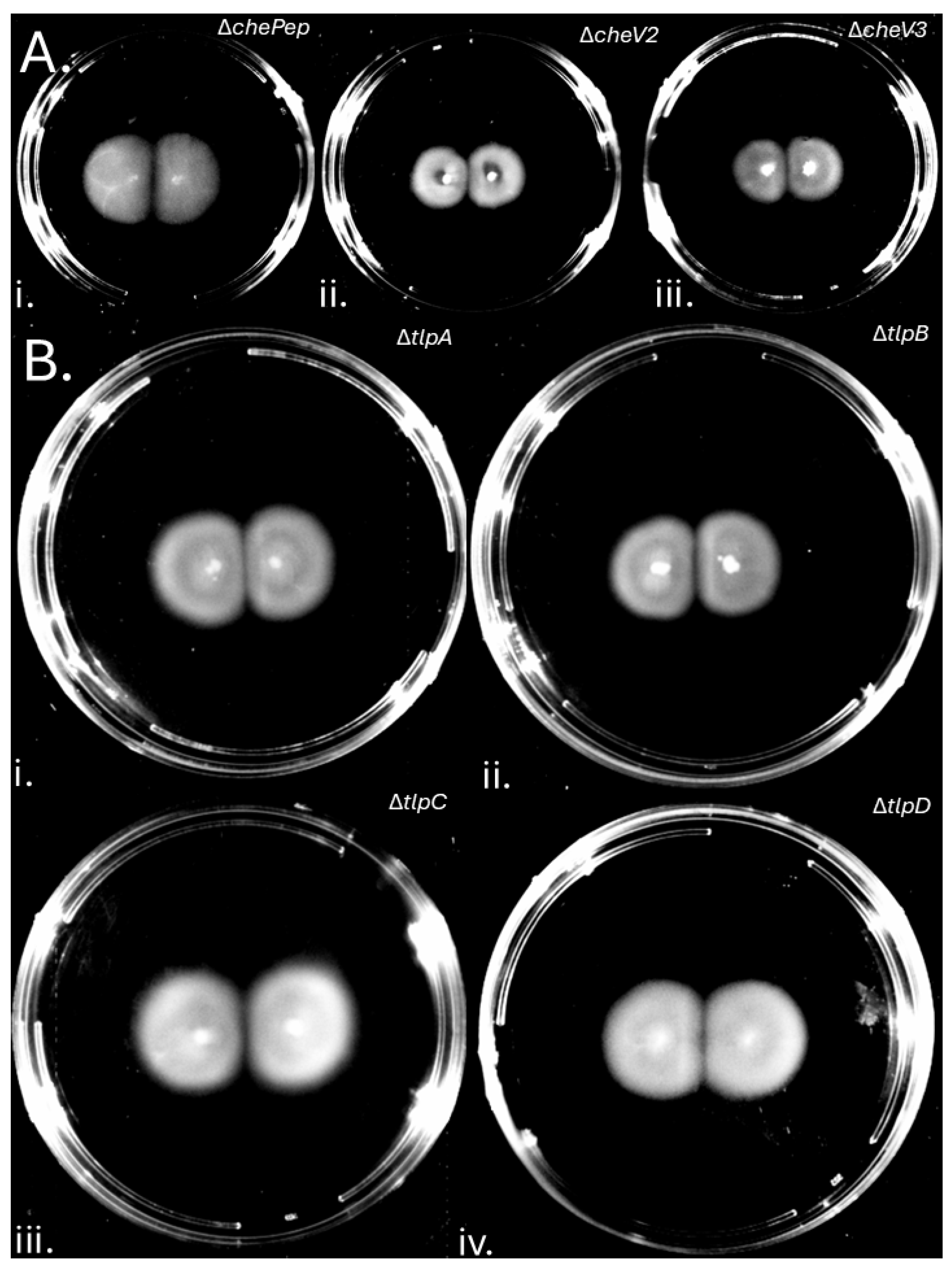
Figure 3.
Gap Formation is Independent of Soft Agar Nutrient and Agar Components. Soft agar plates were inoculated in two places 1 cm apart with H. pylori G27 GFP+ on plates with varying brucella broth (BB) (A) or agar levels (B). (A) (i) 0.5XBB for 7 Days, (ii) 0.75XBB for 7 Days and (iii) 1.5XBB for 9 Days. (B) (i) 0.25%, (ii) 0.3%, (iii) 0.35%, (iv) 0.4% and (v) 0.45% agar. .
Figure 3.
Gap Formation is Independent of Soft Agar Nutrient and Agar Components. Soft agar plates were inoculated in two places 1 cm apart with H. pylori G27 GFP+ on plates with varying brucella broth (BB) (A) or agar levels (B). (A) (i) 0.5XBB for 7 Days, (ii) 0.75XBB for 7 Days and (iii) 1.5XBB for 9 Days. (B) (i) 0.25%, (ii) 0.3%, (iii) 0.35%, (iv) 0.4% and (v) 0.45% agar. .
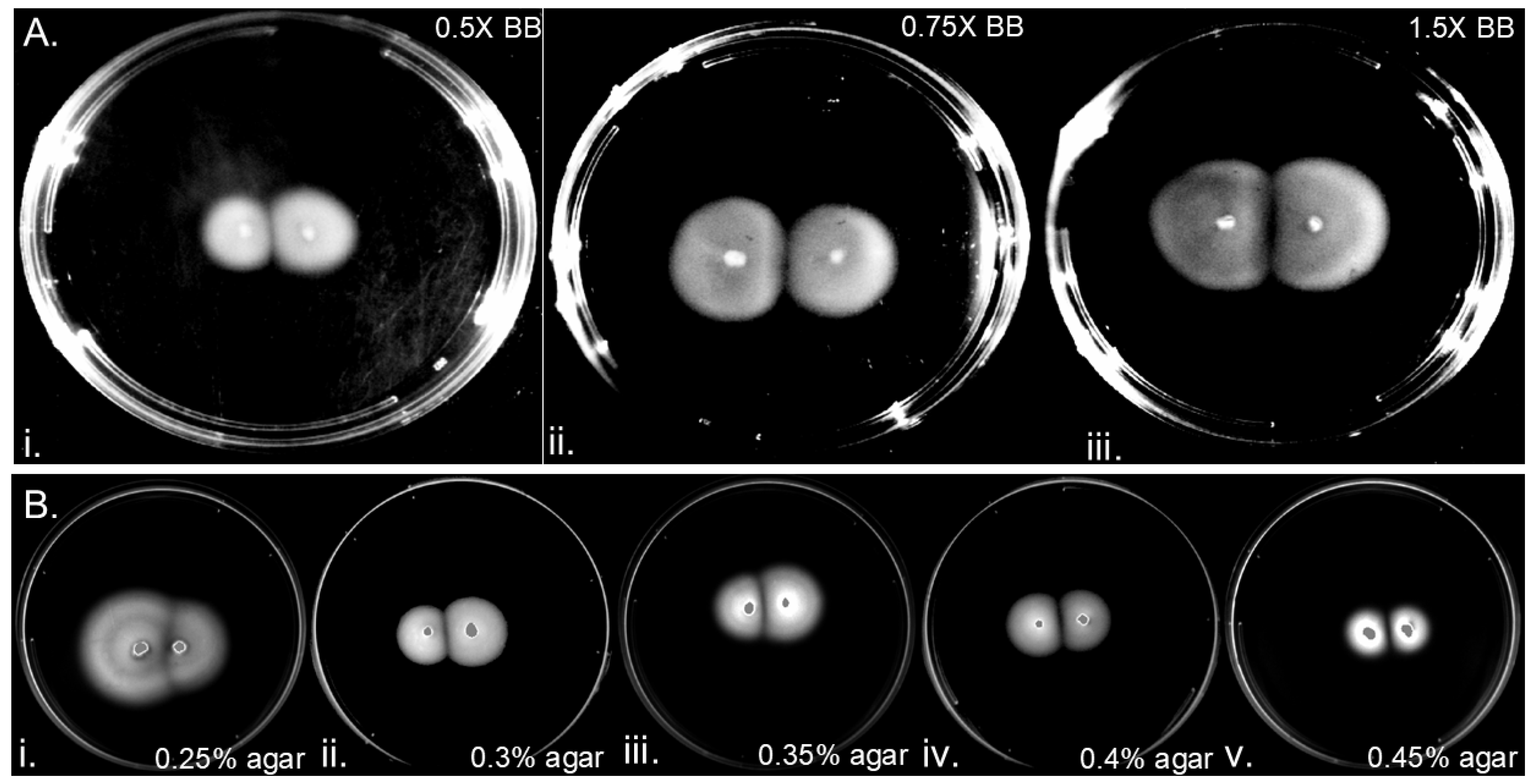
Figure 4.
Gaps of clearance contain H. pylori with spiral morphology at a low but non-zero density.H. pylori G27 soft agar plates were inoculated in two spots 1 cm apart and samples from the resulting colonies imaged after 6 days using microscopy. A 10 uL aliquot of soft agar was taken from either (A) from within the gap region (top panels) or (B) from within the colony (bottom panels) and imaged with phase contrast microscopy, at (i) 200X magnification, (ii) 400X magnification, or (iii) 1000X magnification. Images are indicative of three biological replicates. (C) Whatman paper was inserted into the clear gap of inoculated plates (i) was then exposed to soft agar for 6 days showed bacterial growth in comparison to a negative control, a Whatman paper that was inserted in uninoculated agar (ii).
Figure 4.
Gaps of clearance contain H. pylori with spiral morphology at a low but non-zero density.H. pylori G27 soft agar plates were inoculated in two spots 1 cm apart and samples from the resulting colonies imaged after 6 days using microscopy. A 10 uL aliquot of soft agar was taken from either (A) from within the gap region (top panels) or (B) from within the colony (bottom panels) and imaged with phase contrast microscopy, at (i) 200X magnification, (ii) 400X magnification, or (iii) 1000X magnification. Images are indicative of three biological replicates. (C) Whatman paper was inserted into the clear gap of inoculated plates (i) was then exposed to soft agar for 6 days showed bacterial growth in comparison to a negative control, a Whatman paper that was inserted in uninoculated agar (ii).
Figure 5.
Analysis of Clear Gap Effect on Growth.H. pylori G27 pTM115 (KanR) liquid cultures were incubated for seven hours in microaerobic conditions with material collected from (A) clear gap extracted from between WT H. pylori G27 colonies (B) uninoculated agar or (C) no treatment control. After treatment, samples were serially diluted, and spot plated. (D) CFU data calculated from 3 biological replicates; shown are averages with error bars representing SEM of samples treated as in A-C. (n.s., p-value= 0.0646; *, p-value < 0.05) using Students T-test analysis. .
Figure 5.
Analysis of Clear Gap Effect on Growth.H. pylori G27 pTM115 (KanR) liquid cultures were incubated for seven hours in microaerobic conditions with material collected from (A) clear gap extracted from between WT H. pylori G27 colonies (B) uninoculated agar or (C) no treatment control. After treatment, samples were serially diluted, and spot plated. (D) CFU data calculated from 3 biological replicates; shown are averages with error bars representing SEM of samples treated as in A-C. (n.s., p-value= 0.0646; *, p-value < 0.05) using Students T-test analysis. .
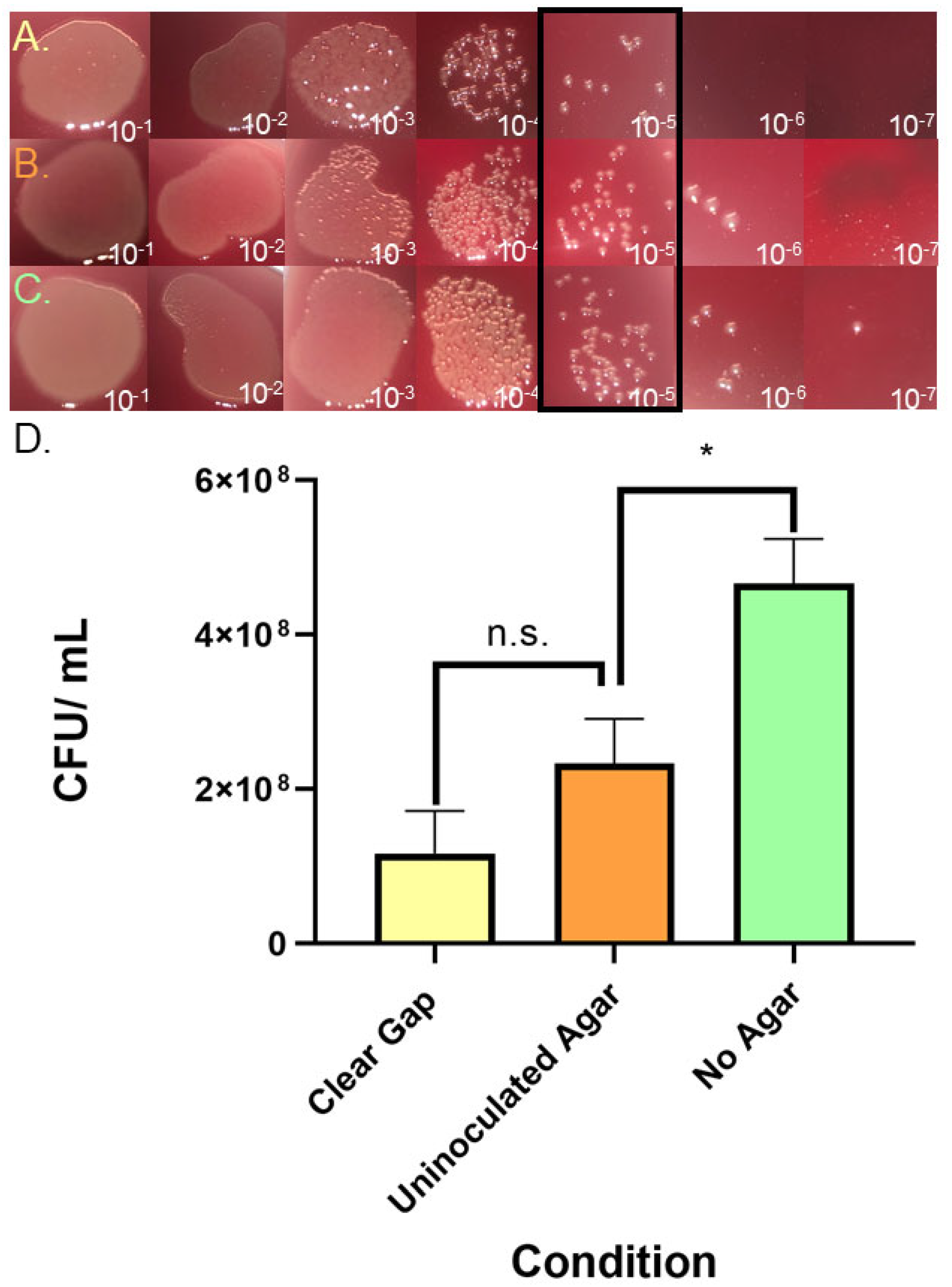
Figure 6.
Isolation of genetic mutations that cause H. pylori to lose gap formation. (A) A H. pylori G27 Tn7 pool [33] was plated at positions 1 cm apart for 6 days on standard soft agar (i). Isolates were taken from the non-competitive front, the competitive front and the gap regions. Example phenotypes observed in Tn7 mutants isolated from within the colony at the non-competitive front (ii), the competitive front (iii) or from the gap region (iv). (B) Merging phenotypes observed in (i) Tn-7 mutants (ii) ∆luxS, and the merging phenotypes can be recapitulated when transforming Tn-7 genomic DNA into (iii) G27 WT or (iv) G27 GFP+. .
Figure 6.
Isolation of genetic mutations that cause H. pylori to lose gap formation. (A) A H. pylori G27 Tn7 pool [33] was plated at positions 1 cm apart for 6 days on standard soft agar (i). Isolates were taken from the non-competitive front, the competitive front and the gap regions. Example phenotypes observed in Tn7 mutants isolated from within the colony at the non-competitive front (ii), the competitive front (iii) or from the gap region (iv). (B) Merging phenotypes observed in (i) Tn-7 mutants (ii) ∆luxS, and the merging phenotypes can be recapitulated when transforming Tn-7 genomic DNA into (iii) G27 WT or (iv) G27 GFP+. .
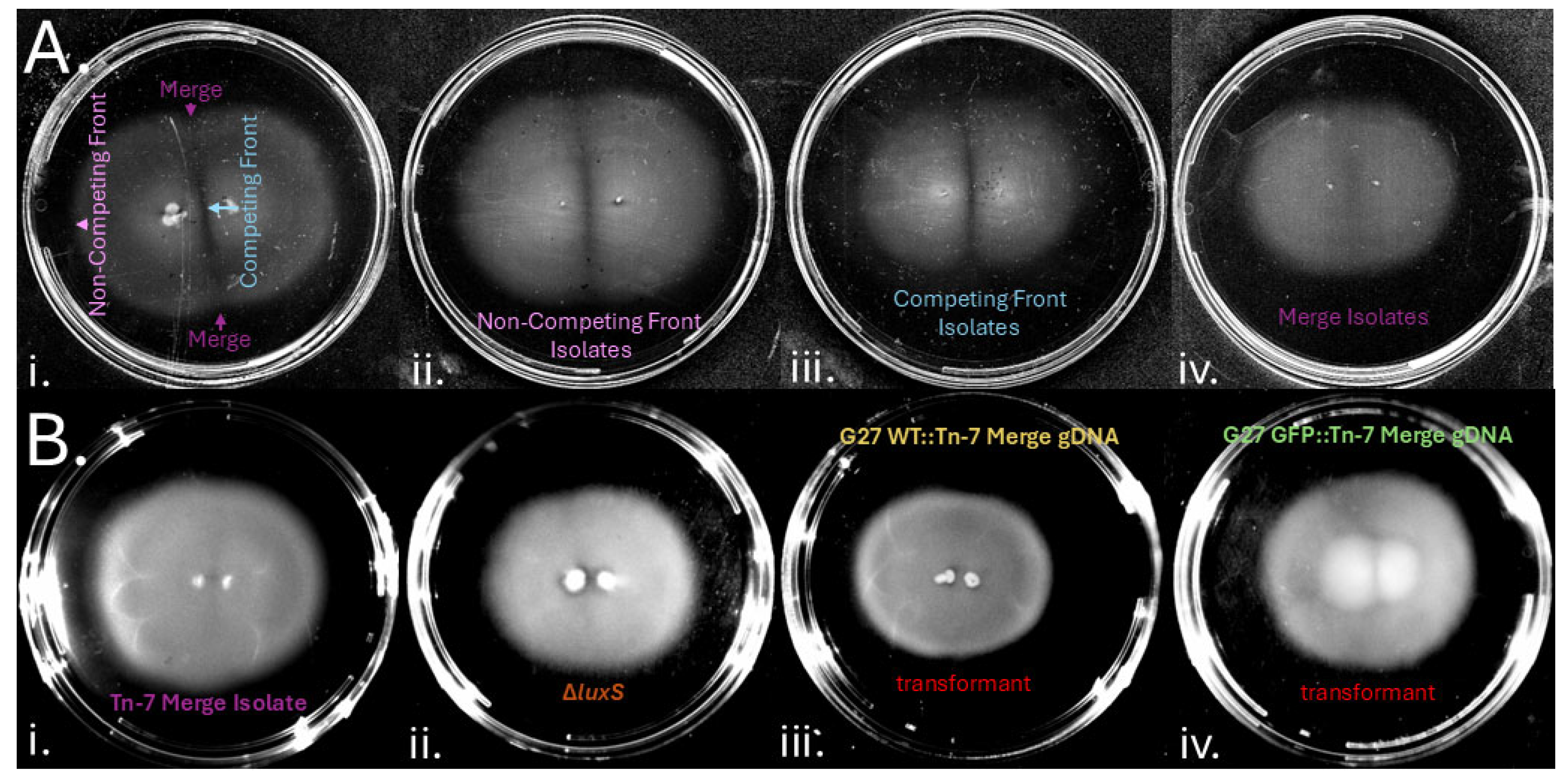

Table 1.
Genes with Tn-7 insertion in merging mutants.
| Gene Disrupted by Tn-7 | Gene Function | Number of Isolates |
|---|---|---|
| HPG27_843 Intergenic HPG27_843&844 HPG27_147 |
Outer membrane protein HopC/AlpA | 3 |
| Outer membrane protein HopCB/AlpAB locus | 2 | |
| cysteine-rich protein D | 1 | |
|
HPG27_412 HPG27_413 |
phosphatidyl glycerophosphate synthase | 1 |
| Hypothetical protein | 1 | |
|
HPG27_462 HPG27_1061 HPG27_1062 HPG27_1299 |
Hypothetical protein | 1 |
| Cysteine-rich protein X | 1 | |
| Hypothetical Protein | 1 | |
| Type II Endorestriction Nuclease HpyAIV | 2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



