
Submitted:
01 April 2025
Posted:
03 April 2025
You are already at the latest version
Abstract
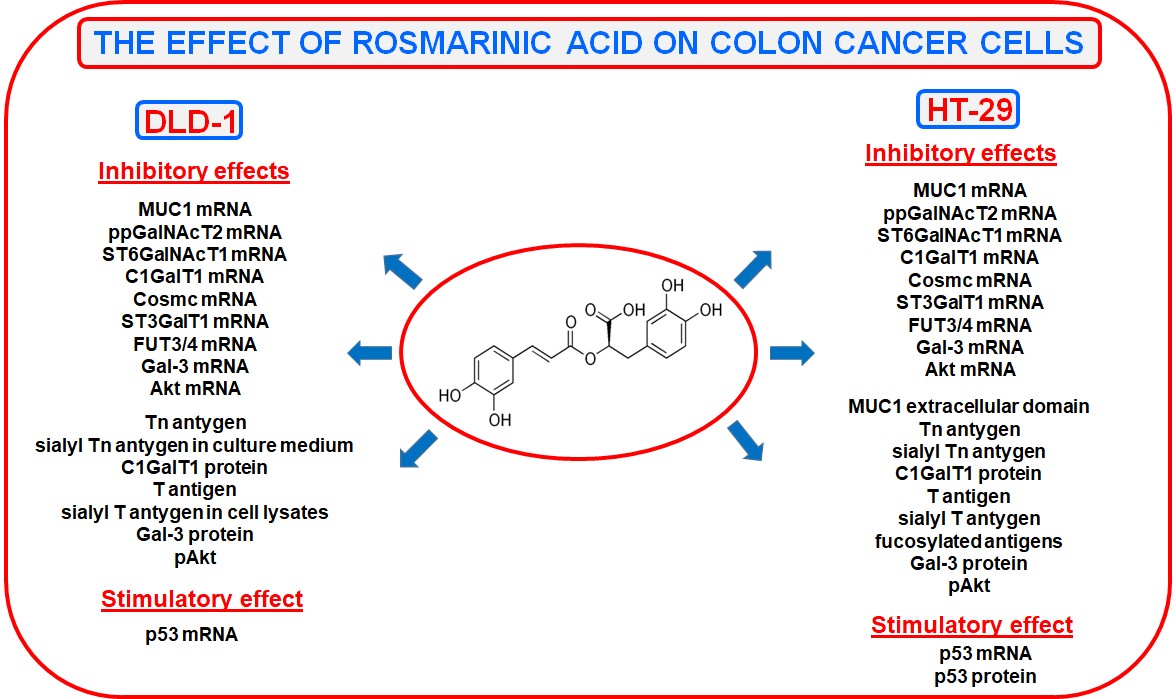
Rosmarinic acid (RA) is polyphenolic compound with beneficial, health-promoting effects. Due to limited studies regarding the influence of RA on colon cancer cells, we examined how this acid influences selected glycoforms, enzymes participating in their formation and Gal-3, Akt, p53 in DLD-1 and HT-29 colon cancer cell lines. To determine the expression of studied factors, qRT-PCR, Western blotting and ELISA tests were applied. We revealed inhibitory effect of RA in both cell lines on MUC1, ppGalNAcT2, ST6GalNAcT1, C1GalT1, Cosmc, ST3GalT1, FUT3/4, Gal-3, and Akt mRNAs. p53 mRNA was increased. Moreover, we observed the suppression of Tn, T antigen as well as C1GalT1 and pAkt protein expression as the result of RA action. Apart from that, in HT-29 colon cancer cells, rosmarinic acid inhibited MUC1 extracellular domain, sialyl Tn, sialyl T, fucosylated antigens, and Gal-3 expression. p53 protein was elevated upon RA action. In DLD-1 cancer cells, RA diminished the expression of sialyl Tn in culture medium and sialyl T in cell lysates. The data suggest potential usefulness of RA as a complementary agent supporting chemotherapy in colon in cancer treatment.
Keywords:
colon cancer
; glycosylation
; MUC1
; rosmarinic acid
1. Introduction
Colon cancer is estimated to be one of the most dangerous malignant tumors which often spreads to the lungs, ovaries, liver and other parts of the gastrointestinal system, and also is the world’s second notable cause of cancer related deaths. According to GLOBOCAN 2020 there will be about 1.92 million new cases of colon cancer worldwide in 2040 [1,2,3,4]. Surgery and chemotherapy are the main tools of this cancer therapy, with much higher effectiveness at early stages of the disease. However, such treatment is very often linked to high cytotoxicity of chemotherapeutics as well as resistance to healing [2]. New therapies are introduced, but 5-year relative survival still remains at only about 65% [5]. An increasing number of reports shows that natural medications produced from plants, used by about 70% of the world population, may be applied in the healing of many cancers, including colon ones [2,6]. It has been demonstrated that polyphenolic compounds, characterized as derivatives of 2-phenyl-benzo-γ-pyrone, extracted from many herbs, are responsible for anti-cancer effects through induction of apoptosis and autophagy, cell cycle arrest, modulation of tumor-suppressive microRNA or by triggering different signaling pathways [6,7]. Using them as potential anti-cancer agents is especially important because of their very few side effects, unlike many chemotherapeutics.
The impact on specifically altered glycoforms of cancer cells, inherently associated with the development of tumors, can be one more, less explored goal of bioactive molecules. Alterations in mucin-type O-linked glycosylation have been related to cancer development and poor prognosis [8,9,10]. Glycosylation-mediated progress of cancer is based on mediation of glycans in oncogenic signal transduction, ligand-receptor interaction, cell-cell, and cell-matrix adhesion in tumor cells [11]. Cancer cells exhibit a considerable range of glycosylation modifications compared to normal ones [12]. Alterations in glycosylation allow malignant cells to stimulate cell mobility, cell adhesion, receptor activation, and by that contributing to the invasive phenotype [13]. Transmembrane MUC1 mucin is one of the main carriers of tumor associated glycoforms. It is reported that MUC1 has minimal or even absent expression in healthy colonic tissues while it is upregulated in about 55 – 100% of colorectal cancers [14,15]. Aberrant, tumor-associated carbohydrate antigens (TACAs), presented especially by cancerous MUC1, include prematurely terminated monosaccharides (Tn antigens) or disaccharides (T antigen), their sialylated forms (sialyl Tn and sialyl T) as well as Lewis antigens [12].
Rosmarinic acid (RA), an ester of caffeic acid and 3-(3,4-dihydroxyphenyl)lactic acid (Figure 1), polyphenol present in medicinal plants, herbs, and species, is a compound with many beneficial, health-promoting effects, including anti-cancer [16,17]. RA has been reported to act by different mechanisms such as reduction of TNF-α, COX-2, IL-6 levels, modulation of p65 expression, stimulation apoptosis by modifying various gene expression, and involvement in apoptosis regulation [18,19]. Recently we have reported promising results revealing potential of rosmarinic acid against AGS gastric cancer cells. RA was able to suppress expression of MUC1 mucin, specific cancer-related carbohydrate antigens, enzymes responsible for their formation as well as some other factors responsible for cancer development [20,21]. Upon these encouraging outcomes we decided to examine the action of rosmarinic acid towards glycoforms of DLD-1 and HT-29 colon cancer cells. According to our knowledge there are no reports considering such effects.
2. Results
2.1. Viability of DLD-1 and HT-29 Colon Cancer Cells in the Presence of Rosmarinic Acid
MTT test was applied to determine the effect of rosmarinic acid action on DLD-1 and HT-29 colon cancer cells. It was revealed that 50 – 400 μM concentrations of RA exerted very low cytotoxicity on examined cells (Figure 2). Viability of the cells was not lower than 80% compared with control where no acid was added. In our experiments we decided to choose 200 and 400 μM RA concentration.
2.2. The Effect of Rosmarinic Acid on MUC1 Expression
As MUC1 is the main carrier of sugar antigens examined in the study, at first, we decided to check the influence of RA on this mucin. The expression of MUC1 mRNA as well as the level of extracellular domain of this glycoprotein culture medium were assessed. Quantitative real time PCR (qRT-PCR) revealed inhibitory effect of RA on MUC1 mRNA in both examined cancer cells, with stronger result in DLD-1 cell line (Figure 3A). However, inhibitory effect of rosmarinic acid on MUC1extracellular domain released to the culture medium is seen only in DLD-1 cells (Figure 3B).
2.3. The Effect of RA on ppGalNAcT2, ST6GalNAcT1, Tn, and Sialyl Tn Antigens Expression
Tn antigen (GalNAcα-Ser/Thr) and its sialyl form (sTn; Neu5Acα2-6GalNAcα-Ser/Thr) are the results of the action of proper polypeptide α-N-acetylgalactosaminyltransferase (ppGalNAcT) and sialyltransferase (ST6GalNAcT), respectively. qRT-PCR assays demonstrated that RA with 200 and 400 μM concentration effectively suppressed the expression of ppGalNAcT2 mRNA in both cell lines (Figure 4A). However, the examination of Tn antigen by ELISA test with biotinylated VVA lectin didn’t reveal such inhibitory effect in DLD-1 cell line (Figure 4B). In case of HT-29 colon cancer cells, suppression of Tn antigen was confirmed in cell lysates, as the effect of 400 μM RA action and in culture medium as the effect of two applied concentrations of acid (Figure 4C). RA with both concentrations decreased the expression of ST6GalNAcT1 on mRNA level in DLD-1 cells, but in case of HT-29 line, such effect was revealed only with higher concentration of rosmarinic acid (Figure 4D). In ELISA test for DLD-1 cell line, performed with biotinylated SNA lectin detecting sialyl Tn antigen, inhibitory effect of RA was observed only in culture medium (Figure 4E). In HT-29, such suppression was revealed in culture medium as well as in cell lysates only by 400 μM rosmarinic acid (Figure 4F).
2.4. The Effect of RA on C1GalT1, Cosmc, ST3GalT1, T, and Sialyl T Antigens Expression
T (TF) sugar structure (Galβ1-3GalNAcα-Ser/Thr) and sialyl T (Neu5Acα2-3Galβ1-3GalNAcα-Ser/Thr) are formed upon action of core 1 β1-3galactosyltransferase (T synthase; C1GalT1) and β-galactoside α2-3sialyltransferase (ST3GalT1), respectively. T synthase needs unique molecular endoplasmic reticulum chaperone (Cosmc; C1GalT1C1) which is required to synthetize active form of this enzyme. C1GalT1 mRNA was significantly suppressed by the action of both RA concentrations in DLD-1 cells, and by 400 μM in HT-29 cells (Figure 5A). Similar inhibitory effect was revealed for the enzyme on the protein level in DLD-1 cells; in case of HT-29, only as the result of 200 μM RA action (Figure 5B). Cosmc mRNA in both cancer cells was suppressed by both concentrations of RA (Figure 5C). We revealed also inhibition of T antigen expression, in ELISA test with biotinylated PNA lectin, by both concentrations of RA in culture medium of DLD-1 cells, and by 400 μM RA in cell lysates (Figure 5D). In HT-29 colon cancer cells, suppression of this antigen expression was observed in cell lysates and culture medium by both RA concentrations (Figure 5E). ST3GalT1 on mRNA level was inhibited in two cell lines by two applied doses of rosmarinic acid (Figure 5F). In ELISA test with biotinylated MAAII lectin we revealed that RA suppressed sialyl T antigen expression in cell lysates of DLD-1 cells (Figure 5G), and in cell lysates and culture medium of HT-29 cells (Figure 5H).
2.5. The Effect of RA on FUT3, FUT4, and Fucosylated Antigens Expression
FUT3 and FUT4 are fucosyltransferases responsible for fucose addition to Gal or GlcNAc and Lewis antigens formation. FUT3 and FUT4 on mRNA level were suppressed by rosmarinic acid in two colon cancer cell lines (Figure 6A,B). However, in ELISA tests, Lewis antigens recognized by LTA lectin were not influenced by RA in DLD-1 cell lines, and inhibited in HT-29 cells (Figure 6C,D).
2.6. The Effect of RA on Gal-3 Expression
Gal-3 is multifunctional, oncogenic protein with high affinity to T carbohydrate antigen. Both applied concentrations of rosmarinic acid suppressed Gal-3 mRNA expression in two colon cancer cell lines (Figure 7A). Moreover, Gal-3 protein expression was inhibited by 200 and 400 μM RA in DLD-1 cells and by 400 μM RA in HT-29 cells (Figure 7B).
2.7. The Effect of RA on Akt Expression
Akt is protein kinase playing critical role in promoting cell survival and apoptosis. Rosmarinic acid inhibited Akt mRNA in both colon cancer cell lines (Figure 8A). In addition, active, phosphorylated form of Akt was suppressed by both concentrations of RA in DLD-1 cells as well as in HT-29 after action of 400 μM RA (Figure 8B).
2.8. The Effect of RA on p53 Expression
In Figure 9A we demonstrate that p53 mRNA, tumor suppressor protein, was stimulated by 200 μM RA concentration in both colon cancer cell lines. 400 μM concentration of rosmarinic acid didn’t influence this factor in DLD-1 cells, and surprisingly inhibited p53 in HT-29 cells. p53 protein expression was stimulated only in HT-29 cells by lower concentration of RA, and inhibited by higher concentration of acid (Figure 9B).
3. Discussion
It has been revealed that altered glycosylation is a common feature in various steps of malignant transformation and progression. Very often glycosylation variations are relatively specific for the type and stage of cancer, thereby making glycans as potential cancer biomarkers as well as targets for anticancer therapy [10,11,12,22]. There are many ways by which O-glycans can regulate tumorigenesis. Carbohydrates on a receptor protein tend to affect its properties, activity, expression level, and stability. Glycans can directly mediate oncogenic signal transduction, ligand-receptor interactions, cell-cell, and cell-matrix adhesion in malignant cells, and in this way are associated with cancer progression and metastasis which correlate with poor prognosis and high mortality [11,12,23,24]. There are also reports concluding, that expression of specific glycans is associated with tumor escape from immune defenses [25,26].
MUC1 is main, heavily O-glycosylated protein of gastrointestinal epithelium, overexpressed and associated with progression in diverse cancers, such as colon, breast, lung, pancreatic, and prostate [14,23,24,27,28,29,30,31]. Thus, glycosylation of epithelium origin cancer cells is said to be mucin-type O-linked glycosylation [9,11,22]. MUC1 is considered as oncogene that regulates cell growth, proliferation, metastasis, apoptosis by participating in various signaling pathways [32]. Moreover, aberrantly glycosylated MUC1 can be recognized by B cell-derived immunoglobulins and afterwards evokes tumor-specific adaptive immune feedback by the activation of tumor antigen-specific cytotoxic T lymphocytes [33]. Thus, cancer-derived O-glycan signatures have been highlighted as potential tumor-specific therapeutic targets, as well as biomarkers for clinical outcome and therapeutic response [10]. Main altered glycosylation features include: hypoglycosylation and exposition of specific tumor-associated carbohydrate antigens (TACAs), like prematurely terminated, short Tn or T antigens, their sialylated forms as well as fucosylated Lewis antigens [8,9,23,27,34,35]. Such truncated O-glycans are observed especially at the earliest stages of cellular malignant transformation [29]. The accurate molecular mechanism by which abnormal glycosylation is able to induce adverse metabolic and cellular signaling leading towards tumor progression, is still a subject of survey.
Recently, an emphasis has been put down on examining the effects of various natural compounds at different steps of cancer development. One of such compounds is rosmarinic acid, plant polyphenol, frequently found in Laminaceae and Boraginaceae families [36]. There are some reports revealing anti-cancer action of RA in colon cancer cells [37]. It has been demonstrated that rosmarinic acid induced apoptosis, inhibited metastatic potential, regulated EMT, suppressed MMPs, mainly by AMPK activation [38], by p38 MAPK/AP-1 signaling [39] or by inhibiting ERKs phosphorylation [40,41,42]. Other authors evidenced anti-inflammatory action of RA by inhibition of the expression of COX-2 [43,44]. Targeting Warburg metabolism by rosmarinic acid has been also indicated as promising action of this acid in colon cancer treatment [45,46]. However, to date, very little attention has been given to the influence of rosmarinic acid on glycoforms of colon cancer cells. Thus, the examining the impact of RA on specific carbohydrate structures in HT-29 and DLD-1 colon cancer cells, was the main aim of our study. Our concept was based on formerly received, promising results concerning anti-cancer action of rosmarinic acid in AGS gastric cancer cells [20,21].
In the presented study we revealed that rosmarinic acid had potential to inhibit MUC1 expression as well as some specific cancerous glycoforms in colon cancer cells DLD-1 and HT-29. We postulate that such action of RA can be considered as kind of therapeutic approach.
It has been reported that truncated Tn and sialyl Tn antigens are highly expressed in colon carcinomas. They appear to be colon-cancer associated antigens acting as markers of poorly differentiated cancers [34,47]. Yamamoto et al. [48] have been suggested that Tn carbohydrate antigen overexpression directly promoted oncogenic hallmark including enhanced cell proliferation, decreased apoptosis, increased adhesion as well as migration capacities. The authors have reported recently that enhanced Tn expression coincided with intensified tumor growth, particularly at later steps of tumor development. Sialyl-Tn antigen has been also described to promote tumor progression. In gastric cancer cells, this epitope has been reported to modulate cell cycle, apoptosis, adhesion, cell-cell aggregation and invasion [48,49]. Apart from that, sialyl Tn antigen in bladder cancer, has been described to be related with high-grade tumors, invasion, poor survival, tumor dissemination and metastasis [50]. We revealed that RA had the potential to inhibit enzymes responsible for Tn and sialyl Tn antigens formation, ppGalNAcT and ST6GalNAcT mRNA, respectively. However, the expression of Tn antigen was suppressed mainly in HT-29 cells in culture medium.
T antigen, highly expressed in carcinomas, is associated with invasiveness, tumor growth, and high metastatic potential. It is said to play a key role in the adhesion of cancer cells to the endothelium via interaction with galectin-3, and by that promoting metastasis [51]. It has been reported that T antigens and Gal-3 are positively associated with colon cancer metastasis [11,52]. In colon cancer, MUC1 is often decorated with T epitopes [53]. It has been revealed that association between Gal-3 and T antigen on MUC1 can lead to EGFR related carcinogenesis, cancer progression as well as cell homotypic aggregation what results in promoting of cancer cells survival by inhibition of cellular anoikis [25]. C1GalT1, favored by molecular chaperone COSMC, is enzyme responsible for T antigen formation. It is considered as the critical enzyme implicated in O-glycans synthesis [11,54]. C1GalT1 expression is upregulated in most cancers, what is firmly associated with malignant behavior of tumors, including proliferation, invasion or tumor spread [55]. Our results concerning T and Tn antigens, as well as enzymes responsible for their production, prove the potential of rosmarinic acid in anti-cancer therapy, as we revealed the suppression of the most mentioned factors.
The next, crucial in tumorigenesis carbohydrate structures are the blood group fucosylated Lewis antigens. Fucose containing glycans and enzymes implicated in fucosylation process are involved in gaining the functional features of cancer cells such as unlimited cell progression, tissue invasion, metastasis or immune evasion [56]. It has been reported that such fucosylated antigens can serve as ligands for e.g., E-, P-selectins of vascular endothelial cells and as result of such potential interactions, could promote extravasation and metastasis of cancer cells [12,21]. In our study we demonstrate inhibitory effect of rosmarinic acid on FUT3 and FUT4 in both examined cell lines as well as Lewis structures in HT-29 cells. Such action of RA allows to treat it as attractive candidate for anticancer agent.
Inhibiting of Akt expression and stimulating of p53 by RA also confirm its validity in cancer treatment. It is well documented that overexpression of phosphorylated Akt is among major factors initiating tumor development and allowing resistance to the conventional chemotherapy in various cancers [57]. p53, a powerful tumor suppressor is considered as a crucial barrier against cancer initiation and progression. It reveals its powerful action by controlling the cell cycle, apoptosis, DNA repair as well as angiogenesis [58]. Thus, the agents suppressing Akt, triggering p53, like rosmarinic acid, are among potential candidates in supporting of cancer treatment.
4. Materials and Methods
4.1. Cell Culture
DLD-1 (CCL-221) and HT-29 (HTB-38) human colorectal adenocarcinoma cells were obtained from American Type Culture Collection (ATCC, Manassas, VA, USA). DLD-1 cells were cultured in RPMI 1640 medium (ATCC, Manassas, VA, USA), and HT-29 cells in McCoy’s 5a medium (Pan Biotech., Aidenbach, Lower Bavaria, Germany) at 37°C, 5% CO2 in humidified air. Both media were supplemented with Fetal Bovine Serum (10%) (FBS; Gibco, Waltham, MA, USA), streptomycin (100 μg/mL), and penicillin (100 U/mL) (Sigma, St. Louis, MO, USA). Cells were seeded into 6-well plates in growth medium (1 mL), grown to 70% confluency, and then used for the further analyses. Dimethyl sulfoxide (DMSO; Sigma, St. Louis, MO, USA) was applied to dissolve rosmarinic acid (Roth, Karsruhe, Germany). Stock solution of RA was 111 mM. The cells were cultured with for 24 h in proper FBS-free medium with 200 and 400 μM RA. Then the cells were washed with Phosphate Buffered Saline (PBS; Corning, Manassas, VA, USA) and lysed with RIPA buffer (Sigma, St. Louis, MO, USA) for 20 min at 4oC with Protease Inhibitor Cocktail (Sigma, St. Louis, MO, USA) diluted 1:200. Next, intense vortexing and centrifugation at 1,000 x g for 5 min at 4°C were performed. The supernatants of cell lysates and culture media were collected, frozen at -70°C and used for further analyses. BCA Protein Assay Kit (Pierce, USA) was applied for the measurements of protein concentration. For quantitative real-time PCR, the wells were washed 3-times with sterile PBS (10 mM), collected and sonified (Sonics Vibra cell; Sonics & Materials, Leicestershire, UK) (10 W, 3-times for 15 s on ice). For RNA isolation, aliquots of the homogenate were used.
4.2. Cell Viability Test
The measurement of the viability of the cultured colon cancer DLD-1 and HT-29 cells, in the presence of rosmarinic acid (50 – 400 μM), were assessed using were assayed using 3-(4,5-dimethylthiazole-2-yl)-2,5-diphenyltetrazolium bromide (MTT) (Sigma, St. Louis, MO, USA) according to the procedure of Carmichael et al. [59]. Briefly, cells were cultured in six-well plates to get 70% of confluency and incubated for 24 h with different concentration of rosmarinic acid. Next, 1 mL of MTT solution (0.5 mg MTT/mL PBS) was added to each well and incubation at 37 oC in 5% CO2 for 4 h was performed. The absorbance of the converted dye was read at 570 nm. The viability of colon cells with RA was determined as a percentage of control cells without RA (100% cell viability).
4.3. Western Blotting
To detect the expression of C1GalT1, Gal-3, pAkt,, p53 proteins in cell lysates and MUC1 extracellular domain in culture media, electrophoresis on polyacrylamide gels (7.5 – 13%) and Western blotting assays were carried out. The samples containing 20 μg of protein, diluted in probe sample buffer with 2.5% SDS (Sigma, St. Luis, MO, USA) were subjected to electrophoresis and transferred to on Immobilon P (Millipore, Bedford, MA, USA) according to Towbin et al. [60]. 5% skim milk in Tris Buffered Saline (TBS) supplemented with 0.05% Tween 20 (Sigma, St. Luis, MO, USA) was used to block the membranes (after Western blotting) for 1 h at room temperature (RT). Then the membranes were washed with TBS-T and incubated with specific primary antibodies (listed in Table 1) overnight at 4 oC. Proper horseradish peroxidase-conjugated secondary antibodies were applied to detect immunoreactive complexes. To visualize the protein bands, enhanced chemiluminescence procedure with Westar Hypernova, ECL substrate for Western blotting (Cyangen, Bologna, Italy) was performed. The Gene Tools program (Syngene, Frederick, MD, USA) was used to quantify densitometrically the intensity of the bands (normalized for β-actin).
4.4. ELISA
To demonstrate the relative levels of carbohydrate antigens in cell lysates and culture media, ELISA tests with specific biotinylated lectins (Vector, Burlingame, CA, USA) were applied (Table 2). The procedure was performed according to previously described [61]. Briefly, 50 μL of cell lysates or culture medium (100 μg protein/mL) were applied on Microtiter plates (NUNC F96 Maxisorp, Roskilde, Denmark) and incubated overnight at RT. Then, blocking reagent for ELISA (Roche Diagnostics, Mannheim, Germany) was used for 1h at RT. As washing buffer, PBS with 0.05% Tween (100 μL) was applied after every step. Next, the plate was incubated with biotinylated lectins diluted to 5 μg/mL (2h at RT). Then, horseradish peroxidase avidin D (Vector, Burlingame, CA, USA), with high affinity to biotin, was used (1h at RT). As the final step, 100 μL of ABTS (2,2’-azino-bis(3-ethylbenzthiazoline-6-sulfonic acid) (Sigma, St. Luis, MO, USA) was applied to expand the colored reaction. Absorbances were read at 405 nm after 20 – 40 min of incubation at RT. The samples were examined in triplicate.
4.5. RNA Isolation and Quantitative Rreal-Time PCR
Total RNA was isolated applying Total RNA Mini Plus Concentrator (A&A Biotechnology, Gdansk, Poland), according to the instruction. Purity and concentration of RNA was examined spectrophotometrically (Nanodrop 2000, Thermo Scientific, Waltham, MA, USA). The same amounts (1 μg) of total RNA were submitted to reverse transcription using the SensiFASTTM cDNA Synthesis Kit (Bioline, London, UK). The reaction mixture (20 μL) contained RNA template, 1 μl of Reverse Transcriptase, 4 μl of 5xTransAmp Buffer, and DEPC-treated water. The conditions of incubation were 10 min at 25°C, 30 min at 45°C, and 5 min at 70°C were. Amplification of cDNA was carried out using SensiFAST™ SYBR Kit (Bioline, London, UK) in the thermocycler CFX96 real-time system (BioRad, Hercules, CA, USA). Reaction mixture (20 μL) contained 2 μL of cDNA template (diluted 3 times), 0.4 μL of each target-specific primer (10 μmol/L) (Genomed, Warsaw, Poland) (Table 3), 2×SensiFAST SYBR No-ROX Mix (5 μL), and DEPC-treated water. As a reference gene β2-Microglobulin was applied. The qRT-PCR parameters were as follows: 95°C for 2 min (activation of DNA polymerase), followed by 40 cycles of 10 s at 95°C (denaturation), 15 s at 60°C (annealing) and 20 s at 72°C (elongation). Each sample was examined in triplicate. The formation of the reaction products was proved by studies of their melting curves. The ∆∆Ct method was used to normalize the levels of target gene transcripts to β2-Microglobulin.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S3B: The effect of rosmarinic acid on MUC1 extracellular domain expression – original images; Figure S5B: The effect of RA on C1GalT1 protein expression – original images; , Figure S7B: The effect of rosmarinic acid on Gal-3 protein expression – original images; Figure S8B: The effect of RA on pAkt expression – original images; Figure S9B: The effect of RA on p53 protein expression – original images.
Author Contributions
Conceptualization, I.R., K.C., A.G.; methodology, I.R., K.C., B.P.; software, K.C.; validation, I.R., A.G.; formal analysis, K.C., B.P.; investigation, I.R.; resources, I.R..; data curation, I.R.; writing-original draft preparation, I.R.; writing-review and editing, I.R., A.G.; visualization, K.C.; supervision, I.R.; project administration, I.R.; funding acquisition, I.R. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by Medical University of Białystok (Grant No B.SUB.24.170).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data available on request. The data presented in this study are available on request from the corresponding author.
Acknowledgments
We thank J. Wosek for technical support.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Xi, Y.; Xu, P. Global colorectal cancer burden in 2020 and projections to 2040. Transl. Oncol. 2021, 14, 101174. [Google Scholar] [CrossRef]
- Islam, M.R.; Akash, S.; Rahman, M.M.; Nowrin, F.T.; Akter, T.; Shohag, S.; Rauf, A.; Aljohani, A.S.M.; Simal-Gandara, J. Colon cancer and colorectal cancer: Prevention and treatment by potential natural products. Chem. Biol. Interact. 2022, 368, 110170. [Google Scholar] [CrossRef]
- Sanchez-Guillen, L.; Arroyo, A. Immunonutrition in patients with colon cancer. Immunotherapy 2020, 12, 5–8. [Google Scholar] [CrossRef] [PubMed]
- Esmeeta, A.; Adhikary, S.; Dharshanaa, V.; Swarnamughi, P.; Maqsummiya, Z.U.; Banerjee, A.; Pathak, S.; Duttaroy, A.K. Plant-derived bioactive compounds in colon cancer treatment: An updated review. Biomed. Pharmacother. 2022, 153, 113384. [Google Scholar] [CrossRef] [PubMed]
- Fabregas, J.C.; Ramnaraign, B.; George, T.J. Clinical updates for colon cancer care in 2022. Clin. Colorectal Cancer 2022, 21, 198–203. [Google Scholar] [CrossRef]
- Benarba, B.; Pandiella, A. Colorectal cancer and medicinal plants: Principle findings from recent studies. Biomed. Pharmacother. 2018, 107, 408–423. [Google Scholar] [CrossRef]
- Hasibuan, P.A.Z.; Simanjuntak, Y.; Hey-Hawkins, E.; Lubis, M.F.; Rohani, A.S.; Park, M.N.; Kim, B.; Syahputra, R.A. Unlocking the potential of flavonoids: natural solutions in the fight against colon cancer. Biomed. Pharmacother. 2024, 176, 116827. [Google Scholar] [CrossRef]
- Zhang, L.; Hagen, K.G.T. Pleiotropic effects of O-glycosylation in colon cancer. J. Biol. Chem. 2018, 293, 1315–1316. [Google Scholar] [CrossRef]
- Stowell, S.R.; Ju, T.; Cummings, R.D. Protein glycosylation in cancer. Annu. Rev. Pathol. 2015, 10, 473–510. [Google Scholar] [CrossRef]
- Magalhaes, A.; Duarte, H.O.; Reis, C.A. The role of O-glycosylation in human disease. Mol. Aspects Med. 2021, 79, 100964. [Google Scholar] [CrossRef] [PubMed]
- Thomas, D.; Rathinavel, A.K.; Radhakrishnan, P. Altered glycosylation in cancer: A promising target for biomarkers and therapeutics. Biochim. Biophys. Acta Rev. Cancer 2021, 1875, 188464. [Google Scholar] [CrossRef] [PubMed]
- Bangargh, R.; Khatana, C.; Kaur, S.; Sharma, A.; Kaushal, A.; Siwal, S.S.; Tuli, H.S.; Dhama, K.; Thakur, V.K.; Saini, R.V.; Saini, A.K. Aberrant protein glycosylation: Implications on diagnosis and immunotherapy. Biotechnol. Adv. 2023, 66, 108149. [Google Scholar]
- Hauselmann, I.; Borsig, L. Altered tumor-cell glycosylation promotes metastasis. Front. Oncol. 2014, 4, 28. [Google Scholar] [CrossRef] [PubMed]
- Cox, K.E.; Liu, S.; Lwin, T.M.; Hoffmann, R.M.; Batra, S.K.; Bouvet, M. The mucin family of proteins: Candidates as potential biomarkers for colon cancer. Cancers 2023, 15, 1491. [Google Scholar] [CrossRef]
- Wang, H.S.; Wang, L.H. The expression and significance of Gal-3 and MUC1 in colorectal cancer and colon cancer. OncoTargets Ther. 2015, 8, 1893–1898. [Google Scholar] [CrossRef]
- Konstantinou, E.K.; Panagiotopoulos, A.A.; Argyri, K.; Panoutsopoulos, G.I.; Dimitrou, M.; Gioxari, A. Molecular pathways of rosmarinic acid anticancer activity in triple-negative breast cancer cells: A literature review. Nutrients 2024, 16, 2. [Google Scholar] [CrossRef]
- Chaitanya, M.V.N.L.; Ramanunny, A.K.; Babu, M.R.; Gulati, M.; Viskwas, S.; Singh, T.G.; Chellappan, D.K.; Adams, J.; Dua, K.; Singh, S.K. Journey of rosmarinic acid as biomedicine to nano-biomedicine for treating cancer: Current strategies and future perspectives. Pharmaceutics 2022, 14, 2401. [Google Scholar] [CrossRef]
- Mahmoud, M.A.; Okda, T.M.; Omran, G.A.; Abd-Alhaseeb, M.M. Rosmarinic acid suppresses inflammation, angiogenesis, and improves paclitaxel induced apoptosis in a breast cancer model via NF3 κB-p53-caspase-3 pathways modulation. J. Appl. Biomed. 2021, 19, 202–209. [Google Scholar] [CrossRef]
- Jin, B.R.; Chung, K.S.; Hwang, S.; Hwang, S.N.; Rhee, K.J.; Lee, M.; An, H.J. Rosmarinic acid represses colitis-associated colon cancer: A pivotal involvement of the TLR4-mediated NF-κB-STAT3 axis. Neoplasia 2021, 23, 561–573. [Google Scholar] [CrossRef] [PubMed]
- Radziejewska, I.; Supruniuk, K.; Bielawska, A. Anti-cancer effect of combined action of anti-MUC1 and rosmarinic acid in AGS gastric cancer cells. Eur. J. Pharmacol. 2021, 902, 174119. [Google Scholar] [CrossRef]
- Radziejewska, I.; Supruniuk, K.; Nazaruk, J.; Karna, E.; Popławska, B.; Bielawska, A.; Galicka, A. Rosmarinic acid influences collagen, MMPs, TIMPs, glycosylation and MUC1 in CRL-1739 gastric cancer cell line. Biomed. Pharmacol. 2018, 107, 397–407. [Google Scholar] [CrossRef] [PubMed]
- Holst, S.; Wuhrer, M.; Rombouts, Y. Glycosylation characteristics of colorectal cancer. Adv. Cancer Res. 2015, 126, 203–256. [Google Scholar] [PubMed]
- Cascio, S.; Finn, O.J. Intra- and Extra-Cellular Events Related to Altered Glycosylation of MUC1 Promote Chronic Inflammation, Tumor Progression, Invasion, and Metastasis. Biomolecules 2016, 6, 39. [Google Scholar] [CrossRef]
- Hollingsworth, M.A.; Swanson, B.J. Mucins in cancer: Protection and control of the cell surface. Nat. Rev. Cancer 2004, 4, 45–60. [Google Scholar] [CrossRef]
- Beckwith, D.M.; Cudic, M. Tumor-associated O-glycans of MUC1: Carriers of the glycol-code and targets for cancer vaccine design. Semin. Immunol. 2020, 47, 101389. [Google Scholar] [CrossRef]
- He, M.; Zhou, X.; Wang, X. Glycosylation: mechanisms, biological functions and clinical implications. Signal Transduct. Target Ther. 2024, 9, 194. [Google Scholar] [CrossRef]
- Kvorjak, M.; Ahmed, Y.; Miller, M.L.; Sriram, R.; Coronello, C.; Hashash, J.G.; Hartmann, D.J.; Telmer, C.A.; Miskov-Zivanov, N.; Finn, O.J.; Cascio, S. Crosstalk between colon cells and macrophages increases ST6GALNAC1 and MUC1-sTn expression in ulcerative colitis and colitis–associated colon cancer. Cancer Immunol. Res. 2020, 8, 167–178. [Google Scholar] [CrossRef]
- Duncan, T.J.; Watson, N.F.; Al-Attar, A.H.; Scholefield, J.H.; Durrant, L.G. The role of MUC1 and MUC3 in the biology and prognosis of colorectal cancer. World J. Surg. Oncol. 2007, 5, 31. [Google Scholar] [CrossRef]
- Berois, N.; Pittini, A.; Osinaga, E. Targeting tumor glycans for cancer therapy: Successes, limitations, and perspectives. Cancers 2022, 14, 645. [Google Scholar] [CrossRef]
- Horm, T.M.; Schroeder, J.A. MUC1 and metastatic cancer: Expression, function and therapeutic targeting. Cell Adh. Migr. 2013, 7, 187–198. [Google Scholar] [CrossRef]
- Nath, S.; Mukherjee, P. MUC1: a multifaced oncoprotein with a key role in cancer progression. Trends Mol. Med. 2014, 20, 332–342. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Zhang, Z.; Zhang, S.; Zhu, P.; Ko, J.K.S.; Yung, K.K.L. MUC1: Structure, function, and clinic application in epithelial cancers. Int. J. Mol. Sci. 2021, 22, 6567. [Google Scholar] [CrossRef]
- Taylor-Papadimitriou, J.; Burchell, J.M.; Graham, R.; Beatson, R. Latest developments in MUC1 immunotherapy. Biochem. Soc. Trans. 2018, 46, 659–668. [Google Scholar] [CrossRef]
- Krishn, S.R.; Kaur, S.; Smith, L.M.; Johansson, S.L.; Jain, M.; Patel, A.; Gautam, S.K.; Hollingsworth, M.A.; Mandel, U.; Clausen, H.; Lo, W.C.; Fan, W.T.L.; Manne, U.; Batra, S.K. Mucins and associated glycan signatures in colon adenoma-carcinoma sequence: Prospective pathological implication(s) for early diagnosis of colon cancer. Cancer Lett. 2016, 374, 304–314. [Google Scholar] [CrossRef]
- Brockhausen, I. Mucin-type O-glycans in human colon and breast cancer: glycodynamics and functions. EMBO Rep. 2006, 7, 599–604. [Google Scholar] [CrossRef]
- Guan, H.; Luo, W.; Bao, B.; Cao, Y.; Cheng, F.; Yu, S.; Fan, Q.; Zhang, L.; Wu, Q.; Shan, M. A comprehensive review of rosmarinic acid: From phytochemistry to pharmacology and its new insight. Molecules 2022, 27, 3292. [Google Scholar] [CrossRef] [PubMed]
- Czerwińska, K.; Radziejewska, I. Rosmarinic acid: A potential therapeutic agent in gastrointestinal cancer management – a review. Int. J. Mol. Sci. 2024, 25, 11704. [Google Scholar] [CrossRef]
- Zhao, J.; Xu, L.; Jin, D.; Xin, Y.; Tian, L.; Wang, T.; Zhao, D.; Wang, Z.; Wang, J. Rosmarinic acid and related dietary supplements: Potential applications in the prevention and treatment of cancer. Biomolecules 2022, 12, 1410. [Google Scholar] [CrossRef]
- Yang, K.; Shen, Z.; Zou, Y.; Gao, K. Rosmarinic acid inhibits migration, invasion, and p38/AP-1 signaling via miR-1225-5p in colorectal cancer cells. J. Recept. Signal Transduct. Res. 2021, 41, 284–293. [Google Scholar] [CrossRef]
- Xu, Y.; Xu, G.; Liu, L.; Xu, D.; Liu, L. Anti-invasion effect of rosmarinic acid via the extracellular signal-regulated kinase and oxidation-reduction pathway in Ls174-T cells. J. Cell. Biochem. 2010, 111, 370–379. [Google Scholar] [CrossRef]
- Xavier, C.P.R.; Lima, C.F.; Fernandes-Ferreira, M.; Pereira-Wilson, C. Induction of apoptosis and inhibition of proliferation in colon cancer cells by Salvia fruticosa, Salvia officinalis and rosmarinic acid. Planta Med. 2008, 74, PA19. [Google Scholar] [CrossRef]
- Xavier, C.P.R.; Lima, C.F.; Fernandes-Ferreira, M.; Pereira-Wilson, C. Salvia fruticosa, Salvia officinalis, and rosmarinic acid induce apoptosis and inhibit proliferation of human colorectal cell lines: The role in MAPK/ERK pathway. Nutr. Cancer 2009, 61, 564–571. [Google Scholar] [CrossRef]
- Scheckel, K.A.; Degner, S.C.; Romagnolo, D.F. Rosmarinic acid antagonizes activator protein-1–dependent activation of cyclooxygenase-2 expression in human cancer and nonmalignant cell lines. J. Nutr. 2008, 138, 2098–2105. [Google Scholar] [CrossRef]
- Liu, H.; Deng, R.; Zhu, C.W.; Han, H.K.; Zong, G.F.; Ren, L.; Cheng, P.; Wei, Z.H.; Zhao, Y.; Yu, S.Y.; et al. Rosmarinic acid in combination with ginsenoside Rg1 suppresses colon cancer metastasis via co-inhition of COX-2 and PD1/PD-L1 signaling axis. Acta Pharmacol. 2024, 45, 193–208. [Google Scholar] [CrossRef] [PubMed]
- Zhong, X.; He, X.; Wang, Y.; Hu, Z.; Huang, H.; Zhao, S.; Wei, P.; Li, D. Warburg effect in colorectal cancer: The emerging roles in tumor microenvironment and therapeutic implications. J. Hematol. Oncol. 2022, 15, 160. [Google Scholar] [CrossRef]
- Xu, Y.; Han, S.; Lei, K.; Chang, X.; Wang, K.; Li, Z.; Liu, J. Anti-Warburg effect of rosmarinic acid via miR-155 in colorectal carcinoma cells. Eur. J. Cancer Prev. 2016, 25, 481–489. [Google Scholar] [CrossRef]
- Liu, Z.; Liu, J.; Dong, X.; Hu, X.; Jiang, Y.; Li, L.; Du, T.; Yang, L.; Wen, T.; An, G.; Feng, G. Tn antigen promotes human colorectal cancer metastasis via H-Ras mediated epithelial-mesenchymal transition activation. J. Cell. Mol. Med. 2019, 23, 2083–2092. [Google Scholar] [CrossRef]
- Yamamoto, D.; Hongo, H.; Kosaka, T.; Aoki, N.; Oya, M.; Sato, T. The sialyl-Tn antigen synthase genes regulate migration-proliferation dichotomy in prostate cancer cells under hypoxia. Glycoconj. J. 2023, 40, 199–212. [Google Scholar] [CrossRef] [PubMed]
- Pinho, S.; Marcos, N.T.; Ferreira, B.; Carvalho, A.S.; Oliveira, M.J.; Santos-Silva, F.; Harduin-Lepers, A.; Reis, C.A. Biological significance of cancer-associated sialyl-Tn antigen: Modulation of malignant phenotype in gastric carcinoma cells. Cancer Lett. 2007, 249, 157–170. [Google Scholar] [CrossRef]
- Wilczak, M.; Surman, M.; Przybyło, M. Altered glycosylation in progression and management of bladder cancer. Molecules 2023, 28, 3436. [Google Scholar] [CrossRef]
- Fu, C.; Zhao, H.; Wang, Y.; Cai, H.; Xiao, Y.; Zeng, Y.; Chen, H. Tumor-associated antigens: Tn antigen, sTn antigen, and T antigen. HLA 2016, 88, 275–286. [Google Scholar] [CrossRef]
- Sindrewicz, P.; Lian, L.Y.; Yu, L.G. Interaction of the oncofetal Thomsen Friedenreich antigen with galectins in cancer progression and metastasis. Front. Oncol. 2016, 6, 79. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.G.; Andrews, N.; Zhao, Q.; McKean, D.; Williams, J.F.; Connor, L.J.; Gerasimenko, O.V.; Hilkens, J.; Hirabayashi, J.; Kasai, K.; Rhodes, J.M. Galectin-3 interaction with Thomsen-Friedenreich disaccharide on cancer-associatedMUC1 causes increased cancer cell endothelial adhesion. J. Biol. Chem. 2007, 282, 773–781. [Google Scholar] [CrossRef]
- Wan, Y.; Adair, K.; Herrmann, A.; Shan, X.; Xia, L.; Duckworth, C.A.; Yu, L.G. C1GalT1 expression reciprocally controls tumour cell-cell and tumour-macrophage interactions mediated by galectin-3 and MGL with double impact on cancer development and progression. Cell Death Dis. 2023, 14, 547. [Google Scholar] [CrossRef] [PubMed]
- Xia, T.; Xiang, T.; Xie, H. Update on the role of C1GALT1 in cancer (Review). Oncol. Lett. 2022, 23, 97. [Google Scholar] [CrossRef]
- Shan, M.; Yang, D.; Dou, H.; Zhang, L. Chapter four – fucosylation in cancer biology and its clinic applications. Prog. Mol. Biol. Transl. Sci. 2019, 162, 93–119. [Google Scholar] [PubMed]
- Revathidevi, S.; Munirajan, A.K. Akt in cancer: Mediator and more. Semin. Cancer Biol. 2019, 59, 80–91. [Google Scholar] [CrossRef]
- Bhatia, R.; Kumar, B. p53 as a potential target for treatment of cancer: A perspective on recent advancements in small molecules with structural insights and SAR studies. Eur. J. Med. Chem. 2023, 247, 115020. [Google Scholar]
- Carmichael, J.; Degraff, W.; Gazdar, A.; Minna, J.; Mitchell, J. Evaluation of tetrazolium-based semi-automated colorimetric assay: Assessment of chemosensitivity testing. Cancer Res. 1987, 47, 936–942. [Google Scholar]
- Towbin, T.; Stachelin, T.; Gordon, J. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: procedure and some applications. Proc. Natl. Acad. Sci. USA 1979, 76, 4350–4354. [Google Scholar] [CrossRef]
- Radziejewska, I.; Supruniuk, K.; Jakimiuk, K.; Tomczyk, M.; Bielawska, A.; Galicka, A. Tiliroside combined with anti-MUC1 monoclonal antibody as promising anti-cancer strategy in AGS cancer cells. Int. J. Mol. Sci. 2023, 24, 13036. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
T Structure of rosmarinic acid.
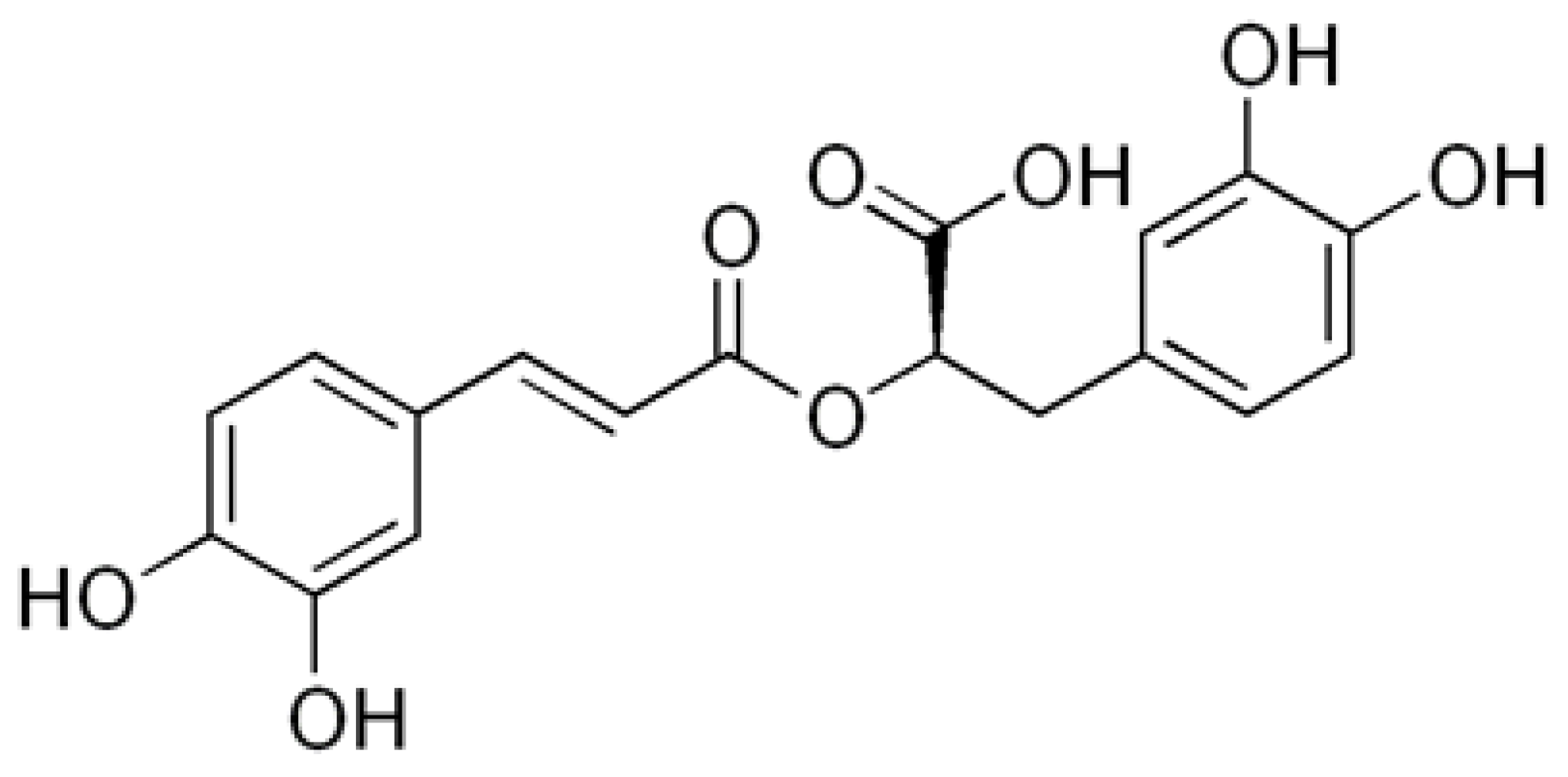
Figure 2.
Viability of DLD-1 and HT-29 colon cancer cells treated for 24 h with 0 – 400 μM concentrations of rosmarinic acid. Mean values ±SD are the mean of triplicate culture.
Figure 2.
Viability of DLD-1 and HT-29 colon cancer cells treated for 24 h with 0 – 400 μM concentrations of rosmarinic acid. Mean values ±SD are the mean of triplicate culture.
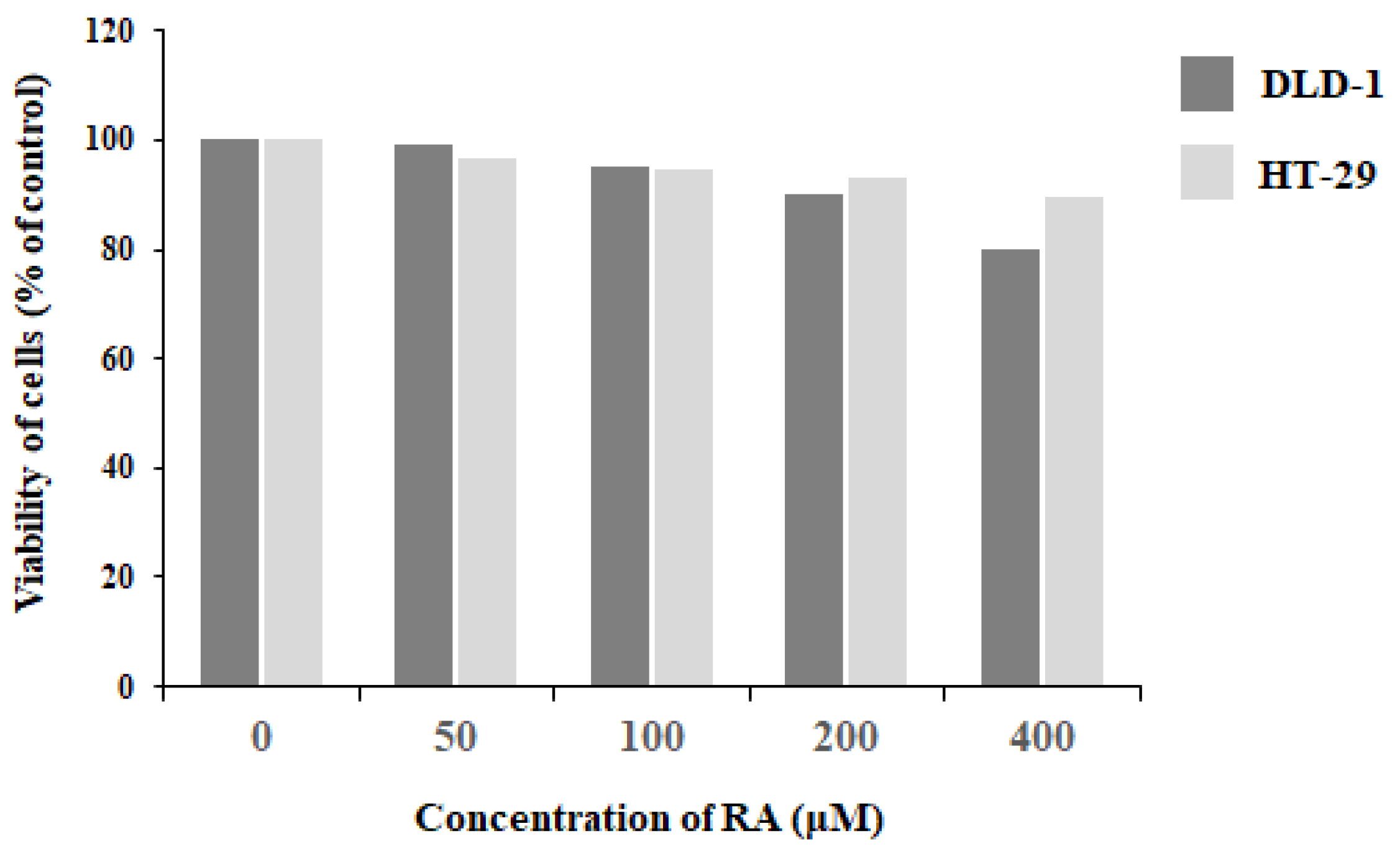
Figure 3.
The effect of rosmarinic acid on MUC1 mRNA and MUC1 extracellular domain expression in DLD-1 and HT-29 colon cancer cells. The cells were incubated for 24 h with 200 and 400 μM RA. mRNA was determined by qRT-PCR (A). The results are presented as a relative fold change in mRNA expression of gene in comparison to the gene in control, where expression was set at 1. ± SD are the mean of triplicate cultures. **p < 0.01, ***p < 0.001. MUC1 extracellular domain in culture medium was assessed by Western blotting (B). The intensities of the bands were quantified by densitometric study. Data represent the mean ± SD of triplicate culture. *p < 0.05, **p < 0.01 compared to control.
Figure 3.
The effect of rosmarinic acid on MUC1 mRNA and MUC1 extracellular domain expression in DLD-1 and HT-29 colon cancer cells. The cells were incubated for 24 h with 200 and 400 μM RA. mRNA was determined by qRT-PCR (A). The results are presented as a relative fold change in mRNA expression of gene in comparison to the gene in control, where expression was set at 1. ± SD are the mean of triplicate cultures. **p < 0.01, ***p < 0.001. MUC1 extracellular domain in culture medium was assessed by Western blotting (B). The intensities of the bands were quantified by densitometric study. Data represent the mean ± SD of triplicate culture. *p < 0.05, **p < 0.01 compared to control.
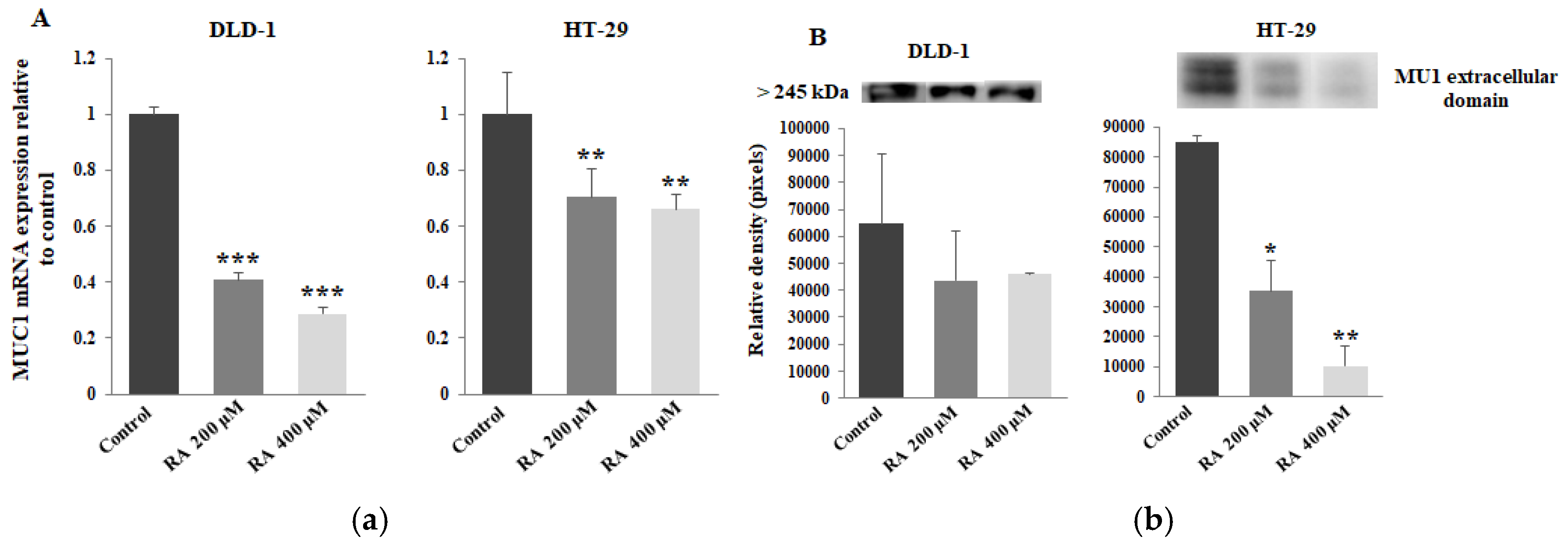
Figure 4.
The effect of RA on ppGalNAcT2 mRNA, ST6GalNAcT1 mRNA, Tn and sialyl Tn antigen expression in cell lysates and culture medium. DLD- and HT-29 colon cancer cells were incubated for 24 h with 200 and 400 μM rosmarinic acid. ppGalNAcT2 mRNA (A) and ST6GalNAcT1 mRNA (D) were assessed by qRT-PCR. The results are presented as a relative fold change in mRNA expression of gene in comparison to the gene in control, where expression was set at 1. ± SD are the mean of triplicate cultures. *p < 0.05, **p < 0.01, ***p < 0.001. Tn and sialyl Tn antigens relative expressions were determined by ELISA tests with specific, biotinylated lectins. VVA was applied to assess Tn antigen (B,C) and SNA was used to assess sialyl Tn (E,F). The results are expressed as absorbance at 405 nm after reactivity with lectins. Values ± SD are the mean from three independent analysis. *p < 0.05, **p < 0.01, ***p < 0.001 compared to untreated control.
Figure 4.
The effect of RA on ppGalNAcT2 mRNA, ST6GalNAcT1 mRNA, Tn and sialyl Tn antigen expression in cell lysates and culture medium. DLD- and HT-29 colon cancer cells were incubated for 24 h with 200 and 400 μM rosmarinic acid. ppGalNAcT2 mRNA (A) and ST6GalNAcT1 mRNA (D) were assessed by qRT-PCR. The results are presented as a relative fold change in mRNA expression of gene in comparison to the gene in control, where expression was set at 1. ± SD are the mean of triplicate cultures. *p < 0.05, **p < 0.01, ***p < 0.001. Tn and sialyl Tn antigens relative expressions were determined by ELISA tests with specific, biotinylated lectins. VVA was applied to assess Tn antigen (B,C) and SNA was used to assess sialyl Tn (E,F). The results are expressed as absorbance at 405 nm after reactivity with lectins. Values ± SD are the mean from three independent analysis. *p < 0.05, **p < 0.01, ***p < 0.001 compared to untreated control.
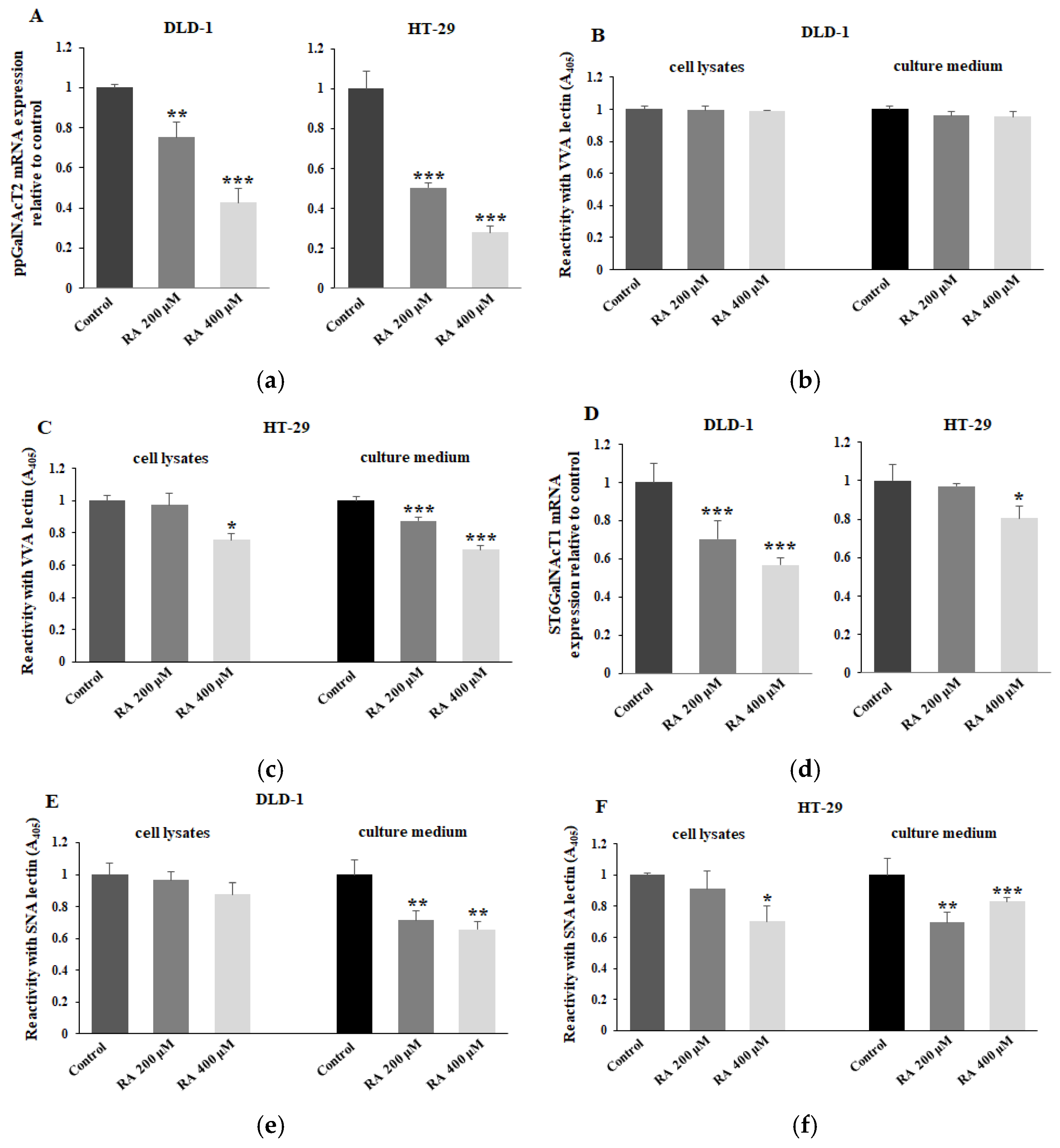
Figure 5.
The effect of rosmarinic acid on C1GalT1 mRNA, C1GalT1 protein, Cosmc mRNA, ST3GalT1 mRNA, T and sialyl T antigens in cell lysates and culture medium of DLD-1 and HT-29 colon cancer cells. The cancer cells were incubated for 24 h with 200 and 400 μM RA. C1GalT1 (A), Cosmc (C), and ST3GalT1 (F) mRNAs were determined by qRT-PCR. The results are presented as a relative fold change in mRNA expression of gene in comparison to the gene in control, where expression was set at 1. ± SD are the mean of triplicate cultures. *p < 0.05, **p < 0.01, ***p < 0.001. C1GalT1 protein (B) was assessed by Western blotting. β-actin served as a protein loading control. The bands were analyzed by densitometric study. Data represent the mean ± SD of triplicate culture. *p < 0.05, ***p < 0.001 compared to control. T and sialyl T antigens expressions were determined by ELISA tests with biotinylated lectins (PNA for T antigen (D,E) and MAAII for sialyl T (G,H)). The results are expressed as absorbance at 405 nm after reactivity with lectins. Values ± SD are the mean from three independent analysis. *p < 0.05, **p < 0.01, ***p < 0.001 compared to untreated control.
Figure 5.
The effect of rosmarinic acid on C1GalT1 mRNA, C1GalT1 protein, Cosmc mRNA, ST3GalT1 mRNA, T and sialyl T antigens in cell lysates and culture medium of DLD-1 and HT-29 colon cancer cells. The cancer cells were incubated for 24 h with 200 and 400 μM RA. C1GalT1 (A), Cosmc (C), and ST3GalT1 (F) mRNAs were determined by qRT-PCR. The results are presented as a relative fold change in mRNA expression of gene in comparison to the gene in control, where expression was set at 1. ± SD are the mean of triplicate cultures. *p < 0.05, **p < 0.01, ***p < 0.001. C1GalT1 protein (B) was assessed by Western blotting. β-actin served as a protein loading control. The bands were analyzed by densitometric study. Data represent the mean ± SD of triplicate culture. *p < 0.05, ***p < 0.001 compared to control. T and sialyl T antigens expressions were determined by ELISA tests with biotinylated lectins (PNA for T antigen (D,E) and MAAII for sialyl T (G,H)). The results are expressed as absorbance at 405 nm after reactivity with lectins. Values ± SD are the mean from three independent analysis. *p < 0.05, **p < 0.01, ***p < 0.001 compared to untreated control.
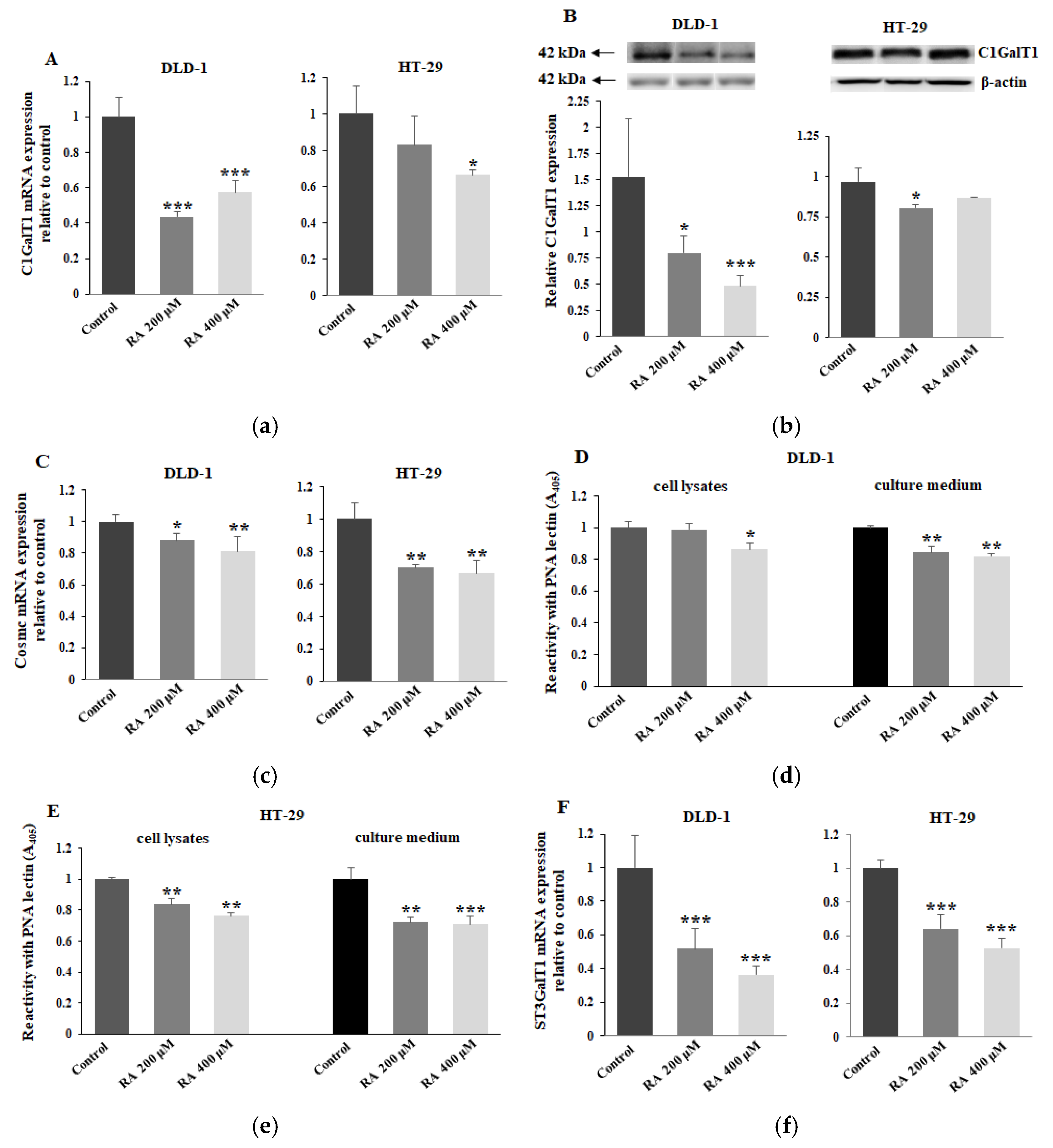
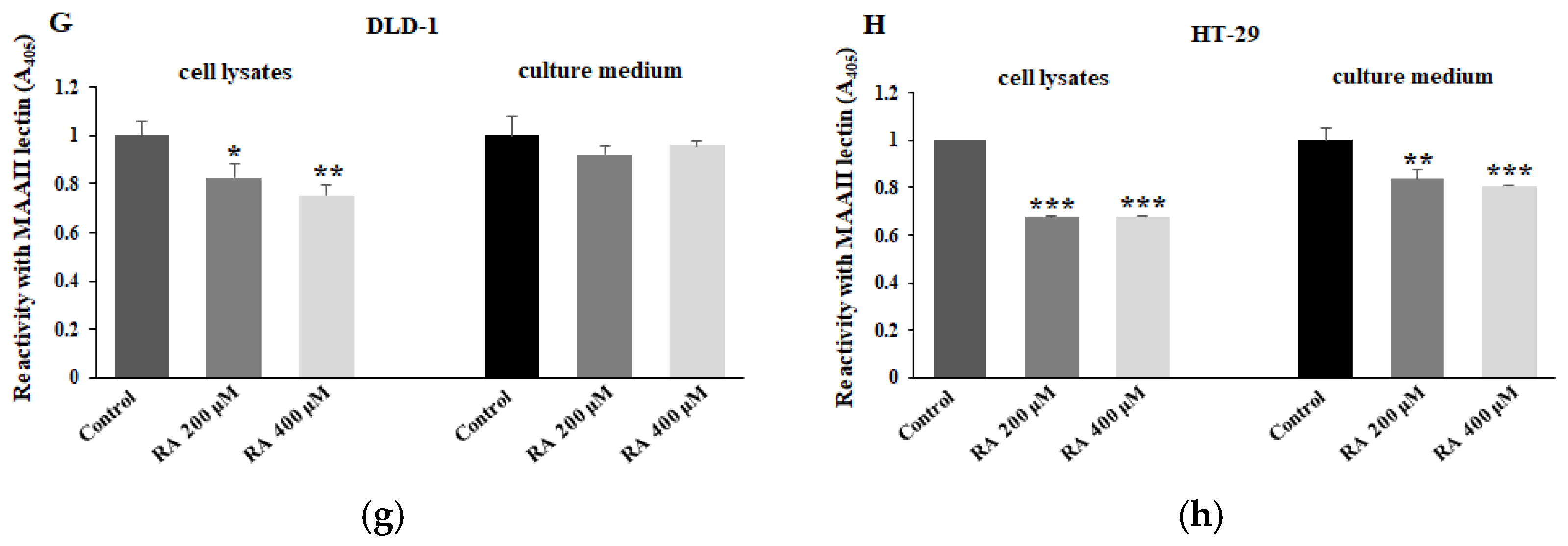
Figure 6.
The effect of rosmarinic acid on FUT3, FUT4 mRNAs and Fucα1,3-GalNAc antigen. DLD-1 and HT-29 colon cancer cells were incubated for 24 h with 200 and 400 μM RA. FUT3 (A) and FUT4 (B) mRNAs were assessed by qRT-PCR analysis. The results are presented as a relative fold change in mRNA expression of gene in comparison to the gene in control, where expression was set at 1. ± SD are the mean of triplicate cultures. ***p < 0.001. Fucα1,3-GalNAc antigen expression in cell lysates and culture medium was determined by ELISA test with biotinylated LTA lectin (C,D). The results are expressed as absorbance at 405 nm after reactivity with lectin. Values ± SD are the mean from three independent analysis. **p < 0.01, ***p < 0.001 compared to untreated control.
Figure 6.
The effect of rosmarinic acid on FUT3, FUT4 mRNAs and Fucα1,3-GalNAc antigen. DLD-1 and HT-29 colon cancer cells were incubated for 24 h with 200 and 400 μM RA. FUT3 (A) and FUT4 (B) mRNAs were assessed by qRT-PCR analysis. The results are presented as a relative fold change in mRNA expression of gene in comparison to the gene in control, where expression was set at 1. ± SD are the mean of triplicate cultures. ***p < 0.001. Fucα1,3-GalNAc antigen expression in cell lysates and culture medium was determined by ELISA test with biotinylated LTA lectin (C,D). The results are expressed as absorbance at 405 nm after reactivity with lectin. Values ± SD are the mean from three independent analysis. **p < 0.01, ***p < 0.001 compared to untreated control.
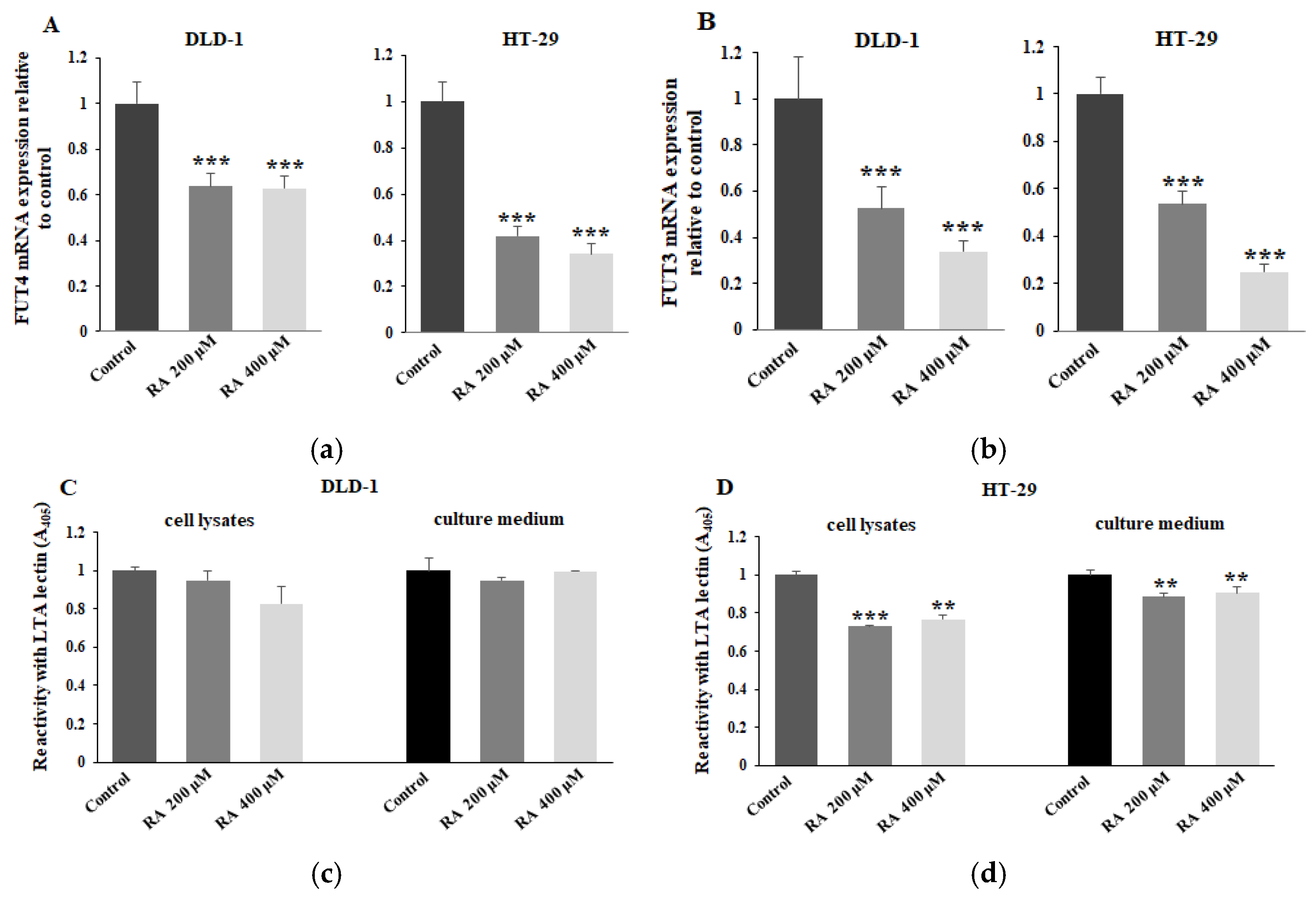
Figure 7.
The effect of rosmarinic acid on Gal-3 mRNA and Gal-3 protein expression in DLD-1 and HT-29 colon cancer cells. Cancer cells were incubated for 24 h with 200 and 400 μM RA. Gal-3 mRNA (A) was assessed by qRT-PCR analysis. The results are presented as a relative fold change in mRNA expression of gene in comparison to the gene in control, where expression was set at 1. ± SD are the mean of triplicate cultures. *p < 0.05, ***p < 0.001. Gal-3 protein in cell lysates (B) was determined by Western blotting. β-actin served as a protein loading control. The bands were analyzed by densitometric study. Data represent the mean ± SD of triplicate culture. *p < 0.05, ***p < 0.001 compared to control.
Figure 7.
The effect of rosmarinic acid on Gal-3 mRNA and Gal-3 protein expression in DLD-1 and HT-29 colon cancer cells. Cancer cells were incubated for 24 h with 200 and 400 μM RA. Gal-3 mRNA (A) was assessed by qRT-PCR analysis. The results are presented as a relative fold change in mRNA expression of gene in comparison to the gene in control, where expression was set at 1. ± SD are the mean of triplicate cultures. *p < 0.05, ***p < 0.001. Gal-3 protein in cell lysates (B) was determined by Western blotting. β-actin served as a protein loading control. The bands were analyzed by densitometric study. Data represent the mean ± SD of triplicate culture. *p < 0.05, ***p < 0.001 compared to control.
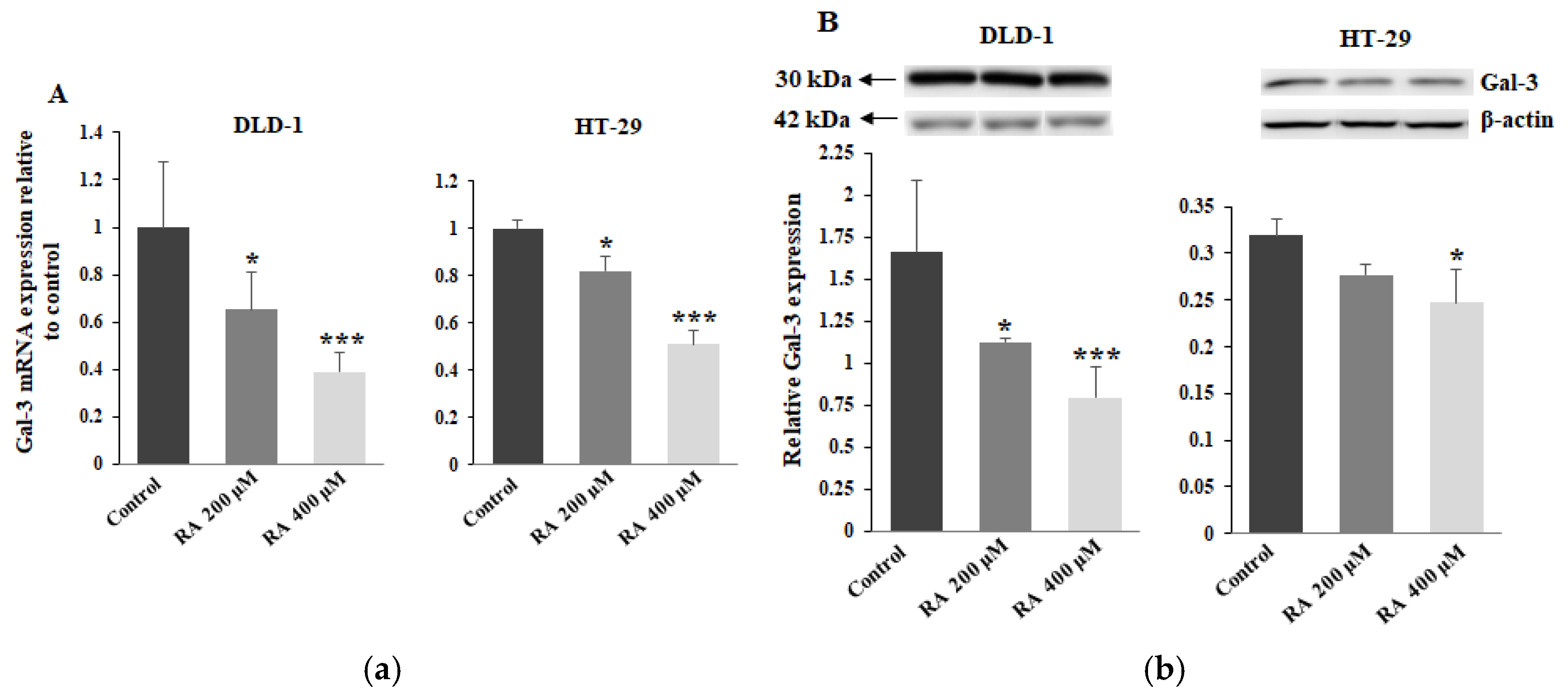
Figure 8.
The effect of RA on Akt mRNA and pAkt protein expression in DLD-1 and HT-29 colon cancer cell lines. Cancer cells were incubated for 24 h with 200 and 400 μM RA. Akt mRNA (A) was analyzed by qRT-PCR analysis. The results are shown as a relative fold change in mRNA expression of gene in comparison to the gene in control, where expression was set at 1. ± SD are the mean of triplicate cultures. **p < 0.01, ***p < 0.001. pAkt in cell lysates (B) was determined by Western blotting. β-actin served as a protein loading control. The bands were assessed by densitometric study. Data represent the mean ± SD of triplicate culture. *p < 0.05, compared to control.
Figure 8.
The effect of RA on Akt mRNA and pAkt protein expression in DLD-1 and HT-29 colon cancer cell lines. Cancer cells were incubated for 24 h with 200 and 400 μM RA. Akt mRNA (A) was analyzed by qRT-PCR analysis. The results are shown as a relative fold change in mRNA expression of gene in comparison to the gene in control, where expression was set at 1. ± SD are the mean of triplicate cultures. **p < 0.01, ***p < 0.001. pAkt in cell lysates (B) was determined by Western blotting. β-actin served as a protein loading control. The bands were assessed by densitometric study. Data represent the mean ± SD of triplicate culture. *p < 0.05, compared to control.
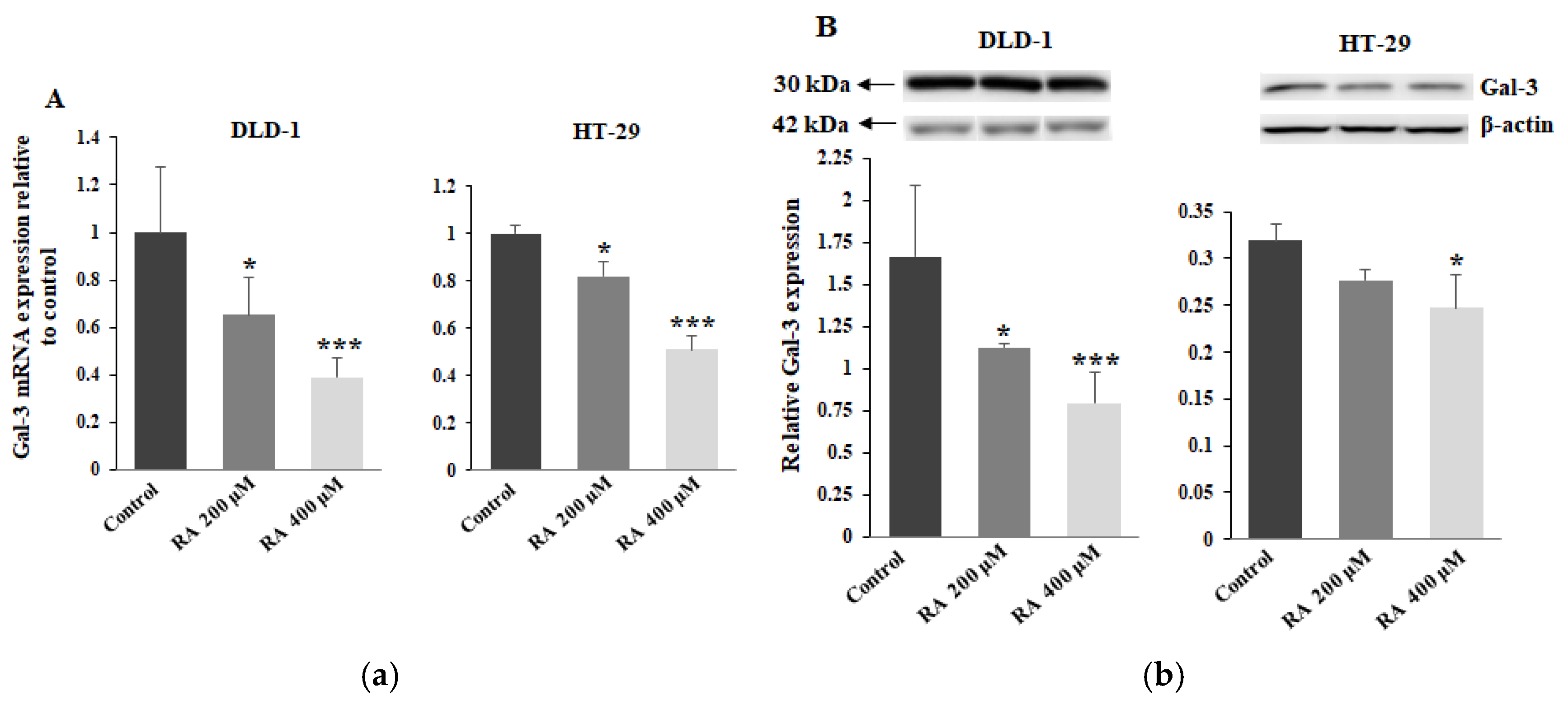
Figure 9.
The effect of RA on p53 mRNA and protein expression in DLD-1 and HT-29 colon cancer cell lines. The cells were incubated for 24 h with 200 and 400 μM RA. p53 mRNA (A) was analyzed by qRT-PCR. The results are shown as a relative fold change in mRNA expression of gene in comparison to the gene in control, where expression was set at 1. ± SD are the mean of triplicate cultures. *p < 0.05, **p < 0.01. p53 in cell lysates (B) was determined by Western blotting. β-actin served as a protein loading control. The bands were assessed by densitometric study. Data represent the mean ± SD of triplicate culture. *p < 0.05, compared to control.
Figure 9.
The effect of RA on p53 mRNA and protein expression in DLD-1 and HT-29 colon cancer cell lines. The cells were incubated for 24 h with 200 and 400 μM RA. p53 mRNA (A) was analyzed by qRT-PCR. The results are shown as a relative fold change in mRNA expression of gene in comparison to the gene in control, where expression was set at 1. ± SD are the mean of triplicate cultures. *p < 0.05, **p < 0.01. p53 in cell lysates (B) was determined by Western blotting. β-actin served as a protein loading control. The bands were assessed by densitometric study. Data represent the mean ± SD of triplicate culture. *p < 0.05, compared to control.
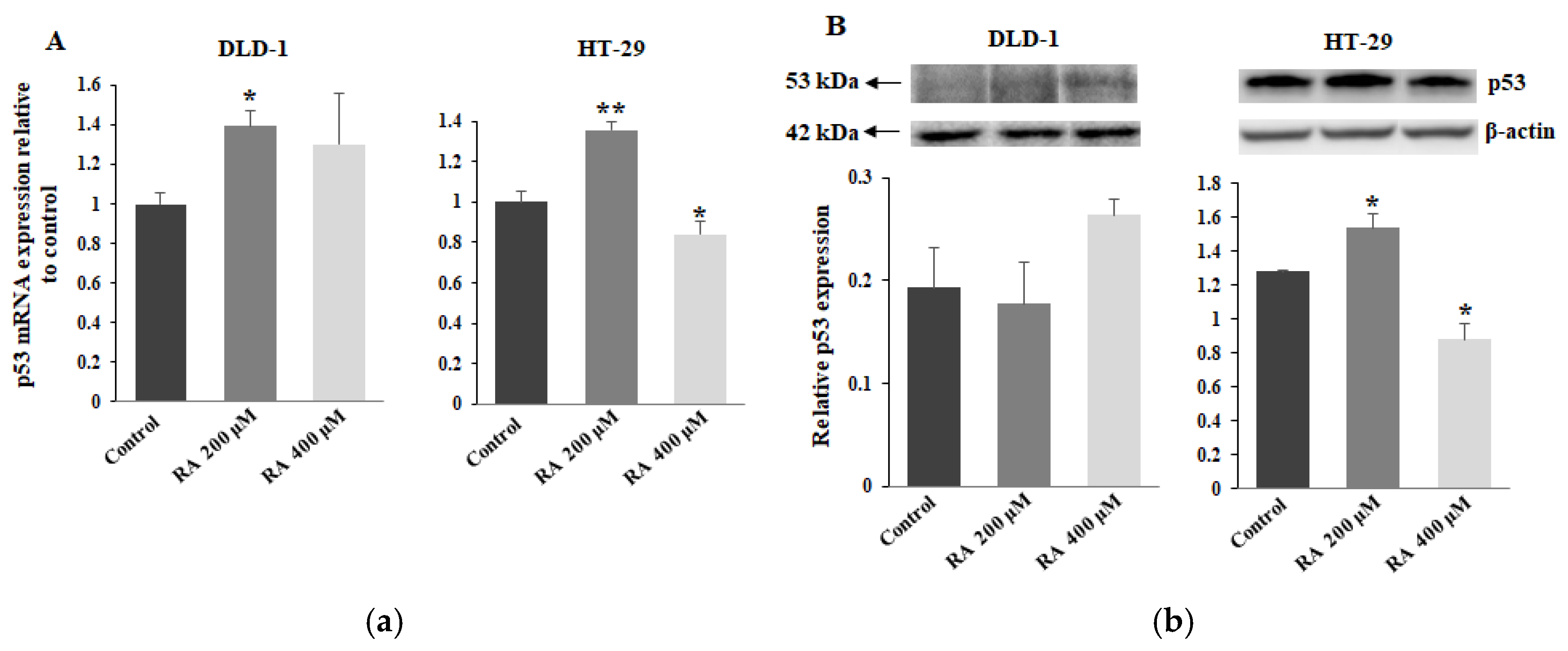

Table 1.
Antibodies used in electrophoresis and Western blotting.
| Antibody | Clone | Source |
|---|---|---|
| Anti-MUC1; extracellular domain (mouse IgG) Anti-C1GalT1 (mouse IgG) Anti-Gal-3 (mouse IgG) Anti-pAkt (rabbit IgG) Anti-p53 (rabbit IgG) Anti-β-actin (rabbit IgG) Anti-mouse IgG peroxidase conjugated Anti-rabbit IgG peroxidase conjugated |
BC2 F-31 B2C10 D9E 7F5 |
Abcam Santa Cruz Santa Cruz Cell Sign Tech Cell Sign Tech Sigma Sigma Sigma |
Table 2.
Biotinylated lectins used in ELISA tests.
| Lectin | Specificity |
|---|---|
| VVA (Vicia villosa agglutinin) SNA (Sambucus nigra agglutinin) PNA (Arachis hypogaea agglutinin (peanut)) MAAII (Maackia amurensis agglutinin) LTA (Lotus tetragonolobus agglutinin) |
Tn antigen (GalNAcα1-O-Ser/Thr) sialyl Tn antigen (NeuAcα2,6-Gal/GalNAc) T antigen (Galβ1,3-GalNAcα1-O-Ser/Thr) sialyl T antigen (NeuAcα2,3-Gal) Lewis structure (Fucα1,3-GlcNAc) |
Table 3.
Primers applied in quantitative Real-Time PCR.
| Gene | Forward primer (5’ → 3’) | Reverse primer (5’ → 3’) |
|---|---|---|
|
MUC1 C1GalT1 Cosmc ppGalNAcT2 ST3GalT1 ST6GalNAcT1 FUT3 FUT4 Gal-3 Akt p53 β2-Microglobulin |
ACAATTGACTCTGGCCTTCC CAAAATACGACCCTGAAGAACC GTTTGCCTGAAATATGCTGGA AAGAAAGACCTTCATCACAGCAATGGAGAA TCGGCCTGGTTCGATGA ACGCAGTCCTGAGGTTTAATGG GCCGACCGCAAGGTGTAC AAGCCGTTGAGGCGGTTT GCAGACAATTTTTCGCTCCATG ACTGTCATCGAACGCACCTT GTTCCGAGAGCTGAATGAGG TTTCTGGCCTGGAGGCTATC |
CAGGTTATATCGAGAGGCTGC GCATCTCCCCAGTGCTAAGTC AATATGCCCAAATGCCCTAAG ATCAAAACCGCCCTTCAAGTCAGCA CGCGTTCTGGGCAGTCA AGTTCATCAGGCGAATGGTAGTTT TGACTTAGGGTTGGACATGATATCC ACAGTTGTGTATGAGATTTGGAAGCT CTGTTGTTCTCATTGAAGCGTG CTCCTCCTCCTCCTGCTTCT TCTGAGTCAGGCCCTTCTCT CATGTCTCCATCCCACTTAACT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



