
Submitted:
01 August 2023
Posted:
03 August 2023
You are already at the latest version
Abstract
(1) Background: Colon cancer is one of the leading causes of cancer morbidity and mortality globally. It is a multi-step process that involves genetic and epigenetic modifications leading to histological and morphological changes. Several complementary therapeutic options have been analyzed, shedding light on plant-based medication as a potential treatment for colon cancer. Flavonoids such as quercetin are known to have anti-cancer and anti-inflammatory properties. This in vitro study examines quercetin's anti-inflammatory, anti-apoptotic, anti-angiogenesis effects and antioxidant properties in colon cancer cells. (2) Methods: The antioxidant capacity of quercetin-treated cells was investigated using biochemical assays, and angiogenesis and cytokine levels were assessed using enzyme-linked immunosorbent assay (ELISA). The epigenetic modulation and differential expression of aging, apoptotic, and proliferation genes, and Histone deacetylases (HDACs) were also investigated. (3) Results: In this study, the quercetin-treated group significantly reduced the antioxidant enzymes, cytokines, and VEGF levels, altering the expression of epigenetic factors. Quercetin also induced significant senescence in colon cancer cells. Moreover, a considerable increase was observed in the apoptotic and hTERT genes. In contrast, a decrease in p53, proliferation genes, and HDACs was observed, providing a basis for the clinical use of quercetin in cancer treatment. (4) Conclusion: In vitro studies showed that quercetin treatment efficiently induces senescence and apoptosis in colon cancer cells. We also found that quercetin effectively modulated the expression of p53, Wnt1, CTNNB1, and HDACs, indicating that it could be used to treat colon cancer.
Keywords:
quercetin
; antioxidant
; apoptotic
; anti-inflammatory activity
; colon cancer
1. Introduction
Colon cancer is known to be one of the most common malignancies, leading up to 1.15 million deaths in 2020 globally [1]. Age, race, personal history, intestinal inflammatory condition, western diet, alcohol consumption, smoking, and obesity are considerable threats to colon cancer [2]. However, oxidative stress, inflammation, and angiogenesis are typical hallmarks of colon cancer [3]. Several treatment approaches, such as surgery, chemotherapy, radiation therapy, targeted drug therapy, combinational therapy, and immunotherapies, are presently accessible for colon cancer [4]. Unfortunately, these therapies have low tumor sensitivity and higher toxicity, which reduces overall therapy effectiveness. The interplay of inflammation in colon cancer is well-established by many epigenetic and pharmacological analyses [5]. Inflammatory cytokines promote tumor cell growth, interrupt differentiation, and aid in cancer cell survival [6]. Inflammation is mediated by cytokines released by tumor cells, suggesting that targeting cytokines may be helpful for both preventive and therapeutic strategies for colon cancer [7]. However, cancer treatment is often ineffective as tumors spread to distant organs. As a result, the discovery of novel therapeutic approaches remains a critical goal in the ongoing fight against colon cancer. Since ancient times, studies have attempted to examine plant-based drugs to treat various diseases as it has fewer side effects [8,9]. Recent research has discovered many plant-based polyphenol compounds with beneficial pharmacological properties and therapeutic value, including anti-tumor activity. Flavonoids are a type of dietary polyphenol widely studied to reduce the risk of cancer development, particularly colon cancer [10]. Evidence supports that polyphenol family members can influence cell proliferation, cellular differentiation, and apoptosis-related gene expression by modulating the associated signaling pathways [11]. One commonly used polyphenol is quercetin which is well known for its anti-proliferative, antioxidant, and anti-inflammatory effects, which make it act as an anti-cancerous agent through broad-spectrum mechanisms from anti-carcinogenesis to anti-metastasis [12,13]. Quercetin can also regulate signaling pathways associated with cellular proliferation and inflammation cascades, including the PI3K, NFκB, MAPK, Wnt pathway, and protein kinase B (Akt). Cytotoxic activity of colon cancer cells is mediated by inhibition of NFκB and induction of apoptosis via p53 activation, suppression of inflammation via downregulation of Cox2, which is commonly upregulated in colon cancer, and cell cycle arrest due to downregulation of cell cycle genes. Another way quercetin may affect colon cancer cell proliferation is by increasing the expression of the G-protein coupled cannabinoid receptor, CB1-R, which can then bind to quercetin, inhibiting cell growth and migration via the Wnt, Akt, PI3K, and STAT3 pathways. In the existence of a CB1-R antagonist, this effect was abrogated [14]. The mechanisms by which flavonoids may decrease colon carcinogenesis are not entirely understood. However, they have been linked to antioxidant, pro-apoptotic, and anti-inflammatory properties. As a result, more research is needed to determine the association between quercetin and colon cancer. Our previous in vitro study demonstrated quercetin's anti-cancer activity and epigenetic regulations in normal, primary, and metastatic colon cancer cells [11]. In this in vitro study, we further focused on analyzing anti-oxidant, anti-inflammatory, apoptosis-induction, and anti-angiogenesis properties of quercetin in normal, primary, and metastatic colon cancer cell lines. Additionally, we have analyzed the effect of quercetin on epigenetic alterations by analyzing Histone deacetylase (HDAC) expression in colon cancer cells. Therefore, this study focused on further detailed parameters validating quercetin's benefits, particularly on the modulation of HDACs and related epigenetic modulators.
2. Materials and Methods
2.1. Chemicals and Reagents
Quercetin and DMSO was acquired from Sigma-Aldrich, USA. Fetal Bovine Serum (FBS), Dulbecco’s Modified Eagle’s Medium (DMEM), phosphate buffer saline (PBS), Trypsin and antibiotic–antimycotic solution (penicillin, amphotericin B, streptomycin was acquired from Gibco, Thermo Fisher Scientific, USA.
2.2. Cell culture
The following colon cell lines were utilized in this study: L132 (Normal lung epithelial cell), HCT 116, Colo 320 (Primary colon cancer cell), and Colo 205 (Metastatic colon cancer cell), all of which were acquired from NCCS, Pune, India. The normoxic condition (37 °C with 5% CO2) was used to maintain the cell lines cultured in DMEM and supplemented with 10% FBS and 1% antibiotic.
2.3. Preparation of quercetin stock solution
20mM Quercetin stock was prepared based on our previous study and stored at 4 °C [11].
2.4. Cell viability analysis
2.4.1. Cell Counting Kit-8 (CCK-8) assay
Based on the method reported in our previous study [11], the cell viability assay was performed using a CCK-8 assay kit (Sigma-Aldrich, USA) for the normal, primary, and metastatic cell lines post-treatment with quercetin. 2×103 cells were seeded into 96-well plates and maintained under normoxic conditions for 24 h. After 24 h, the medium was discarded, and fresh medium was added, along with various concentrations of quercetin (Q20µM, Q40µM, Q80µM, Q120 µM, Q160 µM, Q320µM) and kept for 72 h. Post 72 h of treatment, 10 µl of the CCK-8 reagent was added to every well and kept for 4 h. At 450 nm, the absorbance was analyzed utilizing an ELISA plate reader [11]. Finally, the IC50 value is determined, and the graph was plotted (data not shown).
2.4.2. Experimental groups:
After dose determination, the following experimental groups were taken forward in the present study, L132- (Control, LQ80µM, LQ120 µM), HCT116- (Control, HQ80µM, HQ120 µM), Colo320-(Control, CQ80µM, CQ120 µM) and Colo 205- (Control, CQ80µM, CQ120µM). All the control groups were supplemented with DMEM+10%FBS +DMSO (solvent), and two further groups for each cell line were treated with 80µM and 120µM quercetin concentrations.
2.5. Assessment of the antioxidant activity
2.5.1. Sample preparation
4×104 cells were treated with the preferred doses of quercetin (80µM and 120 µM) for 72 h. After treatment, the cell lysate (the treated cells and control cells were centrifuged at 5000 rpm for 10 mins and the supernatant was collected) for all the cell lines and stored at -20 ºC for the following analysis.
2.5.2. Lipid peroxidase activity
The lipid peroxidase (LPO) assay was analyzed using the spectrophotometric method with minor alterations [15]. The cell lysate (1 mL) was mixed thoroughly with TCA-TBA-HCL buffer (2 mL) in a centrifuge tube and heated for 15 min in a boiling water bath. After cooling, centrifugation was done at 1000 rpm for 10 minutes to remove the precipitate. The absorbance was measured at 535nm using Milli-Q water as a “blank.”
2.5.3. Catalase activity
The catalase activity was evaluated utilizing the spectrophotometric procedure with minimal modification [16]. A reaction mixture comprising phosphate buffer (50mM), H2O2 (15mM), was prepared to which 0.1mL of cell lysate was added. The optical density (O.D.) at 240nm was measured over three minutes at intervals of 30 seconds.
2.5.4. Superoxide dismutase activity
The Superoxide dismutase (SOD) activity was examined using the spectrophotometric method with a minor alteration [17]. 1mL assay mixture was prepared by adding sodium phosphate buffer (0.05 M, pH 8.0), Ethylenediaminetetraacetic acid (EDTA) (0.01M), pyrogallol (0.27 mM solution of pyrogallol was made in 100mM HCl) and cell lysate. The O.D. was analyzed at 420 nm against Tris-EDTA buffer at the start of the experiment and after 1 min of the addition of pyrogallol.
2.5.5. Total thiol content activity
The reaction mixture was prepared by adding 0.1mL Sulfosalicylic acid 4%, 0.5mL sodium phosphate buffer (0.1M). Then, 0.5mL of reaction mixture was incubated for 75 min with 0.65mL of sample and 6.6 µl DTNB at room temperature (RT). After incubation, the test solution vortexed for 15 sec, and the absorbance was measured at 412 nm.
2.6. Assessment of free radical scavenging activity
2.6.1. ABTS free radical scavenging activity
The method for ABTS scavenging activity was followed as per the protocol with slight alterations [18]. A mixture of potassium persulfate (2.45 mM) and the ABTS (7mM, Cat. No. 40157, SRL, India) was prepared as a stock and were in a (1:1, v/v) ratio and maintained in the dark for 16 h at RT. Then, the stock solution was diluted by adding 1 mL of ABTS stock solution in methanol (60 mL), with spectrophotometric absorbance of 0.706 ± 0.01 units at 734 nm. The test samples from various treated groups (10μl) were incubated with diluted ABTS solution (200μl) for 7 min, and the absorbance was taken at 734 nm.
2.6.2. DPPH free radical scavenging activity
DPPH (Cat. No. D9132, Sigma-Aldrich, USA). DPPH stock solutions were prepared in a methanol buffer. Methanol buffer was made by combining 40 mL of 0.1M acetate buffer (0.1 M, pH 5.5) with 60 mL methanol. 100µl cell lysate was incubated with DPPH stock solution in the dark for 30 min at 30 °C. The O.D. was measured spectrophotometrically at 517 nm.
2.7. Senescence-associated β-galactosidase (SA-β-gal) activity
The SA-β-gal analysis uses β-galactosidase to convert X-gal (5-bromo-4-chloro-3-indolyl β-D-galactopyranoside), a colorless substrate, into a blue product that accumulates in senescent cells due to upregulation of β-galactosidase activity. The EZdetectTM cell senescence detection kit (Cat. No- CCK063, HiMedia Ltd, India) was used to measure the activity of SA-β-gal in control and treated cells. The procedure was followed exactly as directed by the manufacturer. The cell senescence was observed, and images were acquired utilizing an inverted microscope.
2.8. Gene expression analysis by qPCR
For extracting the total RNA (L132, HCT116, Colo305, Colo205), the cells were treated with TRIzolTM Reagent (Invitrogen, Thermo Fisher Scientific, Cat. No. 15596026). The purity and concentration of isolated RNA were quantified using a spectrophotometer at 260 nm. cDNA was generated utilizing a reverse transcription kit (Cat. No. RT-RTCK-03, Eurogentec, Belgium). The Syber Green qPCR master mix (Cat.NO. UF- RSMT-B0701, Takyon, Eurogentec, Belgium) was used to perform the qPCR for the following genes (p53, Bax, Bcl-2, CTNNB1(β-catenin), Wnt-1 and GAPDH). ∆Ct values and fold change for each gene were obtained by normalizing the Ct value of the target gene to GAPDH expression, and the graphs were plotted.
2.9. Epigenetic modification assays
2.9.1. HDAC quantification assay
TRIzolTM Reagent (Cat. No. 15596026, Invitrogen, Thermo Fisher Scientific) was utilized to isolate total RNA for HDAC quantification. The purity and concentration of isolated RNA were quantified using a spectrophotometer at 260 nm. cDNA was prepared using a reverse transcription kit (Cat. No. RT-RTCK-03, Eurogentec, Belgium). The RT2 SYBR Green ROX qPCR Mastermix (Cat. No. 330520, Qiagen, USA) and RT2 qPCR primer assay (HDAC1, HDAC2, HDAC4, HDAC5, HDAC8, HDAC10) (Cat. No. 330001, Qiagen, USA) were used to perform qPCR for the epigenetic regulator genes.
2.9.2. hTERT quantification assay
The hTERT expression was quantified utilizing the RT2 first-strand synthesis kit (Cat. No. 330401, Qiagen, USA) and RT2 qPCR hTERT primer assay (Cat. No. 330001, Qiagen, USA) kit by the manufacturer's protocol. The fold expression was analyzed, and graphs were plotted with statistical significance using GraphPad Prism V9.
2.10. Inflammatory cytokines and angiogenic factor expression analysis by ELISA
Protein quantification for the following, Interleukin-6 (IL6), Interleukin-10 (IL10), and Tumor necrosis factor (TNF-α) levels of control and quercetin-treated cells using an IL10 ELISA kit (Cat No. E0102Hu), IL6 ELISA Kit (Cat No. E0090Hu) and TNF-α ELISA Kit (Cat No. E0082Hu, Bioassay Technology Laboratory, China) was conducted by the manufacturer’s instructions.
2.11. Vascular Endothelial Growth Factor (VEGF) expression
The level of VEGF was quantified from the cell lysate obtained from the quercetin-treated cells utilizing the VEGF ELISA Kit (Cat No. E0080Hu, Bioassay Technology Laboratory, China) in accordance with the manufacturer’s instructions.
2.12. Statistical analysis
The GraphPad V9 software was utilized for statistical analysis. Employing the unpaired student’s t-test, the significant difference between the treatment and control groups was determined. Derived from three independent experimental studies, the data are expressed as mean ± standard error. P value =0.05 was considered statistically significant for the present study.
3. Results
3.1. Cell viability analysis
3.1.1. CCK-8 assay
The CCK-8 cell viability assay was carried out in control as well as normal lung epithelial cell, primary and metastatic colon cancer cells to identify and to calculate the IC50 value of quercetin, which has been concurred from our previous study, [11] the dose was determined and taken forward for the current study. Based on the results of the CCK8 assay, two doses of 80µM and 120 µM quercetin were taken for further treatments, which is explained in our previously published in vitro work (data not shown here) [11].
3.2. Assessment of antioxidant activity
3.2.1. Assessment of lipid peroxidation (LPO) assay
The lipid peroxidase activity is elucidated in Figure 1a. The results demonstrated that quercetin at a concentration of 80µM/mL showed decreased LPO activity in HCT116 (p=0.0101), Colo320 (p=0.0007), and Colo 205 respectively, while at a concentration of 120 µM/mL was significantly decreased in Colo320 (p=0.0003) and Colo205 (p=0.0105) when compared to their respective controls. Whereas HCT116 Q120µM (p=0.0011) was slightly increased compared to HCT116 Q80µM. (p=0.0002). Although no significant decrease in LPO level was observed in L132 (LQ80µM, p=0.0101, LQ120µM, p=0.0004) treated groups compared to their respective control group.
3.2.2. Assessment of catalase activity
The catalase activity is represented in Figure 1b. The results shows that catalase activity is significantly decrease in Q120µM of HCT116 (p=0.0034) and Colo 320 cell line (p=0.001) when compared to their respective control groups. Whereas in Q120µM of L132 (p=0.0299) and Q80µM of Colo 205 was seen to be increased compared to its control group.
3.2.3. Assessment of superoxide dismutase activity
The superoxide dismutase activity is demonstrated in Figure 1c. The results shows that SOD levels have significantly decreased in Q120 µM of HCT116 (p=0.0001) and Colo 205 (p=0.0002) cells, respectively, compared to that of their control groups. In L132 cells, both the treatment group showed a significant increase (LQ80µMp=0.0039, LQ120µM p=0.0003) in SOD activity compared to their respective control. Although in Colo 320 cells (CQ80µM p=0.0006, CQ120µM p=0.0006), there were reduced SOD levels, which did not have a significant difference between the treatment groups compared to their control group.
3.2.4. Assessment of total thiol content
The free thiol groups efficiently protect cells from free radical damage. The total thiol content is shown in Figure 1d. The results indicated that total thiol content had been significantly decreased in Colo 320 (CQ80µM, p=0.0001, CQ120µM, p=0.0001) and Colo 205 (CQ80µM, p=0.0001, CQ120µM, p=0.0001) cells when compared to their control groups. Although in HCT116 (HQ80µM, p=0.0001, HQ120µM, p=0.0001) cells thiol content was found to be reduced but did not have a significant difference between the treatment groups when compared to their control group. However, in L132 treated groups, quercetin did not affect the overall thiol content.
3.3. Assessment of free radical scavenging activity
3.3.1. Assessment of ABTS free radical scavenging activity
The ABTS free radical scavenging activity is represented in Figure 1e. Here, a significant inhibition in ABTS free radical scavenging activity in HCT116Q80µM and Q120µM treatment groups (p=0.001) was seen than in their control group. Likewise, there was a considerable reduction in free radical scavenging activity in Colo 320 (p=0.0008), but no changes were observed in Colo 205 groups. In contrast, quercetin administration significantly increased ABTS free radical scavenging capacity in L132 cells (p=0.0005).
3.3.2. Assessment of DPPH free radical scavenging activity
The DPPH scavenging activity is represented in Figure 1f. The DPPH activity was reduced in HCT116 (HQ120µM, p=0.0229) and Colo 320 (Q120µM, p=0.0017) respectively. In contrast, L132 showed an increased level of activity when compared to their control group. However, there was no change in Colo 205 group.
3.4. Assessment of senescence-associated β-galactosidase activity analysis
In SA- β-Gal cell senescence analysis, the number of senescent cells increased significantly in all cell lines upon quercetin treatment. In L132 (Q120 µM, p=0.0091) and HCT116 (Q120 µM, p=0.0005) Q120µM treatment group, the number of senescent cells was much higher than that of their control. Whereas in treated Colo 320 (Q80µM, p=0.0002, Q120µM, p=0.0001), a significantly large number of senescent cells were observed at both doses, Figure 2a. Similarly, a very high number of senescent cells was seen at both concentrations in Colo 205 (Q80µM, p=0.0002, Q120µM, p=0.0024). The senescent cells in each group have been indicated by arrows in Figure 2b. These findings suggest that Q120µM shows a significant increase in cell senescence.
3.5. Gene expression analysis by qPCR
3.5.1. Apoptosis-related and Tumor suppressor gene expression
In this study, quercetin therapy significantly enhanced Bax expression in HCT116 (HQ80µM, p=0.0054, HQ120µM, p=0.0001), Colo320 (CQ80µM, p=0.0016, CQ120µM, p=0.0103) and Colo205 (CQ80µM, p=0.0002, CQ120µM, p=0.0072), whereas there were no significant changes in L132 in comparison to their respective control groups, Figure 3a. In contrast, the Bcl-2 expression was drastically downregulated in all the cell lines with respect to their control groups; L132 (LQ80µM, p=0.0048, LQ120µM, p=0.0305), HCT116 (HQ80µM, p=0.0003, HQ120µM, p=0.0001), Colo320 (CQ80µM, p=0.0068, CQ120µM, p=0.0005) and Colo205 (CQ80µM, p=0.0002, CQ120µM, p=0.0002), Figure 3b. Likewise, the p53 gene was significantly upregulated in quercetin-treated HCT116 (HQ80µM, p=0.0050, HQ120µM, p=0.0031) and Colo 320 (CQ80µM, p=0.0021, CQ120µM, p=0.0009). Whereas no changes in the expression were observed in the Colo 205 and L132 cells, Figure 3c.
3.5.2. Proliferation-related gene expression
The impact of quercetin on primary cell proliferation was found to be more effective than the metastatic cell line. The proliferation-related gene Wnt1 was downregulated in HCT116 (HQ80µM, p=0.0015, HQ120µM, p=0.0006) and Colo320 (CQ80µM, p=0.0010, CQ120µM, p=0.0004) in both treated concentrations than the control group. Likewise, a higher dose of quercetin treatment significantly downregulated CTNNB1 expression in HCT116 (HQ80µM, p=0.0124, HQ120µM, p=0.0325), Colo320 (CQ120µM, p=0.0132) and Colo320 (CQ80µM, p=0.0323, CQ120µM, p=0.0006) cancer cell lines than the control group, respectively Figure 4 a, b. No statistically significant variation in gene expression was seen in either of the genes in L132 cells.
3.6. Epigenetic modification assays
3.6.1. HDAC Quantification assay
HDACs expression analysis was performed to investigate the impact of quercetin on class I HDACs (HDAC1, HDAC2, HDAC8) and class II HDACs (HDAC4, HDAC5, HDAC10), expression in colon cancer cells, where quercetin showed a significant decrease in HDACs expression. Interestingly, we observed that members of both HDACs family, class I and II were downregulated in quercetin-treated colon cancer cells. In brief, quercetin showed significant downregulation in the expression of HDAC1 in L132 (LQ80µM, p=0.0001, LQ120 µM, p = 0.0003), HCT116 (HQ80µM, p=0.0168, HQ120µM, p=0.0010), Colo320 (CQ80µM, p=0.0031, CQ120µM, p=0.034) treatment groups and Colo205 (CQ80µM, p=0.0031, CQ120µM, p=0.0096) cells at both the concentration, Figure 5a. Similarly, HDAC2 expression was reduced in quercetin-treated (L132Q120 µM, p=0.0051), (HCT116Q120 µM, p=0.0269) treatment group, whereas in Colo320 (Q80µM, p=0.0007, Q120 µM, p=0.0003), and Colo205 (Q80µM, p=0.0001, Q120µM, p=0.0001) cells at both the concentration, Figure 5b. In contrast, quercetin treatment reduced the HDAC8 expression in (HCT116Q80µM, p=0.0118), (Colo320Q120µM, p=0.0131), and (Colo205Q120µM, p=0.0007) treatment groups whereas no considerable differences were seen in L132 cells, Figure 5c.
Previous studies showed that HDAC4 plays a significant role in colon cancer maintenance and malignant transformation [19]. Here, quercetin-treatment groups showed a decrease in HDAC4 L132 (LQ120µM, p=0.0482), HCT116 (HQ80µM, p=0.0126, HQ120µM, p=0.0184), Colo320 (CQ80µM, p=0.0067, CQ120µM, p=0.0031) and Colo205 (CQ120µM, p=0.0040) expression in all the cell lines, Figure 6a. A similar pattern of expression inhibition was observed for HDAC10 L132 (LQ80µM, p=0.0001, LQ120µM, p=0.0003), HCT116 (HQ80µM, p=0.0168, HQ120 µM, p=0.0010), Colo320 (CQ80µM, p=0.0031, CQ120µM, p=0.0341) and Colo205 (CQ80µM, p=0.0031, CQ120µM, p=0.0096), Figure 6c. In contrast, HDAC 5 expression was observed to be downregulated in HCT116 (HQ80µM, p=0.0094, HQ120µM, p=0.0058) and Colo 320 (CQ80µM, p=0.0136, CQ120µM, p=0.0035) at both the doses whereas in L132 and Colo 205 cells whereas quercetin treatment did not show any significant changes, Figure 6b.
3.6.2. hTERT quantification assay
In this analysis, quercetin-treated groups showed significant downregulation in the expression of hTERT in (Colo320Q80µM, p=0.0065, Q120µM, p=0.458) and (Colo205Q80µM, p=0.0016) treatment group at a lower dose when compared to higher dose. In HCT116 (HQ80µM p=0.0077, HQ120µM, p=0.0089) quercetin significantly reduced the hTERT expression at both doses. Although a reduction in the hTERT expression was observed only in the L132Q120µM (p=0.0052) group when compared to L132Q80µM, Figure 7.
3.7. Inflammatory cytokines and angiogenic factor expression analysis
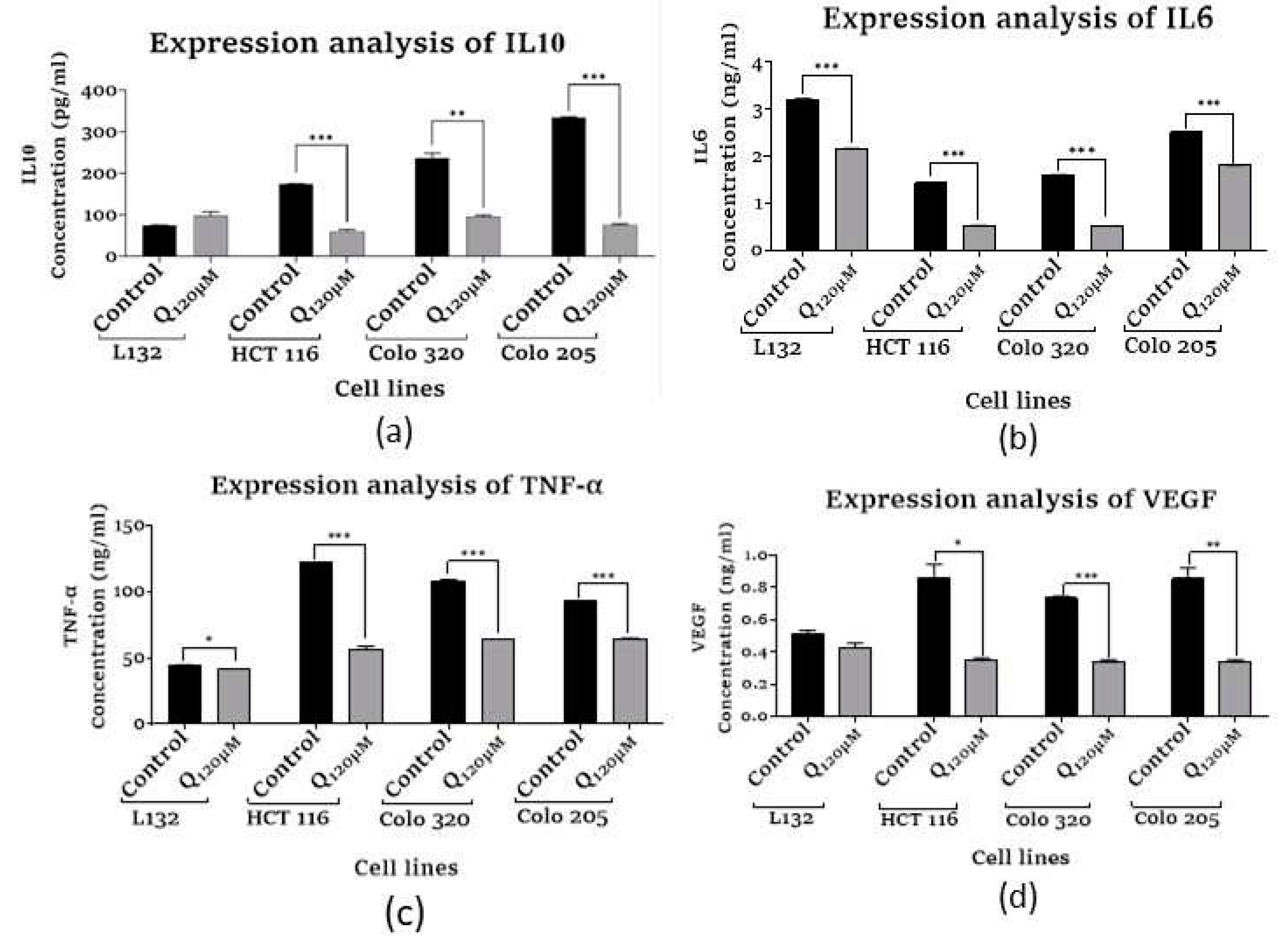
3.7.1. Assessment of Human Interleukin 10
The results showed that there was significant decrease in IL10 levels of HCT116, Colo320 and Colo 205 with their respective control groups. Whereas, we observed mild increase in IL10 level in L132 with respect to its control depicted in Figure 8a.
3.7.2. Assessment of Human Interleukin 6
Interleukin-6 is involved in immune response as well as inflammation. In our investigation, quercetin treatment showed a noticeable decrease in IL6 levels in all the treatment groups (L132Q120 µM, p=0.0006), (HCT116Q120 µM, p=0.0001), (Colo320Q120 µM, p=0.0001) and (Colo205 Q120µM, p=0.0001) to their relevant control groups, Figure 8b.
3.7.3. Assessment of Human Tumor Necrosis Factor-α
Quercetin treated group showed a significant decrease in TNF-α expression in Q120 µM HCT116 (p=0.0007), Colo 320 (p=0.0003), and Colo 205 (p=0.0001) cells at a higher dose of quercetin than the respective control group. In contrast, no significant reduction was observed in L132 (p=0.0156) cells to their control group, Figure 8c.
3.7.4. Assessment of Vascular Endothelial Growth Factor
Quercetin significantly reduced VEGF concentrations in HCT116 (Q120µM, p=0.0123) and Colo320 (Q120µM, p=0.0003) and Colo205 (Q120µM, p=0.0084) cancer cells compared to the relevant control groups. Despite this, no considerable difference was detected in the L132 quercetin-treated group, Figure 8d.
4. Discussion
Quercetin is a plant pigment and secondary metabolite, acts on a multitude of targets involved in the initiation and promotion/progression phases of colorectal carcinogenesis [20]. Due to disease recurrence, the inadequacy of cancer treatment results in suboptimal outcomes. In our investigation, CCK-8 analysis was carried out, which showed that quercetin has a significant cytotoxic impact on normal epithelial, primary, and metastatic colon cancer cell proliferation. The highly quoted antioxidant activity of quercetin, it is noted that it works directly as a ROS scavenger [21]. The cancer cell regulates ROS and antioxidants precisely to survive with high ROS levels. In contrast, antioxidants protect antioxidant defense systems by scavenging reactive oxygen species [22]. When a cell undergoes stress conditions, it starts producing free radicals, accumulating and increasing the ROS level and generating oxidative stress [23]. Antioxidant enzymes such as LPO, catalase, SOD, ABTS, and DPPH neutralize these free radicals and decrease the level of ROS [24]. Our results demonstrated that, quercetin treatment at 120 µM/mL enhanced ROS-generated stress by suppressing cancer cells total thiol, catalase, LPO and SOD activity. Oxidative stress can cause cell cycle arrest, senescence, and finally, apoptosis and necrosis [25]. Similarly, the ABTS and DPPH showed the potential ability of quercetin to scavenge free radicals in primary colon cancer cell line. Therefore, the analysis of the antioxidants suggests that targeting antioxidant enzymes might be a promising approach for generating senescence, thereby apoptosis in cancer cells, based on the above outcome. The inhibition of malignant development may be achieved by the induction of cellular senescence in response to pro-tumorigenic stimuli [26]. SA-β-Galactosidase, a putative hydrolase enzyme, is a well-known biomarker of cell senescence [27]. The activity is driven by residual lysosomal β-galactosidase at pH 6.0 and reflects increased lysosomal content in senescent cells [28]. This study found a concentration-dependent rise in lysosomal-β galactosidase, resulting in a considerable increase in senescent cells in all quercetin-treated cancer cells. Colon cancer commonly undergoes spontaneous apoptosis, possibly due to genetic instability and increased cellular turnover [29]. Bax is a pro-apoptotic protein and an essential gateway of the intrinsic apoptotic pathway [30]. The quercetin treatment showed a potential increase in apoptosis by upregulated Bax expression and down-regulating an anti-apoptotic gene, Bcl-2, in colon cancer cells. Furthermore, the primary tumor suppressor gene p53 is also known as the guardian of the genome due to its primary role in maintaining genome integrity by detecting mutations and inhibiting cell multiplication with mutated genes or damaged DNA [31]. Here, quercetin significantly upregulated the expression of p53. Hence, the reactivation of p53 in cancer cells might be aiding in inducing cell senescence. Wnt/β-catenin mediated signaling pathway is a critical pathway that facilitates colon cancer cell proliferation [32]. Wnt genes are cysteine-rich glycoproteins secreted by cells into the extracellular matrix that activate receptor-mediated signaling, whereas CTNNB1 acts as a signal transducer, and the Wnt/β-catenin signaling system regulates cell proliferation, differentiation, migration, genetic stability, apoptosis, and stem cell renewal [33,34]. By analyzing the expression of both Wnt1 and CTNNB1 in all the treated colon cancer cells, we demonstrated that quercetin substantially inhibits the Wnt1 and CTNNB1 gene expression, thereby Wnt/β-catenin signaling pathway, which has restricted cancer cell proliferation. Epigenetic regulation alludes to the heritable and potentially reversible alterations in the phenotypic variability of the genome that modify gene transcription without altering the DNA sequence [35]. The HDAC protein family is an important regulator of chromatin integrity and governs the consequence of gene expression patterns [36]. In cancer cells, HDAC-mediated gene expression regulation is accomplished by separating the acetyl group from the histone proteins tail to silence the transcription of tumor suppressor genes. Several studies have reported that overexpressed HDACs may promote tumorigenesis [37]. There are 4 classes of HDACs among them, class I HDACs (HDAC1, HDAC2, HDAC8), class II HDACs (HDAC4, HDAC5, HDAC10), and hTERT are directly involved in tumor-related gene expression [38]. Since HDACs were one of the predicted targets of quercetin, the expression analysis of class I and class II HDAC were performed in quercetin-treated colon cancer cells. Quercetin administration reduced the expression of all HDACs and hTERT in colon cancer in a dose-dependent manner, thereby cell senescence and finally apoptosis. Downregulation of HDACs promotes cytochrome C release by activating the intrinsic apoptosis pathway and the activation of caspases by activating the intrinsic apoptosis pathway [39]. Recent studies have found that HDAC inhibitors significantly impact colon cancer cell proliferation and cell cycle. HDAC1 and HDAC2 are reported to be associated with regulating the cell cycle, which is overexpressed in cancer, including colon cancer [40]. Further, the expression of HDAC5 is anomalous in many cancers and is involved in tumor growth, invasion, and immune response, which have been thoroughly investigated [41]. HDAC10 contributes significantly to malignancies by regulating various cellular processes via its epigenetic action on specific signaling pathways. Moreover, the downregulated HDAC10 reduces the Wnt pathway activity, which is also known to promote apoptosis in colon cancer [42]. Hence, the present study signifies that the level of HDACs might be extremely important in revealing their tremendous importance in tumor cells when used with traditional methods [43]. This paradigm is influenced by the abundance of in vitro and in vivo data indicating that epigenetics play a vital role in colon cancer, and their reversible feature decides the rationale behind the current approach [44]. Chronic inflammation in epithelial cells, such as cancer cells, results in epigenetic alterations, such as DNA methylation modifications [45]. IL10 is a critical immunosuppressive agent and pro-inflammatory cytokine implicated in inflammation that promotes cancer development and immune responses against malignancies [46]. Similarly, TNF-α is a pro-inflammatory multifunctional cytokine that plays crucial roles in cellular processes, including cell viability, proliferation, differentiation, and apoptosis [47,48]. In cancer, the production of VEGF and other growth factors induces the 'angiogenic switch,' which causes new vasculature around the tumor, allowing it to grow tremendously [49]. Hence, by ELISA study, we examined the impact of quercetin over the IL10, IL6, TNF-α, and VEGF levels in colon cancer cells, where we found that quercetin significantly reduced the level of IL10, IL6, TNF-α, and VEGF cytokines in all the quercetin treated colon cancer cell lines. TNF-α is a critical inflammatory cytokine of the tumor microenvironment, facilitating colon cancer invasion [29,51]. Further, the expression of VEGF was determined, a homodimer glycoprotein that promotes angiogenesis in embryonic development and wound healing [52,53]. The result of VEGF expression showed that quercetin effectively inhibited expression levels in all colon cancer cell lines.
5. Conclusions
In conclusion, our findings demonstrate that quercetin inhibits proliferation, triggers apoptosis, and suppresses apoptotic genes. The results demonstrated the importance of polyphenols and flavonoids in cancer treatment, and prevention is now widely acknowledged. Based on the gene expression of genetic and epigenetic markers and cytokine analysis, quercetin treatment may significantly prevent inflammation and angiogenesis, which plays a key function in tumor development. Moreover, quercetin shows the potential to modify epigenetic regulation by modulating related signaling pathways like the Wnt/β-catenin signaling pathway, which is a remarkably conserved pathway strongly linked to colon cancer. Hence, a combinational therapy regimen combining quercetin and epigenetic modifiers with current chemotherapeutic medications may assist in slowing the spread of colon cancer and lessen the negative effects of the chemotherapeutic agents.
Author Contributions
AB designed and conceptualized the study. MB conducted general experiments, assays, and in vitro experiments. AB, MB, SMJ, and SP supported data analysis and wide-ranging aspects of the manuscript preparation and development process. SP and AD supported in crucial editing and statistical analysis. AB, AKD, and SFA critically reviewed the draft manuscript and provided feedback on data and analyses. All authors read and approved the final manuscript.
Funding
The work was partially supported by the departmental grants sanctioned to Dr. Antara Banerjee from Chettinad Academy of Research and Education. The APC was supported by the Project Number (RSPD2023R709), King Saud University, Riyadh, Saudi Arabia.
Institutional Review Board Statement
Non-Applicable.
Informed Consent Statement
Non-Applicable. This research does not involve human/animal participants, human/ animal material, or human data.
Data Availability Statement
The entire data set resulting from the studies that have been analyzed for this study are depicted in the figures and are also accessible from the corresponding author.
Acknowledgment
The authors are thankful to the Chettinad Academy of Research and Education (CARE) for providing the infrastructural support to complete this piece of work.
Conflicts of Interest
None. All authors declare that there are no conflicts of / or competing interests.
References
- Sung, H.; Ferlay, J.; Siegel, R. L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA: a cancer journal for clinicians 2021, 71 (3), 209-249. [CrossRef]
- Sawicki T; Ruszkowska M; Danielewicz A; Niedźwiedzka E; Arłukowicz T; Przybyłowicz KE. A Review of Colorectal Cancer in Terms of Epidemiology, Risk Factors, Development, Symptoms and Diagnosis. Cancers (Basel), 2021;13(9):2025. [CrossRef]
- Jung G; Hernández-Illán E; Moreira; Balaguer F; Goel A. Epigenetics of colorectal cancer: biomarker and therapeutic potential. Nat Rev Gastroenterol Hepatol, 2020;17(2):111-130. [CrossRef]
- Sriramulu, S; Malayaperumal, S; Deka, D.; Banerjee, A. and Pathak, S. A General Overview on Causes, Risk Factors, Diagnosis, Treatment, and Role of Oxidative Stress Biomarkers in Colorectal Cancer. Handbook of Oxidative Stress in Cancer: Therapeutic Aspects,2022, 3877-3895. [CrossRef]
- Piotrowski I; Kulcenty K; Suchorska W. Interplay between inflammation and cancer. Rep Pract Oncol Radiother, 2020;25(3):422-427. [CrossRef]
- D'Orazi G; Cordani M; Cirone M. Oncogenic pathways activated by pro-inflammatory cytokines promote mutant p53 stability: clue for novel anticancer therapies. Cell Mol Life Sci,2021;78(5):1853-1860. [CrossRef]
- Singh N; Baby D; Rajguru JP; Patil PB; Thakkannavar SS; Pujari VB. Inflammation and cancer. Ann Afr Med, 2019;18(3):121-126. [CrossRef]
- Fernández J, Silván B, Entrialgo-Cadierno R, et al. Antiproliferative and palliative activity of flavonoids in colorectal cancer. Biomed Pharmacother, 2021;143:112241. [CrossRef]
- Esmeeta A; Adhikary S; Dharshnaa V, et al. Plant-derived bioactive compounds in colon cancer treatment: An updated review. Biomed Pharmacother. 2022;153:113384. [CrossRef]
- Asgharian P; Tazekand AP; Hosseini K; et al. Potential mechanisms of quercetin in cancer prevention: focus on cellular and molecular targets. Cancer Cell Int, 2022;22(1):257. [CrossRef]
- Bhatiya M; Pathak S; Jothimani G; Duttaroy AK; Banerjee A. A Comprehensive Study on the Anti-cancer Effects of Quercetin and Its Epigenetic Modifications in Arresting Progression of Colon Cancer Cell Proliferation. Arch Immunol Ther Exp (Warsz), 2023;71(1):6. [CrossRef]
- Kesavan, P; Banerjee, A; Banerjee, A; Murugesan, R; Marotta, F and Pathak, S. An overview of dietary polyphenols and their therapeutic effects. Polyphenols: Mechanisms of Action in Human Health and Disease, 2018,221-235. [CrossRef]
- Batiha, GE; Beshbishy,AM; Ikram M; et al. The Pharmacological Activity, Biochemical Properties, and Pharmacokinetics of the Major Natural Polyphenolic Flavonoid: Quercetin. Foods, 2020;9(3):374. [CrossRef]
- Li Y; Zhang T; Chen GY. Flavonoids and Colorectal Cancer Prevention. Antioxidants (Basel),2018;7(12):187. [CrossRef]
- Zeb A; Ullah F. A Simple Spectrophotometric Method for the Determination of Thiobarbituric Acid Reactive Substances in Fried Fast Foods. J Anal Methods Chem, 2016;2016:9412767. [CrossRef]
- Hadwan MH; Abed HN. Data supporting the spectrophotometric method for the estimation of catalase activity. Data Brief,2015;6:194-199. [CrossRef]
- Weydert CJ; Cullen JJ. Measurement of superoxide dismutase, catalase and glutathione peroxidase in cultured cells and tissue. Nat Protoc, 2010;5(1):51-66. [CrossRef]
- Majkić N; Berkes I. Determination of free and esterified cholesterol by a kinetic method. I. The introduction of the enzymatic method with 2,2'-azino-di-3[ethyl-benzthiazolin sulfonic acid (6)] (ABTS). Clin Chim Acta, 1977;80(1):121-131. [CrossRef]
- Yuan XG; Huang YR; Yu T; Jiang HW; Xu Y; Zhao XY. Chidamide, a histone deacetylase inhibitor, induces growth arrest and apoptosis in multiple myeloma cells in a caspase-dependent manner. Oncol Lett, 2019;18(1):411-419. [CrossRef]
- Neamtu AA; Maghiar TA; Alaya A, et al. A Comprehensive View on the Quercetin Impact on Colorectal Cancer. Molecules, 2022;27(6):1873. [CrossRef]
- Xu D; Hu MJ; Wang YQ; Cui YL. Antioxidant Activities of Quercetin and Its Complexes for Medicinal Application. Molecules, 2019;24(6):1123. [CrossRef]
- Lo S; Leung E; Fedrizzi B; Barker D. Synthesis, Antiproliferative Activity and Radical Scavenging Ability of 5-O-Acyl Derivatives of Quercetin. Molecules, 2021;26(6):1608. [CrossRef]
- Sadiq IZ. Free Radicals and Oxidative Stress: Signaling Mechanisms, Redox Basis for Human Diseases, and Cell Cycle Regulation. Curr Mol Med, 2023;23(1):13-35. [CrossRef]
- Nwachukwu ID; Sarteshnizi RA; Udenigwe CC; Aluko RE. A Concise Review of Current In Vitro Chemical and Cell-Based Antioxidant Assay Methods. Molecules, 2021;26(16):4865. [CrossRef]
- Di Micco R; Krizhanovsky V; Baker D; d'Adda di Fagagna F. Cellular senescence in ageing: from mechanisms to therapeutic opportunities. Nat Rev Mol Cell Biol, 2021;22(2):75-95. [CrossRef]
- Fakhri S; Zachariah Moradi S; DeLiberto LK; Bishayee A. Cellular senescence signaling in cancer: A novel therapeutic target to combat human malignancies. Biochem Pharmacol, 2022;199:114989. [CrossRef]
- González-Gualda E; Baker AG; Fruk L; Muñoz-Espín D. A guide to assessing cellular senescence in vitro and in vivo. FEBS J,2021;288(1):56-80. [CrossRef]
- Valieva Y, Ivanova E, Fayzullin A, Kurkov A, Igrunkova A. Senescence-Associated β-Galactosidase Detection in Pathology. Diagnostics (Basel), 2022;12(10):2309. [CrossRef]
- Chota A; George BP; Abrahamse H. Interactions of multidomain pro-apoptotic and anti-apoptotic proteins in cancer cell death. Oncotarget, 2021;12(16):1615-1626. [CrossRef]
- Akim AM; Sung YY; Muhammad TST. Cancer and Apoptosis. Methods Mol Biol. 2022;2543:191-210. [CrossRef]
- Ozaki T; Nakagawara A. Role of p53 in Cell Death and Human Cancers. Cancers (Basel),2011;3(1):994-1013. [CrossRef]
- Samatha Jain, M; Makalakshmi, M.K; Deka, D; Pathak, S. and Banerjee, A., 2022. Therapeutic Strategies Targeting Wnt/β-Catenin Signaling Pathway in Stem Cells for ROS-Induced Cancer Progression. In Handbook of Oxidative Stress in Cancer: Therapeutic Aspects (pp. 2133-2151). Singapore: Springer Nature Singapore. [CrossRef]
- Banerjee, A; Rowlo, P; Jothimani, G; Duttaroy, A.K and Pathak, S. Wnt signalling inhibitors potently drive trans-differentiation potential of mesenchymal stem cells towards neuronal lineage. J. Med. Biol. Eng ,2022, 42(5),630-646. [CrossRef]
- Disoma C; Zhou Y; Li S; Peng J; Xia Z. Wnt/β-catenin signaling in colorectal cancer: Is therapeutic targeting even possible?. Biochimie, 2022;195:39-53. [CrossRef]
- Neganova ME; Klochkov SG; Aleksandrova YR; Aliev G. Histone modifications in epigenetic regulation of cancer: Perspectives and achieved progress. Semin Cancer Biol, 2022;83:452-471. [CrossRef]
- Pant K; Peixoto E; Richard S; Gradilone SA. Role of Histone Deacetylases in Carcinogenesis: Potential Role in Cholangiocarcinoma. Cells, 2020;9(3):780. [CrossRef]
- Patra S; Panigrahi DP; Praharaj PP; et al. Dysregulation of histone deacetylases in carcinogenesis and tumor progression: a possible link to apoptosis and autophagy. Cell Mol Life Sci, 2019;76(17):3263-3282. [CrossRef]
- Milazzo G; Mercatelli D; Di Muzio G; et al. Histone Deacetylases (HDACs): Evolution, Specificity, Role in Transcriptional Complexes, and Pharmacological Actionability. Genes (Basel), 2020;11(5):556. [CrossRef]
- Argüelles S; Guerrero-Castilla A; Cano M, Muñoz MF; Ayala A. Advantages and disadvantages of apoptosis in the aging process. Ann N Y Acad Sci, 2019;1443(1):20-33. [CrossRef]
- Lee HY; Tang DW; Liu CY; Cho EC. A novel HDAC1/2 inhibitor suppresses colorectal cancer through apoptosis induction and cell cycle regulation. Chem Biol Interact, 2022;352:109778. [CrossRef]
- Yang J; Gong C; Ke Q; et al. Insights Into the Function and Clinical Application of HDAC5 in Cancer Management. Front Oncol, 2021;11:661620. [CrossRef]
- Cheng F; Zheng B; Wang J; et al. Histone deacetylase 10, a potential epigenetic target for therapy. Biosci Rep. 2021;41(6):BSR20210462. [CrossRef]
- Alseksek RK; Ramadan WS; Saleh E; El-Awady R. The Role of HDACs in the Response of Cancer Cells to Cellular Stress and the Potential for Therapeutic Intervention. Int J Mol Sci, 2022;23(15):8141. [CrossRef]
- Deka D; Scarpa M; Das A; Pathak S; Banerjee A. Current Understanding of Epigenetics Driven Therapeutic Strategies in Colorectal Cancer Management. Endocr Metab Immune Disord Drug Targets, 2021;21(10):1882-1894. [CrossRef]
- Jothimani G; Bhatiya M; Pathak S; Paul S; Banerjee A. Tumor Suppressor microRNAs in Gastrointestinal Cancers: A Mini-Review. Recent Adv Inflamm Allergy Drug Discov, 2022;16(1):5-15. [CrossRef]
- Li L; Yu R; Cai T; et al. Effects of immune cells and cytokines on inflammation and immunosuppression in the tumor microenvironment. Int Immunopharmacol, 2020;88:106939. [CrossRef]
- Dash S; Sahu AK; Srivastava A; Chowdhury R; Mukherjee S. Exploring the extensive crosstalk between the antagonistic cytokines- TGF-β and TNF-α in regulating cancer pathogenesis. Cytokine, 2021;138:155348. [CrossRef]
- Teleanu RI; Chircov C; Grumezescu AM; Teleanu DM. Tumor Angiogenesis and Anti-Angiogenic Strategies for Cancer Treatment. J Clin Med, 2019;9(1):84. [CrossRef]
- Rebecca RJ; Janaki CS; Jayaraman S; et al. Carica Papaya Reduces High Fat Diet and Streptozotocin-Induced Development of Inflammation in Adipocyte via IL-1β/IL-6/TNF-α Mediated Signaling Mechanisms in Type-2 Diabetic Rats. Curr Issues Mol Biol, 2023;18;45(2):852-884. [CrossRef]
- Bhat AA; Nisar S; Singh M; et al. Cytokine- and chemokine-induced inflammatory colorectal tumor microenvironment: Emerging avenue for targeted therapy. Cancer Commun (Lond), 2022;42(8):689-715. [CrossRef]
- V A; Nayar PG; Murugesan R; S S; Krishnan J; Ahmed SS. A systems biology and proteomics-based approach identifies SRC and VEGFA as biomarkers in risk factor mediated coronary heart disease. Mol Biosyst, 2016;12(8):2594-2604. [CrossRef]
- Riccardi C; Napolitano E; Platella C; Musumeci D; Melone MAB; Montesarchio D. Anti-VEGF DNA-based aptamers in cancer therapeutics and diagnostics. Med Res Rev, 2021;41(1):464-506. [CrossRef]
- Subbaraj GK; Masoodi T; Yasam SK; et al. Anti-angiogenic effect of nano-formulated water soluble kaempferol and combretastatin in an in vivo chick chorioallantoic membrane model and HUVEC cells. Biomed Pharmacother, 2023;163:114820. [CrossRef]
Figure 1.
(a-c): Biochemical assays were performed to measure the inhibitory effect of quercetin on antioxidant enzymes (a) Lipid peroxidase activity (b) Catalase activity (c) Superoxide dismutase activity was measured (spectrophotometric method) normal cells (L132), primary, and metastasis colon cancer cells. (d-f). Biochemical assays were performed to measure the inhibitory effect of quercetin on antioxidant enzymes (d) Total thiol content (e) ABTS free radical scavenging activity (f) DPPH free radical scavenging activity was measured (spectrophotometric method) in normal cells (L132), primary, and metastasis colon cancer cells.
Figure 1.
(a-c): Biochemical assays were performed to measure the inhibitory effect of quercetin on antioxidant enzymes (a) Lipid peroxidase activity (b) Catalase activity (c) Superoxide dismutase activity was measured (spectrophotometric method) normal cells (L132), primary, and metastasis colon cancer cells. (d-f). Biochemical assays were performed to measure the inhibitory effect of quercetin on antioxidant enzymes (d) Total thiol content (e) ABTS free radical scavenging activity (f) DPPH free radical scavenging activity was measured (spectrophotometric method) in normal cells (L132), primary, and metastasis colon cancer cells.
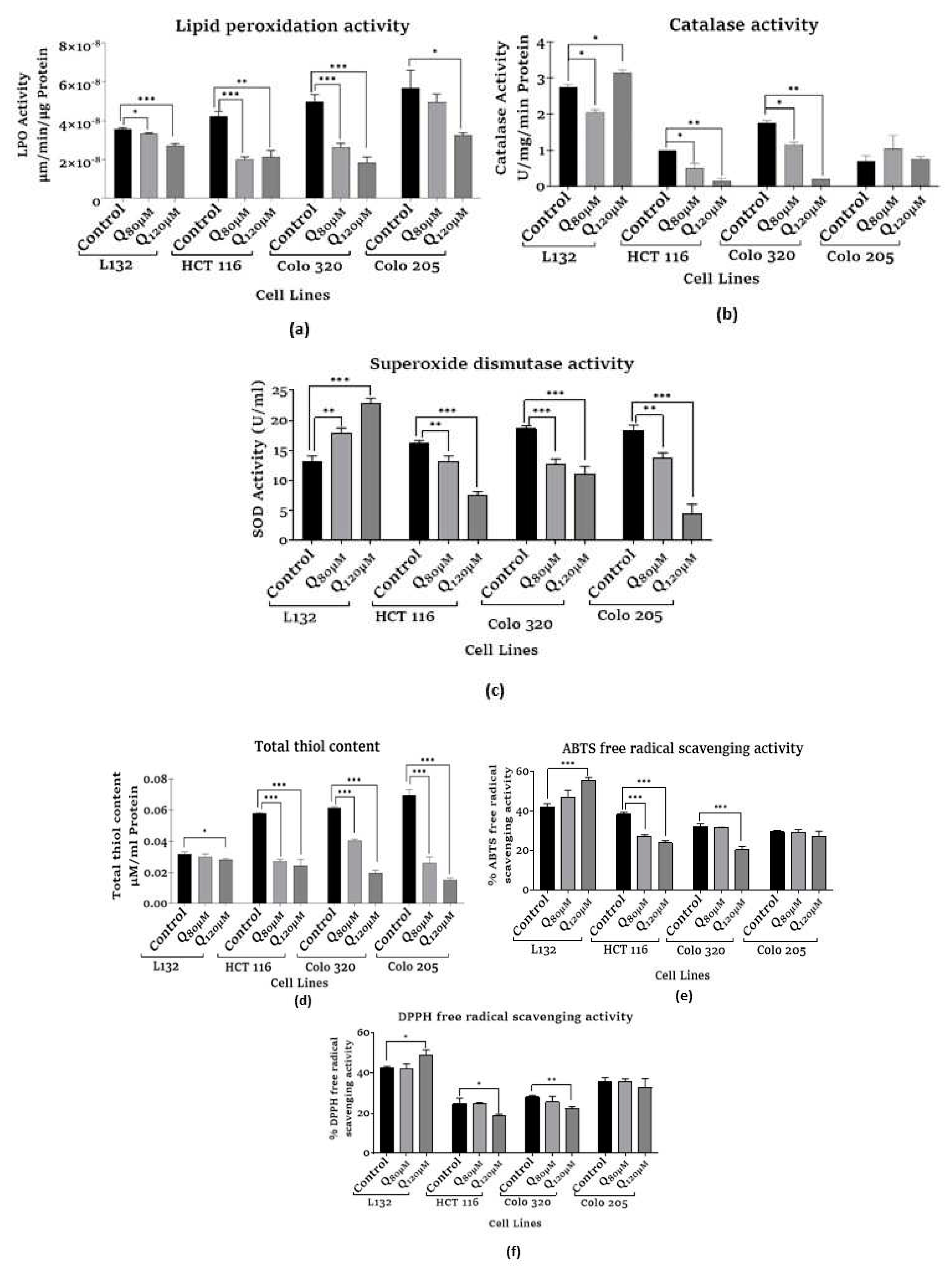
Figure 2.
(a) Graphical presentation, (b) morphological presentation of SA- β-Gal cell senescence analysis in control and quercetin-treated normal cells (L132), primary, and metastasis colon cancer cells.
Figure 2.
(a) Graphical presentation, (b) morphological presentation of SA- β-Gal cell senescence analysis in control and quercetin-treated normal cells (L132), primary, and metastasis colon cancer cells.
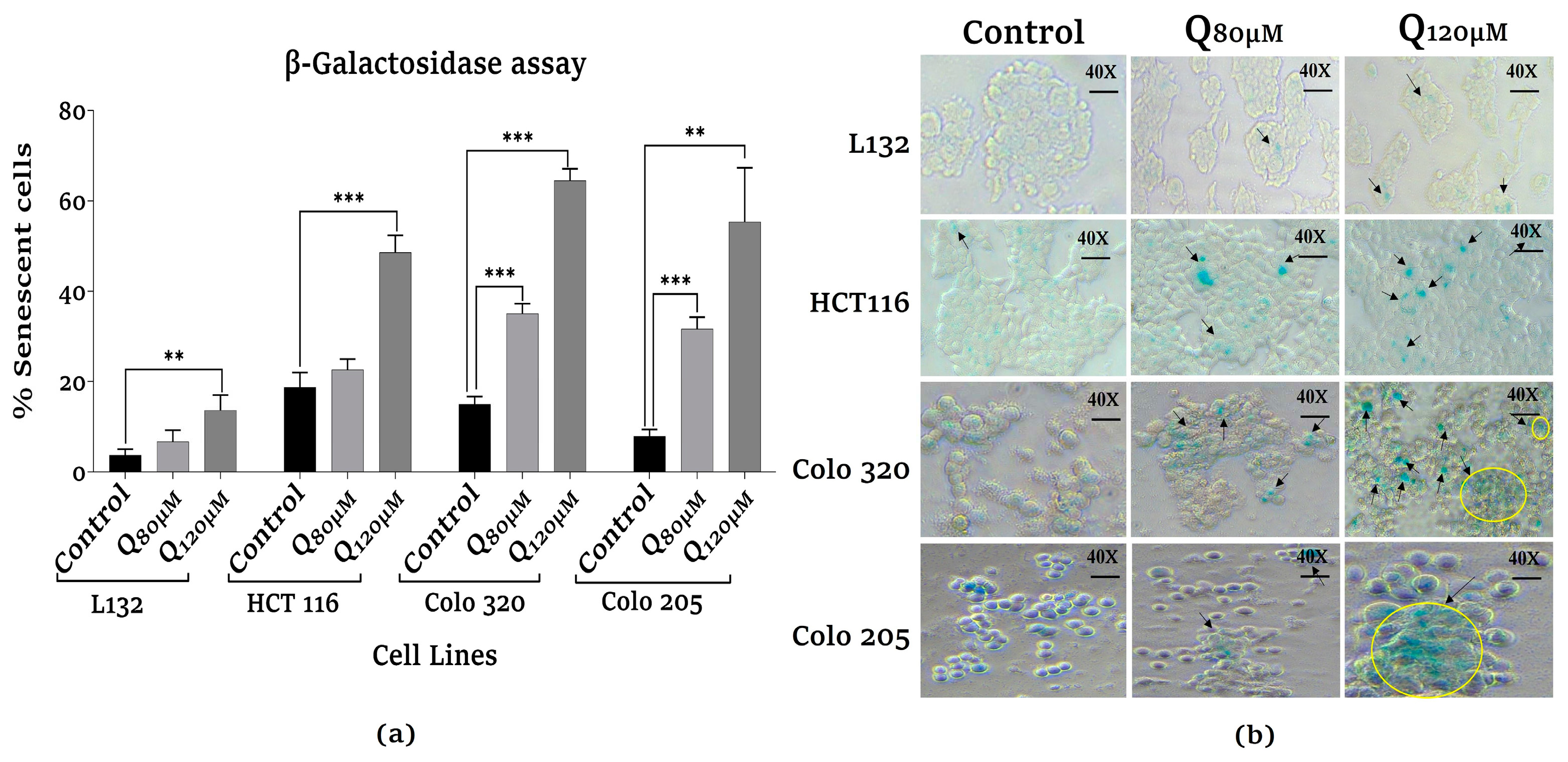
Figure 3.
Analysis of apoptosis-related gene and Tumor suppressor expression (a) Bax, (b) Bcl-2, (c) p53 in control and quercetin-treated normal cells (L132), primary, and metastasis colon cancer cells.
Figure 3.
Analysis of apoptosis-related gene and Tumor suppressor expression (a) Bax, (b) Bcl-2, (c) p53 in control and quercetin-treated normal cells (L132), primary, and metastasis colon cancer cells.
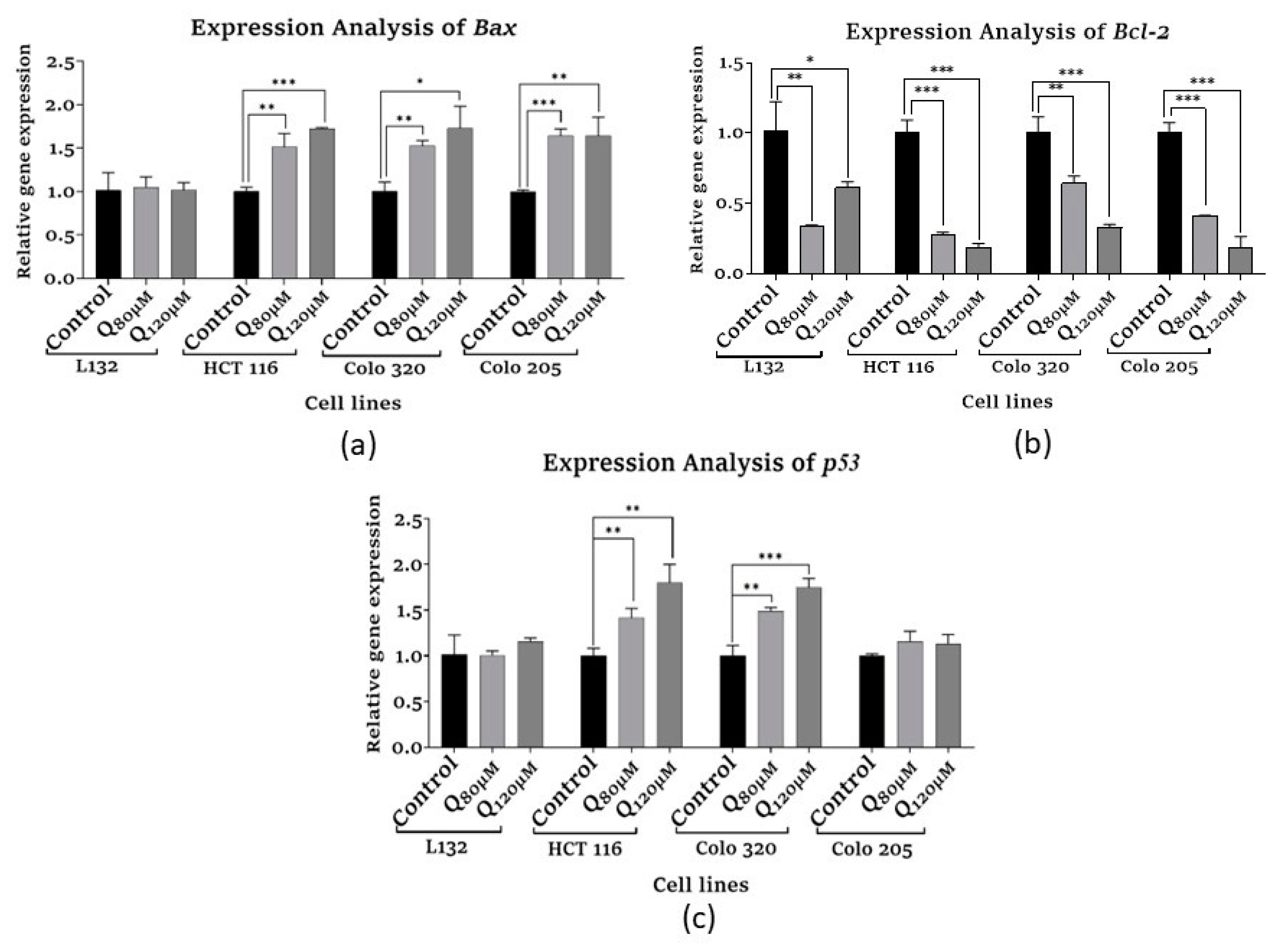
Figure 4.
Analysis of proliferation-related gene expression (a) β-catenin (CTNNB1), (b). Wnt1 in control and quercetin-treated normal cells (L132), primary, and metastasis colon cancer cells.
Figure 4.
Analysis of proliferation-related gene expression (a) β-catenin (CTNNB1), (b). Wnt1 in control and quercetin-treated normal cells (L132), primary, and metastasis colon cancer cells.
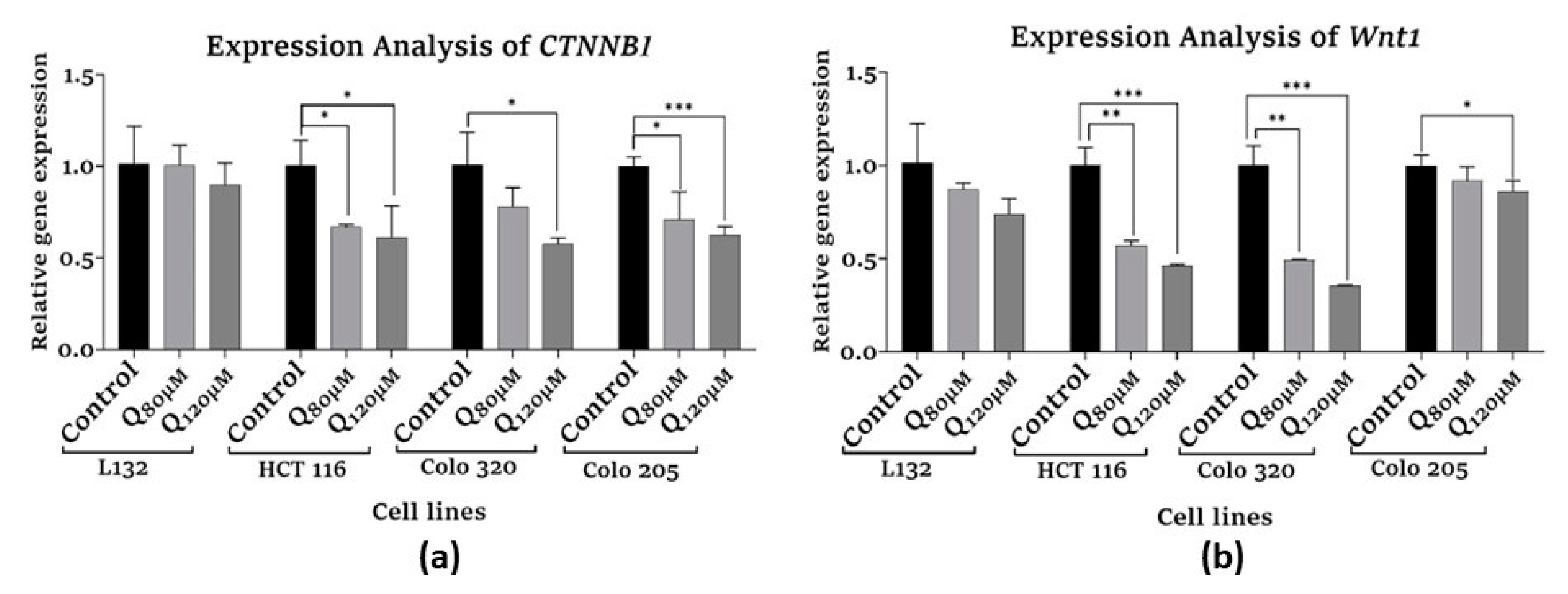
Figure 5.
Epigenetic modification analysis HDAC I family expression (a). HDAC1, (b) HDAC2 (c) HDAC8 in control and quercetin-treated normal cells (L132), primary, and metastasis colon cancer cells.
Figure 5.
Epigenetic modification analysis HDAC I family expression (a). HDAC1, (b) HDAC2 (c) HDAC8 in control and quercetin-treated normal cells (L132), primary, and metastasis colon cancer cells.
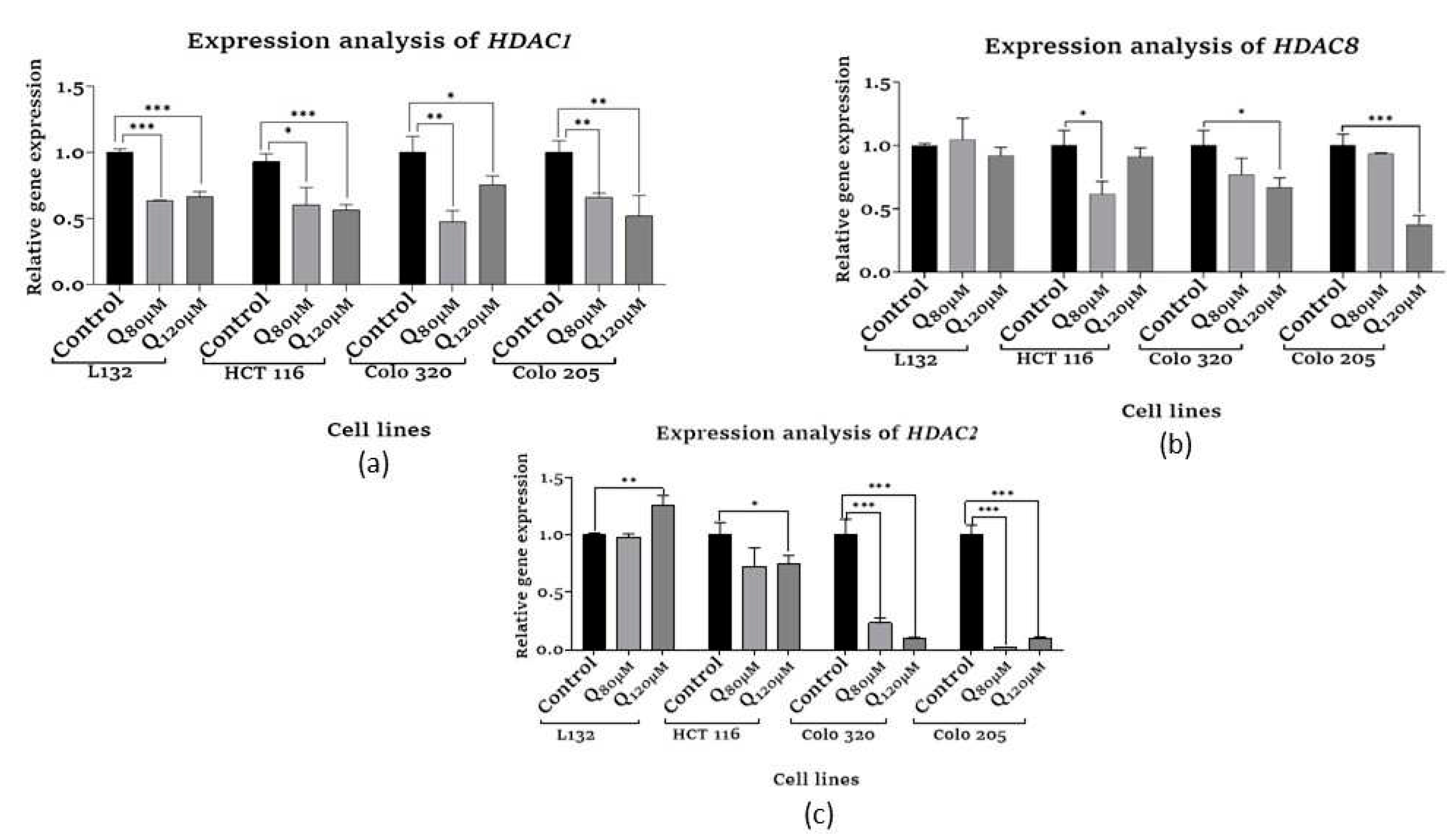
Figure 6.
Epigenetic modification analysis HDAC II family expression (a) HDAC4, (b) HDAC5 (c) HDAC10 in control and quercetin-treated normal cells (L132), primary, and metastasis colon cancer cells.
Figure 6.
Epigenetic modification analysis HDAC II family expression (a) HDAC4, (b) HDAC5 (c) HDAC10 in control and quercetin-treated normal cells (L132), primary, and metastasis colon cancer cells.
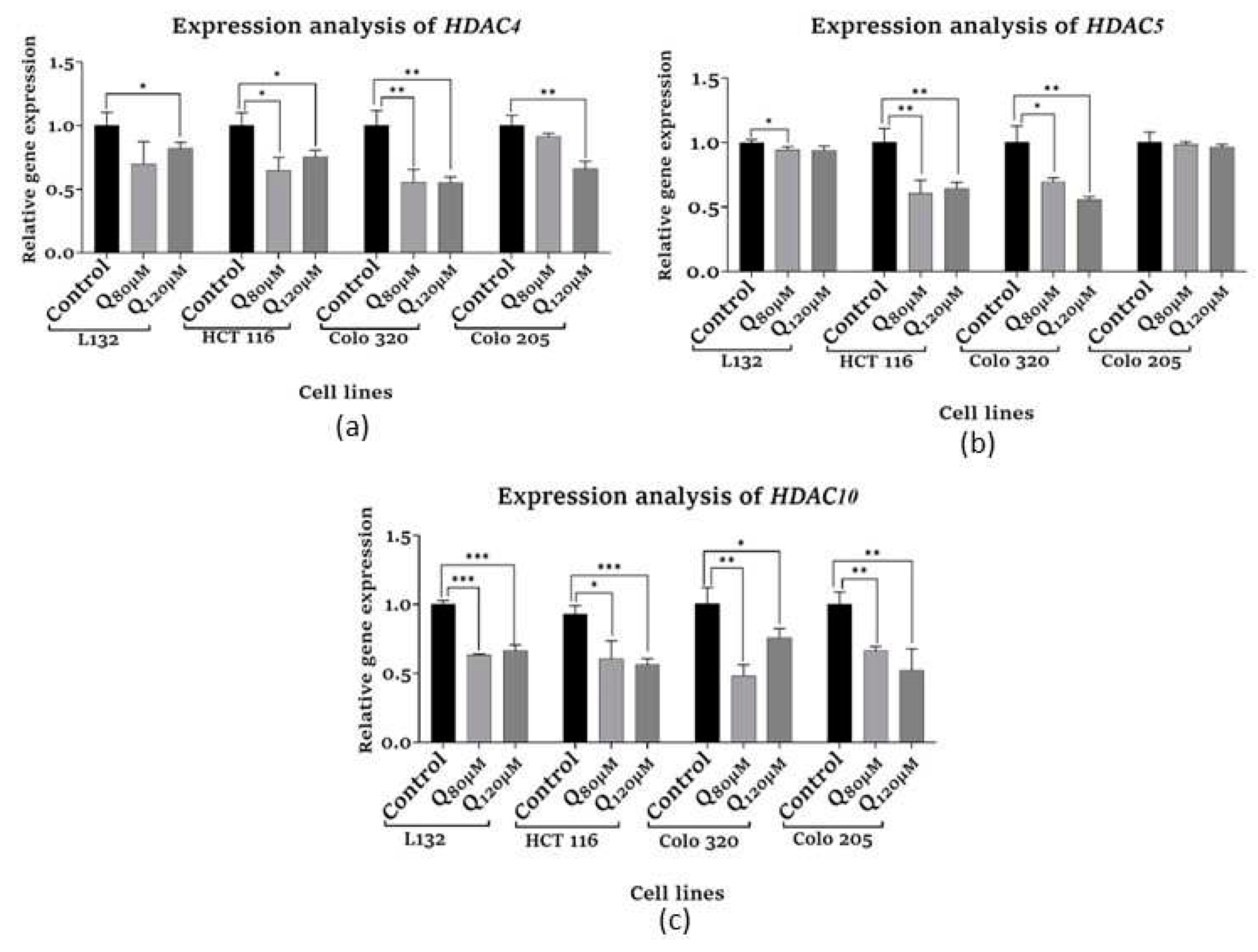
Figure 7.
Epigenetic modification analysis hTERT expression in control and quercetin-treated normal cells (L132), primary, and metastasis colon cancer cells.
Figure 8.
Effect of Quercetin on inflammatory cytokines and angiogenesis factor expression in normal cells (L132), primary, and metastasis colon cancer cells. (a) Human IL10 expression (b) Human IL6 expression (c) Human TNF-α. (d) Human VEGF. .
Figure 8.
Effect of Quercetin on inflammatory cytokines and angiogenesis factor expression in normal cells (L132), primary, and metastasis colon cancer cells. (a) Human IL10 expression (b) Human IL6 expression (c) Human TNF-α. (d) Human VEGF. .
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



