
Submitted:
01 April 2025
Posted:
03 April 2025
You are already at the latest version
Abstract
Background/Objectives: Aminoglycosides (AG) have been extensively used to treat bacterial diseases for decades. Nevertheless, despite their effectiveness, several secondary adverse effects (auditory, renal, and neurological) have been evident. Although the mechanisms of these side effects of AG are still unclear, there is a consensus that megalin is a drug receptor that participates in AG endocytosis. We aimed to explore the interaction between megalin and AGs to explain their ototoxicity side effect by combining in silico studies and cell functionality assays. Methods: Molecular dynamics simulations on the 10th CR domain of human megalin were performed using GROMACS. Seven commonly prescribed AG were docked against the surface of three conformations of this domain megalin derived from the simulations' trajectories. Gentamicin, the AG with the strongest affinity, was selected for cell functionality evaluation using HEK293 cells and Resazurin assays. Results: Docking results revealed that aspartic acids play a crucial role in the binding affinity to AGs, forming key electrostatic interactions. The models of the Megalin-AG complexes allowed the determination of electrostatic and non-electrostatic contributions to the free energy change of binding (ΔGb). Gentamicin, Tobramycin, and Neomycin showed a higher affinity for megalin than Kanamycin, Hygromycin, Paromomycin, and Streptomycin. Cell functionality of HEK293 cells was reduced by Gentamicin. Conclusion: the 10th CR domain of Megalin plays a critical role in AG-induced ototoxicity through electrostatic interactions mainly involving aspartic acid residues. It binds with higher affinity to Gentamicin, Tobramycin, and Neomycin than Kanamycin, Hygromycin, Paromomycin, and Streptomycin. Furthermore, we observed the reduced functionality of HEK293 cells induced by gentamicin. These results highlight megalin as a potential therapeutic target to mitigate/alleviate the adverse side effects of aminoglycoside.
Keywords:
megalin receptor
; aminoglycosides
; molecular dynamics
; docking studies
; binding energies
; functionality assays
1. Introduction
Streptomycin was first isolated eight decades ago from Streptomyces griseus [1], initiating the development of aminoglycoside (AG) antibiotics; since then, thousands of AGs have been either naturally isolated (produced by actinobacteria such as Streptomyces or Micromonospora) or obtained semi-synthetically (chemically modified derivatives of natural aminoglycosides). AG antibiotics are widely used to treat severe bacterial infections caused by Gram-positive and Gram-negative bacteria [2–4}. Despite their efficacy, their clinical use is frequently limited by adverse effects, mainly nephrotoxicity, neurotoxicity, and ototoxicity, which may affect the vestibular and/or cochlear systems. Vestibular toxicity occurs when the vestibular cranial nerve branch responsible for balance is damaged, and cochlear toxicity occurs due to damage to the cochlear Hair Cells (Inner Hair Cells -IHC- and Outer Hair Cells -OHC-) and the cranial nerve branch leading to sensorineural hearing loss [5]. The likelihood of ear damage occurring depends on the dose and duration of treatment with aminoglycosides and genetic susceptibility [6}.
Once AGs reach the inner ear fluid endolymph (fluid surrounding IHC), AG antibiotics enter IHC and OHC. Several mechanisms of AG antibiotics transport into IHC and OHC have been described. The three most important are: 1) via mechano-transduction channels, located at the tips of stereocilia which open and close in response to hair bundle deflection and allow non-specific cation influx {7], 2) by endocytosis, where AG antibiotics entry occurs through cell membrane invagination [8], and 3) by ion channels, like transient receptor potential channels, which are large calcium-permeable cation channels [9].
Like their antibacterial mechanism, AGs target the ribosomes within mammalian inner ear cells, triggering a cascade of deleterious intracellular events. Once aminoglycosides are inside hair cells, increased generation of reactive oxygen species (ROS) or free radicals, activation of stress kinases and proteolytic enzymes have been observed. Through in vivo experiments, it has been shown that AG antibiotics administration leads to activation of the JNK signaling pathway, which triggers pro-apoptotic signals [6,10]. Furthermore, it is suggested that inhibition of mitochondrial protein synthesis by AG antibiotic exposure leads to decreased ATP. Reduced energy production compromises mitochondrial integrity and predisposes to cytochrome C release and caspase activation, suggesting that hair cells undergo caspase-mediated apoptosis [4,11,12,13].
1.1. Megalin
Megalin, also known as Low-Density Lipoprotein Receptor-Related Protein 2, is a multiligand scavenger receptor belonging to the LDL receptor family. It is predominantly expressed by epithelial cells, including those of kidneys (renal proximal tubules) and the cochlea. In the kidney, megalin is essential for the reabsorption of filtered proteins. It interacts with various ligands, including aminoglycosides like gentamicin [14].
In the inner ear, the function of megalin remains less understood. Its expression in the cochlea suggests a potential role in the uptake of specific ligands, which could be relevant to auditory physiology. Structurally, the human megalin receptor is a large glycoprotein whose extracellular domain is composed primarily of complement-type repeat (CR) domains essential for ligand binding. Each CR domain comprises about 40 residues and contains a calcium-binding site and three disulfide bonds [15] . This work focused on the 10th CR domain from the human megalin receptor determined by Dagil et al. 2013. Recently, Goto et al 2024 elucidated the full architecture of megalin purified from rat kidneys using cryoelectron microscopy (Figure 1) [16].
Seven AG antibiotics (Gentamicin, Tobramycin, Kanamycin, Hygromycin, Streptomycin, Neomycin, and Paromomycin) were selected to explore the molecular recognition between megalin-AG antibiotics; all except hygromycin are approved by the Food and Drug Administration (FDA) and are available for clinical use1 Drew RH. Aminoglycosides. 2024, http://www.uptodate.com. Some characteristics and chemical structures of the seven AG studied here are shown in Table 1 and Figure 2.
1.2. Protonation States and pKa Considerations
The protonation state of aminoglycoside amino groups is relevant in receptor binding, given their contribution to electrostatic interactions. At physiological pH, these amino groups of aminoglycoside antibiotics (NH3+) are positively charged, enabling strong electrostatic interactions with negative charges on the receptor's surface. In this work, all aminoglycosides were modeled with protonated amino groups based on prior findings that such interactions contribute significantly to binding affinity and, consequently, to the ototoxic effects [14,17,18]. Furthermore, Zhou et al. (2012) showed a similar behavior for L-dopa, which holds two protonatable groups, the carboxylic moiety (pKa= 2.3) and the amino group (pKa= 8.11), where NH3+/NH2 can be protonated or deprotonated between pH 7.0- 8.0 (17). In addition, unpublished data from our group indicate that non-protonated amino groups might limit or impede the binding of aminoglycosides to megalin, highlighting the importance of pH-dependent protonation equilibria.
2. Results and Discussion
2.1. Molecular Dynamics Simulations and Cluster Analysis
The molecular dynamics simulations of the 10 th CR domain of human megalin was carried out for 200 ns by triplicate, yielding 600 ns of simulation data [19]. Root Mean Square Deviation (RMSD) values for Cα atoms were calculated for all systems (data not shown). Based on the RSMD behavior, we analyzed the final 50 ns from each replica. Cluster analysis allowed us to select the central structure of each replica. The most populated cluster of each experiment contained more than 75% of the analyzed frames. Central structures from the most populated clusters of each cluster were visually compared to the 20 structures determined by NMR from 2M0P [14]. As shown in Figure 3, the most representative structure from each cluster closely resembles the NMR structures.
To identify potential binding sites on the surface of the 10th CR domain of human megalin, blind molecular docking of seven aminoglycosides (Gentamicin, Kanamycin, Hygromycin, Paromomycine, Streptomycin, Tobramycin, and Neomycin) was performed using AutoDock Vina. The three representative Megalin structures (from each cluster) were used for docking experiments. Each ligand was docked 100 [20] independent runs against each cluster. The resulting poses were clustered using a 2 Å RMSD cutoff to obtain the most probable binding sites for each megalin cluster. Binding affinities and binding site occupancy percentages are summarized in Table 2.
2.2. Molecular Docking Analysis
Docking results for the three megalin structures showed two major aminoglycoside binding sites with favorable ΔGb values (Table 4). Figure 4a shows the major bong sites of cluster 1. Figure 4b-4h show representative aminoglycoside-meglin (cluster 1) interactions. Gentamicin formed three hydrogen bonds with Thr 1130, Asn 1132, and Glu1140 (Figure 4b). Tobramycin formed two hydrogen bonds with Thr 1130 and Glu 1140 with Gly 1137 (Figure 4c). Neomycin established four hydrogen bonds: two with Glu1140, one with Asp 1136 and Ser 1145 (Figure 4d). Paromomycin engaged six hydrogen bonds: two with Asp 1111 and one with Cys 1116, Asp 1117, His 1119, and Lys 1124 (Figure 4e). Hygromycin established two hydrogen bonds with Asn 1132 and Glu 1140 (Figure 4f). Streptomycin presented one hydrogen bond with Glu1140 (Fig 4g). Finally, Kanamycin shows five hydrogen bonds: one with Ala 1107, Cys 1109, two with Asp 1111, and one with Gln 1113 (Fig 4h). The amino acids highlighted in bold are consistent with those reported by Dagil et al., 2013 [14]. These authors also performed molecular docking studies of the megalin receptor with gentamicin. Notably, Thr 1130 and Asn 1132 were identified as part of one of the binding sites in both studies {14]. The Supplementary Material presents additional docking results for clusters 2 and 3, where overlapping binding sites were observed across ligands.
Table 3 highlights conserved residues forming the binding sites in each megalin cluster, essential for interacting with the various aminoglycosides.
2.3. Binding Free Energy Determinations
Binding free energy values (ΔGb) for each megalin-aminoglycoside complex was estimated for each representative cluster derived from molecular dynamics simulations (Table 4). The computational results suggested that the binding process of each megalin cluster with antibiotics was governed by electrostatic interactions, mainly by Coulombic forces, and a minor proportion by hydrophobic interactions. It seems important to emphasize that Megalin-Gentamicin, Megalin-Tobramycin, and Megalin-Neomycin systems presented the most favorable ΔGb values due to important electrostatic contacts in the binding, suggesting a stronger binding affinity that may contribute to their higher ototoxic potential.
2.4. Cell Functionality Assays
Resazurin assays in HEK 293 cells revealed that gentamicin reduced cell functionality at concentrations above 10 μM (Figure 5). This correlates with the high binding affinity of Gentamicin observed in docking experiments. Cell functionality decreased in a concentration-dependent manner.
The action mechanism of gentamicin in kidney cells has been reported. This mechanism consists of the antibiotic's internalization via the megalin-cubulin complex, followed by intracellular calcium accumulation. This causes endoplasmic reticulum stress and generation of reactive oxygen species (ROS), which consequently trigger necrosis and the intrinsic apoptosis pathway. It has been reported that the proliferator–activated receptor alpha (PPAR-α) is an important regulator of ROS within the cell. In tubular kidney cells, PPAR-α is less expressed when exposed to gentamicin, which may further enhance sensitivity to its toxicity [16,23].
Notably, gentamicin is frequently prescribed to pediatric patients, and it is reported that 5% of the administered dose of antibiotics accumulates, resulting in non-oliguric acute kidney injury (AKI) for long exposures to this antibiotic [24].
3. Materials and Methods
3.1. Molecular Dynamics Simulations
Molecular dynamics simulations (MDS) of the 10th CR domain of human megalin (PDB ID: 2M0P) were performed using the GROMACS 2019.2 software. Nuclear magnetic resonance (NMR) resolved the human megalin structure, and then the first model, out of twenty-structure ensembles, was used as the initial structure (14). CHARMM-GUI server was assigned hydrogens and charges with CHARMM36 force field [25,26]. Each system was surrounded by 10 Å of TIP3P water from any protein atom to a cubic water box; 0.15 M KCl was added to the simulation box to neutralize the water. All simulations were conducted in triplicate at 310.15 K and 1 atm of pressure for 200 ns, and inputs were used as suggested by the CHARMM-GUI server [19,27].
Trajectories were analyzed using GROMACS 2019.2 utilities. The last 50 ns of each simulation were clustered using the GROMOS method for clustering with a 0.75 Å cutoff (using this value was enough to obtain more than 75% of the structures on the most populated cluster).
3.2. Molecular Docking Assays
The central structure of the most populated cluster was used as the receptor model for blind molecular docking with aminoglycosides (Gentamicin, Kanamycin, Hygromycin, Paromomycin, Streptomycin, Tobramycin, and Neomycin) using Autodock Vina 1.2.5 [20]. Aminoglycosides were obtained from Pubchem Database [28]. Pubchem Database for each ligand were: Gentamicin (3467), Kanamycin (6032), Hygromycin (35766), Paromomycin (165580), Streptomycin (19649), Tobramycin (36294) and Neomycin (8378). Each aminoglycoside was loaded into MarvinSketch, and the best conformer was saved as mol2 and pdb file. Each ligand was docked in 100 blind docking experiments on the surface of each megalin cluster. The resulting poses were clustered based on a 2 Å RMSD threshold. Molecular interactions maps were generated using Maestro from Schrodinger and Visual Molecular Dynamics [22].
3.3. Computational Binding Energy (ΔGb) Calculations
The free energy change of binding (ΔGb) between megalin and its ligands was calculated according to the method described by Baker et al.. using equation 1 [29];
where the free energy change of solvation (ΔGsolv) and coulombic free energy changes ΔGCoul were calculated using the Adaptive Poisson-Boltzmann Solver (APBS) program (http://www.poissonboltzmann.org/) [21].
ΔGb= ΔGsolv + ΔGCoul +ΔGnon-elec,
In Nathan Baker´s script, we assigned dielectric constants of 78 and 4 for water and protein, respectively, for electrostatic determinations. Furthermore, the parameters were taken from the PDB2PQR server (http://server.poissonboltzmann.org/pdb2pqr) [30] as ionic radii and atomic charges for megalin, which were assigned from the CHARMM force field [31] at pH 7.0 [32]. Ligands' atomic charges were assigned from the force field implemented in the Autodock Vina program [20].
Regarding the ΔGnon-elec is the non-electrostatic contribution to the binding energy and was determined by multiplying the change in solvent accessible surface (ΔASA) upon binding by the coefficient interfacial tension (γ=5 cal•mol-1 Å-2) [33]. ASA values were determined by the VMD 1.9.1 program (https://www.ks.uiuc.edu/Research/vmd/) [22]:
ΔGnon-elec = γ (ASAmegalin-ligands complex− ASAmegalin – ASAligands),
3.4. Resazurin Cell Functionality Assays
The blue non-fluorescent dye Resazurin (7-Hydroxy-3H-phenoxazin-3-one 10 oxide), which is irreversibly reduced to the pink-colored fluorescent resorufin by dehydrogenase enzymes in metabolically active cells, was used to determine cytotoxicity on HEK293 cells. These cells could be used as a cellular model of ototoxicity since it has been reported that ototoxicity by aminoglycosides is related to other tissue damages such as the kidney (34). Cells were cultured with Dulbecco's Modified Eagle Medium (DMEM), supplemented with 10% Fetal Bovine Serum, and maintained at 95% humidity and 5% CO2. We used antibiotics with the most favorable affinity energy for the treatments. The concentrations were 10, 25, 50, and 75 mM. Not-treated cells (NT) were positive controls, while cells treated with a stable peroxide solution were negative controls. All data were analysed with one-way ANOVA and post-test Tukey (P < 0.05).
4. Conclusions
The molecular docking results revealed favorable binding interactions with ΔGb<0 between aminoglycosides with the megalin receptor, indicating spontaneous complex formation. Binding affinity was mainly governed by electrostatic interactions and hydrogen bonding, with minor contributions of non-electrostatic forces. Gentamicin, Tobramycin, and Neomycin exhibited the most favorable ΔGb values, suggesting stronger binding to megalin than Kanamycin, Hygromycin, Paromomycin, and Streptomycin. Gentamicin was identified as the aminoglycoside with the strongest binding affinity, and therefore, it was selected for cell functionality assays. HEK293 cells are susceptible to gentamicin by reducing its functionality, supporting its dual role as a potent antibiotic and a potential contributor to organ toxicity when not carefully monitored.
Supplementary Materials
The following supporting information can be downloaded at: www.mdpi.com/xxx/s1, Figure S1: All complexes shown interacting with megalin of cluster B; Figure S2: All complexes shown interacting with megalin of cluster C; Figure S3: Graphic representation of the cytotoxic effect of gentamicin in HEK 293 cells. Data are expressed as mean ± SEM (n = 4), ** P < 0.05 versus the not treated (NT) group; Figure S4: Graphic representation of the cytotoxic effect of gentamicin in HEK 293 cells. Data are expressed as mean ± SEM (n = 4), ** P < 0.05 versus the not treated (NT) group.
Author Contributions
Conceptualization, L.A.C.S. and G.P.H.; Methodology, L.A.C.S., I.N.S., C.M.P., and N.C.V.; Validation, A.L.M. and G.P.H; Formal Analysis, C.M.P., L.A.C.S. and I.N.S.; Investigation, C.B.C and A.Z.C.; Resources, A.L.M. and G.P.H; Data Curation, C.M.P., C.B.C and A. Z.C; Writing—Original Draft Preparation, I.N.S. L.A.C.S., C.M.P and N.C.V.; Writing—Review and Editing, I.N.S., L.A.C.S., C.B.C. and N.C.V; Project Administration, G.P.H and C.B.C.
Funding
This research was funded by Secretaría de Educación, Ciencia y Tecnología de la Ciudad de México (SECTEI) grant number SECTEI/026/2024, Instituto Politécnico Nacional & Universidad Autónoma Metropolitana (108 S264-24).
Institutional Review Board Statement
Not applicable.
Acknowledgments
We thank Laboratorio de Visualización y Cómputo Paralelo at Universidad Autónoma Metropolitana-Iztapalapa (UAM-I) for computing time.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Schatz, A.; Bugie, E.; A Waksman, S.; Hanssen, A.D.; Patel, R.; Osmon, D.R. The Classic: Streptomycin, a Substance Exhibiting Antibiotic Activity against Gram-Positive and Gram-Negative Bacteria. Clin. Orthop. Relat. Res. 6. [CrossRef]
- Krause, K.M.; Serio, A.W.; Kane, T.R.; Connolly, L.E. Aminoglycosides: An overview. Cold Spring Harb Perspect Med. 2016 Jun 1;6(6).
- Tevyashova, A.N.; Shapovalova, K.S. Potential for the Development of a New Generation of Aminoglycoside Antibiotics. Pharm. Chem. J. 2021, 55, 860–875. [Google Scholar] [CrossRef] [PubMed]
- Huth, M.E.; Ricci, A.J.; Cheng, A.G. Mechanisms of Aminoglycoside Ototoxicity and Targets of Hair Cell Protection. Int. J. Otolaryngol. 2011, 2011, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Jospe-Kaufman, M.; Siomin, L.; Fridman, M. The relationship between the structure and toxicity of aminoglycoside antibiotics. Bioorganic Med. Chem. Lett. 2020, 30, 127218–127218. [Google Scholar] [CrossRef]
- Wu, J.; Ye, J.; Kong, W.; Zhang, S.; Zheng, Y. Programmed cell death pathways in hearing loss: A review of apoptosis, autophagy and programmed necrosis. Cell Prolif. 2020, 53, e12915. [Google Scholar] [CrossRef]
- Alharazneh, A.; Luk, L.; Huth, M.; Monfared, A.; Steyger, P.S.; Cheng, A.G.; Ricci, A.J. Functional Hair Cell Mechanotransducer Channels Are Required for Aminoglycoside Ototoxicity. PLOS ONE 2011, 6, e22347. [Google Scholar] [CrossRef]
- Chun, K.-J.; Lee, C.-H.; Kim, K.-W.; Lee, S.-M.; Kim, S.-Y. Effects of Androgen Receptor Inhibition on Kanamycin-Induced Hearing Loss in Rats. Int. J. Mol. Sci. 2021, 22, 5307. [Google Scholar] [CrossRef]
- König, O.; Rüttiger, L.; Müller, M.; Zimmermann, U.; Erdmann, B.; Kalbacher, H.; Gross, M.; Knipper, M. Estrogen and the inner ear: megalin knockout mice suffer progressive hearing loss. FASEB J. 2007, 22, 410–417. [Google Scholar] [CrossRef]
- Lim, H.W.; Pak, K.; Ryan, A.F.; Kurabi, A. Screening Mammalian Cochlear Hair Cells to Identify Critical Processes in Aminoglycoside-Mediated Damage. Front. Cell. Neurosci. 2018, 12, 179. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Wan, P.; Li, P.; Wang, J.; Guo, S.; Zhang, Y.; An, Y.; Ye, C.; Liu, Z.; Gao, J.; et al. Mechanism and Prevention of Ototoxicity Induced by Aminoglycosides. Front. Cell. Neurosci. 2021, 15. [Google Scholar] [CrossRef]
- O’sullivan, M.E.; Perez, A.; Lin, R.; Sajjadi, A.; Ricci, A.J.; Cheng, A.G. Towards the Prevention of Aminoglycoside-Related Hearing Loss. Front. Cell. Neurosci. 2017, 11, 325–325. [Google Scholar] [CrossRef]
- Layman, W.S.; Zuo, J. Preventing ototoxic hearing loss by inhibiting histone deacetylases. Cell Death Dis. 2015, 6, e1882–e1882. [Google Scholar] [CrossRef] [PubMed]
- Dagil, R.; O’Shea, C.; Nykjaer, A.; Bonvin, A.M.J.J.; Kragelund, B.B. Gentamicin binds to the megalin receptor as a competitive inhibitor using the common ligand binding motif of complement type repeats insight from the nmr structure of the wth complement type repeat domain alone and in complex with gentamicin. J. Biol. Chem. 2013;288(6):4424–35.
- Saito, A.; Pietromonaco, S.; Loo, A.K.C.; Farquhar, M.G. Complete cloning and sequencing of rat gp330/’megalin,’ a distinctive member of the low density lipoprotein receptor gene family. Proc Natl Acad Sci U S A. 1994;91(21):9725–9.
- Goto, S.; Tsutsumi, A.; Lee, Y.; Hosojima, M.; Kabasawa, H.; Komochi, K.; Nagatoishi, S.; Takemoto, K.; Tsumoto, K.; Nishizawa, T.; et al. Cryo-EM structures elucidate the multiligand receptor nature of megalin. Proc. Natl. Acad. Sci. 2024, 121. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.Z.; Alany, R.G.; Chuang, V.; Wen, J. Studies of the Rate Constant of l-DOPA Oxidation and Decarboxylation by HPLC. Chromatographia 2012, 75, 597–606. [Google Scholar] [CrossRef]
- Hobbie, S.N.; Akshay, S.; Kalapala, S.K.; Bruell, C.M.; Shcherbakov, D.; Böttger, E.C. Genetic analysis of interactions with eukaryotic rRNA identify the mitoribosome as target in aminoglycoside ototoxicity. Proc Natl Acad Sci U S A. 2008;105:20888–93.
- Berendsen, H.J.C.; Van Der Spoel, D.; Van Drunen, R. GROMACS: A message-passing parallel molecular dynamics implementation PROGRAM SUMMARY Title of program: GROMACS version 1.0 [Internet]. Vol. 91, Comput. Phys. Commun. 1995. Available from: http://rugmd0,chela,rug.
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef]
- Jurrus, E.; Engel, D.; Star, K.; Monson, K.; Brandi, J.; Felberg, L.E.; Brookes, D.H.; Wilson, L.; Chen, J.; Liles, K.; et al. Improvements to the APBS biomolecular solvation software suite. Protein Sci. 2017, 27, 112–128. [Google Scholar] [CrossRef]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef]
- Quiros, Y.; Vicente-Vicente, L.; Morales, A.I.; López-Novoa, J.M.; López-Hernández, F.J. An Integrative Overview on the Mechanisms Underlying the Renal Tubular Cytotoxicity of Gentamicin. Toxicol. Sci. 2011, 119, 245–256. [Google Scholar] [CrossRef]
- McWilliam, S.J.; Antoine, D.J.; Smyth, R.L.; Pirmohamed, M. Aminoglycoside-induced nephrotoxicity in children. Pediatr. Nephrol. 2016, 32, 2015–2025. [Google Scholar] [CrossRef]
- Lee, J.; Cheng, X.; Swails, J.M.; Yeom, M.S.; Eastman, P.K.; Lemkul, J.A.; Wei, S.; Buckner, J.; Jeong, J.C.; Qi, Y.; et al. CHARMM-GUI Input Generator for NAMD, GROMACS, AMBER, OpenMM, and CHARMM/OpenMM Simulations Using the CHARMM36 Additive Force Field. J. Chem. Theory Comput. 2015, 12, 405–413. [Google Scholar] [CrossRef]
- Jo, S.; Kim, T.; Iyer, V.G.; Im, W. CHARMM-GUI: A web-based graphical user interface for CHARMM. J Comput Chem. 2008 Aug;29(11):1859–65.
- Páll, S.; Zhmurov, A.; Bauer, P.; Abraham, M.; Lundborg, M.; Gray, A.; Hess, B.; Lindahl, E. Heterogeneous parallelization and acceleration of molecular dynamics simulations in GROMACS. J. Chem. Phys. 2020, 153, 134110. [Google Scholar] [CrossRef]
- Kim, S.; Chen, J.; Cheng, T.; Gindulyte, A.; He, J.; He, S.; et al. PubChem 2025 update. Nucleic Acids Res. 2025 Jan 6;53(D1):D1516–25.
- Baker, N.A.; Sept, D.; Joseph, S.; Holst, M.J.; McCammon, J.A. Electrostatics of nanosystems: Application to microtubules and the ribosome. Proc. Natl. Acad. Sci. USA 2001, 98, 10037–10041. [Google Scholar] [CrossRef] [PubMed]
- Dolinsky, T.J.; Nielsen, J.E.; McCammon, J.A.; Baker, N.A. PDB2PQR: An automated pipeline for the setup of Poisson-Boltzmann electrostatics calculations. Nucleic Acids Res. 2004, 32, W665–W667. [Google Scholar] [CrossRef] [PubMed]
- Mackerell, A.D.; Bashford, D.; Bellott ⊥, M.; Dunbrack ⊥, R.L.; Evanseck, J.D.; Field, M.J.; et al. All-Atom Empirical Potential for Molecular Modeling and Dynamics Studies of Proteins †. J Phys Chem B. 1998;102(18):3586–616.
- Søndergaard, C.R.; Olsson, M.H.M.; Rostkowski, M.; Jensen, J.H. Improved Treatment of Ligands and Coupling Effects in Empirical Calculation and Rationalization of pKa Values. J. Chem. Theory Comput. 2011, 7, 2284–2295. [Google Scholar] [CrossRef] [PubMed]
- Levy, R.M.; Zhang, L.Y.; Gallicchio, E.; Felts, A.K. On the Nonpolar Hydration Free Energy of Proteins: Surface Area and Continuum Solvent Models for the Solute−Solvent Interaction Energy. J. Am. Chem. Soc. 2003, 125, 9523–9530. [Google Scholar] [CrossRef]
- Ward, D.T.; Maldonado-Pérez, D.; Hollins, L.; Riccardi, D. Aminoglycosides Induce Acute Cell Signaling and Chronic Cell Death in Renal Cells that Express the Calcium-Sensing Receptor. J. Am. Soc. Nephrol. 2005, 16, 1236–1244. [Google Scholar] [CrossRef]
Figure 1.
Megalin protein (ID PDB 8JUU) [16], the main segment of the interaction with aminoglycosides, is shown in red.
Figure 1.
Megalin protein (ID PDB 8JUU) [16], the main segment of the interaction with aminoglycosides, is shown in red.
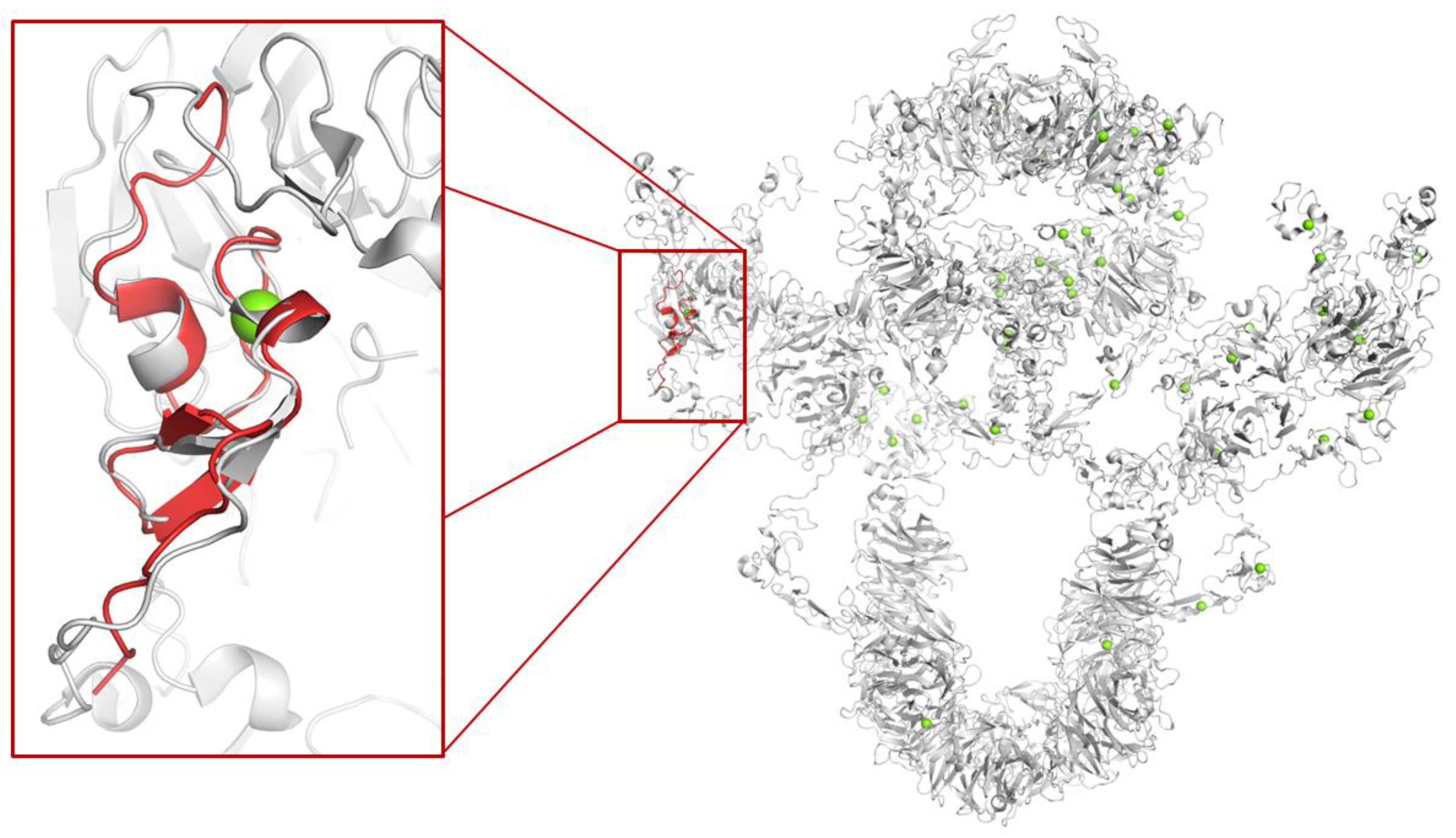
Figure 2.
Chemical structures of aminoglycosides, constructed in ChemDraw (Revvity Signals Software, 1985). a) Kanamycin. b) Paromomycin. c) Neomycin. d) Streptomycin. e) Hygromycin. f) Tobramycin. g) Gentamicin.
Figure 2.
Chemical structures of aminoglycosides, constructed in ChemDraw (Revvity Signals Software, 1985). a) Kanamycin. b) Paromomycin. c) Neomycin. d) Streptomycin. e) Hygromycin. f) Tobramycin. g) Gentamicin.
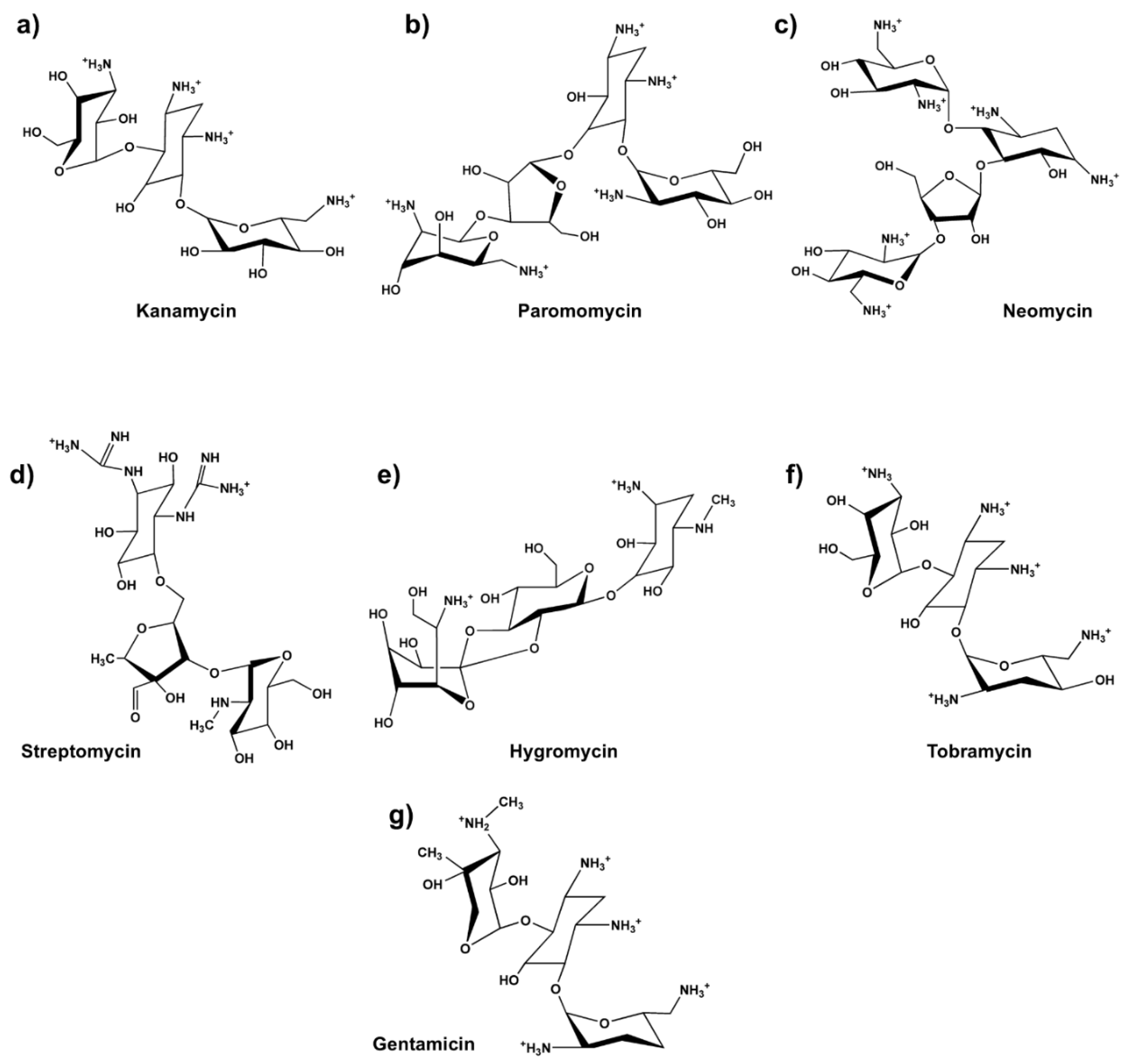
Figure 3.
Megalin structures obtained from clustering analysis. NMR structures are on white. Cluster structures are in red, green, and blue. Cluster structures RMSD against the first structure from 2M0P were 5.87, 5.56 and 5.14 Å and for the structure from 2M0P were 3.76, 3.28 and 4.36 Å. These RMSD values quantify the visual resemble of the cluster and NMR structures (2M0P).
Figure 3.
Megalin structures obtained from clustering analysis. NMR structures are on white. Cluster structures are in red, green, and blue. Cluster structures RMSD against the first structure from 2M0P were 5.87, 5.56 and 5.14 Å and for the structure from 2M0P were 3.76, 3.28 and 4.36 Å. These RMSD values quantify the visual resemble of the cluster and NMR structures (2M0P).
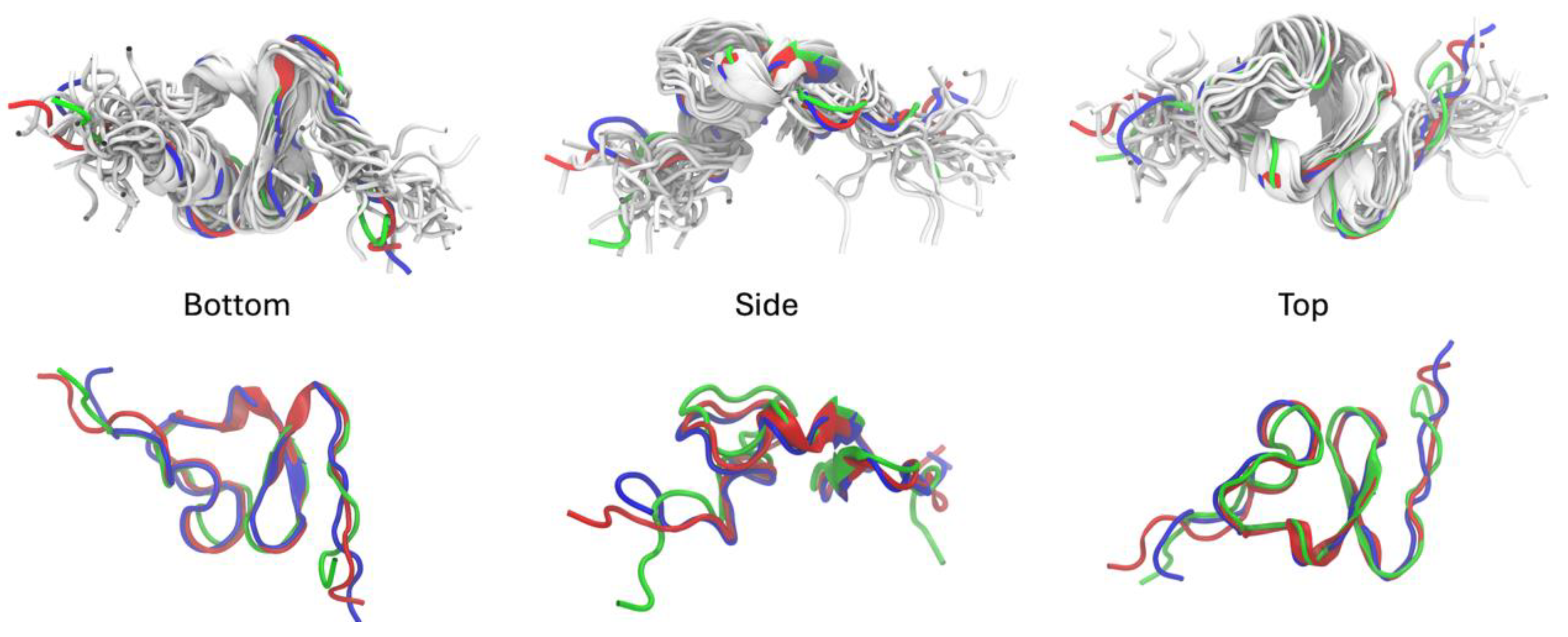
Figure 4.
a) All aminoglycosides interacting with megalin. b) Gentamicin. c) Tobramycin. d) Neomycin. e) Paromomycin. f) Hygromycin. g) Streptomycin. h) Kanamycin. Hydrogen bonds are the cyan lines.
Figure 4.
a) All aminoglycosides interacting with megalin. b) Gentamicin. c) Tobramycin. d) Neomycin. e) Paromomycin. f) Hygromycin. g) Streptomycin. h) Kanamycin. Hydrogen bonds are the cyan lines.
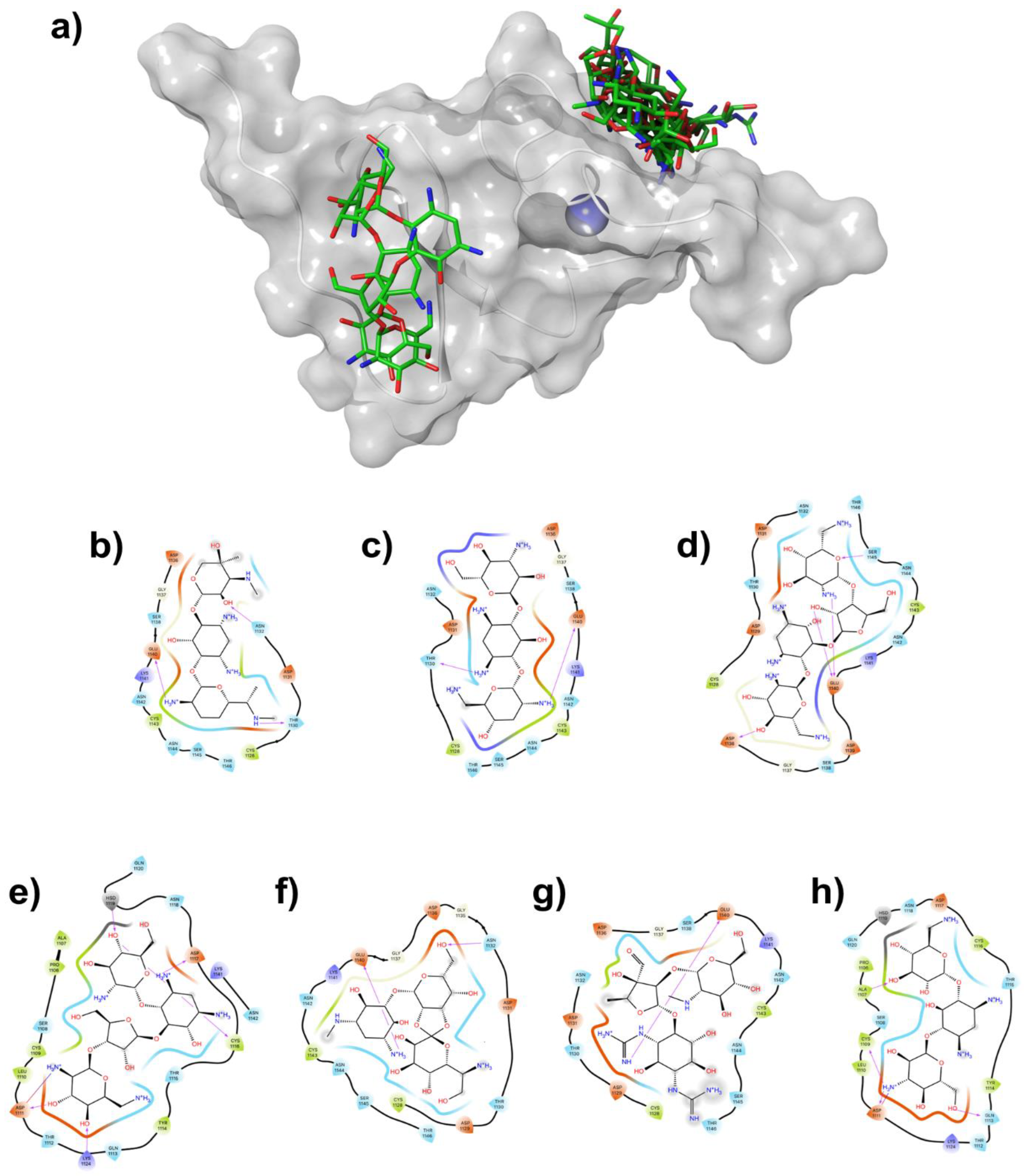
Figure 5.
Graphic representation of the cytotoxic effect of gentamicin in HEK 293 cells. The experiment was performed in triplicate. Data are expressed as mean ± SEM (n = 4), ** P < 0.05 versus the not treated (NT) group.
Figure 5.
Graphic representation of the cytotoxic effect of gentamicin in HEK 293 cells. The experiment was performed in triplicate. Data are expressed as mean ± SEM (n = 4), ** P < 0.05 versus the not treated (NT) group.
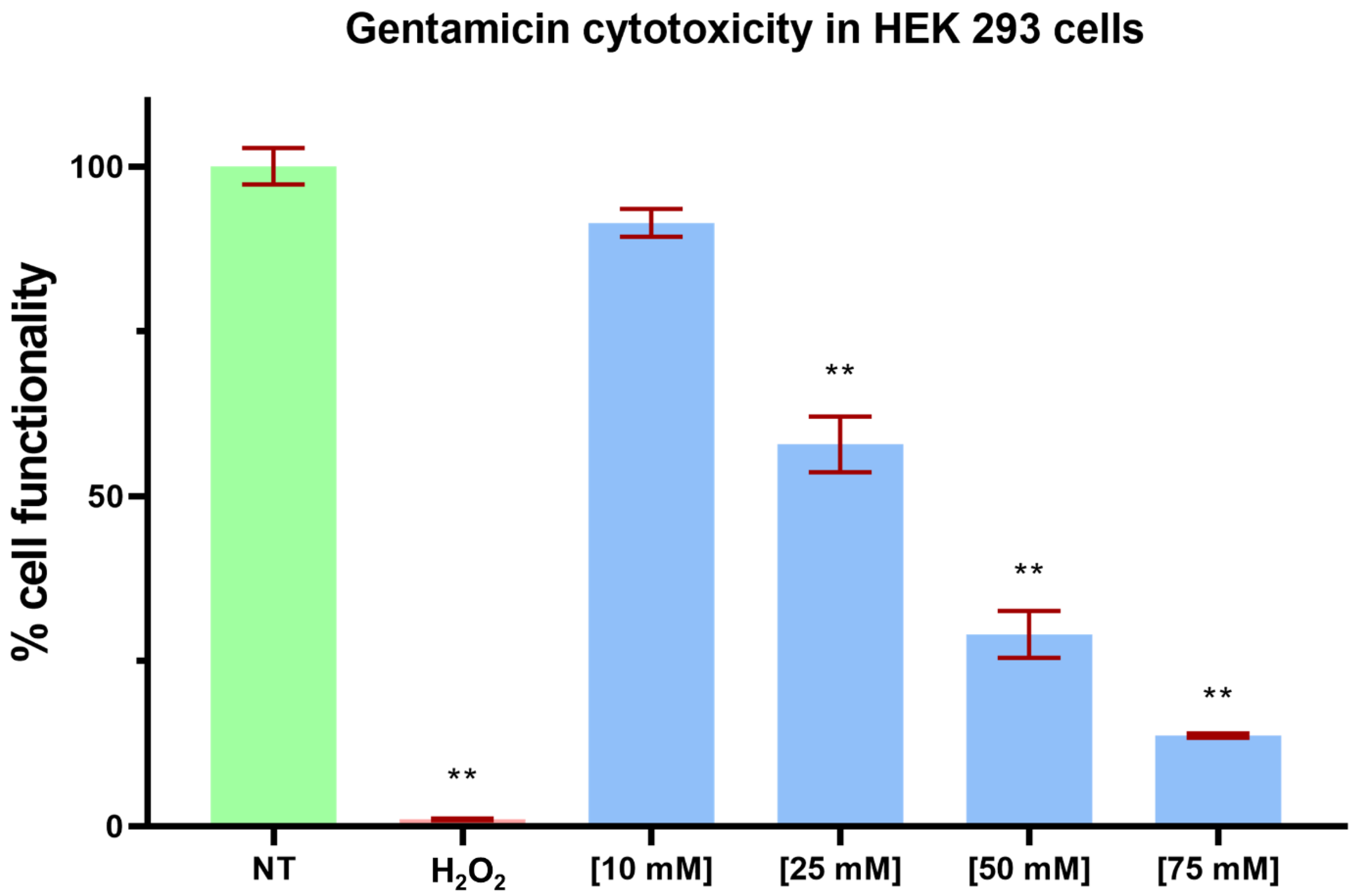

Table 1.
Principal characteristics of the seven AG aminoglycosides included in this work.
| Drug | Produced by (year) | Generation | Prescription | Ototoxicity (Level) |
|---|---|---|---|---|
| Streptomycin | Streptomyces griseus (1943) | I | Tuberculosis, plague, and other serious bacterial infections | Vestibular (intermediate) |
| Neomycin | Streptomyces fradiae (1949) | I | Topical bacterial infections or presurgical preparation of the gastrointestinal tract | Cochlear (high) |
| Kanamycin | Streptomyces kanamyceticus (1957) |
I | Serious bacterial infections, including urinary tract infections and tuberculosis | Cochlear (intermediate) |
| Paromomycin | Streptomyces rimosus (1959) | I | Treatment of intestinal infections caused by amoebiasis and leishmaniasis | Cochlear (intermediate) |
| Gentamicin | Micromonospora (1963) | II | Serious bacterial infections, such as sepsis, urinary tract infections, and respiratory tract infections | Neurotoxicity, Vestibular and Cochlear (intermediate) |
| Hygromycin B | Streptomyces hygroscopicus (1953-1957) |
II | Research used to select resistant cells in cell cultures | Cochlear (Low) |
| Tobramycin | Streptomyces tenebrarius (1967-1970) |
II | Serious bacterial infections, including lung infections in patients with cystic fibrosis | Vestibular and Cochlear (intermediate) |
Table 2.
Affinity energies (kcal/mol) and binding site occupancy, obtained from independent docking runs.
Table 2.
Affinity energies (kcal/mol) and binding site occupancy, obtained from independent docking runs.
| Aminoglycoside | Cluster 1 | Cluster 2 | Cluster 3 |
|---|---|---|---|
| Kanamycin | -5.90 +/- 0.01 (100) | -5.24 +/- 0.01 (100) | -5.12 +/- 0.05 (94) |
| Paromomycin | -5.86 +/- 0.03 (100) | -5.27 +/- 0.11 (44) | -5.27 +/- 0.06 (89) |
| Neomycin | -5.16 +/- 0.02 (93) | -5.30 +/- 0.00 (58) | -5.14 +/- 0.01 (100) |
| Streptomycin | -5.16 +/- 0.08 (92) | -5.39 +/- 0.13 (97) | -5.71 +/- 0.02 (100) |
| Hygromycin | -5.14 +/- 0.02 (52) | -5.67 +/- 0.01 (100) | -5.56 +/- 0.10 (98) |
| Tobramycin | -4.68 +/- 0.01 (100) | -5.16 +/- 0.07 (100) | -5.23 +/- 0.06 (99) |
| Gentamicin | -4.94 +/- 0.01 (100) | -5.16 +/- 0.03 (100) | -4.47 +/- 0.01 (100) |
Numbers in parentheses correspond to the percentage of structures clustered within a 2Å radius.
Table 3.
Residues within the binding site of each megalin cluster with aminoglycosides.
| Aminoglycosides | Cluster 1 | Cluster 2 | Cluster 3 |
|---|---|---|---|
|
Gentamicin |
1128, 1130, 1131 1132, 1136, 1137 1138, 1140, 1141 1142, 1143, 1144 1145, 1146 |
1106, 1107, 1108 1109, 1110, 1111 1112, 1113, 1114 1115, 1116, 1117 1118, 1119, 1124 1138, 1141, 1142 |
1117, 1130, 1131 1132, 1136, 1137 1138, 1139, 1140 1141, 1142, 1143 1144 |
|
Tobramycin |
1128, 1130, 1131 1132, 1136, 1137 1138, 1140, 1141 1142, 1143, 1144 1145, 1146 |
1128, 1129, 1130 1131, 1132, 1136 1137, 1138, 1140 1141, 1142, 1143 1144, 1145, 1146 |
1105, 1106, 1107 1108, 1109, 1114 1115, 1116, 1117 1118, 1119, 1121 1138, 1141, 1142 |
|
Neomycin |
1106, 1107, 1108 1109, 1110, 1111 1112, 1113, 1114 1115, 1116, 1117 1119, 1138, 1141 |
1128, 1129, 1130 1131, 1132, 1136 1137, 1138, 1140 1141, 1142, 1143 1144, 1145, 1146 |
1103, 1105, 1106 1107, 1108, 1109 1113, 1114, 1115 1116, 1117, 1119 1120, 1121, 1124 1138, 1141, 1142 |
|
Paromomycin |
1106, 1107, 1108 1109, 1110, 1111 1112, 1113, 1114 1115, 1116, 1117 1118, 1119, 1120, 1124, 1141, 1142 |
1128, 1130, 1131 1132, 1135, 1136 1137, 1138, 1140 1141, 1142, 1143 1144, 1145 |
1103, 1105, 1106 1107, 1108, 1109 1110, 1111, 1112 1113, 1114, 1115 1116, 1117, 1119 1124, 1138, 1141 1142 |
|
Hygromycin |
1128, 1130, 1131 1132, 1136, 1137 1138, 1140, 1141 1143, 1144, 1145 1146 |
1128, 1129, 1130 1131, 1132, 1136 1137, 1138, 1140 1141, 1142, 1143 1144, 1145, 1146 |
1103, 1105, 1106 1107, 1108, 1109 1114, 1115, 1116 1117, 1118, 1119 1121, 1138, 1141 1142 |
|
Streptomycin |
1128, 1129, 1130 1131, 1132, 1136 1137, 1138, 1140 1141, 1142, 1143 1144, 1145, 1146 |
1128, 1130, 1131 1132, 1136, 1137 1138, 1139, 1140 1141, 1142, 1143 1144, 1145, 1146 |
1103, 1105, 1106 1107, 1108, 1109 1110, 1111, 1112 1113, 1114, 1115 1116, 1117, 1119 1121, 1124, 1138 1141, 1142 |
| Kanamycin | 1106, 1107, 1108 1109, 1110, 1111 1112, 1113, 1114 1115, 1116, 1117 1118, 1119, 1120 1124 |
1103, 1104, 1105 1106, 1107, 1108 1109, 1110, 1113 1118, 1119, 1120 1121 |
1103, 1105, 1106 1108, 1109, 1110 1111, 1112, 1113 1114, 1115, 1116 1117, 1118, 1119 1124, 1138, 1141 |
Table 4.
ΔGb values for the interactions between aminoglycosides and the most representative megalin cluster structures derived from molecular dynamics simulations, calculated at pH 7.0 using APBS [21] and VMD 1.9.1 [22].
| System |
ΔGsolv (kJ/mol) |
ΔGCoul (kJ/mol) |
ΔGnon-elec (kJ/mol) |
ΔGb* (kJ/mol) |
|
|---|---|---|---|---|---|
| Megalin-Gentamicin | |||||
| Cluster 1 | 101 | -1149 | -11 | -1059 | |
| Cluster 2 | 158 | -1071 | -14 | -927 | |
| Cluster 3 | 131 | -949 | -13 | -831 | |
| Mean | 130(29) | -1056(101) | -13(2) | -939(114) | |
| Megalin-Tobramycin | |||||
| Cluster 1 | 138 | -1161 | -10 | -1033 | |
| Cluster 2 | 65 | -1071 | -13 | -1019 | |
| Cluster 3 | 165 | -661 | -12 | -508 | |
| Mean | 123(52) | -833(284) | -12(2) | -722(275) | |
| Megalin-Neomycin | |||||
| Cluster 1 | 165 | -778 | -14 | -627 | |
| Cluster 2 | 166 | -704 | -14 | -552 | |
| Cluster 3 | 191 | -722 | -14 | -545 | |
| Mean | 174(15) | -735(39) | -14(0) | -575(45) | |
| Megalin-Paromomycin | |||||
| Cluster 1 | 118 | -743 | -15 | -640 | |
| Cluster 2 | 105 | -564 | -14 | -473 | |
| Cluster 3 | 217 | -560 | -15 | -358 | |
| Mean | 147(61) | -622(105) | -15(1) | -490(142) | |
| Megalin-Hygromycin | |||||
| Cluster 1 | 116 | -750 | -11 | -645 | |
| Cluster 2 | 60 | -571 | -13 | -524 | |
| Cluster 3 | 96 | -354 | -13 | -271 | |
| Mean | 91(28) | -558(198) | -12(1) | -480(191) | |
| Megalin-Streptomycin | |||||
| Cluster 1 | 76 | -694 | -12 | -630 | |
| Cluster 2 | 49 | -589 | -13 | -553 | |
| Cluster 3 | 115 | -336 | -14 | -235 | |
| Mean | 80(33) | -540(184) | -13(1) | -473(209) | |
| Megalin-Kanamicyn | |||||
| Cluster 1 | 79 | -585 | -13 | -519 | |
| Cluster 2 | 132 | -301 | -13 | -182 | |
| Cluster 3 | 115 | -473 | -13 | -371 | |
| Mean | 109(27) | -453(143) | -13(0) | -357(169) | |
*The binding energy (ΔGb) is given by Eq (1).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



