
Submitted:
31 March 2025
Posted:
01 April 2025
You are already at the latest version
Abstract
Climate change accelerates the competitive displacement of native plants by non-native plants in wetland ecosystems, critically affecting the diversity of wetland plant communities. However, it remains unclear how variation in climate change factors and non-native plants may impact the diversity of native plant communities in the Ertix River Basin wetland ecosystems. As a result, we selected 161 representative wetland plant communities for plot surveys and community classification, including the community structure, species diversity patterns, and the current status of non-native plant species in the Ertix River Basin wetlands, as the region faces serious plant invasion threats in recent decades. We also measured the elevation, mean annual precipitation, and mean annual temperature pattern to assess the impact of environmental factors on the dynamics of relationships between native plant communities and non-native plants. Results of Patrick richness, and Shannon-Wiener indices of native plant communities significantly increased with increasing temperature and elevation of the Ertix River Basin wetlands. The Patrick richness index, Pielou index and Shannon-Wiener index of native plant species positively correlated with the increasing number of naturalized individuals or species. Meanwhile, the number of casual plants, invasive plants, and naturalized plants negatively correlated with the importance value of native plant communities. Overall, results suggest that environmental variability and plant invasion can shift the diversity and distribution pattern of native plant communities in the Ertix River Basin wetlands.
Keywords:
alien plants
; elevational gradient
; environmental factors
; Ertix River Basin
; native plant diversity
; wetland plant communities
Introduction
Climate change accelerates the competitive displacement of native plants by invasive alien plants in many ecosystems, including wetlands [1,2], critically driving biodiversity loss in wetland habitats worldwide [3,4,5]. The colonization of non-native plants in wetland ecosystems has dramatically altered vital attributes of native plant diversity, i.e., species richness and evenness [5,6]. Such invasion-driven effects on native plants have been reported in many wetland ecosystems in China due to favorable and variable climatic conditions [7,8,9], causing enormous ecological and economic losses over the years [10]. For instance, biological invasion has been estimated to result in approximately USD 14.5 billion annual economic cost in China [11]. Despite these negative ramifications on wetland ecosystems in China, how climate change-driven impacts and invasive alien plants may affect the diversity of native plant communities in the Ertix River Basin wetlands remain unclear.
There is a growing awareness of the varying ecological impacts of non-native plants on species composition and diversity of native plant communities [12]. The mechanisms underpinning such ecological impacts of alien plants have been attributed to their superior competitive ability, resource use efficiency, and inherent environmental adaptability [13,14,15]. Additionally, non-native plants are known for their capacity to modify the biotic and abiotic characteristics of invaded soils through priority effects [16] and allelochemical inputs [17,18], creating favorable soil conditions for their growth and fitness. As a result, non-native plants can rapidly colonize, establish, and dominate native plant communities [19,20]. For instance, previous studies indicate that invasive alien plants alter soil biogeochemical processes and functional diversity of soil biota composition of the invaded ecosystems [21,22]. It is worth noting, however, that the resultant effects of the invasibility of native communities also depend on the diversity of native communities, resource availability, and the scale of the experimental study [23,24]. Thus, exhibiting the diversity-invasibility relationship and inconsistent or bi-directional effects of native and invasive plants in natural ecosystems [24,25,26]. While highly diverse native plant communities mediate biotic resistance to alien plant invasion at large scales, the opposite is true for low diverse plant communities at small scales [24,25,26]. Although significant advancements in mechanisms mediating the invasibility of local communities have been studied, how environmental variability contributes to the invasibility of native plant communities in wetland ecosystems remains understudied.
Plants show varying sensitivity and responses to environmental factors such as temperature and precipitation along elevational gradients [27,28], profoundly contributing to the distribution of plant communities [29]. Soil temperature and moisture are known to modulate key soil microbial processes, organic matter decomposition, and nutrient availability [30,31]. For example, Niu et al. found that decreased soil moisture content and increased soil temperature significantly reduced species richness of alpine plant communities [31]. Although climate warming promotes plant biomass [30,32], it significantly reduces species diversity across continental scales among wetland communities [30]. Indeed, as plants, either natives or non-natives, differ in their sensitivity to environmental variation [33,34], responses along elevational gradient may vary accordingly. While invasive plants are known to expand their range towards the north in response to climate warming [35,36], how climate change and invasive alien plants may interactively affect the diversity and distribution of native communities in wetland communities along elevational gradient remain inadequately explored.
Wetland ecosystems are usually located between elevational gradients where the aquatic and terrestrial ecosystems intersect [37], creating unique and distinct soil characteristics supporting the spatial distribution of diverse plant species [37]. The distinctive soil characteristics and favorable environmental conditions also attract many human activities, including agriculture, potentially modifying soil biogeochemical attributes [38,39]. As environmental disturbances are shown to mediate influxes and establishment of invasive species [40,41], human-driven modifications of the wetland ecosystems may likely initiate the interaction of invasive species and native plant communities [40,42]. Similarly, the wetland ecosystems of the Ertix River Basin, with its rich biodiversity, represent a conducive habitat for many plant species [43]. Therefore, under the ongoing rapidly changing global environment, the Ertix River Basin wetlands may be exposed to large influxes of non-native plants, ultimately posing a significant threat to native plant communities. However, the information regarding the magnitude and direction of impacts of non-native plant invasion on the diversity of native plant communities, distribution patterns, and invasion status in the Ertix River Basin wetlands remains limited.
We, therefore, conducted a field study to examine the impact of non-native plants on native plant communities in the Ertix River Basin wetland ecosystems. Specifically, we asked (1) whether non-native plant species affect native plant communities in the wetlands of the Ertix River Basin, (2) what the magnitude and direction of such non-native plants impact the diversity of native plants, (3) whether environmental factors, i.e., mean annual temperature and precipitation, drive non-native plant species impact along elevational gradients in the Ertix River Basin wetland ecosystems.
Results
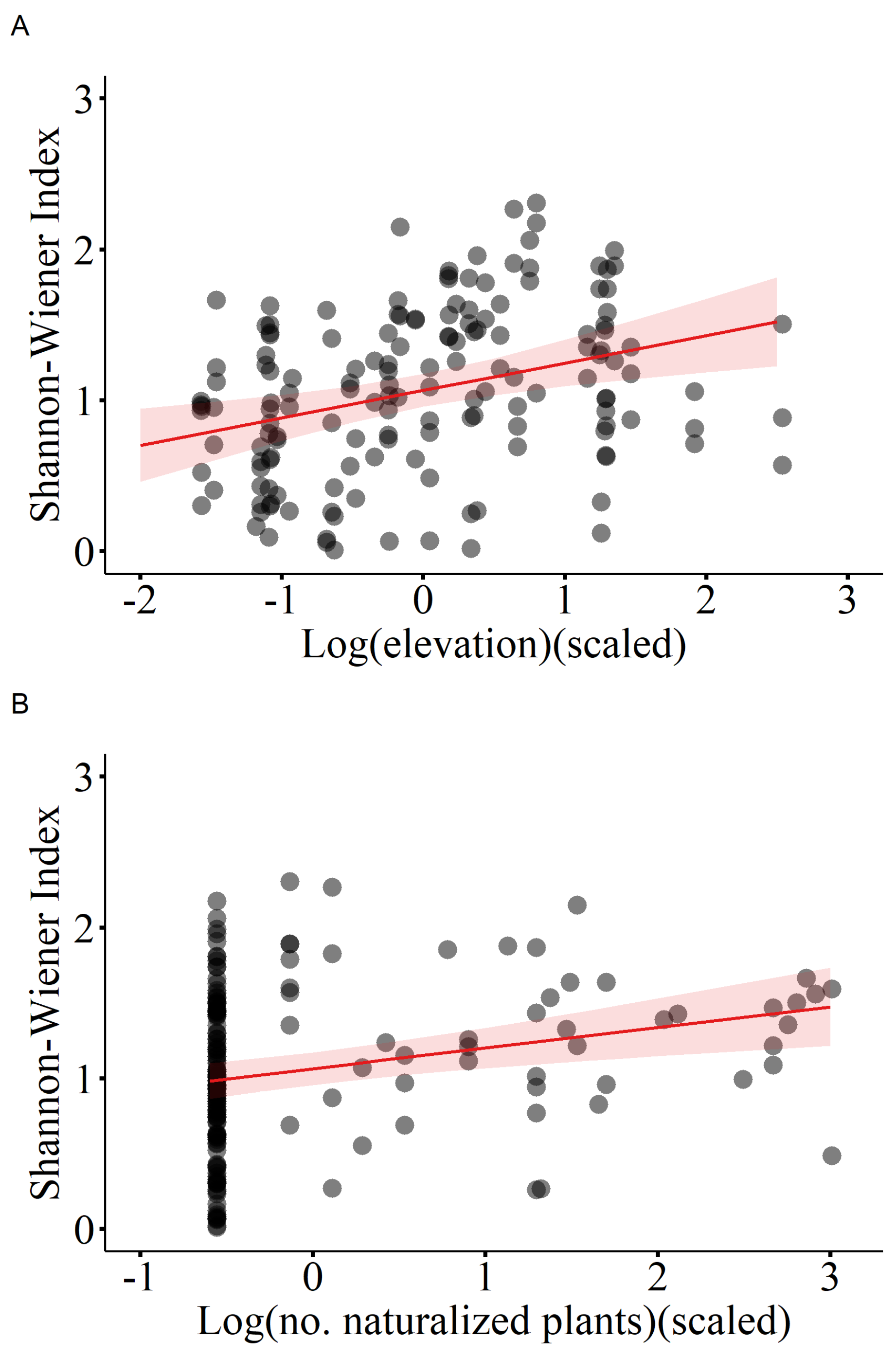
Results of the linear mixed-effects model (LMM) with importance value as the dependent variable indicate that the number of casual plants, invasive plants, and naturalized plants significantly affected the importance value of native plant communities, thus all showing a negative correlation (Table 1, Figure 2). Additionally, elevation was significantly and positively correlated with the importance value (Table 1, Figure 2). The linear mixed-effects model with the Patrick richness index of native plant communities as the dependent variable showed that increases in temperature, elevation, and number of naturalized species significantly enhanced the Patrick richness index of native plant communities (Table 2, Figure 3). The Pielou evenness index of native species communities also increased dramatically with rising number of naturalized plants (Table 3, Figure 4). In the linear mixed-effects model based on the Shannon-Wiener index of native species, elevation and number of naturalized plants were significantly and positively correlated with the Shannon-Wiener index (Table 4, Figure 5).
Figure 1.
Study area and sampling plots. The sampling plots were selected primarily based on the geographical distribution of the Irtysh River Basin and natural vegetation community patterns, with a total of 54 transects established along moisture gradients.
Figure 1.
Study area and sampling plots. The sampling plots were selected primarily based on the geographical distribution of the Irtysh River Basin and natural vegetation community patterns, with a total of 54 transects established along moisture gradients.
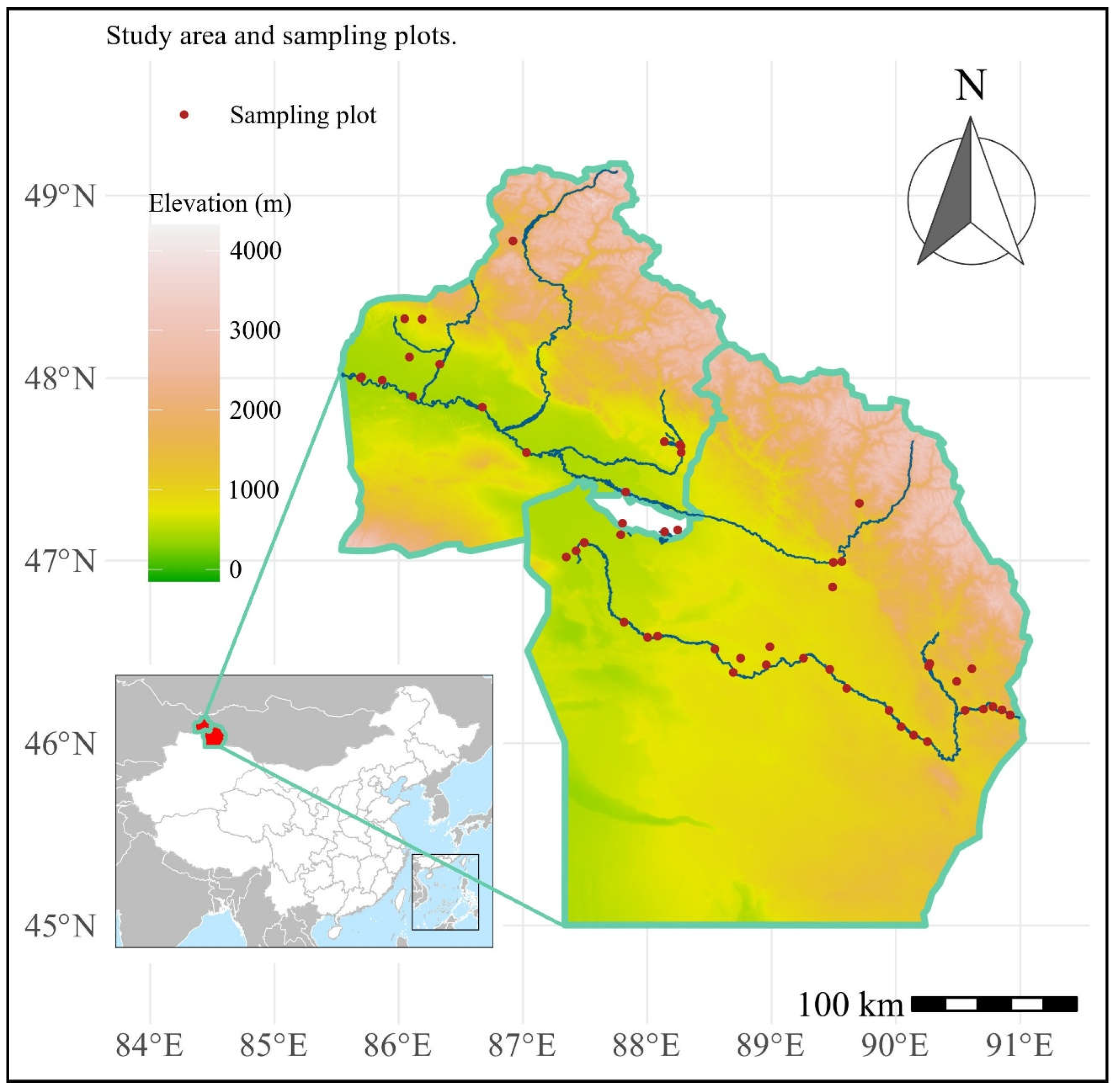
Figure 2.
Importance value of native species in relation to (A) elevation, (B) number of casual plants, (C) number of invasive plants, and (D) number of naturalized plants. The fitted lines with 95% confidence intervals from the minimum adequate linear mixed model (LMM) model are shown.
Figure 2.
Importance value of native species in relation to (A) elevation, (B) number of casual plants, (C) number of invasive plants, and (D) number of naturalized plants. The fitted lines with 95% confidence intervals from the minimum adequate linear mixed model (LMM) model are shown.
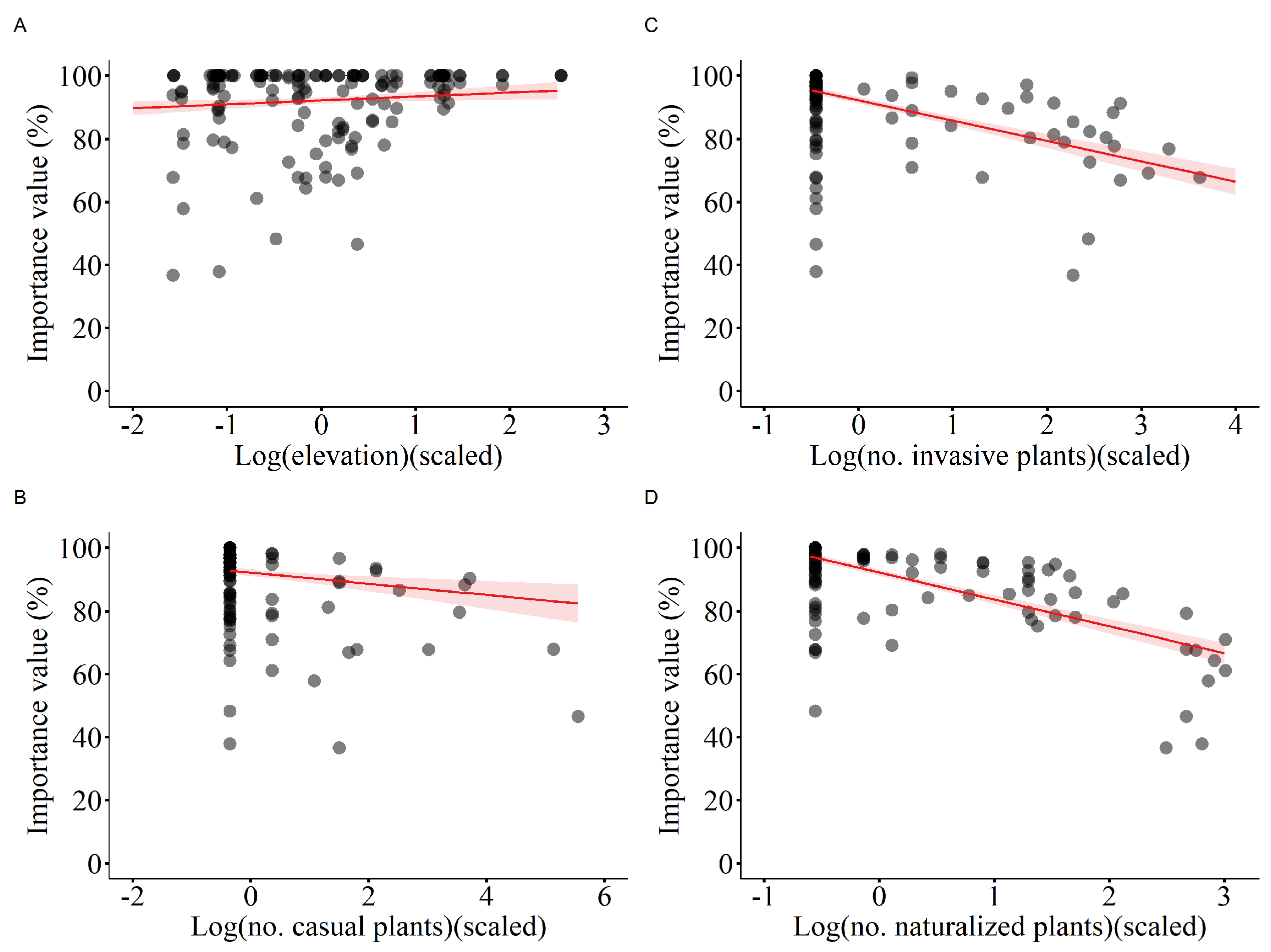
Figure 3.
Richness of native species in relation to (A) elevation, (B) mean annual temperature, and (C) number of naturalized species. The fitted lines with 95% confidence intervals from the minimum adequate LMM model are shown.
Figure 3.
Richness of native species in relation to (A) elevation, (B) mean annual temperature, and (C) number of naturalized species. The fitted lines with 95% confidence intervals from the minimum adequate LMM model are shown.
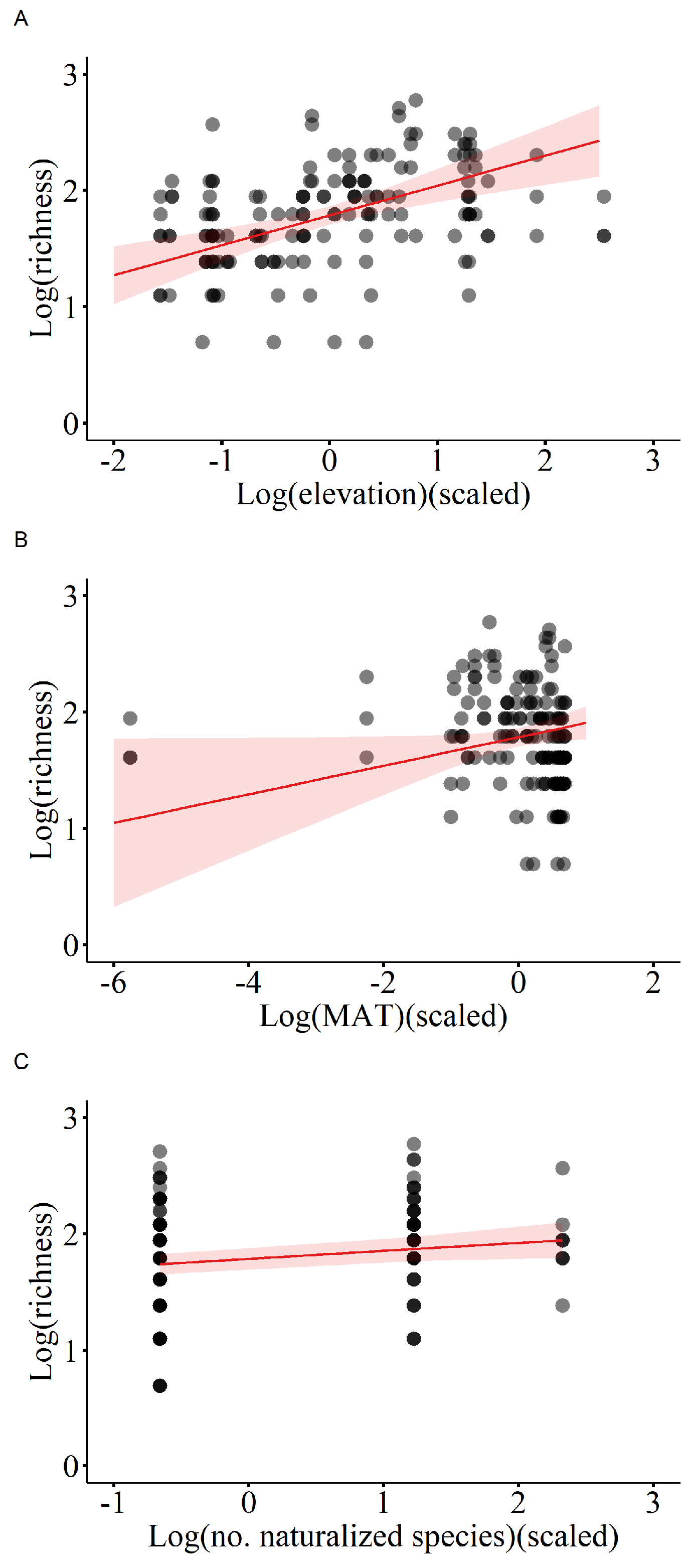
Figure 4.
Evenness of native species in relation to number of naturalized plants. The fitted lines with 95% confidence intervals from the minimum adequate LMM model are shown.
Figure 4.
Evenness of native species in relation to number of naturalized plants. The fitted lines with 95% confidence intervals from the minimum adequate LMM model are shown.
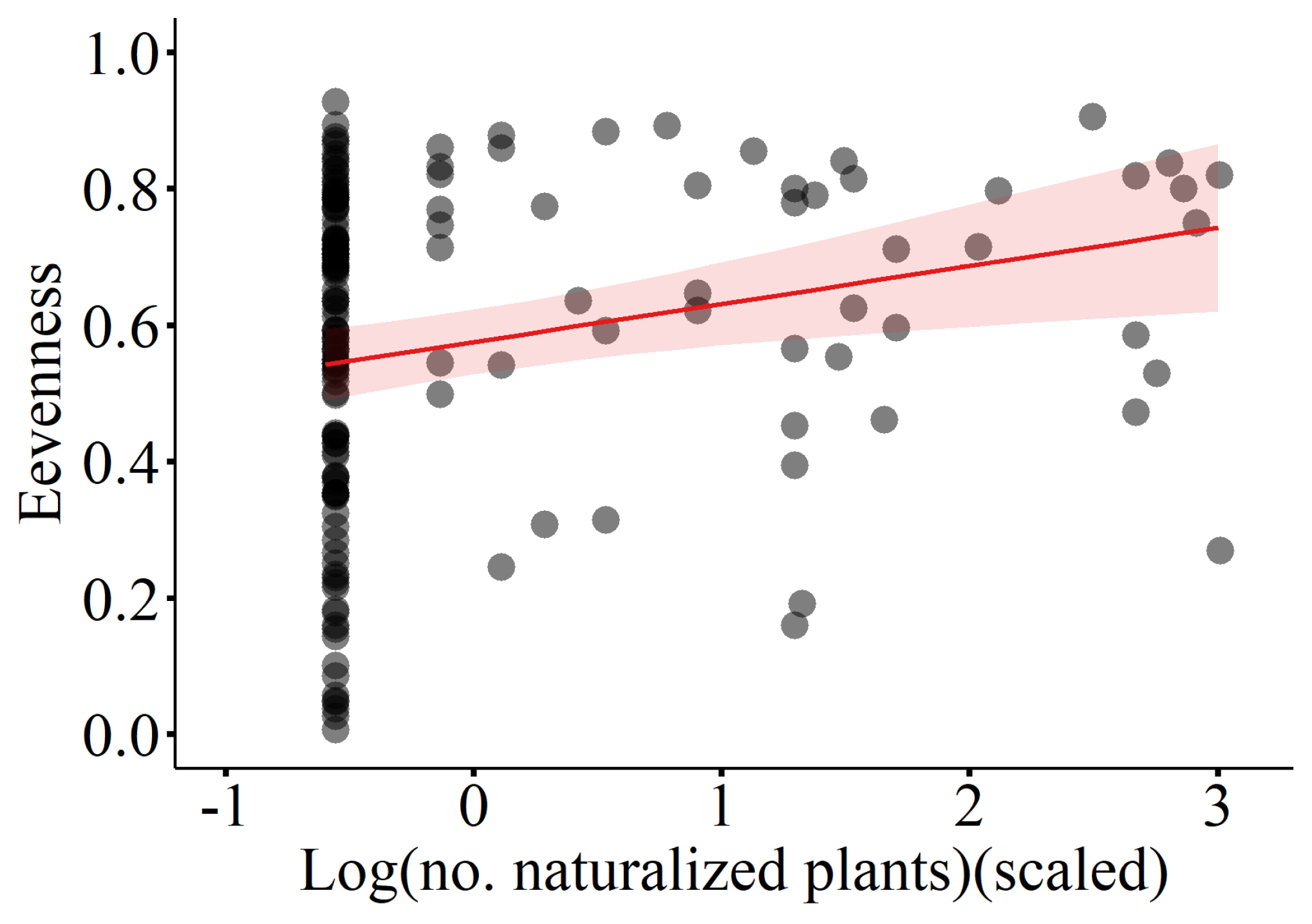
Figure 5.
Shannon of native species in relation to (A) elevation, (B) number of naturalized plants. The fitted lines with 95% confidence intervals from the minimum adequate LMM model are shown.
Figure 5.
Shannon of native species in relation to (A) elevation, (B) number of naturalized plants. The fitted lines with 95% confidence intervals from the minimum adequate LMM model are shown.
Discussion
Historically, non-native plants are known to impact native plant communities in many ecosystems [5,12,14,15,44]. Common natural ecosystems that are mostly invaded include wetlands due to their unique and distinctive environmental conditions [7,45], favoring human activities such as agriculture [46]. Such disturbance-mediated influxes of non-native plants, especially under the ongoing climate change [47], induce negative ramifications on native plant diversity [48]. However, little is known about how climate change and non-native plants affect the diversity and distribution pattern of native plants [49,50] in the Ertix River Basin wetland ecosystems. Our results provide empirical evidence that non-native plants affect native plant communities of the Ertix River Basin wetlands via additive effects of environmental factors and alien plants.
Results indicate that non-native plants affect native plant communities in the Ertix River Basin wetlands as the increase in the number of casual, naturalized, and invasive plants significantly reduced the importance value of native species. These findings consistently confirmed most previously documented findings on the negative impacts of non-native plant invasions on native plants in terrestrial [51,52,53] and wetland ecosystems [1,5,6]. The widely recognized mechanisms underlying such destructive trends in their invaded communities revolved around their inherent superior competitive abilities, resource use dominance, and fast growth rate, usually outperforming the native plant communities [13,14,15].
Most previous studies fail to distinguish casual, naturalized, and invasive species and account for their respective roles in native ecosystems, limiting our capacity to predict their contributions to the invasion processes [54]. Regardless, the disruptive characteristics of non-native plants remain inevitable in many native plant communities [55]. In a meta-analysis of 21 habitat types, for example, two times greater negative impacts of non-native species on native communities and five times higher performance of non-native than native species [55]. Nevertheless, alien plants are reported to play facilitative or complementary roles in native plant communities [56]. For example, in the southern California ecosystem, the native Cryptantha muricata produced more than twice the flowers, reproductive biomass, and total biomass when grown in the presence of non-native grasses than in their absence [57].
Likewise, in the present study, an increasing number of naturalized species showed a significantly positive correlation with the richness of the native plant communities. Also, results showed that the number of naturalized plants was positively correlated with the evenness and Shannon index of native plant communities, which again highlights the promoting effect of alien plants on native plant communities. Similarly, an early-stage mild invasion of the invasive plant Alternanthera philoxeroides increased species richness and evenness among the native community [58]. Such promotional effects of non-native species on native communities have been reported in Solidago canadensis [59] and Ambrosia trifida [60]. Our results suggest that alien plants may promote multi-species coexistence by altering resource utilization or mediating the resistance of native plants against common disturbances within the Ertix River Basin wetland ecosystems.
Furthermore, results indicate that environmental factors (i.e., elevation and temperature) exhibit a more significant correlation with Shannon-Wiener and Patrick richness, than alien plants. The richness of native plants significantly increased with increasing temperature. These results replicate most previous studies that observed significant effects of MAT on the diversity and pattern of distribution of native plant communities [61]. For example, Li et al. found a significant correlation between temperature and the overall richness and abundance of shrubs and trees in montane forests [61].
Environmental factors, such as temperature and precipitation, directly and indirectly affect plants via modulating plant resource uptake and utilization, metabolic and physiological processes, and plant-microorganism interactions [62]. Thus, within the maximum temperature threshold, increasing temperatures can promote plant resource use efficiency and competitive interaction [63,64], critically shaping the life histories and dynamics of native plant communities [61]. Despite temperature and precipitation are vital predictors of plant performance [65,66], both are shown to hinder the migration capacity and adaptability of plants [61], otherwise alien plants, due to climatic variability consequences on species coexistences or interspecific relationships [67,68,69]. Therefore, the observed strong correlation between increasing native plant diversity and rising temperatures along an elevational gradient in the present study may ultimately limit the invasibility of the Ertix River Basin wetland ecosystems. Thus, exhibiting the positive diversity-invasibility or biotic resistance scenario [23,25,26] in the Ertix River Basin wetland ecosystem. As another potent predictor determining climatic impacts on plant growth, precipitation in our study had less effect on native plants than temperature. Such observation was not surprising, especially when the experimental or survey setting is within wetland communities, suggesting that precipitation may least limit native plant diversity [65,66,70].
Plants, natives or invasives, differ in their sensitivity to climatic variability [33,71], substantially exhibiting spatial-structuring patterns of plant diversity and distribution along elevational gradients [71,72,73]. Notably, plant species diversity increases with altitude, especially with the native taxa [71]. Such plants develop unique adaptation strategies, including thick leaves, specialized root systems, and shorter growth forms [74], facilitating their resistance to varying elevation-induced stressors. In the present study, a significant positive correlation between the diversity of native plant communities and increasing elevation was observed, which is consistent with most previous findings [75,76]. For example, as native species richness increased along an increasing gradient in the Mediterranean island, alien plants predominantly occupied the lower elevations [76], suggesting a facilitative role of invasive plants via shielding natives from harsh environmental forces [56].
As indicated earlier, with substantial climatic variability consequences on new arrival species (i.e., alien plants) in mountainous ecosystems [67,68,69,77], native plant diversity and environmental factors can interactively shape the coexistences of alien plants in the Ertix River Basin wetland ecosystems. Overall, results provide empirical evidence that native plants may occupy more ecological niches within the Ertix River Basin wetland communities, enhancing resource use efficiency and providing competitive resistance to alien plant invasions.
Materials and Methods
Study Area
The Ertix River Basin wetlands are located in the northern region of the Xinjiang Uygur Autonomous Region, China. It is bordered by the Altai Mountains to the north, the Sayram Mountains to the southwest, and the Gurbantunggut Desert to the south. The expansive alluvial plain of the Ertix River and the Ulungur River lies at the center of this region. The Ertix River is the second-largest river in Xinjiang and an international river within the Arctic Ocean watershed [78]. The Ertix River Basin spans 85°30′ to 90°30′ E and 46°55′ to 49°10′ N, with elevations ranging from 1,337 m to 3,846 m above sea level. The river system extends over an area of 1,640,000 km² with a length of 4,248 km, of which 633 km flows through China territory, encompassing a basin area of 52,730 km². Located in the northwest arid region of China, the basin is characterized by a temperate continental arid to semi-arid climate and receives an annual mean temperature of 3.3°C and an average annual precipitation of 93.9 mm. [78,79,80].
Field Survey
On 23 June 2022 and 28 July 2022, we conducted a field survey in the Ertix River Basin wetland ecosystems (see Figure 1). The survey site was primarily selected based on the geographical distribution of the Ertix River and natural plant community patterns. By employing a combination of transect and quadrat methods, we established 54 transects with three randomly placed 1 m × 1 m quadrats along each transect to capture different community types across moisture gradients, totaling 161 quadrants. We recorded geographic coordinates and the herbaceous plant species composition for each quadrat, including species names, abundance, and coverage. We calculated the average height of species by summing all individual species’ heights and dividing them by their total number.
Species Name Standardization
Each plant species name was standardized, and their invasion status was determined through a multi-step process. Thus, (1) we identified the species on-site using the Flora of China (FOC; http://www.iplant.cn/) and subsequently verified by botanical experts’ consideration and judgment. (2) We standardized further the names of the species through the Plants of the World Online (POWO; https://powo.science.kew.org/), using the “rWCVP” package in R [81]. (3) For easy categorization, each standardized plant species was classified into four groups and defined as follows: native plants (i.e., the indigenous plant species), naturalized plants (i.e., introduced species that reproduce without human intervention but are not invasive), and casual plants (i.e., alien plants that are not naturalized), as well as those established and confirmed as invasive species. While the alien invasive plants were identified based on the checklist developed by Lin et al. [82], we identified the native plants based on the checklist developed by Lu et al. [83] and cross-checked via the POWO. Additionally, the naturalized and invasive plants were further identified based on the updated checklist from the Naturalized Plant of China (NPC; https://www.iplant.cn/npc/) and the Invasive Alien Species in China (IASC; https://www.plantplus.cn/ias/).
Environmental Factors
Elevation data were extracted from the National DEM dataset (1 km resolution) available through the Resource and Environmental Science Data Platform (https://www.resdc.cn/). We use the “raster” package in R to extract the elevation data for specific coordinate points based on the latitude and longitude of the sampling points. Moreover, the climate data, including monthly temperature and precipitation from 1901 to 2022 (1 km resolution) were obtained from the National Earth System Science Data Center (https://www.geodata.cn) [84]. We use the “ncdf4” package in R to extract the environmental factor data for specific coordinate points based on the latitude and longitude of sampling points. Both temperature and precipitation data from 2012 to 2022 were extracted for each year, and the mean annual temperature (MAT) was calculated by averaging the monthly temperatures. Likewise, we obtained the annual precipitation (MAP) by summing monthly precipitation values.
Species Diversity Metrics
From the field survey data, the relative height, relative abundance, and relative coverage of the native species in each plot were calculated (i.e., the proportion of the sum of the average heights, abundance, and coverage of native species relative to the sum of the average heights, abundance, and coverage of all species in the plot). Also, the importance value of species based on their relative heights, relative abundance, and relative coverage were calculated, further allowing us to determine the alpha diversity indices, i.e., the Patrick Richness Index, Shannon-Wiener Index, and Pielou Evenness Index [85,86]. All calculations were performed using the “vegan” package in R. The formulas are as follows:
Where IV represents importance value,rh is relative height,ra is relative abundance, and rc is relative coverage. Moreover, S denotes the total number of species in the community. Pi is the proportion of individuals of species i to the total number of individuals in the community, calculated as Pi = Ni / N. Ni represents the number of individuals of species i.
Data Analysis
For each quadrant, we recorded the total number of biological species of casual, naturalized, or invasive plants (hereafter referred to as the “number of casual, naturalized, or invasive species”) and the total number of individual casual, naturalized, or invasive plants (hereafter referred to as the “number of casual, naturalized, or invasive plants”). Prior to analysis, we log-transformed and scale-standardized all independent variables, and applied a log transformation to species richness. We used linear mixed-effects model (LMM) to examine the effect of environmental factors (elevation, MAT, and MAP), the number of non-native species (casual, naturalized, and invasive species), and the individual number of non-native plants (casual, naturalized, and invasive plants) on the Shannon index, species richness, importance value and evenness of the native plant community. The optimal model was obtained by manually retaining only significant factors, using the “lmer” function from the “lme4” package [87]. The individual marginal R2 of each fixed factors was estimated using “glmm.hp” function from the “glmm.hp” package[88]. All analyses were performed using R v4.4.0 [89].
Conclusions
We conclude that the invasion of alien plants of the Ertix River Basin wetlands significantly affects the diversity−Shannon, richness and evenness of native plant communities. However, a strong positive correlation between rising temperatures with the diversity of native plant communities along elevational gradients observed in this study may hinder the invasibility of the Ertix River Basin wetland ecosystems. Ultimately, it provides an essential clue of the positive diversity-invasibility or invasion resistance scenario, underscoring the need to bolster the diversity of native plants as management strategies against plant invasions in the Ertix River Basin wetland ecosystems. One caveat is that we did not analyze the taxonomic functional identity (e.g., grasses, forbs, and legumes species) of the various native plant communities, and thus, we cannot provide information regarding species-specific responses to the environmental variability and plant invasions in the Ertix River Basin wetlands. To comprehensively understand the distribution pattern, life-history strategies, and adaptation dynamics of native plants in wetlands and mountainous plant communities, future studies may consider climate- and human-driven disturbances across variable environmental factors when exploring the relationships between alien and native plant species.
Author Contributions
Conceptualization, B.-C.D., J.-Q.G. and F.-H.Y.; methodology, B.-C.D., J.-Q.G. and X.-M.C.; data curation, B.-C.D. and X.-M.C.; formal analysis, B.-C.D. and X.-M.C.; writing-original draft preparation, X.-M.C.; writing-review & editing, B.-C.D., J.-Q.G., O.-A.M., X.-M.C. and F.-H.Y.; supervision, B.-C. D. and J.-Q.G.. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (grant 31500331) and the Third Xinjiang Scientific Expedition Program (grant 2021xjkk0601).
Data Availability Statement
Raw data and R codes are available on request to the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Qiu, S.Y.; Liu, S.S.; Wei, S.J.; Cui, X.H.; Nie, M.; Huang, J.X.; He, Q.; Ju, R.T.; Li, B. Changes in Multiple Environmental Factors Additively Enhance the Dominance of an Exotic Plant with a Novel Trade-off Pattern. J. Ecol. 2020, 108, 1989–1999. [Google Scholar] [CrossRef]
- Ren, G.Q.; Cui, M.M.; Yu, H.C.; Fan, X.; Zhu, Z.Q.; Zhang, H.Y.; Dai, Z.C.; Sun, J.F.; Yang, B.; Du, D.L. Global Environmental Change Shifts Ecological Stoichiometry Coupling between Plant and Soil in Early-Stage Invasions. J. Soil. Sci. Plant Nutr. 2024, 24, 2402–2412. [Google Scholar] [CrossRef]
- Adomako, M.O.; Zhang, Q.; Yu, F.H. Genotypic Differences in Response to Different Patterns of Clonal Fragmentation in the Aquatic Macrophyte Pistia stratiotes. J. Plant Ecol. 2022, 15, 1199–1212. [Google Scholar] [CrossRef]
- Catford, J.A.; Bode, M.; Tilman, D. Introduced Species That Overcome Life History Tradeoffs Can Cause Native Extinctions. Nat. Commun. 2018, 9, 2131. [Google Scholar] [CrossRef]
- Ren, J.; Chen, J.; Xu, C.; Koppel, J. van de; Thomsen, M.S.; Qiu, S.; Cheng, F.; Song, W.; Liu, Q.-X.; Xu, C.; et al. An Invasive Species Erodes the Performance of Coastal Wetland Protected Areas. Sci. Adv. 2021. [Google Scholar] [CrossRef]
- Yang, R.M. Interacting Effects of Plant Invasion, Climate, and Soils on Soil Organic Carbon Storage in Coastal Wetlands. J. Geophys. Res. Biogeosciences 2019, 124, 2554–2564. [Google Scholar] [CrossRef]
- Chen, L. Invasive Plants in Coastal Wetlands: Patterns and Mechanisms. In Wetlands: Ecosystem Services, Restoration and Wise Use; An, S., Verhoeven, J.T.A., Eds.; Springer International Publishing: Cham, 2019; pp. 97–128. ISBN 978-3-030-14861-4. [Google Scholar]
- Yang, Y.B.; Bian, Z.H.; Ren, W.J.; Wu, J.H.; Liu, J.Q.; Shrestha, N. Spatial Patterns and Hotspots of Plant Invasion in China. Glob. Ecol. Conserv. 2023, 43, e02424. [Google Scholar] [CrossRef]
- Dong, B.C.; Dong, R.; Yang, Q.; Kinlock, N.L.; Yu, F.H.; van Kleunen, M. Predicting Invasion Success of Naturalized Cultivated Plants in China. J. Appl. Ecol. 2025, 62, 651–660. [Google Scholar] [CrossRef]
- Fang, X.H.; Chen, X.M.; Liu, R.L.; Zhang, S.H.; Gao, J.Q.; Dong, B.C.; Yu, F.H. Effects of Alligator Weed Invasion on Wetlands in Protected Areas: A Case Study of Lishui Jiulong National Wetland Park. Sci. Total Environ. 2024, 953, 176230. [Google Scholar] [CrossRef]
- Xu, H.G.; Ding, H.; Li, M.; Qiang, S.; Guo, J.Y.; Han, Z.M.; Huang, Z.G.; Sun, H.Y.; He, S.P.; Wu, H.R.; et al. The Distribution and Economic Losses of Alien Species Invasion to China. Biol. Invasions 2006, 8, 1495–1500. [Google Scholar] [CrossRef]
- Early, R.; Bradley, B.A.; Dukes, J.S.; Lawler, J.J.; Olden, J.D.; Blumenthal, D.M.; Gonzalez, P.; Grosholz, E.D.; Ibañez, I.; Miller, L.P.; et al. Global Threats from Invasive Alien Species in the Twenty-First Century and National Response Capacities. Nat. Commun. 2016, 7, 12485. [Google Scholar] [CrossRef] [PubMed]
- Funk, J.L.; Vitousek, P.M. Resource-Use Efficiency and Plant Invasion in Low-Resource Systems. Nature 2007, 446, 1079–1081. [Google Scholar] [CrossRef]
- Li, J.; He, J.Z.; Liu, M.; Yan, Z.Q.; Xu, X.L.; Kuzyakov, Y. Invasive Plant Competitivity Is Mediated by Nitrogen Use Strategies and Rhizosphere Microbiome. Soil Biol. Biochem. 2024, 192, 109361. [Google Scholar] [CrossRef]
- Zhang, Y.L.; Zhang, Y.Y.; Zhang, L.H.; Yang, H.J.; Ma, T.X.; Huang, C.Y.; Hu, L.Y.; Zhang, M.X.; Sang, W.G. Enhanced Competitive Advantage of Invasive Plants by Growth-Defence Trade-off: Evidence from Phytohormone Metabolism and Transcriptomic Analysis. J. Appl. Ecol. 2025, 2025, 1–13. [Google Scholar] [CrossRef]
- Hess, M.C.M.; Mesléard, F.; Buisson, E. Priority Effects: Emerging Principles for Invasive Plant Species Management. Ecol. Eng. 2019, 127, 48–57. [Google Scholar] [CrossRef]
- Zhang, Z.J.; van Kleunen, M. Common Alien Plants Are More Competitive than Rare Natives but Not than Common Natives. Ecol. Lett. 2019, 22, 1378–1386. [Google Scholar] [CrossRef]
- Adomako, M.O.; Ning, L.; Tang, M.; Du, D.L.; van Kleunen, M.; Yu, F.H. Diversity- and Density-Mediated Allelopathic Effects of Resident Plant Communities on Invasion by an Exotic Plant. Plant Soil 2019, 440, 581–592. [Google Scholar] [CrossRef]
- Hou, M.; Wang, J.Y. Functional Traits of Both Specific Alien Species and Receptive Community but Not Community Diversity Determined the Invasion Success under Biotic and Abiotic Conditions. Funct. Ecol. 2023, 37, 2598–2610. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, T.; Liu, Y.J. Effects of Arbuscular Mycorrhizal Fungi on Plant Invasion Success Driven by Nitrogen Fluctuations. J. Appl. Ecol. 2023, 60, 2425–2436. [Google Scholar] [CrossRef]
- Yang, Z.Y.; Yang, L.F. Impacts of invasion plant on soil ecological environment. Adv. Environ. Prot. 2020, 10, 194. [Google Scholar] [CrossRef]
- Wang, X.Y.; Wang, X.; Wang, W.; Wang, J.; Yu, F.H. Effects of Invasive Plant Diversity on Soil Microbial Communities. Diversity 2022, 14, 992. [Google Scholar] [CrossRef]
- Jauni, M.; Hyvönen, T. Positive Diversity–Invasibility Relationships across Multiple Scales in Finnish Agricultural Habitats. Biol. Invasions 2012, 14, 1379–1391. [Google Scholar] [CrossRef]
- Zeiter, M.; Stampfli, A. Positive Diversity–Invasibility Relationship in Species-Rich Semi-Natural Grassland at the Neighbourhood Scale. Ann. Bot. 2012, 110, 1385–1393. [Google Scholar] [CrossRef] [PubMed]
- Lambers, J.H.R.; Harpole, W.S.; Tilman, D.; Knops, J.; Reich, P.B. Mechanisms Responsible for the Positive Diversity–Productivity Relationship in Minnesota Grasslands. Ecol. Lett. 2004, 7, 661–668. [Google Scholar] [CrossRef]
- Fridley, J.D.; Stachowicz, J.J.; Naeem, S.; Sax, D.F.; Seabloom, E.W.; Smith, M.D.; Stohlgren, T.J.; Tilman, D.; Holle, B.V. The Invasion Paradox: Reconciling Pattern and Process in Species Invasions. Ecology 2007, 88, 3–17. [Google Scholar] [CrossRef]
- Souza, L.F.T.; Billings, S.A. Temperature and pH Mediate Stoichiometric Constraints of Organically Derived Soil Nutrients. Glob. Change Biol. 2022, 28, 1630–1642. [Google Scholar] [CrossRef] [PubMed]
- Ren, G.Q.; Du, Y.Z.; Yang, B.; Wang, J.J.; Cui, M.M.; Dai, Z.C.; Adomako, M.O.; Rutherford, S.; Du, D.L. Influence of Precipitation Dynamics on Plant Invasions: Response of Alligator Weed (Alternanthera philoxeroides) and Co-Occurring Native Species to Varying Water Availability across Plant Communities. Biol. Invasions 2023, 25, 519–532. [Google Scholar] [CrossRef]
- Jafari, M.; Chahouki, M.A.Z.; Tavili, A.; Azarnivand, H.; Amiri, Gh.Z. Effective Environmental Factors in the Distribution of Vegetation Types in Poshtkouh Rangelands of Yazd Province (Iran). J. Arid Environ. 2004, 56, 627–641. [Google Scholar] [CrossRef]
- Baldwin, A.H.; Jensen, K.; Schönfeldt, M. Warming Increases Plant Biomass and Reduces Diversity across Continents, Latitudes, and Species Migration Scenarios in Experimental Wetland Communities. Glob. Change Biol. 2014, 20, 835–850. [Google Scholar] [CrossRef]
- Niu, Y.J.; Yang, S.W.; Zhou, J.W.; Chu, B.; Ma, S.J.; Zhu, H.M.; Hua, L.M. Vegetation Distribution along Mountain Environmental Gradient Predicts Shifts in Plant Community Response to Climate Change in Alpine Meadow on the Tibetan Plateau. Sci. Total Environ. 2019, 650, 505–514. [Google Scholar] [CrossRef]
- Lin, D.L.; Xia, J.Y.; Wan, S.Q. Climate Warming and Biomass Accumulation of Terrestrial Plants: A Meta-Analysis. New Phytol. 2010, 188, 187–198. [Google Scholar] [CrossRef] [PubMed]
- Seddon, A.W.R.; Macias-Fauria, M.; Long, P.R.; Benz, D.; Willis, K.J. Sensitivity of Global Terrestrial Ecosystems to Climate Variability. Nature 2016, 531, 229–232. [Google Scholar] [CrossRef]
- Chen, B.-M.; Gao, Y.; Liao, H.X.; Peng, S.L. Differential Responses of Invasive and Native Plants to Warming with Simulated Changes in Diurnal Temperature Ranges. AoB Plants 2017, 9, plx028. [Google Scholar] [CrossRef]
- Iseli, E.; Chisholm, C.; Lenoir, J.; Haider, S.; Seipel, T.; Barros, A.; Hargreaves, A.L.; Kardol, P.; Lembrechts, J.J.; McDougall, K.; et al. Rapid Upwards Spread of Non-Native Plants in Mountains across Continents. Nat. Ecol. Evol. 2023, 7, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Zheng, M.M.; Pyšek, P.; Guo, K.; Hasigerili, *!!! REPLACE !!!*; Guo, W.Y. Clonal Alien Plants in the Mountains Spread Upward More Extensively and Faster than Non-Clonal. NeoBiota 2024, 29–48. [Google Scholar] [CrossRef]
- He, Q.; Li, Z.-A.; Daleo, P.; Lefcheck, J.S.; Thomsen, M.S.; Adams, J.B.; Bouma, T.J. Coastal Wetland Resilience through Local, Regional and Global Conservation. Nat. Rev. Biodivers. 2025, 1, 50–67. [Google Scholar] [CrossRef]
- Amundson, R. Soil Biogeochemistry and the Global Agricultural Footprint. Soil Secur. 2022, 6, 100022. [Google Scholar] [CrossRef]
- Peng, Z.H.; Qian, X.; Liu, Y.; Li, X.M.; Gao, H.; An, Y.N.; Qi, J.J.; Jiang, L.; Zhang, Y.R.; Chen, S.; et al. Land Conversion to Agriculture Induces Taxonomic Homogenization of Soil Microbial Communities Globally. Nat. Commun. 2024, 15, 3624. [Google Scholar] [CrossRef]
- Lembrechts, J.J.; Pauchard, A.; Lenoir, J.; Nuñez, M.A.; Geron, C.; Ven, A.; Bravo-Monasterio, P.; Teneb, E.; Nijs, I.; Milbau, A. Disturbance Is the Key to Plant Invasions in Cold Environments. Proc. Natl. Acad. Sci. 2016, 113, 14061–14066. [Google Scholar] [CrossRef]
- Jo, I.; Bellingham, P.J.; Mason, N.W.H.; McCarthy, J.K.; Peltzer, D.A.; Richardson, S.J.; Wright, E.F. Disturbance-Mediated Community Characteristics and Anthropogenic Pressure Intensify Understorey Plant Invasions in Natural Forests. J. Ecol. 2024, 112, 1856–1871. [Google Scholar] [CrossRef]
- Diniz, É.S.; Dias, F.S.; Borda-de-Água, L.; González, P.M.R. Anthropogenic Disturbance and Alien Plant Invasion Drive the Phylogenetic Impoverishment in Riparian Vegetation. Biodivers. Conserv. 2024, 33, 4237–4256. [Google Scholar] [CrossRef]
- Liu, S.D.; Liu, L.; Zhang, J.J.; Wang, K.; Guo, Y.Q. Study on ecological protection and restoration path of arid area based on improvement of ecosystem service capability, a case of the ecological protection and restoration pilot project aera in Irtysh River basin. Acta Ecol. Sin. 2019, 39, 8998–9007. [Google Scholar]
- Dong, B.C.; Yang, Q.; Kinlock, N.L.; Pouteau, R.; Pyšek, P.; Weigelt, P.; Yu, F.H.; van Kleunen, M. Naturalization of Introduced Plants Is Driven by Life-Form-Dependent Cultivation Biases. Divers. Distrib. 2024, 30, 55–70. [Google Scholar] [CrossRef]
- Adomako, M.O.; Alpert, P.; Du, D.L.; Yu, F.H. Effects of Fragmentation of Clones Compound over Vegetative Generations in the Floating Plant Pistia stratiotes. Ann. Bot. 2021, 127, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Adomako, M.O.; Alpert, P.; Du, D.L.; Yu, F.H. Effects of Clonal Integration, Nutrients and Cadmium on Growth of the Aquatic Macrophyte Pistia stratiotes. J. Plant Ecol. 2020, 13, 765–772. [Google Scholar] [CrossRef]
- Newbold, T.; Hudson, L.N.; Hill, S.L.L.; Contu, S.; Lysenko, I.; Senior, R.A.; Börger, L.; Bennett, D.J.; Choimes, A.; Collen, B.; et al. Global Effects of Land Use on Local Terrestrial Biodiversity. Nature 2015, 520, 45–50. [Google Scholar] [CrossRef]
- Lenda, M.; Skórka, P.; Knops, J.; Żmihorski, M.; Gaj, R.; Moroń, D.; Woyciechowski, M.; Tryjanowski, P. Multispecies Invasion Reduces the Negative Impact of Single Alien Plant Species on Native Flora. Divers. Distrib. 2019, 25, 951–962. [Google Scholar] [CrossRef]
- Turbelin, A.; Catford, J.A. Chapter 25 - Invasive Plants and Climate Change. In Climate Change (Third Edition); Letcher, T.M., Ed.; Elsevier, 2021; pp. 515–539. ISBN 978-0-12-821575-3. [Google Scholar]
- Garces, J.J.C.; Flores, M.J.L. Effects of Environmental Factors and Alien Plant Invasion on Native Floral Diversity in Mt. Manunggal, Cebu Island, Philippines. Curr. World Environ. 2018, 13, 390–402. [Google Scholar] [CrossRef]
- Dong, B.C.; Alpert, P.; Guo, W.; Yu, F.H. Effects of Fragmentation on the Survival and Growth of the Invasive, Clonal Plant Alternanthera philoxeroides. Biol. Invasions 2012, 14, 1101–1110. [Google Scholar] [CrossRef]
- Cheng, C.; Liu, Z.-K.; Song, W.; Chen, X.; Zhang, Z.J.; Li, B.; van Kleunen, M.; Wu, J. Biodiversity Increases Resistance of Grasslands against Plant Invasions under Multiple Environmental Changes. Nat. Commun. 2024, 15, 4506. [Google Scholar] [CrossRef] [PubMed]
- Nunez-Mir, G.C.; McCary, M.A. Invasive Plants and Their Root Traits Are Linked to the Homogenization of Soil Microbial Communities across the United States. Proc. Natl. Acad. Sci. 2024, 121, e2418632121. [Google Scholar] [CrossRef] [PubMed]
- Richardson, D.M.; Pyšek, P. Naturalization of Introduced Plants: Ecological Drivers of Biogeographical Patterns. New Phytol. 2012, 196, 383–396. [Google Scholar] [CrossRef]
- Kuebbing, S.E.; Nuñez, M.A. Invasive Non-Native Plants Have a Greater Effect on Neighbouring Natives than Other Non-Natives. Nat. Plants 2016, 2, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Cavieres, L.A. Facilitation and the Invasibility of Plant Communities. J. Ecol. 2021, 109, 2019–2028. [Google Scholar] [CrossRef]
- Pec, G.J.; Carlton, G.C. Positive Effects of Non-Native Grasses on the Growth of a Native Annual in a Southern California Ecosystem. PLoS One 2014, 9, e112437. [Google Scholar] [CrossRef]
- Wu, H.; Carrillo, J.; Ding, J.Q. Species Diversity and Environmental Determinants of Aquatic and Terrestrial Communities Invaded by Alternanthera philoxeroides. Sci. Total Environ. 2017, 581–582, 666–675. [Google Scholar] [CrossRef]
- Wang, C.Y.; Wu, B.D.; Jiang, K.; Zhou, J.W.; Du, D.L. Canada Goldenrod Invasion Affect Taxonomic and Functional Diversity of Plant Communities in Heterogeneous Landscapes in Urban Ecosystems in East China. Urban For. Urban Green. 2019, 38, 145–156. [Google Scholar] [CrossRef]
- Liu, R.L.; Yang, Y.B.; Lee, B.R.; Liu, G.; Zhang, W.G.; Chen, X.Y.; Song, X.J.; Kang, J.Q.; Zhu, Z.H. The Dispersal-Related Traits of an Invasive Plant Galinsoga quadriradiata Correlate with Elevation during Range Expansion into Mountain Ranges. AoB Plants 2021, 13, plab008. [Google Scholar] [CrossRef]
- Li, T.; Xiong, Q.L.; Luo, P.; Zhang, Y.B.; Gu, X.D.; Lin, B. Direct and Indirect Effects of Environmental Factors, Spatial Constraints, and Functional Traits on Shaping the Plant Diversity of Montane Forests. Ecol. Evol. 2020, 10, 557–568. [Google Scholar] [CrossRef]
- Kong, Z.; Wang, T.-J.; Han, Q.; Dai, Y.B.; Zuo, Y.T.; Wang, L.C.; Lang, Y.C. Impacts of Environmental Factors on Ecosystem Water Use Efficiency: An Insight from Gross Primary Production and Evapotranspiration Dynamics. Agric. For. Meteorol. 2025, 362, 110382. [Google Scholar] [CrossRef]
- Moore, C.E.; Meacham-Hensold, K.; Lemonnier, P.; Slattery, R.A.; Benjamin, C.; Bernacchi, C.J.; Lawson, T.; Cavanagh, A.P. The Effect of Increasing Temperature on Crop Photosynthesis: From Enzymes to Ecosystems. J. Exp. Bot. 2021, 72, 2822–2844. [Google Scholar] [CrossRef] [PubMed]
- Gillies, G.J.; Angert, A.L.; Usui, T. Temperature Dependence and Genetic Variation in Resource Acquisition Strategies in a Model Freshwater Plant. Funct. Ecol. 2024, 38, 1600–1610. [Google Scholar] [CrossRef]
- Moles, A.T.; Perkins, S.E.; Laffan, S.W.; Flores-Moreno, H.; Awasthy, M.; Tindall, M.L.; Sack, L.; Pitman, A.; Kattge, J.; Aarssen, L.W.; et al. Which Is a Better Predictor of Plant Traits: Temperature or Precipitation? J. Veg. Sci. 2014, 25, 1167–1180. [Google Scholar] [CrossRef]
- Butterfield, B.J.; Munson, S.M. Temperature Is Better than Precipitation as a Predictor of Plant Community Assembly across a Dryland Region. J. Veg. Sci. 2016, 27, 938–947. [Google Scholar] [CrossRef]
- Adler, P.B.; HilleRisLambers, J.; Kyriakidis, P.C.; Guan, Q.F.; Levine, J.M. Climate Variability Has a Stabilizing Effect on the Coexistence of Prairie Grasses. Proc. Natl. Acad. Sci. 2006, 103, 12793–12798. [Google Scholar] [CrossRef]
- García-Cervigón, A.I.; Quintana-Ascencio, P.F.; Escudero, A.; Ferrer-Cervantes, M.E.; Sánchez, A.M.; Iriondo, J.M.; Olano, J.M. Demographic Effects of Interacting Species: Exploring Stable Coexistence under Increased Climatic Variability in a Semiarid Shrub Community. Sci. Rep. 2021, 11, 3099. [Google Scholar] [CrossRef]
- Bonato, A.A.E.; Guimarães-Steinicke, C.; Stein, G.; Schreck, B.; Kattenborn, T.; Ebeling, A.; Posch, S.; Denzler, J.; Büchner, T.; Shadaydeh, M.; et al. Seasonal Shifts in Plant Diversity Effects on Above-Ground–below-Ground Phenological Synchrony. J. Ecol. 2025, 113, 472–484. [Google Scholar] [CrossRef]
- Korell, L.; Auge, H.; Chase, J.M.; Harpole, W.S.; Knight, T.M. Responses of Plant Diversity to Precipitation Change Are Strongest at Local Spatial Scales and in Drylands. Nat. Commun. 2021, 12, 2489. [Google Scholar] [CrossRef]
- Zu, K.L.; Chen, F.S.; Huang, C.; Liu, Y.Q.; Wang, F.C.; Zhu, G.J.; Bu, W.S.; Fang, X.M.; Guo, L.P. The Elevational Distribution Patterns of Plant Diversity and Phylogenetic Structure Vary Geographically across Eight Subtropical Mountains. Ecol. Evol. 2024, 14, e70722. [Google Scholar] [CrossRef]
- Chiu, M.C.; Ao, S.C.; He, F.Z.; Resh, V.H.; Cai, Q.H. Elevation Shapes Biodiversity Patterns through Metacommunity-Structuring Processes. Sci. Total Environ. 2020, 743, 140548. [Google Scholar] [CrossRef]
- Zhang, Z.J.; Liu, Y.J.; Hardrath, A.; Jin, H.F.; van Kleunen, M. Increases in Multiple Resources Promote Competitive Ability of Naturalized Non-Native Plants. Commun. Biol. 2022, 5, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Halbritter, A.H.; Fior, S.; Keller, I.; Billeter, R.; Edwards, P.J.; Holderegger, R.; Karrenberg, S.; Pluess, A.R.; Widmer, A.; Alexander, J.M. Trait Differentiation and Adaptation of Plants along Elevation Gradients. J. Evol. Biol. 2018, 31, 784–800. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.N.; Gesang, Q.Z.; Luo, J.F.; Wu, X.L.; Rebi, A.; You, Y.G.; Zhou, J.X. Drivers of Plant Diversification along an Altitudinal Gradient in the Alpine Desert Grassland, Northern Tibetan Plateau. Glob. Ecol. Conserv. 2024, 53, e02987. [Google Scholar] [CrossRef]
- Di Musciano, M.; Calvia, G.; Ruggero, A.; Farris, E.; Ricci, L.; Frattaroli, A.R.; Bagella, S. Elevational Patterns of Plant Species Richness and Phylogenetic Diversity in a Mediterranean Island. Perspect. Plant Ecol. Evol. Syst. 2024, 65, 125815. [Google Scholar] [CrossRef]
- García, F.C.; Bestion, E.; Warfield, R.; Yvon-Durocher, G. Changes in Temperature Alter the Relationship between Biodiversity and Ecosystem Functioning. Proc. Natl. Acad. Sci. 2018, 115, 10989–10994. [Google Scholar] [CrossRef]
- Liu, H.; Hao, H.G.; Zhang, W.G.; Liu, P.; Sun, L.H. Evaluation of ecological protection and restoration effectiveness based on “pattern-quality-service” in Irtysh River basin. Res. Environ. Sci. 2022, 35, 2495–2507. [Google Scholar] [CrossRef]
- Song, J.W.; Xu, Z.R.; Chen, J.X.; Xu, Q.H. Nutrient content of five natural poplar forests in the Irtysh River Basin in Xinjiang. Arid Zone Res. 2021, 38, 1429–1435. [Google Scholar] [CrossRef]
- Zhang, X.M.; Zhang, D.M.; Zhang, W. Effects of human activities on carbon storage in the Irtysh River Basin. Arid Zone Res. 2023, 40, 1333–1345. [Google Scholar] [CrossRef]
- Brown, M.J.M.; Walker, B.E.; Black, N.; Govaerts, R.H.A.; Ondo, I.; Turner, R.; Nic Lughadha, E. rWCVP: A Companion R Package for the World Checklist of Vascular Plants. New Phytol. 2023, 240, 1355–1365. [Google Scholar] [CrossRef]
- Lin, Q.W.; Xiao, C.; Ma, J.S. A dataset on catalogue of alien plants in China. Biodivers. Sci. 2022, 30, 110–117. [Google Scholar] [CrossRef]
- Lu, L.M.; Mao, L.F.; Yang, T.; Ye, J.F.; Liu, B.; Li, H.L.; Sun, M.; Miller, J.T.; Mathews, S.; Hu, H.H.; et al. Evolutionary History of the Angiosperm Flora of China. Nature 2018, 554, 234–238. [Google Scholar] [CrossRef] [PubMed]
- Peng, S.Z.; Ding, Y.X.; Liu, W.Z.; Li, Z. 1 Km Monthly Temperature and Precipitation Dataset for China from 1901 to 2017. Earth Syst. Sci. Data 2019, 11, 1931–1946. [Google Scholar] [CrossRef]
- Wang, G.R.; Sun, Y.; Zhou, M.; Guan, N.Q.; Wang, Y.W.; Jiang, R.H.; Liu, Z.Y.; Wu, M.J.; Xia, F.C. Effect of Thinning Intensity on Understory Herbaceous Diversity and Biomass in Mixed Coniferous and Broad-Leaved Forests of Changbai Mountain. For. Ecosyst. 2021, 8, 53. [Google Scholar] [CrossRef]
- Al-Kofahi, S.D.; Al-Kafawin, A.M.; Al-Gharaibeh, M.M. Investigating Domestic Gardens Landscape Plant Diversity, Implications for Valuable Plant Species Conservation. Environ. Dev. Sustain. 2024, 26, 21259–21279. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Lai, J.S.; Zou, Y.; Zhang, S.; Zhang, X.G.; Mao, L.F. glmm.hp: An R Package for Computing Individual Effect of Predictors in Generalized Linear Mixed Models. J. Plant Ecol. 2022, 15, 1302–1307. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2024. [Google Scholar]

Table 1.
Linear mixed-effects model analysis of the importance value of native plants in the sample plot with environmental factors, the number of alien plant species, and individuals. Adjusted R2 of the model is 0.77. All continuous variables were natural-log transformed and then scaled. The individual R² for each variable and the percentage of R² were given.
Table 1.
Linear mixed-effects model analysis of the importance value of native plants in the sample plot with environmental factors, the number of alien plant species, and individuals. Adjusted R2 of the model is 0.77. All continuous variables were natural-log transformed and then scaled. The individual R² for each variable and the percentage of R² were given.
| Term | Estimate | SE | t-value | P-value | Ind. R2 | Ind. perc |
|---|---|---|---|---|---|---|
| Intercept | 0.92 | 0.01 | 182.87 | <0.001 | / | / |
| Elevation | 0.01 | 0.01 | 2.39 | 0.021 | 0.02 | 2.41 |
| NCP | -0.02 | 0.01 | -3.23 | 0.002 | 0.09 | 12.18 |
| NIP | -0.06 | 0.01 | -12.60 | <0.001 | 0.22 | 29.23 |
| NNP | -0.09 | 0.01 | -16.08 | <0.001 | 0.43 | 56.18 |
NCP represents the number of casual plants. NIP represents the number of invasive plants. NNP represents the number of naturalized plants.
Table 2.
Linear mixed-effects model analysis of the richness of native plants in the sample plot with environmental factors, the number of alien plant species, and individuals. The adjusted R2 of the model is 0.52. All continuous variables were natural-log transformed and then scaled. The individual R² for each variable and the percentage of R² were given.
Table 2.
Linear mixed-effects model analysis of the richness of native plants in the sample plot with environmental factors, the number of alien plant species, and individuals. The adjusted R2 of the model is 0.52. All continuous variables were natural-log transformed and then scaled. The individual R² for each variable and the percentage of R² were given.
| Term | Estimate | SE | t-value | P-value | Ind. R2 | Ind. perc |
|---|---|---|---|---|---|---|
| Intercept | 1.79 | 0.04 | 45.36 | <0.001 | / | / |
| Elevation | 0.26 | 0.06 | 4.27 | <0.001 | 0.16 | 72.29 |
| MAT | 0.12 | 0.06 | 2.02 | 0.048 | 0.03 | 14.23 |
| NNS | 0.07 | 0.03 | 2.40 | 0.018 | 0.03 | 13.48 |
MAT represents the mean annual temperature. NNS represents the number of naturalized species.
Table 3.
Linear mixed-effects model analysis of the Pielou index of native plants in the sample plot with environmental factors, the number of alien plant species, and individuals. The adjusted R2 of the model is 0.38. All continuous variables were natural-log transformed and then scaled.
Table 3.
Linear mixed-effects model analysis of the Pielou index of native plants in the sample plot with environmental factors, the number of alien plant species, and individuals. The adjusted R2 of the model is 0.38. All continuous variables were natural-log transformed and then scaled.
| Term | Estimate | SE | t-value | P-value |
|---|---|---|---|---|
| Intercept | 0.58 | 0.02 | 23.92 | <0.001 |
| NNP | 0.06 | 0.02 | 2.92 | 0.004 |
NNP represents the number of naturalized plants.
Table 4.
Linear mixed-effects model analysis of the Shannon index of native plants in the sample plot with environmental factors, the number of alien plant species, and individuals. Adjusted R2 of the model is 0.54. All continuous variables were natural-log transformed and then scaled. The individual R² for each variable and the percentage of R² were given.
Table 4.
Linear mixed-effects model analysis of the Shannon index of native plants in the sample plot with environmental factors, the number of alien plant species, and individuals. Adjusted R2 of the model is 0.54. All continuous variables were natural-log transformed and then scaled. The individual R² for each variable and the percentage of R² were given.
| Term | Estimate | SE | t-value | P-value | Ind. R2 | Ind. perc |
|---|---|---|---|---|---|---|
| Intercept | 1.07 | 0.05 | 19.41 | <0.001 | / | / |
| Elevation | 0.18 | 0.06 | 3.30 | 0.002 | 0.10 | 63.37 |
| NNP | 0.14 | 0.04 | 3.44 | <0.001 | 0.06 | 36.63 |
NNP represents the number of naturalized plants.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



