Submitted:
31 March 2025
Posted:
01 April 2025
Read the latest preprint version here
Abstract
Background: Quinoline-derived metabolites exhibit notable chemical complexity. What causes minor structural alterations to induce significant changes in disease outcomes? Historically eclipsed by more straightforward scaffolds, these chemicals provide a dynamic nexus in tryptophan metabolism, connecting immunomodulation, excitotoxicity, and cancer. However, they often fail at the blood–brain barrier, and we currently lack understanding of how specific alterations influence bioavailability and off-target effects. Thus, contemporary research highlights halogenation, esterification, and computational modeling to enhance structure–activity relationships. Summary: This narrative review emphasizes the integration of rational drug design, multi-target ligands, and prodrug methods in enhancing quinoline scaffolds. We investigate the potential of each molecule, enhance the potential of each scaffold, and cultivate the potential of each derivative. Translating these laboratory findings into clinical practice, however, remains a formidable challenge. Conclusion: Through the synthesis of findings regarding NMDA receptor antagonism, improved oral bioavailability, and reduced metabolic instability, we demonstrate how single-site changes might modulate excitotoxicity and immunological signaling. Advancing quinoline-based medicines will yield significant advancements in neurology, psychiatry, and oncology. This enlarged framework fosters collaborative discovery, engages various audiences, and advances the field towards next-generation disease-modifying therapies. Robust preclinical validation, patient classification, and comprehensive toxicity evaluations are crucial stages for achieving these extensive endeavors and fostering future therapeutic discoveries globally.
Keywords:
kynurenic acid (KYNA)
; quinoline
; tryptophan (Trp)
; neuroprotection
; neuroinflammation
; excitotoxicity
; blood-brain barrier (BBB)
; pharmacokinetics
; structure-activity relationships (SARs)
; drug design
1. Introduction
Tryptophan (Trp) undergoes a fascinating series of biochemical transformations through kynurenine (KYN) metabolism that leads to the production of various neuroactive and immunomodulatory compounds [1]. At the heart of this metabolic arm lies a cascade of enzymes that generates an array of quinoline-based molecules, each with distinct and sometimes opposing effects on cellular and molecular processes [2]. Among the most scrutinized products are kynurenic acid (KYNA) and quinolinic acid (QUIN), which play critical roles in modulating neurotransmission, safeguarding neurons, and orchestrating immune responses [3]. Under normal circumstances, these molecules contribute to the finely tuned balance between protection and subtle regulation of neural function [4]. However, an overabundance or misregulation of these compounds can fuel diverse pathologies, spanning neurodegenerative conditions, psychiatric disorders, and even certain cancers [5]. It is remarkable that these metabolites, found in species across the evolutionary spectrum, have retained their functional importance over millions of years [6]. Their enduring presence suggests a fundamental role in maintaining physiological homeostasis, yet they can also become potent drivers of disease when their levels shift [7]. This duality underscores the importance of understanding both the biochemical nuances and the broader biological contexts in which these quinoline-based substances operate [3,8].
Figure 1.
The structure of quinoline. The quinoline scaffold, featuring a fused benzene and pyridine ring, forms the structural backbone of many biologically active compounds.
Figure 1.
The structure of quinoline. The quinoline scaffold, featuring a fused benzene and pyridine ring, forms the structural backbone of many biologically active compounds.
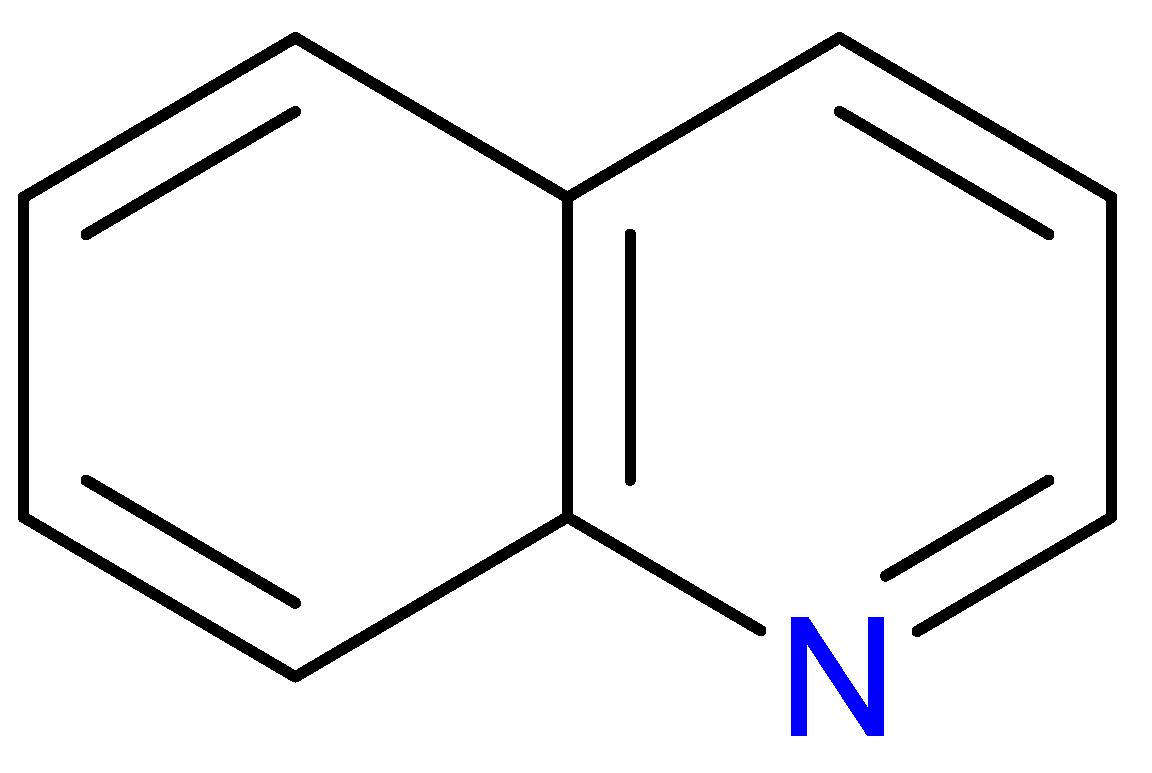
The quinoline skeleton lies at the heart of a wide variety of biologically active molecules, encompassing endogenous metabolites such as KYNA and QUIN, as well as numerous synthetic derivatives found in pharmaceuticals and research tools [9]. Defined by a fused ring structure that merges a benzene ring with a pyridine ring, quinoline offers countless opportunities for chemical substitution [10]. Small modifications, like adding carboxyl or hydroxyl groups, can dramatically alter a molecule’s behavior, influencing everything from its solubility and stability to its affinity for specific protein targets [11]. Halogenation, for instance, may boost lipophilicity and enhance central nervous system penetration, while the presence or absence of an acidic group can shape receptor-binding preferences [12]. In many cases, these structural nuances determine whether a particular quinoline derivative acts as a neuroprotective agent or exacerbates excitotoxic processes through receptors like N-methyl-D-aspartate (NMDA) or G protein-coupled receptor 35 (GPR35) [13]. Endogenous metabolites derived from the Trp degradation route typically exhibit distinct features that help them coordinate intricate cellular signaling, whereas synthetic derivatives, exemplified by quinolone antibiotics, have been optimized for pathogen clearance and bioavailability [14]. Despite their diverse origins, all members of this chemical family share the versatile quinoline core, highlighting the importance of structural considerations in developing targeted interventions for various clinical needs [15].
A hallmark of these Trp-derived quinoline compounds is their profound influence on the delicate balance between neuroprotection and neurotoxicity [16]. Their functions are finely tuned by their relative concentrations, contextual factors, and micro-environment, shaping their cumulative effects [17]. Beyond their influence in the central nervous system, these metabolites also bridge metabolic and immune pathways, as illustrated by enzymes like indoleamine 2,3-dioxygenases (IDOs) and tryptophan 2,3-dioxygenase (TDO) [18]. When activity in this biochemical system goes awry, it can fuel inflammation and shape disease trajectories in conditions as diverse as autoimmunity and cancer [19]. Emerging evidence further suggests crosstalk with the aryl hydrocarbon receptor, hinting at broader physiological roles that extend beyond neuronal circuits [20]. Such multifaceted involvement underscores the need to appreciate how these molecules orchestrate, or disrupt, redox equilibrium, synaptic communication, and immune surveillance in tandem [21]. By simultaneously affecting protective and damaging processes, they embody a paradox: slight imbalances may tip the scale toward pathology [22,23]. Investigating this interplay is vital for deciphering the biology behind complex disorders, as well as for pinpointing novel therapeutic entry points grounded in the chemistry of quinoline-based metabolites.
Therapeutic strategies centered on Trp metabolism have garnered considerable attention in recent years, buoyed by encouraging preclinical results and a surge in clinical trials [24,25]. For instance, inhibitors of IDOs showed early promise in oncology by dampening immunosuppressive pathways and restoring anti-tumor responses [26]. However, some late-stage trials yielded disappointingly modest outcomes, illustrating the complexity of translating metabolic interventions into measurable clinical benefits [27]. Another avenue involves exploiting analogs of KYNA, such as 7-CKA, which demonstrates potential for alleviating schizophrenia-related symptoms by modulating glutamate receptors [28]. While these developments highlight the promise of quinoline-based therapeutics, numerous challenges remain [29]. Achieving adequate penetration across the blood-brain barrier (BBB) can be particularly daunting, as can mitigating off-target effects that may compromise patient safety [30]. Beyond endogenously derived agents, certain quinoline derivatives traditionally used for other indications such as quinine and related molecule, are now being explored for their neuroprotective or anti-inflammatory properties [31]. Repurposing these compounds offers the opportunity to fast-track clinical testing, taking advantage of existing safety and pharmacokinetic data [32]. Yet, optimizing dosage, delivery methods, and specificity remains a priority [33]. Moving forward, interdisciplinary collaborations will likely prove vital, melding synthetic chemistry, pharmacology, and clinical expertise to refine these agents and harness their untapped therapeutic potential [34].
Despite the steady growth of research into quinoline-containing molecules, there are still pressing questions that need answers [35]. One pivotal gap lies in our limited understanding of how specific structural modifications translate into particular biological effects, often termed structure-activity relationships [36]. Such insights are crucial for guiding rational drug design and for predicting how a new derivative might behave in living systems. Another hurdle is the fragmented nature of pharmacokinetic and safety data, which often emerge from small, isolated studies rather than comprehensive analyses [37]. This piecemeal approach complicates efforts to compare efficacy and toxicity across different quinoline scaffolds [37]. Furthermore, there is a noticeable lack of head-to-head evaluations between endogenous Trp-derived metabolites and synthetic or exogenous analogs, preventing a clear assessment of relative benefits and risks. Building on these concerns, the objectives of this review are threefold. First, to systematically evaluate the wealth of endogenous and synthetic quinoline-based compounds for their therapeutic potential in various disease contexts. Second, to synthesize current knowledge about structure-activity relationships with the explicit goal of identifying a candidate inspired by KYNA that might serve as a leading drug template. Lastly, we aim to propose innovative strategies to address translational challenges, including targeted prodrug design and approaches that can engage multiple biological targets simultaneously.
2. Unveiling Endogenous Quinolines in the Kynurenine (KYN) Pathway: A Gateway to Kynurenic Acid (KYNA)–Driven Neuroprotection and Rational Drug Design
The metabolism of Trp produces a striking array of bioactive compounds [38]. KYNA, QUIN, nicotinic acid (NA), picolinic acid, xanthurenic acid (XA), and quinaldic acid each exert profound effects on human physiology [39]. Current research suggests these metabolites act as fragile regulators, holding the potential to tip the balance between neuroprotection and neurotoxicity [40]. KYNA, often described as a safeguard against excitotoxicity, dampens glutamatergic signaling [41]. In contrast, QUIN can escalate excitatory activity to dangerous extremes, contributing to neuronal injury under pathological conditions [42]. NA supports redox homeostasis, while picolinic acid and XA coordinate metal ions, subtly influencing cellular stability [43]. Quinaldic acid remains a shadowed figure in this biochemical network, its roles only beginning to emerge. These metabolites do not function in isolation. They respond to the surrounding microenvironment and reshape it in ways that may ultimately define the trajectory of health and disease (Table 1).
2.1. Structural & Functional Synergy
The quinoline core appears in a range of Trp-derived metabolites, revealing an intriguing contrast in their biological effects [2]. KYNA and QUIN emerge from the same metabolic pathway, yet they are traditionally considered to operate on opposite ends of the spectrum [68]. One protects neurons by dampening excessive excitatory signaling; the other drives excitotoxic damage often linked to inflammation [89]. This shared structural backbone, paired with such starkly different outcomes, suggests a delicate evolutionary balance shaped by minor molecular adjustments [2]. Consider how slight modifications to substituent groups on the quinoline ring can shift a molecule’s interaction with receptors, ion channels, or transporters [90]. Even fluctuations in pH or enzyme activity are enough to sway the system toward protection or harm [91]. There’s more: metal ions and cofactors add another layer of control, either amplifying or suppressing activity [92]. The challenge now lies in identifying how to steer this balance [93]. Could targeted changes to the quinoline scaffold enhance KYNA’s protective roles or curb the destructive potential of QUIN? [94] Understanding these subtle structural cues opens the door to designing molecules with more favorable effects on neural health.
Small structural modifications within quinoline-based metabolites can significantly influence their biological activity and stability [95]. Altering the position of a hydroxyl group, as observed in XA, affects hydrogen bonding capacity and lipophilicity, which may shift receptor affinity and metabolic processing [96]. These effects are particularly relevant when examining interactions with glutamate receptors such as NMDA, where even minor variations can determine whether a metabolite contributes to neuroprotection or excitotoxicity [97]. Similar patterns emerge with GPR35, a receptor that responds differently based on the specific substituents attached to the quinoline scaffold [98]. Variations in these groups regulate receptor engagement and downstream signaling pathways, shaping distinct physiological outcomes [91]. Alongside receptor interactions, metabolic stability is also sensitive to changes in ionization states and enzyme susceptibility, both of which are closely tied to the positioning of carboxyl, hydroxyl, or amino groups [99]. Mapping these structural features across endogenous metabolites provides insights into how they influence pathological processes [100,101]. Examining the relationship between substitution patterns, receptor selectivity, and metabolic half-life may contribute to the design of compounds that adjust the activity of quinoline derivatives in targeted therapeutic contexts.
2.2. Key Gaps in Translation
Despite their diverse biological activities, translating endogenous quinoline-based compounds into therapeutic interventions faces significant hurdles [102]. A chief concern is poor bioavailability, particularly evident in KYNA [103]. Its polar carboxylate group hinders passive diffusion across biological membranes and limits the ability to reach target sites in sufficient concentration [104]. On the other hand, picolinic acid, while exhibiting promising immunomodulatory and metal-chelating activities, undergoes rapid metabolism, leading to short half-life and unstable plasma levels [105]. These kinetic and physicochemical drawbacks are further complicated by the complexity of enzymatic pathways involved, where small fluctuations in metabolic rates can greatly alter the overall balance of protective versus pathogenic metabolites [106]. Such pharmacokinetic challenges underscore the difficulty of achieving consistent dosing in clinical settings [107]. As highlighted in recent efforts to refine KYN system monitoring, relying solely on static concentrations or simplistic ratios may be insufficient. A more nuanced, integrative biomarker framework is needed to capture the dynamic regulation of tryptophan metabolism and guide precision medicine applications in neuropsychiatric and neurodegenerative disorders [108]. Moreover, existing animal models do not always recapitulate human physiology, complicating the interpretation of efficacy and toxicity data [55]. Without efficient strategies to enhance bioavailability—such as prodrug approaches, targeted delivery systems, or structural modifications to increase lipophilicity—these compounds may struggle to meet clinical requirements [109]. As research deepens our understanding of how structural and functional attributes influence pharmacokinetics, efforts to overcome these limitations can be directed more precisely, potentially unlocking the therapeutic promise that many of these metabolites have long suggested.
Translating endogenous quinoline compounds into real-world treatments often involves navigating paradoxical biological roles, as exemplified by NA [110,111]. Widely recognized for its capacity to modulate lipid metabolism and improve cardiovascular profiles, NA also harbors less appreciated functions in inflammatory pathways [112]. This duality creates both opportunity and challenge: on one hand, its impact on lipid-lowering has made it a mainstay in certain therapeutic regimens; on the other hand, emerging studies suggest that NA can engage molecular cascades involved in neuroinflammation, potentially influencing conditions like neurodegenerative diseases [113,114]. Such conflicting actions underscore a larger theme in quinoline-related research, where a single molecule may oscillate between protective and pathogenic effects depending on context and concentration [115]. The incomplete mapping of these dual functionalities hampers our ability to refine dosing strategies or identify the specific patient populations who might benefit the most [91]. Additionally, the diverse array of downstream metabolites and receptors further complicates the picture, making it difficult to anticipate long-term outcomes [116]. As a result, bridging the gap from bench to bedside requires a thorough understanding of how NA’s manifold roles intersect. Only by disentangling these intricate mechanisms can researchers devise targeted interventions that harness the therapeutic benefits of NA while minimizing unanticipated consequences.
A major obstacle in harnessing the therapeutic potential of endogenous quinoline compounds lies in their limited tissue penetration and rapid clearance [49]. KYNA, for example, is known to possess neuroprotective properties but struggles to traverse biological barriers effectively, largely due to its polar carboxyl group [103]. One promising strategy is to employ prodrug approaches such as 4-chlorokynurenine and SZR-104, whereby this polar moiety is temporarily masked through esterification or similar chemical modifications [49,117]. By doing so, researchers can enhance lipophilicity, potentially improving permeability across the gut or BBB. Once inside target tissues, endogenous enzymes would cleave the ester bond, liberating active KYNA precisely where it is needed [118]. Another avenue involves synthesizing hybrid molecules, combining the core quinoline structure with lipid-soluble components [119]. This approach aims not only to improve membrane permeability but also to confer additional functional advantages, such as greater stability or targeted delivery [120]. For instance, merging KYNA with a fatty acid chain could optimize drug distribution in neuronal membranes, where excitotoxic processes frequently unfold [62]. While these strategies are conceptually appealing, they require systematic evaluation to ensure that altered pharmacokinetics do not come at the cost of diminished efficacy or unexpected side effects. Nevertheless, such chemical innovations hold great promise for translating endogenous metabolites into clinically viable treatments.
One approach to bridging the gap between preclinical promise and clinical success lies in developing dual-targeting compounds that can simultaneously influence key enzymes in Trp metabolism and modulate neuroinflammatory pathways [121]. By tackling both processes at once, these agents have the potential to dampen the production of harmful quinoline-based metabolites, such as QUIN, while also curbing inflammatory cascades that can exacerbate neuronal damage [122]. For instance, selective inhibition of enzymes like IDOs or TDO could be paired with suppression of pro-inflammatory cytokines, ultimately reducing excitotoxic insults in conditions such as neurodegeneration or chronic inflammation [123,124,125]. n diseases like multiple sclerosis, recent work emphasizes the critical importance of accurately monitoring redox dynamics as both a diagnostic tool and a therapeutic target, revealing that oxidative imbalance directly influences disease progression and treatment responsiveness [126]. The rationale is to address multiple disease mechanisms in parallel, thereby enhancing therapeutic efficacy and possibly reducing the likelihood of resistance or relapse [127]. However, designing these dual-acting agents demands a nuanced understanding of each target’s regulatory networks, as altering enzyme activity may unintentionally shift the balance toward alternative metabolic routes or trigger compensatory immune responses [128]. Moreover, the BBB remains a formidable challenge, necessitating delivery strategies that ensure sufficient drug concentrations within the central nervous system [129]. Despite these hurdles, dual-targeting compounds offer a forward-thinking strategy, uniting key biochemical nodes into a single intervention that could revolutionize how we harness endogenous quinoline metabolites for clinical benefit.
3. Expanding the Quinoline Landscape: Derivatives Beyond the Kynurenine (KYN) Metabolic Pathway
Emerging evidence supports the repurposing of established pharmacophores with known activities—such as quinine’s antimalarial and 8-hydroxyquinoline’s metal-chelating properties—to modulate the kynurenine pathway through mechanisms like IDO inhibition and redox modulation [130]. However, these compounds may present limitations, including off-target bacterial DNA gyrase inhibition (e.g., nalidixic acid) or toxicity concerns (cinchoninic acid derivatives) [131,132]. Structural homology between isonicotinic acid (INA)’s pyridine ring and KYNA’s quinoline core highlights opportunities for scaffold hybridization and the exploration of underexamined agents, like dipicolinic acid (DPA), in neurodegeneration. Further, systematic SAR studies on broad-spectrum drugs, exemplified by quinidine, could isolate neuroactive fragments for targeted therapeutic applications and ultimately safer use [133].

Table 2.
Structural and pharmacological overview of quinoline, quinolone derivatives, and pyridinecarboxylic acids. Key compounds within these chemical classes are categorized alongside their main therapeutic actions, biological targets, and significant applications. This table traces the progression from classic antimalarials to contemporary antibiotics, emphasizing how structural changes shape efficacy, safety profiles, and clinical relevance.
Table 2.
Structural and pharmacological overview of quinoline, quinolone derivatives, and pyridinecarboxylic acids. Key compounds within these chemical classes are categorized alongside their main therapeutic actions, biological targets, and significant applications. This table traces the progression from classic antimalarials to contemporary antibiotics, emphasizing how structural changes shape efficacy, safety profiles, and clinical relevance.
| Compounds | Main characteristics | Ref. |
|---|---|---|
| Quinoline and quinolone derivatives | ||
 Quinine 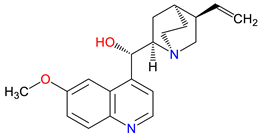 Quinidine |
Quinine: Antimalarial agent – Inhibits Plasmodium pp. by interfering with heme detoxification Historically used to reduce fever and pain Muscle relaxant – Helps alleviate nocturnal leg cramps by modulating ion channels Quinidine: Class I antiarrhythmic – Blocks sodium channels, stabilizing cardiac rhythm Chiral isomer of quinine – Shares structural similarities but has distinct pharmacological effects Proarrhythmic risk – Can prolong QT interval 1, requiring careful clinical use |
[134,135,136] [137,138,139] [140,141] [142,143,144] [145,146,147] [148,149,150] |
 Cinchoninic Acid |
Quinoline derivative – Structurally related to quinine and other cinchona alkaloids Metal chelation – Binds with metal ions, potentially influencing enzymatic activity Pharmacological potential – Investigated for antimicrobial and neuroactive properties |
[141,151,152] [153,154] [155,156,157] |
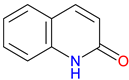 2-Quinolinone 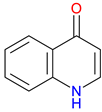 4-Quinolinone |
Core scaffold for quinolone antibiotics – Forms the backbone of fluoroquinolones, targeting bacterial DNA gyrase and topoisomerase IV Broad-spectrum antimicrobial Activity – Effective against Gram-positive and Gram-negative bacteria Synthetic Versatility – Modifiable structure allows for improved potency, pharmacokinetics, and resistance mitigation. |
[158,159,160] [161,162,163] [162,164,165] |
 Nalidixic Acid |
First-generation quinolone antibiotic – Inhibits DNA gyrase, primarily effective against Gram-negative bacteria Limited spectrum & rapid resistance – Narrow activity and high bacterial resistance limit its clinical use Urinary tract infection treatment – Historically used for urinary tract infections, though largely replaced by newer fluoroquinolones |
[166,167,168] [169,170] [171,172,173] |
 Oxolinic Acid |
Early quinolone antibiotic – Inhibits DNA gyrase, effective against Gram-negative bacteria Used in veterinary medicine – Primarily employed for treating bacterial infections in animals Limited clinical use – Replaced by newer fluoroquinolones due to resistance and pharmacokinetic limitations |
[174,175,176] [177,178,179] [180] |
| Pyridinecarboxylic Acids | ||
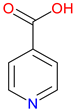 Isonicotinic Acid (INA) |
Pyridine carboxylic acid derivative – Structurally related to NA (vitamin B3). Key precursor for isoniazid – Used in the synthesis of isoniazid, a frontline anti-tuberculosis drug. Pharmacological Potential – Investigated for antimicrobial and metabolic regulatory properties |
[181,182] [183,184] [183,185,186] |
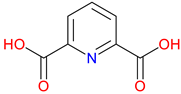 Dipicolinic Acid (DPA) |
Metal ion chelator – Strongly binds calcium and other metal ions, playing a role in metal homeostasis Bacterial spore component – Essential for bacterial endospore resistance and heat stability Potential neuroprotective role – Explored for its effects on metal-related oxidative stress in neurodegeneration |
[187,188] [189] [190,191,192] |
| Hydroxy-Substituted Derivatives | ||
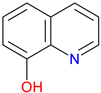 8-Hydroxyquinoline |
Metal clator – Strongly binds iron, copper, and zinc, influencing redox balance and enzymatic activity Antimicrobial and antifungal agent – Exhibits broad-spectrum activity against bacteria and fungi Potential neuroprotective role – Investigated for treating neurodegenerative diseases by regulating metal toxicity |
[193,194,195] [196,197,198] [199,200,201] |
1 QT interval, Time between onset of Q wave and end of T wave on electrocardiogram (ECG), reflecting ventricular depolarization and repolarization. NA, nicotinic acid.
3.1. Repurposing Potential
XA, although not a central intermediate in the KYN pathway, illustrates how precise structural changes can influence receptor interactions and metabolic stability [202]. Introducing a hydroxyl group at specific sites on the quinoline ring modifies hydrogen bonding capacity, which may shift affinity toward NMDA receptors or GPR35 [98]. Such alterations affect not only receptor binding but also downstream processes, including neurotransmission and immune signaling [203]. Adjustments in stereochemistry or the addition of other substituents can further redirect molecular preferences, raising the possibility of selectively targeting distinct physiological pathways [204]. Yet, enhancing receptor specificity often comes with trade-offs [205]. For instance, while certain hydroxylations strengthen binding, they may also increase vulnerability to enzymatic degradation [206]. Strategies such as incorporating protective groups or additional ring modifications can help extend plasma half-life and improve pharmacokinetic profiles [207]. Beyond structural design, patient stratification based on genetic variations in enzymes like IDOs and KMO offers a route to refine therapeutic applications [208]. Identifying metabolic subgroups may improve treatment outcomes and reduce adverse effects, particularly in neurodegenerative and neuroinflammatory conditions, where individual responses vary widely [209].
Quinoline derivatives beyond the KYN pathway present structural opportunities but are often limited by safety concerns that complicate therapeutic development [102]. Nalidixic acid, widely recognized for its antibacterial activity through inhibition of DNA gyrase, raises concerns when considered for neuroprotective or immunomodulatory purposes [210]. Interference with DNA metabolism, while effective against bacterial targets, may introduce risks to human cells, particularly in long-term or systemic applications [211]. Similar challenges appear with cinchoninic acid derivatives, which share structural features with KYNs yet exhibit toxicity that restricts clinical use [212]. In several cases, these effects appear linked to the formation of reactive metabolites or unintended interactions with metabolic enzymes [213]. Even minor modifications to the quinoline scaffold can redirect metabolic pathways, sometimes amplifying toxic outcomes [214]. Addressing these complexities requires a detailed understanding of structure-toxicity relationships and strategies that preserve pharmacological activity while minimizing off-target effects [215]. Approaches such as selective functional group modifications and alternative synthetic routes are under investigation [216]. Advancing these efforts depends on collaboration across medicinal chemistry, toxicology, and clinical research to reduce liabilities and support the development of safer quinoline-based compounds [217].
3.2. Bridging to Kynurenic acid (KYNA)-Based Targets
Structural similarities between INA and KYNA have prompted interest in hybrid molecules that combine elements of both frameworks [218]. Each features a nitrogen-containing heterocycle capable of influencing receptor interactions and electron distribution [219]. By integrating portions of the quinoline core from KYNA with the pyridine ring of INA, researchers aim to enhance selectivity at targets such as NMDA receptors and GPR35. This approach allows for adjustments in ring orientation and substituent placement, offering opportunities to fine-tune receptor engagement while addressing challenges related to metabolic stability [220]. The isonicotinic scaffold, in particular, provides flexibility for introducing functional groups that may improve solubility or permeability [221]. While these modifications hold promise for minimizing off-target effects, the relationship between structural changes and pharmacological outcomes remains complex [222]. Balancing synthetic accessibility with therapeutic potential requires careful evaluation of how hybrid designs alter binding properties and metabolic pathways. Drawing from existing knowledge of INA derivatives, ongoing studies will assess whether these structural combinations can deliver improved performance while limiting adverse effects, especially in applications involving neuroprotective and immunomodulatory pathways.
DPA, a pyridine-2,6-dicarboxylic acid, remains relatively underexplored despite its capacity to bind metal ions commonly implicated in neurodegenerative diseases [223]. Through interactions with iron, zinc, and copper, DPA may contribute to regulating metal balance, a process closely linked to oxidative stress and protein aggregation [224]. Preliminary findings suggest potential roles in modulating inflammatory responses and excitatory signaling, raising questions about its relevance to therapeutic strategies aimed at neuroprotection [225]. Structurally, the resemblance between DPA's pyridine core and the quinoline ring of KYNA presents opportunities to combine metal-chelation with receptor targeting [226]. This dual functionality may prove valuable, particularly in disorders where oxidative damage and glutamatergic dysregulation intersect [227]. Parkinson’s disease and Alzheimer’s disease are two examples where such mechanisms converge, though much remains unknown regarding DPA's pharmacological behavior [228]. Determining its metabolic stability, toxicity profile, and specific enzyme interactions will be essential for guiding further development [229]. By applying knowledge from KYNA analog research, it may be possible to design compounds that extend the benefits of metal-binding while refining receptor selectivity, supporting efforts to address complex neurodegenerative conditions [230].
3.3. Innovative Directions
Identifying neuroactive components within broad-spectrum quinoline derivatives offers a pathway to develop treatments with fewer adverse effects [12]. Quinidine, primarily recognized for its role in modulating cardiac ion channels, provides a useful example [217]. Through structure-activity relationships analysis, specific molecular features can be mapped to isolate those contributing to neurological benefits, such as limiting excitotoxicity, while reducing the risk of arrhythmias [231,232]. Computational docking and high-throughput screening techniques assist in pinpointing functional groups that influence ion channels within the central nervous system, rather than the heart [233]. These insights guide the design of analogues tailored to interact with neuronal targets, including channels and receptors implicated in neurodegenerative disorders [234]. Achieving this balance between therapeutic effect and safety often relies on incremental structural adjustments—altering side chains, introducing new substituents, or modifying ring systems can significantly affect binding behavior and metabolic stability [235]. While quinidine’s properties are well characterized, questions remain about how best to adapt its framework for neurological applications [236]. Ongoing work with related compounds may help define strategies to refine multi-purpose quinoline scaffolds and explore their potential in treating conditions such as epilepsy, chronic pain, and neurodegeneration.
4. Exogenous Horizons: Synthetic Quinoline Scaffolds in Rational Drug Design
Driven by rational design approaches, synthetic modifications to quinoline-based scaffolds have enhanced both target engagement and pharmacokinetic profiles [237]. For example, chlorination in 7-CKA and sulfonamide substitution in Gavestinel improved NMDA receptor blockade and bioavailability, respectively [238,239,240,241]. Despite promising preclinical data, clinical outcomes have not always matched expectations—Gavestinel, for instance, failed in stroke trials [242]. Encouragingly, newer therapies like laquinimod show immunomodulatory benefits in multiple sclerosis [243]. These developments highlight the challenge of balancing selectivity and polypharmacology, exemplified by L-701,324’s potent NMDA antagonism and glutamatergic side effects [244]. Future strategies emphasize BBB-penetrant analogs and multi-target ligands exemplified by tasquinimod, including HDAC (histone deacetylase) 4 inhibition and aryl hydrocarbon receptor modulation.
Table 3.
Synthetic exogenous quinoline-based compounds: structures and pharmacological profiles. Synthetic and exogenous quinoline derivatives are outlined here, with emphasis on their primary mechanisms of action, therapeutic potential, and development hurdles. Each compound is linked to its intended molecular target—from NMDA receptor antagonism to immunomodulatory and anticancer functions—and is accompanied by references for in-depth exploration.
Table 3.
Synthetic exogenous quinoline-based compounds: structures and pharmacological profiles. Synthetic and exogenous quinoline derivatives are outlined here, with emphasis on their primary mechanisms of action, therapeutic potential, and development hurdles. Each compound is linked to its intended molecular target—from NMDA receptor antagonism to immunomodulatory and anticancer functions—and is accompanied by references for in-depth exploration.
| Compounds | Main characteristics | Ref. | |
|---|---|---|---|
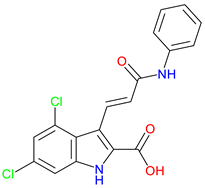
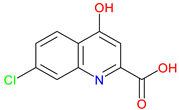 7-Chlorokynurenic Acid (7-CKA) |
NMDA receptor antagonist – Blocks the glycine-binding site, reducing excitotoxicity. Preclinical behavioral actions – Elicits antidepressant-like effects, blocks NMDA-induced convulsions, and attenuates ischemia-induced learning deficits Clinical trials – 4-Clorokynurenin, a prodrug of 7-CKA show no significant antidepressant effects in treatment-resistant depression and launched a clinical trial for neuropathic pain |
[245,246] [247,248,249] [250] |
|
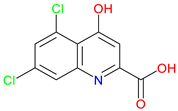 5,7-Dichlorokynurenic Acid |
NMDA receptor antagonist – Blocks the glycine site to reduce excitotoxicity Enhanced potency – More effective than KYNA at inhibiting NMDA receptor activity Preclinical behavioral actions – Show anxiolytic effects and enhance short-term memory and recognition |
[251,252,253] [238,251,253,254] [255,256,257] |
|
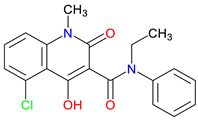
 L-701,324 |
NMDA receptor antagonist – Blocks the glycine site, reducing excitotoxicity Preclinical behavioral evidence – Demonstrates antidepressant-like activity, Reduce anxiety-like behavior Potential clinical application –Epilepsy, schizophrenia, and chronic pain |
[258,259] [257,260] [261,262,263] |
|
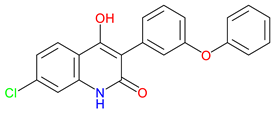
 Gavestinel |
Glycine site NMDA receptor antagonist – Blocks NMDA receptor activity to reduce excitotoxicity Preclinical behavioral findings – potential in reducing ischemic damage and modulating certain NMDA receptor-mediated schizophrenia-like behaviors Stroke neuroprotection Candidate – Investigated for acute stroke, primary intracerebral hemorrhage, acute ischemic stroke, but failed in clinical trials. |
[264,265] [264,266] [264,267,268] |
|
 Laquinimod |
Immunomodulatory agent – Reduces pro-inflammatory cytokines and modulates immune cell activity Preclinical findings – Improve motor function in a Huntington's disease model, activate the AhR in the EAE Model of MS Clinical trials – Modestly reduce relapse rates and disability progression and significantly reduce brain volume change atrophy in relapsing-remitting MS, and show limited efficacy in active non-infectious intermediate, posterior, or panuveitis (NCT02720102) |
[269,270,271] [272,273] [274,275,276] |
|
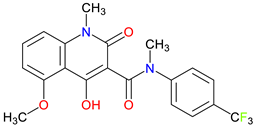 Tasquinimod |
Anti-cancer properties – Inhibits tumor angiogenesis, suppresses myeloid-derived suppressor cells, downregulate immune suppressive pathways and inflammatory cytokine signaling S100A9 modulation – Suppress inflammatory factor expression, potentially inhibit the upregulation of S100A9 in AD, and inhibit MDSC recruitment HDAC4 Modulation – potentially influence epigenetic regulation in neurodegenerative or cognitive disorders and potentially control neuronal memory, plasticity, and learning |
[277,278,279] [280,281,282] [283,284] |
AD, Alzheimer’s disease; HDAC, histone deacetylase; AhR, aryl hydrocarbon receptor; EAE, experimental autoimmune encephalomyelitis, MS: multiple sclerosis; MDSCs, myeloid-derived duppressor cells.
4.1. Rational Design Successes and Failure
Refining quinoline scaffolds through targeted synthetic modifications has proven effective in improving receptor selectivity and pharmacokinetic properties [285]. In the case of 7-CKA, introducing a chlorine atom at the 7-position strengthens antagonism at NMDA receptors, surpassing the activity of KYNA [245]. This substitution not only stabilizes the molecule's conformation at the binding site but also alters its metabolic profile, with potential implications for clearance and half-life [286]. Gavestinel presents a related example, where the incorporation of sulfonamide groups enhances solubility and may support passage across the BBB, features that are critical for central nervous system activity [287]. While such modifications can improve receptor engagement and bioavailability, they also introduce challenges [288]. Altering chemical structures to enhance efficacy may inadvertently affect safety, particularly as compounds advance toward clinical evaluation [289]. Balancing these factors requires careful attention to the relationship between functional groups and biological outcomes [290]. Through studies of compounds like 7-CKA and Gavestinel, medicinal chemistry continues to identify structural features that support NMDA receptor modulation, offering a foundation for the development of new treatments targeting neurological disorders.
The challenges of translating quinoline-derived compounds into clinical success are illustrated by the case of Gavestinel. Developed to limit excitotoxic injury following stroke through NMDA receptor antagonism, Gavestinel advanced from preclinical studies with strong supporting data [291]. Yet, in clinical trials, it failed to produce meaningful improvements in patient outcomes, raising questions about the limitations of commonly used experimental models [292]. Stroke in humans presents variability in onset, severity, and timing that is difficult to replicate in animal studies [293]. Many preclinical trials rely on designs lacking adequate randomization, blinding, or statistical power, factors known to overestimate efficacy in stroke research [294]. Differences in treatment windows also complicate translation [295]. While preclinical models often apply interventions early, real-world scenarios involve delays, with irreversible damage frequently present before therapy begins [295]. Addressing these discrepancies requires trial designs that account for the complexity of human stroke and consider combining neuroprotective strategies with reperfusion approaches [296]. Without such adjustments, promising compounds remain at risk of falling short in clinical settings, regardless of structural refinement or pharmacological rationale.
The failure of IDO inhibitors in oncology trials has raised important considerations for the development of quinoline-based therapies [297]. Epacadostat, an IDO inhibitor evaluated for melanoma, did not meet its clinical endpoints, partly due to limited patient stratification and the absence of reliable biomarkers to predict treatment response [298]. Similar issues have been observed with Gavestinel in stroke studies, where variability in disease onset and delayed intervention may have obscured potential benefits [299]. These examples point to the value of using biomarkers—such as measures of KYN pathway activity or inflammatory cytokines—to guide patient selection and improve trial outcomes [300]. Combining KYNA analogs with immune checkpoint inhibitors or metabolic regulators may also help counteract compensatory mechanisms that reduce therapeutic effectiveness [301]. Trial designs that include real-time pharmacokinetic assessments, like cerebrospinal fluid analysis to verify central nervous system exposure, could support better decision-making during clinical evaluation [302]. Failures such as epacadostat’s also reflect the limitations of animal models, which often fall short of replicating the complexity of human pathophysiology [303,304]. Integrating techniques like metabolomics and single-cell RNA sequencing into preclinical research may help identify specific patient subgroups who are more likely to benefit from targeted quinoline derivatives.
Laquinimod, a quinoline-3-carboxamide derivative, offers a contrasting example of how molecular design can align with disease context to support therapeutic development [305]. In multiple sclerosis, where treatment focuses on managing chronic inflammation leading to neurodegeneration rather than responding to acute injury, laquinimod’s ability to reduce pro-inflammatory cytokines and support neuronal preservation has led to more favorable outcomes compared to agents like Gavestinel [306]. Unlike stroke, where delays in intervention often limit the impact of neuroprotective strategies, the progressive nature of multiple sclerosis provides a broader window in which immunomodulation may alter disease progression [307]. These differences raise an important question: how can molecular targets be matched more effectively to the underlying biology of a condition? Laquinimod’s selective focus on immune pathways contrasts sharply with the NMDA receptor antagonism of Gavestinel, highlighting the value of mechanisms that reflect the complexity of the disease environment. While setbacks in clinical translation are common, cases like laquinimod suggest that carefully tailored approaches, applied to appropriate patient groups, may improve the likelihood of success and offer meaningful advances in treating neurodegenerative disorders [308,309].
4.2. Mechanistic Trade-Offs
L-701,324 has drawn attention for its strong ability to block NMDA receptors and suppress excitatory neurotransmission [310]. In conditions such as epilepsy and neuropathic pain, this level of inhibition may offer therapeutic benefits [311]. Yet, the same mechanism that provides relief can also carry significant risks [312]. Cognitive disturbances, sedation, and symptoms resembling psychosis have been linked to excessive glutamatergic suppression, raising difficult questions about how far receptor blockade should go [313]. Should drug development focus on narrowing selectivity to avoid unintended effects, or might a broader range of activity offer advantages in disorders shaped by multiple interacting pathways? While some support the latter, NMDA receptor antagonists present clear limitations [314]. Even small deviations in targeting can disrupt neural function, underscoring the importance of precise molecular engagement [315]. The case of L-701,324 highlights this tension between therapeutic potential and safety [316]. Adjusting receptor affinity without tipping the balance too far remains a central challenge [314]. As research moves forward, designing compounds that maintain effective modulation while minimizing disruption will be key to expanding the use of NMDA antagonists in clinical settings.
A growing strategy involves developing BBB-penetrant derivatives that build on the neuroprotective potential of KYNA while addressing its limited access to the central nervous system [49]. One example includes modifying KYNA with tert-butyl ester groups, which may improve its transport across the BBB before enzymatic processes release active KYNA within neural tissue [317]. Alongside these efforts, multi-target approaches are attracting interest, particularly for conditions shaped by overlapping biological pathways [318]. Tasquinimod offers a case in point, combining HDAC4 inhibition with aryl hydrocarbon receptor modulation to influence both immune responses and cellular growth [319]. Such dual-action strategies aim to address the complexity of disorders like cancer and autoimmune diseases, where inflammation, proliferation, and tissue damage converge [320]. Whether the goal is to strengthen the delivery of KYNA analogs or to design compounds that coordinate multiple mechanisms, the challenge lies in maintaining therapeutic benefits while controlling off-target effects [321]. Progress will depend on refining these scaffolds through systematic screening, predictive modeling, and careful pharmacokinetic evaluation [322]. Together with biomarker-informed clinical studies, these efforts may guide the development of quinoline-based therapies for neurological, oncological, and inflammatory conditions.
Quinoline derivatives are gaining attention for their ability to influence epigenetic mechanisms, particularly through the modulation of histone deacetylation and DNA methylation [323]. Tasquinimod, targeting HDAC4, has demonstrated potential in addressing both neurodegenerative conditions and cancer, while newer compounds are being designed to expand activity across additional HDAC and DNMT targets [324]. These effects may help restore tumor suppressor functions or regulate inflammatory pathways [325]. Genetic variation within the KYN pathway, including polymorphisms in IDOs and KMO, adds another layer of complexity by shifting the balance between protective and harmful metabolites [326]. Combining genotyping with metabolic profiling may guide treatment strategies [327]. Moving forward, refining these molecules will require careful structure-activity studies and genotype-based clinical trials.
Recent findings also underscore the potential of lipid-based nanoparticle systems to improve the delivery of quinoline derivatives across the BBB [328]. For instance, encapsulating these molecules in lipid vesicles can mitigate their rapid clearance, thereby increasing CNS bioavailability [329]. This strategy not only allows for sustained drug release but also reduces off-target toxicity—advances that could reinvigorate clinical interest in repurposing or refining existing quinoline scaffolds [330]. In parallel, polymeric nanoparticles and biomimetic vesicles such as exosome-based carriers are emerging as next-generation vehicles for transporting hydrophilic or ionizable quinoline species [331]. By tailoring particle size, surface charge, and release kinetics, researchers can customize how these agents distribute and act in vulnerable CNS regions. Nanotechnology offers promising avenues to overcome the delivery challenges of polar quinoline derivatives like KYNA. By encapsulating these molecules in exosomes or polymeric nanoparticles, it is possible to enhance their stability, improve BBB penetration, and achieve targeted release at the site of action. This advanced delivery strategy not only increases local drug concentration but also minimizes systemic exposure, paving the way for more effective neuroprotective therapies. Incorporating such advanced delivery mechanisms will be crucial for translating preclinical ‘hits’ into viable, patient-friendly therapeutics.
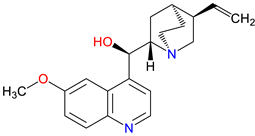
5. Next-Generation Kynurenic Acid (KYNA) Analogues: The SZR Series
Structure–activity relationship (SAR) studies yield SZR derivatives with improved pharmacological profiles [332]. SZR-72 shows enhanced neuroprotection, effective blood–brain barrier (BBB) penetration, and behavioral modulation [333,334,335]. SZR-104 displays high BBB permeability and neuroprotective effects in sepsis models [47,336]. SZRG-21 demonstrates the effect on motor activity and emotional behavior [335]. SZR-109 stands out for its strong BBB penetration, suppression of TNF-α, upregulation of TSG-6, and anti-convulsant activity [49,337,338]. However, SZR-73, despite improving mitochondrial function and reducing systemic inflammation in sepsis, does not restore microcirculation, limiting its utility [221]. SZR-105, while highly BBB-permeable and anti-inflammatory, requires further validation for long-term safety [49,337]. Prioritizing SZR-81 may be premature due to poor BBB penetration, although its antidepressant-like effects and neuroprotective potential are promising [339]. Advancing lead optimization may benefit from synergistic strategies, such as combining these analogs with IDO inhibitors to better modulate the KYN pathway and enhance therapeutic efficacy.
Table 4.
Kynurenic acid (KYNA) analogues from the SZR Series: atructural variations and pharmacological insights. The primary SZR-series analogues derived from KYNA are summarized here, highlighting their structural modifications and core pharmacological properties. The data emphasize their therapeutic relevance in excitotoxicity, neurodegenerative diseases, and related conditions, with references provided for deeper investigation.
Table 4.
Kynurenic acid (KYNA) analogues from the SZR Series: atructural variations and pharmacological insights. The primary SZR-series analogues derived from KYNA are summarized here, highlighting their structural modifications and core pharmacological properties. The data emphasize their therapeutic relevance in excitotoxicity, neurodegenerative diseases, and related conditions, with references provided for deeper investigation.
| Compounds | Main characteristics | Ref. | |
|---|---|---|---|
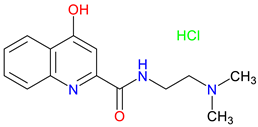 SZR-72 |
BBB penetration – effectively crosses the blood–brain barrier, enhancing its therapeutic potential in the CNS Neuroprotective effect – Offers enhanced neuroprotection, reduces brainstem c-fos, and induces therapeutic hypothermia Behavioral effects – Alters motor and exploratory behaviors, reducing vertical activity and affecting curiosity-linked emotional and motor responses |
[334,335] [333,334] [335] |
|
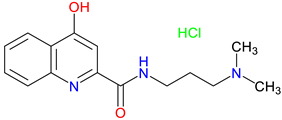 SZR-73 |
Mitochondrial function enhancement – Enhances mitochondrial respiration and ATP production, improving complex I- and II-linked OXPHOS in a rodent sepsis model Systemic inflammatory activation reduction – Reduces systemic inflammation in sepsis, lowering ET-1, IL-6, NT levels, and XOR activity Microcirculatory Effects – Improves mitochondrial function but does not restore microcirculation, unlike KYNA, which improves microvascular perfusion in sepsis |
[221] [221] [221] |
|
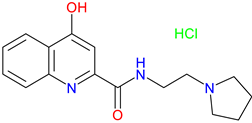 SZR-81 |
Antidepressant-like effects – Reduces immobility and increasing swimming in FST via serotonin 5-HT system involvement BBB permeability – Poor BBB penetration or metabolic alteration before reaching the CNS Neuroprotective potential– To be explored for use in stroke, neurodegenerative diseases, and inflammatory conditions |
[339] [339] [340] |
|
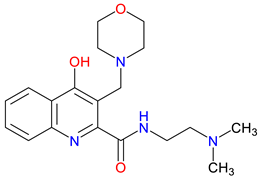 SZR-104 |
High permeability through the blood – Shows high BBB permeability, surpassing KYNA and analogues Neuroprotective effects in sepsis – Demonstrates neuroprotection in septic rodents, reducing BBB disruption and CNS mitochondrial dysfunction Influence on motor behavior – Increases horizontal exploration, indicating BBB penetration and retention of KYNA-like effects on motor-related curiosity and emotional behavior |
[49] [336] [335] |
|
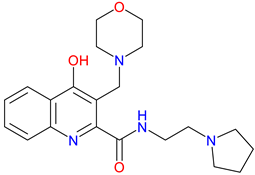 SZR-105 |
High BBB penetrability – Significantly better BBB permeability than KYNA and earlier analogues due to its dual cationic side chains Potent anti-inflammatory effects – Suppresses TNF-α production and strongly induces TSG-6 Neuroprotective activity in CNS models – Reduces cortical spreading depression propagation in migraine models |
[49] [337] [338] |
|
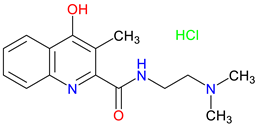
 SZR-109 |
Enhanced BBB penetration – penetrate the BBB more effectively than KYNA Neuroprotective effects –Suppresses pro-inflammatory TNF-α production and upregulates TSG-6 Anti-convulsant activity – prevents or diminishes seizure-like activity in the brain |
[49] [337] [338] |
|
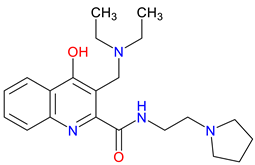
 SZRG-21 |
BBB penetration – Demonstrated efficacy in reducing cytokine levels in colitis models Impact on motor activity – Alter motor activity and exploratory behavior Emotional and behavioral modulation – Affect emotional functions such as motor-associated curiosity and emotions |
[335] [335] [335] |
ATP, adenosine triphosphate; BBB, blood-brain barrier; CNS, central nervous system; ET-1; endothelin-1; FST, forced swim test; 5-HT, 5-hydroxytryptamine; IL-6, interleukin 6; KYNA, kynurenic acid; NT, N-terminal peptide; OXPHOS, oxidative phosphorylation; TNF-α, tumor necrosis factor-alpha; TSG-6, tumor necrosis factor-stimulated gene-6; XOR, xanthine oxidoreductase.
5.1. SAR-Driven Innovations
In-depth SAR investigations reveal how subtle modifications to the KYNA core can profoundly affect receptor affinity and metabolic traits [341]. SZR-72, for instance, features a small methyl group that contributes to its enhanced neuroprotective effects, blood–brain barrier penetration, and behavioral modulation—suggesting that subtle steric modifications may play a significant role in optimizing CNS-targeted activity [335]. Meanwhile, SZR-104's C3 addition of the polar ring system demonstrates how introducing electronegative groups can enhance BBB permeability and support neuroprotection, especially in systemic inflammatory conditions [335]. By tweaking these chemical features—adding a methyl here, attaching a ring system there—researchers can systematically fine-tune key pharmacological properties like potency and half-life [342]. Such directed alterations reflect a deeper understanding of how receptor-ligand interactions depend on precise spatial and electronic configurations . In some cases, these modifications not only boost the BBB permeability but also reduce off-target binding, mitigating potential side effects [39]. Additionally, selecting the right functional group can influence polarity and solubility, traits that shape oral bioavailability [343]. Together, these iterative design strategies exemplify the power of SAR-driven innovations to optimize molecules for both efficacy and safety [344]. Beyond SZR-72 and SZR-104, further explorations could unleash even more advanced analogues that are fine-tuned to specific therapeutic goals. As we learn more, SAR mapping remains a cornerstone of rational drug discovery for KYNA derivatives.
Among the ever-growing list of KYNA analogues, SZRG-21 and SZR-109 stand out for their distinct therapeutic profiles in vivo [335,338]. Meanwhile, SZR-72, SZR-104, and SZRG-21 influence motor-associated curiosity and emotional behavior, implying broader neuromodulatory effects [335]. On the other hand, SZR-109 exhibits potent neuroprotective properties, notably suppressing TNF-α production and upregulating TSG-6 [337]. Its ability to effectively penetrate the blood–brain barrier and its anti-convulsant effects—preventing seizure-like activity—further highlight its therapeutic potential in central nervous system disorders [49,338]. The contrasting profiles of these analogues underscore how strategic SAR modifications can yield compounds tailored to specific pathophysiologies. While SZRG-21 appears suited to modulate behavioral aspects, SZR-109 is structurally optimized for neuroprotection [335,338]. These findings emphasize the flexibility of the KYNA scaffold, which allows for fine-tuned substituents to address diverse pathological targets. Further studies on their binding affinities and pharmacokinetics may unlock new therapeutic applications and refine their clinical relevance. Ultimately, SZRG-21 and SZR-109 exemplify how SAR-guided innovation can push the boundaries of KYNA-based therapeutics.
5.2. Translational Barriers
Despite ongoing refinements to KYNA analogues, multiple translational barriers must be addressed before these compounds can achieve widespread clinical utility. Prodrug strategies or nanoparticle delivery systems could potentially mitigate such clearance problems, yet these approaches introduce their own complexities in formulation and manufacturing [345,346]. On another front, SZR-105 demonstrates considerable promise but poses its own safety concern: off-target kinase inhibition [49,347]. Even minor structural tweaks can inadvertently shift binding toward other enzymes or receptors, jeopardizing patient safety [335]. Balancing pharmacological potency with a clean off-target profile is therefore a central challenge in modern drug development [348,349]. Medicinal chemists must undertake detailed SAR investigations, systematically exploring substituents and functional groups to minimize toxicity while preserving therapeutic efficacy [350]. Such iterative optimization demands close collaboration among chemists, pharmacologists, and toxicologists, each focusing on a different aspect of a molecule’s life cycle [351]. By carefully addressing problems like rapid clearance and unintended kinase inhibition, researchers can pave the way for safer, longer-lasting KYNA-based treatments that ultimately improve patient outcomes. This section may be divided by subheadings. It should provide a concise and precise description of the experimental results, their interpretation, as well as the experimental conclusions that can be drawn.
5.3. Path to Lead Optimization
Achieving an optimal lead among KYNA analogues calls for balancing efficacy, selectivity, and favorable pharmacokinetics [352]. Balanced profiles are critical in translational research, where the nuances of absorption, distribution, metabolism, and excretion (ADME) intersect with complex biological pathways [353]. High bioavailability ensures that the compound can be administered conveniently, improving patient compliance and potentially reducing dosing frequency [354]. Meanwhile, precise receptor selectivity helps mitigate adverse effects by limiting unwanted off-target binding [355]. These attributes underscore the importance of methodical SAR evaluations, wherein subtle modifications to the parent scaffold can bolster both pharmacodynamics and pharmacokinetics [356]. Complementary techniques like computational modeling and in vitro assays further expedite the refinement process, shedding light on metabolic pathways and guiding structural tweaks [357]. By identifying leads in the SZR series, researchers can streamline development cycles and reduce the risk of late-stage failures in clinical trials [358]. Ultimately, focusing on candidates that marry oral bioavailability with selective receptor engagement represents a viable strategy to usher KYNA-based therapies toward their full therapeutic potential.
Adopting combinatorial approaches that integrate multiple therapeutic strategies can heighten the efficacy of KYNA-based treatments [359]. One promising avenue involves combining SZR analogs, designed to selectively modulate GPR35 or NMDA receptors, with established IDO inhibitors that curtail the overall flux through the KYN pathway [360]. By partially blocking the production of downstream metabolites, IDO inhibitors reduce the burden of neurotoxic or immunomodulatory intermediates, thus creating a biochemical context where KYNA analogs may exert stronger protective or immunoregulatory effects [361]. This two-pronged tactic could help maintain stable levels of beneficial metabolites like KYNA, while simultaneously minimizing harmful byproducts, leading to more robust interventions in neurodegenerative or inflammatory conditions [362]. Preclinical data suggests that synergy may stem from the interplay between decreased KYN production and amplified receptor engagement by the analogs [363]. However, identifying optimal drug combinations will require meticulous dose-response studies and toxicological evaluations [364]. In particular, attention to pharmacokinetic profiles, such as how these agents interact or compete for metabolic pathways, is essential to avoid unintended consequences [365]. Ultimately, harnessing a combinatorial approach like SZR analogs plus IDO inhibitors illustrates the potential of integrated interventions targeting multiple points along the KYN pathway [366]. Such strategies may yield tangible clinical outcomes that surpass those achievable by single-agent therapies alone.
3. Discussion
The KYN metabolic system serves as a critical nexus in metabolism, intricately linking Trp catabolism to the production of neuroactive and immunomodulatory quinoline-based compounds [367]. Central to this pathway are endogenous metabolites such as KYNA and QUIN, which exert intrinsically distinct and often opposing effects both individually and in relation to each other, modulating the equilibrium between neuroprotection and excitotoxicity, immune responses, and redox balance [8,368]. Structural nuances within their quinoline scaffolds, including carboxylation, halogenation, and hydroxylation, profoundly influence their receptor specificity (e.g., NMDA, GPR35) and metabolic stability, underscoring the interplay between chemistry and physiology [116]. Despite their therapeutic promise, translational hurdles such as poor bioavailability, rapid clearance, and paradoxical biological effects impede clinical progress [369]. This review systematically evaluates endogenous and synthetic quinoline derivatives, emphasizing their structural diversity, mechanistic trade-offs, and therapeutic potential across neurodegenerative, inflammatory, and oncological contexts. By synthesizing structure-activity relationships and translational challenges, we aim to identify a lead compound inspired by KYNA, prioritizing innovations like prodrug strategies and multi-target engagement to bridge the gap between preclinical promise and clinical efficacy [370,371]. Our findings aim to catalyze rational drug design for quinoline-based therapies in complex diseases.
Building on the evidence from the preceding sections, this discussion underscores how the current knowledge integrates the chemical identity, structural nuances, and biological significance of quinoline-based metabolites to reveal critical therapeutic implications [217]. Across various endogenous and synthetic derivatives, the document emphasizes the delicate interplay between neuroprotective and neurotoxic effects, demonstrating how minor structural shifts can dramatically influence receptor binding, solubility, and metabolic stability [372]. By exploring molecules like KYNA and QUIN alongside analogues engineered for improved pharmacokinetics or selectivity, the synthesis showcases promising targets for conditions spanning neurodegenerative disorders and cancer [373]. Of particular note is the attention placed on bridging preclinical findings with translational barriers, including bioavailability challenges and safety concerns [374]. Overall, this review demonstrates that tuning the quinoline scaffold holds vast potential for advancing novel interventions, underscoring the need for continued interdisciplinary efforts to harness these insights for tangible patient benefits.
Quinoline-metal complexes, particularly 8-hydroxyquinoline chelators, play a crucial role in mitigating oxidative stress in Alzheimer’s disease [125,375,376]. By binding excess metal ions such as copper and zinc, these compounds prevent metal-catalyzed production of reactive oxygen species, thereby reducing neuroinflammation and amyloid aggregation [377]. Additionally, their ability to regulate redox balance and modulate autophagy pathways highlights their potential as multi-target neuroprotective agents [378]. Notably, recent breakthroughs in migraine therapeutics have underscored how targeting neuropeptides such as calcitonin gene-related peptide, pituitary adenylate cyclase-activating polypeptide, and vasoactive intestinal polypeptide can reshape our understanding of neuroinflammation and pain transmission, further emphasizing the need to develop quinoline-based agents with pleiotropic mechanisms [379]. Future research should focus on optimizing their stability and brain permeability to enhance clinical applicability. Structural modifications of KYNA derivatives extend beyond receptor binding, influencing critical downstream pathways such as mTOR and autophagy [380]. Minor alterations in functional groups can modulate phosphorylation events within the mTOR axis, potentially affecting neuronal survival and inflammatory responses [381]. Additionally, shifts in KYNA’s metabolic profile may alter autophagy-related signaling, impacting cellular homeostasis under oxidative stress conditions [382]. These mechanistic insights highlight the need for targeted modifications that optimize KYNA analogs for neuroprotective or immunomodulatory applications. Such synergy underlines the manuscript’s contribution to research.
Although the KYN pathway is often discussed in terms of glutamate receptor modulation, its complex interaction with serotonin metabolism and mitochondrial function deserves greater attention [23,383]. In particular, QUIN—a downstream metabolite—can significantly impact neuronal energy homeostasis by depleting NAD+ through the de novo salvage pathway [384]. When QUIN accumulates, it not only drives excitotoxic damage but also impairs mitochondrial electron transport, exacerbating oxidative stress and triggering deficits in ATP production [385]. This metabolic interplay may be especially critical in neurodegenerative disorders, where chronic inflammation and mitochondrial dysfunction intersect to accelerate disease progression [386]. Meanwhile, upstream metabolites within the KYN pathway compete for Trp, potentially altering serotonin synthesis and influencing mood regulation [383]. Such biochemical crosstalk helps explain why patients with conditions like Parkinson’s disease or Alzheimer’s often present with both cognitive and affective symptoms [387,388]. By illuminating how QUIN’s NAD+-depleting properties converge with disrupted serotonin metabolism, researchers can identify new multi-target strategies—such as modulating key enzymes in each branch of Trp degradation. Ultimately, a deeper appreciation of these interconnected metabolic nodes can refine our understanding of disease mechanisms, paving the way for therapeutic approaches that tackle excitotoxicity, energy deficits, and mood dysregulation in a more holistic manner [389,390].
Beyond exploring prodrug designs and multi-target ligands, integrating emerging technologies—particularly CRISPR-based screening—could elevate the originality and impact of quinoline-focused research [391,392]. CRISPR knockouts or knock-ins enable precise genetic manipulation of targets implicated in the KYN pathway, thereby uncovering how specific enzymes, transporters, or receptors influence the efficacy of quinoline derivatives [393]. This level of control helps delineate off-target effects and refine SAR predictions, ensuring that lead compounds undergo more robust validation before advancing to clinical trials [394]. Additionally, CRISPR-based functional genomics can be paired with advanced phenotypic assays (e.g., high-content imaging or single-cell transcriptomics) to capture subtle changes in cell viability, immune signaling, or neuronal stress responses [395]. Such multimodal approaches can reveal novel synergy points—for instance, identifying an unanticipated metabolic bottleneck that amplifies or diminishes the therapeutic potency of KYNA analogues [396]. By validating these genetic interactions early in the drug development pipeline, investigators can better prioritize candidates with the highest translational potential, thereby reducing costly late-stage failures. [395] Ultimately, the marriage of CRISPR technologies and quinoline-focused medicinal chemistry stands poised to yield a new generation of targeted interventions, leveraging enhanced mechanistic precision to tackle neurodegenerative, oncological, and inflammatory pathologies in a truly innovative manner.
Chemical identity extends well beyond a molecule’s basic formula or nomenclature [397]. Subtle shifts in substituents—such as halogens, hydroxyls, or carboxyl groups—can drastically change a quinoline compound’s lipophilicity, stability, and affinity for target receptors [398]. For instance, introducing a chlorine atom at a strategic position may improve BBB penetration, while adding a bulky moiety might decrease solubility or alter enzyme-mediated metabolism [399]. These seemingly minor alterations can pave the way for novel pharmacological profiles, sometimes swinging the balance between neuroprotection and toxicity [400]. Historically, researchers have leveraged rational design methods to develop promising quinoline analogues with heightened specificity, boosted potency, and reduced side effects [237]. Nonetheless, large-scale synthesis remains challenging when certain derivatives exhibit poor yields or require stringent reaction conditions, thus complicating purification steps [401]. Adding to these hurdles, positional isomerism in the fused ring system can generate multiple byproducts that confound analytical assessments and limit overall compound purity [402]. Recognizing such synthetic barriers is critical for streamlining production pipelines and ensuring that lead candidates meet rigorous quality standards [403]. Overall, mapping out how even the smallest functional group can shift a compound’s fate is key to optimizing existing scaffolds and propelling more targeted therapeutic discoveries.
Biological mechanisms rarely operate in isolation. When a derivative modulates a single receptor or enzyme, it often triggers a ripple effect through multiple pathways, especially in the context of the KYN route [404]. Some compounds, for instance, may tamp down neuroinflammation by directly attenuating excitotoxic signals, only to exert a secondary influence on immune cells by altering cytokine release [405]. Such dual or even multi-pronged interactions underscore the need to see beyond straightforward receptor binding data [406]. For KYN pathway analogues, identifying exact positions in the metabolic cascade reveals potential consequences for redox balance and neuronal homeostasis: a slight shift in enzyme activity could tip the scales from protective antioxidant mechanisms to detrimental oxidative stress [407]. Yet, despite these insights, many indirect targets remain elusive. Off-target interactions—whether they involve uncharacterized receptors or ancillary enzymes—can complicate both therapeutic efficacy and safety [408]. More rigorous mapping of these secondary pathways, using both in vitro and in vivo models, could pinpoint previously overlooked nodes that either amplify or mitigate a compound’s effects [409]. Bridging such gaps would open the door for more targeted preclinical evaluations and eventually guide clinical trials toward meaningful outcomes [410]. Ultimately, unraveling these intricate webs of interaction is crucial for fully harnessing the therapeutic potential of quinoline-based metabolites.
Recent research has propelled several quinoline-based metabolites from exploratory benchwork to more tangible therapeutic leads [242]. In vitro assays frequently highlight their ability to modulate neuroprotective pathways, suppress inflammatory signals, or even arrest tumor cell proliferation [113]. Moving to in vivo models, some of these candidates have displayed meaningful behavioral improvements in neurodegenerative disease paradigms or slowed the progression of certain cancers [411]. However, conflicting data occasionally arise, with a compound that shows potent in vitro efficacy falling short in animal studies due to issues like rapid metabolism or unanticipated toxicity [412]. Such discrepancies underscore the complexity of translating molecular promise into robust, clinically relevant benefits [413]. Despite these hurdles, certain agents continue to stand out [414]. For example, derivatives with enhanced BBB penetration appear especially compelling for psychiatric or neurodegenerative indications, offering a potential edge over older, less permeable analogues [415]. Moreover, targeted delivery systems—ranging from liposomal encapsulations to prodrug designs—are making inroads by bolstering tissue specificity and minimizing off-target effects [416]. Formulation breakthroughs aside, many programs remain in early clinical stages, where gaps in ADME data or sporadic toxicity reports can stall progress [417]. Ultimately, bridging these translational gaps will hinge on ongoing refinements in compound design, alongside more systematic preclinical and clinical assessments.
Though the therapeutic promise of these quinoline derivatives remains compelling, a series of limitations underscores the need for a cautious and well-strategized development path [242]. Low bioavailability, for instance, often impedes their ability to reach efficacious concentrations at the intended site of action, forcing scientists to explore advanced formulation strategies [412]. Prodrug designs and nanoparticle encapsulations stand out as potential solutions, yet they introduce new complexities in manufacturing and regulatory approvals [12]. Additionally, unresolved safety profiles can stall clinical progress, particularly when a compound influences multiple biological pathways that could spark off-target effects [418]. These concerns highlight the importance of rigorous toxicological screening early in the pipeline. Furthermore, complex synthesis methods may hamper large-scale production, driving up costs and limiting accessibility for broader research [419]. Intellectual property barriers can add another layer of difficulty, restricting collaborative efforts and slowing the pace of innovation [420]. In short, no single fix exists, requiring multifaceted strategies and cross-disciplinary collaboration. Recognizing these challenges is essential for shaping the path forward: unless researchers and stakeholders address these technical, financial, and regulatory constraints, the full clinical impact of quinoline-based therapeutics may remain unrealized. By confronting these obstacles head-on, however, the scientific community stands poised to refine existing designs, explore novel delivery routes, and ultimately translate these promising molecules into real-world treatments.
In light of the diverse roles played by KYN pathway enzymes such as IDOs and KMO, incorporating robust biomarker strategies into clinical trial designs becomes essential [421]. This shift aligns with emerging paradigms in mental health diagnostics that emphasize biobehavioral precision over categorical rigidity [422,423]. By measuring baseline gene polymorphisms and enzyme expression levels, clinicians can stratify patients according to their potential responsiveness to quinoline-based agents [424]. This approach mirrors successes in targeted oncology, where genomic profiling matches treatments to precise molecular subtypes [425,426]. In the context of neurological and inflammatory diseases, leveraging a similar paradigm could help avoid the ‘one-size-fits-all’ pitfall, ensuring that individuals most likely to benefit—based on their enzyme activity profiles—are prioritized for quinoline therapies [118]. Such enrichment not only boosts the probability of clinical efficacy but also reduces the risk of adverse reactions in patients whose metabolic idiosyncrasies lead to off-target toxicity [427]. As research into KYN pathway polymorphisms advances, these stratification methods will likely gain further sophistication, enabling personalized interventions that more effectively harness the neuroprotective and anti-inflammatory qualities of quinoline derivatives.
Moreover, polymorphisms in IDOs or KMO can drastically alter flux through the KYN pathway, impacting both the magnitude of neuroprotective metabolites (e.g., KYNA) and the generation of excitotoxic compounds (e.g., QUIN) [428]. This genetic variability underscores why certain patient subgroups respond differently—even within the same clinical trial cohort [429]. Incorporating routine genotyping or enzyme activity assays into trial protocols can clarify which individuals possess ‘favorable’ polymorphisms conducive to enhanced drug efficacy [430]. Conversely, identifying high-risk genotypes might prompt dose adjustments or the addition of secondary agents—like antioxidants or immune modulators—to mitigate potential side effects [431]. By interweaving this biomarker intelligence with standard clinical endpoints, researchers can implement adaptive trial designs that dynamically optimize treatment regimens [432]. This personalized approach stands to accelerate the clinical translation of quinoline-based therapies while reducing attrition rates, effectively bridging the gap between compelling preclinical data and consistent patient outcomes.
Despite this manuscript’s thorough exploration of quinoline-based strategies, a comparative perspective on non-quinoline neuroprotectants—such as indole-based IDO inhibitors or flavonoid antioxidants—could strengthen the rationale for prioritizing quinoline scaffolds [433]. Indole frameworks, while effective at modulating certain enzymes in the KYN pathway, often struggle with limited metabolic stability or suboptimal blood–brain barrier penetration [434]. Likewise, flavonoid antioxidants can offer substantial free-radical scavenging capabilities, yet their broad-spectrum activity and relatively weak target specificity can sometimes lead to off-target effects or insufficient impact on key regulatory nodes [435,436]. In contrast, quinoline cores frequently exhibit a more favorable balance of selective receptor binding and pharmacokinetic versatility, enabling targeted modulation of both excitotoxic and inflammatory cascades [437]. By systematically juxtaposing these scaffold types, researchers would be better equipped to identify which structural features—such as ring fusion motifs or substituent placement—truly confer an edge in neurodegenerative or inflammatory models [438]. Moreover, head-to-head comparisons could clarify cost-effectiveness, synthesis scalability, and the likelihood of off-target liabilities [439]. Ultimately, a robust assessment of non-quinoline contenders would confirm whether quinolines indeed offer the most promising route to next-generation neuroprotectants, thereby ensuring that future research and development efforts are directed toward the scaffolds most likely to yield significant therapeutic benefits.
Looking ahead, several avenues present themselves for propelling this research toward tangible clinical gains. Novel synthetic strategies hold particular promise, as improving the yield or specificity of key intermediates could streamline large-scale production and expedite trials [440]. Equally intriguing is the potential for combining these quinoline derivatives with other pharmacological agents, such as immunomodulators or antioxidants, to tackle multifaceted diseases like neurodegeneration or cancer [441]. These multi-target regimens could not only boost therapeutic impact but also mitigate resistance or relapse [442]. High-priority experiments will likely involve both in vivo and ex vivo models to confirm mechanistic pathways, refine optimal dosing, and uncover synergistic effects [443]. Biomarker-driven assessments that correlate molecular actions with patient outcomes will be vital, offering a clearer rationale for translating preclinical successes into human studies [444]. Ultimately, refining structural features while embracing combination strategies provides a balanced roadmap for researchers eager to unlock the full therapeutic potential of this diverse molecular class.
Each discussion subsection effectively acts as a mini-review that weaves together foundational knowledge, underscores clinical significance, and illuminates new directions. By clustering these quinoline-based compounds according to their structural traits or metabolic origins—be they endogenous, synthetic, or analogues—the overarching narrative becomes clearer. Patterns in receptor engagement, bioavailability, and therapeutic windows emerge, helping researchers pinpoint critical design features for improvement [445,446,447]. This bird’s-eye perspective also reveals shared challenges, such as scalability and off-target toxicity, which might impede clinical translation [448]. Yet, these pitfalls can guide smarter modifications in functional groups or combined treatment strategies, driving progress [446]. Ultimately, evaluating these diverse compounds side by side creates a roadmap of what works, what fails, and how the field can advance. The collaborative spirit behind collecting these insights promises to transform individual leads into robust, evidence-based interventions grounded in the science of quinoline-based therapeutics. Harnessing these consolidated findings can chart new approaches for KYNA–inspired strategies.
5. Conclusions
The structural complexity of quinoline-based metabolites underpins their remarkable versatility across neurodegenerative, inflammatory, and oncological contexts, thereby making them prime candidates for drug development. This review has shown how carefully tuned substitutions—chlorination, fluorination, and esterification—can bolster receptor affinity, metabolic stability, and bioavailability, as demonstrated by derivatives such as 7-CKA (enhanced NMDA blockade) and SZR-81 (improved oral absorption and GPR35 selectivity). Yet, the persistent challenges of limited BBB penetration, off-target interactions (e.g., SZR-105’s kinase inhibition), and rapid clearance (SZR-73) indicate that incremental refinements alone may be insufficient to unlock their full therapeutic value. Moving beyond basic prodrug strategies, a more integrative approach is warranted—one that simultaneously harnesses advanced computational modeling, combinatorial screening, and emerging research on epigenetic modulators. For instance, coupling KYNA analogues with IDO inhibitors could offer synergistic regulation of the KYN pathway, while multi-target ligands like tasquinimod highlight the promise of convergent pharmacological mechanisms. Further, employing novel formulation technologies (e.g., nanoparticle-based delivery) and improved in vivo models can strengthen translational accuracy, fostering therapies that more closely match human pathophysiology. Ultimately, it is by weaving these interdisciplinary strategies into the existing foundation of SAR insights that the next generation of quinoline-based interventions will realize their broad therapeutic potential.
6. Patents
P1500356; PCT/HU2017?000014; EP17759330.8A; US15/082,099: WO 2017149333; A1 2017090. P1500356; PCT/HU2016/050034; US 10,857,236B2 (2020); WO2017021748 (EU).
Author Contributions
“Conceptualization, M.T. and L.V.; methodology, M.T.; software, M.T.; validation, M.T..; formal analysis, M.T.; investigation, M.T.; resources, M.T.; data curation, M.T.; writing—original draft preparation, M.T.; writing—review and editing, M.T.,Á.S, I.S, and L.V.; visualization, M.T.; supervision, L.V.; project administration, L.V.; funding acquisition, L.V. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Research, Development, and Innovation Office—NKFIH K138125, SZTE SZAOK-KKA No: 2022/5S729, and the HUN-REN Hungarian Research Network.
Institutional Review Board Statement
In this section, you should add the Institutional Review Board Statement and approval number, if relevant to your study. You might choose to exclude this statement if the study did not require ethical approval. Please note that the Editorial Office might ask you for further information. Please add “The study was conducted in accordance with the Declaration of Helsinki, and approved by the Institutional Review Board (or Ethics Committee) of NAME OF INSTITUTE (protocol code XXX and date of approval).” for studies involving humans. OR “The animal study protocol was approved by the Institutional Review Board (or Ethics Committee) of NAME OF INSTITUTE (protocol code XXX and date of approval).” for studies involving animals. OR “Ethical review and approval were waived for this study due to REASON (please provide a detailed justification).” OR “Not applicable” for studies not involving humans or animals.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
| BBB | blood-brain barrier |
| 7-CKA | 7-chlorokynurenic acid |
| DPA | dipicolinic acid |
| GPR35 | G protein-coupled receptor 35 |
| HDAC | histone deacetylase |
| IDOs | indoleamine 2,3-dioxygenases |
| INA | isonicotinic acid |
| KYN | kynurenine |
| KYNA | kynurenic acid |
| NA | nicotinic acid |
| NMDA | N-methyl-D-aspartate |
| QUIN | quinolinic acid |
| SAR | structure–activity relationship |
| Trp | tryptophan |
| TDO | tryptophan 2,3-dioxygenase |
| XA | xanthurenic acid |
References
- Stone, T.W.; Williams, R.O. Tryptophan metabolism as a ‘reflex’feature of neuroimmune communication: sensor and effector functions for the indoleamine-2, 3-dioxygenase kynurenine pathway. Journal of Neurochemistry 2024, 168, 3333–3357. [Google Scholar] [CrossRef]
- Lovelace, M.D.; Varney, B.; Sundaram, G.; Lennon, M.J.; Lim, C.K.; Jacobs, K.; Guillemin, G.J.; Brew, B.J. Recent evidence for an expanded role of the kynurenine pathway of tryptophan metabolism in neurological diseases. Neuropharmacology 2017, 112, 373–388. [Google Scholar] [CrossRef] [PubMed]
- Cervenka, I.; Agudelo, L.Z.; Ruas, J.L. Kynurenines: Tryptophan’s metabolites in exercise, inflammation, and mental health. Science 2017, 357, eaaf9794. [Google Scholar] [CrossRef]
- Roth, W.; Zadeh, K.; Vekariya, R.; Ge, Y.; Mohamadzadeh, M. Tryptophan metabolism and gut-brain homeostasis. International journal of molecular sciences 2021, 22, 2973. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Yu, S.; Long, Y.; Shi, A.; Deng, J.; Ma, Y.; Wen, J.; Li, X.; Liu, S.; Zhang, Y. Tryptophan metabolism: Mechanism-oriented therapy for neurological and psychiatric disorders. Frontiers in Immunology 2022, 13, 985378. [Google Scholar] [CrossRef] [PubMed]
- Badawy, A.A. Kynurenine pathway of tryptophan metabolism: regulatory and functional aspects. International journal of tryptophan research 2017, 10, 1178646917691938. [Google Scholar] [CrossRef]
- Wang, Q.; Liu, D.; Song, P.; Zou, M.-H. Deregulated tryptophan-kynurenine pathway is linked to inflammation, oxidative stress, and immune activation pathway in cardiovascular diseases. Frontiers in bioscience (Landmark edition) 2015, 20, 1116. [Google Scholar]
- Tanaka, M.; Szabó, Á.; Vécsei, L. Redefining roles: A paradigm shift in tryptophan–kynurenine metabolism for innovative clinical applications. International journal of molecular sciences 2024, 25, 12767. [Google Scholar] [CrossRef]
- Zhao, Y.-Q.; Li, X.; Guo, H.-Y.; Shen, Q.-K.; Quan, Z.-S.; Luan, T. Application of quinoline ring in structural modification of natural products. Molecules 2023, 28, 6478. [Google Scholar] [CrossRef]
- Ajani, O.O.; Iyaye, K.T.; Ademosun, O.T. Recent advances in chemistry and therapeutic potential of functionalized quinoline motifs–a review. RSC advances 2022, 12, 18594–18614. [Google Scholar] [CrossRef]
- Sehlangia, S.; Nayak, N.; Garg, N.; Pradeep, C.P. Substituent-controlled structural, supramolecular, and cytotoxic properties of a series of 2-Styryl-8-nitro and 2-Styryl-8-hydroxy quinolines. ACS omega 2022, 7, 24838–24850. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Ayala, L.F.; Guzmán-López, E.G.; Galano, A. Quinoline Derivatives: Promising Antioxidants with Neuroprotective Potential. Antioxidants (Basel) 2023, 12. [Google Scholar] [CrossRef]
- Martorana, A.; La Monica, G.; Lauria, A. Quinoline-based molecules targeting c-Met, EGF, and VEGF receptors and the proteins involved in related carcinogenic pathways. Molecules 2020, 25, 4279. [Google Scholar] [CrossRef] [PubMed]
- Shoji, T.; Takeuchi, M.; Uda, M.; Ariga, Y.; Yamazaki, A.; Sekiguchi, R.; Ito, S. Synthesis of Azuleno [2, 1-b] quinolones and Quinolines via Brønsted Acid-Catalyzed Cyclization of 2-Arylaminoazulenes. Molecules 2023, 28, 5785. [Google Scholar] [CrossRef] [PubMed]
- Yadav, V.; Reang, J.; Sharma, V.; Majeed, J.; Sharma, P.C.; Sharma, K.; Giri, N.; Kumar, A.; Tonk, R.K. Quinoline-derivatives as privileged scaffolds for medicinal and pharmaceutical chemists: A comprehensive review. Chemical Biology & Drug Design 2022, 100, 389–418. [Google Scholar]
- Lanz, T.V.; Williams, S.K.; Stojic, A.; Iwantscheff, S.; Sonner, J.K.; Grabitz, C.; Becker, S.; Böhler, L.-I.; Mohapatra, S.R.; Sahm, F. Tryptophan-2, 3-Dioxygenase (TDO) deficiency is associated with subclinical neuroprotection in a mouse model of multiple sclerosis. Scientific reports 2017, 7, 41271. [Google Scholar] [CrossRef] [PubMed]
- Maya, S.; Prakash, T.; Goli, D. Effect of wedelolactone and gallic acid on quinolinic acid-induced neurotoxicity and impaired motor function: Significance to sporadic amyotrophic lateral sclerosis. Neurotoxicology 2018, 68, 1–12. [Google Scholar]
- Platten, M.; Friedrich, M.; Wainwright, D.A.; Panitz, V.; Opitz, C.A. Tryptophan metabolism in brain tumors—IDO and beyond. Current opinion in immunology 2021, 70, 57–66. [Google Scholar] [CrossRef]
- Lovelace, M.D.; Varney, B.; Sundaram, G.; Franco, N.F.; Ng, M.L.; Pai, S.; Lim, C.K.; Guillemin, G.J.; Brew, B.J. Current evidence for a role of the kynurenine pathway of tryptophan metabolism in multiple sclerosis. Frontiers in Immunology 2016, 7, 246. [Google Scholar] [CrossRef]
- Krause, D.; Suh, H.-S.; Tarassishin, L.; Cui, Q.L.; Durafourt, B.A.; Choi, N.; Bauman, A.; Cosenza-Nashat, M.; Antel, J.P.; Zhao, M.-L. The tryptophan metabolite 3-hydroxyanthranilic acid plays anti-inflammatory and neuroprotective roles during inflammation: role of hemeoxygenase-1. The American journal of pathology 2011, 179, 1360–1372. [Google Scholar] [CrossRef]
- Zádori, D.; Veres, G.; Szalárdy, L.; Klivényi, P.; Vécsei, L. Alzheimer’s disease: recent concepts on the relation of mitochondrial disturbances, excitotoxicity, neuroinflammation, and kynurenines. Journal of Alzheimer’s Disease 2018, 62, 523–547. [Google Scholar] [CrossRef] [PubMed]
- Phillips, R.S.; Iradukunda, E.C.; Hughes, T.; Bowen, J.P. Modulation of enzyme activity in the kynurenine pathway by kynurenine monooxygenase inhibition. Frontiers in molecular biosciences 2019, 6, 3. [Google Scholar] [CrossRef]
- Valotto Neto, L.J.; Reverete de Araujo, M.; Moretti Junior, R.C.; Mendes Machado, N.; Joshi, R.K.; Dos Santos Buglio, D.; Barbalho Lamas, C.; Direito, R.; Fornari Laurindo, L.; Tanaka, M.; et al. Investigating the Neuroprotective and Cognitive-Enhancing Effects of Bacopa monnieri: A Systematic Review Focused on Inflammation, Oxidative Stress, Mitochondrial Dysfunction, and Apoptosis. Antioxidants (Basel) 2024, 13. [Google Scholar] [CrossRef] [PubMed]
- Michaudel, C.; Danne, C.; Agus, A.; Magniez, A.; Aucouturier, A.; Spatz, M.; Lefevre, A.; Kirchgesner, J.; Rolhion, N.; Wang, Y. Rewiring the altered tryptophan metabolism as a novel therapeutic strategy in inflammatory bowel diseases. Gut 2023, 72, 1296–1307. [Google Scholar] [CrossRef]
- Opitz, C.A.; Somarribas Patterson, L.F.; Mohapatra, S.R.; Dewi, D.L.; Sadik, A.; Platten, M.; Trump, S. The therapeutic potential of targeting tryptophan catabolism in cancer. British journal of cancer 2020, 122, 30–44. [Google Scholar] [CrossRef] [PubMed]
- Tang, K.; Wu, Y.-H.; Song, Y.; Yu, B. Indoleamine 2, 3-dioxygenase 1 (IDO1) inhibitors in clinical trials for cancer immunotherapy. Journal of Hematology & Oncology 2021, 14, 1–21. [Google Scholar]
- El-Zahabi, M.A.A.; Abouzeid, M.Y.; Eissa, I.H.; Taghour, M.S. DUAL INHIBITORS OF INDOLEAMINE-2, 3-DIOXYGENASE (IDO) AND TRYPTOPHAN-2, 3-DIOXYGENASE (TDO) AS ANTI-TUMOR IMMUNE MODULATORS. Al-Azhar Journal of Pharmaceutical Sciences 2024, 69, 38–61. [Google Scholar] [CrossRef]
- Plitman, E.; Iwata, Y.; Caravaggio, F.; Nakajima, S.; Chung, J.K.; Gerretsen, P.; Kim, J.; Takeuchi, H.; Chakravarty, M.M.; Remington, G. Kynurenic acid in schizophrenia: a systematic review and meta-analysis. Schizophrenia bulletin 2017, 43, 764–777. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.; Zhang, J. Indoleamine 2, 3-dioxygenase (IDO) activity: a perspective biomarker for laboratory determination in tumor immunotherapy. Biomedicines 2023, 11, 1988. [Google Scholar] [CrossRef]
- Stone, T.W. Does kynurenic acid act on nicotinic receptors? An assessment of the evidence. Journal of neurochemistry 2020, 152, 627–649. [Google Scholar] [CrossRef]
- Fukuwatari, T. Possibility of amino acid treatment to prevent the psychiatric disorders via modulation of the production of tryptophan metabolite kynurenic acid. Nutrients 2020, 12, 1403. [Google Scholar] [CrossRef] [PubMed]
- Bauzon, J.; Lee, G.; Cummings, J. Repurposed agents in the Alzheimer’s disease drug development pipeline. Alzheimer's research & therapy 2020, 12, 1–16. [Google Scholar]
- Saranraj, K.; Kiran, P.U. Drug repurposing: Clinical practices and regulatory pathways. Perspectives in Clinical Research 2024, 10.4103.
- Patrignani, P.; Contursi, A.; Tacconelli, S.; Steinhilber, D. The future of pharmacology and therapeutics of the arachidonic acid cascade in the next decade: Innovative advancements in drug repurposing. Frontiers in pharmacology 2024, 15, 1472396. [Google Scholar] [CrossRef] [PubMed]
- PANCHAL, N.B.; VAgHELA, V.M. " From Molecules to Medicine: The Remarkable Pharmacological Odyssey of Quinoline and It's Derivatives". Oriental Journal of Chemistry 2023, 39. [Google Scholar] [CrossRef]
- Atukuri, D.; Vijayalaxmi, S.; Sanjeevamurthy, R.; Vidya, L.; Prasannakumar, R.; Raghavendra, M. Identification of quinoline-chalcones and heterocyclic chalcone-appended quinolines as broad-spectrum pharmacological agents. Bioorganic Chemistry 2020, 105, 104419. [Google Scholar] [CrossRef] [PubMed]
- Elebiju, O.F.; Ajani, O.O.; Oduselu, G.O.; Ogunnupebi, T.A.; Adebiyi, E. Recent advances in functionalized quinoline scaffolds and hybrids—Exceptional pharmacophore in therapeutic medicine. Frontiers in Chemistry 2023, 10, 1074331. [Google Scholar] [CrossRef] [PubMed]
- Fukushima, T.; Umino, M.; Sakamoto, T.; Onozato, M. A review of chromatographic methods for bioactive tryptophan metabolites, kynurenine, kynurenic acid, quinolinic acid, and others, in biological fluids. Biomedical Chromatography 2022, 36, e5308. [Google Scholar] [CrossRef]
- Ostapiuk, A.; Urbanska, E.M. Kynurenic acid in neurodegenerative disorders—unique neuroprotection or double-edged sword? CNS neuroscience & therapeutics 2022, 28, 19–35. [Google Scholar]
- Biernacki, T.; Sandi, D.; Bencsik, K.; Vécsei, L. Kynurenines in the pathogenesis of multiple sclerosis: Therapeutic perspectives. Cells 2020, 9, 1564. [Google Scholar] [CrossRef]
- Tóth, F.; Cseh, E.K.; Vécsei, L. Natural molecules and neuroprotection: kynurenic acid, pantethine and α-lipoic acid. International journal of molecular sciences 2021, 22, 403. [Google Scholar] [CrossRef]
- Ferreira, F.S.; Schmitz, F.; Marques, E.P.; Siebert, C.; Wyse, A.T. Intrastriatal quinolinic acid administration impairs redox homeostasis and induces inflammatory changes: prevention by kynurenic acid. Neurotoxicity Research 2020, 38, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Maitre, M.; Taleb, O.; Jeltsch-David, H.; Klein, C.; Mensah-Nyagan, A.G. Xanthurenic acid: A role in brain intercellular signaling. Journal of Neurochemistry 2024, 168, 2303–2315. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-M.; Huang, C.-Y.; Lai, C.-H.; Chen, Y.-C.; Hwang, Y.-T.; Lin, C.-Y. Neuroprotection effects of kynurenic acid-loaded micelles for the Parkinson’s disease models. Journal of Liposome Research 2024, 34, 593–604. [Google Scholar] [CrossRef] [PubMed]
- Hertelendy, P.; Toldi, J.; Fülöp, F.; Vécsei, L. Ischemic stroke and kynurenines: Medicinal chemistry aspects. Current Medicinal Chemistry 2018, 25, 5945–5957. [Google Scholar] [CrossRef] [PubMed]
- Martín-Hernández, D.; Tendilla-Beltrán, H.; Madrigal, J.L.; García-Bueno, B.; Leza, J.C.; Caso, J.R. Chronic mild stress alters kynurenine pathways changing the glutamate neurotransmission in frontal cortex of rats. Molecular Neurobiology 2019, 56, 490–501. [Google Scholar] [CrossRef]
- Wirthgen, E.; Hoeflich, A.; Rebl, A.; Günther, J. Kynurenic acid: the Janus-faced role of an immunomodulatory tryptophan metabolite and its link to pathological conditions. Frontiers in immunology 2018, 8, 1957. [Google Scholar] [CrossRef]
- Savitz, J. The kynurenine pathway: a finger in every pie. Molecular psychiatry 2020, 25, 131–147. [Google Scholar] [CrossRef]
- Molnár, K.; Lőrinczi, B.; Fazakas, C.; Szatmári, I.; Fülöp, F.; Kmetykó, N.; Berkecz, R.; Ilisz, I.; Krizbai, I.A.; Wilhelm, I. Szr-104, a novel kynurenic acid analogue with high permeability through the blood–brain barrier. Pharmaceutics 2021, 13, 61. [Google Scholar] [CrossRef] [PubMed]
- Juhász, Á.; Ungor, D.; Varga, N.; Katona, G.; Balogh, G.T.; Csapó, E. Lipid-Based nanocarriers for delivery of neuroprotective kynurenic acid: Preparation, characterization, and BBB transport. International Journal of Molecular Sciences 2023, 24, 14251. [Google Scholar] [CrossRef]
- Varga, N.; Csapó, E.; Majláth, Z.; Ilisz, I.; Krizbai, I.A.; Wilhelm, I.; Knapp, L.; Toldi, J.; Vécsei, L.; Dékány, I. Targeting of the kynurenic acid across the blood–brain barrier by core-shell nanoparticles. European Journal of Pharmaceutical Sciences 2016, 86, 67–74. [Google Scholar] [CrossRef]
- Hara, N.; Yamada, K.; Shibata, T.; Osago, H.; Hashimoto, T.; Tsuchiya, M. Elevation of cellular NAD levels by nicotinic acid and involvement of nicotinic acid phosphoribosyltransferase in human cells. Journal of biological chemistry 2007, 282, 24574–24582. [Google Scholar] [CrossRef] [PubMed]
- Xiao, W.; Wang, R.-S.; Handy, D.E.; Loscalzo, J. NAD (H) and NADP (H) redox couples and cellular energy metabolism. Antioxidants & redox signaling 2018, 28, 251–272. [Google Scholar]
- Oyama, T.; Yamamoto, T.; Kameda, T.; Kamiya, T.; Abe, H.; Abe, T.; Tanuma, S.-i. Supplementation of nicotinic acid and its derivatives up-regulates cellular NAD+ level rather than nicotinamide derivatives in cultured normal human epidermal keratinocytes. Life 2024, 14, 413. [Google Scholar] [CrossRef]
- Carlson, L.A. Nicotinic acid: the broad-spectrum lipid drug. A 50th anniversary review. Journal of internal medicine 2005, 258, 94–114. [Google Scholar] [CrossRef] [PubMed]
- Schachter, M. Strategies for modifying high-density lipoprotein cholesterol: a role for nicotinic acid. Cardiovascular drugs and therapy 2005, 19, 415–422. [Google Scholar] [CrossRef]
- Figge, H.L.; Figge, J.; Souney, P.F.; Mutnick, A.H.; Sacks, F. Nicotinic acid: a review of its clinical use in the treatment of lipid disorders. Pharmacotherapy: The Journal of Human Pharmacology and Drug Therapy 1988, 8, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Cheng, K.; Wu, T.-J.; Wu, K.K.; Sturino, C.; Metters, K.; Gottesdiener, K.; Wright, S.D.; Wang, Z.; O’Neill, G.; Lai, E. Antagonism of the prostaglandin D2 receptor 1 suppresses nicotinic acid-induced vasodilation in mice and humans. Proceedings of the National Academy of Sciences 2006, 103, 6682–6687. [Google Scholar] [CrossRef]
- Benyo, Z.; Gille, A.; Bennett, C.L.; Clausen, B.E.; Offermanns, S. Nicotinic acid-induced flushing is mediated by activation of epidermal langerhans cells. Molecular pharmacology 2006, 70, 1844–1849. [Google Scholar] [CrossRef]
- Benyó, Z.; Gille, A.; Kero, J.; Csiky, M.; Suchánková, M.C.; Nüsing, R.M.; Moers, A.; Pfeffer, K.; Offermanns, S. GPR109A (PUMA-G/HM74A) mediates nicotinic acid–induced flushing. The Journal of clinical investigation 2005, 115, 3634–3640. [Google Scholar] [CrossRef]
- Braidy, N.; Grant, R.; Adams, S.; Brew, B.J.; Guillemin, G.J. Mechanism for quinolinic acid cytotoxicity in human astrocytes and neurons. Neurotoxicity research 2009, 16, 77–86. [Google Scholar] [CrossRef]
- Lugo-Huitrón, R.; Ugalde Muñiz, P.; Pineda, B.; Pedraza-Chaverrí, J.; Ríos, C.; Pérez-de la Cruz, V. Quinolinic acid: an endogenous neurotoxin with multiple targets. Oxidative medicine and cellular longevity 2013, 2013, 104024. [Google Scholar] [CrossRef] [PubMed]
- Schurr, A.; West, C.A.; Rigor, B. Neurotoxicity of quinolinic acid and its derivatives in hypoxic rat hippocampal slices. Brain research 1991, 568, 199–204. [Google Scholar] [CrossRef] [PubMed]
- La Cruz, V.P.-D.; Carrillo-Mora, P.; Santamaría, A. Quinolinic acid, an endogenous molecule combining excitotoxicity, oxidative stress and other toxic mechanisms. International Journal of Tryptophan Research 2012, 5, IJTR–S8158. [Google Scholar] [CrossRef] [PubMed]
- Leipnitz, G.; Schumacher, C.; Scussiato, K.; Dalcin, K.B.; Wannmacher, C.M.; Wyse, A.T.; Dutra-Filho, C.S.; Wajner, M.; Latini, A. Quinolinic acid reduces the antioxidant defenses in cerebral cortex of young rats. International Journal of Developmental Neuroscience 2005, 23, 695–701. [Google Scholar] [CrossRef]
- Pérez-Severiano, F.; Rodríguez-Pérez, M.; Pedraza-Chaverrí, J.; Maldonado, P.D.; Medina-Campos, O.N.; Ortíz-Plata, A.; Sánchez-García, A.; Villeda-Hernández, J.; Galván-Arzate, S.; Aguilera, P. S-Allylcysteine, a garlic-derived antioxidant, ameliorates quinolinic acid-induced neurotoxicity and oxidative damage in rats. Neurochemistry international 2004, 45, 1175–1183. [Google Scholar] [CrossRef]
- Schwarcz, R.; Whetsell Jr, W.O.; Mangano, R.M. Quinolinic acid: an endogenous metabolite that produces axon-sparing lesions in rat brain. Science 1983, 219, 316–318. [Google Scholar] [CrossRef] [PubMed]
- Hestad, K.; Alexander, J.; Rootwelt, H.; Aaseth, J.O. The role of tryptophan dysmetabolism and quinolinic acid in depressive and neurodegenerative diseases. Biomolecules 2022, 12, 998. [Google Scholar] [CrossRef] [PubMed]
- Seal, C.J.; Heaton, F.W. Effect of dietary picolinic acid on the metabolism of exogenous and endogenous zinc in the rat. The Journal of nutrition 1985, 115, 986–993. [Google Scholar] [CrossRef]
- Testa, U.; Louache, F.; Titeux, M.; Thomopoulos, P.; Rochant, H. The iron-chelating agent picolinic acid enhances transferrin receptors expression in human erythroleukaemic cell lines. British journal of haematology 1985, 60, 491–502. [Google Scholar] [CrossRef]
- Prodinger, J.; Loacker, L.J.; Schmidt, R.L.; Ratzinger, F.; Greiner, G.; Witzeneder, N.; Hoermann, G.; Jutz, S.; Pickl, W.F.; Steinberger, P. The tryptophan metabolite picolinic acid suppresses proliferation and metabolic activity of CD4+ T cells and inhibits c-Myc activation. Journal of Leucocyte Biology 2016, 99, 583–594. [Google Scholar] [CrossRef]
- Tomioka, H.; Shimizu, T.; Tatano, Y. Effects of picolinic acid on the antimicrobial functions of host macrophages against Mycobacterium avium complex. International journal of antimicrobial agents 2007, 29, 460–464. [Google Scholar] [CrossRef] [PubMed]
- Cai, S.; Sato, K.; Shimizu, T.; Yamabe, S.; Hiraki, M.; Sano, C.; Tomioka, H. Antimicrobial activity of picolinic acid against extracellular and intracellular Mycobacterium avium complex and its combined activity with clarithromycin, rifampicin and fluoroquinolones. Journal of Antimicrobial Chemotherapy 2006, 57, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Bosco, M.C.; Rapisarda, A.; Massazza, S.; Melillo, G.; Young, H.; Varesio, L. The tryptophan catabolite picolinic acid selectively induces the chemokines macrophage inflammatory protein-1α and-1β in macrophages. The Journal of Immunology 2000, 164, 3283–3291. [Google Scholar] [CrossRef] [PubMed]
- Grant, R.; Coggan, S.; Smythe, G.A. The physiological action of picolinic acid in the human brain. International journal of tryptophan research 2009, 2, IJTR–S2469. [Google Scholar] [CrossRef] [PubMed]
- Cockhill, J.; Jhamandas, K.; Boegman, R.; Beninger, R. Action of picolinic acid and structurally related pyridine carboxylic acids on quinolinic acid-induced cortical cholinergic damage. Brain research 1992, 599, 57–63. [Google Scholar] [CrossRef]
- Kalisch, B.E.; Jhamandas, K.; Boegman, R.J.; Beninger, R.J. Picolinic acid protects against quinolinic acid-induced depletion of NADPH diaphorase containing neurons in the rat striatum. Brain Research 1994, 668, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Taleb, O.; Maammar, M.; Klein, C.; Maitre, M.; Mensah-Nyagan, A.G. A role for xanthurenic acid in the control of brain dopaminergic activity. International Journal of Molecular Sciences 2021, 22, 6974. [Google Scholar] [CrossRef]
- Fazio, F.; Lionetto, L.; Curto, M.; Iacovelli, L.; Copeland, C.S.; Neale, S.A.; Bruno, V.; Battaglia, G.; Salt, T.E.; Nicoletti, F. Cinnabarinic acid and xanthurenic acid: Two kynurenine metabolites that interact with metabotropic glutamate receptors. Neuropharmacology 2017, 112, 365–372. [Google Scholar] [CrossRef]
- Murakami, K.; Haneda, M.; Yoshino, M. Prooxidant action of xanthurenic acid and quinoline compounds: role of transition metals in the generation of reactive oxygen species and enhanced formation of 8-hydroxy-2′-deoxyguanosine in DNA. Biometals 2006, 19, 429–435. [Google Scholar] [CrossRef]
- Murakami, K.; Ito, M.; Yoshino, M. Xanthurenic acid inhibits metal ion-induced lipid peroxidation and protects NADP-isocitrate dehydrogenase from oxidative inactivation. Journal of nutritional science and vitaminology 2001, 47, 306–310. [Google Scholar] [CrossRef]
- Lima, V.L.; Dias, F.; Nunes, R.D.; Pereira, L.O.; Santos, T.S.; Chiarini, L.B.; Ramos, T.D.; Silva-Mendes, B.J.; Perales, J.; Valente, R.H. The antioxidant role of xanthurenic acid in the Aedes aegypti midgut during digestion of a blood meal. PloS one 2012, 7, e38349. [Google Scholar] [CrossRef] [PubMed]
- Nowicka-Stążka, P.; Langner, E.; Turski, W.; Rzeski, W.; Parada-Turska, J. Quinaldic acid in synovial fluid of patients with rheumatoid arthritis and osteoarthritis and its effect on synoviocytes in vitro. Pharmacological Reports 2018, 70, 277–283. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, H.; Miyamoto, S.; Mabuchi, H.; Yoneyama, Y.; Takeda, R. Insulin-releasing effect of quinaldic acid and its relatives on isolated Langerhans islets. Biochem Biophys Res Commun 1973, 53, 1297–1303. [Google Scholar] [CrossRef] [PubMed]
- Jhamandas, K.H.; Boegman, R.J.; Beninger, R.J.; Miranda, A.F.; Lipic, K.A. Excitotoxicity of quinolinic acid: modulation by endogenous antagonists. Neurotox Res 2000, 2, 139–155. [Google Scholar] [CrossRef]
- Bala, M.; Radhakrishnan, T.; Kumar, A.; Mishra, G.P.; Dobraia, J.R.; Kirti, P.B. Erratum: Overexpression of a fusion defensin gene from radish and fenugreek improves resistance against leaf spot diseases caused by Cercospora arachidicola and Phaeoisariopsis personata in peanut. Turk J Biol 2019, 43, 154. [Google Scholar] [CrossRef]
- Zheng, Q.; Wang, Q.; Wang, S.; Wu, J.; Gao, Q.; Liu, W. Thiopeptide Antibiotics Exhibit a Dual Mode of Action against Intracellular Pathogens by Affecting Both Host and Microbe. Chem Biol 2015, 22, 1002–1007. [Google Scholar] [CrossRef]
- Jhamandas, K.; Boegman, R.J.; Beninger, R.J.; Bialik, M. Quinolinate-induced cortical cholinergic damage: modulation by tryptophan metabolites. Brain Res 1990, 529, 185–191. [Google Scholar] [CrossRef]
- Anderson, E.W.; Fishbein, J.; Hong, J.; Roeser, J.; Furie, R.A.; Aranow, C.; Volpe, B.T.; Diamond, B.; Mackay, M. Quinolinic acid, a kynurenine/tryptophan pathway metabolite, associates with impaired cognitive test performance in systemic lupus erythematosus. Lupus Sci Med 2021, 8. [Google Scholar] [CrossRef]
- Sawada, Y.; Kayakiri, H.; Abe, Y.; Mizutani, T.; Inamura, N.; Asano, M.; Hatori, C.; Aramori, I.; Oku, T.; Tanaka, H. Discovery of the first non-peptide full agonists for the human bradykinin B(2) receptor incorporating 4-(2-picolyloxy)quinoline and 1-(2-picolyl)benzimidazole frameworks. J Med Chem 2004, 47, 2853–2863. [Google Scholar] [CrossRef]
- Martorana, A.; La Monica, G.; Lauria, A. Quinoline-Based Molecules Targeting c-Met, EGF, and VEGF Receptors and the Proteins Involved in Related Carcinogenic Pathways. Molecules 2020, 25. [Google Scholar] [CrossRef]
- Ghorai, P.; Pal, K.; Karmakar, P.; Saha, A. The development of two fluorescent chemosensors for the selective detection of Zn(2+) and Al(3+) ions in a quinoline platform by tuning the substituents in the receptor part: elucidation of the structures of the metal-bound chemosensors and biological studies. Dalton Trans 2020, 49, 4758–4773. [Google Scholar] [CrossRef] [PubMed]
- Mikata, Y. Quinoline- and isoquinoline-derived ligand design on TQEN (N,N,N',N'-tetrakis(2-quinolylmethyl)ethylenediamine) platform for fluorescent sensing of specific metal ions and phosphate species. Dalton Trans 2020, 49, 17494–17504. [Google Scholar] [CrossRef]
- Peterson, L.R. Quinolone molecular structure-activity relationships: what we have learned about improving antimicrobial activity. Clin Infect Dis 2001, 33 Suppl 3, S180–186. [Google Scholar] [CrossRef]
- Wang, Y.; Xiao, F.; Jin, G. Structural basis of quinolone derivatives, inhibition of type I and II topoisomerases and inquiry into the relevance of bioactivity in odd or even branches with molecular docking study. J Mol Struct 2020, 1221, 128869. [Google Scholar] [CrossRef] [PubMed]
- Murakami, K.; Haneda, M.; Yoshino, M. Prooxidant action of xanthurenic acid and quinoline compounds: role of transition metals in the generation of reactive oxygen species and enhanced formation of 8-hydroxy-2'-deoxyguanosine in DNA. Biometals 2006, 19, 429–435. [Google Scholar] [CrossRef] [PubMed]
- Fray, M.J.; Bull, D.J.; Carr, C.L.; Gautier, E.C.; Mowbray, C.E.; Stobie, A. Structure-activity relationships of 1,4-dihydro-(1H,4H)-quinoxaline-2,3-diones as N-methyl-D-aspartate (glycine site) receptor antagonists. 1. Heterocyclic substituted 5-alkyl derivatives. J Med Chem 2001, 44, 1951–1962. [Google Scholar] [CrossRef]
- Maitre, M.; Taleb, O.; Jeltsch-David, H.; Klein, C.; Mensah-Nyagan, A.G. Xanthurenic acid: A role in brain intercellular signaling. J Neurochem 2024, 168, 2303–2315. [Google Scholar] [CrossRef] [PubMed]
- Hou, Z.; Vanecek, A.S.; Tepe, J.J.; Odom, A.L. Synthesis, structure, properties, and cytotoxicity of a (quinoline)RuCp(+) complex. Dalton Trans 2023, 52, 721–730. [Google Scholar] [CrossRef]
- Liu, J.Y.; Wellen, K.E. Advances into understanding metabolites as signaling molecules in cancer progression. Curr Opin Cell Biol 2020, 63, 144–153. [Google Scholar] [CrossRef]
- Liloia, D.; Zamfira, D.A.; Tanaka, M.; Manuello, J.; Crocetta, A.; Keller, R.; Cozzolino, M.; Duca, S.; Cauda, F.; Costa, T. Disentangling the role of gray matter volume and concentration in autism spectrum disorder: A meta-analytic investigation of 25 years of voxel-based morphometry research. Neuroscience & Biobehavioral Reviews 2024, 105791.
- Boros, F.; Vécsei, L. Progress in the development of kynurenine and quinoline-3-carboxamide-derived drugs. Expert Opin Investig Drugs 2020, 29, 1223–1247. [Google Scholar] [CrossRef]
- Stone, T.W. Neuropharmacology of quinolinic and kynurenic acids. Pharmacol Rev 1993, 45, 309–379. [Google Scholar] [CrossRef] [PubMed]
- Stone, T.W. Kynurenines in the CNS: from endogenous obscurity to therapeutic importance. Prog Neurobiol 2001, 64, 185–218. [Google Scholar] [CrossRef] [PubMed]
- Stone, T.W.; Stoy, N.; Darlington, L.G. An expanding range of targets for kynurenine metabolites of tryptophan. Trends Pharmacol Sci 2013, 34, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Lamson, A.R.; Shelley, M.; Troyanskaya, O. Interpretable neural architecture search and transfer learning for understanding CRISPR-Cas9 off-target enzymatic reactions. Nat Comput Sci 2023, 3, 1056–1066. [Google Scholar] [CrossRef]
- Vieira, P.A.; Shin, C.B.; Arroyo-Currás, N.; Ortega, G.; Li, W.; Keller, A.A.; Plaxco, K.W.; Kippin, T.E. Ultra-High-Precision, in-vivo Pharmacokinetic Measurements Highlight the Need for and a Route Toward More Highly Personalized Medicine. Front Mol Biosci 2019, 6, 69. [Google Scholar] [CrossRef]
- Tanaka, M.; Vécsei, L. Monitoring the kynurenine system: Concentrations, ratios or what else? Adv Clin Exp Med 2021, 30, 775–778. [Google Scholar] [CrossRef]
- Zeng, M.; Yang, L.; He, D.; Li, Y.; Shi, M.; Zhang, J. Metabolic pathways and pharmacokinetics of natural medicines with low permeability. Drug Metab Rev 2017, 49, 464–476. [Google Scholar] [CrossRef]
- Matada, B.S.; Pattanashettar, R.; Yernale, N.G. A comprehensive review on the biological interest of quinoline and its derivatives. Bioorg Med Chem 2021, 32, 115973. [Google Scholar] [CrossRef] [PubMed]
- Carlson, L.A. Nicotinic acid: the broad-spectrum lipid drug. A 50th anniversary review. J Intern Med 2005, 258, 94–114. [Google Scholar] [CrossRef]
- Siddique, S.; Hussain, K.; Shehzadi, N.; Arshad, M.; Arshad, M.N.; Iftikhar, S.; Saghir, F.; Shaukat, A.; Sarfraz, M.; Ahmed, N. Design, synthesis, biological evaluation and molecular docking studies of quinoline-anthranilic acid hybrids as potent anti-inflammatory drugs. Org Biomol Chem 2024, 22, 3708–3724. [Google Scholar] [CrossRef]
- Arabiyat, S.; Alzoubi, A.; Al-Daghistani, H.; Al-Hiari, Y.; Kasabri, V.; Alkhateeb, R. Evaluation of Quinoline-Related Carboxylic Acid Derivatives as Prospective Differentially Antiproliferative, Antioxidative, and Anti-Inflammatory Agents. Chem Biol Drug Des 2024, 104, e14615. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Xie, Y.; Liu, X.; Xu, H.; Ren, H.; Tang, S.; Liu, Q.; Huang, M.; Shao, X.; Li, C.; et al. Discovery of Novel Imidazo[4,5-c]quinoline Derivatives to Treat Inflammatory Bowel Disease (IBD) by Inhibiting Multiple Proinflammatory Signaling Pathways and Restoring Intestinal Homeostasis. J Med Chem 2022, 65, 11949–11969. [Google Scholar] [CrossRef] [PubMed]
- Shang, X.F.; Morris-Natschke, S.L.; Liu, Y.Q.; Guo, X.; Xu, X.S.; Goto, M.; Li, J.C.; Yang, G.Z.; Lee, K.H. Biologically active quinoline and quinazoline alkaloids part I. Med Res Rev 2018, 38, 775–828. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, C.; Liu, W.; Guo, Z.; Zhang, Y.; Wu, Y.; Wei, C.; Wu, J.; Yang, X. Quinolines: A Promising Heterocyclic Scaffold for Cancer Therapeutics. Curr Med Chem 2025, 32, 958–973. [Google Scholar] [CrossRef]
- Park, L.T.; Kadriu, B.; Gould, T.D.; Zanos, P.; Greenstein, D.; Evans, J.W.; Yuan, P.; Farmer, C.A.; Oppenheimer, M.; George, J.M. A randomized trial of the N-methyl-d-aspartate receptor glycine site antagonist prodrug 4-chlorokynurenine in treatment-resistant depression. International Journal of Neuropsychopharmacology 2020, 23, 417–425. [Google Scholar] [CrossRef] [PubMed]
- Heyes, M.P.; Saito, K.; Crowley, J.S.; Davis, L.E.; Demitrack, M.A.; Der, M.; Dilling, L.A.; Elia, J.; Kruesi, M.J.; Lackner, A.; et al. Quinolinic acid and kynurenine pathway metabolism in inflammatory and non-inflammatory neurological disease. Brain 1992, 115 ( Pt 5) Pt 5, 1249–1273. [Google Scholar] [CrossRef]
- Kadela-Tomanek, M.; Jastrzębska, M.; Chrobak, E.; Bębenek, E. Lipophilicity and ADMET Analysis of Quinoline-1,4-quinone Hybrids. Pharmaceutics 2022, 15. [Google Scholar] [CrossRef] [PubMed]
- Elebiju, O.F.; Ajani, O.O.; Oduselu, G.O.; Ogunnupebi, T.A.; Adebiyi, E. Recent advances in functionalized quinoline scaffolds and hybrids-Exceptional pharmacophore in therapeutic medicine. Front Chem 2022, 10, 1074331. [Google Scholar] [CrossRef] [PubMed]
- Platten, M.; Nollen, E.A.A.; Röhrig, U.F.; Fallarino, F.; Opitz, C.A. Tryptophan metabolism as a common therapeutic target in cancer, neurodegeneration and beyond. Nat Rev Drug Discov 2019, 18, 379–401. [Google Scholar] [CrossRef]
- Krause, D.; Suh, H.S.; Tarassishin, L.; Cui, Q.L.; Durafourt, B.A.; Choi, N.; Bauman, A.; Cosenza-Nashat, M.; Antel, J.P.; Zhao, M.L.; et al. The tryptophan metabolite 3-hydroxyanthranilic acid plays anti-inflammatory and neuroprotective roles during inflammation: role of hemeoxygenase-1. Am J Pathol 2011, 179, 1360–1372. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Zhang, S.; Fang, X.; Guo, L.; Hu, N.; Guo, Z.; Li, X.; Yang, S.; He, J.C.; Kuang, C.; et al. N-Benzyl/Aryl Substituted Tryptanthrin as Dual Inhibitors of Indoleamine 2,3-Dioxygenase and Tryptophan 2,3-Dioxygenase. J Med Chem 2019, 62, 9161–9174. [Google Scholar] [CrossRef] [PubMed]
- Dolšak, A.; Gobec, S.; Sova, M. Indoleamine and tryptophan 2,3-dioxygenases as important future therapeutic targets. Pharmacol Ther 2021, 221, 107746. [Google Scholar] [CrossRef] [PubMed]
- Barbalho, S.M.; Leme Boaro, B.; da Silva Camarinha Oliveira, J.; Patočka, J.; Barbalho Lamas, C.; Tanaka, M.; Laurindo, L.F. Molecular Mechanisms Underlying Neuroinflammation Intervention with Medicinal Plants: A Critical and Narrative Review of the Current Literature. Pharmaceuticals 2025, 18, 133. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Vécsei, L. Monitoring the Redox Status in Multiple Sclerosis. Biomedicines 2020, 8. [Google Scholar] [CrossRef]
- Löscher, W.; Gericke, B. Novel Intrinsic Mechanisms of Active Drug Extrusion at the Blood-Brain Barrier: Potential Targets for Enhancing Drug Delivery to the Brain? Pharmaceutics 2020, 12. [Google Scholar] [CrossRef]
- Zuchero, Y.J.; Chen, X.; Bien-Ly, N.; Bumbaca, D.; Tong, R.K.; Gao, X.; Zhang, S.; Hoyte, K.; Luk, W.; Huntley, M.A.; et al. Discovery of Novel Blood-Brain Barrier Targets to Enhance Brain Uptake of Therapeutic Antibodies. Neuron 2016, 89, 70–82. [Google Scholar] [CrossRef]
- Terstappen, G.C.; Meyer, A.H.; Bell, R.D.; Zhang, W. Strategies for delivering therapeutics across the blood-brain barrier. Nat Rev Drug Discov 2021, 20, 362–383. [Google Scholar] [CrossRef]
- Dounay, A.B.; Tuttle, J.B.; Verhoest, P.R. Challenges and Opportunities in the Discovery of New Therapeutics Targeting the Kynurenine Pathway. J Med Chem 2015, 58, 8762–8782. [Google Scholar] [CrossRef]
- Shen, L.L.; Pernet, A.G. Mechanism of inhibition of DNA gyrase by analogues of nalidixic acid: the target of the drugs is DNA. Proc Natl Acad Sci U S A 1985, 82, 307–311. [Google Scholar] [CrossRef]
- Walker, H.A.; Wilson, S.; Farrar, C.; Richardson, A.P. The reflex respiratory and circulatory actions of some cinchoninic acid derivatives and a number of unrelated compounds. J Pharmacol Exp Ther 1952, 104, 211–218. [Google Scholar] [CrossRef]
- de Lima, E.P.; Laurindo, L.F.; Catharin, V.C.S.; Direito, R.; Tanaka, M.; Jasmin Santos German, I.; Lamas, C.B.; Guiguer, E.L.; Araújo, A.C.; Fiorini, A.M.R. Polyphenols, Alkaloids, and Terpenoids Against Neurodegeneration: Evaluating the Neuroprotective Effects of Phytocompounds Through a Comprehensive Review of the Current Evidence. Metabolites 2025, 15, 124. [Google Scholar] [CrossRef] [PubMed]
- Herraiz, T.; Guillén, H.; González-Peña, D.; Arán, V.J. Antimalarial Quinoline Drugs Inhibit β-Hematin and Increase Free Hemin Catalyzing Peroxidative Reactions and Inhibition of Cysteine Proteases. Sci Rep 2019, 9, 15398. [Google Scholar] [CrossRef] [PubMed]
- Kapishnikov, S.; Staalsø, T.; Yang, Y.; Lee, J.; Pérez-Berná, A.J.; Pereiro, E.; Yang, Y.; Werner, S.; Guttmann, P.; Leiserowitz, L.; et al. Mode of action of quinoline antimalarial drugs in red blood cells infected by Plasmodium falciparum revealed in vivo. Proc Natl Acad Sci U S A 2019, 116, 22946–22952. [Google Scholar] [CrossRef] [PubMed]
- Foley, M.; Tilley, L. Quinoline antimalarials: mechanisms of action and resistance and prospects for new agents. Pharmacol Ther 1998, 79, 55–87. [Google Scholar] [CrossRef] [PubMed]
- Currie, J.P.; Brown, R.A.; Will, G. Observations on the treatment of rheumatoid arthritis with butazolidin. Ann Rheum Dis 1953, 12, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Gelfman, D.M. Reflections on quinine and its importance in dermatology today. Clin Dermatol 2021, 39, 900–903. [Google Scholar] [CrossRef] [PubMed]
- Mukusheva, G.K.; Zhasymbekova, A.R.; Seidakhmetova, R.B.; Nurkenov, O.A.; Akishina, E.A.; Petkevich, S.K.; Dikusar, E.A.; Potkin, V.I. Quinine Esters with 1,2-Azole, Pyridine and Adamantane Fragments. Molecules 2022, 27. [Google Scholar] [CrossRef]
- Man-Son-Hing, M.; Wells, G.; Lau, A. Quinine for nocturnal leg cramps: a meta-analysis including unpublished data. J Gen Intern Med 1998, 13, 600–606. [Google Scholar] [CrossRef]
- Gisselmann, G.; Alisch, D.; Welbers-Joop, B.; Hatt, H. Effects of Quinine, Quinidine and Chloroquine on Human Muscle Nicotinic Acetylcholine Receptors. Front Pharmacol 2018, 9, 1339. [Google Scholar] [CrossRef]
- Li, Z.; Jin, X.; Wu, T.; Huang, G.; Wu, K.; Lei, J.; Pan, X.; Yan, N. Structural Basis for Pore Blockade of the Human Cardiac Sodium Channel Na(v) 1.5 by the Antiarrhythmic Drug Quinidine*. Angew Chem Int Ed Engl 2021, 60, 11474–11480. [Google Scholar] [CrossRef]
- Hill, R.J.; Duff, H.J.; Sheldon, R.S. Class I antiarrhythmic drug receptor: biochemical evidence for state-dependent interaction with quinidine and lidocaine. Mol Pharmacol 1989, 36, 150–159. [Google Scholar] [CrossRef] [PubMed]
- Kodama, I.; Toyama, J.; Yamada, K. Open and inactivated sodium channel block by class-I antiarrhythmic drugs. Jpn Heart J 1986, 27 Suppl 1, 83–89. [Google Scholar] [CrossRef]
- Conn, H.L., Jr.; Luchi, R.J. SOME CELLULAR AND METABOLIC CONSIDERATIONS RELATING TO THE ACTION OF QUINIDINE AS A PROTOTYPE ANTIARRHYTHMIC AGENT. Am J Med 1964, 37, 685–699. [Google Scholar] [CrossRef] [PubMed]
- Yan, M.; Fan, P.; Shi, Y.; Feng, L.; Wang, J.; Zhan, G.; Li, B. Stereoselective Blockage of Quinidine and Quinine in the hERG Channel and the Effect of Their Rescue Potency on Drug-Induced hERG Trafficking Defect. Int J Mol Sci 2016, 17. [Google Scholar] [CrossRef]
- Brocks, D.R.; Mehvar, R. Stereoselectivity in the pharmacodynamics and pharmacokinetics of the chiral antimalarial drugs. Clin Pharmacokinet 2003, 42, 1359–1382. [Google Scholar] [CrossRef]
- Malik, M.; Camm, A.J. Evaluation of drug-induced QT interval prolongation: implications for drug approval and labelling. Drug Saf 2001, 24, 323–351. [Google Scholar] [CrossRef] [PubMed]
- Haeusler, I.L.; Chan, X.H.S.; Guérin, P.J.; White, N.J. The arrhythmogenic cardiotoxicity of the quinoline and structurally related antimalarial drugs: a systematic review. BMC Med 2018, 16, 200. [Google Scholar] [CrossRef] [PubMed]
- Reiter, M.J.; Higgins, S.L.; Payne, A.G.; Mann, D.E. Effects of quinidine versus procainamide on the QT interval. Am J Cardiol 1986, 58, 512–516. [Google Scholar] [CrossRef]
- Wang, X.; Zeng, Y.; Sheng, L.; Larson, P.; Liu, X.; Zou, X.; Wang, S.; Guo, K.; Ma, C.; Zhang, G.; et al. A Cinchona Alkaloid Antibiotic That Appears To Target ATP Synthase in Streptococcus pneumoniae. J Med Chem 2019, 62, 2305–2332. [Google Scholar] [CrossRef]
- Boratyński, P.J.; Zielińska-Błajet, M.; Skarżewski, J. Cinchona Alkaloids-Derivatives and Applications. Alkaloids Chem Biol 2019, 82, 29–145. [Google Scholar] [CrossRef]
- Chajdaś, Z.; Kucharska, M.; Wesełucha-Birczyńska, A. Two-Dimensional Correlation Spectroscopy (2D-COS) Tracking of the Formation of Selected Transition Metal Compounds Cu(II) and Cd(II) With Cinchonine and Their Impact on Model Components of Erythrocytes. Appl Spectrosc 2024, 37028241279434. [Google Scholar] [CrossRef] [PubMed]
- Jabłońska-Trypuć, A.; Wydro, U.; Wołejko, E.; Świderski, G.; Lewandowski, W. Biological Activity of New Cichoric Acid-Metal Complexes in Bacterial Strains, Yeast-Like Fungi, and Human Cell Cultures In Vitro. Nutrients 2020, 12. [Google Scholar] [CrossRef] [PubMed]
- Ramić, A.; Skočibušić, M.; Odžak, R.; Čipak Gašparović, A.; Milković, L.; Mikelić, A.; Sović, K.; Primožič, I.; Hrenar, T. Antimicrobial Activity of Quasi-Enantiomeric Cinchona Alkaloid Derivatives and Prediction Model Developed by Machine Learning. Antibiotics (Basel) 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Parveen, S.; Maurya, N.; Meena, A.; Luqman, S. Cinchonine: A Versatile Pharmacological Agent Derived from Natural Cinchona Alkaloids. Curr Top Med Chem 2024, 24, 343–363. [Google Scholar] [CrossRef] [PubMed]
- Kalluraya, B.; Sreenivasa, S. Synthesis and pharmacological properties of some quinoline derivatives. Farmaco 1998, 53, 399–404. [Google Scholar] [CrossRef]
- Chen, C.R.; Malik, M.; Snyder, M.; Drlica, K. DNA gyrase and topoisomerase IV on the bacterial chromosome: quinolone-induced DNA cleavage. J Mol Biol 1996, 258, 627–637. [Google Scholar] [CrossRef]
- Drlica, K.; Zhao, X. DNA gyrase, topoisomerase IV, and the 4-quinolones. Microbiol Mol Biol Rev 1997, 61, 377–392. [Google Scholar] [CrossRef]
- Spencer, A.C.; Panda, S.S. DNA Gyrase as a Target for Quinolones. Biomedicines 2023, 11. [Google Scholar] [CrossRef]
- Fu, H.G.; Li, Z.W.; Hu, X.X.; Si, S.Y.; You, X.F.; Tang, S.; Wang, Y.X.; Song, D.Q. Synthesis and Biological Evaluation of Quinoline Derivatives as a Novel Class of Broad-Spectrum Antibacterial Agents. Molecules 2019, 24. [Google Scholar] [CrossRef]
- Khamkhenshorngphanuch, T.; Kulkraisri, K.; Janjamratsaeng, A.; Plabutong, N.; Thammahong, A.; Manadee, K.; Na Pombejra, S.; Khotavivattana, T. Synthesis and Antimicrobial Activity of Novel 4-Hydroxy-2-quinolone Analogs. Molecules 2020, 25. [Google Scholar] [CrossRef]
- Kania, A.; Tejchman, W.; Pawlak, A.M.; Mokrzyński, K.; Różanowski, B.; Musielak, B.M.; Greczek-Stachura, M. Preliminary Studies of Antimicrobial Activity of New Synthesized Hybrids of 2-Thiohydantoin and 2-Quinolone Derivatives Activated with Blue Light. Molecules 2022, 27. [Google Scholar] [CrossRef] [PubMed]
- Kostopoulou, I.; Tzani, A.; Chronaki, K.; Prousis, K.C.; Pontiki, E.; Hadjiplavlou-Litina, D.; Detsi, A. Novel Multi-Target Agents Based on the Privileged Structure of 4-Hydroxy-2-quinolinone. Molecules 2023, 29. [Google Scholar] [CrossRef] [PubMed]
- Gach-Janczak, K.; Piekielna-Ciesielska, J.; Waśkiewicz, J.; Krakowiak, K.; Wtorek, K.; Janecka, A. Quinolin-4-ones: Methods of Synthesis and Application in Medicine. Molecules 2025, 30. [Google Scholar] [CrossRef]
- Ahmed, M.; Kelley, S.O. Enhancing the Potency of Nalidixic Acid toward a Bacterial DNA Gyrase with Conjugated Peptides. ACS Chem Biol 2017, 12, 2563–2569. [Google Scholar] [CrossRef] [PubMed]
- Naeem, A.; Badshah, S.L.; Muska, M.; Ahmad, N.; Khan, K. The Current Case of Quinolones: Synthetic Approaches and Antibacterial Activity. Molecules 2016, 21, 268. [Google Scholar] [CrossRef]
- Fàbrega, A.; Madurga, S.; Giralt, E.; Vila, J. Mechanism of action of and resistance to quinolones. Microb Biotechnol 2009, 2, 40–61. [Google Scholar] [CrossRef]
- Fernandes, P.B. Mode of action, and in vitro and in vivo activities of the fluoroquinolones. J Clin Pharmacol 1988, 28, 156–168. [Google Scholar] [CrossRef] [PubMed]
- Duckworth, G.J.; Williams, J.D. Frequency of appearance of resistant variants to norfloxacin and nalidixic acid. J Antimicrob Chemother 1984, 13 Suppl B, 33–38. [Google Scholar] [CrossRef]
- Majalekar, P.P.; Shirote, P.J. Fluoroquinolones: Blessings Or Curses. Curr Drug Targets 2020, 21, 1354–1370. [Google Scholar] [CrossRef]
- Barlow, A.M. NALIDIXIC ACID IN INFECTIONS OF URINARY TRACT. LABORATORY AND CLINICAL INVESTIGATIONS. Br Med J 1963, 2, 1308–1310. [Google Scholar] [CrossRef]
- Reeves, D.S.; Lacey, R.W.; Mummery, R.V.; Mahendra, M.; Bint, A.J.; Newsom, S.W. Treatment of acute urinary infection by norfloxacin or nalidixic acid/citrate: a multi-centre comparative study. J Antimicrob Chemother 1984, 13 Suppl B, 99–105. [Google Scholar] [CrossRef]
- Engle, E.C.; Manes, S.H.; Drlica, K. Differential effects of antibiotics inhibiting gyrase. J Bacteriol 1982, 149, 92–98. [Google Scholar] [CrossRef]
- Mizuuchi, K.; O'Dea, M.H.; Gellert, M. DNA gyrase: subunit structure and ATPase activity of the purified enzyme. Proc Natl Acad Sci U S A 1978, 75, 5960–5963. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.L.; Heller, K.; Gellert, M.; Zubay, G. Differential sensitivity of gene expression in vitro to inhibitors of DNA gyrase. Proc Natl Acad Sci U S A 1979, 76, 3304–3308. [Google Scholar] [CrossRef] [PubMed]
- Palma, E.; Tilocca, B.; Roncada, P. Antimicrobial resistance in veterinary medicine: An overview. International journal of molecular sciences 2020, 21, 1914. [Google Scholar] [CrossRef] [PubMed]
- Martinez, M.N.; Watts, J.L.; Gilbert, J.M. Questions associated with the development of novel drugs intended for the treatment of bacterial infections in veterinary species. The Veterinary Journal 2019, 248, 79–85. [Google Scholar] [CrossRef]
- Pomba, C.; Rantala, M.; Greko, C.; Baptiste, K.E.; Catry, B.; Van Duijkeren, E.; Mateus, A.; Moreno, M.A.; Pyörälä, S.; Ružauskas, M. Public health risk of antimicrobial resistance transfer from companion animals. Journal of Antimicrobial Chemotherapy 2017, 72, 957–968. [Google Scholar] [CrossRef]
- Wolfson, J.S.; Hooper, D.C. The fluoroquinolones: structures, mechanisms of action and resistance, and spectra of activity in vitro. Antimicrob Agents Chemother 1985, 28, 581–586. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Xie, Y.; Li, Z. Room-Temperature Phosphorescence of Nicotinic Acid and Isonicotinic Acid: Efficient Intermolecular Hydrogen-Bond Interaction in Molecular Array. J Phys Chem Lett 2022, 13, 1652–1659. [Google Scholar] [CrossRef]
- Rajan, K.; Jaw, R.; Grecz, N. Role of chelation and water binding of calcium in dormancy and heat resistance of bacterial endospores. Bioinorganic Chemistry 1978, 8, 477–491. [Google Scholar] [CrossRef]
- Ragab, A.E.; Badawy, E.T.; Aboukhatwa, S.M.; Abdel-Aziz, M.M.; Kabbash, A.; Abo Elseoud, K.A. Isonicotinic acid N-oxide, from isoniazid biotransformation by Aspergillus niger, as an InhA inhibitor antituberculous agent against multiple and extensively resistant strains supported by in silico docking and ADME prediction. Nat Prod Res 2023, 37, 1687–1692. [Google Scholar] [CrossRef] [PubMed]
- Metcalfe, C.; Macdonald, I.K.; Murphy, E.J.; Brown, K.A.; Raven, E.L.; Moody, P.C. The tuberculosis prodrug isoniazid bound to activating peroxidases. J Biol Chem 2008, 283, 6193–6200. [Google Scholar] [CrossRef] [PubMed]
- Volynets, G.P.; Tukalo, M.A.; Bdzhola, V.G.; Derkach, N.M.; Gumeniuk, M.I.; Tarnavskiy, S.S.; Yarmoluk, S.M. Novel isoniazid derivative as promising antituberculosis agent. Future Microbiol 2020, 15, 869–879. [Google Scholar] [CrossRef] [PubMed]
- Judge, V.; Narasimhan, B.; Ahuja, M.; Sriram, D.; Yogeeswari, P.; De Clercq, E.; Pannecouque, C.; Balzarini, J. Synthesis, antimycobacterial, antiviral, antimicrobial activity and QSAR studies of N(2)-acyl isonicotinic acid hydrazide derivatives. Med Chem 2013, 9, 53–76. [Google Scholar] [CrossRef] [PubMed]
- Lewis, J.C. Germination of bacterial spores by calcium chelates of dipicolinic acid analogues. Journal of Biological Chemistry 1972, 247, 1861–1868. [Google Scholar] [CrossRef]
- Hintze, P.E.; Nicholson, W.L. Single-spore elemental analyses indicate that dipicolinic acid-deficient Bacillus subtilis spores fail to accumulate calcium. Archives of microbiology 2010, 192, 493–497. [Google Scholar] [CrossRef]
- Chen, A.Y.; Thomas, P.W.; Stewart, A.C.; Bergstrom, A.; Cheng, Z.; Miller, C.; Bethel, C.R.; Marshall, S.H.; Credille, C.V.; Riley, C.L.; et al. Dipicolinic Acid Derivatives as Inhibitors of New Delhi Metallo-β-lactamase-1. J Med Chem 2017, 60, 7267–7283. [Google Scholar] [CrossRef]
- Church, B.D.; Halvorson, H. Dependence of the heat resistance of bacterial endospores on their dipicolinic acid content. Nature 1959, 183, 124–125. [Google Scholar] [CrossRef]
- Grecz, N.; Tang, T. Relation of dipicolinic acid to heat resistance of bacterial spores. J Gen Microbiol 1970, 63, 303–310. [Google Scholar] [CrossRef]
- Setlow, B.; Atluri, S.; Kitchel, R.; Koziol-Dube, K.; Setlow, P. Role of dipicolinic acid in resistance and stability of spores of Bacillus subtilis with or without DNA-protective alpha/beta-type small acid-soluble proteins. J Bacteriol 2006, 188, 3740–3747. [Google Scholar] [CrossRef]
- Summers, K.L.; Pushie, M.J.; Sopasis, G.J.; James, A.K.; Dolgova, N.V.; Sokaras, D.; Kroll, T.; Harris, H.H.; Pickering, I.J.; George, G.N. Solution Chemistry of Copper(II) Binding to Substituted 8-Hydroxyquinolines. Inorg Chem 2020, 59, 13858–13874. [Google Scholar] [CrossRef] [PubMed]
- Pierre, J.L.; Baret, P.; Serratrice, G. Hydroxyquinolines as iron chelators. Curr Med Chem 2003, 10, 1077–1084. [Google Scholar] [CrossRef] [PubMed]
- Pape, V.F.S.; May, N.V.; Gál, G.T.; Szatmári, I.; Szeri, F.; Fülöp, F.; Szakács, G.; Enyedy É, A. Impact of copper and iron binding properties on the anticancer activity of 8-hydroxyquinoline derived Mannich bases. Dalton Trans 2018, 47, 17032–17045. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Wu, H.; Wang, J.; Ji, Z.; Fang, T.; Lu, H.; Yan, L.; Shen, F.; Zhang, D.; Jiang, Y.; et al. Discovery of Novel 8-Hydroxyquinoline Derivatives with Potent In Vitro and In Vivo Antifungal Activity. J Med Chem 2023, 66, 16364–16376. [Google Scholar] [CrossRef] [PubMed]
- Shen, A.Y.; Chen, C.P.; Roffler, S. A chelating agent possessing cytotoxicity and antimicrobial activity: 7-morpholinomethyl-8-hydroxyquinoline. Life Sci 1999, 64, 813–825. [Google Scholar] [CrossRef] [PubMed]
- Cherdtrakulkiat, R.; Boonpangrak, S.; Sinthupoom, N.; Prachayasittikul, S.; Ruchirawat, S.; Prachayasittikul, V. Derivatives (halogen, nitro and amino) of 8-hydroxyquinoline with highly potent antimicrobial and antioxidant activities. Biochem Biophys Rep 2016, 6, 135–141. [Google Scholar] [CrossRef]
- Prati, F.; Bergamini, C.; Fato, R.; Soukup, O.; Korabecny, J.; Andrisano, V.; Bartolini, M.; Bolognesi, M.L. Novel 8-Hydroxyquinoline Derivatives as Multitarget Compounds for the Treatment of Alzheimer's Disease. ChemMedChem 2016, 11, 1284–1295. [Google Scholar] [CrossRef]
- Fernández-Bachiller, M.I.; Pérez, C.; González-Muñoz, G.C.; Conde, S.; López, M.G.; Villarroya, M.; García, A.G.; Rodríguez-Franco, M.I. Novel tacrine-8-hydroxyquinoline hybrids as multifunctional agents for the treatment of Alzheimer's disease, with neuroprotective, cholinergic, antioxidant, and copper-complexing properties. J Med Chem 2010, 53, 4927–4937. [Google Scholar] [CrossRef]
- Prachayasittikul, V.; Prachayasittikul, S.; Ruchirawat, S.; Prachayasittikul, V. 8-Hydroxyquinolines: a review of their metal chelating properties and medicinal applications. Drug Des Devel Ther 2013, 7, 1157–1178. [Google Scholar] [CrossRef]
- Fazio, F.; Lionetto, L.; Curto, M.; Iacovelli, L.; Cavallari, M.; Zappulla, C.; Ulivieri, M.; Napoletano, F.; Capi, M.; Corigliano, V.; et al. Xanthurenic Acid Activates mGlu2/3 Metabotropic Glutamate Receptors and is a Potential Trait Marker for Schizophrenia. Sci Rep 2015, 5, 17799. [Google Scholar] [CrossRef]
- Fazio, F.; Lionetto, L.; Curto, M.; Iacovelli, L.; Copeland, C.S.; Neale, S.A.; Bruno, V.; Battaglia, G.; Salt, T.E.; Nicoletti, F. Cinnabarinic acid and xanthurenic acid: Two kynurenine metabolites that interact with metabotropic glutamate receptors. Neuropharmacology 2017, 112, 365–372. [Google Scholar] [CrossRef] [PubMed]
- Taleb, O.; Maammar, M.; Brumaru, D.; Bourguignon, J.J.; Schmitt, M.; Klein, C.; Kemmel, V.; Maitre, M.; Mensah-Nyagan, A.G. Xanthurenic acid binds to neuronal G-protein-coupled receptors that secondarily activate cationic channels in the cell line NCB-20. PLoS One 2012, 7, e48553. [Google Scholar] [CrossRef] [PubMed]
- Haruki, H.; Hovius, R.; Pedersen, M.G.; Johnsson, K. Tetrahydrobiopterin Biosynthesis as a Potential Target of the Kynurenine Pathway Metabolite Xanthurenic Acid. J Biol Chem 2016, 291, 652–657. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, L.; Larsson, A.; Begum, A.; Iakovleva, I.; Carlsson, M.; Brännström, K.; Sauer-Eriksson, A.E.; Olofsson, A. Modifications of the 7-Hydroxyl Group of the Transthyretin Ligand Luteolin Provide Mechanistic Insights into Its Binding Properties and High Plasma Specificity. PLoS One 2016, 11, e0153112. [Google Scholar] [CrossRef] [PubMed]
- Kontermann, R.E. Half-life extended biotherapeutics. Expert Opin Biol Ther 2016, 16, 903–915. [Google Scholar] [CrossRef] [PubMed]
- Contella, L.; Farrell, C.L.; Boccuto, L.; Litwin, A.; Snyder, M.L. Gene Variant Frequencies of IDO1, IDO2, TDO, and KMO in Substance Use Disorder Cohorts. Genes (Basel) 2024, 15. [Google Scholar] [CrossRef]
- Procaccini, C.; Santopaolo, M.; Faicchia, D.; Colamatteo, A.; Formisano, L.; de Candia, P.; Galgani, M.; De Rosa, V.; Matarese, G. Role of metabolism in neurodegenerative disorders. Metabolism 2016, 65, 1376–1390. [Google Scholar] [CrossRef]
- Andrade, J.C.O.; do Vale, T.M.; Gomes, R.L.M.; Forezi, L.; de Souza, M.; Batalha, P.N.; Boechat, F. Exploring 4-quinolone-3-carboxamide derivatives: A versatile framework for emerging biological applications. Bioorg Chem 2025, 157, 108240. [Google Scholar] [CrossRef]
- Verma, S.; Lal, S.; Narang, R.; Sudhakar, K. Quinoline Hydrazide/Hydrazone Derivatives: Recent Insights on Antibacterial Activity and Mechanism of Action. ChemMedChem 2023, 18, e202200571. [Google Scholar] [CrossRef]
- Stone, T.W.; Forrest, C.M.; Darlington, L.G. Kynurenine pathway inhibition as a therapeutic strategy for neuroprotection. Febs j 2012, 279, 1386–1397. [Google Scholar] [CrossRef]
- Liu, X.; Lv, H.; Guo, Y.; Teka, T.; Wang, X.; Huang, Y.; Han, L.; Pan, G. Structure-Based Reactivity Profiles of Reactive Metabolites with Glutathione. Chem Res Toxicol 2020, 33, 1579–1593. [Google Scholar] [CrossRef] [PubMed]
- Musiol, R. An overview of quinoline as a privileged scaffold in cancer drug discovery. Expert Opin Drug Discov 2017, 12, 583–597. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Yang, Y.; Wang, Z.; Li, H.; Dong, C.; Zhang, X. Recent Advances in Pro-PROTAC Development to Address On-Target Off-Tumor Toxicity. J Med Chem 2023, 66, 8428–8440. [Google Scholar] [CrossRef] [PubMed]
- Robles, O.; Romo, D. Chemo- and site-selective derivatizations of natural products enabling biological studies. Nat Prod Rep 2014, 31, 318–334. [Google Scholar] [CrossRef] [PubMed]
- Ajani, O.O.; Iyaye, K.T.; Ademosun, O.T. Recent advances in chemistry and therapeutic potential of functionalized quinoline motifs - a review. RSC Adv 2022, 12, 18594–18614. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Fang, J.J.; Wang, Z.Y.; Xie, Y.P.; Lu, X. Structural diversity of copper(I) alkynyl cluster-based coordination polymers utilizing bifunctional pyridine carboxylic acid ligands. Nanoscale 2024, 16, 17817–17824. [Google Scholar] [CrossRef]
- Vitaku, E.; Smith, D.T.; Njardarson, J.T. Analysis of the structural diversity, substitution patterns, and frequency of nitrogen heterocycles among U.S. FDA approved pharmaceuticals. J Med Chem 2014, 57, 10257–10274. [Google Scholar] [CrossRef]
- Garcia, C. Tuning receptor signaling through ligand engineering. The FASEB Journal 2020, 34, 1–1. [Google Scholar] [CrossRef]
- Abraham, M.H.; Acree Jr, W.E. On the solubility of nicotinic acid and isonicotinic acid in water and organic solvents. The Journal of Chemical Thermodynamics 2013, 61, 74–78. [Google Scholar] [CrossRef]
- Kotammagari, T.K.; Saleh, L.Y.; Lönnberg, T. Organometallic modification confers oligonucleotides new functionalities. Chem Commun (Camb) 2024, 60, 3118–3128. [Google Scholar] [CrossRef]
- MURAKAMI, K.; UEDA, T.; MORIKAWA, R.; ITO, M.; HANEDA, M.; YOSHINO, M. Antioxidant effect of dipicolinic acid on the metal-catalyzed lipid peroxidation and enzyme inactivation. Biomedical Research 1998, 19, 205–208. [Google Scholar] [CrossRef]
- Dattatray Shinde, S.; Kumar Behera, S.; Kulkarni, N.; Dewangan, B.; Sahu, B. Bifunctional backbone modified squaramide dipeptides as amyloid beta (Aβ) aggregation inhibitors. Bioorg Med Chem 2024, 97, 117538. [Google Scholar] [CrossRef] [PubMed]
- Bhat, S.A.; Henry, R.J.; Blanchard, A.C.; Stoica, B.A.; Loane, D.J.; Faden, A.I. Enhanced Akt/GSK-3β/CREB signaling mediates the anti-inflammatory actions of mGluR5 positive allosteric modulators in microglia and following traumatic brain injury in male mice. J Neurochem 2021, 156, 225–248. [Google Scholar] [CrossRef]
- Jiang, J.; Dingledine, R. Prostaglandin receptor EP2 in the crosshairs of anti-inflammation, anti-cancer, and neuroprotection. Trends Pharmacol Sci 2013, 34, 413–423. [Google Scholar] [CrossRef]
- Falcucci, R.M.; Wertz, R.; Green, J.L.; Meucci, O.; Salvino, J.; Fontana, A.C.K. Novel Positive Allosteric Modulators of Glutamate Transport Have Neuroprotective Properties in an in Vitro Excitotoxic Model. ACS Chem Neurosci 2019, 10, 3437–3453. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Li, W.X.; Dai, S.X.; Guo, Y.C.; Han, F.F.; Zheng, J.J.; Li, G.H.; Huang, J.F. Meta-Analysis of Parkinson's Disease and Alzheimer's Disease Revealed Commonly Impaired Pathways and Dysregulation of NRF2-Dependent Genes. J Alzheimers Dis 2017, 56, 1525–1539. [Google Scholar] [CrossRef]
- Yang, L.; Nao, J.; Dong, X. The Therapeutic Potential of Hydroxycinnamic Acid Derivatives in Parkinson's Disease: Focus on In Vivo Research Advancements. J Agric Food Chem 2023, 71, 10932–10951. [Google Scholar] [CrossRef] [PubMed]
- Pagotto, G.L.d.O.; Santos, L.M.O.d.; Osman, N.; Lamas, C.B.; Laurindo, L.F.; Pomini, K.T.; Guissoni, L.M.; Lima, E.P.d.; Goulart, R.d.A.; Catharin, V.M.S. Ginkgo biloba: a leaf of hope in the fight against Alzheimer’s dementia: clinical trial systematic review. Antioxidants 2024, 13, 651. [Google Scholar] [CrossRef]
- Salomé, C.; Salomé-Grosjean, E.; Park, K.D.; Morieux, P.; Swendiman, R.; DeMarco, E.; Stables, J.P.; Kohn, H. Synthesis and anticonvulsant activities of (R)-N-(4'-substituted)benzyl 2-acetamido-3-methoxypropionamides. J Med Chem 2010, 53, 1288–1305. [Google Scholar] [CrossRef]
- Sharifi, M.; Buzatu, D.; Harris, S.; Wilkes, J. Development of models for predicting Torsade de Pointes cardiac arrhythmias using perceptron neural networks. BMC Bioinformatics 2017, 18, 497. [Google Scholar] [CrossRef]
- Assis de Oliveira, B.; Gonçalves de Oliveira, F.; de Assis Cruz, O.; Mendonça de Assis, P.; Glanzmann, N.; David da Silva, A.; Rezende Barbosa Raposo, N.; Zabala Capriles Goliatt, P.V. In Silico and In Vitro Evaluation of Quinoline Derivatives as Potential Inhibitors of AChE, BACE1, and GSK3β for Neurodegenerative Diseases Treatment. Chem Biodivers 2025, 22, e202401629. [Google Scholar] [CrossRef] [PubMed]
- Zindo, F.T.; Joubert, J.; Malan, S.F. Propargylamine as functional moiety in the design of multifunctional drugs for neurodegenerative disorders: MAO inhibition and beyond. Future Med Chem 2015, 7, 609–629. [Google Scholar] [CrossRef] [PubMed]
- Janse van Rensburg, H.D.; Terre'Blanche, G.; van der Walt, M.M.; Legoabe, L.J. 5-Substituted 2-benzylidene-1-tetralone analogues as A(1) and/or A(2A) antagonists for the potential treatment of neurological conditions. Bioorg Chem 2017, 74, 251–259. [Google Scholar] [CrossRef]
- Bongarzone, S.; Bolognesi, M.L. The concept of privileged structures in rational drug design: focus on acridine and quinoline scaffolds in neurodegenerative and protozoan diseases. Expert Opin Drug Discov 2011, 6, 251–268. [Google Scholar] [CrossRef] [PubMed]
- Jun, J.V.; Petersson, E.J.; Chenoweth, D.M. Rational Design and Facile Synthesis of a Highly Tunable Quinoline-Based Fluorescent Small-Molecule Scaffold for Live Cell Imaging. J Am Chem Soc 2018, 140, 9486–9493. [Google Scholar] [CrossRef] [PubMed]
- Foster, A.C.; Kemp, J.A.; Leeson, P.D.; Grimwood, S.; Donald, A.E.; Marshall, G.R.; Priestley, T.; Smith, J.D.; Carling, R.W. Kynurenic acid analogues with improved affinity and selectivity for the glycine site on the N-methyl-D-aspartate receptor from rat brain. Mol Pharmacol 1992, 41, 914–922. [Google Scholar] [CrossRef] [PubMed]
- Kemp, J.A.; Priestley, T. Effects of (+)-HA-966 and 7-chlorokynurenic acid on the kinetics of N-methyl-D-aspartate receptor agonist responses in rat cultured cortical neurons. Mol Pharmacol 1991, 39, 666–670. [Google Scholar] [CrossRef]
- Kloog, Y.; Lamdani-Itkin, H.; Sokolovsky, M. The glycine site of the N-methyl-D-aspartate receptor channel: differences between the binding of HA-966 and of 7-chlorokynurenic acid. J Neurochem 1990, 54, 1576–1583. [Google Scholar] [CrossRef]
- Wu, H.Q.; Salituro, F.G.; Schwarcz, R. Enzyme-catalyzed production of the neuroprotective NMDA receptor antagonist 7-chlorokynurenic acid in the rat brain in vivo. Eur J Pharmacol 1997, 319, 13–20. [Google Scholar] [CrossRef]
- Yadav, P.; Shah, K. Quinolines, a perpetual, multipurpose scaffold in medicinal chemistry. Bioorg Chem 2021, 109, 104639. [Google Scholar] [CrossRef]
- Dolšak, A.; Švajger, U.; Lešnik, S.; Konc, J.; Gobec, S.; Sova, M. Selective Toll-like receptor 7 agonists with novel chromeno[3,4-d]imidazol-4(1H)-one and 2-(trifluoromethyl)quinoline/ quinazoline-4-amine scaffolds. Eur J Med Chem 2019, 179, 109–122. [Google Scholar] [CrossRef] [PubMed]
- Konieczny, J.; Ossowska, K.; Schulze, G.; Coper, H.; Wolfarth, S. L-701,324, a selective antagonist at the glycine site of the NMDA receptor, counteracts haloperidol-induced muscle rigidity in rats. Psychopharmacology (Berl) 1999, 143, 235–243. [Google Scholar] [CrossRef]
- Kemp, J.A.; Foster, A.C.; Leeson, P.D.; Priestley, T.; Tridgett, R.; Iversen, L.L.; Woodruff, G.N. 7-Chlorokynurenic acid is a selective antagonist at the glycine modulatory site of the N-methyl-D-aspartate receptor complex. Proc Natl Acad Sci U S A 1988, 85, 6547–6550. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, J.C.; Procureur, D.; Wood, P.L. 7-Chlorokynurenate prevents NMDA-induced and kainate-induced striatal lesions. Brain Res 1993, 620, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.L.; Wang, S.J.; Liu, M.M.; Shi, H.S.; Zhang, R.X.; Liu, J.F.; Ding, Z.B.; Lu, L. Glycine site N-methyl-D-aspartate receptor antagonist 7-CTKA produces rapid antidepressant-like effects in male rats. J Psychiatry Neurosci 2013, 38, 306–316. [Google Scholar] [CrossRef]
- Koek, W.; Colpaert, F.C. Selective blockade of N-methyl-D-aspartate (NMDA)-induced convulsions by NMDA antagonists and putative glycine antagonists: relationship with phencyclidine-like behavioral effects. J Pharmacol Exp Ther 1990, 252, 349–357. [Google Scholar] [CrossRef] [PubMed]
- Wood, E.R.; Bussey, T.J.; Phillips, A.G. A glycine antagonist 7-chlorokynurenic acid attenuates ischemia-induced learning deficits. Neuroreport 1993, 4, 151–154. [Google Scholar] [CrossRef] [PubMed]
- Park, L.T.; Kadriu, B.; Gould, T.D.; Zanos, P.; Greenstein, D.; Evans, J.W.; Yuan, P.; Farmer, C.A.; Oppenheimer, M.; George, J.M.; et al. A Randomized Trial of the N-Methyl-d-Aspartate Receptor Glycine Site Antagonist Prodrug 4-Chlorokynurenine in Treatment-Resistant Depression. Int J Neuropsychopharmacol 2020, 23, 417–425. [Google Scholar] [CrossRef] [PubMed]
- Baron, B.M.; Harrison, B.L.; Miller, F.P.; McDonald, I.A.; Salituro, F.G.; Schmidt, C.J.; Sorensen, S.M.; White, H.S.; Palfreyman, M.G. Activity of 5,7-dichlorokynurenic acid, a potent antagonist at the N-methyl-D-aspartate receptor-associated glycine binding site. Mol Pharmacol 1990, 38, 554–561. [Google Scholar] [CrossRef]
- Baron, B.M.; Siegel, B.W.; Slone, A.L.; Harrison, B.L.; Palfreyman, M.G.; Hurt, S.D. [3H]5,7-dichlorokynurenic acid, a novel radioligand labels NMDA receptor-associated glycine binding sites. Eur J Pharmacol 1991, 206, 149–154. [Google Scholar] [CrossRef] [PubMed]
- McNamara, D.; Smith, E.C.; Calligaro, D.O.; O'Malley, P.J.; McQuaid, L.A.; Dingledine, R. 5,7-Dichlorokynurenic acid, a potent and selective competitive antagonist of the glycine site on NMDA receptors. Neurosci Lett 1990, 120, 17–20. [Google Scholar] [CrossRef] [PubMed]
- Hurt, S.D.; Baron, B.M. [3H] 5,7-dichlorokynurenic acid, a high affinity ligand for the NMDA receptor glycine regulatory site. J Recept Res 1991, 11, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Corbett, R.; Dunn, R.W. Effects of 5,7 dichlorokynurenic acid on conflict, social interaction and plus maze behaviors. Neuropharmacology 1993, 32, 461–466. [Google Scholar] [CrossRef] [PubMed]
- Plaznik, A.; Palejko, W.; Nazar, M.; Jessa, M. Effects of antagonists at the NMDA receptor complex in two models of anxiety. Eur Neuropsychopharmacol 1994, 4, 503–512. [Google Scholar] [CrossRef]
- Hlinák, Z.; Krejci, I. Kynurenic acid and 5,7-dichlorokynurenic acids improve social and object recognition in male rats. Psychopharmacology (Berl) 1995, 120, 463–469. [Google Scholar] [CrossRef]
- Priestley, T.; Laughton, P.; Macaulay, A.J.; Hill, R.G.; Kemp, J.A. Electrophysiological characterisation of the antagonist properties of two novel NMDA receptor glycine site antagonists, L-695,902 and L-701,324. Neuropharmacology 1996, 35, 1573–1581. [Google Scholar] [CrossRef] [PubMed]
- Dannhardt, G.; Kohl, B.K. The glycine site on the NMDA receptor: structure-activity relationships and possible therapeutic applications. Curr Med Chem 1998, 5, 253–263. [Google Scholar] [CrossRef] [PubMed]
- Kotlinska, J.; Liljequist, S. A characterization of anxiolytic-like actions induced by the novel NMDA/glycine site antagonist, L-701,324. Psychopharmacology (Berl) 1998, 135, 175–181. [Google Scholar] [CrossRef]
- Potschka, H.; Löscher, W.; Wlaź, P.; Behl, B.; Hofmann, H.P.; Treiber, H.J.; Szabo, L. LU 73068, a new non-NMDA and glycine/NMDA receptor antagonist: pharmacological characterization and comparison with NBQX and L-701,324 in the kindling model of epilepsy. Br J Pharmacol 1998, 125, 1258–1266. [Google Scholar] [CrossRef]
- Bristow, L.J.; Flatman, K.L.; Hutson, P.H.; Kulagowski, J.J.; Leeson, P.D.; Young, L.; Tricklebank, M.D. The atypical neuroleptic profile of the glycine/N-methyl-D-aspartate receptor antagonist, L-701,324, in rodents. The Journal of pharmacology and experimental therapeutics 1996, 277, 578–585. [Google Scholar] [CrossRef]
- Christoph, T.; Reissmüller, E.; Schiene, K.; Englberger, W.; Chizh, B.A. Antiallodynic effects of NMDA glycine(B) antagonists in neuropathic pain: possible peripheral mechanisms. Brain Res 2005, 1048, 218–227. [Google Scholar] [CrossRef] [PubMed]
- Lees, K.R.; Asplund, K.; Carolei, A.; Davis, S.M.; Diener, H.-C.; Kaste, M.; Orgogozo, J.-M.; Whitehead, J. Glycine antagonist (gavestinel) in neuroprotection (GAIN International) in patients with acute stroke: a randomised controlled trial. The Lancet 2000, 355, 1949–1954. [Google Scholar] [CrossRef]
- Banks, P.; Franks, N.P.; Dickinson, R. Competitive inhibition at the glycine site of the N-methyl-D-aspartate receptor mediates xenon neuroprotection against hypoxia-ischemia. Anesthesiology 2010, 112, 614–622. [Google Scholar] [CrossRef] [PubMed]
- Benneyworth, M.A.; Basu, A.C.; Coyle, J.T. Discordant behavioral effects of psychotomimetic drugs in mice with altered NMDA receptor function. Psychopharmacology 2011, 213, 143–153. [Google Scholar] [CrossRef]
- Haley Jr, E.C.; Thompson, J.L.; Levin, B.; Davis, S.; Lees, K.R.; Pittman, J.G.; DeRosa, J.T.; Ordronneau, P.; Brown, D.L.; Sacco, R.L. Gavestinel does not improve outcome after acute intracerebral hemorrhage: an analysis from the GAIN International and GAIN Americas studies. Stroke 2005, 36, 1006–1010. [Google Scholar] [CrossRef] [PubMed]
- Sacco, R.L.; DeRosa, J.T.; Haley Jr, E.C.; Levin, B.; Ordronneau, P.; Phillips, S.J.; Rundek, T.; Snipes, R.G.; Thompson, J.L.; Investigators, G.A. Glycine antagonist in neuroprotection for patients with acute stroke: GAIN Americas: a randomized controlled trial. Jama 2001, 285, 1719–1728. [Google Scholar] [CrossRef]
- Jiang, T.; Zhang, W.; Wang, Z. Laquinimod protects against TNF-α-induced attachment of monocytes to human aortic endothelial cells (HAECs) by increasing the expression of KLF2. Drug Design, Development and Therapy 2020, 1683-1691.
- Nessler, S.; Ott, M.; Wegner, C.; Hayardeny, L.; Ullrich, E.; Bruck, W. Laquinimod suppresses CNS autoimmunity by activation of natural killer cells (P1. 161). Neurology 2015, 84, P1–161. [Google Scholar] [CrossRef]
- Ott, M.; Avendaño-Guzmán, E.; Ullrich, E.; Dreyer, C.; Strauss, J.; Harden, M.; Schön, M.; Schön, M.P.; Bernhardt, G.; Stadelmann, C. Laquinimod, a prototypic quinoline-3-carboxamide and aryl hydrocarbon receptor agonist, utilizes a CD155-mediated natural killer/dendritic cell interaction to suppress CNS autoimmunity. Journal of Neuroinflammation 2019, 16, 1–14. [Google Scholar] [CrossRef]
- Ellrichmann, G.; Blusch, A.; Fatoba, O.; Brunner, J.; Reick, C.; Hayardeny, L.; Hayden, M.; Sehr, D.; Winklhofer, K.F.; Saft, C. Laquinimod treatment in the R6/2 mouse model. Scientific reports 2017, 7, 4947. [Google Scholar] [CrossRef]
- Rothhammer, V.; Kenison, J.E.; Li, Z.; Tjon, E.; Takenaka, M.C.; Chao, C.-C.; Alves de Lima, K.; Borucki, D.M.; Kaye, J.; Quintana, F.J. Aryl hydrocarbon receptor activation in astrocytes by laquinimod ameliorates autoimmune inflammation in the CNS. Neurology: Neuroimmunology & Neuroinflammation 2021, 8, e946. [Google Scholar]
- Comi, G.; Jeffery, D.; Kappos, L.; Montalban, X.; Boyko, A.; Rocca, M.A.; Filippi, M. Placebo-controlled trial of oral laquinimod for multiple sclerosis. New England Journal of Medicine 2012, 366, 1000–1009. [Google Scholar] [CrossRef] [PubMed]
- Vollmer, T.; Sorensen, P.; Selmaj, K.; Zipp, F.; Havrdova, E.; Cohen, J.; Sasson, N.; Gilgun-Sherki, Y.; Arnold, D.; Group, B.S. A randomized placebo-controlled phase III trial of oral laquinimod for multiple sclerosis. Journal of neurology 2014, 261, 773–783. [Google Scholar] [CrossRef] [PubMed]
- Comi, G.; Dadon, Y.; Sasson, N.; Steinerman, J.R.; Knappertz, V.; Vollmer, T.L.; Boyko, A.; Vermersch, P.; Ziemssen, T.; Montalban, X. CONCERTO: a randomized, placebo-controlled trial of oral laquinimod in relapsing-remitting multiple sclerosis. Multiple Sclerosis Journal 2022, 28, 608–619. [Google Scholar] [CrossRef] [PubMed]
- Isaacs, J.T. The long and winding road for the development of tasquinimod as an oral second-generation quinoline-3-carboxamide antiangiogenic drug for the treatment of prostate cancer. Expert opinion on investigational drugs 2010, 19, 1235–1243. [Google Scholar] [CrossRef]
- Shen, L.; Sundstedt, A.; Ciesielski, M.; Miles, K.M.; Celander, M.; Adelaiye, R.; Orillion, A.; Ciamporcero, E.; Ramakrishnan, S.; Ellis, L. Tasquinimod modulates suppressive myeloid cells and enhances cancer immunotherapies in murine models. Cancer immunology research 2015, 3, 136–148. [Google Scholar] [CrossRef]
- Wobus, M.; Balaian, E.; Oelschlaegel, U.; Towers, R.; Möbus, K.; Habermann, I.; Wehner, R.; Stölzel, F.; Chavakis, T.; Törngren, M. Targeting the Inflammatory Niche in MDS By Tasquinimod Restores Hematopoietic Support and Suppresses Immune-Checkpoint Expression in Vitro. Blood 2021, 138, 2596. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, X.; Liu, C.; Peng, Q.; Wu, Y.; Wen, Y.; Zheng, R.; Yan, Q.; Ma, J. Macrophage-Biomimetic Nanoparticles Ameliorate Ulcerative Colitis through Reducing Inflammatory Factors Expression. J Innate Immun 2022, 14, 380–392. [Google Scholar] [CrossRef]
- Cristóvão, J.S.; Gomes, C.M. S100 Proteins in Alzheimer's Disease. Front Neurosci 2019, 13, 463. [Google Scholar] [CrossRef]
- Leimkühler, N.B.; Gleitz, H.F.E.; Ronghui, L.; Snoeren, I.A.M.; Fuchs, S.N.R.; Nagai, J.S.; Banjanin, B.; Lam, K.H.; Vogl, T.; Kuppe, C.; et al. Heterogeneous bone-marrow stromal progenitors drive myelofibrosis via a druggable alarmin axis. Cell Stem Cell 2021, 28, 637–652.e638. [Google Scholar] [CrossRef]
- Xu, K.; Dai, X.L.; Huang, H.C.; Jiang, Z.F. Targeting HDACs: a promising therapy for Alzheimer's disease. Oxid Med Cell Longev 2011, 2011, 143269. [Google Scholar] [CrossRef]
- Ganai, S.A.; Ramadoss, M.; Mahadevan, V. Histone Deacetylase (HDAC) Inhibitors - emerging roles in neuronal memory, learning, synaptic plasticity and neural regeneration. Curr Neuropharmacol 2016, 14, 55–71. [Google Scholar] [CrossRef] [PubMed]
- Yadav, P.; Shah, K. Quinolines, a perpetual, multipurpose scaffold in medicinal chemistry. Bioorganic Chemistry 2021, 109, 104639. [Google Scholar] [CrossRef] [PubMed]
- Kleckner, N.; Dingledine, R. Selectivity of quinoxalines and kynurenines as antagonists of the glycine site on N-methyl-D-aspartate receptors. Molecular pharmacology 1989, 36, 430–436. [Google Scholar] [CrossRef]
- Culletta, G.; Tutone, M.; Zappalà, M.; Almerico, A.M. Sulfonamide moiety as “molecular chimera” in the design of new drugs. Current Medicinal Chemistry 2023, 30, 128–163. [Google Scholar] [CrossRef]
- Brnardic, E.J.; Ye, G.; Brooks, C.; Donatelli, C.; Barton, L.; McAtee, J.; Sanchez, R.M.; Shu, A.; Erhard, K.; Terrell, L. Discovery of pyrrolidine sulfonamides as selective and orally bioavailable antagonists of transient receptor potential vanilloid-4 (TRPV4). Journal of Medicinal Chemistry 2018, 61, 9738–9755. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Wang, C.; Zhang, X.; Zhu, J.; Wang, L.; Ji, M.; Zhang, Z.; Ji, X.-M.; Wang, S.-L. Perfluorooctane sulfonate disrupts the blood brain barrier through the crosstalk between endothelial cells and astrocytes in mice. Environmental Pollution 2020, 256, 113429. [Google Scholar] [CrossRef]
- Ma, H.; Brust, T.; Frankowski, K.J.; Lovell, K.M.; Cameron, M.D.; Bohn, L.M.; Aube, J. Advances in Sulfonamide Kappa Opioid Receptor Antagonists: Structural Refinement and Evaluation of CNS Clearance. ACS Chemical Neuroscience 2022, 13, 1315–1332. [Google Scholar] [CrossRef] [PubMed]
- Zong, P.; Feng, J.; Yue, Z.; Li, Y.; Wu, G.; Sun, B.; He, Y.; Miller, B.; Albert, S.Y.; Su, Z. Functional coupling of TRPM2 and extrasynaptic NMDARs exacerbates excitotoxicity in ischemic brain injury. Neuron 2022, 110, 1944–1958. [Google Scholar] [CrossRef]
- Shen, Z.; Xiang, M.; Chen, C.; Ding, F.; Wang, Y.; Shang, C.; Xin, L.; Zhang, Y.; Cui, X. Glutamate excitotoxicity: Potential therapeutic target for ischemic stroke. Biomedicine & pharmacotherapy 2022, 151, 113125. [Google Scholar]
- Butcher, S.P.; Henshall, D.C.; Teramura, Y.; Iwasaki, K.; Sharkey, J. Neuroprotective actions of FK506 in experimental stroke: in vivo evidence against an antiexcitotoxic mechanism. Journal of Neuroscience 1997, 17, 6939–6946. [Google Scholar] [CrossRef]
- Singh, D.; Wasan, H.; Reeta, K. Preclinical stroke research and translational failure: a bird’s eye view on preventable variables. Cellular and Molecular Neurobiology 2022, 42, 2003–2017. [Google Scholar] [CrossRef]
- Cui, L.l.; Golubczyk, D.; Tolppanen, A.M.; Boltze, J.; Jolkkonen, J. Cell therapy for ischemic stroke: are differences in preclinical and clinical study design responsible for the translational loss of efficacy? Annals of neurology 2019, 86, 5–16. [Google Scholar] [CrossRef] [PubMed]
- Dirnagl, U.; Fisher, M. International, multicenter randomized preclinical trials in translational stroke research: it's time to act. 2012, 32, 933-935.
- Le Naour, J.; Galluzzi, L.; Zitvogel, L.; Kroemer, G.; Vacchelli, E. Trial watch: IDO inhibitors in cancer therapy. Oncoimmunology 2020, 9, 1777625. [Google Scholar] [CrossRef] [PubMed]
- Van den Eynde, B.J.; van Baren, N.; Baurain, J.-F. Is there a clinical future for IDO1 inhibitors after the failure of epacadostat in melanoma? Annual Review of Cancer Biology 2020, 4, 241–256. [Google Scholar] [CrossRef]
- Muller, A.J.; Manfredi, M.G.; Zakharia, Y.; Prendergast, G.C. Inhibiting IDO pathways to treat cancer: lessons from the ECHO-301 trial and beyond. In Proceedings of the Seminars in immunopathology; 2019; pp. 41–48. [Google Scholar]
- Fujiwara, Y.; Kato, S.; Nesline, M.K.; Conroy, J.M.; DePietro, P.; Pabla, S.; Kurzrock, R. Indoleamine 2, 3-dioxygenase (IDO) inhibitors and cancer immunotherapy. Cancer treatment reviews 2022, 110, 102461. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Kumar, A.B.; Finnes, H.; Markovic, S.N.; Park, S.; Dronca, R.S.; Dong, H. Combining immune checkpoint inhibitors with conventional cancer therapy. Frontiers in immunology 2018, 9, 1739. [Google Scholar] [CrossRef]
- Desnoyer, A.; Broutin, S.; Delahousse, J.; Maritaz, C.; Blondel, L.; Mir, O.; Chaput, N.; Paci, A. Pharmacokinetic/pharmacodynamic relationship of therapeutic monoclonal antibodies used in oncology: Part 2, immune checkpoint inhibitor antibodies. European Journal of Cancer 2020, 128, 119–128. [Google Scholar] [CrossRef]
- Metropulos, A.E.; Munshi, H.G.; Principe, D.R. The difficulty in translating the preclinical success of combined TGFβ and immune checkpoint inhibition to clinical trial. EBioMedicine 2022, 86. [Google Scholar] [CrossRef]
- Tanaka, M.; Vécsei, L. From Lab to Life: Exploring Cutting-Edge Models for Neurological and Psychiatric Disorders. Biomedicines 2024, 12. [Google Scholar] [CrossRef]
- Giacomini, P.S.; Bar-Or, A. Laquinimod in multiple sclerosis. Clinical Immunology 2012, 142, 38–43. [Google Scholar] [CrossRef]
- Brück, W.; Zamvil, S.S. Laquinimod, a once-daily oral drug in development for the treatment of relapsing–remitting multiple sclerosis. Expert review of clinical pharmacology 2012, 5, 245–256. [Google Scholar] [CrossRef] [PubMed]
- Haggiag, S.; Ruggieri, S.; Gasperini, C. Efficacy and safety of laquinimod in multiple sclerosis: current status. Therapeutic advances in neurological disorders 2013, 6, 343–352. [Google Scholar] [CrossRef]
- Ahmad, S.R.; Zeyaullah, M.; Khan, M.S.; AlShahrani, A.M.; Altijani, A.A.G.; Ali, H.; Dawria, A.; Mohieldin, A.; Alam, M.S.; Mohamed, A.O.A. Pharmacogenomics for neurodegenerative disorders-a focused review. Frontiers in Pharmacology 2024, 15, 1478964. [Google Scholar] [CrossRef] [PubMed]
- Toader, C.; Tataru, C.P.; Munteanu, O.; Serban, M.; Covache-Busuioc, R.-A.; Ciurea, A.V.; Enyedi, M. Decoding Neurodegeneration: A Review of Molecular Mechanisms and Therapeutic Advances in Alzheimer’s, Parkinson’s, and ALS. International Journal of Molecular Sciences 2024, 25, 12613. [Google Scholar] [CrossRef] [PubMed]
- Priestley, T.; Laughton, P.; Macaulay, A.J.; Hill, R.G.; Kemp, J.A. Electrophysiological characterisation of the antagonist properties of two novel NMDA receptor glycine site antagonists, L-695,902 and L-701,324. Neuropharmacology 1996, 35, 1573–1581. [Google Scholar] [CrossRef] [PubMed]
- Zellinger, C.; Salvamoser, J.D.; Soerensen, J.; van Vliet, E.A.; Aronica, E.; Gorter, J.; Potschka, H. Pre-treatment with the NMDA receptor glycine-binding site antagonist L-701,324 improves pharmacosensitivity in a mouse kindling model. Epilepsy research 2014, 108, 634–643. [Google Scholar] [CrossRef]
- Konieczny, J.; Ossowska, K.; Schulze, G.; Coper, H.; Wolfarth, S. L-701,324, a selective antagonist at the glycine site of the NMDA receptor, counteracts haloperidol-induced muscle rigidity in rats. Psychopharmacology 1999, 143, 235–243. [Google Scholar] [CrossRef]
- Obrenovitch, T.P.; Hardy, A.M.; Zilkha, E. Effects of L-701,324, a high-affinity antagonist at the N-methyl-D-aspartate (NMDA) receptor glycine site, on the rat electroencephalogram. Naunyn-Schmiedeberg's archives of pharmacology 1997, 355, 779–786. [Google Scholar] [CrossRef]
- Egunlusi, A.O.; Joubert, J. NMDA receptor antagonists: emerging insights into molecular mechanisms and clinical applications in neurological disorders. Pharmaceuticals 2024, 17, 639. [Google Scholar] [CrossRef]
- Lotti, J.S.; Jones, J.; Farnsworth, J.C.; Yi, F.; Zhao, F.; Menniti, F.S.; Volkmann, R.A.; Clausen, R.P.; Hansen, K.B. Evaluation of allosteric N-methyl-d-aspartate receptor modulation by GluN2A-selective antagonists using pharmacological equilibrium modeling. Molecular pharmacology 2025, 107, 100004. [Google Scholar] [CrossRef]
- Ahmed, H.; Haider, A.; Ametamey, S.M. N-Methyl-D-Aspartate (NMDA) receptor modulators: a patent review (2015-present). Expert opinion on therapeutic patents 2020, 30, 743–767. [Google Scholar] [CrossRef]
- Verma, S.; Ambatwar, R.; Datusalia, A.K.; Khatik, G.L. Convenient One-Pot Synthesis of Kynurenic Acid Ethyl Ester and Exploration to Direct Synthesis of Neuroprotective Kynurenic Acid and Amide Derivatives. The Journal of Organic Chemistry 2023, 88, 10494–10500. [Google Scholar] [CrossRef] [PubMed]
- Xiong, B.; Wang, Y.; Chen, Y.; Xing, S.; Liao, Q.; Chen, Y.; Li, Q.; Li, W.; Sun, H. Strategies for structural modification of small molecules to improve blood–brain barrier penetration: a recent perspective. Journal of Medicinal Chemistry 2021, 64, 13152–13173. [Google Scholar] [CrossRef] [PubMed]
- Isaacs, J.T.; Antony, L.; Dalrymple, S.L.; Brennen, W.N.; Gerber, S.; Hammers, H.; Wissing, M.; Kachhap, S.; Luo, J.; Xing, L. Tasquinimod is an allosteric modulator of HDAC4 survival signaling within the compromised cancer microenvironment. Cancer research 2013, 73, 1386–1399. [Google Scholar] [CrossRef]
- Raymond, E.; Dalgleish, A.; Damber, J.-E.; Smith, M.; Pili, R. Mechanisms of action of tasquinimod on the tumour microenvironment. Cancer Chemotherapy and Pharmacology 2014, 73, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Joseph, J.; Gonzalez-Lopez, M.; Galang, C.; Garcia, C.; Lemar, H.; Lu, J.; Vaidya, K.; Fischer, M.; Frey, C.; Alam, M. Small-molecule antagonists of the aryl hydrocarbon receptor (AhR) promote activation of human PBMCs in vitro and demonstrate significant impact on tumor growth and immune modulation in vivo. Cancer research 2018, 78, 4719–4719. [Google Scholar] [CrossRef]
- Musiol, R. An overview of quinoline as a privileged scaffold in cancer drug discovery. Expert Opinion on Drug Discovery 2017, 12, 583–597. [Google Scholar] [CrossRef]
- Zheng, H.; Dai, Q.; Yuan, Z.; Fan, T.; Zhang, C.; Liu, Z.; Chu, B.; Sun, Q.; Chen, Y.; Jiang, Y. Quinazoline-based hydroxamic acid derivatives as dual histone methylation and deacetylation inhibitors for potential anticancer agents. Bioorganic & Medicinal Chemistry 2022, 53, 116524. [Google Scholar]
- Ma, C.; Taghour, M.S.; Belal, A.; Mehany, A.B.M.; Mostafa, N.; Nabeeh, A.; Eissa, I.H.; Al-Karmalawy, A.A. Design and Synthesis of New Quinoxaline Derivatives as Potential Histone Deacetylase Inhibitors Targeting Hepatocellular Carcinoma: In Silico, In Vitro, and SAR Studies. Front Chem 2021, 9, 725135. [Google Scholar] [CrossRef]
- Sforzini, L.; Nettis, M.A.; Mondelli, V.; Pariante, C.M. Inflammation in cancer and depression: a starring role for the kynurenine pathway. Psychopharmacology (Berl) 2019, 236, 2997–3011. [Google Scholar] [CrossRef] [PubMed]
- Maddison, D.C.; Giorgini, F. The kynurenine pathway and neurodegenerative disease. Semin Cell Dev Biol 2015, 40, 134–141. [Google Scholar] [CrossRef] [PubMed]
- Isaacs, J.T.; Antony, L.; Dalrymple, S.L.; Brennen, W.N.; Gerber, S.; Hammers, H.; Wissing, M.; Kachhap, S.; Luo, J.; Xing, L.; et al. Tasquinimod Is an Allosteric Modulator of HDAC4 survival signaling within the compromised cancer microenvironment. Cancer Res 2013, 73, 1386–1399. [Google Scholar] [CrossRef] [PubMed]
- Correia, A.C.; Monteiro, A.R.; Silva, R.; Moreira, J.N.; Sousa Lobo, J.M.; Silva, A.C. Lipid nanoparticles strategies to modify pharmacokinetics of central nervous system targeting drugs: Crossing or circumventing the blood-brain barrier (BBB) to manage neurological disorders. Adv Drug Deliv Rev 2022, 189, 114485. [Google Scholar] [CrossRef] [PubMed]
- Khare, P.; Edgecomb, S.X.; Hamadani, C.M.; Tanner, E.E.L.; D, S.M. Lipid nanoparticle-mediated drug delivery to the brain. Adv Drug Deliv Rev 2023, 197, 114861. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, M.I.; Lopes, C.M.; Amaral, M.H.; Costa, P.C. Surface-modified lipid nanocarriers for crossing the blood-brain barrier (BBB): A current overview of active targeting in brain diseases. Colloids Surf B Biointerfaces 2023, 221, 112999. [Google Scholar] [CrossRef] [PubMed]
- Rahimi, S.; Khoee, S.; Ghandi, M. Preparation and characterization of rod-like chitosan-quinoline nanoparticles as pH-responsive nanocarriers for quercetin delivery. Int J Biol Macromol 2019, 128, 279–289. [Google Scholar] [CrossRef]
- Ren, B.; Guo, C.; Liu, R.-Z.; Bian, Z.-Y.; Liu, R.-C.; Huang, L.-F.; Tang, J.-J. Imidazolylacetophenone oxime-based multifunctional neuroprotective agents: Discovery and structure-activity relationships. European journal of medicinal chemistry 2022, 228, 114031. [Google Scholar] [CrossRef] [PubMed]
- Knyihar-Csillik, E.; Mihaly, A.; Krisztin-Peva, B.; Robotka, H.; Szatmari, I.; Fulop, F.; Toldi, J.; Csillik, B.; Vecsei, L. The kynurenate analog SZR-72 prevents the nitroglycerol-induced increase of c-fos immunoreactivity in the rat caudal trigeminal nucleus: comparative studies of the effects of SZR-72 and kynurenic acid. Neuroscience research 2008, 61, 429–432. [Google Scholar] [CrossRef]
- Kassai, F.; Kedves, R.; Gyertyán, I.; Tuka, B.; Fülöp, F.; Toldi, J.; Lendvai, B.; Vécsei, L. Effect of a kynurenic acid analog on home-cage activity and body temperature in rats. Pharmacological Reports 2015, 67, 1188–1192. [Google Scholar] [CrossRef]
- Martos, D.; Lőrinczi, B.; Szatmári, I.; Vécsei, L.; Tanaka, M. The impact of C-3 side chain modifications on Kynurenic Acid: a behavioral analysis of its analogs in the Motor Domain. International journal of molecular sciences 2024, 25, 3394. [Google Scholar] [CrossRef]
- Poles, M.Z.; Nászai, A.; Gulácsi, L.; Czakó, B.L.; Gál, K.G.; Glenz, R.J.; Dookhun, D.; Rutai, A.; Tallósy, S.P.; Szabó, A. Kynurenic acid and its synthetic derivatives protect against sepsis-associated neutrophil activation and brain mitochondrial dysfunction in rats. Frontiers in Immunology 2021, 12, 717157. [Google Scholar] [CrossRef] [PubMed]
- Mándi, Y.; Endrész, V.; Mosolygó, T.; Burián, K.; Lantos, I.; Fülöp, F.; Szatmári, I.; Lőrinczi, B.; Balog, A.; Vécsei, L. The opposite effects of kynurenic acid and different kynurenic acid analogs on tumor necrosis factor-α (TNF-α) production and tumor necrosis factor-stimulated gene-6 (TSG-6) expression. Frontiers in Immunology 2019, 10, 1406. [Google Scholar] [CrossRef] [PubMed]
- Knapp, L.; Szita, B.; Kocsis, K.; Vécsei, L.; Toldi, J. Nitroglycerin enhances the propagation of cortical spreading depression: comparative studies with sumatriptan and novel kynurenic acid analogues. Drug Design, Development and Therapy 2016, 27-34.
- Tanaka, M.; Szabó, Á.; Lőrinczi, B.; Szatmári, I.; Fülöp, F.; Vécsei, L.; Tanaka, M.; Szabó, Á.; Lőrinczi, B.; Szatmári, I. Antidepressant-like Effects of Kynurenic Acid Analogues. In Proceedings of the 1st International Electronic Conference on Biomedicine; 2021; p. 26. [Google Scholar]
- Evelin, H.L.F. Kinurénsav-analógok in vitro elektrofiziológiai vizsgálata az ischaemiás stroke szempontjából. Szegedi Tudomanyegyetem (Hungary), 2022.
- Leeson, P.D.; Baker, R.; Carling, R.W.; Curtis, N.R.; Moore, K.W.; Williams, B.J.; Foster, A.C.; Donald, A.E.; Kemp, J.A.; Marshall, G.R. Kynurenic acid derivatives. Structure-activity relationships for excitatory amino acid antagonism and identification of potent and selective antagonists at the glycine site on the N-methyl-D-aspartate receptor. Journal of medicinal chemistry 1991, 34, 1243–1252. [Google Scholar] [CrossRef] [PubMed]
- Vasilopoulos, A.; Krska, S.W.; Stahl, S.S. C (sp3)–H methylation enabled by peroxide photosensitization and Ni-mediated radical coupling. Science 2021, 372, 398–403. [Google Scholar] [CrossRef]
- Comeau, C.; Ries, B.; Stadelmann, T.; Tremblay, J.; Poulet, S.; Frohlich, U.; Côté, J.; Boudreault, P.-L.; Derbali, R.M.; Sarret, P. Modulation of the passive permeability of semipeptidic macrocycles: N-and C-methylations fine-tune conformation and properties. Journal of Medicinal Chemistry 2021, 64, 5365–5383. [Google Scholar] [CrossRef]
- Jóźwiak, K.; Płazińska, A. Structural insights into ligand—receptor interactions involved in biased agonism of G-protein coupled receptors. Molecules 2021, 26, 851. [Google Scholar] [CrossRef]
- Liu, L.; Ge, J.; Fu, H.; Wang, J.; Chen, Y.; Sun, Y.; Zhao, C.; Li, Y.; Li, Y. A fluorinated prodrug strategy enhances the therapeutic index of nanoparticle-delivered hydrophilic drugs. Colloids Surf B Biointerfaces 2025, 249, 114539. [Google Scholar] [CrossRef]
- Chen, K.J.; Plaunt, A.J.; Leifer, F.G.; Kang, J.Y.; Cipolla, D. Recent advances in prodrug-based nanoparticle therapeutics. Eur J Pharm Biopharm 2021, 165, 219–243. [Google Scholar] [CrossRef]
- Tanaka, M.; Tuka, B.; Vécsei, L. Navigating the Neurobiology of Migraine: From pathways to potential therapies. 2024, 13, 1098.
- Deora, G.S.; Kantham, S.; Chan, S.; Dighe, S.N.; Veliyath, S.K.; McColl, G.; Parat, M.O.; McGeary, R.P.; Ross, B.P. Multifunctional Analogs of Kynurenic Acid for the Treatment of Alzheimer's Disease: Synthesis, Pharmacology, and Molecular Modeling Studies. ACS Chem Neurosci 2017, 8, 2667–2675. [Google Scholar] [CrossRef]
- Yoshida, Y.; Fujigaki, H.; Kato, K.; Yamazaki, K.; Fujigaki, S.; Kunisawa, K.; Yamamoto, Y.; Mouri, A.; Oda, A.; Nabeshima, T.; et al. Selective and competitive inhibition of kynurenine aminotransferase 2 by glycyrrhizic acid and its analogues. Sci Rep 2019, 9, 10243. [Google Scholar] [CrossRef]
- Agrafiotis, D.K.; Shemanarev, M.; Connolly, P.J.; Farnum, M.; Lobanov, V.S. SAR maps: a new SAR visualization technique for medicinal chemists. Journal of Medicinal Chemistry 2007, 50, 5926–5937. [Google Scholar] [CrossRef] [PubMed]
- Lipinski, C.A. Overview of hit to lead: The medicinal chemist's role from HTS retest to lead optimization hand off. Lead-seeking approaches 2010, 1-24.
- Segall, M.D.; Beresford, A.P.; Gola, J.M.; Hawksley, D.; Tarbit, M.H. Focus on success: using a probabilistic approach to achieve an optimal balance of compound properties in drug discovery. Expert Opinion on Drug Metabolism & Toxicology 2006, 2, 325–337. [Google Scholar]
- Ward, K.W. Optimizing pharmacokinetic properties and attaining candidate selection. Reducing Drug Attrition 2014, 73–95. [Google Scholar]
- Qin, D.; Lin, X.; Liu, Z.; Chen, Y.; Zhang, Z.; Wu, C.; Liu, L.; Pan, Y.; Laquerre, S.; Emery, J. Discovery of orally bioavailable ligand efficient quinazolindiones as potent and selective tankyrases inhibitors. ACS Medicinal Chemistry Letters 2021, 12, 1005–1010. [Google Scholar] [CrossRef] [PubMed]
- Morphy, R.; Kay, C.; Rankovic, Z. From magic bullets to designed multiple ligands. Drug discovery today 2004, 9, 641–651. [Google Scholar] [CrossRef]
- Mager, D.E. Quantitative structure–pharmacokinetic/pharmacodynamic relationships. Advanced drug delivery reviews 2006, 58, 1326–1356. [Google Scholar] [CrossRef] [PubMed]
- Guha, R. On exploring structure–activity relationships. In silico models for drug discovery 2013, 81–94. [Google Scholar]
- Yan, G.; Rose, J.; Ellison, C.; Mudd, A.M.; Zhang, X.; Wu, S. Refine and strengthen SAR-based read-across by considering bioactivation and modes of action. Chemical Research in Toxicology 2023, 36, 1532–1548. [Google Scholar] [CrossRef]
- Brogi, S.; Tabanelli, R.; Calderone, V. Combinatorial approaches for novel cardiovascular drug discovery: a review of the literature. Expert Opinion on Drug Discovery 2022, 17, 1111–1129. [Google Scholar] [CrossRef]
- Zhang, C.; Chen, Y.-Y.; Chen, S.; Wang, Y.; Yuan, Y.; Yang, X.; Hu, W.; Chen, B.; Qi, Z.; Huse, J.T. Characterization of an Enhancer RNA Signature Reveals Treatment Strategies for Improving Immunotherapy Efficacy in Cancer. Cancer Research 2025. [Google Scholar] [CrossRef]
- Günther, J.; Däbritz, J.; Wirthgen, E. Limitations and off-target effects of tryptophan-related IDO inhibitors in cancer treatment. Frontiers in immunology 2019, 10, 1801. [Google Scholar] [CrossRef] [PubMed]
- Qian, S.; Zhang, M.; Chen, Q.; He, Y.; Wang, W.; Wang, Z. IDO as a drug target for cancer immunotherapy: recent developments in IDO inhibitors discovery. RSC Advances 2016, 6, 7575–7581. [Google Scholar] [CrossRef]
- Tanaka, M.; Bohar, Z.; Vecsei, L. Are kynurenines accomplices or principal villains in dementia? Maintenance of kynurenine metabolism. Molecules 2020, 25, 564. [Google Scholar] [CrossRef]
- Papathanasiou, T.; Strathe, A.; Hooker, A.C.; Lund, T.M.; Overgaard, R.V. Feasibility of exposure-response analyses for clinical dose-ranging studies of drug combinations. The AAPS Journal 2018, 20, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, S.; Michor, F. Pharmacokinetics and drug interactions determine optimum combination strategies in computational models of cancer evolution. Cancer research 2017, 77, 3908–3921. [Google Scholar] [CrossRef] [PubMed]
- Nowak-Sliwinska, P.; Weiss, A.; Ding, X.; Dyson, P.J.; Van Den Bergh, H.; Griffioen, A.W.; Ho, C.-M. Optimization of drug combinations using Feedback System Control. Nature protocols 2016, 11, 302–315. [Google Scholar] [CrossRef]
- Tanaka, M.; Tóth, F.; Polyák, H.; Szabó, Á.; Mándi, Y.; Vécsei, L. Immune Influencers in Action: Metabolites and Enzymes of the Tryptophan-Kynurenine Metabolic Pathway. Biomedicines 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Perez-Castro, L.; Garcia, R.; Venkateswaran, N.; Barnes, S.; Conacci-Sorrell, M. Tryptophan and its metabolites in normal physiology and cancer etiology. Febs j 2023, 290, 7–27. [Google Scholar] [CrossRef]
- Zuccarello, E.; Zhang, H.; Acquarone, E.; Pham, D.; Staniszewski, A.; Deng, S.X.; Landry, D.W.; Arancio, O.; Fiorito, J. Optimizing metabolic stability of phosphodiesterase 5 inhibitors: Discovery of a potent N-(pyridin-3-ylmethyl)quinoline derivative targeting synaptic plasticity. Bioorg Med Chem Lett 2023, 92, 129409. [Google Scholar] [CrossRef]
- de Lima, E.; Tanaka, M.; Lamas, C.; Quesada, K.; Detregiachi, C.; Araújo, A.; Guiguer, E.; Catharin, V.; de Castro, M.; Junior, E. Vascular Impairment, Muscle Atrophy, and Cognitive Decline: Critical Age-Related Conditions. Biomedicines. 2024; 12: 2096.
- Tanaka, M.; Battaglia, S.; Giménez-Llort, L.; Chen, C.; Hepsomali, P.; Avenanti, A.; Vécsei, L. Innovation at the Intersection: Emerging Translational Research in Neurology and Psychiatry. Cells 2024, 13. [Google Scholar] [CrossRef]
- Jena, S.; Gonzalez, G.; Vítek, D.; Kvasnicová, M.; Štěpánková, Š.; Strnad, M.; Voller, J.; Chanda, K. Novel neuroprotective 5,6-dihydropyrido[2',1':2,3]imidazo[4,5-c]quinoline derivatives acting through cholinesterase inhibition and CB2 signaling modulation. Eur J Med Chem 2024, 276, 116592. [Google Scholar] [CrossRef] [PubMed]
- Ostapiuk, A.; Urbanska, E.M. Kynurenic acid in neurodegenerative disorders-unique neuroprotection or double-edged sword? CNS Neurosci Ther 2022, 28, 19–35. [Google Scholar] [CrossRef]
- Shen, H.; Xu, X.; Bai, Y.; Wang, X.; Wu, Y.; Zhong, J.; Wu, Q.; Luo, Y.; Shang, T.; Shen, R.; et al. Therapeutic potential of targeting kynurenine pathway in neurodegenerative diseases. Eur J Med Chem 2023, 251, 115258. [Google Scholar] [CrossRef] [PubMed]
- Pavlidis, N.; Kofinas, A.; Papanikolaou, M.G.; Miras, H.N.; Drouza, C.; Kalampounias, A.G.; Kabanos, T.A.; Konstandi, M.; Leondaritis, G. Synthesis, characterization and pharmacological evaluation of quinoline derivatives and their complexes with copper(ΙΙ) in in vitro cell models of Alzheimer's disease. J Inorg Biochem 2021, 217, 111393. [Google Scholar] [CrossRef] [PubMed]
- Barbalho, S.M.; Laurindo, L.F.; de Oliveira Zanuso, B.; da Silva, R.M.S.; Gallerani Caglioni, L.; Nunes Junqueira de Moraes, V.B.F.; Fornari Laurindo, L.; Dogani Rodrigues, V.; da Silva Camarinha Oliveira, J.; Beluce, M.E. AdipoRon’s Impact on Alzheimer’s Disease—A Systematic Review and Meta-Analysis. International Journal of Molecular Sciences 2025, 26, 484. [Google Scholar] [CrossRef] [PubMed]
- Adlard, P.A.; Cherny, R.A.; Finkelstein, D.I.; Gautier, E.; Robb, E.; Cortes, M.; Volitakis, I.; Liu, X.; Smith, J.P.; Perez, K.; et al. Rapid restoration of cognition in Alzheimer's transgenic mice with 8-hydroxy quinoline analogs is associated with decreased interstitial Abeta. Neuron 2008, 59, 43–55. [Google Scholar] [CrossRef]
- Huang, W.; Liang, M.; Li, Q.; Zheng, X.; Zhang, C.; Wang, Q.; Tang, L.; Zhang, Z.; Wang, B.; Shen, Z. Development of the "hidden" multifunctional agents for Alzheimer's disease. Eur J Med Chem 2019, 177, 247–258. [Google Scholar] [CrossRef]
- Tanaka, M.; Szabó, Á.; Körtési, T.; Szok, D.; Tajti, J.; Vécsei, L. From CGRP to PACAP, VIP, and Beyond: Unraveling the Next Chapters in Migraine Treatment. Cells 2023, 12. [Google Scholar] [CrossRef]
- Sarkar, S. Regulation of autophagy by mTOR-dependent and mTOR-independent pathways: autophagy dysfunction in neurodegenerative diseases and therapeutic application of autophagy enhancers. Biochem Soc Trans 2013, 41, 1103–1130. [Google Scholar] [CrossRef]
- Yin, S.; Liu, L.; Gan, W. The Roles of Post-Translational Modifications on mTOR Signaling. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef]
- Filomeni, G.; De Zio, D.; Cecconi, F. Oxidative stress and autophagy: the clash between damage and metabolic needs. Cell Death Differ 2015, 22, 377–388. [Google Scholar] [CrossRef] [PubMed]
- Correia, A.S.; Vale, N. Tryptophan Metabolism in Depression: A Narrative Review with a Focus on Serotonin and Kynurenine Pathways. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef] [PubMed]
- Pathak, S.; Nadar, R.; Kim, S.; Liu, K.; Govindarajulu, M.; Cook, P.; Watts Alexander, C.S.; Dhanasekaran, M.; Moore, T. The Influence of Kynurenine Metabolites on Neurodegenerative Pathologies. Int J Mol Sci 2024, 25. [Google Scholar] [CrossRef] [PubMed]
- Schwarcz, R.; Stone, T.W. The kynurenine pathway and the brain: Challenges, controversies and promises. Neuropharmacology 2017, 112, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Sas, K.; Robotka, H.; Toldi, J.; Vécsei, L. Mitochondria, metabolic disturbances, oxidative stress and the kynurenine system, with focus on neurodegenerative disorders. J Neurol Sci 2007, 257, 221–239. [Google Scholar] [CrossRef]
- Tanaka, M.; Vécsei, L. Revolutionizing our understanding of Parkinson’s disease: Dr. Heinz Reichmann’s pioneering research and future research direction. Journal of Neural Transmission 2024, 1-21.
- Battaglia, S.; Avenanti, A.; Vécsei, L.; Tanaka, M. Neurodegeneration in Cognitive Impairment and Mood Disorders for Experimental, Clinical and Translational Neuropsychiatry. Biomedicines 2024, 12. [Google Scholar] [CrossRef] [PubMed]
- Battaglia, S.; Avenanti, A.; Vécsei, L.; Tanaka, M. Neural Correlates and Molecular Mechanisms of Memory and Learning. Int J Mol Sci 2024, 25. [Google Scholar] [CrossRef] [PubMed]
- Jászberényi, M.; Thurzó, B.; Bagosi, Z.; Vécsei, L.; Tanaka, M. The Orexin/Hypocretin System, the Peptidergic Regulator of Vigilance, Orchestrates Adaptation to Stress. Biomedicines 2024, 12. [Google Scholar] [CrossRef]
- Manetti, F.; Maresca, L.; Crivaro, E.; Pepe, S.; Cini, E.; Singh, S.; Governa, P.; Maramai, S.; Giannini, G.; Stecca, B.; et al. Quinolines and Oxazino-quinoline Derivatives as Small Molecule GLI1 Inhibitors Identified by Virtual Screening. ACS Med Chem Lett 2022, 13, 1329–1336. [Google Scholar] [CrossRef]
- Szabó, Á.; Galla, Z.; Spekker, E.; Szűcs, M.; Martos, D.; Takeda, K.; Ozaki, K.; Inoue, H.; Yamamoto, S.; Toldi, J.; et al. Oxidative and Excitatory Neurotoxic Stresses in CRISPR/Cas9-Induced Kynurenine Aminotransferase Knockout Mice: A Novel Model for Despair-Based Depression and Post-Traumatic Stress Disorder. Front Biosci (Landmark Ed) 2025, 30, 25706. [Google Scholar] [CrossRef]
- Wang, S.W.; Gao, C.; Zheng, Y.M.; Yi, L.; Lu, J.C.; Huang, X.Y.; Cai, J.B.; Zhang, P.F.; Cui, Y.H.; Ke, A.W. Current applications and future perspective of CRISPR/Cas9 gene editing in cancer. Mol Cancer 2022, 21, 57. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Ye, F.; Wang, Y.; Sun, X.; Chen, H.; Chen, T.; Gao, Y.; Chen, H. Synthesis, structure-activity relationship, and biological evaluation of quinolines for development of anticancer agents. Arch Pharm (Weinheim) 2023, 356, e2200673. [Google Scholar] [CrossRef]
- Behan, F.M.; Iorio, F.; Picco, G.; Gonçalves, E.; Beaver, C.M.; Migliardi, G.; Santos, R.; Rao, Y.; Sassi, F.; Pinnelli, M.; et al. Prioritization of cancer therapeutic targets using CRISPR-Cas9 screens. Nature 2019, 568, 511–516. [Google Scholar] [CrossRef] [PubMed]
- Han, K.; Jeng, E.E.; Hess, G.T.; Morgens, D.W.; Li, A.; Bassik, M.C. Synergistic drug combinations for cancer identified in a CRISPR screen for pairwise genetic interactions. Nat Biotechnol 2017, 35, 463–474. [Google Scholar] [CrossRef] [PubMed]
- Ngai, C.; Sevian, H. Probing the Relevance of Chemical Identity Thinking in Biochemical Contexts. CBE Life Sci Educ 2018, 17, ar58. [Google Scholar] [CrossRef] [PubMed]
- Günther, D.; Boto, R.A.; Contreras-Garcia, J.; Piquemal, J.P.; Tierny, J. Characterizing Molecular Interactions in Chemical Systems. IEEE Trans Vis Comput Graph 2014, 20, 2476–2485. [Google Scholar] [CrossRef]
- Gallegos, L.C.; Luchini, G.; St John, P.C.; Kim, S.; Paton, R.S. Importance of Engineered and Learned Molecular Representations in Predicting Organic Reactivity, Selectivity, and Chemical Properties. Acc Chem Res 2021, 54, 827–836. [Google Scholar] [CrossRef]
- Guan, K.; Wang, H.; Zhuang, L.; Sun, Y. Multi-granularity physicochemical-inspired molecular representation learning for property prediction. Expert Systems with Applications 2025, 266, 126115. [Google Scholar] [CrossRef]
- Weyesa, A.; Mulugeta, E. Recent advances in the synthesis of biologically and pharmaceutically active quinoline and its analogues: a review. RSC Adv 2020, 10, 20784–20793. [Google Scholar] [CrossRef]
- Palani, V.; Wendlandt, A.E. Strain-Inducing Positional Alkene Isomerization. J Am Chem Soc 2023, 145, 20053–20061. [Google Scholar] [CrossRef]
- Ishida, K.; Higashino, T.; Wada, Y.; Kaji, H.; Saeki, A.; Imahori, H. Thiophene-Fused Naphthodiphospholes: Modulation of the Structural and Electronic Properties of Polycyclic Aromatics by Precise Fusion of Heteroles. Chempluschem 2021, 86, 130–136. [Google Scholar] [CrossRef] [PubMed]
- Stone, T.W.; Williams, R.O. Modulation of T cells by tryptophan metabolites in the kynurenine pathway. Trends Pharmacol Sci 2023, 44, 442–456. [Google Scholar] [CrossRef] [PubMed]
- Mithaiwala, M.N.; Santana-Coelho, D.; Porter, G.A.; O'Connor, J.C. Neuroinflammation and the Kynurenine Pathway in CNS Disease: Molecular Mechanisms and Therapeutic Implications. Cells 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Erhardt, S.; Schwieler, L.; Imbeault, S.; Engberg, G. The kynurenine pathway in schizophrenia and bipolar disorder. Neuropharmacology 2017, 112, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Zádori, D.; Veres, G.; Szalárdy, L.; Klivényi, P.; Vécsei, L. Alzheimer's Disease: Recent Concepts on the Relation of Mitochondrial Disturbances, Excitotoxicity, Neuroinflammation, and Kynurenines. J Alzheimers Dis 2018, 62, 523–547. [Google Scholar] [CrossRef]
- Sutherland, J.J.; Yonchev, D.; Fekete, A.; Urban, L. A preclinical secondary pharmacology resource illuminates target-adverse drug reaction associations of marketed drugs. Nat Commun 2023, 14, 4323. [Google Scholar] [CrossRef] [PubMed]
- Casas, A.I.; Hassan, A.A.; Larsen, S.J.; Gomez-Rangel, V.; Elbatreek, M.; Kleikers, P.W.M.; Guney, E.; Egea, J.; López, M.G.; Baumbach, J.; et al. From single drug targets to synergistic network pharmacology in ischemic stroke. Proc Natl Acad Sci U S A 2019, 116, 7129–7136. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhang, Y.; Lian, X.; Li, F.; Wang, C.; Zhu, F.; Qiu, Y.; Chen, Y. Therapeutic target database update 2022: facilitating drug discovery with enriched comparative data of targeted agents. Nucleic Acids Res 2022, 50, D1398–d1407. [Google Scholar] [CrossRef]
- Ruiz, M.C.; Resasco, A.; Di Virgilio, A.L.; Ayala, M.; Cavaco, I.; Cabrera, S.; Aleman, J.; León, I.E. In vitro and in vivo anticancer effects of two quinoline-platinum(II) complexes on human osteosarcoma models. Cancer Chemother Pharmacol 2019, 83, 681–692. [Google Scholar] [CrossRef]
- Saxena, A.; Majee, S.; Ray, D.; Saha, B. Inhibition of cancer cells by Quinoline-Based compounds: A review with mechanistic insights. Bioorg Med Chem 2024, 103, 117681. [Google Scholar] [CrossRef]
- Pich, O.; Bailey, C.; Watkins, T.B.K.; Zaccaria, S.; Jamal-Hanjani, M.; Swanton, C. The translational challenges of precision oncology. Cancer Cell 2022, 40, 458–478. [Google Scholar] [CrossRef]
- Zugazagoitia, J.; Guedes, C.; Ponce, S.; Ferrer, I.; Molina-Pinelo, S.; Paz-Ares, L. Current Challenges in Cancer Treatment. Clin Ther 2016, 38, 1551–1566. [Google Scholar] [CrossRef] [PubMed]
- Malone, E.R.; Oliva, M.; Sabatini, P.J.B.; Stockley, T.L.; Siu, L.L. Molecular profiling for precision cancer therapies. Genome Med 2020, 12, 8. [Google Scholar] [CrossRef] [PubMed]
- Ioannidis, J.P.A.; Kim, B.Y.S.; Trounson, A. How to design preclinical studies in nanomedicine and cell therapy to maximize the prospects of clinical translation. Nat Biomed Eng 2018, 2, 797–809. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Sahakian, D.C.; de Morais, S.M.; Xu, J.J.; Polzer, R.J.; Winter, S.M. The role of absorption, distribution, metabolism, excretion and toxicity in drug discovery. Curr Top Med Chem 2003, 3, 1125–1154. [Google Scholar] [CrossRef]
- Liu, J.; Gui, Y.; Rao, J.; Sun, J.; Wang, G.; Ren, Q.; Qu, N.; Niu, B.; Chen, Z.; Sheng, X.; et al. In silico off-target profiling for enhanced drug safety assessment. Acta Pharm Sin B 2024, 14, 2927–2941. [Google Scholar] [CrossRef] [PubMed]
- David, P.O. Abstract A001: Navigating the future of therapeutic development: innovations in safety and efficacy through advanced research methodologies and nanotechnology. Molecular Cancer Therapeutics 2024, 23, A001–A001. [Google Scholar] [CrossRef]
- Son, A.; Park, J.; Kim, W.; Yoon, Y.; Lee, S.; Ji, J.; Kim, H. Recent Advances in Omics, Computational Models, and Advanced Screening Methods for Drug Safety and Efficacy. Toxics 2024, 12. [Google Scholar] [CrossRef]
- Mor, A.; Tankiewicz-Kwedlo, A.; Ciwun, M.; Lewkowicz, J.; Pawlak, D. Kynurenines as a Novel Target for the Treatment of Inflammatory Disorders. Cells 2024, 13. [Google Scholar] [CrossRef]
- Tanaka, M. Beyond the boundaries: Transitioning from categorical to dimensional paradigms in mental health diagnostics. Advances in Clinical and Experimental Medicine 2024, 33, 1295–1301. [Google Scholar] [CrossRef]
- Tanaka, M.; Vécsei, L. A decade of dedication: pioneering perspectives on neurological diseases and mental illnesses. 2024, 12, 1083.
- Ryan, K.M.; Corrigan, M.; Murphy, T.M.; McLoughlin, D.M.; Harkin, A. Gene expression of kynurenine pathway enzymes in depression and following electroconvulsive therapy. Acta Neuropsychiatr 2024, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Wang, X.; Wang, L.; Ma, X.; Gong, Z.; Zhang, S.; Li, Y. Targeting the IDO1 pathway in cancer: from bench to bedside. J Hematol Oncol 2018, 11, 100. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M. From Serendipity to Precision: Integrating AI, Multi-Omics, and Human-Specific Models for Personalized Neuropsychiatric Care. Biomedicines 2025, 13, 167. [Google Scholar] [CrossRef]
- Cadilla, R.; Deaton, D.N.; Do, Y.; Elkins, P.A.; Ennulat, D.; Guss, J.H.; Holt, J.; Jeune, M.R.; King, A.G.; Klapwijk, J.C.; et al. The exploration of aza-quinolines as hematopoietic prostaglandin D synthase (H-PGDS) inhibitors with low brain exposure. Bioorg Med Chem 2020, 28, 115791. [Google Scholar] [CrossRef]
- Boros, F.A.; Bohár, Z.; Vécsei, L. Genetic alterations affecting the genes encoding the enzymes of the kynurenine pathway and their association with human diseases. Mutat Res Rev Mutat Res 2018, 776, 32–45. [Google Scholar] [CrossRef] [PubMed]
- Tamimou, R.; Montout, C.; Mura, T.; Conejero, I.; Evrard, A.; Courtet, P.; Bonilla-Escribano, P.; Riaza, C.; Vaquero-Lorenzo, C.; Baca-Garcia, E.; et al. Genetic association of the kynurenine pathway to suicidal behavior. Brain Behav Immun Health 2024, 42, 100903. [Google Scholar] [CrossRef]
- Nemkov, T.; Stephenson, D.; Erickson, C.; Dzieciatkowska, M.; Key, A.; Moore, A.; Earley, E.J.; Page, G.P.; Lacroix, I.S.; Stone, M.; et al. Regulation of kynurenine metabolism by blood donor genetics and biology impacts red cell hemolysis in vitro and in vivo. Blood 2024, 143, 456–472. [Google Scholar] [CrossRef] [PubMed]
- Martin, J.H.; Galettis, P.; Flynn, A.; Schneider, J. Phenotype versus genotype to optimize cancer dosing in the clinical setting-focus on 5-fluorouracil and tyrosine kinase inhibitors. Pharmacol Res Perspect 2024, 12, e1182. [Google Scholar] [CrossRef]
- Swenberg, J.A.; Fryar-Tita, E.; Jeong, Y.C.; Boysen, G.; Starr, T.; Walker, V.E.; Albertini, R.J. Biomarkers in toxicology and risk assessment: informing critical dose-response relationships. Chem Res Toxicol 2008, 21, 253–265. [Google Scholar] [CrossRef]
- Cores, Á.; Carmona-Zafra, N.; Clerigué, J.; Villacampa, M.; Menéndez, J.C. Quinones as Neuroprotective Agents. Antioxidants (Basel) 2023, 12. [Google Scholar] [CrossRef]
- Wang, Z.; Hu, J.; Yang, X.; Feng, X.; Li, X.; Huang, L.; Chan, A.S.C. Design, Synthesis, and Evaluation of Orally Bioavailable Quinoline-Indole Derivatives as Innovative Multitarget-Directed Ligands: Promotion of Cell Proliferation in the Adult Murine Hippocampus for the Treatment of Alzheimer's Disease. J Med Chem 2018, 61, 1871–1894. [Google Scholar] [CrossRef] [PubMed]
- Pietta, P.G. Flavonoids as antioxidants. J Nat Prod 2000, 63, 1035–1042. [Google Scholar] [CrossRef] [PubMed]
- Nunes, Y.C.; Mendes, N.M.; Pereira de Lima, E.; Chehadi, A.C.; Lamas, C.B.; Haber, J.F.; dos Santos Bueno, M.; Araújo, A.C.; Catharin, V.C.S.; Detregiachi, C.R.P. Curcumin: A golden approach to healthy aging: A systematic review of the evidence. Nutrients 2024, 16, 2721. [Google Scholar] [CrossRef] [PubMed]
- Galleano, M.; Verstraeten, S.V.; Oteiza, P.I.; Fraga, C.G. Antioxidant actions of flavonoids: thermodynamic and kinetic analysis. Arch Biochem Biophys 2010, 501, 23–30. [Google Scholar] [CrossRef]
- Venkanna, A.; Cho, K.H.; Dhorma, L.P.; Kumar, D.N.; Hah, J.M.; Park, H.G.; Kim, S.Y.; Kim, M.H. Chemistry-oriented synthesis (ChOS) and target deconvolution on neuroprotective effect of a novel scaffold, oxaza spiroquinone. Eur J Med Chem 2019, 163, 453–480. [Google Scholar] [CrossRef] [PubMed]
- Karale, U.B.; Shinde, A.; Gaikwad, V.R.; Kalari, S.; Gourishetti, K.; Radhakrishnan, M.; Poornachandra, Y.; Amanchy, R.; Chakravarty, S.; Andugulapati, S.B.; et al. Iron mediated reductive cyclization/oxidation for the generation of chemically diverse scaffolds: An approach in drug discovery. Bioorg Chem 2023, 139, 106698. [Google Scholar] [CrossRef]
- Kim, S.; Kim, D.; Hong, S.Y.; Chang, S. Tuning Orbital Symmetry of Iridium Nitrenoid Enables Catalytic Diastereo- and Enantioselective Alkene Difunctionalizations. J Am Chem Soc 2021, 143, 3993–4004. [Google Scholar] [CrossRef]
- Cubillos-Ruiz, A.; Guo, T.; Sokolovska, A.; Miller, P.F.; Collins, J.J.; Lu, T.K.; Lora, J.M. Engineering living therapeutics with synthetic biology. Nat Rev Drug Discov 2021, 20, 941–960. [Google Scholar] [CrossRef]
- Liu, Z.; Cao, S.; Liu, M.; Kang, W.; Xia, J. Self-Assembled Multienzyme Nanostructures on Synthetic Protein Scaffolds. ACS Nano 2019, 13, 11343–11352. [Google Scholar] [CrossRef]
- Siefen, T.; Lokhnauth, J.; Liang, A.; Larsen, C.C.; Lamprecht, A. An ex-vivo model for transsynovial drug permeation of intraarticular injectables in naive and arthritic synovium. J Control Release 2021, 332, 581–591. [Google Scholar] [CrossRef]
- Lavallie, E.R.; Dorner, A.J.; Burczynski, M.E. Use of ex vivo systems for biomarker discovery. Curr Opin Pharmacol 2008, 8, 647–653. [Google Scholar] [CrossRef] [PubMed]
- Stefaniak, J.; Huber, K.V.M. Importance of Quantifying Drug-Target Engagement in Cells. ACS Med Chem Lett 2020, 11, 403–406. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.; Kwon, S.H.; Zhou, X.; Fuller, C.; Wang, X.; Vadgama, J.; Wu, Y. Overcoming Challenges in Small-Molecule Drug Bioavailability: A Review of Key Factors and Approaches. Int J Mol Sci 2024, 25. [Google Scholar] [CrossRef] [PubMed]
- Stefanov, B.A.; Fussenegger, M. Biomarker-driven feedback control of synthetic biology systems for next-generation personalized medicine. Front Bioeng Biotechnol 2022, 10, 986210. [Google Scholar] [CrossRef] [PubMed]
- Hassanzadeh, P.; Atyabi, F.; Dinarvand, R. Technical and engineering considerations for designing therapeutics and delivery systems. J Control Release 2023, 353, 411–422. [Google Scholar] [CrossRef]
Table 1.
Biochemical properties of key kynurenine (KYN) pathway metabolites and nicotinic acid (NA). This table presents the primary physiological roles and biochemical features of these compounds, underscoring their relevance to neurotransmission, immunological regulation, and metabolic pathways.
Table 1.
Biochemical properties of key kynurenine (KYN) pathway metabolites and nicotinic acid (NA). This table presents the primary physiological roles and biochemical features of these compounds, underscoring their relevance to neurotransmission, immunological regulation, and metabolic pathways.
| Compounds | Main characteristics | Ref. |
|---|---|---|
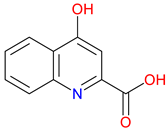 Kynurenic Acid |
Neuroprotective – Blocks NMDA and α7nAChR to reduce excitotoxicity KYN pathway metabolite – Modulates neurotransmission and immune response Poor BBB permeability – Limited CNS access, driving prodrug development |
[30,44,45] [46,47,48] [49,50,51] |
 Nicotinic Acid (NA) |
Essential vitamin (B3) – Crucial for NAD+/NADP+ synthesis, supporting cellular metabolism Lipid-lowering effects – Reduces LDL and triglycerides while increasing HDL cholesterol Modulation of Nicotinic Acid-Induced Flushing – GPR109A and Langerhans cells drive flushing; DP1 antagonists suppress it |
[52,53,54] [55,56,57] [58,59,60] |
 Quinolinic Acid (QUIN) |
Excitotoxic agent: Acts as a potent NMDA receptor agonist, leading to neuronal overexcitation and potential cell death Oxidative stress contributor: Promotes free radical generation, exacerbating cellular damage Neurodegenerative implications: Linked to the progression of neurological disorders through its detrimental effects on brain tissue |
[61,62,63] [64,65,66] [62,67,68] |
 Picolinic Acid |
Metal ion chelator – Binds zinc, iron, and other metals, influencing cellular metabolism Immune Modulator – Enhances macrophage activity and antimicrobial defense Neuroactive Compound – Involved in neurotransmission and potential neuroprotective effects |
[69,70,71] [72,73,74] [75,76,77] |
 Xanthurenic Acid (XA) |
Neuromodulatory effects: Influences neuronal signaling and may modulate receptor activity Putative neuroactive role – Modulates glutamate signaling and may influence neurotransmission Metal chelating properties – Binds with zinc and other metal ions, potentially affecting oxidative stress |
[43,78,79] [43,78,79] [80,81,82] |
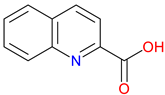 Quinaldic Acid |
Quinoline derivative – Structurally related to other KYN pathway metabolites Antimicrobial properties – Exhibits activity against certain bacteria and fungi Potential neuroactive role – May influence neurotransmission, though its biological significance remains underexplored |
[83,84,85] [86,87] [85,88] |
α7nAChR, α7 nicotinic acetylcholine receptor; BBB, blood-brain barrier; CNS, central nervous system; DP1, prostaglandin D₂ receptor 1; GPR109A, G protein-coupled receptor 109A; HDL, high-density lipoprotein; LDL, low-density lipoprotein; NAD+/NADP+, reduced nicotinamide adenine dinucleotide/reduced nicotinamide adenine dinucleotide phosphate; NMDA, N-methyl-D-aspartate.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



