
Submitted:
29 March 2025
Posted:
31 March 2025
You are already at the latest version
Abstract
The biotechnological applications of chitinases are diverse, they are used in industrial sectors such as pharmaceutical and textile, as well as in agriculture, including the use of recombinant chitinases for pest control, since traditional treatments affect and contaminate hive products. Bacillus licheniformis UV01 is an bacterium of interest that expresses genes for different enzymes, including chitinase. The Chibluv01 gene was cloned into the pHTP8 vector with a His/tag, for purification by affinity chromatography. Showing a specific activity of 115 U/mL, the optimal pH of the enzyme was pH 7.5, and an optimal temperature of 42 ºC. The choleoptericidal activity of the enzyme was evaluated in larvae and adult beetles of Aethina tumida treated by immersion in a purified enzyme extract at different concentrations, verifying mortality at 24, 48 and 72 hours. Within 24 hours of application, there was a higher mortality of 50 % in the larval stage and 56.6 % in adult beetles compared to the control groups. LC50 and LC90 were obtained: 104.05 U/mL and 234.36 U/mL in larvae and 92.99 U/mL and 211.14 U/mL in adults. This results indicate the potential of the application od ChiBlUV02 chitinase in pest control.
Keywords:
Bacillus licheniformis
; Biocontrol
; ChiBlUV02 chitinase
; Aethina tumida
1. Introduction
The chitinases (E.C. 3.2.2.14) are a group of enzymes that have a wide range of applications due to their ability to degrade chitin obtaining biotechnologically important polymers such as chitosan. Chitinases catalyze the cleavage of β(1-4)-linked in native chitin [1], releasing N-acetyl-β-D-glucosamine units in the process. Likewise, chitinases play an important role in several organisms such as the wall of fungi and insects primarily at the remodeling of the exoskeletons and the acquisition of carbon sources, as well as the collection of energy for microorganisms that feed by means of the hydrolysis of this polymer [2].
The research, development and optimization of these enzymes represents a significant advance in the area of biotechnology, the obtained parameters such as pH or working temperature are fundamental data that when obtained can provide advantages for their application. Such as the management and control of some pests in the agricultural area, for example: some phytopathogenic fungi such as Botrytis cinerea, or insect pests such as Helicoverpa armigera that cause millionaire losses yearly in this sector. In addition, chitinases can be used to manufacture biofuels using chitin as raw material, and to produce fermentable sugars on the conversion of waste from crustaceans [3,4,5,6].
Obtaining chitinases with industrial value has generated benefits in different sectors. For example, in the food industry are used for chitosan production. Chitosan is a polymer derived from chitin that is used as an antimicrobial agent and as food preservative [3]. There are also reports of effects on insects of the orders Lepidoptera and Diptera [7].
Chitinases of bacterial origin can act as endochitinases and exochitinases depending on the site in the chitin chain where they act, they also present a broad range of molecular weight, from 20 to 80 kDa. Among the bacterial genera that produce chitinases, the Bacillus is one of the most interesting, species such as Bacillus thuringiensis possess a gen named bt15a3 that encodes for the protein called Chia1 which possesses chitinolytic activity inhibiting significantly the spore germination of four species of fungi. [8]
Bacillus cereus strain NK91 produces an extracellular chitinase, the enzyme was tested against three phytopathogenic fungi where it exhibited higher antifungal activity towards Fusarium oxysporum (66.7%), Rhizoctonia solani (64.6%) and Colletotrichum gloeosporioides (63%) [9]. Native chitinase from Bacillus licheniformis NK-7 was tested as an antifungal agent against phytopathogenic fungi and for the isolation and formation of fungal protoplasts of Aspergillus niger showing inhibition of radial growth in relation to the control of 69.44 % [10].
Bacillus licheniformis is a Gram-positive, facultative anaerobic bacterium that requires a simple medium for growth. It is used in industry to produce endogenous and exogenous enzymes due to its thermophilic bacterial characteristics and great variety of genes that express enzymes of different characteristics such as proteases, amylases, lipases, and chitinases [11,12,13]. Slimene et al. reports that Bacillus licheniformis S213 produces a chitinase with an effect phytopathogenic on some fungi as Phoma medicaginis [14]. Kwon et al. indicates that Bacillus licheniformis PR2 exhibits antifungal activity against certain fungi vegetable [15]. Bacillus licheniformis UV01 isolated in thermal waters of San José Purua Michoacán has a gene called Chibluv01 that codes for the chitinase ChiBlUV02. This enzyme showed a choleoptericidal effect on larvae of the species Aethina tumida.
Aethina tumida or small hive beetle was discovered in 1867 [16], it is a scavenging and parasitic insect native to Africa and has become a pest that causes catastrophic damage to Apis mellifera hives outside its natural range [17]. The beetles infest hives where they feed, develop and oviposit. They feed primarily on bee brood (larvae and pupae), as well as honey, pollen and dead bees [18]. Precise figures on monetary losses are not available due to a lack of recent and specific data. However, reports from previous years give an idea of the magnitude of economic losses in the international beekeeping sector.
United States reported losses of $428 million in 2023 [19]. and the European Union reported losses of €1.4 billion due to pests and factors related to climate change [20].
This study focuses on the purification and characterization of chitinase ChiBlUV02, and the evaluation of the choleoptericidal effect against the Aethina tumida species, thus increasing its efficacy as a biological control agent. The results obtained for the recombinant chitinase ChiBlUV02 from Bacillus licheniformis UV01 showed that they can contribute to the development of strategies for biotechnologies for the control of agricultural pests and provide a solid foundation for the for future research in the field of chitinases and their application in different industrial sectors.
2. Materials and Methods
2.1. Reactivation and Growth of Bacillus licheniformis UV01
Bacillus licheniformis strain UV01 was reactivated from a preserved culture at -80 °C in 30 % glycerol which was inoculated in Luria Bertani medium (LB). The inoculate was grown at 50°C under orbital shaking at 100 rpm for 24 h.
2.2. Chitinase Gene Amplification
The reaction was carried out using polymerase chain reaction (PCR) with the chromosomal DNA of Bacillus licheniformis UV01 and the designed oligonucleotides previously: FChiBLNd01 sequence (5'->3'): GGAATTCCATATATGAAGAAAGCCGCTTCATC- and RChiBLBa02 sequence (5'->3'): -CGGGATCCAATTTCCTTTAAGCCTGTACTTT-. The reaction was carried out in a C1000™ Touch Thermal Cycler (Bio-Rad®). The PCR conditions comprised a single cycle of 1 minute at 94°C, 30 cycles of 1 minute at 94°C, 1 minute at 58°C, 1 minute at 72°C, and a final single cycle of 5 minutes at 72°C. The PCR products were stored at -20°C until further use.
2.3. Purification and Quantification of PCR Product
The purification of PCR products was accomplished through the utilization of the Wizard® SV Gel™ kit and the PCR Clean-Up System™ (Promega®), in accordance with the established protocols. The quantification of purity and concentration was using the NanoDrop® ND 2000 (Thermo Scientific™, Waltham, Massachusetts, USA), adhering to the established protocols. The concentration was determined based on the absorption ratio at 260 nanometers, while the purity was gauged based on the ratio at 260/280 nanometers.
2.4. Cloning of the Chitinase Gene Chibluv01
Cloning of the chitinase gene was performed in the pHTP8 His-tag NZYEasy vector Cloning & Expression kit®. From concentrations and purities estimated by Lehninger spectrometry [21]. With a purity ratio for the gene of 1.8 to 2.0, proceeded in a single ligase-independent reaction mediated by the NZYEasy, enzyme mixture according to the user's manual; the digested vector and insert were ligated in a 3:1 ratio and incubated at temperature cycles of 37°C/60 minutes, 80°C/10 minutes, and 30°C/10 minutes. The resulting ligation product was used as a template for transformation of competent E. coli BL 21 (DE3) cells and plated on LB agar plates containing kanamycin (50 µg/mL).
2.5. Recombinant Protein Expression
Expression of recombinant cells was performed in liquid culture media. LB-kanamycin (50 µg/mL). These previously activated cultures were incubated 8 hours at 37 °C under agitation at 150 rpm with 0.8 mM IPTG. At the end of incubation, the cell culture was centrifuged at 14,000 xg for 10 minutes at 4 °C, and the cell package was collected and resuspended in phosphate buffer (pH 6.0, 100mM), and subsequently sonicated in a sonicator Yucheng Tech® (20 kHz) for 20 cycles alternating 15 seconds and 30 seconds All steps of the protocol were carried out in cold. The cell lysate was centrifuged under the same conditions and the supernatant was recovered and stored under at 4 °C.
To determine the molecular weight of the enzyme SDS-PAGE was performed (12%) The bands of protein were stained with Coomassie Brilliant blue G 250 Sigma/Aldrich® according to the protocol described [22]. The banding pattern was observed using a white light transilluminator.
2.6. Preparation of Colloidal Chitin
Sigma/Aldrich brand colloidal chitin® was prepared through a modification of method of Hsu and Lockwood [23]. The paste obtained was sterilized for 15 min at 121 °C and 15 psi. It was stored at 4 °C until further use.
2.7. Quantification of Concentration Protein
The protein concentration of the crude and purified extracts was measured using the spectrophotometric method described by Bradford [24]. A calibration curve with bovine serum albumin (BSA) and phosphate buffer pH 7 100mM, with concentrations between 0.0 and 1.0 mg/mL, the absorbances were read in a Beckman Coulter TM spectrophotometer® at 595 nm.
2.8. Enzymatic Assay
For the determination of the enzymatic activity, 750 μL of 1% colloidal chitin were incubated with 750 µL of purified extract, the reaction was incubated at 42 °C for 1 hour. Then, the reaction mixture was centrifuged at 12,000 xg, for 10 min, and the release of N-acetyl-D-glucosamine was determined in 300 μL of supernatant by the DNS protocol (3,5-dinitrosalicylic acid) [25]. The absorbances were measured at 540 nm. A unit of chitinolytic activity was defined as the amount of enzyme that releases 1 µM of sugar per 1 minute at 42 °C (pH 6 100mM).
2.9. Purification by Affinity Chromatography
The purification was performants using Ni-NTA Agarose (QIAGEN®) Nickel-NTA. The column preparation was realized using the manufacturer's recommended protocol. The activated Ni-NTA Agarose was packed in 1 x 5 cm column and equilibrated with 3 volumes of lysis buffer solution (100mM/L NaH2PO4 10 mM Tris-Cl and 10 mM/L Imidazole. pH 8). The cells extract was mixed with lysis buffer and Ni-NTA agarose resin for 60 minutes at 200 rpm at 4 °C. The mixture was poured in the column and the filtrated was recovered for SDS-PAGE analysis.
Column washing was performed using 4 mL of the wash solution (100 mM/L NaH2PO4, 10 mM Tris-Cl, and 20 mM/L imidazole), and 1 mL of wash fractions were collected. The elution fractions were performed with elution buffer (100 mM/L NaH2PO4, 10 mM Tris-Cl, and 250 mM/L imidazole) up to 500 μL aliquots. This step was followed by polyacrylamide gel electrophoresis with 12% SDS, under dissociation conditions according to the technique described by Gallagher. [26].
2.10. Determination of Optimal Temperature and pH for Chitinolytic Activity
The determination of the temperature for the maximum chitinolytic activity was evaluated between 25 and 50 °C. The reactions were performed as previously described. To determine the maximum pH, the assay was performed at maximum activity temperature (42 °C) using acetate and phosphate buffer systems at pH 4.0 to 9.0, 100 mM.
2.11. Adaptation of Aethina tumida.
Adult Aethina tumida beetles were collected from apiaries in the Sotavento region of central Veracruz state. For the maintenance of the biological cycle of A. tumida were conditioned in rearing chambers at 28± 2 °C and 80 % relative humidity by Haas et al. [27]. Beetles were deposited in 60 ×15 mm Petri dishes (Scientific Senna®) with a central perforation of 2 cm in diameter, covered with nylon mesh to allow ventilation. Bettles were feeded with 500 mg of a mixture of honey, pollen, soybean and barley (1:1:1:1:1) every third day. After oviposition, the beetles were relocated in new conditioned Petri dishes, and the hatched larvae were feed with the same food mixture, humidity and temperature conditions until the start of their normal biological cycle. For this last phase, plastic pupating chambers of 1 liter were designed, ¾ parts of the chambers were filled with a mix of soils of sand, clay and silt. A Petri dish was adapted at the upper part of the chamber with a perforation at the base in oder to allow that the larvae had descend and initiate its pupation stage. For the choleoptericidal activity assays were used larvae and adult beetles of the third generation for 3 to 4 days after their hatching.
2.12. Choleoptericidal on Activity Aethina tumida
For the evaluation of the choleoptericidal activity of ChiBlUV02, five concentrations of extract were used (10, 30, 45, 70 and 115 U/mL) and a negative control without. Purified recombinant chitinase was administered by the immersion method, which consists of using 10 beetles or 10 larvae, depending on the stage evaluated, for 30 seconds for the tests. Acute effect conditions were determined at 24, 48, and 72 hours. All assays were carried out in triplicate. Any specimen that did not walk or did not respond to manipulation with mechanic manipulation was considered dead. In all assays, beetles and larvae treated were kept at temperature of 28± 2 °C in darkness. The response variable was the mortality percentage of beetles and larvae in 24, 48 and 72 hours after the assay conditions.
The damages in the exoskeleton of the beetles were observed with micrographs that were taken with an EBTOOL® digital stereoscopic microscope.
3. Results
3.1. Recovery of the Gene chiBLUV01
The PCR reaction was carried out using chromosomal DNA from Bacillus licheniformis UV01 as a template. The amplicon of approximately 2,000 bp was corroborated through a 1% agarose gel. (Figure 1). The sequence corresponds a chitinase gene previously reported by Berka et al. (2004) (GeneBank CP000002.3) (data not shown).
3.2. Identification Transformed Cells
The construction of the pHTP-Chibluv01 vector was verified through the amplification of the pHTP-chibluv01 by PCR of the chibluv01 gene using plasmid. DNA as a template the transformed strains were analyzed on 1% agarose gels Figure 2 shows amplification of the chibluv01 gene present in Escherichia coli BL21 cells (DE3) transformed.
3.3. Induction of chibluv01 Gene Expression
Gene expression was induced according to the methodology previously described. The cells were harvested at different times after the induction and were lysed. A prominent band will be observed on the polyacrylamide gel at approximately 80 kilodaltons (kDa), corresponding to the expected molecular weight of the Interest protein in the overexpressed sample. The highest expression was determined using polyacrylamide SDS-PAGE under dissociating conditions (Figure 3).
3.4. Determination pH and Temperature Maximum
The optimal enzyme temperature was 42 °C and the optimal pH of activity was 7.5. The changes in incubation temperature within the range studied (20 - 80 °C) showed relevance to the performance of recombinant chitinase. Produced by B. licheniformis UV01. It was found that the maximum production was 115 U/mL between 40 and 42 °C (Figure 4).
The effect of different pH on chitinase ChiBlUV02 activity performance was measured from pH 4.0 up to pH 9.0 (100mM). The maximum value was found at pH 7.5 with an activity of 115 U\mL (defined as 100% relative activity), while the minimum activity was 3 U\mL at pH 4 (3.5 % relative activity) as shown in Figure 5.
3.5. ChiBlUV02 Chitinase Purification
The extracts obtained by treatment with lysis buffer (100 mM/L NaH2PO4 10 mM Tris-Cl and 10 mM/L Imidazole), were clarified by centrifugation and adjusted to at pH 8.0 and loaded on to a Kimble Flex-Column®. The matrix used was Ni-NTA agarose from Qiagen™. The column matrix was washed with the respective buffer (100 mM/L NaH2PO4 10 mM/L Tris-Cl and 20 mM/L Imidazole), the retained enzyme on the matrix was recovered using the elution buffer (100 mM/L NaH2PO4 10 mM Tris-Cl and 250 mM/L Imidazole). The data from the purification showed that the fraction corresponding to ChiBlUV02 from the protein (Figure 6 has an increase of 90% with respect to the crude extract and 23% of yield.
3.6. Choleoptericidal Activity on Larvae and Adults of Aethina tumida
Table 1 shows the mortality of larvae treated with purified enzyme by immersion for 30 Seconds. The maximum mortality was observed at 24 hours post-treatment, which might suggest that the enzyme loses its choleoptericidal effect after that time following its administration.
Table 1 Choleoptericidal activity of purified extract of the enzyme ChiBlUV02 on larvae of Aethina tumida.
The Table 2 Shows the results of adult beetle mortality in the 30 seconds immersion assays. See observed that the enzyme, as with larvae of Aethina tumida, manifested its maximum efficiency 24 hours post treatment.
Table 2 Choleoptericidal activity of purified extract of the enzyme ChiBlUV02 on adult of Aethina tumida.
The calculations of the LC50 and LC90 for this method of application are shown in the Figure 7 and Figure 8. The tests of mortality on larvae of A. tumida showed differences statistically significant (P≤ 0.05) with the different activity units evaluated.
Mortality rate increased as enzymatic activity increased obtaining the best result when using purified chitinase at 115 U/mL which promoted 50% mortality of larvae at 24 h (Table 2) and 56% of mortality in adult beetles of Aethina tumida Table 2). The units of activity evaluated in the present study did not allow us to obtain 100% of the mortality of larvae and adult beetles but the LC50 and LC90 could be calculated to determine the enzyme activity necessary to promote the mortality at 50% and 90% of the population, respectively.
Probit analysis was performed on data obtained and the units were calculated of chitinolytic activity required to promote 50 (LC50) and 90 mortality % (LC 90) of the population of adult beetles which were 97.58 U/mL and 222.94 U/mL for beetle adult and in larvae 104.05 U/mL and 234.36 U/mL, (Figure 7 and 8).
The choleoptericidal effect of the ChiBlUV02 enzyme was also observed in the architecture of exoskeleton of larvae and adults of Aethina tumida. A change in the structure of the exoskeleton was observed in the larvae (Figure 9).
Digital microscopic observation revealed changes at the structural level that could the results are attributable to the action of the chitinase contained in the purified. Extract At the examined area was in detail it was observed that the cause of the wear of the exoskeleton in the adult beetle may be associated with the hydrolysis of chitin present in the in the beetle (Figure 9).
The egg-laying rate was also impacted, with a 15-day prolongation relative to the control group, possibly attributable to the purified enzyme (Figure 10). Aethina tumida eggs post enzymatic treatment of adult beetles The Figure 10 a) shows how the larvae are damaged, interrupting their life. Cycle (a) change was observed in the coloration of the eggshell becoming darker (b) Furthermore, the cuticle was visually perceived as thinner, suggesting a possible alteration of the in its structure or composition compared to the control (c and d).
4. Discussion
Bacillus licheniformis is a bacterium that produces an important variety of enzymes including chitinases given its saprophytic nature [28], In the present study, the recombinant chitinase named ChiBlUV02 presented a maximum activity of 115 U/mL with a temperature of 42 °C and pH 7.5. Jayanthi et al. Performed the characterization of a thermostable chitinase from Bacillus licheniformis B2, observing optimal activity at 50 °C and enzymatic stability up to 70 °C; with maximum activity at pH 7.0. As a result of these findings, the authors consider that the enzyme can be employed in the recycling of chitin residues [29]. Hassiba et al. cloned, purified, characterized an extracellular chitinase of Bacillus licheniformis LHH100 isolated from wastewater samples in Algeria. The enzyme has optimal activity at pH 4.0 at 75 °C to be used in the bioconversion of chitin residues [30].
In their study, Menghiu et al. characterized the biochemical properties of a recombinant chitinase from Bacillus licheniformis DSM8785, expressed in Pichia pastoris KM71H. The optimal activity levels of the enzyme were reported with 140 U/mL at pH 4.0 to 5.0, and in a temperature range of 50 to 60°C [31].
The temperature as well as the host for enzyme expression is another variant that can be manipulated to reduce or increase chitinolytic activity, Narasimhan and Shivakumar reported the maximum activity of a chitinase from Bacillus subtilis at 30 °C [32], whereas another report provides data that, to produce chitinases from Bacillus licheniformis, the optimum temperature for expression is approximately 35 °C Jholapara et al. [33]. Similar to what was reported in this research where the optimum temperature is 42 °C. Similar research refers to the differences in the pH of different native chitinases from different microorganisms of the genus Bacillus. This was observed in reports of a chitinase from Bacillus turingiensis [34], which showed optimal activity at pH 7.0, and Bacillus laterosporous with optimal activity at pH 8.0 [35]. However, the chitinase ChiBlUV02 showed a pH different atthe range reported for enzymes chitinolytic Bacillus genus with regard to biological. The control chitinases with insecticidal capacity have been used for tested on various fungal and insect. The research of infestations by Brzezinska reports that a chitinase produced by the Aspergillus niger LOCK 62 inhibited the growth of the fungi phytopathogens: Fusarium culmorum, Fusarium solani and Rhizoctonia [36]. Aoki et al. reported that they expressed a chitinase produced by Trichoderma sp that was tested against Botrytis cinerea by inhibiting the growth of this fungus on cucumber leaves [37]. Essghaier et al. they obtained that the strain J24 from Bacillus licheniformis expresses a gene coding for a chitinase, where showed a fungicidal effect on Fusarium mangiferae [38]. Moon et al. investigated the efficacy of the chitinase, and the proteas produced by Bacillus licheniformis PR2 on the termite Reticulitermes speratus in conditions of laboratory, presenting a mortality of 88.9 % [39]. The purified enzyme ChiBlUV02 obtained a mortality rate of 50 and 56 % on two phases (larvae and adults) respectively of Aethina tumida. The effectiveness may be attributed to the sweeping of the cuticular relief of the exoskeleton and increased permeability, due to the hydrolyzed chitin, in addition to the acute effect caused by a possible atrophy due to enzymatic, action which is manifested in the increase of days for laying, as demonstrated by the results presented. Biochemical parameters such as temperature and pH of the recombinant chitinase ChiBlUV02 allowed the establishment of initial conditions for the application of the recombinant enzyme in pest control as a bioinsecticide, and therefore this represents an important possibility for the use of these enzymatic technologies as an alternative to control infestations such as that represented by Aethina tumida in the beekeeping, sectoring addition to not having the negative effect of traditional insecticides with chemicals, the chitinases ChiBlUV02 exhibit residual activity of approximately 80 % at the temperature conditions that have the hives (approximately 34 °C), so it would require any not conditions special application.
5. Conclusions
This study presents the development of an alternative treatment using recombinant chitinase ChiBlUV02 and its effects on Aethina tumida at two different life stages. In Mexico, there is currently no approved method for controlling this pest, which poses a significant threat to apiculture. The proposed approach offers a promising alternative to conventional chemical treatments, which can harm honeybees (Apis mellifera) and contaminate hive products, leading to economic losses for beekeepers.
The study found that ChiBlUV02 exhibited an enzymatic activity of 115 U/mL at 42 °C and pH 7.5, demonstrating a choleoptericidal effect on Aethina tumida, with a mortality rate of 50% in larvae and 56% in adults. Additionally, structural changes in the beetle's exoskeleton were observed, suggesting enzymatic degradation. These findings support the potential of recombinant chitinase as a novel biological control agent specifically targeting Aethina tumida. Future research should focus on optimizing the formulation and delivery of this enzymatic treatment to enhance its efficacy under field conditions. Some further studies on large-scale application are the stability in different environmental conditions, and potential synergistic effects with other biocontrol agents will be crucial for technology transfer and its eventual implementation in apiculture. The successful development of this biotechnological approach has the potential to provide a sustainable, environmentally friendly solution for Aethina tumida management, thereby benefiting the beekeeping industry.
Author Contributions
Deny de Jesús Velasco-Vique carried out the experimental, work analyzed the data and wrote the article. Argel Flores-Primo analyzed data and supervised the research. Sóstenes R. Rodríguez-Dehaibes collaborated in the collection and management of (Aethina tumida) María Guadalupe Sánchez-Otero and Violeta T. Pardío-Sedas analyzed the data obtained. Rosa María Oliart-Ros reviewed the experimental part of the protein purification, Gabriela Blasco-López contributed to the determination of chitinolytic activity and data analysis and Rodolfo Quintana-Castro provided the initial idea of the project, designed the study and revised the manuscript. All authors read and approved the final version of the manuscript.
Data Availability Statement
All the data are also available from the corresponding author on reasonable request.
Acknowledgments
This work was carried out with the support of the (SECIHTI) Secretariat of Science, Humanities, Technology and Innovation for doctoral grant No. 1075199.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Langner, T.; Göhre, V. Fungal chitinases: function, regulation, and potential roles in plant/pathogen interactions. Curr Genet. 2016, 62, 243–254. [Google Scholar] [CrossRef] [PubMed]
- Salazar, U. Production of chitinases from by-products. of the food: industry application to edible mushrooms and crustaceans. Seville; 2019.
- Bhattacharya, D.; Nagpure, A.; Gupta, R.K. Bacterial chitinases: properties and potential. Crit Rev Biotechnol. 2007, 27, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Swiontek Brzezinska, M.; Jankiewicz, U.; Burkowska, A.; Walczak, M. Chitinolytic microorganisms and their possible application in environmental protection. Curr Microbiol. 2014, 68, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Ramírez, M.V.; Calzadíaz, L. Industrial enzymes and metabolites from Actinobacteria in food and medicine industry. Actinobacteria - Basics and Biotechnological Applications; InTech, 2016. [Google Scholar] [CrossRef]
- Yang, S.; Fu, X.; Yan, Q.; Guo, Y.; Liu, Z.; Jiang, Z. Cloning, expression, purification and application of a novel chitinase from a thermophilic marine bacterium Paenibacillus barengoltzii. Food Chem. 2016, 192, 1041–1048. [Google Scholar] [CrossRef]
- Reverchon, F.; Diyarza Sandoval, N.A. Potential biological control agents against Fusarium spp. in Mexico: current situation, challenges and perspectives. 2021. [Google Scholar] [CrossRef]
- Liu, D.; Cai, J.; Xie, C.-C.; Liu, C.; Chen, Y.-H. Purification and partial characterization of a 36-kDa chitinase from Bacillus thuringiensis subsp. Colmeri and its biocontrol potential. Enzyme Microb Technol. 2010, 46, 252–256. [Google Scholar] [CrossRef]
- Thakur, N.; Nath, A.K.; Chauhan, A.; Gupta, R. Purification, characterization, and antifungal activity of Bacillus cereus strain NK91 chitinase from rhizospheric soil samples of Himachal Pradesh, India. Biotechnol Appl Biochem. 2022, 69, 1830–1842. [Google Scholar] [CrossRef]
- Verma, K.; Garg, N. antifungal activity and protoplast formation by chitinase produced from Bacillus licheniformis NK-7. Biology Fascicle/Analele Universităţii din Oradea. 2022;(1). https://www.bioresearch.ro/2022-1/061-067-AUOFB.29.1.2022-VERMA.K.-Antifungal.activity.and.protoplast.
- Borgi, M.A.; Khila, M.; Boudebbouze, S.; Aghajari, N.; Szukala, F.; Pons, N.; et al. The attractive recombinant phytase from Bacillus licheniformis: biochemical and molecular characterization. Appl Microbiol Biotechnol. 2014, 98, 5937–5947. [Google Scholar] [CrossRef]
- Gong, C.; Maquat, L.E. lncRNAs transactivate STAU1-mediated mRNA decay by duplexing with 3’ UTRs via Alu elements. Nature 2011, 470, 284–288. [Google Scholar] [CrossRef]
- Jellouli, K.; Ghorbel-Bellaaj, O.; Ayed, H.B.; Manni, L.; Agrebi, R.; Nasri, M. Alkaline-protease from Bacillus licheniformis MP1: Purification, characterization and potential application as a detergent additive and for shrimp waste deproteinization. Process Biochem. 2011, 46, 1248–1256. [Google Scholar] [CrossRef]
- Slimene, I.B.; Tabbene, O.; Gharbi, D.; Mnasri, B.; Schmitter, J.M.; Urdaci, M.-C.; et al. Isolation of a chitinolytic Bacillus licheniformis S213 strain exerting a biological control against Phoma medicaginis infection. Appl Biochem Biotechnol. 2015, 175, 3494–3506. [Google Scholar] [CrossRef]
- Kwon, J.-H.; Won, S.-J.; Moon, J.-H.; Lee, U.; Park, Y.-S.; Maung, C.E.H.; et al. Bacillus licheniformis PR2 controls fungal diseases and increases production of jujube fruit under field conditions. Horticulturae. 2021, 7, 49. [Google Scholar] [CrossRef]
- Murray, A.; Li, X. List of Coleoptera received from old calabar, on the west coast of Africa. Ann Mag Nat Hist. 1867, 20, 314–323. [Google Scholar] [CrossRef]
- Neumann, P.; Elzen, P.J. The biology of the small hive beetle (Aethina tumida, Coleoptera: Nitidulidae): Gaps in our knowledge of an invasive species. Apidologie. 2004, 35, 229–247. [Google Scholar] [CrossRef]
- Hood, W.M. The small hive beetle, Aethina tumida: a review. Bee World. 2004, 85, 51–59. [Google Scholar] [CrossRef]
- Richardson, K. Beekeeping role in enhancing food security and environmental public health. Health Economics and Management Review 2023, 4, 69–79. [Google Scholar] [CrossRef]
- Nikolova, T.; Dimitrova, I.; Teneva, A. The development of beekeeping in Bulgaria and the European Union in the last ten years. An overview. Bulgarian Journal of Animal Husbandry/Životnov Dni Nauki 2023. [Google Scholar]
- Lehninger Principles of Biochemistry 7th ed, W.H. Freeman, 2017.
- Syrový, I.; Hodný, Z. Staining and quantification of proteins separated by polyacrylamide gel electrophoresis. Journal of Chromatography B: Biomedical Sciences and Applications 1991, 569, 175–196. [Google Scholar] [CrossRef]
- Hsu, S.C.; Lockwood, J.L. Powdered chitin agar as a selective medium for enumeration of Actinomycetes in water and Soil1. Appl Microbiol. 1975, 29, 422–426. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Monreal, J.; Reese, E.T. The chitinase of Serratia marcescens. Can J Microbiol. 1969, 15, 689–696. [Google Scholar] [CrossRef]
- Gallagher, S.R. SDS-polyacrylamide gel electrophoresis (SDS-PAGE). Current Protocols Essential Laboratory Techniques 2012, 6, 7–10. [Google Scholar] [CrossRef]
- Haas Jorge Ismael Tucuch et, a.l. Alternative control of Aethina tumida Murray (Coleoptera: Nitidulidae) with plant powders. Acta Agrícola y Pecuaria 2020, 6, 1–9. [Google Scholar] [CrossRef]
- Muras, A.; Romero, M.; Mayer, C.; Otero, A. Biotechnological applications of Bacillus licheniformis. Crit Rev Biotechnol. 2021, 41, 609–627. [Google Scholar] [CrossRef]
- Jayanthi, N.; et al. Characterization of thermostable chitinase from Bacillus licheniformis B2. In IOP Conference Series: Earth and Environmental Science; IOP Publishing, 2019; Volume 293. [Google Scholar] [CrossRef]
- Laribi-Habchi, H.; Bouanane-Darenfed, A.; Drouiche, N.; Pauss, A.; Mameri, N. Purification, characterization, and molecular cloning of an extracellular chitinase from Bacillus licheniformis stain LHH100 isolated from wastewater samples in Algeria. Int J Biol Macromol. 2015, 72, 1117–1128. [Google Scholar] [CrossRef] [PubMed]
- Menghiu, G.; Ostafe, V.; Prodanovic, R.; Fischer, R.; Ostafe, R. Biochemical characterization of chitinase A from Bacillus licheniformis DSM8785 expressed in Pichia pastoris KM71H. Protein Expr Purif. 2019, 154, 25–32. [Google Scholar] [CrossRef]
- Narasimhan Ashwini Srividya Shivakumar "Optimization of chitinase produced by a biocontrol strain of Bacillus subtilis using Plackett-Burman design" Eur, J. Exp. Biol 2012, 2, 861–865.
- Jholapara, R.J.; Radhika, S.M.; Chhaya, S.S. Optimization of cultural conditions for chitinase production from chitinolytic bacterium isolated from soil sample. Int. J. Pharm. Biol. Sci 2013, 4, 464–471. [Google Scholar]
- Gomaa, E.Z. Chitinase production by Bacillus thuringiensis and Bacillus licheniformis: their potential in antifungal biocontrol. J Microbiol. 2012, 50, 103–111. [Google Scholar] [CrossRef]
- Shanmugaiah, V.; Mathivanan, N.; Balasubramanian, N.; Manoharan, P.T. Optimization of cultural conditions for production of chitinase by Bacillus laterosporous MML2270 isolated from rice rhizosphere soil. African Journal of Biotechnology 2008, 7, 2562–2568. [Google Scholar]
- Brzezinska, M.S.; Jankiewicz, U. Production of antifungal chitinase by Aspergillus niger LOCK 62 and its potential role in the biological control. Curr Microbiol. 2012, 65, 666–672. [Google Scholar] [CrossRef]
- Aoki, Y.; Haga, S.; Suzuki, S. Direct antagonistic activity of chitinase produced by Trichoderma sp. SANA20 as biological control agent for grey mould caused by Botrytis cinerea. Cogent Biol. 2020, 6, 1747903. [Google Scholar] [CrossRef]
- Essghaier, B.; Zouaoui, M.; Najjari, A.; Sadfi, N. Potentialities and characterization of an antifungal chitinase produced by a halotolerant Bacillus licheniformis. Curr Microbiol. 2021, 78, 513–521. [Google Scholar] [CrossRef] [PubMed]
- Moon, J.-H.; Ajuna, H.B.; Won, S.-J.; Choub, V.; Choi, S.-I.; Yun, J.-Y.; et al. The Anti-Termite Activity of Bacillus licheniformis PR2 against the Subterranean Termite, Reticulitermes speratus kyushuensis Morimoto (Isoptera: Rhinotermitidae). Forests. 2023, 14, 1000. [Google Scholar] [CrossRef]
Figure 1.
Visualized on 1% agarose gel. Lane (1) the corresponding band of the Chibluv01 gene with an approximate weight of 2000 bp; M: Molecular marker of 1 Kpb.
Figure 1.
Visualized on 1% agarose gel. Lane (1) the corresponding band of the Chibluv01 gene with an approximate weight of 2000 bp; M: Molecular marker of 1 Kpb.
Figure 2.
PCR products on agarose gel. Lanes (1 to 6) recombinant strains confirming the presence of the chibluv01 gene; lane M: Molecular marker 1 Kb.
Figure 2.
PCR products on agarose gel. Lanes (1 to 6) recombinant strains confirming the presence of the chibluv01 gene; lane M: Molecular marker 1 Kb.
Figure 3.
Polyacrylamide gel corresponding to the pHTP-Chibluv01 vector. Lane (M) corresponds to the protein weight marker, lane (1) and (2) refer to the negative induction control, lane (3) and (4) correspond to the ChiBlUV02 chitinase induction labeled in the box in lane 4.
Figure 3.
Polyacrylamide gel corresponding to the pHTP-Chibluv01 vector. Lane (M) corresponds to the protein weight marker, lane (1) and (2) refer to the negative induction control, lane (3) and (4) correspond to the ChiBlUV02 chitinase induction labeled in the box in lane 4.
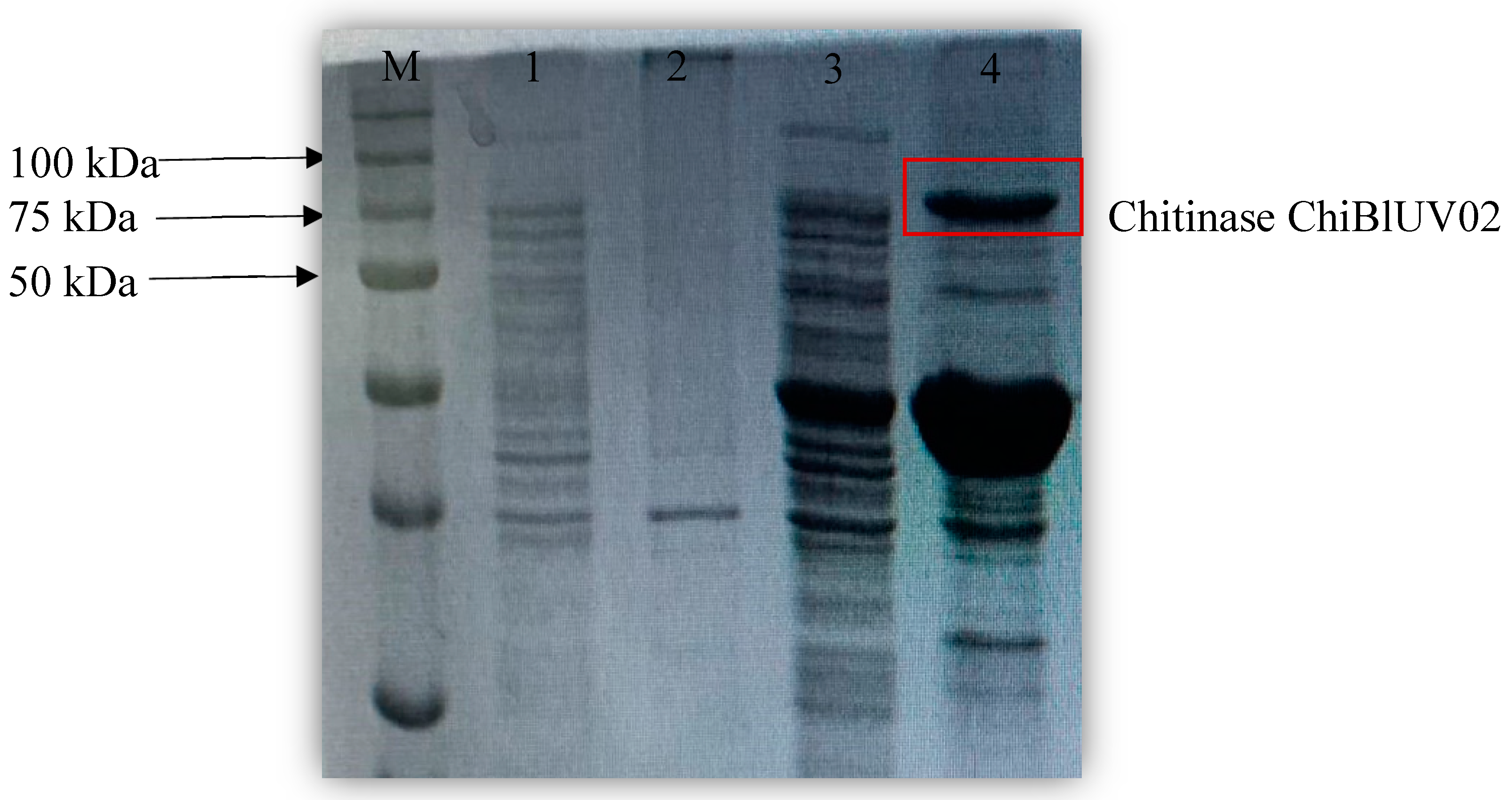
Figure 4.
Effect of temperature on chitinase activity ChiBlUV02.
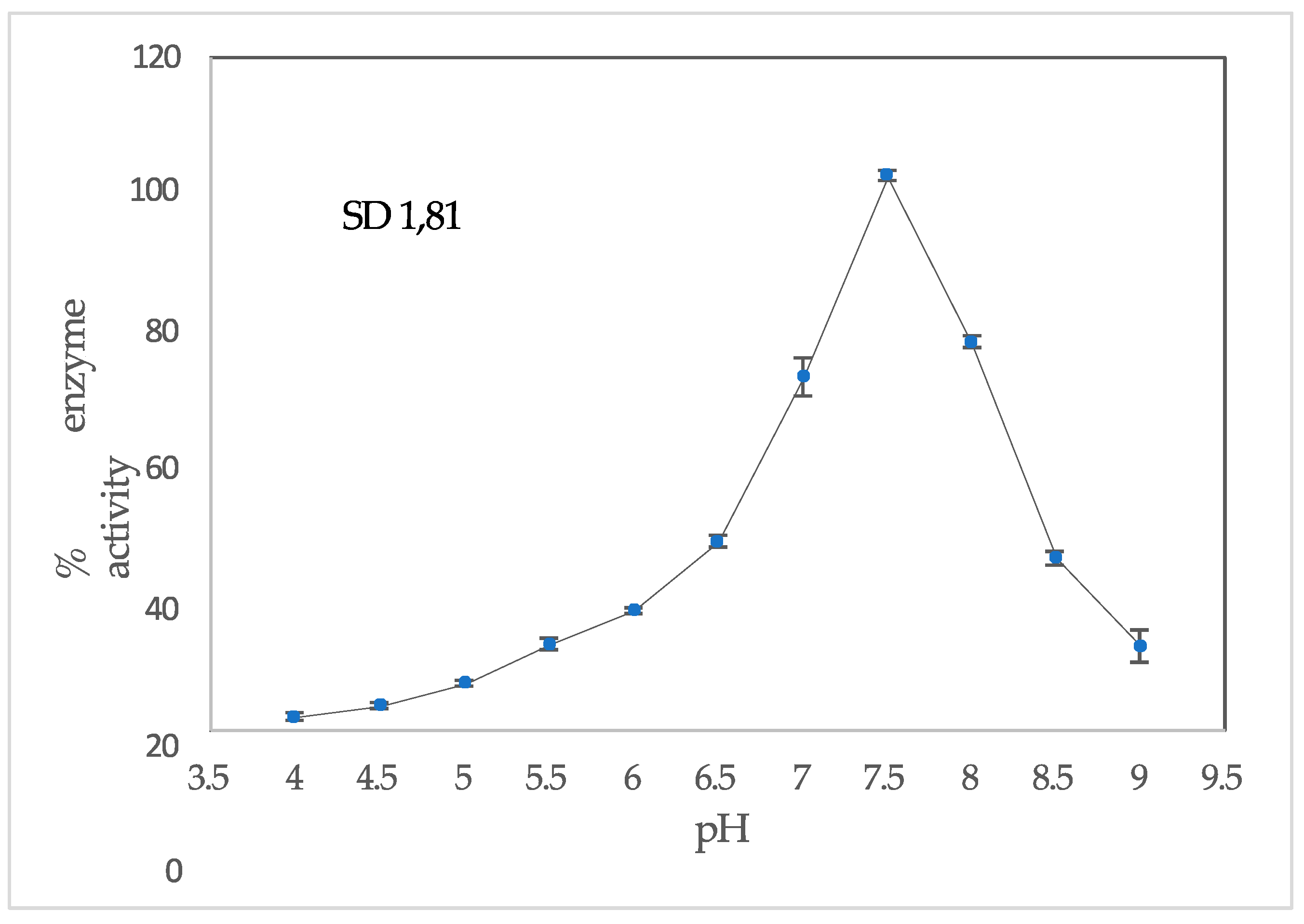
Figure 5.
Effect of pH on the activity of the purified chitinase ChiBlUV02.
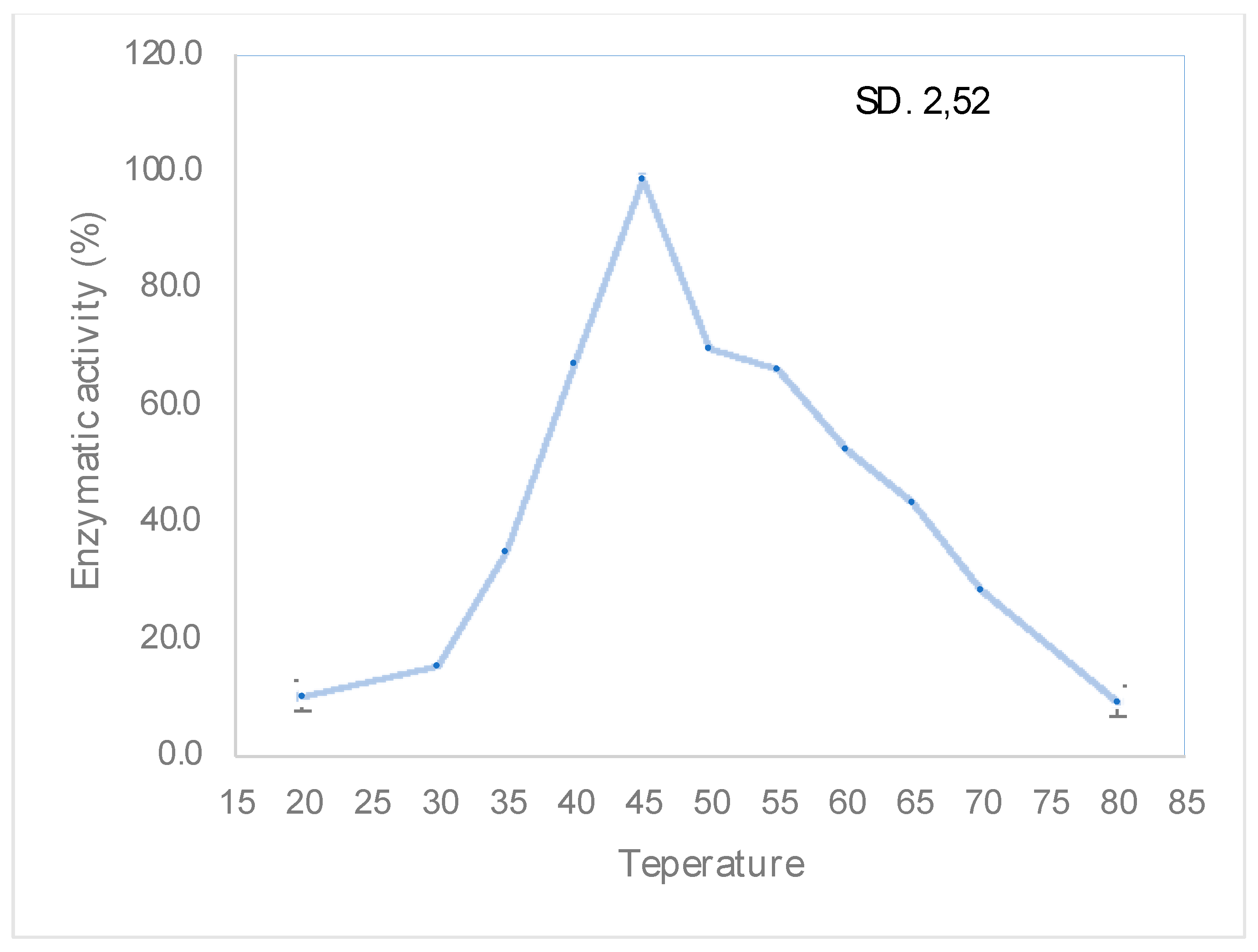
Figure 6.
SDS-PAGE polyacrylamide gel showing the purification of recombinant ChiBlUV02 protein by affinity chromatography with an approximate weight of 80 kDa Lane (1), Ni-resin retained fraction, lane (2) and (3) resin wash fractions and lane (4) resin elution fraction.
Figure 6.
SDS-PAGE polyacrylamide gel showing the purification of recombinant ChiBlUV02 protein by affinity chromatography with an approximate weight of 80 kDa Lane (1), Ni-resin retained fraction, lane (2) and (3) resin wash fractions and lane (4) resin elution fraction.

Figure 7.
Determination of LC50 and LC90 of purified chitinase ChiBlUV02 on adult Aethina tumida beetles.
Figure 7.
Determination of LC50 and LC90 of purified chitinase ChiBlUV02 on adult Aethina tumida beetles.
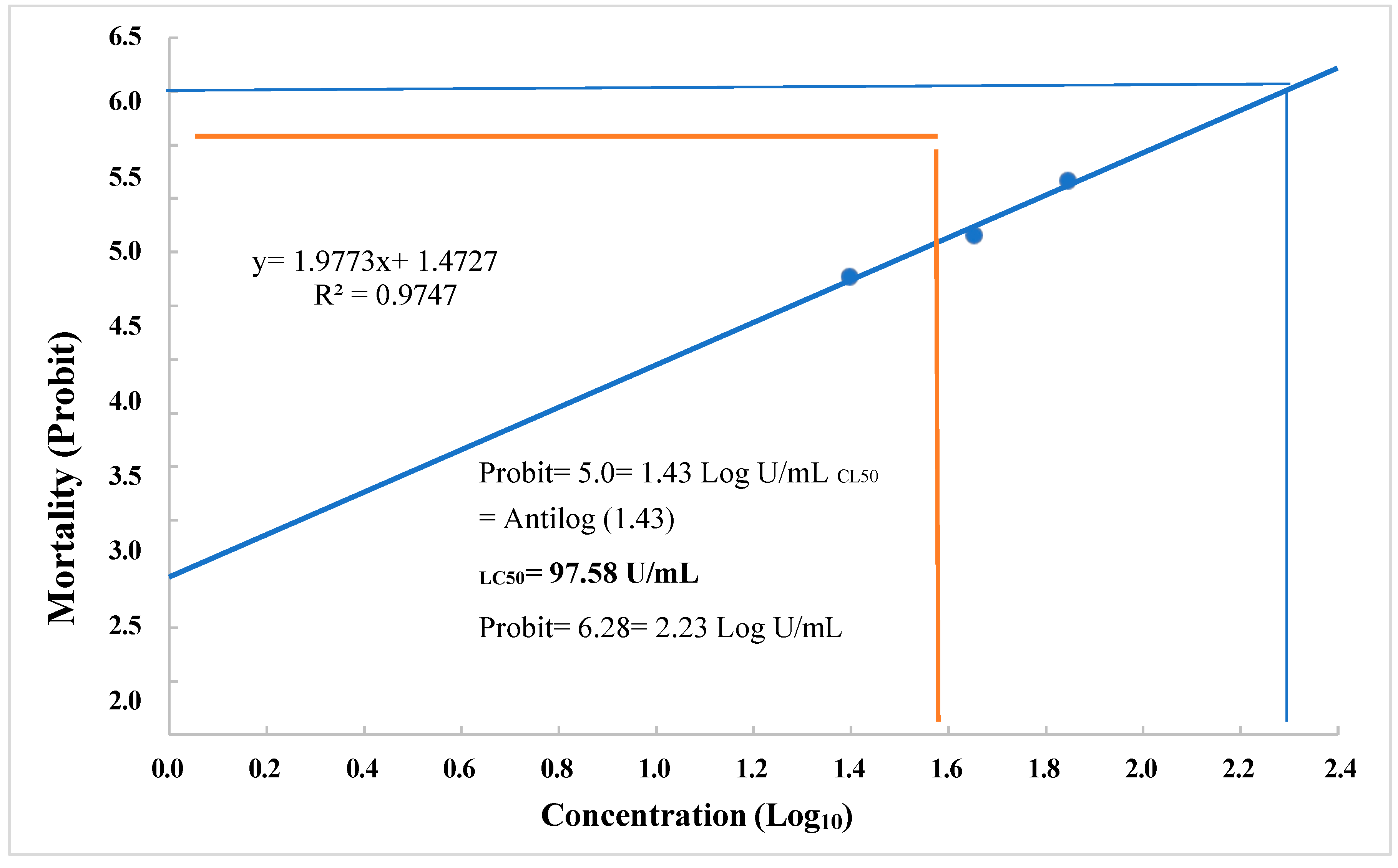
Figure 8.
Calculation of LC50 and LC90 of purified crude extract on Aethina tumida larvae.
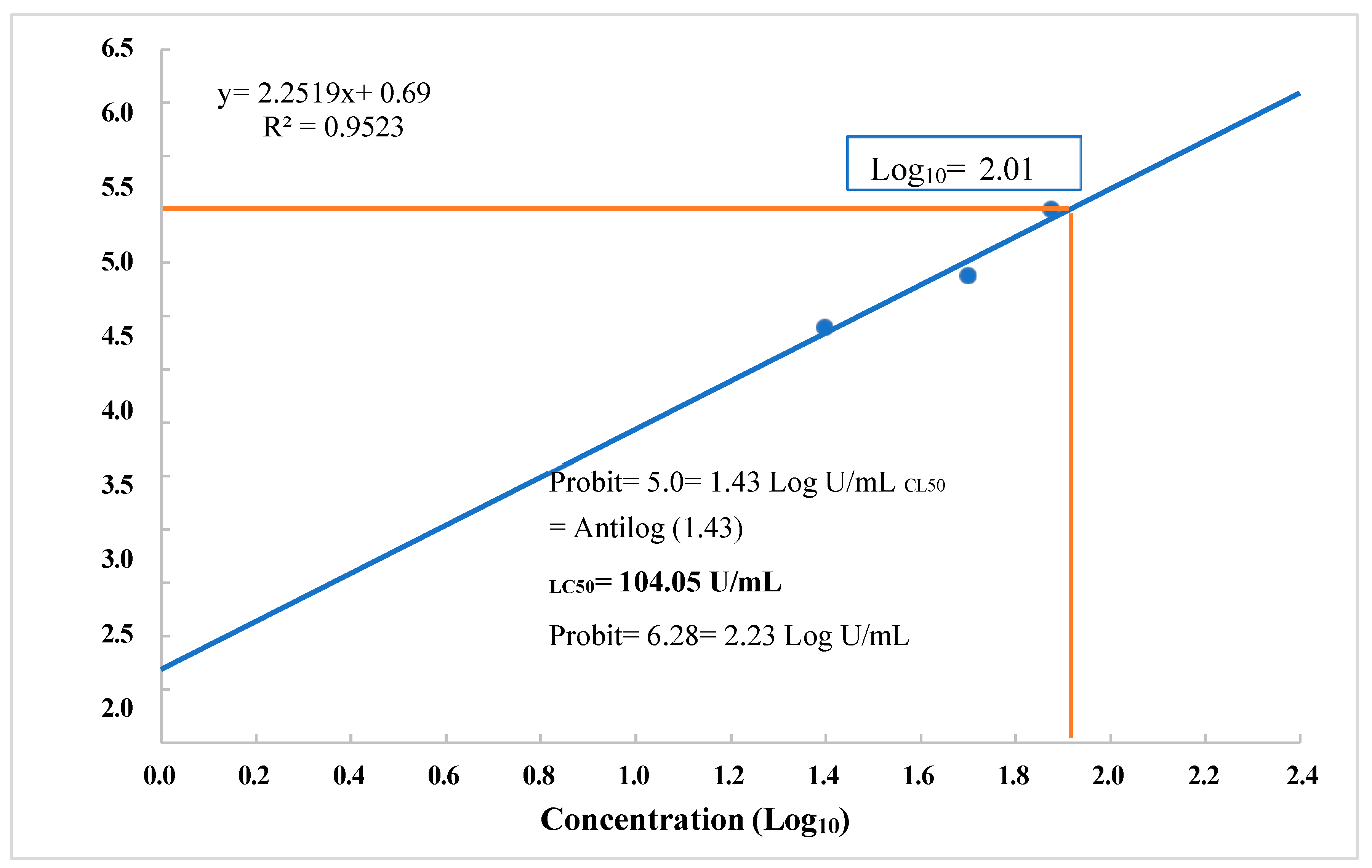
Figure 9.
Under digital optical stereoscopy, of the effect of the chitinases ChiBlUV02 at a concentration of 115 U/mL on the exoskeleton of adult beetles of Aethina tumida (a) control treatment; (b) post-treatment. Wear and deformation of the exoskeleton cuticle is observed.
Figure 9.
Under digital optical stereoscopy, of the effect of the chitinases ChiBlUV02 at a concentration of 115 U/mL on the exoskeleton of adult beetles of Aethina tumida (a) control treatment; (b) post-treatment. Wear and deformation of the exoskeleton cuticle is observed.
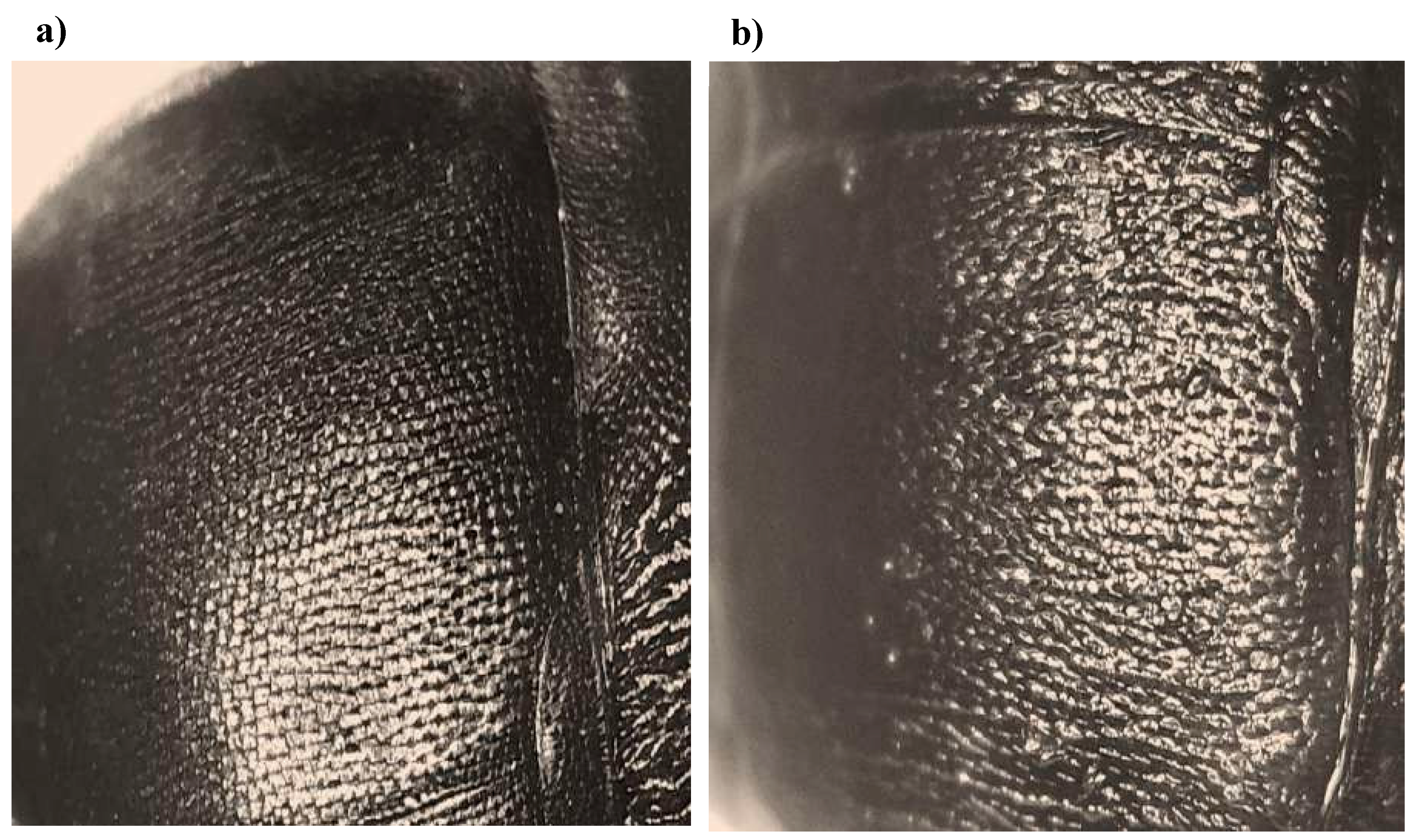
Figure 10.
Eggs of under digital optical stereoscopy. a) and b) post- effect of chitinase ChiBlUV02 at a concentration of 115 U/mL; c) and d) untreated control.
Figure 10.
Eggs of under digital optical stereoscopy. a) and b) post- effect of chitinase ChiBlUV02 at a concentration of 115 U/mL; c) and d) untreated control.
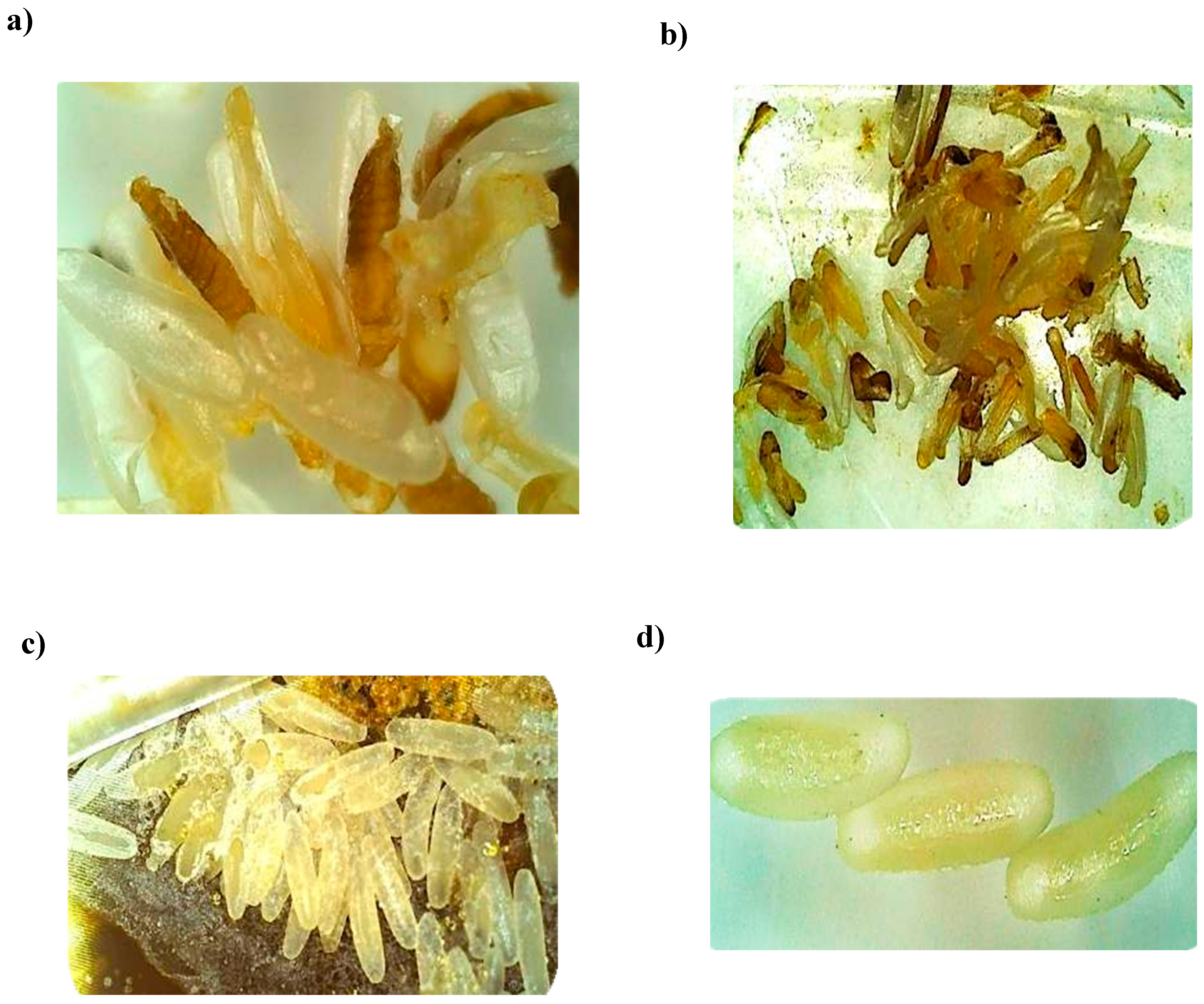

Table 1.
Choleoptericidal effect of the purified enzyme ChiBlUV02 at different time periods and concentrations of treatments on Aethina tumida larvae.
Table 1.
Choleoptericidal effect of the purified enzyme ChiBlUV02 at different time periods and concentrations of treatments on Aethina tumida larvae.
| 24 h | 48h | 72h | |||||
| Concentración (U/mL) | Live | Dead | Live | Dead | Live | Dead | Mortality |
| Control | 30 | 0 | 30 | 0 | 30 | 0 | 0 |
| 10 | 30 | 0 | 30 | 0 | 30 | 0 | 0 |
| 30 | 26 | 4 | 26 | 26 | 26 | 3 | 13 % |
| 45 | 22 | 8 | 21 | 21 | 21 | 7 | 30 % |
| 70 | 19 | 11 | 18 | 18 | 18 | 12 | 43 % |
| 115 | 15 | 15 | 15 | 15 | 15 | 15 | 50 % |
Table 2.
Choleoptericidal effect of the purified enzyme ChiBlUV02 at different time periods and concentrations of treatments on the adult stage of Aethina tumida.
Table 2.
Choleoptericidal effect of the purified enzyme ChiBlUV02 at different time periods and concentrations of treatments on the adult stage of Aethina tumida.
| 24 h | 48h | 72h | |||||
| Concentración (U/mL) | Live | Dead | Live | Dead | Live | Dead | Mortality |
| Control | 30 | 0 | 30 | 0 | 30 | 0 | 0 |
| 10 | 30 | 0 | 30 | 0 | 30 | 0 | 0 |
| 30 | 27 | 3 | 27 | 3 | 27 | 3 | 10 % |
| 45 | 23 | 7 | 23 | 7 | 23 | 7 | 23 % |
| 70 | 19 | 11 | 18 | 12 | 18 | 12 | 40 % |
| 115 | 13 | 17 | 13 | 17 | 13 | 17 | 56 % |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



