
Submitted:
19 March 2025
Posted:
20 March 2025
You are already at the latest version
Abstract
Rice blast caused by Magnaporthe oryzae pathotype is the worst disease that leads to serious food insecurity globally. Understanding rice blast disease pathogenesis is therefore essential for development of blast disease mitigation strategy. Reverse phosphorylation mediated by phosphatases perform a vital function in the activation of diverse biological mechanisms within eukaryotic. However, little has been reported on the roles of PP2Cs in the virulence of blast fungus. In this current work, we deployed functional genomics and biochemical approach to characterize type 2C protein phosphatase MoPtc6 in blast fungus. Deletion of MoPTC6 led to drastic reduction in conidiophore development, conidia production, hyphal growth, and stresses tolerance. Western blotting assay demonstrated that phosphorylation level of MoOsm1 was decreased while MoMps1 was increased in the MoPtc6 deletion mutant, comparative transcriptome assay revealed higher number of expressed genes between mutant and wild type. Localization assay confirmed that MoPtc6 is sub localized in the cytoplasm of mycelia, spores, and in the appressoria of M. oryzae. Furthermore, disruption of MoPTC6 impaired appressoria turgor pressure and glycogen utilization; more findings revealed attenuation of hyphal penetration and virulence upon deletion of MoPTC6. Generally, present findings suggested the role of MoPtc6 in the growth and virulence of M. oryzae.
Keywords:
Magnaporthe oryzae
; phosphorylation
; protein phosphatases
; stress tolerance
; virulence
1. Introduction
Magnaporthe oryzae is a filamentous phytopathogen which infects many cereals worldwide, especially rice [1]. Due to its adaptability and amenability to genetic manipulation, M. oryzae has been chosen by several researchers as a model fungal-plant pathosystem [2]. M. oryzae can reproduce asexually by forming appressoria, which facilitate the fungus to penetrate the host’s epidermal cells [3]. Blast disease affects rice production especially in temperate areas and has already been detected in more than eighty countries [4].
Type 2C protein phosphatases (PP2C) are potent serine/threonine protein phosphorylation and dephosphorylation factors and play important function in the regulation of diverse cellular biological mechanisms [5]. PP2C proteins belong to the family of metal-dependent protein phosphatases (PPM) that orientate towards several metal ions, including Mn2+, Mg2+, and Ca2 [6,7,8]. These metal ions are situated in the middle of the enzyme to link up to the phosphorus atoms and improve the precision of dephosphorylation assay [9].
In yeast PP2C proteins reported to perform indispensable role in diverse biological processes and pathways, such as cell wall maintenance by mediating Slt2 functions, high osmolality glycerol, rapamycin functions, and cell wall remodeling pathways [10,11,12,13]. Specifically, PTC6 is required in the activation and regulation of Slt2 during the dephosphorylation and vacuole formation under various stress factors, such as cell wall stress drugs in yeast cells [11]. In fungal pathogens, including Fusarium graminaerum, Magnaporthe oryzae, Fusarium oxysporum and Botrytis cinerea early findings revealed that PP2Cs are important in fungal pathogenesis [14,15,16]. Previous report disclosed that Ptc6 involved in the pathogenicity and cell wall conservation in F.oxysporum [15]. Disruption of PTC1 and PTC2 affected vegetative growth and virulence in B.cinerea [16]. In F. graminearum, loss of Ptc1 decreased virulence in wheat at diverse growth stages [17]. in M. oryzae, PTC5 and PTC7 regulate MoMps1 and MoOsm1 phosphorylation levels, cell wall integrity, appressorium turgor generation, and glycogen mobilization [18]. Moreover, rice blast PTC1 and PTC2 deletion mutants influenced appressorium morphogenesis, growth, and ultimately reduced pathogenicity [19,20]. Recently, PTC1 and PTC2 were demonstrated to influence conidia formation, seed infection, aflatoxin synthesis by regulating phosphoglycerate kinase1, and autophagic vesicles development in Aspergillus flavus [21]. Thus, these findings suggest that the functions of PP2Cs are highly related to the development and pathogenesis of diverse model fungi [22,23,24,25]. In this study, we examined the functional roles of MoPtc6 in relation to the growth and virulence of M.oryzae. The results demonstrated that Ptc6 mutants have weakened conidia production ability, perturbed vegetative growth, cell wall development, osmotic and oxidative stress tolerance, and regulation of MoMps1 and MoOsm1 phosphorylation. Taken together, findings provided evidence that the protein phosphatase MoPtc6 participates in fungal virulence and development.
2. Materials and Methods
2.1. Strain and Culture Conditions
Pathogenic fungus M. oryzae Guy11 was acquired from fungal Genetics Stock Centre (FGSC 9462) and was employed to generate mutant in this work. The WT Guy11, ∆Moptc6 and ∆Moptc6_C were spotted on conidiation inducing - media, rice bran media for ten days at 110rpm under 28oC, lastly were scratched and incubated for 3-5 days under constant light to produce conidia. For hyphal growth mycelia plugs were cultured on solid complete media (6 g of yeast extract, 6 g of casamino acid, 1L of ddH2O,20g of agar) for ten days at 110rpm under 28oC and diameter measurements were taken respectively. For protein, DNA and RNA extraction mycelia were oscillated in complete media for three days at 110rpm under 28oC. All the strains were kept on autoclaved dried papers for storage under 4oC for further uses.
2.2. Strategies for Generation of Mutant and Complementation Assays
To generate MoPTC6 deletion mutant, split marker homologous recombination approach was deployed [26]. Plasmid Vectors for knockout (split-marker) were constructed to delete MoPTC6 in M. oryzae genome. Flanking fragments A and B were amplified with appropriate primers designed, both fragments upstream and downstream were separately ligated with hygromycin transferase (hph) on pCX62 vector using overlapping polymerase chain reaction[27,28]. The PCR product used for generation of deletion mutants, were amplified for the constructed pCX62 vector using primer pair MoPTC6-BR+HY-F and MoPTC6-AF+YG-R. M. oryzae (Guy11) protoplast preparation and genetic transformation was conducted following the procedures described by [29,30,31]. Screening of putative transformants were performed by PCR using appropriate primers (MoPtc6_OR,MoPtc6_OF and MoPtc6_UA) and southern blotting assay to confirm the positive candidates (mutant) [32]. For complementation assays, The MoPTC6 fragment comprising of its native promoter and the open reading frame (ORF) genomic region was cloned behind GFP in the PKNTG vector. The MoPTC6-GFP construct was then transformed in ∆Moptc6 protoplasts and transformants were selected by PCR and GFP signals were visualized with a confocal microscope.
2.3. Southern Blotting Assay
Genomic DNA for southern blot analysis was extracted following cetyltrimethylammonium bromide (CTAB method) protocol [33,34]. Genomic DNA quality was examined by running on agarose, and the DNA concentration measured using Nano drop (Gene group campany Ltd, China). The mutant and the WT genomic DNA were digested by restriction enzyme (Cla I), this was followed by running the digested DNA on agarose gel to separate the bands under 120V for 10min then changed to 25V overnight. The gel was denatured by soaking it 0.5M NAOH and transfer to a positively charged nylon membrane which have binding capacity. The membrane was treated by UV crosslinker, prehybridation was performed under 42oC for 45min-2hours to block non-targets sites on the membrane which followed by hybridation (Probing the membrane) overnight to facilitate the binding of complementary bands in HL-2000 hybrilinker (LABrepco.Ltd, Shanghai, China). Lastly, the membrane was washed by 2X SSC+SDS, maleic acid solutions. Further membrane washing was conducted following starter kit I instructions and images were taken by Tanon imaging system.
2.4. RNA Extraction and qPCR Assays
For RNA extraction, the WT Guy11 and ∆Moptc6 mutant mycelia were cultured in complete media and oscillated at 110 rpm under 28oC for 72 hours. Mycelial samples were washed twice using double distilled water and filtered using sterilized filter paper. Total RNA was extracted following the instruction of Eastep super kit. cDNA was prepared by using Evo M-MLV RT kit. The quantitative polymerase chain reaction (q-PCR) was performed using Cham Q universal SYBR qPCR master mix and Eppendorf Realplex2 master (Eppendorf AG 22331 Hamburg) , genes expression were analyzed by the (2 − ΔΔCT) formula [35].
2.5. Colony Growth, Conidiation Formation and Conidiophores Development
To examine the fungal growth, mycelia plugs of mutant, Guy11 strain were spotted on solid complete media and kept in the incubator for 10 days under 28oC. The diameters were then measured using a ruler. For conidiation assay strains were spotted on conidia-inducing media (RBM), the hyphae were then scratched off on days 10 after inoculation and incubated in light chamber for an additional 72 hours. Double distilled water was used to wash conidia and counted under microscope. To examine the conidiophore development, strains were grown on RBM (Rice Bran Media), this was followed by scratching the medium and then, cut and transferred on microscopic slides. Conidiophore were observed under microscope at different time points (12h, 24h and 36h).
2.6. Evaluation of Pathogenicity, Hyphae Penetration and Incipient Cytorryhsis Assays
For pathogenicity evaluation and rice infection, spores were harvested from fungal cultured on rice bran media by using double distilled water containing 1% tween 20 solution then filtered with double layer of clean paper. Concentration of conidia adjusted up to (5×104) conidia/mL and used to spray 3 weeks old rice (Oryzae sativa). The sprayed plants were kept for 24 hours in the dark, then moved and incubated in the chamber where light is on for 12 hours and 12 hours of dark cycle. Disease development was monitored every day and images taken after seven days of inoculation. For penetration and invasive development assays, conidial suspension was applied on 10-day old detached barley leaves then the sheath peeled off. The invasive hyphae were examined and photograph was taken using Nikon microscope during different time points 30,48 and 60 hours respectively. To study incipient cytorryhsis assay (Cell collapse), 10 µL (5×104) conidia/mL suspension were added on the surface which is hydrophobic and kept in the incubator for 24 hours. that the water around spores was sucked and then exchanged with an equal volume of 1 M, 2M, and 3 M glycerol concentrations. The collapsed appressorium were recorded and viewed under microscope for each replicate.
2.7. Cell Sensitivity Assay
To evaluate influence of diverse stresses such as, osmotic cell wall reagents on ∆Moptc6 and Guy11. Fungi were cultured on complete media contained 200 µg/ mL Congo red, 0.01% SDS, 200 µg/mL CFW, 1 M NaCl, 1 M KCl, and 10mM hydrogen peroxide (H2O2) separately to stress the fungus as well. Later, the mutant and WT were cultured and kept at 28℃ for ten days, growth inhibition was calculated and photograph were taken respectively.
2.8. Total Protein Extraction and Western Blotting Assays
To extract total protein, ∆Moptc6 and Guy11 were inoculated in complete media and shaken at 110rpm under 28oC for 3 days. Then mycelia were harvested and grounded under liquid nitrogen then collected into 2mL tubes containing (1mL of protein extraction buffer containing 10µL of proteinase inhibitor and 10 µL of phenylmethyl sulphonyl fluoride). Centrifugation was conducted at 14000 rpm for 15 mins at 4°C. The upper clear supernatant liquid was pipetted carefully into flesh 2 mL tubes and SDS buffer added. The proteins were then denatured at 100oC for 10 minutes. The denatured proteins were analyzed by separating them on the SDS-PAGE and then sent to a nitrocellulose paper membrane. The targeted proteins on the membrane were detected with P-p38 MAPK, and P44/42MAPK primary antibody along with Goat antirabbit& mouse IgG-HRP second antibody. Actin mab was used as the reference antibody A chemiluminescent detection reagents, western brightTm (stabilized peroxide solution and lumino enhancer solution) was added and signal detected by western blotting Kit (Advanta). Finally, photograph was taken by Tanon-5200.
2.9. Microscopy Assays
Nikon confocal microscope was used to visualize and imaging of the green fluorescent protein light (GFP) photos. Conidia morphology, conidiophores development, appressorium development and glycogen dynamics from conidium to appressorium were observed by Olympus DP80 (Olympus co. Ltd, China, Beijing).
3.10. RNA Sequencing and Analysis
To extract RNA, mycelia derived from the ∆Moptc6 mutant and WT Guy11 strain were cultured in complete liquid media (CM) and shaken for three 3 days at 110rpm. The mycelia were then gathered by filtrating using autoclaved filter paper and washed with double distilled paper. The wet mycelia were then dried by squeezing out water using dry autoclaved filter, and crushed by using mortar and pestle in liquid nitrogen. Eastep super kit was used for total RNA extraction. cDNA was prepared following the instruction of Evo M-MLV RT kit. cDNA library was sequenced and cleaned, after that reads were produced. Lastly, the alignment of reads to the reference genome of M.oryzae by using Hist2 was performed. [36]. Differential expressed genes (DEGs) were examined with by DESeq. Further analysis were done by genes ontology and KEGG [37].
3. Results
3.1. Identification, Domain Architecture, and Phylogenetic Analysis of MoPtc6
To Identify MoPtc6 in Magnaporthe oryzae, we performed a homology BLAST search using Ptc6 protein sequence from Saccharomyces cerevisiae (https://blast.ncbi.nlm.nih.gov/Blast.cgi). The BLASTp hit reveal MoPTC6 (MGG_03918) gene in M. oryzae genome. Domain prediction and architecture revealed that PP2C_SIG domain is present in six MoPtc6 orthologs while the PP2C domain is conserved in four orthologs (Figure 1a). Phylogenetic analyses indicated that MoPtc6 shares the same ancestry as its orthologs in different fungi, including Fusarium gramineum, Fusarium oxysporum, Neurospora crassa and Botrytis cinerea and displayed an identity of a 83% with N. crassa and 97% with, F. graminearum, F. oxysporum and B. cinerea (Figure 1b). Next, we generated the 3D structure of MoPtc6 from the Swiss Model Software (https://www.genecards.org/.) (Figure 1c). Overall, these results suggest that Ptc6 protein phosphatase is conserved in pathogenic fungus.
3.2. Expression Profile of MoPTC6 During Pathogen-Host Interaction and Subcellular Localization
To explore the importance of MoPTC6 gene on fungal development. We first checked its expression during host pathogen interaction conidia were harvested from WT, Guy11 and used to spray on rice seedlings, samples were collected at different time points (4 h, 8 h, 12 h, 24 h, 48 h, and 72 h) and total RNA were extracted for qRT-PCR assay. The expression of MoPTC6 was upregulated at 48 and 72 hours post inoculation (hpi), suggesting that MoPTC6 could be crucial in hyphae invasion of host tissues (Figure 2a). We next investigated the co-localization of MoPtc6-GFP by using confocal scanning microscope. Results showed MoPtc6 is sub-localized in the cytoplasm of conidia, mycelia and in appressoria (Figure 2b).
3.3. Effects of MoPTC6 Deletion on Fungal Growth, Conidiophore Development, and Sporulation
To investigate the roles of MoPtc6 in fungal growth and conidiation of blast fungus, we monitored the growth of ∆Moptc6 , WT Guy11 on CM medium, Growth of ∆Moptc6 was remarkably attenuated compared to the WT strain (Figure 3a,b). For conidiation we harvested and quantified the amount of conidia generated by the mutant and WT grown on rice bran media and found that the ∆Moptc6 mutant generated few numbers of conidia (Figure 3c). Also, conidiophore assay revealed that the ∆Moptc6 produces few conidiophores structures than the WT Guy11 strain; the mutant conidiophores bear one or two conidia, unlike those of the WT Guy11 that had a bunch of conidia per conidiophore (Figure 3d). Moreover, we analyzed the expression levels of conidiation encoding genes such as MoCON1, MoCON6, MoCON7, MoFLBA, and MoFLB, the results indicated that all the genes were downregulated (Figure 3e). Overall, these findings revealed that MoPtc6 is important for conidiation and growth rate in M. oryzae.
3.4. Sensitivity of ΔMoptc6 Mutant to Stress-Inducing Agents
To evaluate the role of MoPtc6 on stress resistance, we cultured the WT, ΔMoptc6 mutant and ΔMoptc6_C strains on CM media with 200 µg/mL Congo red (CR), 200 µg/mL Calcofluor white (CFW), 1M sodium chloride (NaCl), 10mM hydrogen peroxide (H2O2), 1M potassium chloride (KCl) and 0.02% (SDS) as supplements. The ΔMoptc6 showed significant growth inhibition compared to the WT Guy11 due to the stresses agents (Figure 4a,b). These findings imply that MoPtc6 is important for cell wall integrity and osmoregulation. To further unveil the mechanism of stress resistance by MoPtc6, we examined the phosphorylation levels of stress-regulating proteins Mps1 and Osm1 in the various strains. Western blotting assay revealed that Osm1 phosphorylation decreased in ∆Moptc6 (Figure 4c,d). However, the phosphorylation level of Mps1 was expended in the ∆Moptc6 mutant in comparison to the WT (Figure 4e,f), suggesting the crucial role of MoPtc6 in the phosphorylation level of Mps1 and Osm1 proteins in blast fungus.
3.5. Deletion of MoPTC6 Attenuates Appressorium Turgor Pressure
For successful penetration, the blast fungus develops a critical structure called appressorium on the plant surface, which accumulates high turgor pressure used to forcefully puncture and penetrate the harder cuticle layer of the host [38]. To investigate the cause of reduced virulence in ∆Moptc6, we investigated and compared the appressorium turgor pressure of the various strains by harvesting fresh spores from the fungal strains and subjecting them to incipient cytorryhsis assay. Most of the ΔMoptc6 mutants were found to produce collapsed appressoria with significantly reduced turgor pressure as indicated by their percentage incipient cytorryhsis (Figure 5a,b). Because turgor generation depends on glycogen mobilization from conidia to appressoria and its subsequent degradation in the appressoria [39], we monitored the spatiotemporal mobilization and degradation of glycogen at different timepoints by potassium iodide (KI) staining. Within 24 h of KI staining, all the fungal strains had glycogen accumulated in their appressoria (Figure 5c,d). From 8 h through 12 h to 24 h of staining, the glycogen contents of these appressoria reduced with time, but the ΔMoptc6 mutant significantly retained more glycogen in the appressoria than the WT and ΔMoptc6_C strains, indicating a weaker glycogen degradation rate in the mutant, which results in weak turgor generation.
3.6. ∆Moptc6 Play a Significant Role in M. oryzae Virulence
To analyze the influence of MoPtc6 in the pathogenicity of M.oryzae , we sprayed rice seedlings with conidia suspension derived from the ΔMoptc6 mutant, Guy11 and the complemented strain. At 1-week post infection, the ΔMoptc6 leaves displayed a significantly reduced pathogenicity (Figure 6a,b). Similarly, we infected barley with mycelial plugs of ΔMoptc6 and WT strains and results revealed that the infection rate of the ∆Moptc6 was decreased (Figure 6c,d). We next evaluate the fungal hyphal invasion within barley epidermal cells at different time points 24h, 48hand 72h post inoculation. Results demonstrated that loss of MoPTC6 weakened the fungal invasive growth in the barley cells (Figure 6e). Taken together, this result indicates that MoPTC6 is required in the pathogenesis of M. oryzae.
3.7. Influence of MoPTC6 Gene Deletion on Global Gene Expression
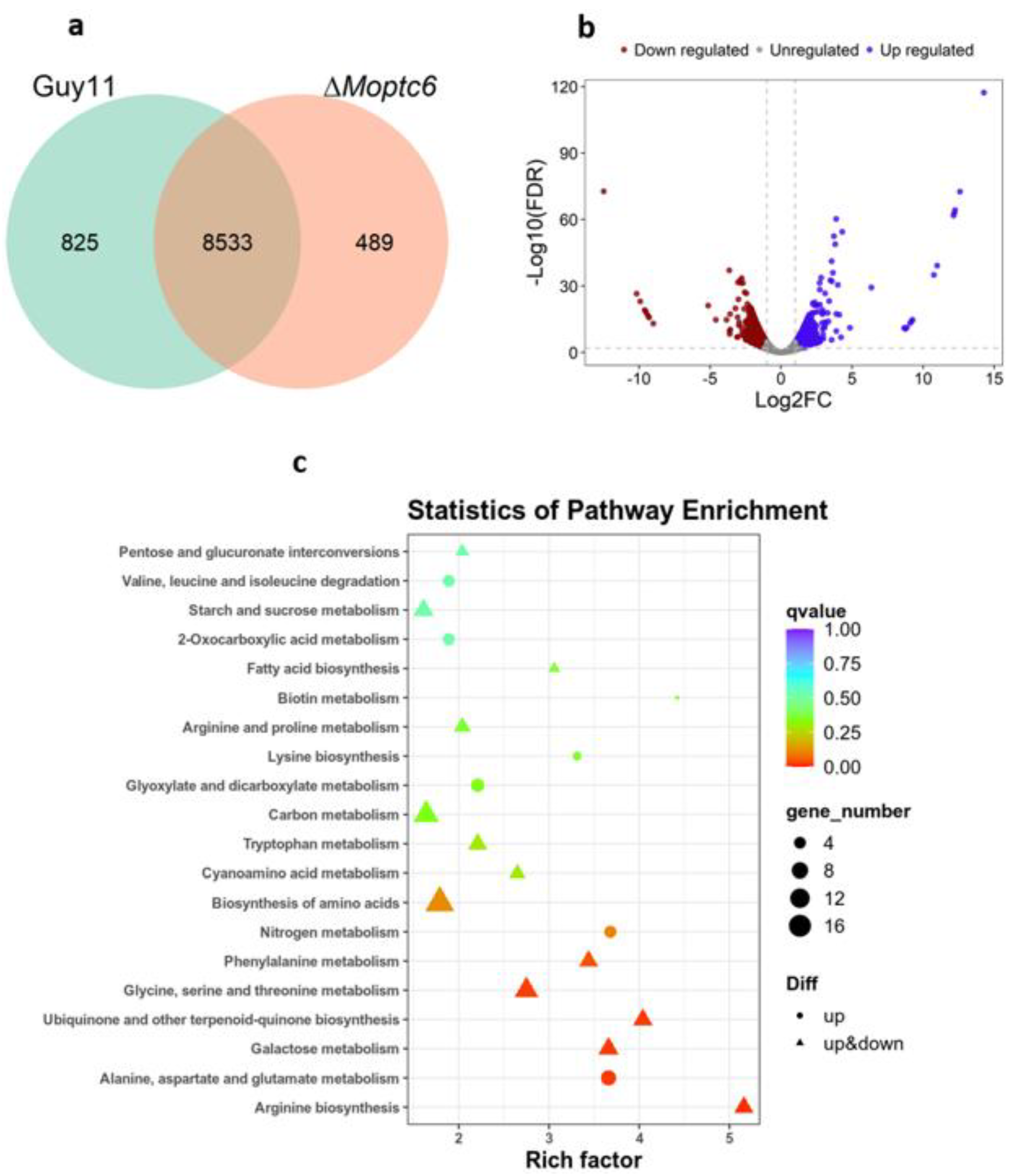
Due to the significant phenotype resulting from the loss of MoPTC6, we conducted RNA sequencing of the ∆Moptc6 compared to the WT (Guy11). This assay aimed to gain insight into differentially expressed genes (DEGs), GO, and KEGG that are enriched by expressed genes upon deletion of MoPTC6. The gene expression analysis revealed 8533 genes were expressed in both the WT and ∆Moptc6 mutant, with 825 and 489 genes expressed in the WT and ∆Moptc6 (Figure 7a,b). Next, we analyzed all DEGs based GO enrichment (P≤0.01). A total of 43 GO terms were enriched in biological processes (17 terms), cellular components (14 terms) and molecular functions (12 terms) (Figure S2). The KEGG enrichments of DEGs were also analyzed and the results showed that many pathways were associated with metabolic processes (Figure 7c).
4. Discussion
Type 2C protein phosphatases are important for diverse signaling and biological pathways in eukaryotes. Furthermore, their contributions in the physiology and development of M. oryzae remain poorly analyzed. The various PP2C have already been discussed in Saccharomyces cerevisiae [40,41]. The number of phosphatases identified in M. oryzae are in consistent with other filamentous model fungi including, Fusarium oxysporum, Botrytis cinerea and Fusarium gramineurum [14,15,40]. In this work, we explored the functions of PP2Cs MoPtc6 in fungal growth, stress tolerance, and virulence of rice blast. Firstly, we evaluated the transcription expression pattern of MoPtc6 in different developmental stages of the disease cycle and found MoPtc6 was up-regulated at 48, 72h post-inoculation (hpi). Indicating a possibly role in host colonization during plant-pathogen interaction. Since the localization of a protein is related to function, we decided to check the localization of MoPtc6 at all fungal developmental stages and the findings demonstrated that MoPtc6 is sub localized in the cytoplasm of hypha, spores, and appressoria respectively. These findings suggest that MoPtc6 might be required in different subcellular processes during fungal development and pathogenesis.
Conidiation is crucial for the pathogenesis of filamentous fungus such as Fusarium averaceum,Neurospora crassa and, Aspergillus nidulans [42,43]. Previous studies demonstrated that protein phosphatases are involved in asexual reproduction in M.oryzae [44]. This study further strengthens the involvement of protein phosphatases in reproduction process of rice blast fungus as the ∆Moptc6 produced few spores and conidiophores structures compared to the WT Guy11 strain which might be due to the weak expression of conidiation encoding genes [45].
Abiotic stresses such as osmotic and oxidative stressors were previously discussed and elaborated in S. cerevisiae [46,47]. Cell wall maintenance pathways are required in the stress adaptation mechanisms, cell anatomy and, virulence of M. oryzae [48]. In this work, the ∆Moptc6 growth was strongly inhibited by cell wall stresses agents such as congo red compared to the WT. thus, indication MoPTC6 important for tolerance to the cell wall stresses reagents in M. oryzae development which is in agreement with previous studies cited [49,50]. Ultimately, our findings revealed that mutant exhibited defect in osmotic stresses including, NACL and KCL, this suggested the involvement of Ptc6 in responses to hyperosmotic stress responses in M.oryzae and is in consistent with [51,52,53]. The variation of antioxidants and free radicals lead to oxidative stress in eukaryotes [54]. Furthermore, disruption of MoPTC6 increased sensitivity towards oxidative stress such as hydrogen peroxide (H2O2) this means that probably Ptc6 could be recruited in regulation of fungal oxidant-sensing pathways [55,56]. Osm1 which is commonly known as Hog1 protein is required in stress responses and phosphorylated under diverse osmotic, oxidative stresses, and fungicides [57,58,59]. Various cell wall stresses regulate Mps1 phosphorylation and MAP kinase pathway as earlier discussed [60,61,62,63]. Moreover, western blot assay demonstrated that deletion of MoPtc6 is important in the protein post-modification and phosphorylation of the MoMps1 and MoOsm1 in M. oryzae , these findings are in line with [15,64]. We suggested that MoPTC6 enhanced the activation of Slt2-mediated cell wall maintenance and regulate proteins responsible for stress in M. oryzae. Taken together, MoPTC6 Plays a key function in responses to the abiotic, biotic stresses, and homeostasis in M. oryzae development.
Appressoria turgor pressure is induced by higher concentration of metabolites which is essential for penetration and invasion of fungal hyphal into the host tissue [65,66,67]. Here, we demonstrated through biochemical assay (incipient cytorryhsis) that MoPTC6 potentially is important for appressorium turgor pressure, which confirmed by many appressoria collapsed in ∆Moptc6 after being treated with diverse concentration of glycerol. Moreover, ∆Moptc6 delayed glycogen transport and degradation from conidium to appressorium which showed that MoPtc6 is exclusively required in the trafficking of glycogen utilization in M. oryzae. our results are in line with previous publications where they detailed on Appressorium turgor during infection process in M.oryzae [68,69]. Here, we presumed that MoPTC6 participated in the glycogen metabolism and turgor pressure maintenance in M. oryzae.
For host infection many filamentous fungi, including rice M.oryzae need develop a structure called appressorium to gain entry into host organisms [70]. Through comprehensive analysis of ∆Moptc6 findings led to the reduction of virulence on the rice and on barley as well these findings are in line with early report on M.oryzae [71]. From these results, we proposed that MoPTC6 may serve as essential for pathogenicity and related pathways. Taken together, we suggested that PP2Cs are involved in the virulence mechanisms of M. oryzae.
5. Conclusion
Overall, we employed the filamentous fungus M. oryzae as a model pathosystem to identify and characterize the protein phosphatase MoPtc6 and revealed that the activities of MoPtc6 contribute to fungal virulence, conidia production, vegetative growth, appressorium turgor pressure generation, multi-stresses tolerance and transcripts expressed level. Based on the findings, we expect that new insights contribute to combat blast fungus extension.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Z.W., W.T., F.J and J. B. Designed this Research in details, funding acquisation , and manuscript revision; F. J., H. M., M. C., A. M. W. conducted the experiments and discussed about methodology; F.J., performed datas analysis and wrote the draft manuscript; J.B., Y.A.A.F; N.O; O.A. worked on editing of the final manuscript copy and referencing. All authors have critically read and approved the final manuscripts.
Funding
This work was supported by grants from the Central Guidance on Local Science and Technology Development Fund of Fujian Province (2022L3088 ), the Natural Science Foundation of Fujian Province (2022J01129), Fujian Provincial Science and Technology Key Project (2022NZ030014).
Acknowledgments
The authors are grateful to the Natural Science Foundation of Fujian Province (2022J01129), Fujian Provincial Science and Technology Key Project (2022NZ030014) for supporting our project.
Competing Interests
The authors declare that the research was conducted without any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Baudin, M.; Le Naour-Vernet, M.; Gladieux, P.; Tharreau, D.; Lebrun, M.H.; Lambou, K.; Leys, M.; Fournier, E.; Césari, S.; Kroj, T. Pyricularia oryzae: Lab star and field scourge. Mol Plant Pathol. 2024, 25, e13449.
- Wilson, R.A.; Talbot, N.J. Under pressure: investigating the biology of plant infection by Magnaporthe oryzae. Nat Rev Microbiol. 2009, 7, 185–195. [Google Scholar] [PubMed]
- Anjago, W.M.; Zhou, T.; Zhang, H.; Shi, M.; Yang, T.; Zheng, H.; Wang, Z. Regulatory network of genes associated with stimuli sensing, signal transduction and physiological transformation of appressorium in Magnaporthe oryzae. Mycol 2018, 9, 211–222. [Google Scholar]
- Eseola, A.B.; Ryder, L.S.; Osés-Ruiz, M.; Findlay, K.; Yan, X.; Cruz-Mireles, N.; Molinari, C.; Garduño-Rosales, M.; Talbot, N.J. Investigating the cell and developmental biology of plant infection by the rice blast fungus Magnaporthe oryzae. Fungal Genet Biol. 2021, 154, 103562. [Google Scholar]
- Escalona-Montaño, A.R.; Zuñiga-Fabián, M.; Cabrera, N.; Mondragón-Flores, R.; Gómez-Sandoval, J.N.; Rojas-Bernabé, A.; González-Canto, A.; Gutierrez-Kobeh, L.; Perez-Montfort, R.; Becker, I. Protein serine/threonine phosphatase type 2C of Leishmania mexicana. Front Cell infect Microbiol. 2021, 11, 641356. [Google Scholar]
- Shi, Y. Serine/threonine phosphatases: mechanism through structure. Cell 2009, 139, 468–484. [Google Scholar] [PubMed]
- Bradshaw, N.; Levdikov, V.M.; Zimanyi, C.M.; Gaudet, R.; Wilkinson, A.J.; Losick, R. A widespread family of serine/threonine protein phosphatases shares a common regulatory switch with proteasomal proteases. Elife. 2017, 6, e26111. [Google Scholar]
- Yang, Z.; Yang, J.; Wang, Y.; Wang, F.; Mao, W.; He, Q.; Xu, J.; Wu, Z.; Mao, C. Protein phosphatase95 regulates phosphate homeostasis by affecting phosphate transporter trafficking in rice. Plant Cell. 2020, 32, 740–757. [Google Scholar]
- Rej, R.; Bretaudiere, J.P. Effects of metal ions on the measurement of alkaline phosphatase activity. Clin Chem. 1980, 26, 423–428. [Google Scholar]
- Warmka, J.; Hanneman, J.; Lee, J.; Amin, D.; Ota, I. Ptc1, a type 2C Ser/Thr phosphatase, inactivates the HOG pathway by dephosphorylating the mitogen-activated protein kinase Hog1. Mol Cell Biol. 2001, 21, 51–60. [Google Scholar]
- Sharmin, D.; Sasano, Y.; Sugiyama, M.; Harashima, S. Type 2C protein phosphatase Ptc6 participates in activation of the Slt2-mediated cell wall integrity pathway in Saccharomyces cerevisiae. J Biosci Bioeng. 2015, 119, 392–398. [Google Scholar]
- Ruiz, A.; Xu, X.; Carlson, M. Ptc1 protein phosphatase 2C contributes to glucose regulation of SNF1/AMP-activated protein kinase (AMPK) in Saccharomyces cerevisiae. JBiol Chem. 2013, 288, 31052–31058. [Google Scholar]
- González, A.; Ruiz, A.; Serrano, R.; Ariño, J.; Casamayor, A. Transcriptional profiling of the protein phosphatase 2C family in yeast provides insights into the unique functional roles of Ptc1. J Biol Chem. 2006, 281, 35057–35069. [Google Scholar]
- Jiang, J.; Yun, Y.; Yang, Q.; Shim, W.-B.; Wang, Z.; Ma, Z. A type 2C protein phosphatase FgPtc3 is involved in cell wall integrity, lipid metabolism, and virulence in Fusarium graminearum. PloS one. 2011, 6, e25311. [Google Scholar]
- Nunez-Rodriguez, J.C.; Ruiz-Roldán, C.; Lemos, P.; Membrives, S.; Hera, C. The phosphatase Ptc6 is involved in virulence and MAPK signalling in Fusarium oxysporum. Mol Plant pathol. 2020, 21, 206–217. [Google Scholar]
- Yang, Q.; Jiang, J.; Mayr, C.; Hahn, M.; Ma, Z. Involvement of two type 2 C protein phosphatases B c P tc1 and B c P tc3 in the regulation of multiple stress tolerance and virulence of Botrytis cinerea. Envir Microbiol. 2013, 15, 2696–2711. [Google Scholar]
- Jiang, L.; Yang, J.; Fan, F.; Zhang, D.; Wang, X. The Type 2C protein phosphatase FgPtc1p of the plant fungal pathogen Fusarium graminearum is involved in lithium toxicity and virulence. Molecular plant pathology 2010, 11, 277–282. [Google Scholar]
- Biregeya, J.; Anjago, W.M.; Pan, S.; Zhang, R.; Yang, Z.; Chen, M.; Felix, A.; Xu, H.; Lin, Y.; Nkurikiyimfura, O. Type 2C Protein Phosphatases MoPtc5 and MoPtc7 Are Crucial for Multiple Stress Tolerance, Conidiogenesis and Pathogenesis of Magnaporthe oryzae. J Fungi 2023, 9, 1. [Google Scholar]
- Anjago, W.M.; Biregeya, J.; Shi, M.; Chen, Y.; Wang, Y.; Wang, Z.; Hong, Y.; Chen, M. The Calcium Chloride Responsive Type 2C Protein Phosphatases Play Synergistic Roles in Regulating MAPK Pathways in Magnaporthe oryzae. J Fungi 2022, 8, 1287. [Google Scholar]
- Cai, Y.; Liu, X.; Shen, L.; Wang, N.; He, Y.; Zhang, H.; Wang, P.; Zhang, Z. Homeostasis of cell wall integrity pathway phosphorylation is required for the growth and pathogenicity of Magnaporthe oryzae. Mol Plant Pathol 2022, 2022. 23, 1214–1225. [Google Scholar]
- Zhu, Z.; Yang, M.; Yang, G.; Zhang, B.; Cao, X.; Yuan, J.; Ge, F.; Wang, S. PP2C phosphatases Ptc1 and Ptc2 dephosphorylate PGK1 to regulate autophagy and aflatoxin synthesis in the pathogenic fungus Aspergillus flavus. Mbio. 2023, e00977–00923. [Google Scholar]
- Winkelströter, L.K.; Bom, V.L.P.; de Castro, P.A.; Ramalho, L.N.Z.; Goldman, M.H.S.; Brown, N.A.; Rajendran, R.; Ramage, G.; Bovier, E.; Dos Reis, T.F. H igh osmolarity glycerol response PtcB phosphatase is important for Aspergillus fumigatus virulence. Mol Microbiol. 2015, 96, 42–54. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Qin, Y.; Zhu, D.; Shan, A.; Feng, J. Identification and characterization of PP2C phosphatase SjPtc1 in Schistosoma japonicum. Parasitol intern 2018, 67, 213–217. [Google Scholar] [CrossRef] [PubMed]
- Kamada, R.; Kudoh, F.; Ito, S.; Tani, I.; Janairo, J.I.B.; Omichinski, J.G.; Sakaguchi, K. Metal-dependent Ser/Thr protein phosphatase PPM family: Evolution, structures, diseases and inhibitors. Pharmacol Therap 2020, 215, 107622. [Google Scholar] [CrossRef] [PubMed]
- Rogers, J.P.; Beuscher; Flajolet, M.; McAvoy, T.; Nairn, A.C.; Olson, A.J.; Greengard, P. Discovery of protein phosphatase 2C inhibitors by virtual screening. J Med Chem. 2006, 49, 1658-1667.
- Peri, K.V.R.; Faria-Oliveira, F.; Larsson, A.; Plovie, A.; Papon, N.; Geijer, C. Split-marker-mediated genome editing improves homologous recombination frequency in the CTG clade yeast Candida intermedia. FEMS yeast Res. 2023, 23. [Google Scholar] [CrossRef]
- Shen, B.; Xiao, J.; Dai, L.; Huang, Y.; Mao, Z.; Lin, R.; Yao, Y.; Xie, B. Development of a high-efficiency gene knockout system for Pochonia chlamydosporia. Microbiol Res. 2015, 170, 18–26. [Google Scholar] [CrossRef]
- Bryksin, A.V.; Matsumura, I. Overlap extension PCR cloning: a simple and reliable way to create recombinant plasmids. Biotech. 2010, 48, 463–465. [Google Scholar] [CrossRef]
- Ning, Y.; Hu, B.; Yu, H.; Liu, X.; Jiao, B.; Lu, X. Optimization of Protoplast Preparation and Establishment of Genetic Transformation System of an Arctic-Derived Fungus Eutypella sp. Front Microbiol. 2022, 13, 769008. [Google Scholar] [CrossRef]
- Talbot, N.J.; Ebbole, D.J.; Hamer, J.E. Identification and characterization of MPG1, a gene involved in pathogenicity from the rice blast fungus Magnaporthe grisea. Plant Cell. 1993, 5, 1575–1590. [Google Scholar]
- Nakayashiki, H.; Hanada, S.; Quoc, N.B.; Kadotani, N.; Tosa, Y.; Mayama, S. RNA silencing as a tool for exploring gene function in ascomycete fungi. Fungal Genet Biol. 2005, 42, 275–283. [Google Scholar] [CrossRef]
- Hattori, N.; Ushijima, T. Analysis of gene-specific DNA methylation. Handbook of epigenetics 2017, 113–123. [Google Scholar]
- Xu, J.-R.; Hamer, J.E. MAP kinase and cAMP signaling regulate infection structure formation and pathogenic growth in the rice blast fungus Magnaporthe grisea. Genes Dev. 1996, 10, 2696–2706. [Google Scholar] [CrossRef] [PubMed]
- Tripathy, S.K.; Maharana, M.; Ithape, D.M.; Lenka, D.; Mishra, D.; Prusti, A.; Swain, D.; Mohanty, M.R.; Raj, K.R. Exploring rapid and efficient protocol for isolation of fungal DNA. Intern Curr Microbiol Appl Sci. 2017, 6, 951–960. [Google Scholar] [CrossRef]
- Huhndorf, S.M.; Greif, M.; Mugambi, G.K.; Miller, A.N. Two new genera in the Magnaporthaceae, a new addition to Ceratosphaeria and two new species of Lentomitella. Mycol. 2008, 100, 940–955. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 1–21. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: a fast spliced aligner with low memory requirements. Nat Meth. 2015, 12, 357–360. [Google Scholar] [CrossRef]
- Ryder, L.S.; Talbot, N.J. Regulation of appressorium development in pathogenic fungi. Curr opin Plant Biol. 2015, 26, 8–13. [Google Scholar] [CrossRef]
- Aron, O.; Otieno, F.J.; Tijjani, I.; Yang, Z.; Xu, H.; Weng, S.; Guo, J.; Lu, S.; Wang, Z.; Tang, W. De novo purine nucleotide biosynthesis mediated by MoAde4 is required for conidiation, host colonization and pathogenicity in Magnaporthe oryzae. Appl Microbiol Biotechnol. 2022, 106, 5587–5602. [Google Scholar] [CrossRef]
- Arino, J.; Casamayor, A.; González, A. Type 2C protein phosphatases in fungi. Eukaryot Cell. 2011, 10, 21–33. [Google Scholar] [CrossRef]
- Albataineh, M.T.; Kadosh, D. Regulatory roles of phosphorylation in model and pathogenic fungi. Sabouraudia. 2015, 54, 333–352. [Google Scholar] [CrossRef]
- Su, Y.-Y.; Qi, Y.-L.; Cai, L. Induction of sporulation in plant pathogenic fungi. Mycol. 2012, 3, 195–200. [Google Scholar] [CrossRef]
- Pagliarulo, C.; Sateriale, D.; Scioscia, E.; De Tommasi, N.; Colicchio, R.; Pagliuca, C.; Scaglione, E.; Jussila, J.; Makkonen, J.; Salvatore, P. Growth, survival and spore formation of the pathogenic aquatic oomycete Aphanomyces astaci and fungus Fusarium avenaceum are inhibited by Zanthoxylum rhoifolium bark extracts in vitro. Fishes. 2018, 3, 12. [Google Scholar]
- Huang, Z.; Wang, Q.; Li, Y.; Huang, P.; Liao, J.; Wang, J.; Li, H.; Cai, Y.; Wang, J.; Liu, X. A multilayered regulatory network mediated by protein phosphatase 4 controls carbon catabolite repression and de-repression in Magnaporthe oryzae. Commun Biol. 2025, 8, 130. [Google Scholar]
- Kim, K.S.; Lee, Y.H. Gene expression profiling during conidiation in the rice blast pathogen Magnaporthe oryzae. PLoS One. 2012, 7, e43202. [Google Scholar]
- Brown, A.J.; Budge, S.; Kaloriti, D.; Tillmann, A.; Jacobsen, M.D.; Yin, Z.; Ene, I.V.; Bohovych, I.; Sandai, D.; Kastora, S. Stress adaptation in a pathogenic fungus. J Exper Biol. 2014, 217, 144–155. [Google Scholar]
- Nikolaou, E.; Agrafioti, I.; Stumpf, M.; Quinn, J.; Stansfield, I.; Brown, A.J. Phylogenetic diversity of stress signalling pathways in fungi. BMC Evol Biol. 2009, 9, 1–18. [Google Scholar]
- Feng, W.; Yin, Z.; Wu, H.; Liu, P.; Liu, X.; Liu, M.; Yu, R.; Gao, C.; Zhang, H.; Zheng, X.; et al. Balancing of the mitotic exit network and cell wall integrity signaling governs the development and pathogenicity in Magnaporthe oryzae. PLoS Pathog. 2021, 17, e1009080. [Google Scholar]
- Su, X.; Yan, X.; Chen, X.; Guo, M.; Xia, Y.; Cao, Y. Calcofluor white hypersensitive proteins contribute to stress tolerance and pathogenicity in entomopathogenic fungus, Metarhizium acridum. Pest manag sci. 2021, 77, 1915–1924. [Google Scholar] [CrossRef]
- Yoshimi, A.; Miyazawa, K.; Kawauchi, M.; Abe, K. Cell Wall Integrity and Its Industrial Applications in Filamentous Fungi. J Fungi. 2022, 8. [Google Scholar]
- Calcáneo-Hernández, G.; Landeros-Jaime, F.; Cervantes-Chávez, J.A.; Mendoza-Mendoza, A.; Esquivel-Naranjo, E.U. Osmotic Stress Responses, Cell Wall Integrity, and Conidiation Are Regulated by a Histidine Kinase Sensor in Trichoderma atroviride. J Fungi. 2023, 9. [Google Scholar]
- Liu, Y.; Gong, X.; Li, M.; Si, H.; Zhou, Q.; Liu, X.; Fan, Y.; Zhang, X.; Han, J.; Gu, S.; et al. Effect of Osmotic Stress on the Growth, Development and Pathogenicity of Setosphaeria turcica. Front Microbiol. 2021, 12, 706349. [Google Scholar]
- Duran, R.; Cary, J.W.; Calvo, A.M. Role of the osmotic stress regulatory pathway in morphogenesis and secondary metabolism in filamentous fungi. Toxins. 2010, 2, 367–381. [Google Scholar] [PubMed]
- Chaudhary, P.; Janmeda, P.; Docea, A.O.; Yeskaliyeva, B.; Abdull Razis, A.F.; Modu, B.; Calina, D.; Sharifi-Rad, J. Oxidative stress, free radicals and antioxidants: potential crosstalk in the pathophysiology of human diseases. Front Chem. 2023, 11, 1158198. [Google Scholar]
- Bai, Z.; Harvey, L.M.; McNeil, B. Oxidative stress in submerged cultures of fungi. Crit Rev Biotechnol. 2003, 23, 267–302. [Google Scholar] [PubMed]
- Simaan, H.; Lev, S.; Horwitz, B.A. Oxidant-Sensing Pathways in the Responses of Fungal Pathogens to Chemical Stress Signals. Front Microbiol. 2019, 10, 567. [Google Scholar]
- Segmüller, N.; Ellendorf, U.; Tudzynski, B.; Tudzynski, P. BcSAK1, a stress-activated mitogen-activated protein kinase, is involved in vegetative differentiation and pathogenicity in Botrytis cinerea. Eukaryot Cell 2007, 6, 211–221. [Google Scholar]
- Tatebayashi, K.; Yamamoto, K.; Tomida, T.; Nishimura, A.; Takayama, T.; Oyama, M.; Kozuka-Hata, H.; Adachi-Akahane, S.; Tokunaga, Y.; Saito, H. Osmostress enhances activating phosphorylation of Hog1 MAP kinase by mono-phosphorylated Pbs2 MAP2K. Embo j 2020, 39, e103444. [Google Scholar]
- Stępień, Ł.; Lalak-Kańczugowska, J. Signaling pathways involved in virulence and stress response of plant-pathogenic Fusarium species. Fungal Biol Rev. 2021, 35, 27–39. [Google Scholar]
- Yin, Z.; Tang, W.; Wang, J.; Liu, X.; Yang, L.; Gao, C.; Zhang, J.; Zhang, H.; Zheng, X.; Wang, P.; et al. Phosphodiesterase MoPdeH targets MoMck1 of the conserved mitogen-activated protein (MAP) kinase signalling pathway to regulate cell wall integrity in rice blast fungus Magnaporthe oryzae. Mol Plant Pathol 2016, 17, 654–668. [Google Scholar]
- Zhang, X.; Wang, Z.; Jiang, C.; Xu, J.R. Regulation of biotic interactions and responses to abiotic stresses by MAP kinase pathways in plant pathogenic fungi. Stress Biol. 2021, 1, 5. [Google Scholar]
- Wang, Y.; Cui, X.; Xiao, J.; Kang, X.; Hu, J.; Huang, Z.; Li, N.; Yang, C.; Pan, Y.; Zhang, S. A novel MAP kinase-interacting protein MoSmi1 regulates development and pathogenicity in Magnaporthe oryzae. Mol Plant Pathol. 2024, 25, e13493. [Google Scholar]
- Abah, F.; Kuang, Y.; Biregeya, J.; Abubakar, Y.S.; Ye, Z.; Wang, Z. Mitogen-Activated Protein Kinases SvPmk1 and SvMps1 Are Critical for Abiotic Stress Resistance, Development and Pathogenesis of Sclerotiophoma versabilis. J fungi. 2023, 9. [Google Scholar] [CrossRef]
- Angelova, M.B.; Pashova, S.B.; Spasova, B.K.; Vassilev, S.V.; Slokoska, L.S. Oxidative stress response of filamentous fungi induced by hydrogen peroxide and paraquat. Mycoll Res. 2005, 109, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Chumley, F.G.; Valent, B. Genetic analysis of melanin-deficient, nonpathogenic mutants of Magnaporthe grisea. Mol. Plant-Microbe Interact. 1990, 3, 135–143. [Google Scholar] [CrossRef]
- Liu, X.-H.; Liang, S.; Wei, Y.-Y.; Zhu, X.-M.; Li, L.; Liu, P.-P.; Zheng, Q.-X.; Zhou, H.-N.; Zhang, Y.; Mao, L.-J. Metabolomics analysis identifies sphingolipids as key signaling moieties in appressorium morphogenesis and function in Magnaporthe oryzae. MBio 2019, 10, 10.1128. [Google Scholar] [CrossRef] [PubMed]
- Wengler, M.R.; Talbot, N.J. Mechanisms of regulated cell death during plant infection by the rice blast fungus Magnaporthe oryzae. Cell Death Differ. 2025. [Google Scholar] [CrossRef]
- Liu, C.; Shen, N.; Zhang, Q.; Qin, M.; Cao, T.; Zhu, S.; Tang, D.; Han, L. Magnaporthe oryzae transcription factor MoBZIP3 regulates Appressorium turgor pressure formation during pathogenesis. Intern J Mol Sci. 2022, 23, 881. [Google Scholar] [CrossRef]
- Lin, L.; Cao, J.; Du, A.; An, Q.; Chen, X.; Yuan, S.; Batool, W.; Shabbir, A.; Zhang, D.; Wang, Z. eIF3k domain-containing protein regulates conidiogenesis, appressorium turgor, virulence, stress tolerance, and physiological and pathogenic development of Magnaporthe oryzae. Front Plant Sci. 2021, 12, 748120. [Google Scholar] [CrossRef]
- Xu, J.R.; Hamer, J.E. MAP kinase and cAMP signaling regulate infection structure formation and pathogenic growth in the rice blast fungus Magnaporthe grisea. Genes Dev. 1996, 10, 2696–2706. [Google Scholar] [CrossRef]
- Aron, O.; Wang, M.; Lin, L.; Batool, W.; Lin, B.; Shabbir, A.; Wang, Z.; Tang, W. MoGLN2 Is Important for Vegetative Growth, Conidiogenesis, Maintenance of Cell Wall Integrity and Pathogenesis of Magnaporthe oryzae. J Fungi. 2021, 7. [Google Scholar] [CrossRef]
Figure 1.
Domain structure and phylogenetic of MoPtc6 and its orthologs. a Represent the domains architecture of Ptc6 proteins from different fungi (PP2Cs domain and PP2C_SIG domain) the architecture was built using IBS software (IBS.biocuckoo.org).b Analysis of the phylogenetic relationship of Ptc6 proteins among different pathogenic fungi. The tree was produced by using MEGA X software.c 3D structure of MoPtc6 generated from Swiss-Model Software https://www.genecards.org/. Mo: Magnaporthe oryzae; Fo: Fusarium oxysporum; Fg: Fusarium gramineurum; Sc: Saccharomyces Cerevisiae ; Bc: Botrytis Cinerea.
Figure 1.
Domain structure and phylogenetic of MoPtc6 and its orthologs. a Represent the domains architecture of Ptc6 proteins from different fungi (PP2Cs domain and PP2C_SIG domain) the architecture was built using IBS software (IBS.biocuckoo.org).b Analysis of the phylogenetic relationship of Ptc6 proteins among different pathogenic fungi. The tree was produced by using MEGA X software.c 3D structure of MoPtc6 generated from Swiss-Model Software https://www.genecards.org/. Mo: Magnaporthe oryzae; Fo: Fusarium oxysporum; Fg: Fusarium gramineurum; Sc: Saccharomyces Cerevisiae ; Bc: Botrytis Cinerea.
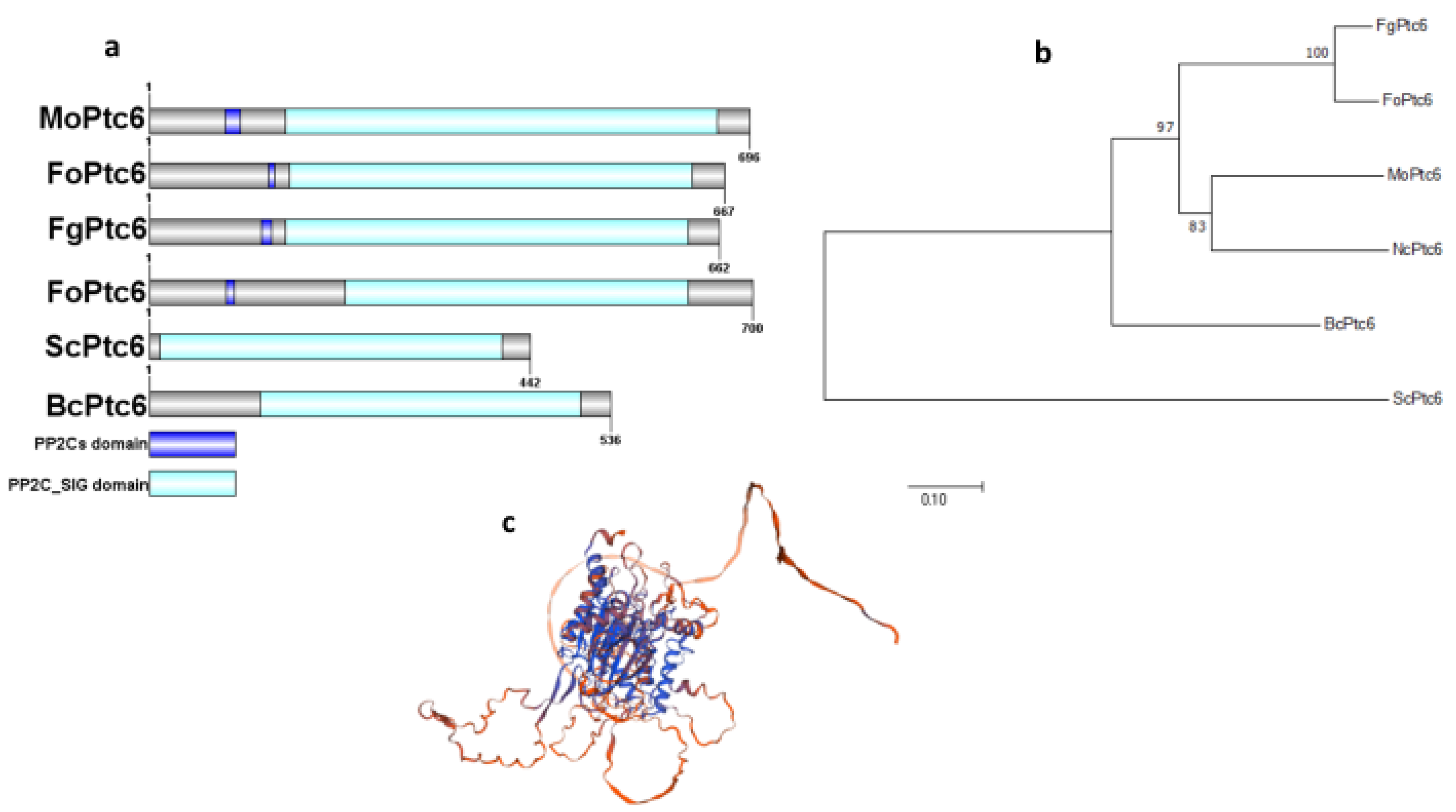
Figure 2.
Expression pattern and Localization of MoPTC6 in M. oryzae. a Transcription patterns of MoPTC6 during interaction with the host, three weeks rice seedlings were sprayed with Guy11 and the samples were harvested at different time points (4 h,8 h,12 h,24 h,48 h, and 72 h).b Displayed Sub localization of MoPtc6-GFP in the mycelia, conidium and appressoria. Images were taken by using confocal scanning microscope. (** p < 0.01).
Figure 2.
Expression pattern and Localization of MoPTC6 in M. oryzae. a Transcription patterns of MoPTC6 during interaction with the host, three weeks rice seedlings were sprayed with Guy11 and the samples were harvested at different time points (4 h,8 h,12 h,24 h,48 h, and 72 h).b Displayed Sub localization of MoPtc6-GFP in the mycelia, conidium and appressoria. Images were taken by using confocal scanning microscope. (** p < 0.01).
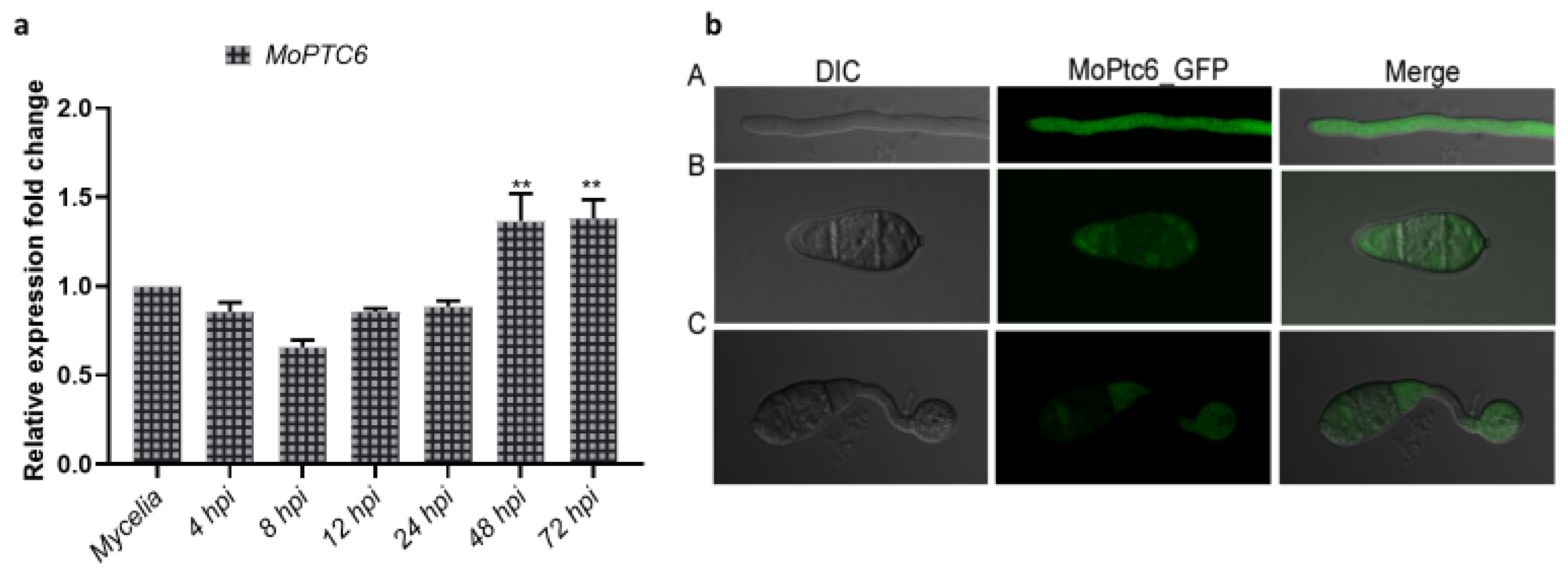
Figure 3.
MoPtc6 is important for growth and conidiation in M. oryzae. a, b Growth and colony diameters of the mutant and WT grown on solid complete media at 28oC for ten days. c, d Quantification of conidia and conidiophores development in the ∆Moptc6 and WT (E) Expression levels of conidiation related genes in ∆Moptc6 compared to the WT. Asterisks show statistically significant difference analyzed by ANOVA (**, P < 0.01).
Figure 3.
MoPtc6 is important for growth and conidiation in M. oryzae. a, b Growth and colony diameters of the mutant and WT grown on solid complete media at 28oC for ten days. c, d Quantification of conidia and conidiophores development in the ∆Moptc6 and WT (E) Expression levels of conidiation related genes in ∆Moptc6 compared to the WT. Asterisks show statistically significant difference analyzed by ANOVA (**, P < 0.01).
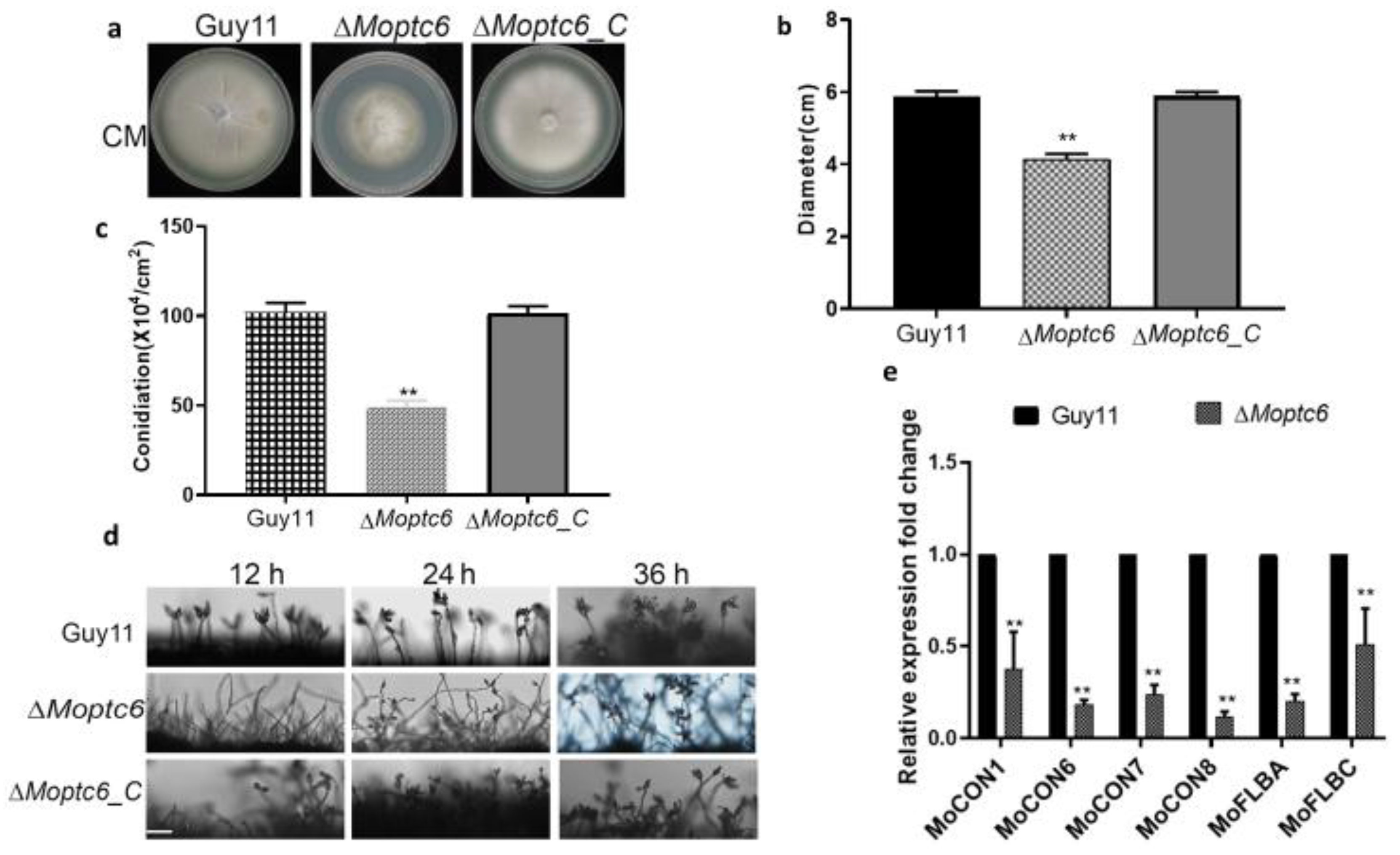
Figure 4.
Sensitivity of ∆Moptc6 mutant and WT to various stresses. a Growth of the strains on CM media with 200µg/mL CR, 0.01% SDS, 200 µg/mL CFW, 1M KCl, 1M NaCl and 10Mm H2O2 at 28oC for ten days. b inhibition of growth of the strains due to the diverse fungal stressors. c, d Show Osm1 phosphorylation level, MoOsm1 were bind to P38 Thr180/Tyr182 antibody and actin was used as a control, quantification of bands intensity was done with ImageJ software. e, f Indicated Mps1 phosphorylation level, MoMps1 were detected with antiphospho P44/42 antibody and actin was used as a control. (F) Displayed bands intensity in mutant and WT. Asterisks show statistically significant difference (*, P < 0.05 ; **, P < 0.01).
Figure 4.
Sensitivity of ∆Moptc6 mutant and WT to various stresses. a Growth of the strains on CM media with 200µg/mL CR, 0.01% SDS, 200 µg/mL CFW, 1M KCl, 1M NaCl and 10Mm H2O2 at 28oC for ten days. b inhibition of growth of the strains due to the diverse fungal stressors. c, d Show Osm1 phosphorylation level, MoOsm1 were bind to P38 Thr180/Tyr182 antibody and actin was used as a control, quantification of bands intensity was done with ImageJ software. e, f Indicated Mps1 phosphorylation level, MoMps1 were detected with antiphospho P44/42 antibody and actin was used as a control. (F) Displayed bands intensity in mutant and WT. Asterisks show statistically significant difference (*, P < 0.05 ; **, P < 0.01).
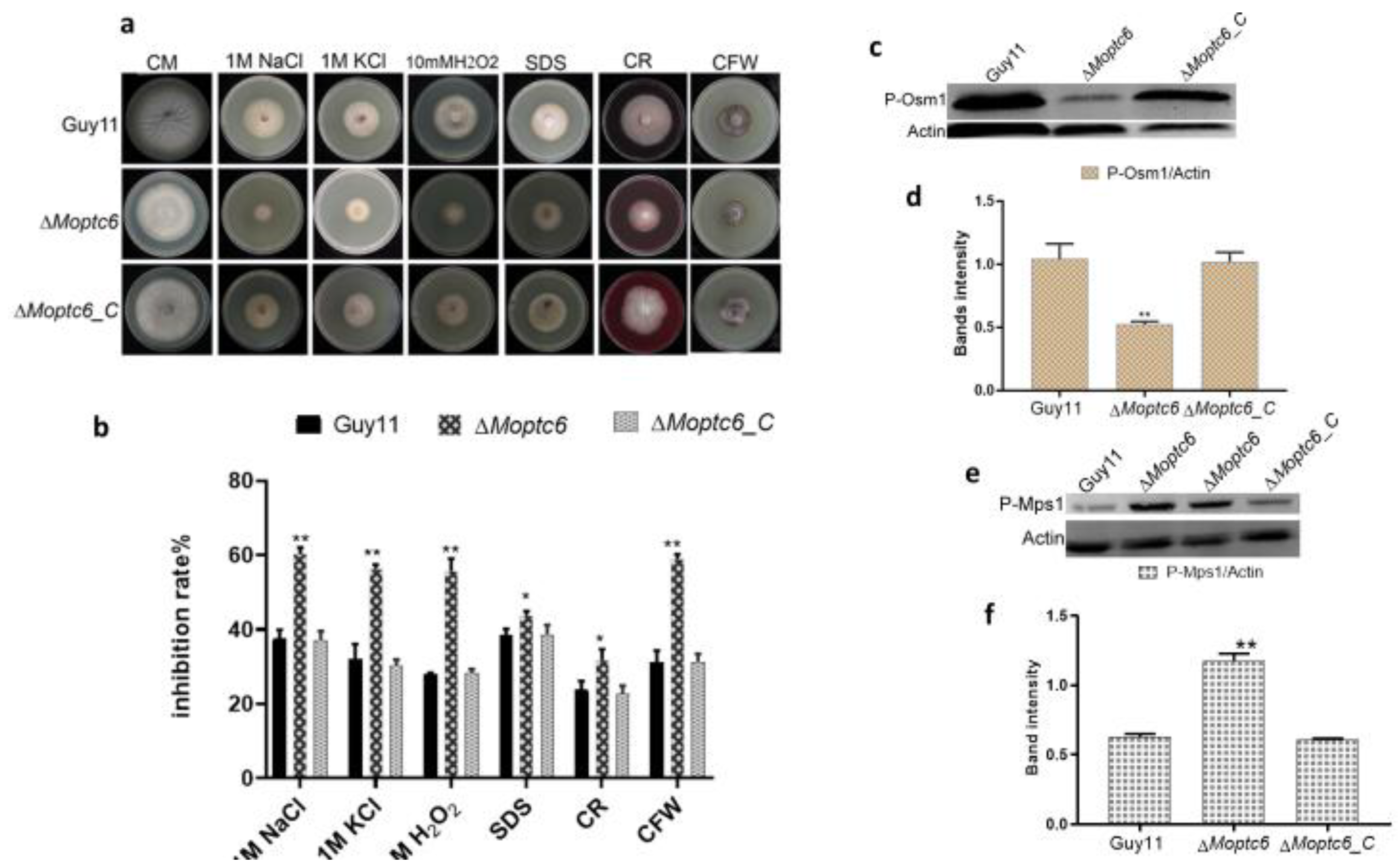
Figure 5.
∆Moptc6 promote appressoria turgor pressure and mobilization of glycogen. (A, B) Indicates incipient cytorryhsis conducted to examine appressorium turgor pressure for ∆Moptc6, WT and complementation strains, appressoria were incubated for 24 h period and tested by various concentration of glycerol 1 ,2, and 3 M 5 min before viewed by using microscope. (C, D) Micrograph displayed glycogen utilization from conidium to appressorium and percentages of glycogen levels in conidium and in appressorium at different time points 4 h,8 h,12 h and 24 h in mutant compared to WT. Asterisks Displays statistically significant (*, P < 0.05 ; **, P < 0.01).
Figure 5.
∆Moptc6 promote appressoria turgor pressure and mobilization of glycogen. (A, B) Indicates incipient cytorryhsis conducted to examine appressorium turgor pressure for ∆Moptc6, WT and complementation strains, appressoria were incubated for 24 h period and tested by various concentration of glycerol 1 ,2, and 3 M 5 min before viewed by using microscope. (C, D) Micrograph displayed glycogen utilization from conidium to appressorium and percentages of glycogen levels in conidium and in appressorium at different time points 4 h,8 h,12 h and 24 h in mutant compared to WT. Asterisks Displays statistically significant (*, P < 0.05 ; **, P < 0.01).
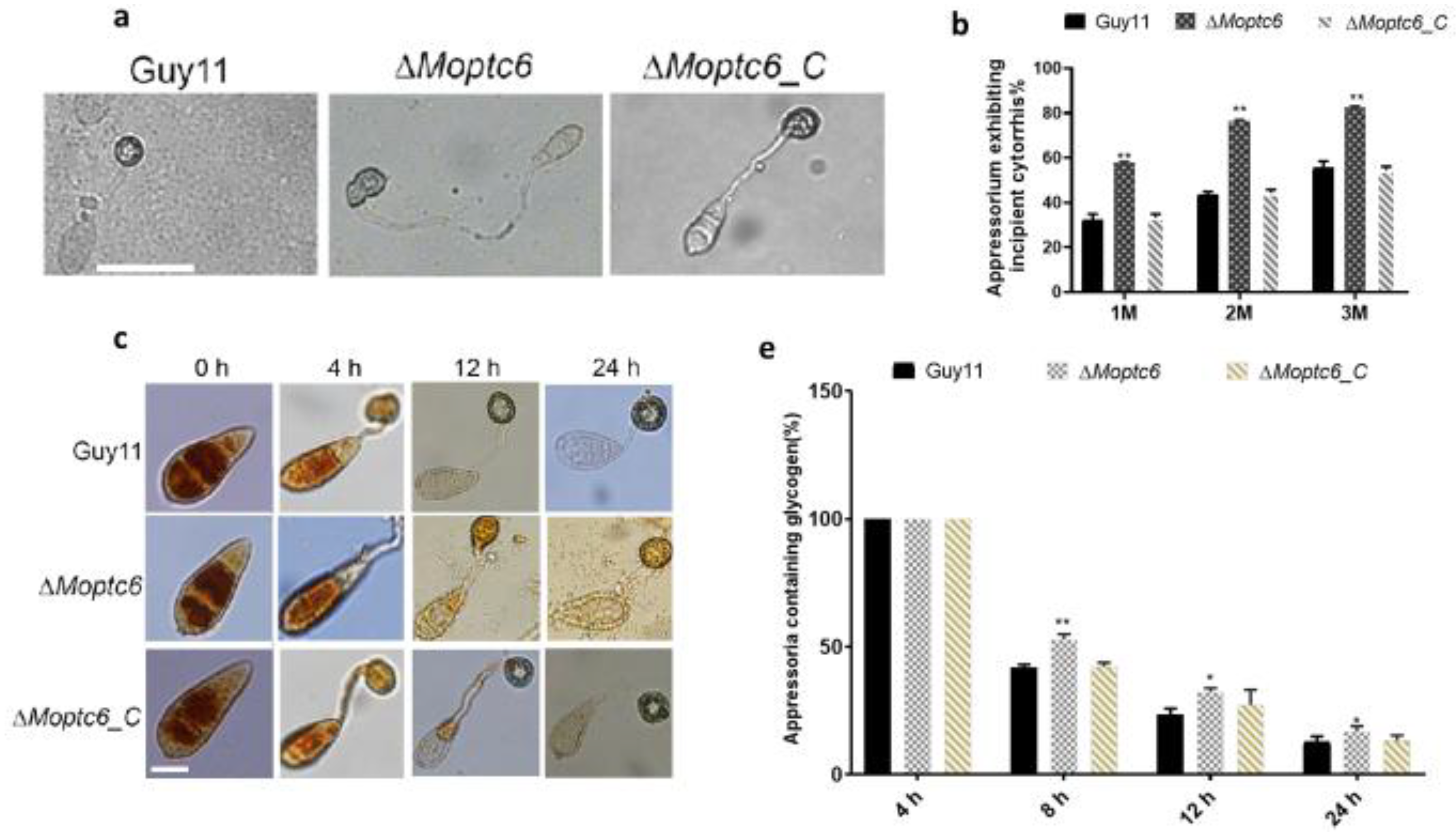
Figure 6.
Virulence analysis of MoPtc6 in M. oryzae. a, b The figures displayed the pathogenicity analysis and lesions quantification assay for ∆Moptc6, WT and complementation strain, images were captured one-week post spraying. c, d Indicate the lesions area on barley leaves induced by ΔMoptc6, ΔMoptc6-com and the WT strains mycelia plugs. e Observation of invasive hyphae in ten- days old barley leaves infected with 5×104 spores/mL and photographed after 24h, 48h, and 72h post inoculation. Asterisks Displays statistically significant (*, P < 0.05 ; **, P < 0.01).
Figure 6.
Virulence analysis of MoPtc6 in M. oryzae. a, b The figures displayed the pathogenicity analysis and lesions quantification assay for ∆Moptc6, WT and complementation strain, images were captured one-week post spraying. c, d Indicate the lesions area on barley leaves induced by ΔMoptc6, ΔMoptc6-com and the WT strains mycelia plugs. e Observation of invasive hyphae in ten- days old barley leaves infected with 5×104 spores/mL and photographed after 24h, 48h, and 72h post inoculation. Asterisks Displays statistically significant (*, P < 0.05 ; **, P < 0.01).
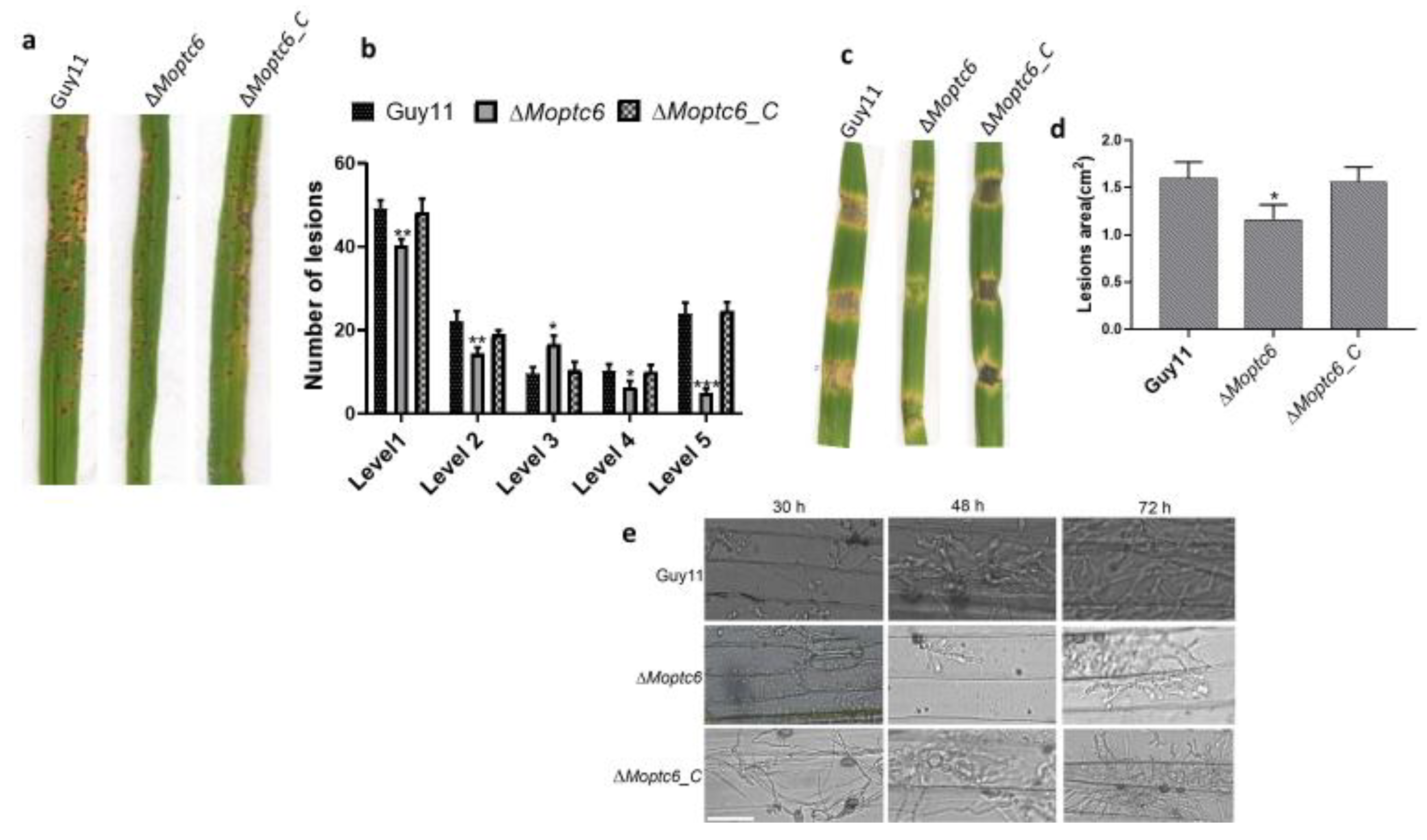
Figure 7.
Expression of genes between ∆Moptc6 and WT. a Global view of genes expression levels in ∆Moptc6 and WT, this figure was generated by Jvenn software b Volcano figure Plotted by using R software, indicating differential expressed genes (DEGs) distribution patterns in ∆Moptc6 mutant and WT. Red dot demonstrate downregulated genes, blue dots represent up-regulated genes. c Displaying the KEGG pathways enriched by differential expressed genes (DEGs) (P < 0.01).
Figure 7.
Expression of genes between ∆Moptc6 and WT. a Global view of genes expression levels in ∆Moptc6 and WT, this figure was generated by Jvenn software b Volcano figure Plotted by using R software, indicating differential expressed genes (DEGs) distribution patterns in ∆Moptc6 mutant and WT. Red dot demonstrate downregulated genes, blue dots represent up-regulated genes. c Displaying the KEGG pathways enriched by differential expressed genes (DEGs) (P < 0.01).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



