
Submitted:
15 February 2024
Posted:
16 February 2024
You are already at the latest version
Abstract
The multifunctional carbon-catabolite repression-negative on TATA box less complex (CCR4-NOT) is a multi-subunit complex present in all eukaryotes, including fungi. This complex plays an essential role in gene expression; however, a functional study of the CCR4-NOT complex in the rice blast fungus Magnaporthe oryzae has not been conducted. Seven genes encoding the putative CCR4-NOT complex were identified in the M. oryzae genome. Among these, a homologous gene, MoNOT3, was overexpressed during appressorium development in a previous study. Deletion of MoNOT3 in M. oryzae resulted in a significant reduction in hyphal growth, conidiation, abnormal septation in conidia, conidial germination, and appressorium formation compared to the wild-type. Transcriptional analyses suggested that the MoNOT3 gene affects conidiation and conidial morphology by regulating COS1 and COM1 in M. oryzae. Furthermore, monot3 exhibited a lack of pathogenicity, both with and without wounding, which is attributable to deficiencies in the development of invasive growth in planta. This result was also observed in onion epidermal cells, which are non-host plants. In addition, the MoNOT3 gene was involved in cell wall stress responses and heat shock. Taken together, these observations suggest that the MoNOT3 gene is required for fungal infection-related cell development and stress responses in M. oryzae.
Keywords:
appressorium
; CCR4-NOT complex
; conidiation
; functional analysis
; Magnaporthe oryzae
; pathogenicity
1. Introduction
While the canonical TATAAA sequence is a prevalent core promoter element in eukaryotes, not all core promoters contain this sequence, leading to the identification of "TATA-less" promoters [1]. One such regulator is the carbon-catabolite repression-negative on TATA box less (Ccr4-Not) complex. The CCR4-Not complex is a highly conserved multi-subunit protein found in eukaryotic cells, including fungi, and plays an essential role in regulating gene expression. These functions include interacting with transcription factors, initiating transcription, elongating transcription, exporting messenger RNA (mRNA), degrading mRNA, deadenylating mRNA, and repressing mRNA translation. The complex is consistently found in the nucleus and cytoplasm and is present at all stages of gene expression throughout the life cycle, indicating its importance [2,3,4].
In yeast, a classic model fungus, the CCR4-Not complex is structurally composed of nine conserved subunits, including Ccr4 and associated factor Caf genes (Caf130, Caf40, and Caf1) as well as five Not genes (Not1, Not2, Not3 (SIG1), Not4 (Mot2), Not5) [2,4]. Through genetic analysis of yeast mutants, the CCR4 protein was identified as a positive regulator of glucose-repressible alcohol dehydrogenase (ADH2) genes [5], and the CCR4-associated factor (Caf) gene was identified as a catalytic subunit [2,6].
NOT1 proteins act as scaffolds for assembling the complex, providing binding sites for the NOT2-NOT3/5 subunits, CAF130, CAF40, and the catalytic module [7]. However, the precise function of NOT3 in yeast remains unclear. In humans, CNOT3 has been implicated in gene expression, regulation, mRNA surveillance, and export. This suggests that this gene plays a crucial role in post-transcriptional gene regulation and is essential for normal cell function and viability [8]. In most eukaryotes, except for yeast and Candida albicans, NOT3 genes are orthologous to yeast NOT3 and NOT5. In human fungal pathogens, such as Cryptococcus neoformans and Candida albicans, CaNOT5 is involved in morphogenesis, virulence, and cell wall structure [9,10].
Similarly, in plant pathogenic fungi, the CCR4-NOT complex has been found to play a crucial role in regulating virulence-related functions. In the case of Fusarium oxysporum f. sp. niveum, which causes Fusarium wilt in watermelon, the FoNOT2 gene plays diverse roles, including mycelial growth, conidiation, virulence, cell wall integrity, and the production of secondary metabolites [11]. Fusarium graminearum, the pathogen responsible for Fusarium head blight in wheat, has also been studied previously. FgNOT3 is involved in pleiotropic defects in vegetative growth, sexual reproduction, secondary metabolite production, and virulence [12]. However, to date, there have been no reports on the gene functional analysis of the NOT complex, including NOT3, in Magnaporthe oryzae, which causes rice blast disease.
Rice blast caused by M. oryzae is one of the most destructive diseases affecting cultivated rice worldwide. M. oryzae has been used as a model to understand fungal diseases [13]. To infect a host cell, M. oryzae undergoes several developmental processes, including attachment, conidial germination, appressorium formation, direct penetration using turgor pressure, and invasive growth into the rice host cell. As an initial step in invading a host plant, three-celled asexual conidia attach to the rice leaves after landing on the host surface [14]. When conidia are attached to a leaf with mucilage, they germinate within 3–4 h and form an appressorium within 4–6 h. A specialised structure called an appressorium directly penetrates the host cell with an enormous internal turgor pressure estimated to be as high as 8.0 MPa [15]. Several studies have reported that signalling-related pathways are involved in appressorium formation, including the cAMP-related pathway [16], MAP kinase pathway [17], and calcium/calmodulin-dependent signalling [18,19]. Genes related to signalling regulate the formation of appressoria, as well as penetration and invasive growth [15,20].
Invasive hyphae colonise host cells and express numerous genes associated with pathogenicity [21]. Subsequently, conidia of M. oryzae emerge on the infected rice cells to reinitiate the disease cycle. Conidia produced from conidiophores are controlled by several genes, including Com1, Con7, and Con1 [20,22,23]. Developmental processes, such as conidiation, conidial attachment, germination, appressorium formation, and pathogenicity, are regulated by genes, including transcription factors and protein complexes [24,25,26,27]. However, the pathosystem mechanisms between the host cells and M. oryzae need to be elucidated. Understanding the genetic mechanisms underlying host infections is a priority for the development of new disease-control strategies.
In a previous study, we identified a gene, MGG_08101, which was specifically abundant during appressorium development compared to mycelial growth in M. oryzae [28]. The gene encoding putative NOT3/5 was named MoNOT3/5. In this study, we conducted a functional analysis of and MoNOT3/5 in M. oryzae. To analyse the function of this gene under infection-related conditions, we generated MoNOT3/5 deletion mutants. Our data show that MoNOT3/5 is necessary for the overall fungal development of M. oryzae infection, particularly for pathogenicity and thermal stress response.
2. Materials and Methods
2.1. Fungal Isolate and Culture Conditions
M. oryzae wild-type (WT) strain KJ201 and all the generated transformants in this study were cultivated on V8-Juice agar medium (8% Campbell’s V8-Juice, 1.5% agar, pH adjusted to 7 using 0.1 N NaOH) or oatmeal agar medium (50 g oatmeal per litre) at 25°C under constant light. To extract genomic DNA and RNA, mycelia were isolated from fungal mycelia collected from 4-day-old cultures cultivated in liquid complete medium (CM, 0.6% yeast extract, 0.6% casamino acid, and 1% sucrose) for 5 days.
To assess mycelial growth, we used modified complete agar medium (mCM), minimal agar medium (MM), carbon-starved medium based on MM (MM-C), and nitrogen-starved medium based on MM (MM-N), as previously described [29]. Genomic DNA was isolated from mycelia collected from 4-day-old cultures grown in liquid CM.
CaCl2 (0.2 M), NaCl (0.5 M), MnCl2 (0.02 M), sodium dodecyl sulphate (SDS) (0.01%), Congo red (200 ppm), calcofluor white (200 ppm), and H2O2 (2 mM) were added to CM agar to determine their effects on fungal mycelial growth. Thermal stress response assays were performed at 25°C and 32°C on CM. The inoculum was prepared with 5-mm agar plugs from 7-day-old strains on CM, and the plugs were then transferred upside down onto each medium. Mycelial growth was measured after incubation for 9 days, and the percentage of inhibition was calculated by comparing the mycelial growth on the CM.
2.2. Computational and Phylogenetic Analyses
The MoNOT3/5 protein sequence (MGG_08101) and sequences of NOT3 various fungi were obtained from the Comparative Fungal Genomics Platform (http://www.cfgp.snu.ac.kr) [30]. Homology searches for DNA and protein sequences were performed using BLAST at the National Center for Biotechnology Information (NCBI). Sequence analysis and domain architecture of NOT3/5 were conducted using InterPro Scan, which is available at the European Bioinformatics Institute (http://www.ebi.ac.uk/interpro/). The amino acid sequence of NOT3 was aligned using ClustalW (http://align.genome.jp/). Phylogenetic trees were constructed using the maximum likelihood method in the MEGA program [31]. All sequence alignments were tested using the bootstrap method with 1,000 repetitions.
2.3. Vector Construction and Fungal Transformation
The construction of the MoNOT3/5 gene knockout mutant was performed using double-joint polymerase chain reaction (PCR) [32]. Fragments corresponding to 1.5 kb upstream and downstream of MoNOT3/5 were amplified using primers MoNOT3/5_5UF/5UR and MoNOT3/5_DF/DR, respectively. A 1.4-kb hph cassette was amplified with primers HygB_F and HygB_R (Table 1) from pBCATPH, which contains the hygromycin phosphotransferase gene (hph) under the control of the Aspergillus nidulans TrpC promoter [33]. The MoNOT3/5 gene disruption construct was amplified with primers nested MoNOT3/5_5UF/3DR using the fused products as a template and was directly transformed into the protoplasts of strain WT.
Protocols for protoplast preparation and fungal transformation were adapted from a previous study [34]. Hygromycin-resistant transformants were screened by PCR using primers MoNOT3/5_ORF_F/R (Table 1).
For complementation, a 4-kb fragment containing the native promoter and ORF of MoNOT3/5 was amplified with primers MoNOT3/5_5UF and 3DR (Table 1) from genomic DNA of the WT. This fragment was co-transformed into protoplasts from the MoNOT3/5 gene deletion mutant using pll99, which contains a gene that confers resistance to geneticin. Transformants were selected based on their ability to grow in the presence of geneticin (800 μg/mL).
2.4. Nucleic acid Manipulation and Southern Blot Analysis
Genomic DNA was isolated from using the NucleoSpin® Plant II Kit (MachereyNagel, Düren, Germany). The DNA concentration was estimated using a NanoDrop ND-1000 spectrophotometer (NanoDrop Technologies, Inc. Wilmington, NC, USA). Putative MoNOT3/5 gene deletion mutants were confirmed by Southern blot analysis. For Southern blotting, genomic DNA (approximately 5 μg) was digested with EcoRI and fractionated on a 0.7% agarose gel at 40 V for 4 h in 1% Tris-acetate-EDTA buffer. The fractionated DNA was transferred onto Hydond N+ nylon membranes (Amersham International, Little Chalfont, England) using 10× SSC (1× SSC comprised 0.15 M NaCl plus 0.015 M sodium citrate). The probe, designed from the 3’-flanking region, was labelled with alkaline phosphate using AlkPhos Direct Labelling Reagents (Amersham Biosciences, UK). Hybridisation, washing, and detection were performed according to the instruction manual of the Alkphos Direct Labelling and Detection System using CDP Star (Amersham Pharmacia).
Briefly, hybridisation was performed at 55°C in Gene Images AlkPhos Direct hybridisation buffer, including 0.5 M NaCl and 4% blocking reagent. After hybridisation, the membrane was washed twice in primary wash buffer at 55°C for 10 min and then washed twice in secondary wash buffer at room temperature for 5 min. Target DNA was detected using CDP star reagent (Amersham International plc, Buckinghamshire, UK) according to the manufacturer’s instructions.
2.5. Conidiation, Conidial Morphology, Conidial Germination, and Appressorium Formation
To measure conidiation and conidial morphology, conidia were collected from 7-day sporulating cultures on V8-Juice agar media by gently rubbing the mycelia with sterilised distilled water, followed by filtration through Miracloth (Calbiochem, San Diego, USA). The number of conidial suspensions was counted using a haemocytometer (BRAND counting chamber, BLAUBRAND, Brand GMBH, Germany). Conidial size was measured using a microscope, and the conidia were stained with calcofluor white and KOH to observe conidial morphology.
Conidial germination and appressorium formation were measured using hydrophobic coverslips. A total of 50 μL of a conidial suspension (5 × 104 conidia/mL) was placed on a coverslip and incubated in the moist box at 25°C for 8 h and 24 h. The percentage of conidia with germ tubes and appressoria was determined by microscopic examination of at least 100 conidia per replicate. All experiments were performed in triplicate.
2.6. Plant Infection Assays
For the pathogenicity assay, conidia were harvested from 7-day-old cultures on V8 agar plates, and 10 mL of a conidial suspension (1 × 105 conidia/mL) containing 250 ppm Tween 20 was sprayed onto 4-week-old susceptible rice seedlings (Oryza sativa cv. Nakdongbyeo) at the three-to-four-leaf stage. For the detached leaf assay, 50 μL of conidial suspension (2 × 104 conidia/mL) was dropped on three points per leaf of 4-week-old rice plans. For the infiltration infection assay, 100 μL of conidial suspension (2 × 104 conidia/mL) was injected into three points per leaf of 4-week-old rice plants. All inoculated plants were kept in a dew chamber at 25°C for 24 h in the dark and then moved to a growth chamber with a photoperiod of 16 h under fluorescent lights. Disease severity was measured 5 days after inoculation.
Plant penetration assays were performed using onion epidermis and rice sheaths, as previously described [35]. Briefly, 4-week-old rice sheaths of cv. Nakdongbyeo and conidia (1 × 104 conidia/mL) from 7-day-old cultures on V8-Juice agar media were used for onion epidermis and rice sheaths. The inoculated plants were maintained in a moist box at 25°C for 36 h in the dark. An infection assay using onion epidermis was performed using fresh onion epidermis. All experiments were performed in triplicate.
2.7. RNA Isolation, Quantitative Real-Time (qRT)-PCR, and Gene Expression Analysis
Total RNA was prepared using the Easy-Spin Total RNA Extraction Kit (iNtRON Biotechnology, Sungnam, Korea) following the manufacturer’s instructions. Total RNA was extracted from the fungus at several developmental stages under stress conditions and used as a template for reverse transcription (RT)-PCR. Total RNA was extracted from mycelia in liquid mCM for expression analysis of the COS1, Com1, Con7, and MoHox2 genes.
To obtain samples during conidiogenesis, WT and gene-deletion mutants were inoculated into liquid mCM and incubated at 25°C on a 120 rpm orbital shaker for 3 days. The resulting mycelia were fragmented using a spatula and filtered through a single-layer cheesecloth. Mycelia were collected using one layer of Miracloth (Calbiochem, California, USA) in 2 mL of sterilised distilled water. Then, 400 μL of the mycelial suspension was spread onto individual 0.45-μm pore cellulose nitrate membrane filters (Whatman, Maidstone, England) placed on V8-Juice agar plates using a loop. The plates were dried and incubated at 25°C with constant light. After 12 h of inoculation, whole mycelia on cellulose nitrate membrane filters were collected using a disposable scraper (iNtRON Biotechnology, Seoul, Korea) [27].
qRT-PCR reactions were performed using a 96-well reaction plate and a CFX96 Touch™ Real-Time PCR Detection System (Bio-Rad Laboratories, Hercules, CA, USA). Each well contained 5 μL of SYBR Green I Master mix (Bio-Rad Laboratories, Hercules, CA, USA), 2 μL of cDNA (12.5 ng), and 15 pmol of each primer. The thermal cycling conditions were 10 min at 94°C, followed by 40 cycles of 15 s at 94°C and 1 min at 60°C. All amplification curves were analysed with a normalised reporter threshold of 17 to obtain the threshold cycle (Ct) values.
The primer sequences used for each gene are listed in Table 1. To compare the relative abundance of the target gene transcripts, the average Ct value was normalised to that of -tubulin (MGG00604) for each of the treated samples as 2-Ct, where Ct = (Ct, target gene - Ct, -tubulin). Fold changes during fungal development and infectious growth in liquid CM were calculated as 2-Ct, where Ct = (Ct, target gene - Ct, -tubulin)test condition - (Ct, WT - Ct, -tubulin)CM [10]. qRT-PCR was performed using three independent tissue pools with two sets of experimental replicates.
3. Results
3.1. Molecular Characterisation of MoNOT3/5
The CCR4-NOT complex consists of CCR4 and NOT proteins in eukaryotes, except for Saccharomycetaceae and Candida albicans, which contain additional Caf protein subunits. Members of NOT3/5 possess conserved components, including the CCR4-NOT complex, N-terminal (IPR007207), NOT2, NOT3, and NOT5 (IPR007282). In a previous study, MGG_08101 was upregulated during appressorium development. This gene was identified as an ortholog of NOT3/5 in Saccharomyces cerevisiae using a BLAST search. Thus, we named this gene MoNOT3/5 (Magnaporthe oryzae NOT3/5). The predicted amino acid sequences of MoNOT3/5 exhibited high levels of similarity to filamentous ascomycetes, showing high homology with EWZ90470 of Fusarium oxysporum f. sp. lycopersici (65.87%), XP_011394869 of Neurospora crassa (63.24%), XP_011319317 of F. graminearum (61.35%), and XP_024550716 of Botrytis cinerea (57.06%). Therefore, they formed a phylogenetic clade (Figure 1a). Phylogenetic analyses of NOT3/5 homologues showed that they were conserved in eukaryotes and clustered into a group related to fungi, animals, and plants, with the exception of Saccharomycetaceae (Figure 1a).
To investigate the interaction of MoNOT3/5 with M. oryzae, 48 genes that predicted the first neighbours of MoNOT3/5 were listed in a diverse database. MoNOT3/5 interacts with many genes involved in transport, mycelium development, metabolism, signalling, transcription, protein localisation and modification, translation, DNA replication and chromatin-related processes, mRNA processing, exo- and endocytosis, and pathogenesis. The results showed that MoNOT3/5 genes were most closely related to MGG_06958, MGG_01228, MGG_07928, and MGG_06044 (Figure 1b).
qRT-PCR was used to examine the expression pattern of MoNOT3/5 from the mycelia to the infection stages. The MoNOT3/5 gene was constitutively expressed at all stages (Figure 2c). This pattern of MoNOT3/5 expression appears to correlate with its functional role in fungal development and pathogenicity.
3.2. Targeted Replacement of MoNOT3/5
Deletion mutants were generated via homologous recombination using a resistance marker cassette (Figure 2a). The protoplasts were transformed using genetic constructs. Two Δmonot3/5 deletion mutants were identified based on Southern blot analysis. Genomic DNA samples were digested with EcoRI and probed with the 1.5 kb 3’-flanking regions (Figure 2a). The 3.9-kb band was detected in Δmonot3/5-1 and Δmonot3/5-2, but not in WT KJ201 (Figure 2b). The MoNOT3/5 transcript was detected in the RT-PCR and qRT-PCR analyses of the WT and complemented strains MoNOT3/5c-1 and MoNOT3/5c-2 but was not detected in Δmonot3/5-1 and Δmonot3/5-2 (Figure 2c).
3.3. MoNOT3/5 is Required for Vegetative Growth and Conidiogenesis
To characterise the functional role of MoNOT3/5, mycelial growth of the WT, mutant strain (MoNOT3/5), and the complement strain (MoNOT3/5c) was compared on CM, MM, MM-N, and MM-C. Mycelial growth rates were significantly reduced in the mutants monot3/5-1 and monot3/5-2 compared to the WT on all four media. The complements (MoNOT3/5c-1 and MoNOT3/5c-2) were completely recovered, as shown in Figure 3.
The mutant showed a significant (>95%) reduction in conidiation compared to the WT on oatmeal agar medium (Figure 4a). Microscopic observations indicated that the WT strain formed dense conidiophores bearing conidia in a sympodial pattern, whereas the Δmonot3/5-1 and Δmonot3/5-2 mutants produced rare conidiophores, each consisting of a single conidium. These results indicate that MoNOT3/5 is required for both conidiophore and conidial development. Both Δmonot3/5-1 and Δmonot3/5-2 mutants showed a significant reduction in the production of conidia compared with the WT. However, conidiation fully recovered to the WT levels after complementation with MoNOT3/5 in the MoNOT3/5c-1 and MoNOT3/5c-2 strains (Figure 4b).
Microscopic observations revealed that the gene deletion mutants (Δmonot3/5-1 and Δmonot3/5-2) produced conidia with abnormal shapes and septation defects (Figure 4c). While WT conidia were typically three-celled, the Δmonot3/5-1 and Δmonot3/5-2 strains produced more than 50% and 45% of the one-celled and two-celled conidia, respectively (Figure 4d). Furthermore, quantification of the abnormal shape defect revealed that the length and width of conidia in Δmonot3/5-1 and Δmonot3/5-2 strains were significantly smaller than those in the WT and complements (Figure 4e). These data indicate that MoNOT3/5 is required for proper asexual reproduction in M. oryzae.
3.4. monot3/5 is Required for Conidial Germination and Appressorium Formation
Conidial germination and appressorium formation decreased in both Δmonot3/5-1 and Δmonot3/5-2 mutants (Figure 5a–c). Approximately 90% of WT conidia germinated at 8 h and 24 h following inoculation on a coverslip, whereas only 30% and less than 50% of Δmonot3/5-1 and Δmonot3/5-2 mutant conidia germinated after 8 h and 24 h of incubation, respectively. Conidial germination of the mutant did not fully recover even after culturing on a coverslip for a longer period (Figure 5a,b).
Over 95% of the germinated conidia formed appressoria at 8 h and 24 h in the WT and complemented strains, and the majority of germinated conidia in Δmonot3/5-1 and Δmonot3/5-2 failed to form appressoria (Figure 5a). After 24 h, the germ tubes grew longer and did not form appressoria in either the Δmonot3/5-1 or Δmonot3/5-2 mutants (Figure 5a,c). The complemented strains fully recovered germination and appressorium formation, similar to the WT strain.
3.5. MoNOT3/5 is Required for Penetration and Virulence in M. oryzae
The results of the spray inoculation test showed that WT plants exhibited typical diamond-shaped lesions. In contrast, the Δmonot3/5-1 and Δmonot3/5-2 mutants exhibited complete failure of disease development. Complements containing the monot3/5 gene were fully recovered (Figure 6a, left). The detached leaf test also produced results identical to those of spray inoculation (Figure 6a, middle). To gain a deeper understanding of this mechanism, we investigated invasive growth by injecting a conidial suspension into rice seedlings via infiltration. The mutants showed no blast symptoms (Figure 6a, right).
To assess the penetration ability of MoNOT3/5 in plants, a suspension of conidia was applied to the onion epidermis (Figure 6b) and inoculated onto rice sheath cells (Figure 6c). The mutants showed no penetration or invasive hyphae on either non-host or host plants (Figure 6b,c). The complemented strains showed fully restored pathogenicity, invasive growth, and penetration, similar to the WT.
3.6. MoNOT3/5 is Important for Stress Tolerance
In yeast, mutants lacking NOT are defective under thermal and cell wall stress conditions. To investigate the role of MoNOT3/5 in various stress-resistance mechanisms, we measured the inhibition rates of mycelial growth compared to growth on CM. Nine days after inoculation, the mycelial growth of Δmonot3/5-1 and Δmonot3/5-2 mutants was significantly reduced compared to that of the WT on CM containing Congo red and SDS. The mutant was sensitive to cell wall-damaging agents in the presence of Congo red and SDS.
To examine the role of MoNOT3/5 under thermal stress conditions, we analysed the mycelial growth at an optimal temperature of 25°C and a maximum temperature of 32°C. The WT grew normally at 32°C, while both Δmonot3/5-1 and Δmonot3/5-2 mutants did not. The mycelial growth of Δmonot3/5-1 and Δmonot3/5-2 mutants exhibited a significant decrease compared to the WT at 32°C. These results indicate that Δmonot3/5 is involved in thermal stress resistance (Figure 7 and Table 2).
3.7. Analysis of MoNOT3/5 Gene Expression Patterns
To identify MoNOT3/5 genes that potentially control conidiation, we analysed the expression patterns of conidia-related genes compared to the WT during conidiation. The expression of Con1-related conidiophore stalk and Con7-related conidial morphology was downregulated during conidiation (Figure 8a). However, transcription factor genes related to conidia were not affected by MoNOT3/5. These results suggest that conidiation and conidial morphology are affected by MoNOT3/5. As mentioned earlier, we found that MoNOT3/5 interacts with numerous genes involved in all stages.
To identify protein interactions involving Δmonot3/5, we quantified the transcripts of four genes associated with ubiquitin proteins. Polyubiquitin expression increased during mycelial growth (Figure 8b). These results suggest that MoNOT3/5 regulates ubiquitination.
4. Discussion
CCR4-NOT complexes are essential proteins that regulate and coordinate diverse biological processes. While numerous new studies have been published on the functional analyses of CCR4-NOT complexes in yeast, humans, human pathogenic fungi, and some phytopathogenic fungi, the characterisation of CCR4-NOT complexes in phytopathogenic fungi is still required [2,6]. In this study, we characterised the crucial regulatory roles of MoNOT3/5 in the developmental stages of M. oryzae. The absence of MoNOT3/5 gene led to multiple defects in mycelial growth, conidiation, conidial morphology, germination, appressorium formation, cell wall stress response, and virulence.
The CCR4-NOT complexes are well conserved in M. oryzae (Figure 1a). The MoNOT3/5 gene is homologous to the yeast genes ScNOT3 and ScNOT5, human gene CNOT3, C. albicans gene CaNOT5, and F. graminearum gene FgNOT3 [12]. In C. albicans, CaNOT5 is involved in spore attachment and pathogenicity [9,10]. In F. graminearum, FgNOT3 is involved in mycelial growth, conidiation, sexual reproduction, and the production of secondary metabolites [12]. In the present study, we discovered that MoNOT3/5 is involved in infection-related cell development, including conidiation, conidial germination, appressorium formation, invasive growth, spore germination, and appressorium formation. These results indicate that MoNOT3/5 may be involved in all stages of fungal development.
Interestingly, we found that MoNOT3/5 contributes to conidiation-related morphogenesis, including the development of conidiophore stalks, conidial morphology, and conidial production. Except for germination, these phenotypes were similar to those observed in other fungi [12]. The deletion of FgNOT3 results in defective conidial morphology and germination [12]. However, the germination of ΔFgnot3 fully recovered over time, whereas that of Δmonot3/5 did not, even with additional time. These differences may indicate species divergence among filamentous fungi. Because delayed mycelial growth, germination, and abnormal septation are related to cell cycle progression, the observed phenotypes in the mutants suggest that MoNOT3/5 is involved in regulating the cell cycle.
Previous studies have reported that the mutants of C. albicans and F. graminearum were deficient in virulence. The deletion of CaNOT5 does not induce a switch to a hyphal transition related to virulence [9]. In F. graminearum, deletion of FoNOT3 resulted in an increase in secondary metabolites but did not lead to disease development [11]. The mutants were unable to form mats or aspersoria-like structures to penetrate the host cell wall. Unlike F. graminearum, in this study, we found that MoNOT3/5 was associated with appressorium formation and invasive hyphae, demonstrating its crucial role in penetration. This is the first report indicating that MoNOT3/5 is involved in the formation of the appressorium and invasive hyphae for penetration.
In yeast, double knockout mutants of NOT1, NOT2, and NOT3/NOT5 are sensitive to heat stress [36]. In F. graminearum, NOT module mutants exhibit sensitivity to heat stress [12]. CaNOT5 in C. albicans was sensitive to calcofluor white, whereas FoNOT2 was sensitive to calcofluor white, Congo red, and oxidative stress. Δmonot3/5 mutants were sensitive to heat stress, SDS, and Congo red. However, there was no difference in the other cell wall stress conditions. These differences may indicate species divergence among filamentous fungi. Other modules appear to play a more significant role in cell wall stress conditions in fungi than Δmonot3/5.
To understand these phenotypes, we identified the proteins that interact with MoNOT3/5. These results suggest that MoNOT3/5 interacts with ubiquitin, which plays an important role in various cellular processes and host–pathogen interactions [37]. The levels of polyubiquitin transcripts increased in the mutant. Mono-ubiquitination may negatively affect polyubiquitin levels, and our transcript analyses demonstrated interactions among these proteins. In M. oryzae, Ssb1, Ssz, Zuo1, and Mkk1, which are associated with heat shock proteins, are involved in growth, pathogenicity, cell wall integrity, and conidiation [38]. The loss of Ssb1 resulted in protein misfolding. When comparing the phenotype of Δmonot3/5 with ΔMoSsb1, it is necessary to confirm the interactions with heat shock proteins. Monosodium triphosphate may affect the expression of heat shock proteins, which are crucial regulators.
5. Conclusion
In our previous study, we analyzed the function of the MoNOT3/5 (MGG_08101) genes, which were up-regulated during appressorium formation in M. oryzae. For functional analysis, gene deletion mutants were generated and phenotype analysis was conducted. As a result, the mycelial growth was retarded compared to WT. In particular, it was found to be involved in various functions related to conidiation, including conidial morphology and conidial germination. Furthermore, it appears to have played a crucial role in the formation of appressoria and invasive growth, which are essential for invading the host plant. This suggests that MoNOT3/5 may be specifically involved in the process of disease development. Since this protein was found to be involved in heat shock in F. graminearum, it was also found to be involved in various stress conditions in M. oryzae. Through transcriptional analysis in WT and Δmonot3/5 mutant strains, the results indicate that the MoNOT3/5 gene regulates COS1 and COM1, which are involved in conidiation. Taken together, MoNOT3/5 is presumed to be a gene involved in various aspects of the disease development process of M. oryzae, as well as necessary for temperature regulation and ionic stress.
Supplementary Materials
Not applicable.
Author Contributions
Conceptualization, Y.K., K.-T.K. and S.-Y.P.; methodology, Y.K., Y.L. and M.-H.J.; validation, Y.K., M.J., S. A. and S.-Y.P.; formal analysis, Y.K., Y.L. and M.-H.J.; investigation, Y.K., M.J., S. A., Y.L., E.D.C., M.-H.J. and K.-T.K.; resources, S.-Y.P.; data curation, Y.K. and S.-Y.P.; writing—original draft preparation, Y.K. and S.-Y.P.; writing—review and editing, Y.K. and S.-Y.P.; visualization, Y.K., K.-T.K. and S.-Y.P.; supervision, S.-Y.P.; project administration, S.-Y.P.; funding acquisition, S.-Y.P. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by a research promotion program of SCNU.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Iyer, V.; Struhl, K. Mechanism of differential utilization of the his3 TR and TC TATA elements. Mol. Cell Biol. 1995, 15, 7059–7066. [Google Scholar] [CrossRef]
- Collart, M.A.; Panasenko, O.O. The Ccr4–not complex. Gene 2012, 492, 42–53. [Google Scholar] [CrossRef]
- Collart, M.A.; Struhl, K. NOT1 (CDC39), NOT2 (CDC36), NOT3, and NOT4 encode a global-negative regulator of transcription that differentially affects TATA-element utilization. Genes Dev. 1994, 8, 525–537. [Google Scholar] [CrossRef]
- Miller, J.E.; Reese, J.C. Ccr4-Not complex: the control freak of eukaryotic cells. Crit. Rev. Biochem. Mol. Biol. 2012, 47, 315–333. [Google Scholar] [CrossRef]
- Malvar, T.; Biron, R.W.; Kaback, D.B.; Denis, C.L. The CCR4 protein from Saccharomyces cerevisiae contains a leucine-rich repeat region which is required for its control of ADH2 gene expression. Genetics 1992, 132, 951–962. [Google Scholar] [CrossRef]
- Collart, M.A. The Ccr4-Not complex is a key regulator of eukaryotic gene expression. Wiley Interdiscip. Rev. RNA 2016, 7, 438–454. [Google Scholar] [CrossRef]
- Lau, N.-C.; Kolkman, A.; van Schaik, F.M.A.; Mulder, K.W.; Pijnappel, W.W.M.P.; Heck, A.J.R.; Timmers, H.T.M. Human Ccr4-Not complexes!contain!variable deadenylase subunits. Biochem. J. 2009, 422, 443–453. [Google Scholar] [CrossRef]
- Collart, M.A.; Panasenko, O.O.; Nikolaev, S.I. The Not3/5 subunit of the Ccr4-Not complex: a central regulator of gene expression that integrates signals between the cytoplasm and the nucleus in eukaryotic cells. Cell Signal. 2013, 25, 743–751. [Google Scholar] [CrossRef]
- Cheng, S.; Clancy, C.J.; Checkley, M.A.; Handfield, M.; Hillman, J.D.; Progulske-Fox, A.; Lewin, A.S.; Fidel, P.L.; Nguyen, M.H. Identification of Candida albicans genes induced during thrush offers insight into pathogenesis. Mol. Microbiol. 2003, 48, 1275–1288. [Google Scholar] [CrossRef]
- Panepinto, J.C.; Heinz, E.; Traven, A. The cellular roles of Ccr4-NOT in model and pathogenic fungi–implications for fungal virulence. Frontiers in Genetics 2013, 4, 302. [Google Scholar] [CrossRef]
- Dai, Y.; Cao, Z.; Huang, L.; Liu, S.; Shen, Z.; Wang, Y.; Wang, H.; Zhang, H.; Li, D.; Song, F. CCR4-Not complex subunit Not2 plays critical roles in vegetative growth, conidiation and virulence in watermelon Fusarium wilt pathogen Fusarium oxysporum f. sp. niveum. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef]
- Bui, D.-C.; Son, H.; Shin, J.Y.; Kim, J.-C.; Kim, H.; Choi, G.J.; Lee, Y.-W. The FgNot3 subunit of the Ccr4-Not complex regulates vegetative growth, sporulation, and virulence in Fusarium graminearum. PloS One 2016, 11, e0147481. [Google Scholar] [CrossRef]
- Dean, R.A.; Talbot, N.J.; Ebbole, D.J.; Farman, M.L.; Mitchell, T.K.; Orbach, M.J.; Thon, M.; Kulkarni, R.; Xu, J.-R.; Pan, H. The genome sequence of the rice blast fungus Magnaporthe grisea. Nat. 2005, 434, 980–986. [Google Scholar] [CrossRef]
- Talbot, N.J. Having a blast: exploring the pathogenicity of Magnaporthe grisea. Trends Microbiol. 1995, 3, 9–16. [Google Scholar] [CrossRef]
- Thines, E.; Weber, R.W.; Talbot, N.J. MAP kinase and protein kinase A–dependent mobilization of triacylglycerol and glycogen during appressorium turgor generation by Magnaporthe grisea. Plant Cell 2000, 12, 1703–1718. [Google Scholar]
- Mitchell, T.K.; Dean, R.A. The cAMP-dependent protein kinase catalytic subunit is required for appressorium formation and pathogenesis by the rice blast pathogen Magnaporthe grisea. Plant Cell 1995, 7, 1869–1878. [Google Scholar] [CrossRef]
- Xu, J.R.; Hamer, J.E. MAP kinase and cAMP signaling regulate infection structure formation and pathogenic growth in the rice blast fungus Magnaporthe grisea. Genes Dev. 1996, 10, 2696–2706. [Google Scholar] [CrossRef]
- Choi, J.; Kim, Y.; Kim, S.; Park, J.; Lee, Y.H. MoCRZ1, a gene encoding a calcineurin-responsive transcription factor, regulates fungal growth and pathogenicity of Magnaporthe oryzae. Fungal. Genet Biol. 2009, 46, 243–254. [Google Scholar] [CrossRef]
- Kim, S.; Hu, J.; Oh, Y.; Park, J.; Choi, J.; Lee, Y.H.; Dean, R.A.; Mitchell, T.K. Combining ChIP-chip and expression profiling to model the MoCRZ1 mediated circuit for Ca/calcineurin signaling in the rice blast fungus. PLoS Pathog. 2010, 6, e1000909. [Google Scholar] [CrossRef]
- Li, G.; Zhou, X.; Xu, J.-R. Genetic control of infection-related development in Magnaporthe oryzae. Curr. Opin. Microbiol. 2012, 15, 678–684. [Google Scholar] [CrossRef]
- Ebbole, D.J. Magnaporthe as a model for understanding host-pathogen interactions. Annu. Rev. Phytopathol. 2007, 45, 437–456. [Google Scholar] [CrossRef]
- Odenbach, D.; Breth, B.; Thines, E.; Weber, R.W.; Anke, H.; Foster, A.J. The transcription factor Con7p is a central regulator of infection-related morphogenesis in the rice blast fungus Magnaporthe grisea. Mol. Microbiol. 2007, 64, 293–307. [Google Scholar] [CrossRef]
- Yang, J.; Zhao, X.; Sun, J.; Kang, Z.; Ding, S.; Xu, J.-R.; Peng, Y.-L. A novel protein Com1 is required for normal conidium morphology and full virulence in Magnaporthe oryzae. Mol. Plant-Microbe Interact. 2010, 23, 112–123. [Google Scholar] [CrossRef]
- Cao, Y.; Chen, J.; Xie, X.; Liu, S.; Jiang, Y.; Pei, M.; Wu, Q.; Qi, P.; Du, L.; Peng, B.; et al. Characterization of two infection-induced transcription factors of Magnaporthe oryzae reveals their roles in regulating early infection and effector expression. Mol. Plant Pathol. 2022, 23, 1200–1213. [Google Scholar] [CrossRef]
- Kim, S.; Park, S.Y.; Kim, K.S.; Rho, H.S.; Chi, M.H.; Choi, J.; Park, J.; Kong, S.; Park, J.; Goh, J.; et al. Homeobox transcription factors are required for conidiation and appressorium development in the rice blast fungus Magnaporthe oryzae. PLoS Genet 2009, 5, e1000757. [Google Scholar] [CrossRef]
- Matheis, S.; Yemelin, A.; Scheps, D.; Andresen, K.; Jacob, S.; Thines, E.; Foster, A.J. Functions of the Magnaporthe oryzae Flb3p and Flb4p transcription factors in the regulation of conidiation. Microbiol. Res. 2017, 196, 106–117. [Google Scholar] [CrossRef]
- Park, S.-Y.; Choi, J.; Lim, S.-E.; Lee, G.-W.; Park, J.; Kim, Y.; Kong, S.; Kim, S.R.; Rho, H.-S.; Jeon, J. Global expression profiling of transcription factor genes provides new insights into pathogenicity and stress responses in the rice blast fungus. PLoS Pathog. 2013, 9, e1003350. [Google Scholar] [CrossRef]
- Chi, M.-H.; Park, S.-Y. Identification of differentially expressed genes by cDNA-AFLP in Magnaporthe oryzae. Res. Plant Dis. 2019, 25, 205–212. [Google Scholar]
- Talbot, N.J.; Ebbole, D.J.; Hamer, J.E. Identification and characterization of MPG1, a gene involved in pathogenicity from the rice blast fungus Magnaporthe grisea. Plant Cell 1993, 5, 1575–1590. [Google Scholar]
- Choi, J.; Cheong, K.; Jung, K.; Jeon, J.; Lee, G.-W.; Kang, S.; Kim, S.; Lee, Y.-W.; Lee, Y.-H. CFGP 2.0: a versatile web-based platform for supporting comparative and evolutionary genomics of fungi and Oomycetes. Nucleic Acids Res. 2013, 41, D714–D719. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.-H.; Hamari, Z.; Han, K.-H.; Seo, J.-A.; Reyes-Domínguez, Y.; Scazzocchio, C. Double-joint PCR: a PCR-based molecular tool for gene manipulations in filamentous fungi. Fungal. Genet. Biol. 2004, 41, 973–981. [Google Scholar] [CrossRef] [PubMed]
- Gritz, L.; Davies, J. Plasmid-encoded hygromycin B resistance: the sequence of hygromycin B phosphotransferase gene and its expression in Escherichia coli and Saccharomyces cerevisiae. Gene 1983, 25, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Leung, H.; Lehtinen, U.; Karjalainen, R.; Skinner, D.; Tooley, P.; Leong, S.; Ellingboe, A. Transformation of the rice blast fungus Magnaporthe grisea to hygromycin B resistance. Curr. Genet. 1990, 17, 409–411. [Google Scholar] [CrossRef]
- Koga, H.; Dohi, K.; Nakayachi, O.; Mori, M. A novel inoculation method of Magnaporthe grisea for cytological observation of the infection process using intact leaf sheaths of rice plants. Physiol. Mol. Plant Pathol. 2004, 64, 67–72. [Google Scholar] [CrossRef]
- Oberholzer, U.; Collart, M. Characterization of NOT5 that encodes a new component of the Not protein complex. Gene 1998, 207, 61–69. [Google Scholar] [CrossRef]
- Oh, Y.; Franck, W.L.; Han, S.-O.; Shows, A.; Gokce, E.; Muddiman, D.C.; Dean, R.A. Polyubiquitin is required for growth, development and pathogenicity in the rice blast fungus Magnaporthe oryzae. PloS One 2012, 7, e42868. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Liu, M.; Liu, X.; Yin, Z.; Sun, Y.; Zhang, H.; Zheng, X.; Wang, P.; Zhang, Z. Heat-shock proteins MoSsb1, MoSsz1, and MoZuo1 attenuate MoMkk1-mediated cell-wall integrity signaling and are important for growth and pathogenicity of Magnaporthe oryzae. Mol. Plant-Microbe Interact. 2018, 31, 1211–1221. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Phylogenetic trees, putative protein–protein interaction (PPI) network, and the expression patterns of MoNOT3/5. (a) Phylogenetic analysis of NOT3/5 from Magnaporthe oryzae with eukaryotes. A maximum likelihood tree was constructed based on the amino acid sequence using MEGA X. All sequence alignments were tested with a bootstrap method using 1,000 repetitions. The domain architecture of NOT3/5s was determined by InterProScan. The scale bar shows the number of amino acid differences per site. Subclades containing MoNOT3/5 are shaded, in which MoNOT3/5 and characterised genes from other fungi are shown in bold in red or black, respectively. The text with numbers in parentheses represents the National Center for Biotechnology Information (NCBI) accession numbers of the protein sequences. (b) The PPI network of MoNOT3/5. Only the first neighbours of MoNOT3/5 are shown. The circle size of the nodes represents the number of edges (interactions) connected to each node. The putative function of each node represented by the GO term is colour coded. (c) The expression patterns of MoNOT3/5 and its PPI neighbours. The expression patterns from mycelia to the infection stages of MoNOT3/5 (red line) and its 47 neighbours are shown.
Figure 1.
Phylogenetic trees, putative protein–protein interaction (PPI) network, and the expression patterns of MoNOT3/5. (a) Phylogenetic analysis of NOT3/5 from Magnaporthe oryzae with eukaryotes. A maximum likelihood tree was constructed based on the amino acid sequence using MEGA X. All sequence alignments were tested with a bootstrap method using 1,000 repetitions. The domain architecture of NOT3/5s was determined by InterProScan. The scale bar shows the number of amino acid differences per site. Subclades containing MoNOT3/5 are shaded, in which MoNOT3/5 and characterised genes from other fungi are shown in bold in red or black, respectively. The text with numbers in parentheses represents the National Center for Biotechnology Information (NCBI) accession numbers of the protein sequences. (b) The PPI network of MoNOT3/5. Only the first neighbours of MoNOT3/5 are shown. The circle size of the nodes represents the number of edges (interactions) connected to each node. The putative function of each node represented by the GO term is colour coded. (c) The expression patterns of MoNOT3/5 and its PPI neighbours. The expression patterns from mycelia to the infection stages of MoNOT3/5 (red line) and its 47 neighbours are shown.
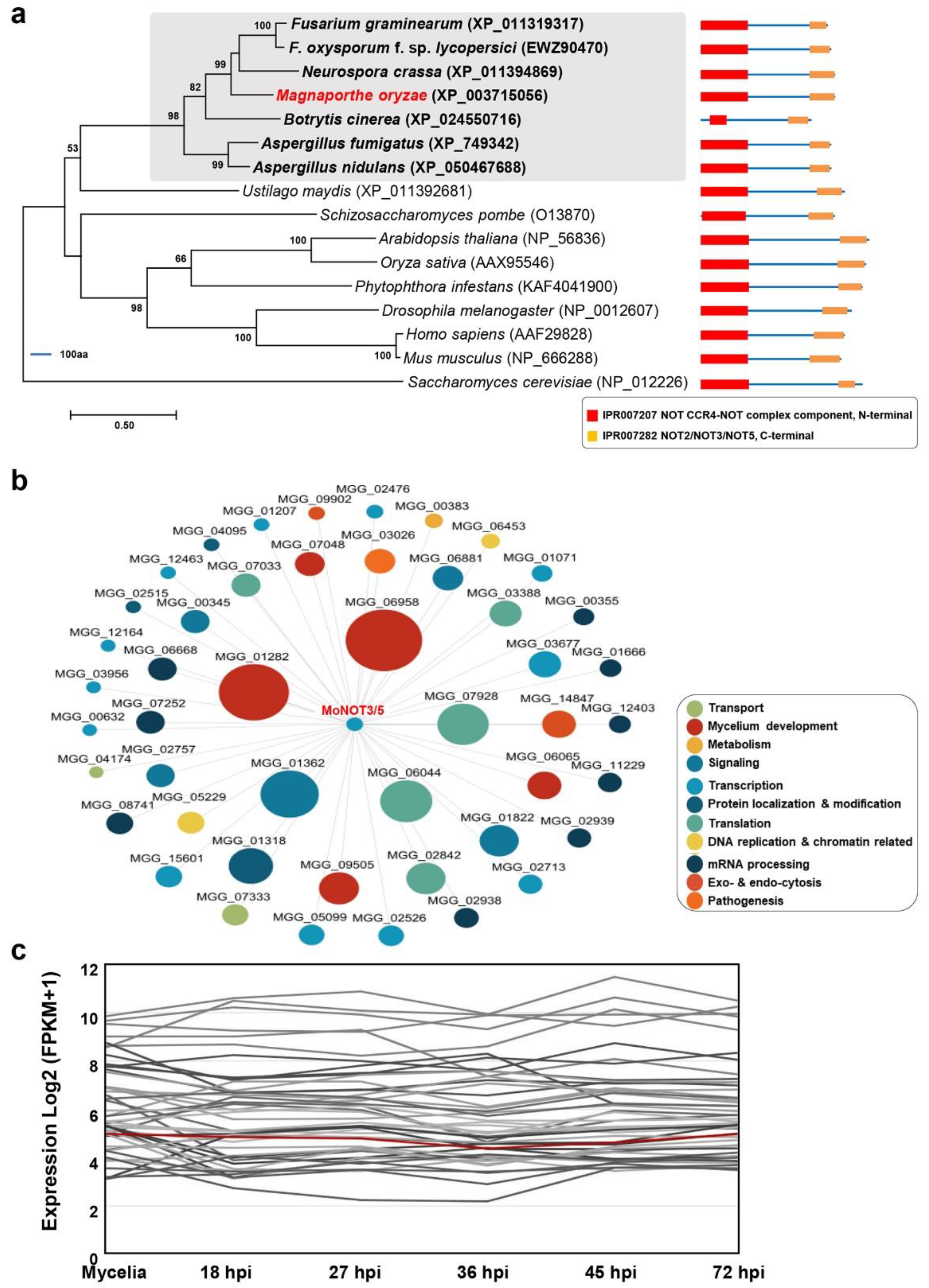
Figure 2.
Generation of gene deletion mutants for the MoNOT3/5 gene. (a) The Δmonot3/5 knockout fragment was constructed by PCR amplification. The 1.5 kb upstream and downstream flanking sequences were amplified with primers MoNOT3/5_UF/UR and MoNOT3/5_DF/DR, respectively, and ligated with the hph gene cassette. (b) Southern blot analysis of EcoRI-digested genomic DNA of the wild-type (WT) and the Δmonot3/5 mutants, Δmonot3/5-1 and Δmonot3/5-2. The blot was hybridized with the probe shown in Figure 2a. (c) Expression analysis of ΜοΝOΤ3/5 in the WT strain, Δmonat3/5 mutants, and the complemented transformants, MoNOT3/5c-1 and MoNOT3/5c-2. Primers, MoNOT3/5c_qRT_F/R (Table 1), were used to amplify the Δmonot3/5 transcript using RT-PCR.
Figure 2.
Generation of gene deletion mutants for the MoNOT3/5 gene. (a) The Δmonot3/5 knockout fragment was constructed by PCR amplification. The 1.5 kb upstream and downstream flanking sequences were amplified with primers MoNOT3/5_UF/UR and MoNOT3/5_DF/DR, respectively, and ligated with the hph gene cassette. (b) Southern blot analysis of EcoRI-digested genomic DNA of the wild-type (WT) and the Δmonot3/5 mutants, Δmonot3/5-1 and Δmonot3/5-2. The blot was hybridized with the probe shown in Figure 2a. (c) Expression analysis of ΜοΝOΤ3/5 in the WT strain, Δmonat3/5 mutants, and the complemented transformants, MoNOT3/5c-1 and MoNOT3/5c-2. Primers, MoNOT3/5c_qRT_F/R (Table 1), were used to amplify the Δmonot3/5 transcript using RT-PCR.
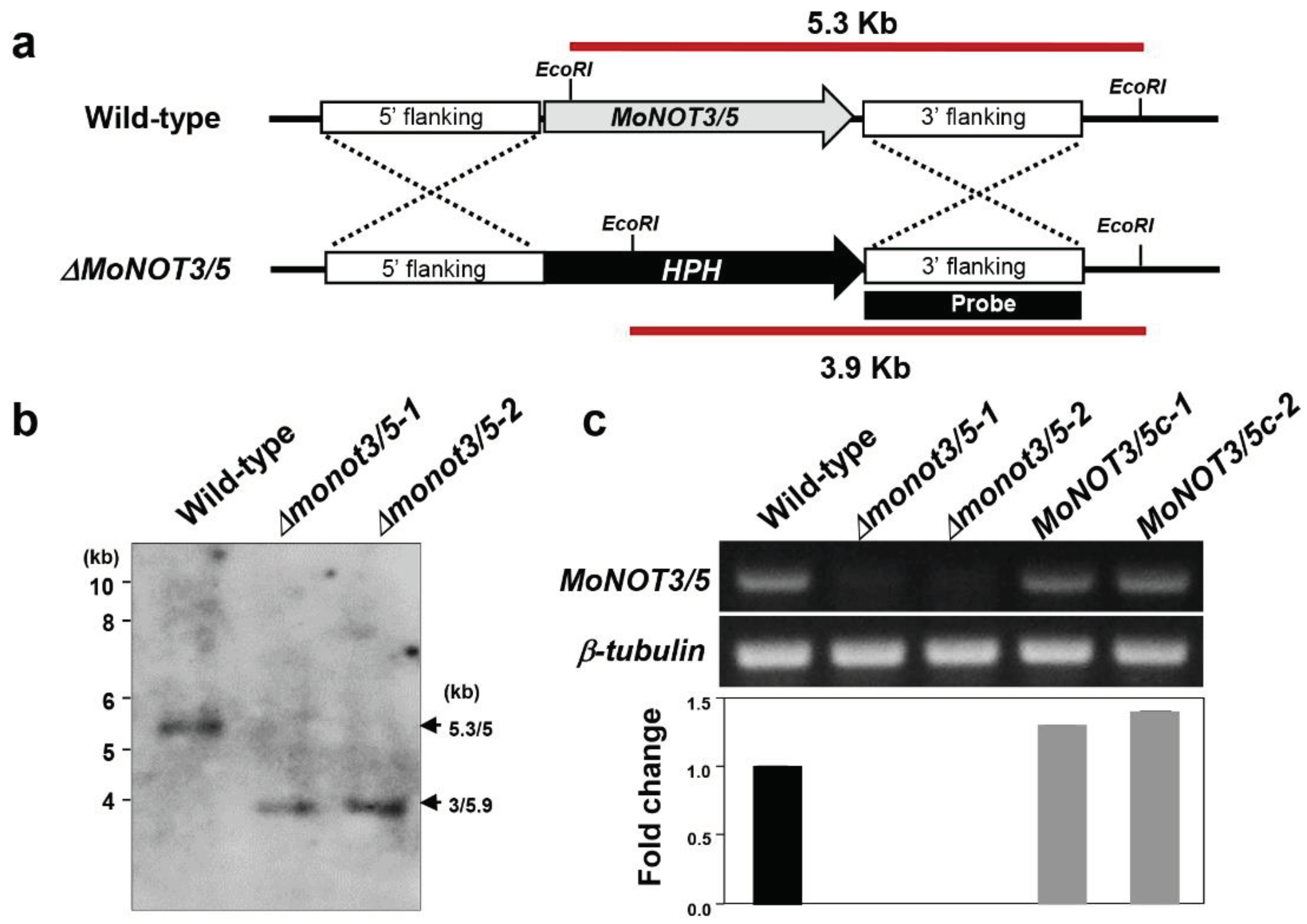
Figure 3.
Hyphal growth of M. oryzae WT, Δmonot3/5, and MoNOT3/5c on various media. (a) Hyphal growth of M. oryzae was measured on complete medium (CM), minimal medium (MM), carbon-starved medium based on MM (MM-C), and nitrogen-starved medium based on MM (MM-C). (b) Mycelial growth (mm) was measured at 9 days post-inoculation on the different types of media. Significance was determined by Tukey’s test (P = 0.01).
Figure 3.
Hyphal growth of M. oryzae WT, Δmonot3/5, and MoNOT3/5c on various media. (a) Hyphal growth of M. oryzae was measured on complete medium (CM), minimal medium (MM), carbon-starved medium based on MM (MM-C), and nitrogen-starved medium based on MM (MM-C). (b) Mycelial growth (mm) was measured at 9 days post-inoculation on the different types of media. Significance was determined by Tukey’s test (P = 0.01).
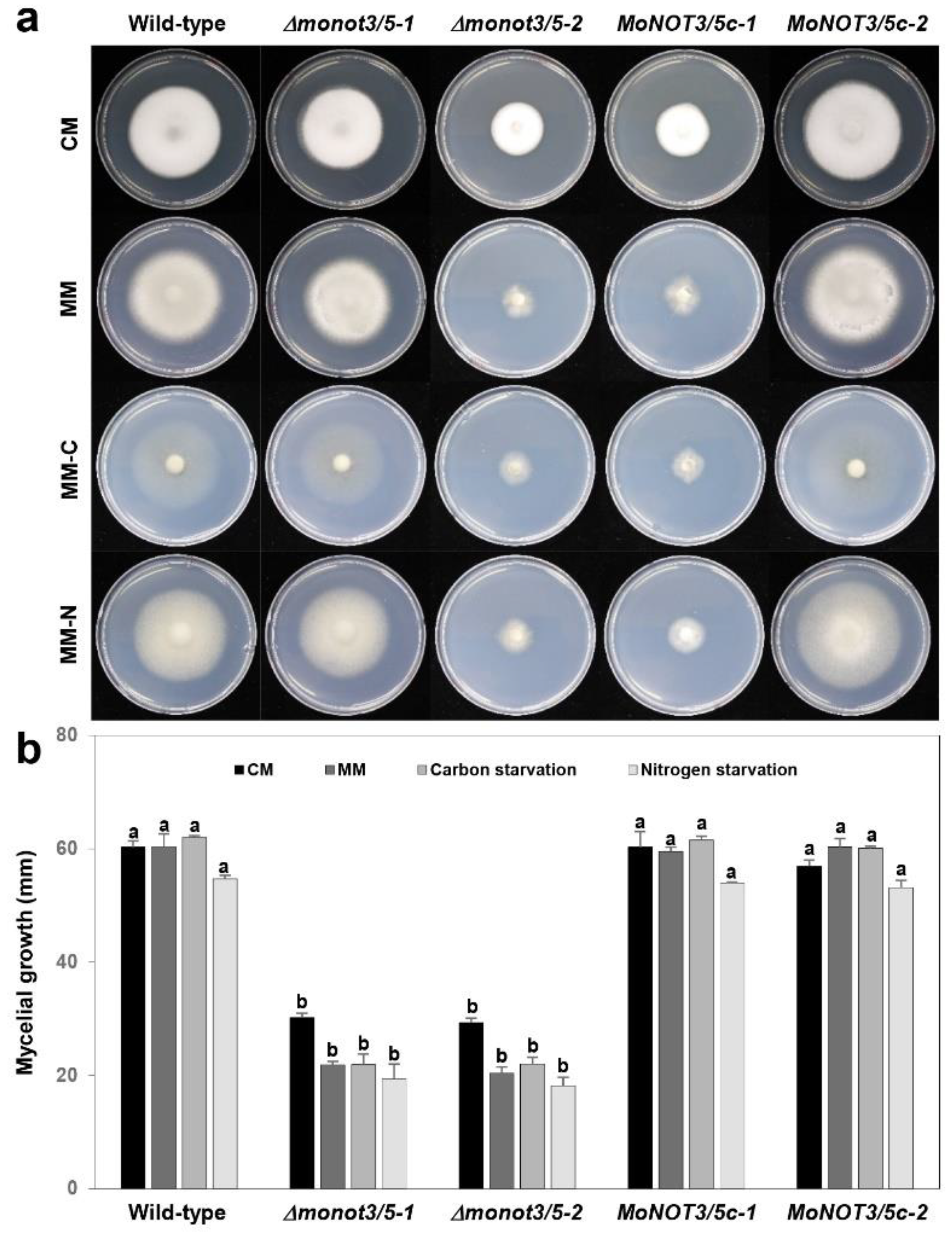
Figure 4.
Effect of MoNOT3/5 disruption on conidiation, production of conidia, and conidial morphology of M. oryzae. (a) Development of conidiophores and conidia was visualised in M. oryzae WT, Δmonat3/5, and MoNOT3/5c strains grown on oatmeal agar media. Conidiogenesis on conidiophores was observed under a microscope. Scale bar, 100 μm. (b) Production of conidia numbers in M. oryzae WT, Δmonat3/5, and MoNOT3/5c strains. Conidia of the WT, deletion mutants, and complemented strains were collected from V8 agar after incubation for 7 days. (c) Conidial shape and septum formation were analysed in M. oryzae WT, Δmonat3/5, and MoNOT3/5c strains. Conidia were stained by calcofluor white with KOH. (d) Distribution of conidia cell numbers in the control and Δmonat3/5 mutant strains. Conidia (>100) of each strain were counted by microscopic observation. (c) Conidial size of the WT, Δmonat3/5, and MoNOT3/5c strains. Values are the means±SD from >100 conidia of each strain, which were measured using the Axiovision image analyser. Tukey’s test was used to determine significance at the 95% probability level. The same letters show no significant differences.
Figure 4.
Effect of MoNOT3/5 disruption on conidiation, production of conidia, and conidial morphology of M. oryzae. (a) Development of conidiophores and conidia was visualised in M. oryzae WT, Δmonat3/5, and MoNOT3/5c strains grown on oatmeal agar media. Conidiogenesis on conidiophores was observed under a microscope. Scale bar, 100 μm. (b) Production of conidia numbers in M. oryzae WT, Δmonat3/5, and MoNOT3/5c strains. Conidia of the WT, deletion mutants, and complemented strains were collected from V8 agar after incubation for 7 days. (c) Conidial shape and septum formation were analysed in M. oryzae WT, Δmonat3/5, and MoNOT3/5c strains. Conidia were stained by calcofluor white with KOH. (d) Distribution of conidia cell numbers in the control and Δmonat3/5 mutant strains. Conidia (>100) of each strain were counted by microscopic observation. (c) Conidial size of the WT, Δmonat3/5, and MoNOT3/5c strains. Values are the means±SD from >100 conidia of each strain, which were measured using the Axiovision image analyser. Tukey’s test was used to determine significance at the 95% probability level. The same letters show no significant differences.
Figure 5.
Effect of MoNOT3/5 deletion on conidial germination and appressorium formation. (a) Microscopic observation of appressorium development on germ tubes. Appressorium formation was examined at 8 and 24 h after incubation of the conidial suspension on hydrophobic cover slips. Black arrowheads in Δmonot3/5 indicate the occurrence of swellings and hooks in a germ tube. Percentage of conidial germination (b) and appressorium formation (c) on a hydrophobic coverslip was measured at 8 and 24 h after incubation of the conidial suspension under a light microscope using conidia harvested from 6-day-old V8-Juice agar medium. Tukey’s test was used to determine significance at the 95% probability level. The same letters show no significant differences.
Figure 5.
Effect of MoNOT3/5 deletion on conidial germination and appressorium formation. (a) Microscopic observation of appressorium development on germ tubes. Appressorium formation was examined at 8 and 24 h after incubation of the conidial suspension on hydrophobic cover slips. Black arrowheads in Δmonot3/5 indicate the occurrence of swellings and hooks in a germ tube. Percentage of conidial germination (b) and appressorium formation (c) on a hydrophobic coverslip was measured at 8 and 24 h after incubation of the conidial suspension under a light microscope using conidia harvested from 6-day-old V8-Juice agar medium. Tukey’s test was used to determine significance at the 95% probability level. The same letters show no significant differences.
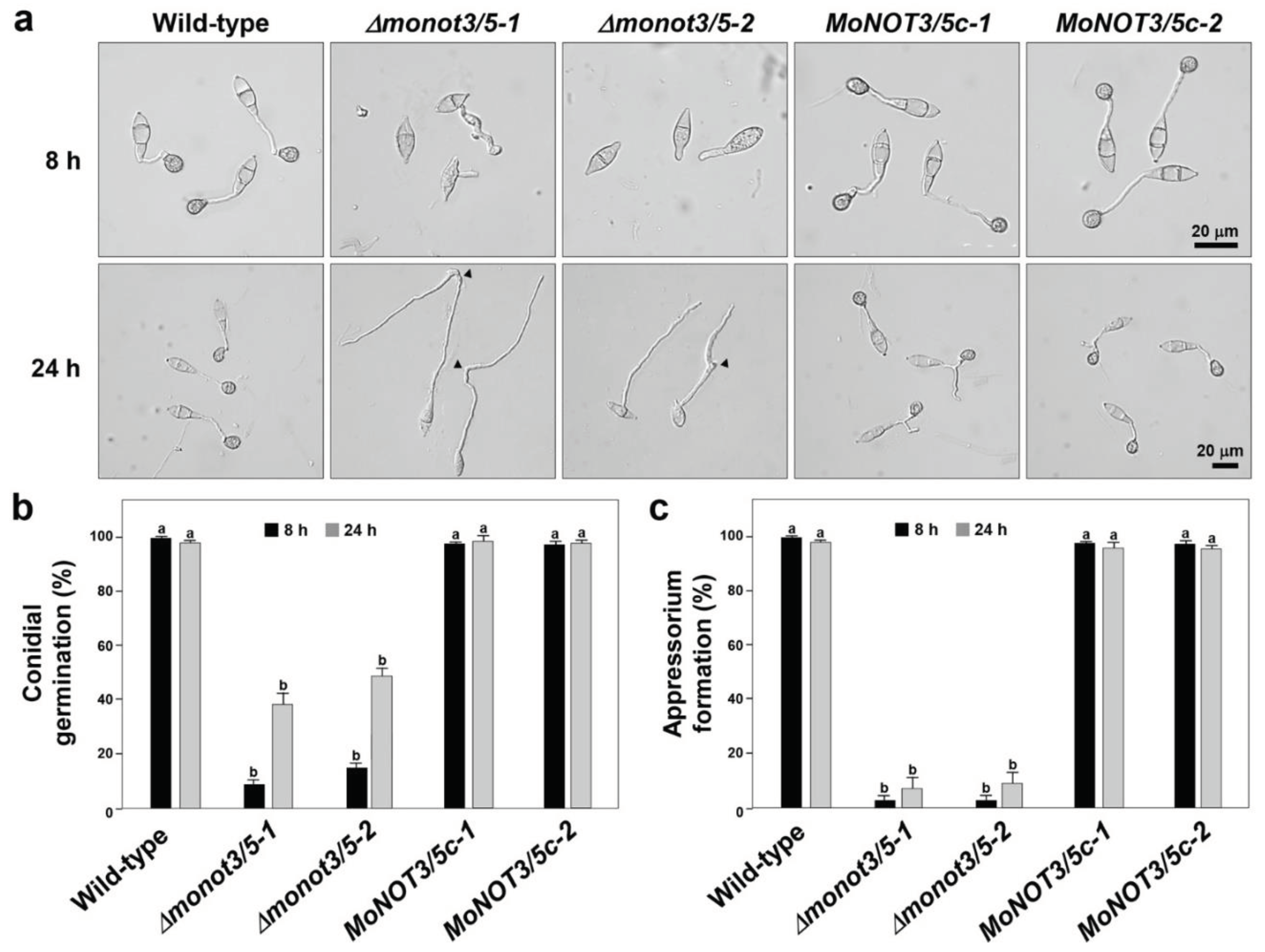
Figure 6.
Pathogenicity of Δmonot3/5. (a) Pathogenicity assay results. Spray inoculation (left). Conidial suspensions (1 × 105/mL) were sprayed onto 4-week-old rice seedlings, and lesions were observed at 5 days post-inoculation. Detached leaf inoculation (middle). Conidial suspensions (2 × 104/mL) were dropped onto 4-week-old rice seedlings. Infiltration inoculation (right). Conidial suspensions (2 × 104/mL) were injected onto 4-week-old rice seedlings. (b) Onion epidermal cell infection 24 and 48 h after inoculation. (c) Rice sheath infection 48 h after inoculation. The growth of invasive hyphae was observed under a microscope at 48 h post-inoculation. Arrows indicate appressoria.
Figure 6.
Pathogenicity of Δmonot3/5. (a) Pathogenicity assay results. Spray inoculation (left). Conidial suspensions (1 × 105/mL) were sprayed onto 4-week-old rice seedlings, and lesions were observed at 5 days post-inoculation. Detached leaf inoculation (middle). Conidial suspensions (2 × 104/mL) were dropped onto 4-week-old rice seedlings. Infiltration inoculation (right). Conidial suspensions (2 × 104/mL) were injected onto 4-week-old rice seedlings. (b) Onion epidermal cell infection 24 and 48 h after inoculation. (c) Rice sheath infection 48 h after inoculation. The growth of invasive hyphae was observed under a microscope at 48 h post-inoculation. Arrows indicate appressoria.
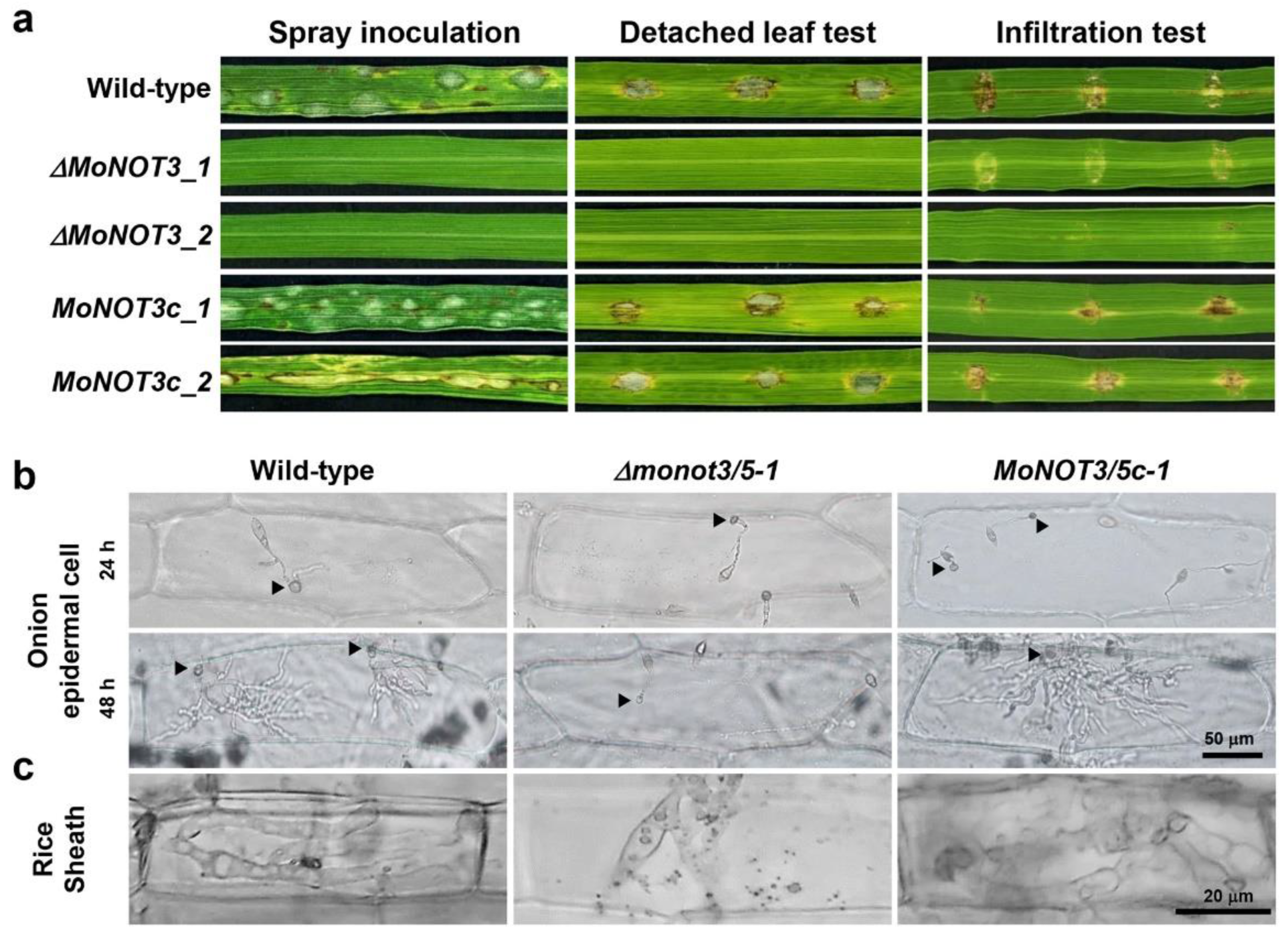
Figure 7.
Effect of stress conditions on growth rates of Δmonot3/5. The WT, Δmonot3/5 (Δmonot3/5-1 and Δmonot3/5-2), and complemented (MoNOT3/5c-1 and MoNOT3/5c-2) strains were grown for 9 days on CM and CM containing CaCl2 (0.2 M), NaCl (0.5 M), MnCl2 (0.02 M), sodium dodecyl sulphate (SDS) (0.01%), Congo red (200 ppm), calcofluor white (CFW) (200 ppm), and H2O2 (2 mM) at a temperature of 32°C. The 5 mm-diameter mycelial blocks were transferred onto the media and incubated for 9 days at 25°C.
Figure 7.
Effect of stress conditions on growth rates of Δmonot3/5. The WT, Δmonot3/5 (Δmonot3/5-1 and Δmonot3/5-2), and complemented (MoNOT3/5c-1 and MoNOT3/5c-2) strains were grown for 9 days on CM and CM containing CaCl2 (0.2 M), NaCl (0.5 M), MnCl2 (0.02 M), sodium dodecyl sulphate (SDS) (0.01%), Congo red (200 ppm), calcofluor white (CFW) (200 ppm), and H2O2 (2 mM) at a temperature of 32°C. The 5 mm-diameter mycelial blocks were transferred onto the media and incubated for 9 days at 25°C.
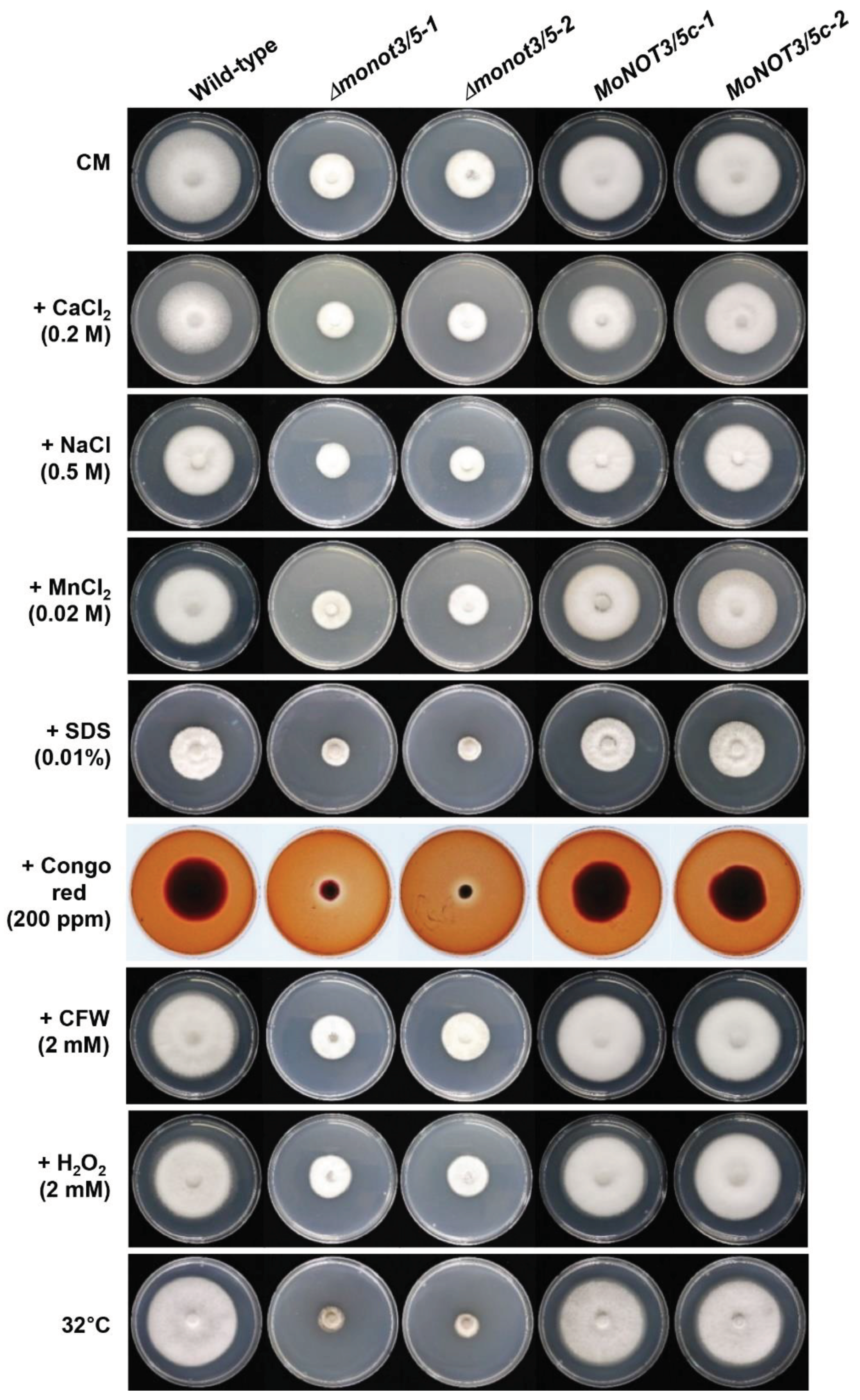
Figure 8.
Expression of the WT and Δmonot3/5 in M. oryzae. (a) Relative expression of conidiation-related genes during conidiation in the Δmonot3/5. (b) Relative expression of ubiquitin-related genes during mycelial growth in the Δmonot3/5. Transcript expression was quantified by qRT-PCR analysis, and values were normalized to the expression of β-tubulin. Relative expression levels, presented as fold changes (2-ΔΔct), were calculated by comparison with the WT strain.
Figure 8.
Expression of the WT and Δmonot3/5 in M. oryzae. (a) Relative expression of conidiation-related genes during conidiation in the Δmonot3/5. (b) Relative expression of ubiquitin-related genes during mycelial growth in the Δmonot3/5. Transcript expression was quantified by qRT-PCR analysis, and values were normalized to the expression of β-tubulin. Relative expression levels, presented as fold changes (2-ΔΔct), were calculated by comparison with the WT strain.
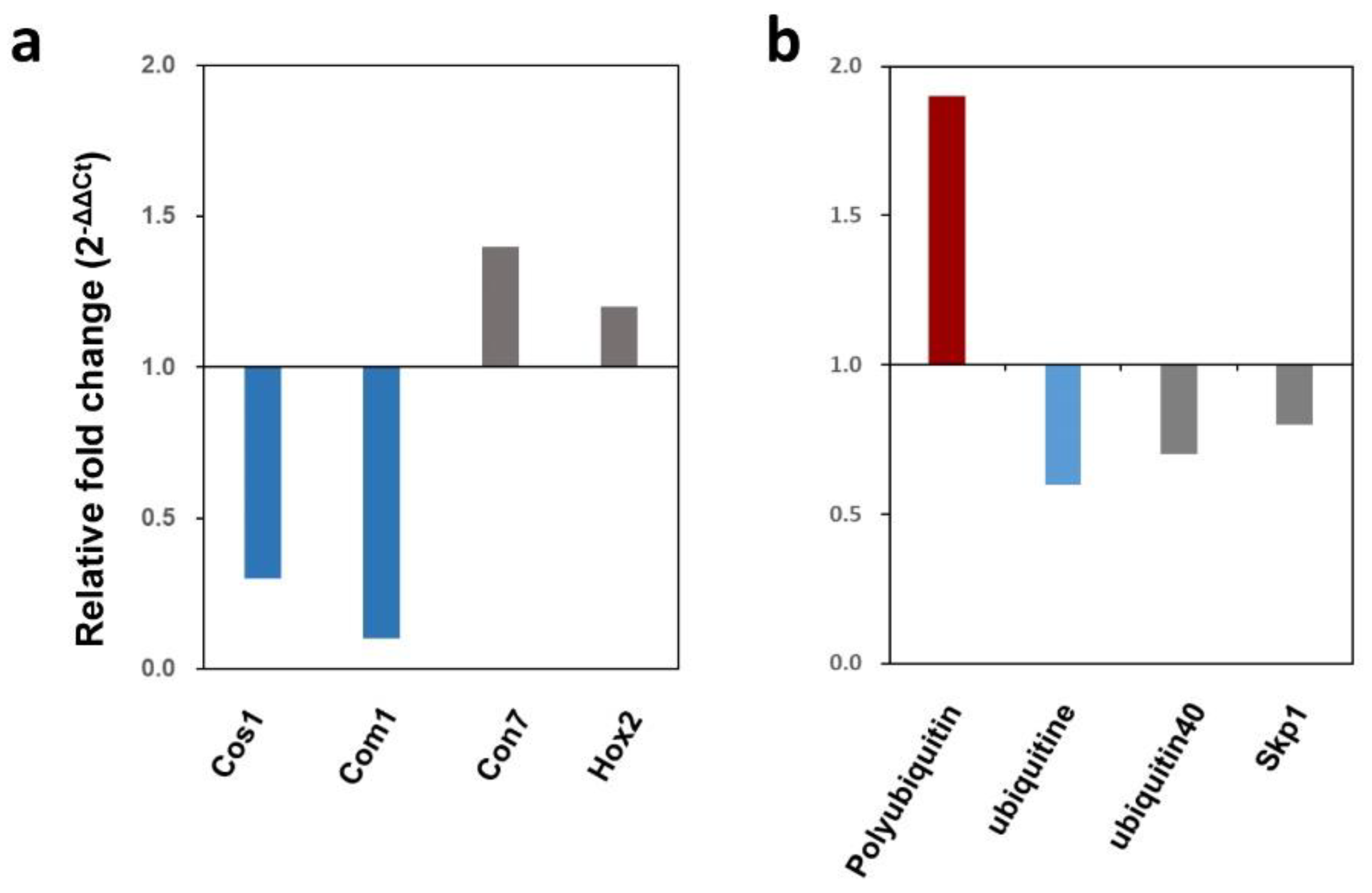

Table 1.
Primers used in this study.
| Target | Name | Sequence (5′-3′) |
|---|---|---|
| Knockout | MoNOT3/5_5UF | ATATACAGGAGGCGGGGTCAGAGT |
| construct | MoNOT3/5_5UR | CCTCCACTAGCTCCAGCCAAGCCGGCTAGTTGTTGTTTCGGATGTCT |
| MoNOT3/5_3DF | GTTGGTGTCGATGTCAGCTCCGGAGGAGTGAATACGGCCAATAGC | |
| MoNOT3/5_3DR | GCACAGAGCCTAACATCAAACCCC | |
| MoNOT3/5_5UF_nested | GACACCATGAAACCACGCACTCTA | |
| MoNOT3/5_3DR_nested | GGACCAACAAGCTCCTCTCA | |
| hph gene | HygB_F | TCAGCTTCGATGTAGGAGGG |
| HygB_R | TTCTACACAGCCATCGGTCC | |
| Screening of | MoNOT3/5_ORF_F | TACATACGCCACTACCAACCATTA |
| transformant | MoNOT3/5_ORF_R | GATGCTGGAAGGTGGTAGATAGAT |
| qRT-PCR | Mo β-tubulin_F | ACAACTTCGTCTTCGGTCAG |
| Mo β-tubulin_R | GTGATCTGGAAACCCTGGAG | |
| MoNOT3/5_qRT_F | CAAATGGAGAGGTTCAAAGCG | |
| MoNOT3/5_qRT_R | TGTCGATCATGTTCCCAAGG | |
| COS1_qRT_F | TGCACCACGATCCCAGAGA | |
| COS1_qRT_R | GCGATGTTGTGCCGTTGTTCC | |
| COM1_qRT_F | GCCAGAGGTCCGCTATCAAA | |
| COM1_qRT_R | CGGGATCTCGTCACTGGATT | |
| CON7_qRT_F | TAAGGAGATCCGCAAAGAGT | |
| CON7_qRT_R | TAGCGTTGTAGTCGGGGAGT | |
| HOX2_qRT_F | TGGGGTTCTGCAGCCATGTT | |
| HOX2_qRT_R | GTCCCGTGGTGTTACGTTCTGG | |
| Polyubiqutin_qRT-F | CAACGCCTTATTTTCGCTGG | |
| Polyubiqutin_qRT-R | TCTTGCCCGTCAAAGTCTTG | |
| Ubiqutin_qRT-F | GCAAGTTCAACTGCGACAAG | |
| Ubiqutin_qRT-R | TCCACACTTTCTCTTCCTGC | |
| Ubiqutin40_qRT-F | AGTTGGCTGTGCTCAAGTAC | |
| Ubiqutin40_qRT-R | GTAGGTCAGGTGGCAACG | |
| Skp1-qRT-F | TGGGATCAGAAGTTCATGCAG | |
| Skp1-qRT-R | ATATCAAGGTAGTTGCTCGCC |
Table 2.
Hyphal growth of M. oryzae strains under diverse stress conditions.
| Strain | Hyphal growtha (mm) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| CM | CaCl2 (0.2 M) |
NaCl (0.5 M) |
MnCl2 (0.02 M) |
SDS (0.01%) |
Congo red (200 ppm) |
CFW (2 mM) |
H2O2 (2 mM) |
32°C | |
| Wild-type | 69.3±0.4ab | 50.9±1.9a | 44.4±1.7a | 53.0±1.7a | 30.6±0.3a | 45.4±0.8a | 68.3±0.2a | 58.9±1.5a | 65.3±0.2a |
| Δmonot3/5-1 | 34.3±0.3b | 26.0±0.4b | 22.2±0.5b | 27.7±0.8b | 11.3±0.6b | 12.2±0.2b | 33.2±0.4b | 29.8±1.0b | 17.4±0.4b |
| Δmonot3/5-2 | 32.7±0.6b | 25.9±0.4b | 23.8±0.8b | 28.1±0.2b | 14.0±0.5b | 11.4±0.6b | 33.7±0.5b | 30.1±0.1b | 17.3±0.3b |
| MoNOT3c-1 | 68.3±0.6a | 49.9±1.3a | 44.2±1.3a | 52.8±1.1a | 32.1±0.6a | 42.3±1.9a | 67.2±0.4a | 59.1±0.2a | 65.2±0.3a |
| MoNOT3c-2 | 67.7±0.2a | 46.7±0.8a | 43.0±1.4a | 50.8±1.1a | 30.0±1.0a | 45.7±1.2a | 68.0±0.5a | 58.9±1.4a | 65.3±0.3a |
a A 5 mm diameter mycelial plug was inoculated to complete media (CM) or CM supplemented with the indicated chemicals and incubated at 25°C. Radial growth was measured at 9 days; b Tukey’s test was used to determine significance at the 95% probability level. The same letters in a column showed no significant difference.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



