
Submitted:
17 March 2025
Posted:
19 March 2025
You are already at the latest version
Abstract
The enzyme 5,10-methylenetetrahydrofolate reductase (MTHFR) catalyzes the conversion of 5,10-methylenetetrahydrofolate to 5-methyltetrahydrofolate, a process essential for the methylation of homocysteine to methionine. Polymorphisms in the MTHFR gene can reduce enzyme activity, disrupting the folate cycle and leading to hyperhomocysteinemia. The two most common polymorphisms associated with this gene are 667C>T (rs1801133) and 1298A>C (rs1801131). Background: This review provides a comprehensive summary of current knowledge regarding MTHFR polymorphisms, with a particular focus on their potential impact on disease susceptibility. We hope this review will serve as a valuable resource for understanding the significance of MTHFR polymorphisms and their complex relationships with various diseases. Methods: For this review, we prioritized recent evidence, focusing on reviews and meta-analyses published between 2015 and 2025, sourced from PubMed and Google Scholar. Results: We explore the connection between these polymorphisms and a broad spectrum of medical conditions, including: cardiovascular diseases and oxidative stress pathology, neurological and psychiatric disorders, such as Autism Spectrum Disorder, Alzheimer’s disease, Schizophrenia, and Major Depressive Disorder, fertility, pregnancy, and neonatal complications, including recurrent pregnancy loss, preeclampsia, preterm birth, low birth weight, and neural tube defects, metabolic disorders, such as diabetes mellitus, inflammatory bowel disease, and non-alcoholic fatty liver disease, oncological conditions, including breast, prostate, and ovarian cancers, as well as leukemia, autoimmune diseases, particularly rheumatoid arthritis. Conclusions: While some diseases have a well-established association with MTHFR polymorphisms, others require further investigation. Our analysis highlights the crucial role of environmental factors, such as ethnic background and dietary folate intake, in influencing study outcomes.
Keywords:
MTHFR
; mutations
; polymorphisms
; homocysteine
; folate
; atherosclerosis
; cancerogenesis
; Alzheimer’s disease
; diabetes
; NTDs
1. Introduction
Methylenetetrahydrofolate reductase (MTHFR) is an enzyme responsible for converting 5,10-methylenetetrahydrofolate (5,10-methylene-THF) to 5-methyltetrahydrofolate (5-methyl-THF). MTHFR is a flavoprotein that facilitates NADPH-dependent reduction, utilizing FAD as a cofactor [1]. This conversion is a critical step in the folate cycle [Figure 1], with 5-methyl-THF subsequently serving as a substrate for the methylation of homocysteine (Hcy) to methionine (Met). This reaction is catalyzed by methionine synthase (MS), with vitamin B12 acting as a coenzyme.
The direct products and further derivatives of this reaction serve as one-carbon donors, which are essential for numerous metabolic pathways [2]. These pathways include the synthesis of amino acids, purines, deoxythymidine monophosphate (dTMP), creatine, and epinephrine, as well as DNA and protein methylation [3]. Given the wide range of these processes, MTHFR plays a crucial role in cellular metabolism.
The MTHFR gene is located on chromosome 1p36.22 [4]. It is transcribed alongside other enzymes involved in the folate cycle, such as cystathionine β-synthase (CBS) and methionine synthase (MS), as they share a common promoter region. Alternative splicing has been observed in humans [1]. To date, approximately 247 single nucleotide polymorphisms (SNPs) [5,6] and over 200 pathogenic variants [6,7] of the MTHFR gene have been identified. Although 34 rare polymorphisms have been linked to severe MTHFR deficiency, research on this topic remains limited, as only around 50 patients have been diagnosed with one of these polymorphisms [8].
Two of the nine known MTHFR gene polymorphisms, 677C>T and 1298A>C, are the most common and widely studied. These are the only two polymorphisms currently known to have a confirmed effect on enzyme activity [8]. The 677C>T (rs1801133) polymorphism is a point mutation that results in the substitution of alanine (Ala) at position 222 with valine (Val) in the enzyme’s catalytic domain (A222V) [1]. This polymorphism increases thermolability [9] and reduces the enzyme’s affinity for FAD, leading to a 35% decrease in enzyme efficiency per mutated allele [1]. The prevalence of this polymorphism in human populations averages 30–40%, but it varies from 0% to over 40%, depending on geographic location, with minimal difference between genders [2]. Homozygous individuals have been diagnosed with mild hyperhomocysteinemia. The 1298A>C (rs1801131) polymorphism is a point polymorphism that causes a substitution of glutamate (Glu) at position 429 with alanine (Ala) (E429A) [9]. Unlike 677C>T, carrying the 1298A>C mutated allele alone does not lead to hyperhomocysteinemia. However, individuals who are 677C>T heterozygotes and also carry the 1298A>C allele exhibit enzyme function similar to 677C>T homozygotes [1]. Both polymorphisms can result in decreased MTHFR activity, potentially disrupting the folate cycle and related metabolic processes.
Hyperhomocysteinemia (HHcy) is a condition characterized by elevated blood levels of homocysteine (Hcy), a non-proteinogenic, sulfur-containing amino acid. It can result from disruptions in the folate cycle, such as MTHFR gene polymorphisms, which impair the methylation of Hcy to methionine (Met). A compensatory defense mechanism exists, in which excess Hcy is converted to cysteine through the transsulfuration pathway [10]. However, this process appears insufficient to fully prevent HHcy.
The study conducted by Nishio et al. suggests that homozygous individuals with the TT genotype (resulting from the 677C>T polymorphism) have 20% lower folate levels compared to individuals without the polymorphism, despite having the same folate intake [11]. This may be particularly significant during periods of high folate demand, such as pregnancy, lactation, or infancy [12]. Individuals with this homozygous polymorphism may require not only folic acid supplementation but also active forms of folate. Single-carbon units, such as tetrahydrofolate (THF) and S-adenosylmethionine (SAM), are derivatives of the folate cycle [3]. These molecules serve as methyl group donors, and their deficiency can lead to: DNA, protein, and phospholipid hypomethylation, degradation of telomere sequences, and disrupted amino acid and nucleotide metabolism [13]. These effects highlight the critical role of folate metabolism in maintaining genetic stability and cellular function.
The toxic effects of Hcy, which primarily impact the nervous and circulatory systems, are attributed to multiple mechanisms [Figure 2]. These mechanisms highlight the widespread impact of HHcy on cellular and systemic functions, particularly in the cardiovascular and neurological systems [13,14].
The folate cycle has been extensively researched, particularly in recent years. However, most studies focus either on specific aspects of the metabolic pathway or on the correlation between folate cycle disruptions and particular medical conditions. There appear to be few comprehensive reviews that summarize folate cycle knowledge in a practical, multidisciplinary manner across various medical fields.
In this article, we aim to highlight the medical disciplines most relevant to the folate cycle and its associated research. Therefore, the purpose of this review is to examine the most common MTHFR gene polymorphisms, explore their implications across selected medical fields and provide a comprehensive summary of current knowledge on the topic. By doing so, we hope to provide a broad yet practical perspective on the role of MTHFR polymorphisms in health and disease.
2. MTHFR in Cardiology
MTHFR polymorphisms and disorders of folate metabolism are the underlying causes of hyperhomocysteinemia (HHcy). Impaired methylation of homocysteine (Hcy) leads to its accumulation in the body, which has been identified as an independent risk factor for cardiovascular diseases. Studies show that up to 40% of patients diagnosed with premature coronary artery disease, vascular disease or recurrent venous thrombosis have elevated homocysteine levels [15].
Epithelial damage occurs due to Hcy's tendency to auto-oxidize and the impairment of the vasodilatory response associated with nitric oxide (NO). Hcy also exerts a pro-inflammatory effect on endothelial cells (EC), inducing the secretion of pro-inflammatory cytokines (IL-1β, IL-6, IL-8, and TNF-α). These cytokines are linked to reactive oxygen species (ROS) formation and the activation of nuclear factor kappa B (NF-κB) [16]. Cytokines recruit monocytes, which differentiate into pro-inflammatory macrophages that transform into foam cells, contributing to plaque formation. Furthermore, the presence of Hcy affects endothelial cell (EC) aging, promoting apoptosis or other forms of cell death, such as gasdermin D-dependent pyroptosis [17].
HHcy is also involved in cellular damage by affecting respiratory chain proteins [18]. Evidence shows that HHcy reduces the activity of complexes II and III in cardiac cells of rats [19]. The reactive metabolite of Hcy, thiolactone, can bind to the amino groups of lysine in susceptible proteins, including those in the cytochrome c complex, which is involved in transferring electrons from complex III to IV [20]. Studies indicate that Hcy increases the leakage of superoxide, which is normally produced at low levels, but when elevated, it leads to the formation of ROS [21]. The cited research also presents mechanisms of ROS formation in the presence of copper and suggests that Hcy influences Ca²⁺ homeostasis, which is linked to ischemia-reperfusion injury in hypoxic heart cells during myocardial infarction [18,22]. In summary, Hcy affects cell metabolism through oxidative stress, inhibits respiratory chain proteins, reduces ATP production, induces protein damage, and has pro-inflammatory effects on cardiovascular cells, contributing to vascular dysfunction and increasing the risk of a heart disease [Figure 3].
Recent reviews have summarized numerous studies and meta-analyses indicating cardiovascular complications associated with the 677C>T polymorphism, including an increased risk of developing cardiovascular disease (CVD). 677C>T homozygotes of the MTHFR gene are at a higher risk of developing CVD [23]. However, the same review also included studies that did not find a correlation between the 677C>T variant and CVD in patients with diabetes, though this study had a small sample size of 107 participants [23]. A large number of studies have shown an increased risk of ischemic stroke (IS) in individuals with the MTHFR gene polymorphism. For example, a study by Moti Wala et al. found an association between MTHFR and carotid and spinal artery dissection, which significantly affects the risk of IS at a young age. The review included 11 articles, comprising three case reports and one experimental study with a sample size of 4840 patients [24]. Hyperhomocysteinemia (HHcy) in homozygotes has been shown to be a risk factor for vascular events, including myocardial infarction [25]. Arterial dissection refers to structural damage to the arterial wall with intimal hemorrhage, which forms an intramural hematoma (IMH), resulting in a bulge inside the vessel and the potential for thrombus formation. This condition is also associated with Hcy-related damage mechanisms. IMH hinders perfusion, increasing the risk of embolism [26]. A retrospective study by Ahmed et al., involving 4055 patients, provides evidence of a correlation between vitamin B12 deficiency (present in folate cycle disorders) and ischemic stroke [27]. Another study by Park et al. on 146 patients with thromboembolic disease also found a correlation between thrombosis, HHcy, and the 677C>T polymorphism. The authors concluded that MTHFR gene polymorphism testing could aid in the early diagnosis of arterial dissection, which often cannot be diagnosed based on symptoms like headache or migraine [28].
Polymorphisms that reduce the enzymatic activity of MTHFR cause folate deficiency, which is also an independent risk factor for stroke. The correlation between the 677C>T mutation and stroke has proven challenging due to inconsistent results from studies in different populations [29]. The study by Zhao et al. updated previous meta-analyses on this topic by including clinical and cohort studies investigating MTHFR 677C>T polymorphism and stroke susceptibility, focusing on patients diagnosed with stroke for the first time. Non-original studies, duplicate articles, and those where the Hardy-Weinberg equilibrium value was <0.05 were excluded. In total, 96 clinical-cohort studies from different populations and age groups, comprising 314,814 patients, were selected for meta-analysis. In the dominant model, the MTHFR 677C>T polymorphism significantly increased the risk of ischemic stroke (OR = 1.47, 95% CI = 1.33-1.61). A correlation was observed in all age groups except the younger population. In the recessive model, similar results were obtained (OR = 1.52, 95% CI = 1.38-1.81), with a significant increase in risk observed in older individuals. Both heterozygous and homozygous models showed similar results [30]. These findings were consistent with previous analyses [31]. Similarly, a meta-analysis including nine studies (3,337 patients) showed a statistically significant increase in the risk of ischemic stroke in Chinese cohorts, with a similar trend observed in other populations [32]. The study by Qin et al. indicates reduced efficacy of Hcy-lowering treatment in individuals with the 677C>T polymorphism. However, the results should be interpreted with caution due to the lack of prospective studies and the fact that not all study groups were age-specific [29].
3. MTHFR in Oncology
Reduced enzymatic activity of methylenetetrahydrofolate reductase (MTHFR) caused by the 677C>T and 1298A>C polymorphisms has been observed to increase the propensity for cancer development. Both polymorphisms are associated with hyperhomocysteinemia, decreased blood folate levels [33], and disturbances in one-carbon metabolism, which hinder proper DNA methylation, thus contributing to genomic instability [34,35]. The reduced folate supply is also linked to impaired nucleotide incorporation and inhibited DNA repair [36,37]. These factors, together, make both the 677C>T and 1298A>C polymorphisms particularly relevant to the study of various cancers, including breast, prostate, ovarian cancers, as well as leukemia [38,39,40,41,42,43,44].
3.1. Breast Cancer
A review by Petrone et al. examined 19 studies on the prevalence of MTHFR polymorphisms in breast cancer (BC) across different populations. Although the results were inconclusive, 677C>T and, to a lesser extent, 1298A>C were found to increase the risk of BC [45]. Notably, the 1298A>C polymorphism was observed to possess a protective role against BC among Kazakh women [46]. Moreover the 677CT 1298AA diplotype was found to decrease BC susceptibility in the South Asian population [47]. The combined effect of the 677C>T SNP and risk factors, such as alcohol consumption and low folate intake, was also found to increase BC susceptibility [48]. Both polymorphisms were associated with more aggressive cancer biophenotypes (such as HER-2 and TN), and the 1298A>C polymorphism was found to be a risk factor for increased lymph node metastasis, although these studies had sample sizes of less than 300 [49]. These results align with a meta-analysis of 75 studies by Kumar et al., which concluded a moderate overall association between the 677C>T polymorphism and susceptibility to BC. In the Asian population, an increased risk was found across all genetic models. In contrast, in the Caucasian population, a statistically significant association was observed only in the allele contrast model (C vs. T). In the mixed subgroup, significant correlations were observed in the allele contrast, dominant, and co-dominant models. Additionally, the allele contrast model revealed a significantly stronger association between the 677C>T polymorphism and BC in hospital-based studies compared to population-based studies. An analysis of 18 studies with varying results found no overall association between menopausal status and the effect of the 677C>T polymorphism on BC risk [50].
3.2. Prostate Cancer
A systematic review by You et al. aimed to clarify the effect of MTHFR polymorphisms on the prevalence of prostate cancer (PC) in men of Asian, Caucasian, and mixed descent. The overall study revealed no significant association between the 677C>T polymorphism and an increased risk of PC. However, an analysis of the Asian subgroup revealed a decrease in PC susceptibility across all five genetic models used [51]. On the other hand, a collective study of 6,396 cases and 8,913 controls with the 1298A>C polymorphism—while not showing any statistically significant overall increase in susceptibility—revealed an increased risk of PC among American men of mixed descent. The authors speculate that this may be due to region-based risk factors [52,53,54].
3.3. Ovarian Cancer
A meta-analysis by Xiong et al. incorporated 16 studies on both MTHFR polymorphisms and their influence on ovarian cancer (OC) susceptibility. No significant overall correlation was established regarding the 677C>T SNP, though it was found to be a risk factor among the Asian subgroup. Since Asians made up only a fraction of the examined participants, the authors speculate that a more balanced distribution of participants might alter the overall results. In terms of the 1298A>C polymorphism, the study (with 6,860 participants) differentiated only between "overall" and "Caucasian" groups, with no significant increase in risk observed in either [55].
3.4. Leukemia
Leukemia has not been proven to have a specific cause and is believed to result from a combination of environmental, genetic, and epigenetic factors [35,56,57]. However, multiple studies have suggested that, through disturbances in folate metabolism, MTHFR gene polymorphisms likely negatively affect hematopoietic progenitor cells, triggering leukemia [35,58,59]. Four common types of leukemia have been studied: chronic lymphocytic leukemia (CLL), acute lymphocytic leukemia (ALL), acute myeloblastic leukemia (AML), and chronic myeloblastic leukemia (CML) [60,61].
A meta-analysis by Raoufi et al. included a total of 16 studies on the association of common MTHFR polymorphisms and the risk of CLL. The results failed to show a significant association between the 677C>T variation and CLL in any of the five models used. However, the allelic model (A vs. C) revealed a correlation between increased CLL susceptibility and the 1298A>C polymorphism. It is important to note that, although the individual studies were conducted in different geographical regions, the participants were not grouped based on ethnicity. Moreover, environmental factors and their role in the development of CLL were not considered [60]. A comprehensive meta-analysis by Lien et al. reviewed a total of 97 studies to assess the association between MTHFR polymorphisms and ALL, AML, and CML among participants divided by region and age. Pooled data analysis revealed an overall protective role of the 677C>T CT and TT polymorphisms against the development of leukemia, while subgroup analysis specified the findings. No significant association was found regarding the prevalence of leukemia in adults. Among American children, the TT genotype decreased the risk of ALL, while the CC and TT genotypes increased it. In children from East Asia and Europe, both mutated variants lowered the risk of leukemia, while the CC wildtype was a risk factor among the latter. On the other hand, a leukemogenic effect was discovered among African and South Asian populations. MTHFR 1298A>C polymorphisms were also found to have a protective role against certain types of leukemia, both overall and in specific subgroups. Among adults and children worldwide, the AC and CC genotypes lowered the risk of CML, while the AA wildtype increased it. Furthermore, both mutated variants displayed a protective effect against ALL in American children, with the CC wildtype showing the opposite effect [61].
Though collectively, the findings of the reviewed studies are generally consistent, a notable discrepancy exists between individual papers, which can be attributed to the specific factors considered, sample sizes, and other conditions. Furthermore, the interaction between environmental factors and MTHFR polymorphisms is only explored in a fraction of the articles. Overall, more comprehensive and precise studies are needed to clarify the complex relationship between the discussed polymorphisms and carcinogenesis in individuals.
4. MTHFR in Neurology and Psychiatry
The relationship between MTHFR polymorphisms and neurological and psychiatric disorders is quite complex. HHcy is suspected to cause neurotoxicity [62,63], and most neurological and psychiatric disorders appear to result from a combination of various genetic and environmental factors [64]. MTHFR polymorphisms have been extensively studied in connection with different neurological and psychiatric conditions [65,66]. The aim of this chapter is to summarize some of the findings on the subject.
4.1. Autism Spectrum Disorder (ASD)
Autism Spectrum Disorder (ASD) is a group of complex neurodevelopmental disorders characterized by repetitive behaviors, restricted interests, social and communication impairments, sensory hypersensitivity, and difficulty adjusting to changes [64,67,68]. Symptoms related to ASD usually manifest between 12 and 18 months after birth and the diagnosis is typically made at 2 years of age [68]. The male-to-female ratio is thought to be 4:1 [67,69]. The prevalence of ASD is estimated to be around 1% of the child population [64,67,69,70,71]. The etiology of ASD has been widely researched and is currently attributed to a combination of environmental, genetic, and epigenetic factors [64,70,71]. Suggested environmental risk factors include maternal age exceeding 35, chronic hypertension, pre-eclampsia, gestational hypertension, being overweight before or during pregnancy, medication and toxin exposure, socioeconomic factors, and complications during pregnancy and birth [64,67,68]. Studies have shown that monozygotic twins share ASD in about 90% of cases [71], and heritability is estimated at 50% [68]. Over 600 genes have been linked to ASD, with only a few (such as Fragile X, SHANK 3, and CASPR 2) showing a statistically significant connection [64]. The suspected negative influence of MTHFR polymorphisms and HHcy on the nervous system has sparked research into their potential connection to ASD [69]. This is further supported by the fact that epigenetic processes, such as DNA methylation and histone modifications, may also be disrupted by MTHFR polymorphisms [70]. Additionally, it has been noted that high concentrations of methyl donors are required during neurogenesis, and MTHFR polymorphisms may negatively impact their availability [64].
There is some variability in the findings of the analyzed studies. However, there seems to be a trend that the 677C>T polymorphism has a significant link to ASD, while the 1298A>C polymorphism does not [68,69,70]. A systematic review by Roufael et al. suggests that low levels of folic acid and vitamin B12 are associated with ASD [67]. A systematic review by Hoxha et al. adds that scientific focus should be placed on modulating folate uptake at the blood-brain barrier, via RFC1, folate receptor alpha (FRα; antibodies against it have been found in the cerebrospinal fluid of ASD children), and proton-coupled folate transporter (PCFT), as a way of managing the effects of MTHFR polymorphisms [64]. Additionally, some studies have investigated supplying folic or folinic acid to ASD children or pregnant mothers. Maternal prenatal supplementation of 600µg of folic acid a day is believed to reduce the risk of developing ASD in children [64]. Tests involving supplementation for children have shown some improvement in cognitive functions, as cited by Lam et al. [72].
4.2. Alzheimer’s Disease
Alzheimer's disease (AD) is the most common neurodegenerative disorder, accounting for about 60-80% of dementia cases. Those affected suffer from progressive memory decline and worsening cognitive function, which significantly impacts their quality of life [73]. AD is divided into early-onset and late-onset (EOAD/LOAD), with the age of differentiation being 65 years [74]. The etiology of AD is complex and not yet fully understood. Over 50 genes have been linked to AD so far. Familial EOAD is believed to be caused mainly by polymorphisms in the amyloid precursor protein gene (APP), presenilin1 gene (PSEN1), and presenilin2 gene (PSEN2). Sporadic LOAD is associated with various genetic and environmental factors, with apolipoprotein E (APOE) being the most significant one [74]. HHcy is considered a risk factor for AD, as it affects processes such as DNA methylation and repair, oxidative stress, amyloid β aggregation, tau phosphorylation, vascular endothelial dysfunction, and the secretion of inflammatory mediators like tumor necrosis factor α (TNFα), nuclear factor κB, interleukin 6 (IL-6), and interleukin 8 (IL-8) [73]. Since MTHFR gene polymorphisms may lead to HHcy, several studies have focused on determining the connection between MTHFR polymorphisms and AD, particularly the 677C>T (rs1801133), 1298A>C (rs1801131), and 1793A>G (rs2274976) polymorphisms [74].
A study by Jiang et al., involving 721 AD patients and 365 controls from China, linked the 677C>T and 1298A>C polymorphisms to AD. The 677C>T polymorphism was also associated with an earlier onset, higher Hcy levels, and more severe white matter lesions [74]. Research has also investigated the connection between MTHFR polymorphisms and (amnestic) Mild Cognitive Impairment (aMCI), which is considered a transitional stage between aging and the pathological state of dementia [73,75]. The risk of MCI progressing into AD is estimated at 10-15% per year, compared to 1-2% for unaffected elders [73]. In China and the USA, about 20% of people over 70 years old are affected, with about 40% of them showing potentially reversible changes [75]. A 2021 meta-analysis by Sun et al., involving 8 articles and 2,175 patients, did not link the 677C>T polymorphism to MCI susceptibility. However, the authors noted that the small sample size might have affected the results [75]. Another study by You et al., including 60 aMCI patients and 30 control patients, did find a link between the 677C>T polymorphism and structural changes in the patients' right precunei. Again, the small sample size may have influenced the results [73].
4.3. Schizophrenia
Schizophrenia (SCZ) is a psychiatric condition affecting about 1% of the population. Its symptoms are usually divided into positive and negative. The heritability of SCZ is estimated at about 64-81%, and some studies suggest a strong impact of environmental factors in the development of the condition [76]. Schizophrenia negatively affects the quality of life, and those affected have a higher mortality rate than the general population, largely due to high suicide rates [66].
Attempts to link SCZ to folate and vitamin B12 levels have yielded conflicting results. The correlation between MTHFR polymorphisms and SCZ is also being explored due to the former's potential neurodegenerative effects. The 677C>T and 1298A>C MTHFR polymorphisms both appeared in the Schizophrenia gene database as part of 24 genetic variants associated with SCZ risk [76]. The reviewed studies indicate some interesting findings. Yoo et al. conducted a meta-analysis of 34 case-control studies and found a link between both of the mentioned polymorphisms and an increased risk of schizophrenia, without significant differences between age, sex, and ethnicity [76,77]. Some studies, however, only found a significant link between the 677C>T polymorphism and the risk of SCZ [66]. Zhang et al. researched depressive (negative) symptoms of schizophrenia in relation to MTHFR polymorphisms. Their research included 715 patients with stable SCZ. There was no difference in the occurrence of depressive symptoms in patients with and without MTHFR polymorphisms. Interestingly, those with depressive symptoms did have higher average levels of Hcy, which was concluded to be a possible risk factor for SCZ depressive symptoms rather than the MTHFR polymorphisms [63]. Liu et al. tested 957 affected patients and 576 controls and did not find a link between the 677C>T polymorphism and schizophrenia but did find a correlation between the polymorphism and suicidal behavior in SCZ patients. The study, however, was claimed to be limited by sample size and ethnic inconsistencies [78]. Lam et al. reported a link between the 677C>T polymorphism and the risk and age of onset of schizophrenia. Folate deficiency is reported to aggravate negative symptoms, which is consistent with the results of other studies. Folate treatments for SCZ patients have been tested with inconsistent results [72].
4.4. Major Depressive Disorder
Major Depressive Disorder (MDD) is a psychiatric condition characterized by at least two weeks of low mood or loss of interest, which interferes with daily activities [65]. It decreases the overall quality of life, increases healthcare costs, and carries an increased risk of suicide. About 30% of cases are resistant to pharmacological treatment, classifying them as Treatment-Resistant Depression (TRD) [79]. Similarly to other disorders in this chapter, the development of MDD is caused by multiple factors [65].
Several studies have researched the link between MTHFR polymorphisms and depressive disorders, including MDD. A 2022 meta-analysis by Zhang et al. found a link between MDD and the 677C>T MTHFR polymorphism [77]. Halaris et al. mentioned that folic acid and L-methylfolate have been shown to be effective in treating depression, with L-methylfolate being one of the substances licensed by the FDA for this purpose [79]. Asif et al. also speculated that MTHFR polymorphisms might influence migraine with comorbid depression (MID), as both conditions are linked to the 677C>T polymorphism [80].
4.5. Other Neurological and Psychiatric Disorders
MTHFR polymorphisms have also been studied in relation to a range of conditions not yet mentioned. A considerable amount of research has connected the 677C>T and, in some cases, the 1298A>C polymorphisms with bipolar disorder [65,66,77]. MTHFR polymorphisms can lead to HHcy, which is considered a risk factor for Parkinson’s disease (PD) and complications associated with its L-DOPA treatment [81]. However, Periñán et al. did not find a direct link between the polymorphisms and PD [62]. ADHD has also been linked to MTHFR polymorphisms, with varying results. Sadeghiyeh et al. and Meng et al. found a connection between ADHD and the 1298A>C polymorphism [66,82]. A study by Aguoulea et al. revealed that women with anorexia nervosa were 2.66 times more likely to carry either the 677C>T or the 1298A>C MTHFR polymorphism compared to unaffected individuals [65,83]. The link between MTHFR polymorphisms and neurological and psychiatric disorders remains uncertain as of the time of writing. However, the topic appears to contain some interesting associations that require further research. There is a lack of broad, multinational studies on this subject, as well as research on MTHFR polymorphisms beyond 677C>T and 1298A>C.
5. MTHFR in Gastroenterology and Diabetology
5.1. Type 2 Diabetes Mellitus
Diabetes mellitus (DM) is a worldwide disease that affected 10.5% (approximately 537 million) of the global adult population in 2021, according to the International Diabetes Federation. It is estimated that by 2030, this number will rise to 643 million [84]. DM includes several types, each varying in causes, pathomechanisms, and the onset of symptoms [85]. The most prevalent type is type 2 DM (T2DM), which is diagnosed in 90% of diabetic patients. One of the factors responsible for T2DM is genetic predisposition [86]. T2DM is associated with insulin resistance (IR). HHcy has been suggested to contribute to IR by activating a cascade of reactions that leads to the inhibition of the insulin signaling pathway. In these reactions, activated c-Jun N-terminal kinase is involved in the deactivation of insulin receptor substrate-1 through phosphorylation. HHcy is also connected with oxidative stress through its influence on the production of free radicals and the activity of antioxidative enzymes [1]. Oxidative stress is considered one of the pathomechanisms of T2DM and its complications [87]. β-cells, similarly to the endothelium and neurons, are vulnerable to reactive oxygen species because of their low antioxidant defense efficiency [88]. T2DM causes microvascular and neurological complications in numerous cases. The most common are diabetic nephropathy, diabetic retinopathy, and diabetic neuropathy. It is known that HHcy, which may be caused by MTHFR polymorphisms, is associated with vascular damage [23]. Elevated levels of Hcy, along with the oxidative stress connected to it, are also responsible for disturbances in neuronal function [89].
MTHFR 677C>T and 1298A>C polymorphisms are associated with HHcy [1]. Because of this, these polymorphisms are thought to be risk factors for the development of T2DM through the aforementioned pathomechanisms. This topic has been explored in several studies. Meng et al. conducted a meta-analysis consisting of 68 studies that found a strong association between the 677C>T polymorphism and T2DM prevalence in the Asian population, but not in Caucasian and African populations [90]. Studies from India and Iran also suggest that this polymorphism may be a risk factor for T2DM [91,92]. However, a recent study from Brazil, performed on 286 patients, suggests that the 677C>T polymorphism is not associated with T2DM [93]. Studies are also inconsistent regarding whether the 1298A>C polymorphism increases the risk of T2DM. Yan et al. observed an association between this polymorphism and an elevated risk of T2DM, especially in the Asian population [94]. However, in a prospective study performed on Chinese adults, the authors concluded that the AC + CC genotypes might have a protective effect against T2DM development [95].
Guan et al. analyzed 28 studies with a total of 4,633 Caucasian and 4,153 Asian patients to explore the potential association between the 677C>T polymorphism and the risk of diabetic nephropathy (DN). The results suggested the existence of such an association in both ethnic groups [96]. However, Elqadi et al. could not link the 677C>T polymorphism with DN risk, although the study had limited statistical power due to factors such as a small sample size (208 patients) and potential issues with generalizability [97].
Settin et al. conducted research on Egyptian patients and observed that the presence of the 677T allele and the 1298C allele is associated with susceptibility to diabetic retinopathy (DR) [98]. Although another study found no such association [97], the results of a meta-analysis performed on 18 studies indicate a significant relationship between the 677C>T polymorphism and DR risk in the Asian population [99].
Zhao et al. conducted an extensive meta-analysis on the influence of eight polymorphisms on the occurrence of diabetic peripheral neuropathy (DPN) and found no relationship between the 677C>T polymorphism and the risk of diabetic neuropathy in Asian and Caucasian populations [89]. In contrast, a study with a smaller sample size (90 patients) concluded that such an association existed [100]. However, Zhao et al. suggested that the 1298A>C polymorphism is associated with DPN in Caucasians [89].
5.2. Gestational Diabetes Mellitus
Gestational diabetes mellitus (GDM) is a condition of impaired glucose tolerance detected for the first time during pregnancy. Its pathomechanisms include insulin resistance (IR) and disturbed proliferation of β-cells. In pregnancy, the main cause of IR is elevated levels of placental hormones [101]. Studies conflict regarding whether HHcy is related to GDM risk [102]. Nonetheless, increased Hcy levels might negatively influence IR, and through this pathway, HHcy could potentially be associated with GDM development [1]. The role of MTHFR polymorphisms, which are responsible for HHcy, has been explored in several studies.
A meta-analysis performed on 17 studies, including 12,345 Chinese patients, shows that the MTHFR 677C>T polymorphism might be positively associated with GDM risk in the southern Chinese population [103]. However, Pham et al., who explored the influence of this polymorphism among Vietnamese pregnant women in their first trimester, concluded that such an association did not exist. However, the sample size of their study was significantly smaller (210 participants) [104]. Dias et al., in their study performed on South African women, also failed to link this polymorphism to GDM, but they found an association between the T allele and higher levels of fasting insulin. The authors suggest that the MTHFR 677C>T polymorphism might participate, among other factors, in the pathomechanisms related to GDM development [105].
5.3. Metabolic Syndrome
Metabolic syndrome (MS) is a combination of five pathologies: insulin resistance, central obesity, hyperglycemia, dyslipidemia, and hypertension. HHcy is considered a risk factor for MS through several pathways, such as oxidative stress and cytotoxicity [106]. One of the main pathomechanisms of MS is associated with insulin resistance (IR) [106], and as mentioned earlier, HHcy contributes to IR [1].
One of the polymorphisms that might be a risk factor for MS development is 677C>T in the MTHFR gene. Azizi et al., in their meta-analysis comprising seven studies conducted between 2002 and 2012, concluded that this polymorphism did not have a significant impact on MS risk [107]. However, a Chinese control study presents conflicting results [108]. The reason for such disagreement may lie in the epigenetics of individuals [107].
5.4. Obesity
Obesity is a chronic disease characterized by excessive fat accumulation, which can lead to various disorders, such as type 2 diabetes mellitus (T2DM). HHcy may be associated with the risk of obesity through processes that disrupt lipid metabolism [109]. Hcy could potentially cause these impairments both directly and indirectly, through endoplasmic reticulum (ER) stress or epigenetic mechanisms [109,110].
Fu et al. analyzed 20 studies involving 38,317 patients and found that the MTHFR 677C>T TT genotype is significantly associated with HHcy, and this homozygous gene variant could be a risk factor for obesity development [110]. However, a study from Mexico, involving 316 young participants (average age 19 years), failed to link the 677C>T polymorphism to obesity. Additionally, authors suggested that this polymorphism might have a protective effect against obesity development, due to lower cholesterol and triglyceride levels in TT carriers [111].
5.5. Non-Alcoholic Fatty Liver Disease
Non-alcoholic fatty liver disease (NAFLD) is a condition characterized by hepatic steatosis, which may progress to non-alcoholic steatohepatitis (NASH) with more severe liver damage [112]. Alcohol consumption is excluded as a cause, as metabolic disturbances primarily drive the pathogenesis of NAFLD. These disturbances include impairments in lipid metabolism pathways that lead to fat accumulation and oxidative stress related to dysfunctional mitochondria [112,113]. Additionally, elevated levels of homocysteine (Hcy) may increase the risk of NAFLD [114]. HHcy, along with epigenetic mechanisms, could be involved in the development of NAFLD [114,115]. Increased Hcy levels contribute to lipid accumulation, endoplasmic reticulum stress, endothelial damage — processes responsible for liver injury [115]. Insulin resistance (IR), which is also linked to NAFLD, might be a consequence of HHcy [1,115]. One potential cause of HHcy could be MTHFR polymorphisms.
A meta-analysis suggests that the 677C>T polymorphism may be a global risk factor for NAFLD, while the 1298A>C polymorphism is associated with NAFLD risk in Caucasians [116]. Similarly, a study of 4096 Chinese patients indicates an increased risk of the disease in individuals with the MTHFR CT and CC genotypes [117]. However, a study involving 1786 patients from Italy and Finland (European population) found no association between these polymorphisms and NAFLD risk [118]. Wang et al. also suggest that the 677C>T polymorphism is not a risk factor for NAFLD, although it does influence Hcy levels. They propose that whether this polymorphism increases the risk of NAFLD may depend on ethnic, geographical, and individual factors in the populations studied [119].
5.6. Inflammatory Bowel Disease
Inflammatory bowel disease (IBD) is a condition characterized by chronic intestinal inflammation, which can be divided into three phenotypes: Crohn's disease (CD), ulcerative colitis (UC), and IBD-unclassified (U-IBD) [120]. Environmental factors, such as smoking and diet, may influence the risk of CD and UC [121]. IBD can also have polygenic, monogenic, or pharmacogenetic origins. Monogenic IBD is more common in children than in adults. However, in a gene panel consisting of 75 genes responsible for monogenic IBD, MTHFR was not included [122]. Despite this, polymorphisms, including those in the MTHFR gene, may be associated with IBD in the adult population, and this relationship has been explored in numerous studies. HHcy, potentially caused by MTHFR polymorphisms, triggers increased expression of molecules involved in inflammatory processes and hypomethylation of proinflammatory genes, resulting in their activation. Elevated Hcy also disrupts cellular antioxidant defense and may be associated with neoangiogenesis in the intestinal mucosa [123]. The results of studies investigating the relationship between MTHFR polymorphisms and IBD are inconsistent. Varzari et al. concluded that the 677C>T and 1298A>C polymorphisms were not associated with the risk of UC in the Moldavian population. However, they found that the 677 TT genotype might be associated with a less severe course of UC. These findings conflicted with previous studies. The authors also found that the 1298AC genotype, but not the 1298CC genotype, was correlated with a more severe subtype of UC [124]. According to Karban et al., among the Non-Ashkenazi Jewish population, the 677C>T polymorphism was found to increase the risk of CD, but not UC [125].
6. MTHFR in Pregnancy and Neonatology
6.1. Infertility
Correct cell division during spermatogenesis and oogenesis requires THF for dTMP synthesis [126], while genomic imprinting depends on SAM and DNA methyltransferases (DNMTs) to create a parent-specific DNA methylation pattern [127]. These epigenetic marks influence zygotic development, fetal growth, and are maintained throughout adult life. Furthermore, histone tail modification and chromatin remodeling are also dependent on folate cycle derivatives. Therefore, MTHFR polymorphisms may contribute to infertility [127]. More et al. examined 127 infertile Indian men, of whom 9.7% were 677C>T mutants, compared to only 1.6% in the fertile control group [128]. Another study, conducted by Ménézo et al., shows that the prevalence of 677C>T and 1298A>C homozygous or compound heterozygous polymorphisms reaches 40% in infertility consultation patients [129]. These patients also have higher Hcy levels, which, according to the authors, increase oxidative stress and methylation errors during gametogenesis [129,130]. On the other hand, Ren et al. demonstrated a weak association between MTHFR polymorphisms and semen abnormalities in a study group of 1,049 men [131]. A meta-analysis by Liu et al. suggests an association between the 677C>T polymorphism and male infertility, but with inconclusive results in European and African populations [130]. Gong et al. show a correlation between increased semen abnormalities and the 677C>T polymorphism in Asians and Europeans [132]. Notably, previous meta-analyses have been inconsistent, and no further meta-analyses have been published in the last 5 years. Further studies are needed to assess whether the findings depend on ethnic background and dietary folate intake, as is strongly suggested.
6.2. Recurrent Pregnancy Loss
HHcy, which frequently occurs in patients with MTHFR polymorphisms [1], is a risk factor for several pregnancy complications [102]. Notably, Hcy levels are lower during pregnancy than in non-pregnant women due to an increase in glomerular filtration rate, plasma volume, and enhanced fetal absorption [133]. Recurrent pregnancy loss (RPL), defined as two or more early miscarriages, can be caused by HHcy, among other factors [102,134]. Hcy induces vascular inflammation and damages placental and uterine blood vessels, leading to thrombosis and subsequent miscarriage [134]. Placental abruption and the resulting fetal hypoxia are often the causes of a miscarriage [133]. A prospective study by Basha et al. found that the prevalence of 677C>T homozygous polymorphisms was significantly higher in women with RPL than in controls (10% vs 2.5%). The 1298A>C polymorphism was also more common in the treatment group (13.3% vs 5%) [135]. The same association was described by Mehta et al. in a meta-analysis that included 5,888 cases and 8,401 controls. Interestingly, the 1298A>C polymorphism was identified as a risk factor for RPL in Caucasians but not in Asians [136]. However, some previous studies have found the association between MTHFR polymorphisms and RPL to be insignificant [137,138].
6.3. Preeclampsia
Endothelial inflammation occurring in HHcy is a potential cause of preeclampsia (PE), a condition with a complex etiology [102,139,140]. It is unclear whether HHcy is a consequence of MTHFR polymorphisms [141], as it can also be associated with hypertension, folate deficiency, drug use, impaired renal function, alcoholism, and smoking [140]. Zhang et al. reported a significant association between HHcy and PE, but MTHFR polymorphisms did not appear to increase Hcy levels [142]. Previous studies have been inconclusive, and few studies have been conducted in the last 10 years [143].
6.4. Preterm Birth and Low Birth Weight
Preterm birth (PTB), defined as delivery before 37 weeks of gestation, may be associated with HHcy and MTHFR polymorphisms [102]. Abnormal placental vasculature and increased hormone secretion in HHcy may accelerate delivery [102,133]. Elevated Hcy levels in preterm newborns seem to support this theory [144]. A study by Huang et al. involving 1,995 pregnant Chinese women revealed a higher prevalence of PTB in mothers who were homozygous for the 677C>T polymorphism. The authors consider hypertensive disorders and gestational diabetes mellitus as potential causes of these PTBs [145], both of which may be caused by HHcy and pose a threat to pregnancy [133]. On the other hand, Wang et al. found MTHFR polymorphisms to be insignificant in the incidence of PTB [146]. Research by Dević Pavlić et al. found no relevant DNA methylation disruptions in MTHFR mutants, although the influence of HHcy was not studied [147]. A meta-analysis conducted by Wu et al. associates PTB with the maternal 677C>T MTHFR polymorphism, particularly in homozygous cases. Notably, a similar association with neonatal polymorphisms has not been established [148]. Some authors suggest that maternal Hcy levels may serve as a potential marker for preterm delivery [144].
Low birth weight (LBW) in newborns may also be associated with MTHFR polymorphisms in the infant or mother, although conclusive evidence has not yet been found. The influence of Hcy on LBW, however, is clearer [133]. Wu et al. found no association between these polymorphisms and LBW [148]. On the other hand, Dipitika et al. described the birth weights of full-term and preterm infants, noting a significant difference between those born to mothers with and without the 677C>T polymorphism [149].
6.5. Neural Tube Defects
The development and closure of the neural tube require derivatives from the folate cycle [150,151]. Mothers with MTHFR polymorphisms are likely to have reduced folate levels, which may result in their children developing neural tube defects (NTDs), such as anencephaly or spina bifida. NTDs can lead to fetal death or lifelong disability [152]. It is also suspected that DNA damage repair systems depend on the folate cycle, and their failure may contribute to the occurrence of NTDs [151]. A meta-analysis of 42 studies by Almekkawi et al. confirms a strong correlation between the MTHFR 677C>T polymorphism and the incidence of NTDs [153]. Furthermore, Li et al. show an association between both maternal and fetal 677C>T polymorphisms and NTDs [154]. On the other hand, a meta-analysis by Soleimani-Jadidi et al. found no association between the maternal MTHFR 1298A>C polymorphism and any type of NTDs [155].
The neural tube closes during the third and fourth weeks of gestation. Therefore, to prevent NTDs, it is important to start folic acid supplementation (400 μg/day) when attempting conception [152]. Folic acid has been successfully used for NTD prevention. However, supplementation with 5-MTHF (the active form of folate) instead of folic acid bypasses the MTHFR block, which is crucial for patients with MTHFR polymorphisms [126]. High-dose folic acid supplementation is also a risk factor for the development of UMFA (unmetabolized folic acid) syndrome, which involves the accumulation of excess folate that could potentially have an oxidizing effect [156,157]. 5-MTHF supplementation is also less likely to cause hematological symptoms associated with vitamin B12 deficiency [157].
7. MTHFR in Rheumatoid Arthritis
Rheumatoid arthritis (RA) is an autoimmune disease characterized by synovitis, which leads to damage to joint cartilage and bone. The etiology of the disease is not precisely defined, but it is generally agreed that both environmental and genetic factors can influence its onset and progression. The disease is more common in women (especially after the age of 65), with a female-to-male ratio of 3:1 [159]. Regarding the genetic factors that affect the risk of RA, MTHFR polymorphisms 677C>T and 1298A>C have been implicated, with their prevalence varying by population. The presence of the 677C>T polymorphism is estimated at 34.1% in Europe, 19.7% in Asia, and 10.3% in Africa [160]. For the 1298A>C polymorphism, the prevalence in Caucasians is 34%, while in Africans it is 9% [161]. A consensus has been reached regarding the impact of the MTHFR 1298A>C polymorphism on an increased risk of RA in the general population. However, the association between the 677C>T polymorphism and RA is inconsistent. The impact of MTHFR polymorphisms was addressed in a meta-analysis by Cen et al., who concluded that the MTHFR 677C>T polymorphism increases genetic susceptibility to RA in Asians, while the MTHFR 1298A>C polymorphism increases susceptibility in the general population [162].
Another meta-analysis found that the MTHFR 677C>T polymorphism is associated with a higher risk of RA in the African population, but no association was found in Caucasians and Asians. In the case of MTHFR 1298A>C, an association with higher susceptibility to RA in the general population was observed. However, the same association was not found in each racial group [163]. Yi et al. indicated that the MTHFR 677C>T and 1298A>C polymorphisms result in higher susceptibility to RA in the general population. When broken down into subgroups, they noted an increased risk of RA in Caucasians and Africans, but not in Asians with the 677C>T polymorphism. With the 1298A>C polymorphism, subgroup analysis showed no significant association with RA in Caucasians and Africans [164].
8. Conclusions
In this review, we have summarized the current state of knowledge on MTHFR polymorphisms and the diseases associated with them. Some of these conditions have been clearly linked to MTHFR polymorphisms, while others require further research. It is important to remember that ethnic background and a traditional diet rich in folate have a significant impact on the results of current studies. In many areas, additional prospective studies are needed to determine whether MTHFR polymorphisms influence disease development and treatment. It has not yet been established whether routine genetic testing for these polymorphisms is necessary or if current treatment regimens should be altered accordingly. Some of the assumptions made in this article are certainly worth pursuing in future studies.
Author Contributions
Conceptualization, D.J.-L., A.F.A., K.J; methodology, A.F.A., K.J., B.B., S.K., D.J.-L.; software, K.J., S.K., P.W.; validation A.F.A., K.J., B.B., S.K., P.W., M.S., D.J.-L.; formal analysis, A.F.A., K.J., B.B., S.K., P.W., M.S., D.J.-L.; investigation A.F.A., K.J., B.B., S.K., P.W., M.S.; writing—original draft preparation A.F.A., K.J., B.B., S.K., P.W., M.S.; writing—review and editing, A.F.A., K.J., B.B., S.K., P.W., M.S., D.J.-L.; visualization A.F.A., K.J., S.K., M.S.; supervision, D.J.-L.; project administration, D.J.-L.; All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The authors of the manuscript hereby formally declare that the research presented herein is based primarily upon publicly accessible scientific sources and scholarly materials acquired through institutional university subscriptions. Furthermore, should any challenges or ambiguities arise regarding access to this manuscript or the source materials referenced within, the authors commit to providing the full text of the work and offering comprehensive assistance to ensure the transparency and accessibility of the research. This declaration is made in adherence to principles of scholarly integrity and with a commitment to facilitating the dissemination and verification of scientific knowledge.
Acknowledgments
Special thanks to Martyna Kubiak for providing support with figure design.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| MDPI | Multidisciplinary Digital Publishing Institute |
| MTHFR | 5,10-methylenetetrahydrofolate reductase; Methylenetetrahydrofolate reductase |
| NTDs | Neural tube defects |
| 5,10-methylene-THF | 5,10-methylenetetrahydrofolate |
| 5-methyl-THF | 5-methyltetrahydrofolate |
| NADPH | Nicotinamide adenine dinucleotide phosphate |
| FAD | Flavin adenine dinucleotide |
| Hcy | Homocysteine |
| Met | Methionine |
| MS | Methionine synthase |
| THF | Tetrahydrofolate |
| 5,10-MTHF | 5,10-methylenetetrahydrofolate |
| 5-MTHF | 5-methyltetrahydrofolate |
| SAM | S-Adenosyl-L-methionine |
| SAH | S-Adenosyl-L-homocysteine |
| dTMP | Deoxythymidine monophosphate |
| DNA | Deoxyribonucleic acid |
| CBS | Cystathionine β-synthase |
| SNPs | Single nucleotide polymorphisms |
| Ala | Alanine |
| Val | Valine |
| Glu | Glutamine |
| HHcy | Hyperhomocysteinemia |
| A | Adenine |
| C | Cytosine |
| T | Thymine |
| G | Guanine |
| NO | Nitric Oxide |
| NMDA | N-methyl-D-aspartate |
| PP2A | Protein phosphatase 2A |
| ALP1 | Apolipoprotein 1 |
| EC | Endothelial cells |
| IL-1β | Interleukin 1 beta |
| IL-6 | Interleukin 6 |
| IL-8 | Interleukin 8 |
| TNF-α | Tumor necrosis factor |
| ROS | Reactive oxygen species |
| NF-κB | Nuclear factor kappa B |
| CVD | Cardiovascular disease |
| IMH | Intramural hematoma |
| OR | Odds Ratio |
| CI | Confidence Interval |
| BC | Breast cancer |
| HER-2 | Receptor tyrosine-protein kinase erbB-2 |
| PC | Prostate cancer |
| OC | Ovarian cancer |
| CLL | Chronic lymphocytic leukemia |
| ALL | Acute lymphocytic leukemia |
| AML | Acute myeloblastic leukemia |
| CML | Chronic myeloblastic leukemia |
| ASD | Autism Spectrum Disorder |
| Fragile X | Fragile X Messenger Ribonucleoprotein 1 |
| SHANK 3 | SH3 and multiple ankyrin repeat domains 3 |
| CASPR 2 | Contactin-associated protein-like 2 |
| RFC1 | Replication factor C subunit 1 |
| FRα | Folate receptor alpha |
| PCFT | Proton-coupled folate transporter |
| AD | Alzheimer's disease |
| EOAD | Early-onset Alzheimer's disease |
| LOAD | Late-onset Alzheimer's disease |
| APP | Amyloid precursor protein |
| PSEN1 | Presenilin 1 |
| PSEN2 | Presenilin 2 |
| APOE | Apolipoprotein E |
| aMCI | amnestic Mild Cognitive Impairment |
| SCZ | Schizophrenia |
| MDD | Major Depressive Disorder |
| TRD | Treatment-Resistant Depression |
| FDA | The United States Food and Drug Administration |
| MID | Migraine with comorbid depression |
| PD | Parkinson’s disease |
| L-DOPA | L-3,4-dihydroxyphenylalanine |
| ADHD | Attention deficit hyperactivity disorder |
| DM | Diabetes mellitus |
| T2DM | Type 2 diabetes mellitus |
| IR | Insulin resistance |
| DN | Diabetic nephropathy |
| DR | Diabetic retinopathy |
| DPN | Diabetic Peripheral Neuropathy |
| GDM | Gestational diabetes mellitus |
| MS | Metabolic syndrome |
| ER | Endoplasmic reticulum |
| NAFLD | Non-alcoholic fatty liver disease |
| NASH | Non-alcoholic steatohepatitis |
| IBD | Inflammatory bowel disease |
| CD | Crohn's disease |
| UC | Ulcerative colitis |
| U-IBD | Unclassified inflammatory bowel disease |
| DNMTs | DNA methyltransferases |
| RPL | Recurrent pregnancy loss |
| PE | Preeclampsia |
| PTB | Preterm Birth |
| LBW | Low birth weight |
| UMFA | Unmetabolized folic acid |
| RA | Rheumatoid arthritis |
References
- Zarembska, E.; Ślusarczyk, K.; Wrzosek, M. The Implication of a Polymorphism in the Methylenetetrahydrofolate Reductase Gene in Homocysteine Metabolism and Related Civilisation Diseases. Int J Mol Sci 2023, 25, 193. [CrossRef]
- Botto, L.D.; Yang, Q. 5,10-Methylenetetrahydrofolate Reductase Gene Variants and Congenital Anomalies: A HuGE Review. Am J Epidemiol 2000, 151, 862–877. [CrossRef]
- Froese, D.S.; Fowler, B.; Baumgartner, M.R. Vitamin B12 , Folate, and the Methionine Remethylation Cycle—Biochemistry, Pathways, and Regulation. J of Inher Metab Disea 2019, 42, 673–685. [CrossRef]
- 4. MTHFR Methylenetetrahydrofolate Reductase [Homo Sapiens (Human)] - Gene - NCBI. Available online: https://www.ncbi.nlm.nih.gov/gene/4524 (accessed on 12 March 2025).
- Mallhi, T.H.; Shahid, M.; Rehman, K.; Khan, Y.H.; Alanazi, A.S.; Alotaibi, N.H.; Akash, M.S.H.; Butt, M.H. Biochemical Association of MTHFR C677T Polymorphism with Myocardial Infarction in the Presence of Diabetes Mellitus as a Risk Factor. Metabolites 2023, 13, 251. [CrossRef]
- Singh, J.; Wilkins, G.; Goodman-Vincent, E.; Chishti, S.; Bonilla Guerrero, R.; McFadden, L.; Zahavi, Z.; Santosh, P. Co-Occurring Methylenetetrahydrofolate Reductase (MTHFR) Rs1801133 and Rs1801131 Genotypes as Associative Genetic Modifiers of Clinical Severity in Rett Syndrome. Brain Sciences 2024, 14, 624. [CrossRef]
- Barretta, F.; Uomo, F.; Fecarotta, S.; Albano, L.; Crisci, D.; Verde, A.; Fisco, M.G.; Gallo, G.; Dottore Stagna, D.; Pricolo, M.R.; et al. Contribution of Genetic Test to Early Diagnosis of Methylenetetrahydrofolate Reductase (MTHFR) Deficiency: The Experience of a Reference Center in Southern Italy. Genes 2023, 14, 980. [CrossRef]
- Leclerc, D.; Sibani, S.; Rozen, R. Molecular Biology of Methylenetetrahydrofolate Reductase (MTHFR) and Overview of Mutations/Polymorphisms. Madame Curie Bioscience Database [Internet]. Austin (TX): Landes Bioscience; 2000-2013. Available online: https://www.ncbi.nlm.nih.gov/books/NBK6561/ (accessed on 12 March 2025).
- Yamada, K.; Chen, Z.; Rozen, R.; Matthews, R.G. Effects of Common Polymorphisms on the Properties of Recombinant Human Methylenetetrahydrofolate Reductase. Proc Natl Acad Sci U S A 2001, 98, 14853–14858. [CrossRef]
- Bhatia, P.; Singh, N. Homocysteine Excess: Delineating the Possible Mechanism of Neurotoxicity and Depression. Fundamemntal Clinical Pharma 2015, 29, 522–528. [CrossRef]
- Nishio, K.; Goto, Y.; Kondo, T.; Ito, S.; Ishida, Y.; Kawai, S.; Naito, M.; Wakai, K.; Hamajima, N. Serum Folate and Methylenetetrahydrofolate Reductase (MTHFR) C677T Polymorphism Adjusted for Folate Intake. J Epidemiol 2008, 18, 125–131. [CrossRef]
- Bjørke-Monsen, A.-L.; Ueland, P.M. Folate – a Scoping Review for Nordic Nutrition Recommendations 2023. Food Nutr Res 2023, 67, 10.29219/fnr.v67.10258. [CrossRef]
- Román, G.C.; Mancera-Páez, O.; Bernal, C. Epigenetic Factors in Late-Onset Alzheimer’s Disease: MTHFR and CTH Gene Polymorphisms, Metabolic Transsulfuration and Methylation Pathways, and B Vitamins. Int J Mol Sci 2019, 20, 319. [CrossRef]
- Smith, A.D.; Refsum, H. Homocysteine, B Vitamins, and Cognitive Impairment. Annual Review of Nutrition 2016, 36, 211–239. [CrossRef]
- Austin, R.C.; Lentz, S.R.; Werstuck, G.H. Role of Hyperhomocysteinemia in Endothelial Dysfunction and Atherothrombotic Disease. Cell Death & Differentiation 2004, 11, S56–S64. [CrossRef]
- Yuan, D.; Chu, J.; Lin, H.; Zhu, G.; Qian, J.; Yu, Y.; Yao, T.; Ping, F.; Chen, F.; Liu, X. Mechanism of Homocysteine-Mediated Endothelial Injury and Its Consequences for Atherosclerosis. Front Cardiovasc Med 2022, 9, 1109445. [CrossRef]
- Sun, T.; Ghosh, A.K.; Eren, M.; Miyata, T.; Vaughan, D.E. PAI-1 Contributes to Homocysteine-Induced Cellular Senescence. Cell Signal 2019, 64, 109394. [CrossRef]
- Kaplan, P.; Tatarkova, Z.; Sivonova, M.K.; Racay, P.; Lehotsky, J. Homocysteine and Mitochondria in Cardiovascular and Cerebrovascular Systems. Int J Mol Sci 2020, 21, 7698. [CrossRef]
- Timkova, V.; Tatarkova, Z.; Lehotsky, J.; Racay, P.; Dobrota, D.; Kaplan, P. Effects of Mild Hyperhomocysteinemia on Electron Transport Chain Complexes, Oxidative Stress, and Protein Expression in Rat Cardiac Mitochondria. Molecular and Cellular Biochemistry 2016, 411, 261–270. [CrossRef]
- Perła-Kaján, J.; Marczak, Ł.; Kaján, L.; Skowronek, P.; Twardowski, T.; Jakubowski, H. Modification by Homocysteine Thiolactone Affects Redox Status of Cytochrome c. Biochemistry 2007, 46, 6225–6231. [CrossRef]
- Chang, L.; Geng, B.; Yu, F.; Zhao, J.; Jiang, H.; Du, J.; Tang, C. Hydrogen Sulfide Inhibits Myocardial Injury Induced by Homocysteine in Rats. Amino Acids 2008, 34, 573–585. [CrossRef]
- Schwarz, K.; Siddiqi, N.; Singh, S.; Neil, C.J.; Dawson, D.K.; Frenneaux, M.P. The Breathing Heart - Mitochondrial Respiratory Chain Dysfunction in Cardiac Disease. Int J Cardiol 2014, 171, 134–143. [CrossRef]
- Raghubeer, S.; Matsha, T.E. Methylenetetrahydrofolate (MTHFR), the One-Carbon Cycle, and Cardiovascular Risks. Nutrients 2021, 13, 4562. [CrossRef]
- Moti Wala, S.; AlEdani, E.M.; Samuel, E.A.; Ahmad, K.; Manongi, N.J.; Rajapandian, R.; Khan, S. Exploring the Nexus: A Systematic Review on the Interplay of the Methylenetetrahydrofolate Reductase (MTHFR) Gene C677T Genotype, Hyperhomocysteinemia, and Spontaneous Cervical/Vertebral Artery Dissection in Young Adults. Cureus 2024, 16, e60878. [CrossRef]
- Liao, S.; Guo, S.; Ma, R.; He, J.; Yan, Y.; Zhang, X.; Wang, X.; Cao, B.; Guo, H. Association between Methylenetetrahydrofolate Reductase (MTHFR) C677T Polymorphism and H-Type Hypertension: A Systematic Review and Meta-Analysis. Ann Hum Genet 2022, 86, 278–289. [CrossRef]
- Bax, M.; Romanov, V.; Junday, K.; Giannoulatou, E.; Martinac, B.; Kovacic, J.C.; Liu, R.; Iismaa, S.E.; Graham, R.M. Arterial Dissections: Common Features and New Perspectives. Front Cardiovasc Med 2022, 9, 1055862. [CrossRef]
- Ahmed, S.; Bogiatzi, C.; Hackam, D.G.; Rutledge, A.C.; Sposato, L.A.; Khaw, A.; Mandzia, J.; Azarpazhoo, M.R.; Hachinski, V.; Spence, J.D. Vitamin B 12 Deficiency and Hyperhomocysteinaemia in Outpatients with Stroke or Transient Ischaemic Attack: A Cohort Study at an Academic Medical Centre. BMJ Open 2019, 9, e026564. [CrossRef]
- Park, W.-C.; Chang, J.-H. Clinical Implications of Methylenetetrahydrofolate Reductase Mutations and Plasma Homocysteine Levels in Patients with Thromboembolic Occlusion. Vasc Specialist Int 2014, 30, 113–119. [CrossRef]
- Qin, X.; Spence, J.D.; Li, J.; Zhang, Y.; Li, Y.; Sun, N.; Liang, M.; Song, Y.; Zhang, Y.; Wang, B.; et al. Interaction of Serum Vitamin B12 and Folate with MTHFR Genotypes on Risk of Ischemic Stroke. Neurology 2020, 94, e1126–e1136. [CrossRef]
- Zhao, L.; Li, T.; Dang, M.; Li, Y.; Fan, H.; Hao, Q.; Song, D.; Lu, J.; Lu, Z.; Jian, Y.; et al. Association of Methylenetetrahydrofolate Reductase (MTHFR) Rs1801133 (677C>T) Gene Polymorphism with Ischemic Stroke Risk in Different Populations: An Updated Meta-Analysis. Front Genet 2022, 13, 1021423. [CrossRef]
- Song, Y.; Li, B.; Wang, C.; Wang, P.; Gao, X.; Liu, G. Association between 5,10-Methylenetetrahydrofolate Reductase C677T Gene Polymorphism and Risk of Ischemic Stroke: A Meta-Analysis. Journal of Stroke and Cerebrovascular Diseases 2016, 25, 679–687. [CrossRef]
- Chang, G.; Kuai, Z.; Wang, J.; Wu, J.; Xu, K.; Yuan, Y.; Hu, Y. The Association of MTHFR C677T Variant with Increased Risk of Ischemic Stroke in the Elderly Population: A Meta-Analysis of Observational Studies. BMC Geriatrics 2019, 19, 331. [CrossRef]
- Pieroth, R.; Paver, S.; Day, S.; Lammersfeld, C. Folate and Its Impact on Cancer Risk. Curr Nutr Rep 2018, 7, 70–84. [CrossRef]
- Jacques, P.F.; Bostom, A.G.; Williams, R.R.; Ellison, R.C.; Eckfeldt, J.H.; Rosenberg, I.H.; Selhub, J.; Rozen, R. Relation between Folate Status, a Common Mutation in Methylenetetrahydrofolate Reductase, and Plasma Homocysteine Concentrations. Circulation 1996, 93, 7–9. [CrossRef]
- Aly, R.M.; Taalab, M.M.; Ghazy, H.F. MTHFR A1298C and C677T Gene Polymorphisms and Susceptibility to Chronic Myeloid Leukemia in Egypt. Int J Clin Exp Pathol 2014, 7, 2571–2578.
- Wainfan, E.; Poirier, L.A. Methyl Groups in Carcinogenesis: Effects on DNA Methylation and Gene Expression. Cancer Res 1992, 52, 2071s–2077s.
- Sanjoaquin, M.A.; Allen, N.; Couto, E.; Roddam, A.W.; Key, T.J. Folate Intake and Colorectal Cancer Risk: A Meta-Analytical Approach. Int J Cancer 2005, 113, 825–828. [CrossRef]
- Zara-Lopes, T.; Silva Galbiatti-Dias, A.L.; Urbanin Castanhole-Nunes, M.M.; Padovani-Júnior, J.A.; Maniglia, J.V.; Pavarino, E.C.; Goloni-Bertollo, E.M. Polymorphisms in MTHFR, MTR, RFC1 and CßS Genes Involved in Folate Metabolism and Thyroid Cancer: A Case-Control Study. aoms 2019, 15, 522–530. [CrossRef]
- Zhao, T.; Gu, D.; Xu, Z.; Huo, X.; Shen, L.; Wang, C.; Tang, Y.; Wu, P.; He, J.; Gong, W.; et al. Polymorphism in One-Carbon Metabolism Pathway Affects Survival of Gastric Cancer Patients: Large and Comprehensive Study. Oncotarget 2015, 6, 9564–9576. [CrossRef]
- Yi, K.; Yang, L.; Lan, Z.; Xi, M. The Association between MTHFR Polymorphisms and Cervical Cancer Risk: A System Review and Meta Analysis. Arch Gynecol Obstet 2016, 294, 579–588. [CrossRef]
- Purnomo, A.F.; Daryanto, B.; Seputra, K.P.; Budaya, T.N.; Lutfiana, N.C.; Nurkolis, F.; Chung, S.; Suh, J.Y.; Park, M.N.; Seo, B.-K.; et al. Methylenetetrahydrofolate Reductase C677T (Rs1801133) Polymorphism Is Associated with Bladder Cancer in Asian Population: Epigenetic Meta-Analysis as Precision Medicine Approach. Cancers (Basel) 2023, 15, 4402. [CrossRef]
- Niu, Y.-M.; Deng, M.-H.; Chen, W.; Zeng, X.-T.; Luo, J. MTHFR C677T Gene Polymorphism and Head and Neck Cancer Risk: A Meta-Analysis Based on 23 Publications. Dis Markers 2015, 2015, 681313. [CrossRef]
- Jamshidi, M.; Mohammadi Pour, S.; Mahmoudian-Sani, M.-R. Single Nucleotide Variants Associated with Colorectal Cancer Among Iranian Patients: A Narrative Review. Pharmgenomics Pers Med 2020, 13, 167–180. [CrossRef]
- Wang, Y.; Huo, L.; Yang, C.; He, X. Methylenetetrahydrofolate Reductase C677T and A1298C Polymorphisms and Gastric Cancer Susceptibility: An Updated Meta-Analysis. Biosci Rep 2023, 43, BSR20222553. [CrossRef]
- Petrone, I.; Bernardo, P.S.; Dos Santos, E.C.; Abdelhay, E. MTHFR C677T and A1298C Polymorphisms in Breast Cancer, Gliomas and Gastric Cancer: A Review. Genes (Basel) 2021, 12, 587. [CrossRef]
- Akilzhanova, A.; Nurkina, Z.; Momynaliev, K.; Ramanculov, E.; Zhumadilov, Z.; Rakhypbekov, T.; Hayashida, N.; Nakashima, M.; Takamura, N. Genetic Profile and Determinants of Homocysteine Levels in Kazakhstan Patients with Breast Cancer. Anticancer Res 2013, 33, 4049–4059.
- Ajaz, S.; Zaidi, S.-Z.; Mehboob Ali, S.; Siddiqa, A.; Memon, M.A.; Abid, A.; Khaliq, S. Independent, Diplotype and Haplotype Association Analyses of the Selected MTHFR SNPs with the Risk of Breast Cancers in a South-Asian Population. medRxiv 2021. [CrossRef]
- Zara-Lopes, T.; Gimenez-Martins, A.P.A.; Nascimento-Filho, C.H.V.; Castanhole-Nunes, M.M.U.; Galbiatti-Dias, A.L.S.; Padovani-Júnior, J.A.; Maniglia, J.V.; Francisco, J.L.E.; Pavarino, E.C.; Goloni-Bertollo, E.M. Role of MTHFR C677T and MTR A2756G Polymorphisms in Thyroid and Breast Cancer Development. Genet. Mol. Res. 2016, 15. [CrossRef]
- Castiglia, P.; Sanna, V.; Azara, A.; De Miglio, M.R.; Murgia, L.; Pira, G.; Sanges, F.; Fancellu, A.; Carru, C.; Bisail, M.; et al. Methylenetetrahydrofolate Reductase (MTHFR) C677T and A1298C Polymorphisms in Breast Cancer: A Sardinian Preliminary Case-Control Study. Int J Med Sci 2019, 16, 1089–1095. [CrossRef]
- Kumar, P.; Yadav, U.; Rai, V. Methylenetetrahydrofolate Reductase Gene C677T Polymorphism and Breast Cancer Risk: Evidence for Genetic Susceptibility. Meta Gene 2015, 6, 72–84. [CrossRef]
- You, J.; Huang, Y.; Shen, X.; Chen, Y.; Ding, X. Associations between MTHFR Gene Polymorphisms (C677T and A1298C) and Genetic Susceptibility to Prostate Cancer: A Systematic Review and Meta-Analysis. Front Genet 2024, 15, 1343687. [CrossRef]
- Cicek, M.S.; Nock, N.L.; Li, L.; Conti, D.V.; Casey, G.; Witte, J.S. Relationship between Methylenetetrahydrofolate Reductase C677T and A1298C Genotypes and Haplotypes and Prostate Cancer Risk and Aggressiveness. Cancer Epidemiol Biomarkers Prev 2004, 13, 1331–1336.
- Singal, R.; Ferdinand, L.; Das, P.; Reis, I.; Schlesselman, J. Polymorphisms in the Methylenetetrahydrofolate Reductase Gene and Prostate Cancer Risk. Int J Oncol 2004. [CrossRef]
- Stevens, V.L.; Rodriguez, C.; Sun, J.; Talbot, J.T.; Thun, M.J.; Calle, E.E. No Association of Single Nucleotide Polymorphisms in One-Carbon Metabolism Genes with Prostate Cancer Risk. Cancer Epidemiol Biomarkers Prev 2008, 17, 3612–3614. [CrossRef]
- Xiong, Y.; Bian, C.; Lin, X.; Wang, X.; Xu, K.; Zhao, X. Methylenetetrahydrofolate Reductase Gene Polymorphisms in the Risk of Polycystic Ovary Syndrome and Ovarian Cancer. Biosci Rep 2020, 40, BSR20200995. [CrossRef]
- Qin, Y.-T.; Zhang, Y.; Wu, F.; Su, Y.; Lu, G.-N.; Wang, R.-S. Association between MTHFR Polymorphisms and Acute Myeloid Leukemia Risk: A Meta-Analysis. PLoS One 2014, 9, e88823. [CrossRef]
- Sazawal, S.; Chaubey, R.; Kaur, P.; Chikkara, S.; Kumar, B.; Bakshi, S.; Arya, L.S.; Raina, V.; Das Gupta, A.; Saxena, R. MTHFR Gene Polymorphisms and the Risk of Acute Lymphoblastic Leukemia in Adults and Children: A Case Control Study in India. Indian J Hematol Blood Transfus 2014, 30, 219–225. [CrossRef]
- Kim, H.N.; Kim, Y.-K.; Lee, I.-K.; Yang, D.-H.; Lee, J.-J.; Shin, M.-H.; Park, K.-S.; Choi, J.-S.; Park, M.R.; Jo, D.Y.; et al. Association between Polymorphisms of Folate-Metabolizing Enzymes and Hematological Malignancies. Leuk Res 2009, 33, 82–87. [CrossRef]
- Pereira, T.V.; Rudnicki, M.; Pereira, A.C.; Pombo-de-Oliveira, M.S.; Franco, R.F. 5,10-Methylenetetrahydrofolate Reductase Polymorphisms and Acute Lymphoblastic Leukemia Risk: A Meta-Analysis. Cancer Epidemiol Biomarkers Prev 2006, 15, 1956–1963. [CrossRef]
- Raoufi, A.; Rahimi Kelarijani, B.; Ahadi, H.R.; Hassani Derakhshandeh, B.; Nooroollahzadeh, Z.; Hajifathali, A. Association of MTHFR C677T and A1298C Polymorphisms with Susceptibility to Chronic Lymphocytic Leukemia: A Systematic Review and Meta-Analysis. Iran J Public Health 2021, 50, 83–92. [CrossRef]
- Lien, S.-Y.A.; Young, L.; Gau, B.-S.; K Shiao, S.P. Meta-Prediction of MTHFR Gene Polymorphism-Mutations, Air Pollution, and Risks of Leukemia among World Populations. Oncotarget 2017, 8, 4387–4398. [CrossRef]
- Periñán, M.T.; Macías-García, D.; Jesús, S.; Martín-Rodríguez, J.F.; Muñoz-Delgado, L.; Jimenez-Jaraba, M.V.; Buiza-Rueda, D.; Bonilla-Toribio, M.; Adarmes-Gómez, A.D.; Gómez-Garre, P.; et al. Homocysteine Levels, Genetic Background, and Cognitive Impairment in Parkinson’s Disease. J Neurol 2023, 270, 477–485. [CrossRef]
- Zhang, Y.; Zhao, J.; Wang, W.; Fan, W.; Tang, W.; Zhang, C. Homocysteine, but Not MTHFR Gene Polymorphism, Influences Depressive Symptoms in Patients with Schizophrenia. Journal of Affective Disorders 2020, 272, 24–27. [CrossRef]
- Hoxha, B.; Hoxha, M.; Domi, E.; Gervasoni, J.; Persichilli, S.; Malaj, V.; Zappacosta, B. Folic Acid and Autism: A Systematic Review of the Current State of Knowledge. Cells 2021, 10, 1976. [CrossRef]
- Khan, S.; Naeem, A.; Fritts, A.; Cummins, M.; Kayes, C.; Fang, W. Discovery of Methylenetetrahydrofolate Reductase (MTHFR) Deficiency in Individuals With Common Psychiatric Comorbidities: A Retrospective Case Review. Cureus 2024, 16, e58122. [CrossRef]
- Meng, X.; Zheng, J.-L.; Sun, M.-L.; Lai, H.-Y.; Wang, B.-J.; Yao, J.; Wang, H. Association between MTHFR (677C>T and 1298A>C) Polymorphisms and Psychiatric Disorder: A Meta-Analysis. PLoS One 2022, 17, e0271170. [CrossRef]
- Roufael, M.; Bitar, T.; Sacre, Y.; Andres, C.; Hleihel, W. Folate-Methionine Cycle Disruptions in ASD Patients and Possible Interventions: A Systematic Review. Genes (Basel) 2023, 14, 709. [CrossRef]
- Qiu, S.; Qiu, Y.; Li, Y.; Cong, X. Genetics of Autism Spectrum Disorder: An Umbrella Review of Systematic Reviews and Meta-Analyses. Transl Psychiatry 2022, 12, 249. [CrossRef]
- Li, Y.; Qiu, S.; Shi, J.; Guo, Y.; Li, Z.; Cheng, Y.; Liu, Y. Association between MTHFR C677T/A1298C and Susceptibility to Autism Spectrum Disorders: A Meta-Analysis. BMC Pediatr 2020, 20, 449. [CrossRef]
- Razi, B.; Imani, D.; Hassanzadeh Makoui, M.; Rezaei, R.; Aslani, S. Association between MTHFR Gene Polymorphism and Susceptibility to Autism Spectrum Disorders: Systematic Review and Meta-Analysis. Research in Autism Spectrum Disorders 2020, 70, 101473. [CrossRef]
- Tisato, V.; Silva, J.A.; Longo, G.; Gallo, I.; Singh, A.V.; Milani, D.; Gemmati, D. Genetics and Epigenetics of One-Carbon Metabolism Pathway in Autism Spectrum Disorder: A Sex-Specific Brain Epigenome? Genes (Basel) 2021, 12, 782. [CrossRef]
- Lam, N.S.K.; Long, X.X.; Li, X.; Saad, M.; Lim, F.; Doery, J.C.; Griffin, R.C.; Galletly, C. The Potential Use of Folate and Its Derivatives in Treating Psychiatric Disorders: A Systematic Review. Biomed Pharmacother 2022, 146, 112541. [CrossRef]
- You, M.; Zhou, X.; Yin, W.; Wan, K.; Zhang, W.; Li, C.; Li, M.; Zhu, W.; Zhu, X.; Sun, Z. The Influence of MTHFR Polymorphism on Gray Matter Volume in Patients With Amnestic Mild Cognitive Impairment. Front. Neurosci. 2021, 15, 778123. [CrossRef]
- Jiang, Y.; Xiao, X.; Wen, Y.; Wan, M.; Zhou, L.; Liu, X.; Wang, X.; Guo, L.; Liu, H.; Zhou, Y.; et al. Genetic Effect of MTHFR C677T, A1298C, and A1793G Polymorphisms on the Age at Onset, Plasma Homocysteine, and White Matter Lesions in Alzheimer’s Disease in the Chinese Population. Aging (Albany NY) 2021, 13, 11352–11362. [CrossRef]
- Sun, J.; Jiang, X.; Zhao, M.; Ma, L.; Pei, H.; Liu, N.; Li, H. Association of Methylenetetrahydrofolate Reductase C677T Gene Polymorphisms with Mild Cognitive Impairment Susceptibility: A Systematic Review and Meta-Analysis. Behav Neurol 2021, 2021, 2962792. [CrossRef]
- Yoo, S.; Montazeri, A.; McNulty, H.; Kent, M.P.; Little, J. Is There a Causal Link between Folate Status and Schizophrenia? Evidence from Genetic Association Studies. Nutr Neurosci 2024, 1–12. [CrossRef]
- Zhang, Y.-X.; Yang, L.-P.; Gai, C.; Cheng, C.-C.; Guo, Z.-Y.; Sun, H.-M.; Hu, D. Association between Variants of MTHFR Genes and Psychiatric Disorders: A Meta-Analysis. Front Psychiatry 2022, 13, 976428. [CrossRef]
- Liu, J.H.; Zhu, C.; Zheng, K.; Tang, W.; Gao, L.L.; Trihn, T.H.; Emily Wu, H.; Chen, D.C.; Hong Xiu, M.; Yang Zhang, X. MTHFR Ala222Val Polymorphism and Clinical Characteristics Confer Susceptibility to Suicide Attempt in Chronic Patients with Schizophrenia. Sci Rep 2020, 10, 5008. [CrossRef]
- Halaris, A.; Sohl, E.; Whitham, E.A. Treatment-Resistant Depression Revisited: A Glimmer of Hope. J Pers Med 2021, 11, 155. [CrossRef]
- Asif, N.; Patel, A.; Vedantam, D.; Poman, D.S.; Motwani, L. Migraine With Comorbid Depression: Pathogenesis, Clinical Implications, and Treatment. Cureus 2022, 14, e25998. [CrossRef]
- Vuletić, V.; Rački, V.; Papić, E.; Peterlin, B. A Systematic Review of Parkinson’s Disease Pharmacogenomics: Is There Time for Translation into the Clinics? Int J Mol Sci 2021, 22, 7213. [CrossRef]
- Sadeghiyeh, T.; Dastgheib, S.A.; Lookzadeh, M.H.; Noori-Shadkam, M.; Akbarian-Bafghi, M.J.; Zare-Shehneh, M.; Poursharif, Z.; Neamatzadeh, H. Association of MTHFR 677C > T and 1298A > C Polymorphisms with Susceptibility to Attention Deficit and Hyperactivity Disorder. Fetal and Pediatric Pathology 2020, 39, 422–429. [CrossRef]
- Augoulea, A.; Armeni, E.; Deligeoroglou, E.; Paschou, S.A.; Papadimitriou, G.; Stergioti, E.; Karountzos, V.; Tsitsika, A.; Panoulis, K.; Economou, E.; et al. MTHFR Polymorphisms in Girls with Anorexia Nervosa: Implications on Body Weight. Endocrine Research 2021, 46, 80–85. [CrossRef]
- Global diabetes data report 2000 — 2045. Available online: https://diabetesatlas.org/data/en/world/ (accessed on 12 March 2025).
- Cai, Y.; Liu, B.; Zhang, Y.; Zhou, Y. MTHFR Gene Polymorphisms in Diabetes Mellitus. Clin Chim Acta 2024, 561, 119825. [CrossRef]
- Diabetes Facts and Figures. Available online: https://idf.org/about-diabetes/diabetes-facts-figures/ (accessed on 12 March 2025).
- Bigagli, E.; Lodovici, M. Circulating Oxidative Stress Biomarkers in Clinical Studies on Type 2 Diabetes and Its Complications. Oxid Med Cell Longev 2019, 2019, 5953685. [CrossRef]
- Schwartz, S.S.; Epstein, S.; Corkey, B.E.; Grant, S.F.A.; Gavin Iii, J.R.; Aguilar, R.B.; Herman, M.E. A Unified Pathophysiological Construct of Diabetes and Its Complications. Trends Endocrinol Metab 2017, 28, 645–655. [CrossRef]
- Zhao, Y.; Zhu, R.; Wang, D.; Liu, X. Genetics of Diabetic Neuropathy: Systematic Review, Meta-Analysis and Trial Sequential Analysis. Ann Clin Transl Neurol 2019, 6, 1996–2013. [CrossRef]
- Meng, Y.; Liu, X.; Ma, K.; Zhang, L.; Lu, M.; Zhao, M.; Guan, M.; Qin, G. Association of MTHFR C677T Polymorphism and Type 2 Diabetes Mellitus (T2DM) Susceptibility. Molec Gen & Gen Med 2019, 7, e1020. [CrossRef]
- Pathak, D.; Shrivastav, D.; Verma, A.K.; Alsayegh, A.A.; Yadav, P.; Khan, N.H.; Al-Harbi, A.I.; Khan, M.I.; Bihade, K.; Singh, D.D.; et al. Role of Metabolizing MTHFR Gene Polymorphism (Rs1801133) and Its mRNA Expression among Type 2 Diabetes. J Diabetes Metab Disord 2022, 21, 511–516. [CrossRef]
- Rahimi, A.; Moridi, N.; Golestani, A.; Anani-Sarab, G.; Salmani, F.; Yaqubi, G.; Mesbahzadeh, B.; Jalalifar, M.A.; Malekaneh, M.; Sajjadi, S.M. Factor V Leiden, MTHFR, and FXIIIVal34Leu Gene Polymorphisms and Their Association with Clinical Features and Risk of Diabetic Retinopathy in Patients with Type 2 Diabetes. Caspian J Intern Med 2024, 15, 101–108. [CrossRef]
- Santos, K.F.; Assunção, L.P.; Santos, R.S.; Reis, A. a. S. Machine Learning Approaches and Genetic Determinants That Influence the Development of Type 2 Diabetes Mellitus: A Genetic Association Study in Brazilian Patients. Braz J Med Biol Res 2024, 57, e13957. [CrossRef]
- Yan, Y.; Liang, H.; Yang, S.; Wang, J.; Xie, L.; Qin, X.; Li, S. Methylenetetrahydrofolate Reductase A1298C Polymorphism and Diabetes Risk: Evidence from a Meta-Analysis. Ren Fail 2014, 36, 1013–1017. [CrossRef]
- Lu, X.; Huang, R.; Li, S.; Fang, A.; Chen, Y.; Chen, S.; Wang, F.; Lin, X.; Liu, Z.; Zhu, H. Associations between Serum Betaine, Methyl-Metabolizing Genetic Polymorphisms and Risk of Incident Type 2 Diabetes: A Prospective Cohort Study in Community-Dwelling Chinese Adults. Nutrients 2022, 14, 362. [CrossRef]
- Guan, H.; Xia, M.-D.; Wang, M.; Guan, Y.-J.; Lyu, X.-C. Methylenetetrahydrofolate Reductase Genetic Polymorphism and the Risk of Diabetic Nephropathy in Type 2 Diabetic Patients. Medicine (Baltimore) 2020, 99, e21558. [CrossRef]
- Elqadi, M.; Eweidat, K.; Abu Sabha, M.; Yagmour, A.; Sabarneh, A.; Nasereddin, A.; Ereqat, S. Methylenetetrahydrofolate Reductase C677T Gene Polymorphism and the Association with Dyslipidemia in Type 2 Diabetic Palestinian Patients. J Clin Lab Anal 2021, 35, e23994. [CrossRef]
- Settin, A.; El-Baz, R.; Ismaeel, A.; Tolba, W.; Allah, W.A. Association of ACE and MTHFR Genetic Polymorphisms with Type 2 Diabetes Mellitus: Susceptibility and Complications. J Renin Angiotensin Aldosterone Syst 2015, 16, 838–843. [CrossRef]
- Luo, S.; Wang, F.; Shi, C.; Wu, Z. A Meta-Analysis of Association between Methylenetetrahydrofolate Reductase Gene (MTHFR) 677C/T Polymorphism and Diabetic Retinopathy. Int J Environ Res Public Health 2016, 13, 806. [CrossRef]
- Yehya, A.; Altaany, Z.; Beni-Yonis, O. Dual Mapping of MTHFR C677T (A1298C) and BDNF G196A (Val66Met) Polymorphisms in Patients with Diabetic Peripheral Neuropathy. Eur Rev Med Pharmacol Sci 2023, 27, 6682–6690. [CrossRef]
- Sweeting, A.; Wong, J.; Murphy, H.R.; Ross, G.P. A Clinical Update on Gestational Diabetes Mellitus. Endocr Rev 2022, 43, 763–793. [CrossRef]
- Dai, C.; Fei, Y.; Li, J.; Shi, Y.; Yang, X. A Novel Review of Homocysteine and Pregnancy Complications. Biomed Res Int 2021, 2021, 6652231. [CrossRef]
- Tan, X.; Chen, H. Association between MTHFR Gene C677T Polymorphism and Gestational Diabetes Mellitus in Chinese Population: A Meta-Analysis. Front Endocrinol (Lausanne) 2023, 14, 1273218. [CrossRef]
- Pham, N.T.N.; Huynh, C.T.N.; Nguyen, A.T.T.; Ho, C.Q.; Duong, L.M.; Bui, D.T.; Nguyen, H.H. Pre-Gestational Diabetes Mellitus, Gestational Diabetes Mellitus, and Its Association with the MTHFR C677T Polymorphism. Medicine (Baltimore) 2024, 103, e38648. [CrossRef]
- Dias, S.; Adam, S.; Rheeder, P.; Pheiffer, C. No Association Between ADIPOQ or MTHFR Polymorphisms and Gestational Diabetes Mellitus in South African Women. Diabetes Metab Syndr Obes 2021, 14, 791–800. [CrossRef]
- Ulloque-Badaracco, J.R.; Hernandez-Bustamante, E.A.; Alarcon-Braga, E.A.; Al-Kassab-Córdova, A.; Cabrera-Guzmán, J.C.; Herrera-Añazco, P.; Benites-Zapata, V.A. Vitamin B12, Folate, and Homocysteine in Metabolic Syndrome: A Systematic Review and Meta-Analysis. Front Endocrinol (Lausanne) 2023, 14, 1221259. [CrossRef]
- Azizi, S.; Shamshirian, A.; Alizadeh-Navaei, R.; Jafarpour, H.; Asemi, Z.; Tamtaji, O.R.; Vaziri, M.S.; Homayounfar, R.; Rezaei Shahmirzadi, A.; Alipoor, R. A Genetic Association Study of MTHFR C677T Polymorphism with Risk of Metabolic Syndrome: A Systematic Review and Meta-Analysis. Galen Med J 2019, 8, e1472. [CrossRef]
- Wang, J.; Xu, L.; Xia, H.; Li, Y.; Tang, S. Association of MTHFR C677T Gene Polymorphism with Metabolic Syndrome in a Chinese Population: A Case-Control Study. J Int Med Res 2018, 46, 2658–2669. [CrossRef]
- Wang, J.; You, D.; Wang, H.; Yang, Y.; Zhang, D.; Lv, J.; Luo, S.; Liao, R.; Ma, L. Association between Homocysteine and Obesity: A Meta-Analysis. J Evid Based Med 2021, 14, 208–217. [CrossRef]
- Fu, L.; Li, Y.-N.; Luo, D.; Deng, S.; Hu, Y.-Q. Plausible Relationship between Homocysteine and Obesity Risk via MTHFR Gene: A Meta-Analysis of 38,317 Individuals Implementing Mendelian Randomization. Diabetes Metab Syndr Obes 2019, 12, 1201–1212. [CrossRef]
- Leal-Ugarte, E.; Peralta-Leal, V.; Meza-Espinoza, J.P.; Durán-González, J.; Macías-Gómez, N.; Bocanegra-Alonso, A.; Lara-Ramos, J.R. Association of the MTHFR 677C>T Polymorphism with Obesity and Biochemical Variables in a Young Population of Mexico. J Med Biochem 2019, 38, 461–467. [CrossRef]
- Pouwels, S.; Sakran, N.; Graham, Y.; Leal, A.; Pintar, T.; Yang, W.; Kassir, R.; Singhal, R.; Mahawar, K.; Ramnarain, D. Non-Alcoholic Fatty Liver Disease (NAFLD): A Review of Pathophysiology, Clinical Management and Effects of Weight Loss. BMC Endocr Disord 2022, 22, 63. [CrossRef]
- Nassir, F. NAFLD: Mechanisms, Treatments, and Biomarkers. Biomolecules 2022, 12, 824. [CrossRef]
- Yuan, S.; Chen, J.; Dan, L.; Xie, Y.; Sun, Y.; Li, X.; Larsson, S.C. Homocysteine, Folate, and Nonalcoholic Fatty Liver Disease: A Systematic Review with Meta-Analysis and Mendelian Randomization Investigation. Am J Clin Nutr 2022, 116, 1595–1609. [CrossRef]
- Dai, Y.; Zhu, J.; Meng, D.; Yu, C.; Li, Y. Association of Homocysteine Level with Biopsy-Proven Non-Alcoholic Fatty Liver Disease: A Meta-Analysis. J. Clin. Biochem. Nutr. 2016, 58, 76–83. [CrossRef]
- Sun, M.-Y.; Zhang, L.; Shi, S.-L.; Lin, J.-N. Associations between Methylenetetrahydrofolate Reductase (MTHFR) Polymorphisms and Non-Alcoholic Fatty Liver Disease (NAFLD) Risk: A Meta-Analysis. PLoS One 2016, 11, e0154337. [CrossRef]
- Hao, X.; Ma, C.; Xiang, T.; Ou, L.; Zeng, Q. Associations Among Methylene Tetrahydrofolate Reductase Rs1801133 C677T Gene Variant, Food Groups, and Non-Alcoholic Fatty Liver Disease Risk in the Chinese Population. Front Genet 2021, 12, 568398. [CrossRef]
- De Vincentis, A.; Mancina, R.M.; Pihlajamäki, J.; Männistö, V.; Petta, S.; Dongiovanni, P.; Fracanzani, A.L.; Valenti, L.; Tavaglione, F.; Romeo, S.; et al. Genetic Variants in the MTHFR Are Not Associated with Fatty Liver Disease. Liver Int 2020, 40, 1934–1940. [CrossRef]
- Wang, X.; Zhou, Y.; Zhang, M.; Wang, Y.; Qin, B. The Methylenetetrahydrofolate Reductase Genotype 677CT and Non-Alcoholic Fatty Liver Disease Have a Synergistic Effect on the Increasing Homocysteine Levels in Subjects from Chongqing, China. Genes Dis 2019, 6, 88–95. [CrossRef]
- Levine, A.; Koletzko, S.; Turner, D.; Escher, J.C.; Cucchiara, S.; de Ridder, L.; Kolho, K.-L.; Veres, G.; Russell, R.K.; Paerregaard, A.; et al. ESPGHAN Revised Porto Criteria for the Diagnosis of Inflammatory Bowel Disease in Children and Adolescents. J Pediatr Gastroenterol Nutr 2014, 58, 795–806. [CrossRef]
- Mak, W.Y.; Zhao, M.; Ng, S.C.; Burisch, J. The Epidemiology of Inflammatory Bowel Disease: East Meets West. J Gastroenterol Hepatol 2020, 35, 380–389. [CrossRef]
- Uhlig, H.H.; Charbit-Henrion, F.; Kotlarz, D.; Shouval, D.S.; Schwerd, T.; Strisciuglio, C.; de Ridder, L.; van Limbergen, J.; Macchi, M.; Snapper, S.B.; et al. Clinical Genomics for the Diagnosis of Monogenic Forms of Inflammatory Bowel Disease: A Position Paper From the Paediatric IBD Porto Group of European Society of Paediatric Gastroenterology, Hepatology and Nutrition. J Pediatr Gastroenterol Nutr 2021, 72, 456–473. [CrossRef]
- Peyrin-Biroulet, L.; Rodriguez-Guéant, R.-M.; Chamaillard, M.; Desreumaux, P.; Xia, B.; Bronowicki, J.-P.; Bigard, M.-A.; Guéant, J.-L. Vascular and Cellular Stress in Inflammatory Bowel Disease: Revisiting the Role of Homocysteine. Am J Gastroenterol 2007, 102, 1108–1115. [CrossRef]
- Varzari, A.; Deyneko, I.V.; Tudor, E.; Turcan, S. Polymorphisms of Glutathione S-Transferase and Methylenetetrahydrofolate Reductase Genes in Moldavian Patients with Ulcerative Colitis: Genotype-Phenotype Correlation. Meta Gene 2016, 7, 76–82. [CrossRef]
- Karban, A.; Feldman, T.; Waterman, M.; Leiba, R.; Efrati, E. The Association of the MTHFR C677T Polymorphism with Inflammatory Bowel Diseases in the Israeli Jewish Population: An Example of Genetic Heterogeneity. Medicine (Baltimore) 2016, 95, e5611. [CrossRef]
- Servy, E.J.; Jacquesson-Fournols, L.; Cohen, M.; Menezo, Y.J.R. MTHFR Isoform Carriers. 5-MTHF (5-Methyl Tetrahydrofolate) vs Folic Acid: A Key to Pregnancy Outcome: A Case Series. J Assist Reprod Genet 2018, 35, 1431–1435. [CrossRef]
- Erdoğan, K.; Sanlier, N.T.; Sanlier, N. Are Epigenetic Mechanisms and Nutrition Effective in Male and Female Infertility? J Nutr Sci 2023, 12, e103. [CrossRef]
- More, A.; Anjankar, N.; Shrivastava, J.; Nair, N.; Jadhav, R. Correlation of MTHFR C677T Polymorphism with Male Infertility among Indian Population: Case-Control Study. J Pharm Bioallied Sci 2024, 16, S2809–S2814. [CrossRef]
- Ménézo, Y.; Patrizio, P.; Alvarez, S.; Amar, E.; Brack, M.; Brami, C.; Chouteau, J.; Clement, A.; Clement, P.; Cohen, M.; et al. MTHFR (Methylenetetrahydrofolate Reductase: EC 1.5.1.20) SNPs (Single-Nucleotide Polymorphisms) and Homocysteine in Patients Referred for Investigation of Fertility. J Assist Reprod Genet 2021, 38, 2383–2389. [CrossRef]
- Liu, K.; Zhao, R.; Shen, M.; Ye, J.; Li, X.; Huang, Y.; Hua, L.; Wang, Z.; Li, J. Role of Genetic Mutations in Folate-Related Enzyme Genes on Male Infertility. Sci Rep 2015, 5, 15548. [CrossRef]
- Ren, F.; Fang, G.; Zhang, Z. Association between Methylenetetrahydrofolate Reductase C677T Polymorphisms and Male Oligozoospermia, Asthenozoospermia or Oligoasthenozoospermia: A Case–Control Study. Sci Rep 2024, 14, 25219. [CrossRef]
- Gong, M.; Dong, W.; He, T.; Shi, Z.; Huang, G.; Ren, R.; Huang, S.; Qiu, S.; Yuan, R. MTHFR 677C>T Polymorphism Increases the Male Infertility Risk: A Meta-Analysis Involving 26 Studies. PLoS One 2015, 10, e0121147. [CrossRef]
- Thakur, P.; Bhalerao, A. High Homocysteine Levels During Pregnancy and Its Association With Placenta-Mediated Complications: A Scoping Review. Cureus 2023, 15, e35244. [CrossRef]
- Ota, K.; Takahashi, T.; Han, A.; Damvaeba, S.; Mizunuma, H.; Kwak-Kim, J. Effects of MTHFR C677T Polymorphism on Vitamin D, Homocysteine and Natural Killer Cell Cytotoxicity in Women with Recurrent Pregnancy Losses. Hum Reprod 2020, 35, 1276–1287. [CrossRef]
- Basha, A.; Alkhatib, Y.; Tashtoush, T.; Yousef, M.; Oweidi, L.; Alkhatib, M.; Al-Aqrabawi, S.; Jarrar, Y.; Awidi, A. Recurrent Early Pregnancy Loss and Congenital Thrombophilia: A Prospective Study. J Clin Med 2024, 13, 6871. [CrossRef]
- Mehta, P.; Vishvkarma, R.; Singh, K.; Rajender, S. MTHFR 1298A>C Substitution Is a Strong Candidate for Analysis in Recurrent Pregnancy Loss: Evidence from 14,289 Subjects. Reprod Sci 2022, 29, 1039–1053. [CrossRef]
- Pritchard, A.M.; Hendrix, P.W.; Paidas, M.J. Hereditary Thrombophilia and Recurrent Pregnancy Loss. Clin Obstet Gynecol 2016, 59, 487–497. [CrossRef]
- Chen, H.; Yang, X.; Lu, M. Methylenetetrahydrofolate Reductase Gene Polymorphisms and Recurrent Pregnancy Loss in China: A Systematic Review and Meta-Analysis. Arch Gynecol Obstet 2016, 293, 283–290. [CrossRef]
- Memon, S.I.; Acharya, N.S. The Association Between Serum Homocysteine Levels and Placenta-Mediated Complications: A Narrative Review. Cureus 2022, 14, e31305. [CrossRef]
- Dymara-Konopka, W.; Laskowska, M. The Role of Nitric Oxide, ADMA, and Homocysteine in The Etiopathogenesis of Preeclampsia—Review. Int J Mol Sci 2019, 20, 2757. [CrossRef]
- Raijmakers, M.T.; Zusterzeel, P.L.; Steegers, E.A.; Peters, W.H. Hyperhomocysteinaemia: A Risk Factor for Preeclampsia? Eur J Obstet Gynecol Reprod Biol 2001, 95, 226–228. [CrossRef]
- Zhang, C.; Hu, J.; Wang, X.; Gu, H. High Level of Homocysteine Is Associated with Pre-Eclampsia Risk in Pregnant Woman: A Meta-Analysis. Gynecol Endocrinol 2022, 38, 705–712. [CrossRef]
- Mahmood, S.; Younas, H.; Younus, A.; Nathenial, S. A Narrative Review on the Role of Folate-Mediated One-Carbon Metabolism and Its Associated Gene Polymorphisms in Posing Risk to Preeclampsia. Clin Exp Hypertens 2021, 43, 487–504. [CrossRef]
- Qiu, X.; Gao, F.; Qiu, Y.; Bao, J.; Gu, X.; Long, Y.; Liu, F.; Cai, M.; Liu, H. Association of Maternal Serum Homocysteine Concentration Levels in Late Stage of Pregnancy with Preterm Births: A Nested Case-Control Study. J Matern Fetal Neonatal Med 2018, 31, 2673–2677. [CrossRef]
- Huang, L.-L.; Tong, J.-R.; Huang, Y.; Wei, Y.-N.; Chen, H.-F.; Chen, Y.; Su, J.-Y.; Deng, L. Association of MTHFR Gene C677T Polymorphism with Pregnancy Outcome. Eur Rev Med Pharmacol Sci 2023, 27, 166–171. [CrossRef]
- Wang, B.J.; Liu, M.J.; Wang, Y.; Dai, J.R.; Tao, J.Y.; Wang, S.N.; Zhong, N.; Chen, Y. Association between SNPs in Genes Involved in Folate Metabolism and Preterm Birth Risk. Genet Mol Res 2015, 14, 850–859. [CrossRef]
- Dević Pavlić, S.; Šverko, R.; Barišić, A.; Mladenić, T.; Vraneković, J.; Stanković, A.; Peterlin, A.; Peterlin, B.; Ostojić, S.; Pereza, N. MTHFR Gene Polymorphisms and DNA Methylation in Idiopathic Spontaneous Preterm Birth. Medicina (Kaunas) 2024, 60, 2028. [CrossRef]
- Wu, H.; Zhu, P.; Geng, X.; Liu, Z.; Cui, L.; Gao, Z.; Jiang, B.; Yang, L. Genetic Polymorphism of MTHFR C677T with Preterm Birth and Low Birth Weight Susceptibility: A Meta-Analysis. Arch Gynecol Obstet 2017, 295, 1105–1118. [CrossRef]
- Tiwari, D.; Bose, P.D.; Das, S.; Das, C.R.; Datta, R.; Bose, S. MTHFR (C677T) Polymorphism and PR (PROGINS) Mutation as Genetic Factors for Preterm Delivery, Fetal Death and Low Birth Weight: A Northeast Indian Population Based Study. Meta Gene 2015, 3, 31–42. [CrossRef]
- Steele, J.W.; Kim, S.-E.; Finnell, R.H. One Carbon Metabolism and Folate Transporter Genes: Do They Factor Prominently in the Genetic Etiology of Neural Tube Defects? Biochimie 2020, 173, 27–32. [CrossRef]
- Wang, X.; Yu, J.; Wang, J. Neural Tube Defects and Folate Deficiency: Is DNA Repair Defective? Int J Mol Sci 2023, 24, 2220. [CrossRef]
- Kakebeen, A.D.; Niswander, L. Micronutrient Imbalance and Common Phenotypes in Neural Tube Defects. Genesis 2021, 59, e23455. [CrossRef]
- Almekkawi, A.K.; AlJardali, M.W.; Daadaa, H.M.; Lane, A.L.; Worner, A.R.; Karim, M.A.; Scheck, A.C.; Frye, R.E. Folate Pathway Gene Single Nucleotide Polymorphisms and Neural Tube Defects: A Systematic Review and Meta-Analysis. J Pers Med 2022, 12, 1609. [CrossRef]
- Li, J.; Feng, D.; He, S.; Yang, H.; Su, Z.; Ye, H. Association of MTHFR 677C > T Gene Polymorphism with Neonatal Defects: A Meta-Analysis of 81444 Subjects. J Obstet Gynaecol 2022, 42, 1811–1822. [CrossRef]
- Soleimani-Jadidi, S.; Meibodi, B.; Javaheri, A.; Tabatabaei, R.S.; Hadadan, A.; Zanbagh, L.; Abbasi, H.; Bahrami, R.; Mirjalili, S.R.; Karimi-Zarchi, M.; et al. Association between Fetal MTHFR A1298C (Rs1801131) Polymorphism and Neural Tube Defects Risk: A Systematic Review and Meta-Analysis. Fetal Pediatr Pathol 2022, 41, 116–133. [CrossRef]
- Menezo, Y.; Elder, K.; Clement, A.; Clement, P. Folic Acid, Folinic Acid, 5 Methyl TetraHydroFolate Supplementation for Mutations That Affect Epigenesis through the Folate and One-Carbon Cycles. Biomolecules 2022, 12, 197. [CrossRef]
- Ferrazzi, E.; Tiso, G.; Di Martino, D. Folic Acid versus 5- Methyl Tetrahydrofolate Supplementation in Pregnancy. Eur J Obstet Gynecol Reprod Biol 2020, 253, 312–319. [CrossRef]
- Samaniego-Vaesken, M. de L.; Morais-Moreno, C.; Carretero-Krug, A.; Puga, A.M.; Montero-Bravo, A.M.; Partearroyo, T.; Gregorio, V.-M. Supplementation with Folic Acid or 5-Methyltetrahydrofolate and Prevention of Neural Tube Defects: An Evidence-Based Narrative Review. Nutrients 2024, 16, 3154. [CrossRef]
- Radu, A.-F.; Bungau, S.G. Management of Rheumatoid Arthritis: An Overview. Cells 2021, 10, 2857. [CrossRef]
- Yadav, U.; Kumar, P.; Gupta, S.; Rai, V. Distribution of MTHFR C677T Gene Polymorphism in Healthy North Indian Population and an Updated Meta-Analysis. Ind J Clin Biochem 2017, 32, 399–410. [CrossRef]
- Rosenberg, N.; Murata, M.; Ikeda, Y.; Opare-Sem, O.; Zivelin, A.; Geffen, E.; Seligsohn, U. The Frequent 5,10-Methylenetetrahydrofolate Reductase C677T Polymorphism Is Associated with a Common Haplotype in Whites, Japanese, and Africans. The American Journal of Human Genetics 2002, 70, 758–762. [CrossRef]
- Cen, H.; Huang, H.; Zhang, L.-N.; Liu, L.-Y.; Zhou, L.; Xin, X.-F.; Zhuo, R.-J. Associations of Methylenetetrahydrofolate Reductase (MTHFR) C677T and A1298C Polymorphisms with Genetic Susceptibility to Rheumatoid Arthritis: A Meta-Analysis. Clin Rheumatol 2017, 36, 287–297. [CrossRef]
- Bagheri-Hosseinabadi, Z.; Imani, D.; Yousefi, H.; Abbasifard, M. MTHFR Gene Polymorphisms and Susceptibility to Rheumatoid Arthritis: A Meta-Analysis Based on 16 Studies. Clin Rheumatol 2020, 39, 2267–2279. [CrossRef]
- Yuan, Y.; Shao, W.; Li, Y. Associations between C677T and A1298C Polymorphisms of MTHFR and Susceptibility to Rheumatoid Arthritis: A Systematic Review and Meta-Analysis. Rheumatol Int 2017, 37, 557–569. [CrossRef]
Figure 1.
Graphic representation of the reactions of the folate cycle with information specified in the introduction, the destination of the indicated products is shown.
Figure 1.
Graphic representation of the reactions of the folate cycle with information specified in the introduction, the destination of the indicated products is shown.
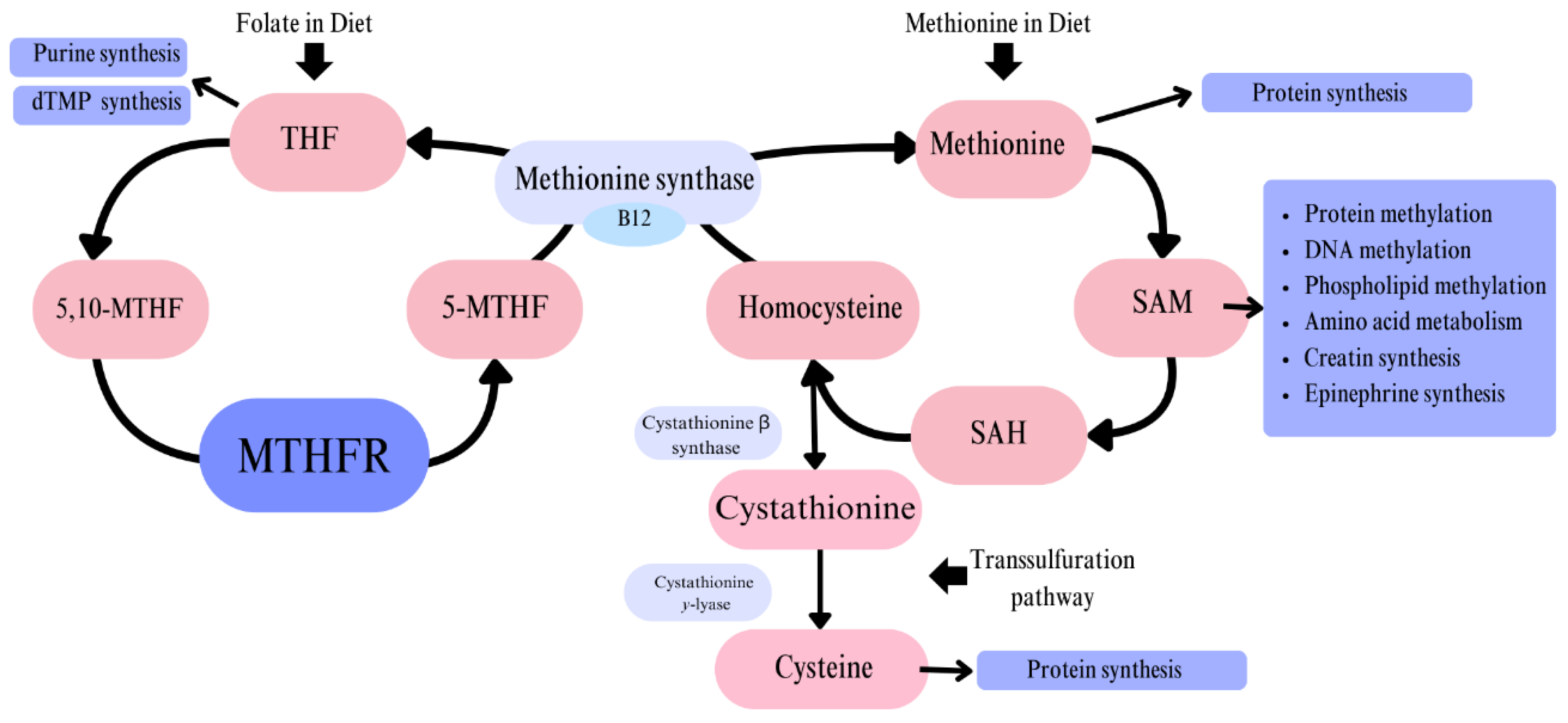
Figure 2.
Illustrating the effects of hyperhomocysteinemia/elevated homocysteine levels on the cardiovascular system. The figure depicts the effects of homocysteine on various components and parameters of the cardiovascular system, highlighting the endothelium based on the information provided in the introduction. [13,14].
Figure 2.
Illustrating the effects of hyperhomocysteinemia/elevated homocysteine levels on the cardiovascular system. The figure depicts the effects of homocysteine on various components and parameters of the cardiovascular system, highlighting the endothelium based on the information provided in the introduction. [13,14].
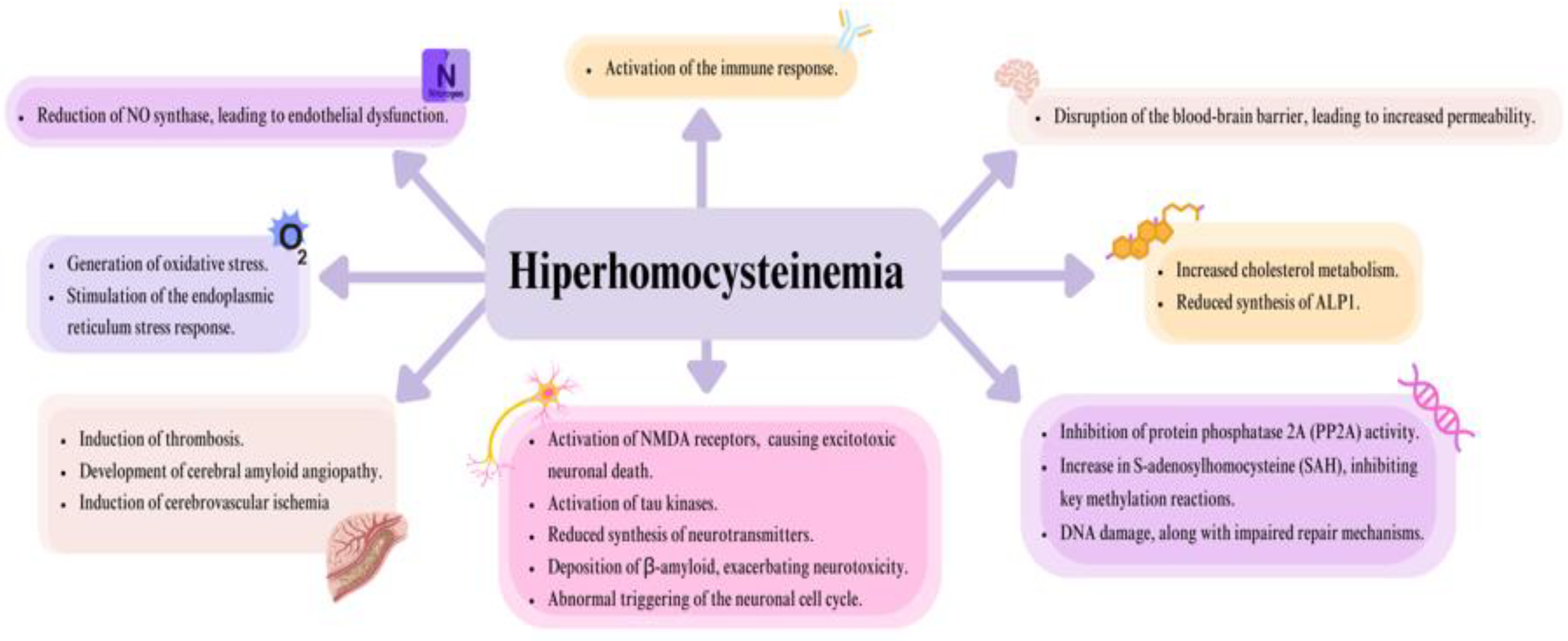
Figure 3.
The figure shows the discussed pathomechanism of epithelial damage and contribution to atherosclerotic plaque formation in hyperhomocysteinemia.
Figure 3.
The figure shows the discussed pathomechanism of epithelial damage and contribution to atherosclerotic plaque formation in hyperhomocysteinemia.
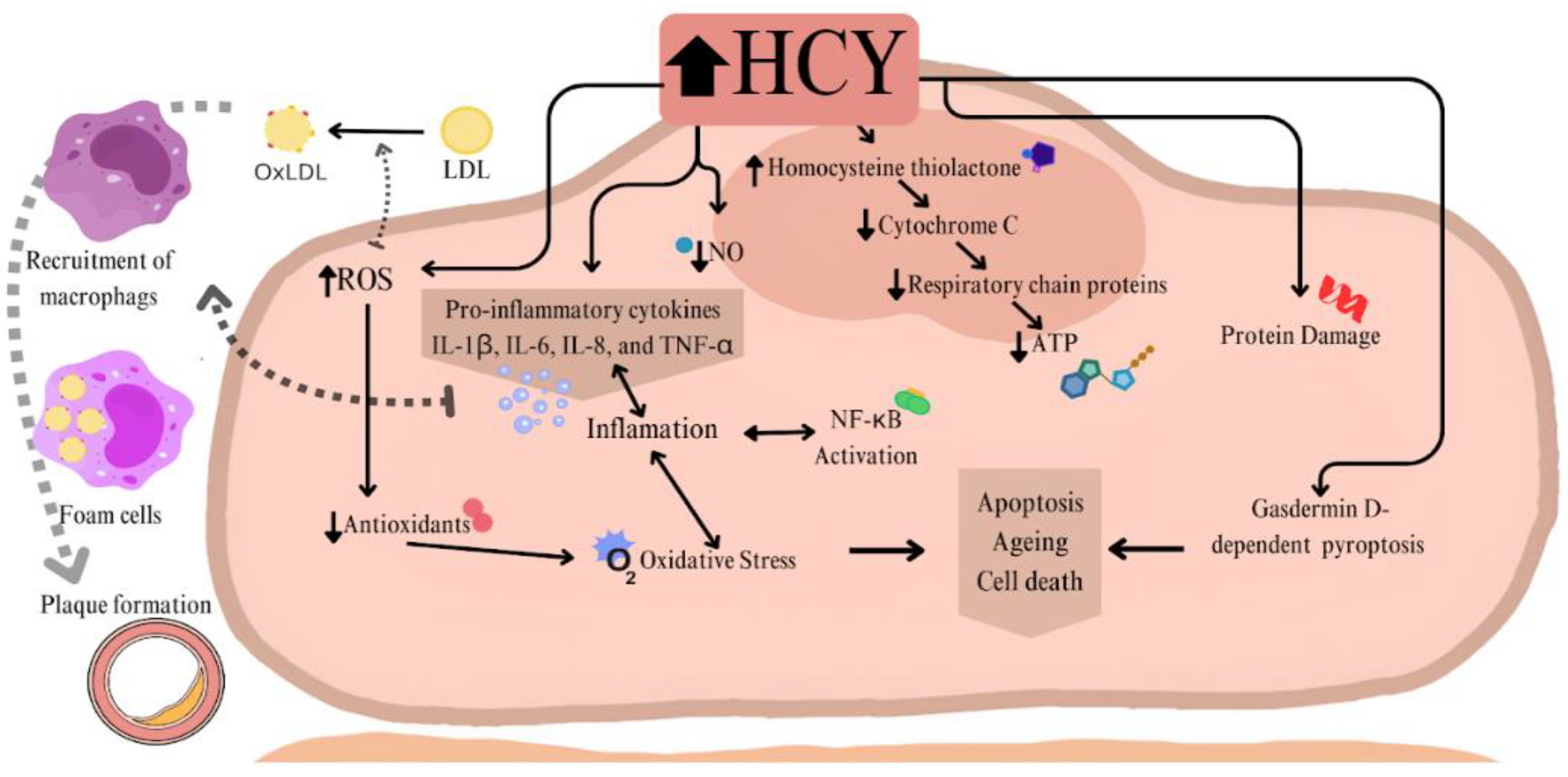
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



