Submitted:
15 March 2025
Posted:
17 March 2025
You are already at the latest version
Abstract
The canonical model of vertebrate sex chromosome evolution predicts one way of trend toward degradation. However, most sex chromosomes in lower vertebrates are homomorphic. Recent progress in studies of sex determination resulted in the discovery of more than 30 master sex determination (MSD) genes, most of which were from teleost fish. Analysis of MSD gene acquisition, recombination suppression, and sex chromosome-specific sequences revealed correlation of the modes of MSD gene acquisition and evolution of sex chromosomes: Sex chromosomes remain homomorphic with MSD genes acquired by simple mutations, gene duplications, allelic variations or neofunctionalization; in contrast, they become heteromorphic with MSD genes acquired by chromosomal inversions, fusions and fissions. There is no recombination suppression with sex chromosomes carrying MSD genes gained through simple mutations. In contrast, there is extensive recombination suppression with sex chromosomes carrying MSD genes gained through chromosome inversion. There is limited recombination suppression with sex chromosomes carrying MSD genes gained through transpositions or translocations. We proposed the cause-effect model that predicts sex chromosomes evolution being consequential of the acquisition modes of MSD genes, which explains evolution of sex chromosomes in various vertebrates. A key factor determining the trend of sex chromosome evolution is if non-homologous regions are created during the acquisition of MSD genes. Chromosome inversion creates inversely homologous but directly non-homologous sequences which lead to recombination suppression but remain recombination potential. Over time, recurrent recombination in the inverted regions causes degradation of sex chromosomes. Depending on the nature of deletions in the inverted regions, sex chromosomes may evolve with dosage compensation or mechanisms to retain haploinsufficient genes.
Keywords:
Sex determination
; Aquaculture
; Sex differentiation
; fish
; recombination suppression
; chromosome inversion
1. Introduction
The mechanism of sex determination in vertebrates is enormously diverse, especially with teleost fish [1,2], ranging from unisexuals, hermaphroditism, environmental sex determination, to genetic sex-determination [3]. With genetic sex determination, 30 distinct MSD genes have been identified from vertebrates under various sex determination (SD) systems, XY, ZW, and multiple sex chromosomes [1,4]. Despite such diversities, the molecular pathways and downstream players are generally conserved (Table 1). The basic operation of the sex determination network is through the dynamics of the opposing male and female pathways to ensure 1:1 male and female ratio [5], thus demanding the SD gene to be expressed in a temporal, spatial, dose- and temperature-sensitive fashion. The gene products in the male pathway and female pathway are antagonistic, and their expression is negatively regulated against each other (Figure 1).
As with the MSD genes, sex chromosomes in vertebrates have evolved with various pace to various degrees, ranging from entirely homomorphic in many species of lower vertebrates to highly degraded heteromorphic in higher vertebrates. The canonical model of sex chromosome evolution predicts one way of trend toward degeneration and degradation of the sex chromosome carrying the MSD gene [6,7,8,9]. However, empirical evidence in support of this theory does not go beyond mammals and birds. The sex chromosomes in reptiles, amphibians, and teleost fish can be homomorphic or heteromorphic, and most of them are homomorphic. A popular explanation is that sex chromosomes in lower vertebrates are young, but both homomorphic and heteromorphic sex chromosomes are present across all lower vertebrates ranging from Cartilaginous fish, teleost fish, amphibians, and reptiles; some homomorphic sex chromosomes are older than some highly differentiated heteromorphic sex chromosomes, directly challenging the canonical theory of sex chromosome evolution. The availability of genome sequences and known MSD genes from various vertebrate species, especially from lower vertebrates, made it possible to determine: 1) How MSD genes were acquired; 2) How sex chromosomes evolve in relation to the modes of MSD gene acquisition; 3) How MSD genes affect the evolution of sex determination systems. Here we present the cause-effect model that states the mode of MSD acquisition determines the evolution of sex chromosomes. This model explains for evolution of vertebrate sex chromosomes in relation to the MSD genes and their chromosome karyotypes.
Only one MSD gene and one SD system has evolved from homeothermic mammals and birds, respectively, but multiple MSD genes and SD systems have evolved from poikilothermic vertebrates (Figure 1). In mammals, Sry is both necessary and sufficient for sex determination [10]. In birds, dmrt1 works in a dose-sensitive fashion, where two copies of dmrt1 make a male, while one copy makes a female [11]. Apparently, the two-fold difference in dmrt1 expression is sufficient to reach the threshold for the sex phenotype in birds. With reptiles, ZW system was believed to be the only genetic SD system for over 50 years until the recent demonstration of the XY SD system in boa and python snakes [12]. Similarly, with amphibians, DM-W in African clawed frog, Xenopus laevis, was the only MSD gene known from 8,470 amphibian species until the recent discovery of MSD gene bot1l in the European green toad, Bufo viridis, with an XY system [13].
As poikilotherms living in the aquatic environments, teleost fish must delicately respond to broad temperature variations in the SD process. In correlation, 28 distinct MSD genes have evolved in teleost fish (Figure 1), including transcriptional factors, TGF-β cytokines, steroidogenesis genes, and many “newcomers”, all are downstream players in the SD pathways as compared to the MSD gene Sry in mammals. While Sry functions as a decisive MSD gene, most other MSD genes are quantitative in response to dose and temperature sensitivity.
2. Molecular Mechanisms for the Acquisition of MSD Genes
The first step of sex chromosome evolution is the emergence of MSD genes. To serve as a sex chromosome, one gene on one of the autosome pairs must initially gain new functions or new expression patterns in favor of male or female pathway, thereby becoming a master switch turning on the genes for either the male or female development. As shown in Figure 2, different mechanisms of MSD gene acquisition determine the pathway of sex chromosome evolution. In the context of sex chromosome evolution, we classify the mechanism of MSD gene acquisition into four categories: 1) Simple mutations, such as allelic variations (including base substitutions, neofunctionalization and subfunctionalization), gene duplications, and small deletions or insertions [14]. This category accounts for the largest numbers of known MSD genes in vertebrates (Table 2). Sex chromosomes in this category is characterized by not harboring any non-homologous sequences and, therefore, they stay homomorphic. Examples of this category include missense mutations in the coding regions, mutations in the regulatory sequences such as promoters, enhancers, silencers, and splicing junctions, as well as gene duplications; 2) Translocations or transpositions of DNA carrying the MSD gene, or small-sized inversions. Sex chromosomes of this category carry a limited region of non-homologous sequences that are not recombining, but they stay homomorphic. Examples of this category include MSD gene DM-W in African clawed frog, sdY in salmonids, and amhr2 among silurid catfishes; 3) Large chromosomal inversions. An MSD gene is activated due to juxtaposition of the MSD gene to a new regulatory context that lead to changes in expression profiles, and or temporal or spatial regulation in favor of male or female pathway. Sex chromosomes of this category carry large non-homologous regions that are inverted between the sex chromosomes, which reduce crossovers, but retain the recombination potential upon formation of the recombination loop [15]. Reduced recombination leads to sequence degeneration, but any recombination and recurrent recombination in the inverted region would lead to duplication in one and deletion in the other chromosome [16,17], and most likely requires double crossovers to survive. As a result, sex chromosomes of this category become heteromorphic; 4) Chromosome fusions or fissions. Chromosome fusions or fissions lead to non-homologous regions of large sizes, and often the multi-chromosome SD system.
3. The Diversity of Master Sex Determination Genes in Vertebrates
Since the discovery of SRY gene in humans [10], it took 12 years to find the second vertebrate MSD gene, dmY, from medaka [18]. However, recent advances in sequencing technologies drastically accelerated the pace. Now 30 distinct MSD genes have been identified from vertebrates (Table 2), with the vast majority of these being identified from teleost fish. Only two MSD genes have been identified from amphibians, DM-W from African clawed frog and bod1l from the European green toad [13]. Although both XY and ZW SD systems have been found in reptiles, no MSD genes have yet been identified.
In teleost fish, 28 distinct MSD genes have been identified, including transcriptional factors, TGF-β cytokines, genes involved in steroidogenesis, and many “newcomers” (Table 2). Of the transcriptional factors, dmrt1 is the most popular although sox family of transcription factors including sox2, sox3, and sox7 also serve as MSD genes. In addition, transcriptional factors FIGLA-like and ptf1a were identified to be the MSD genes. Of these, dmrt1 and sox genes were well known as the “usual suspects” [19], but FIGLA-like and ptf1a are newcomers. FIGLA-like gene is the MSD gene for Nile tilapia [20]; it encodes a protein of 99 amino acids including a 45-amino-acid basic helix-loop-helix domain and specifically expressed in testis. While the autosomal FIGLA gene is a femaleness gene promoting ovary formation, FIGLA-like gene on the Y interferes with the functions of the autosomal FIGLA gene, leading to testis development [20]. Similarly, a truncated form of Ptf1a (pancreas transcriptional factor 1 alpha), named ptf1aY, was identified as MSD gene in Chinese longsnout catfish [21]. Five TGF-β genes including amh, amhr2, gdf6, gsdf, and bmpr1b have been identified as MSD genes in teleost fish. In approximately a dozen species, genes involved in steroidogenesis have been identified as MSD genes, including hsd17b1, cyp19a1, hsdl1, sult1st6y, and fshrY. Of these, the functions of hsd17b1 and cyp19a1 are well known: 17β-Hydroxysteroid dehydrogenase 1 oxidizes or reduces the C17 hydroxy/keto group of androgens and estrogens and, hence, regulates the potency of these sex steroids, while cyp19a1 is a temperature-sensitive aromatase that convert androgens into estrogens. Therefore, these are generally regarded as femaleness genes, and they serve as MSD genes mostly in ZW systems (Table 2). In addition to these three groups, over a dozen of newcomers have been identified as MSD genes (Table 2). These include sdY, cephx1Y, paics, banf2, RIN3, zkY, pfpdz1, hydin, and gipc1 (a pdz domain-containing gene). The pathways for sex determination of these genes are unknown, with exception of sdY that is a truncated form of interferon regulatory factor 9; sdY functions as sex determination gene in salmonids as a dominant negative regulator through its interaction with FOXL2 [22].
4. Convergent and Divergent Evolution of MSD Genes
Despite convergent evolution of MSD genes in mammals and birds, most known MSD genes from lower vertebrates evolve independently. The identification of 30 distinct MSD genes indicated unlimited options of MSD genes, and the options go far beyond the usual suspects, with a trend of moving from upstream “master” to downstream players. Limited information is available from amphibians and reptiles, but MSD genes evolve mostly independently in lower vertebrates, with some local convergence. As shown in Figure 3, many orders involve more than one MSD genes, reflecting divergent evolution of MSD genes. Even if a specific MSD gene is found in multiple orders, the molecular mechanisms of their acquisition were different (Table 2). Of all teleost orders, Cichliformes has the largest known number of seven MSD genes, involving transcription factors (banf2 and FIGLA), TGF-β members (amh, amhr2, gsdf), and the newcomers category of unknown pathways (paics and RIN3), followed by Beloniformes and Pleuronectiformes, each with six known MSD genes, and then by Perciformes and Siluriformes, each with five known MSD genes. Convergent evolution, mostly through shared ancestry [23], does exist locally, mostly within the orders. The most dramatic is the conservation of sdY as the MSD gene across the entire salmonids [24,25]. The common GTF-β factors could be shared by several genera, or families, but none goes beyond the scope of the order.
5. The Canonical Model of Sex Chromosome Evolution
The canonical model of sex-chromosome evolution includes four consecutive phases: 1) First, a MSD gene acquires sex-determining function by mutation from a pair of autosomes; 2) Recombination suppression between the sex chromosomes; 3) Degeneration of the sex chromosome with accumulation of deleterious mutations and TE in non-recombinational regions of the sex chromosomes (Y or W); 4) Deletions of the degenerated sex chromosome lead to the decay of the sex chromosome, and evolution of dosage compensation [6,7,8,9]. The diversity and evolutionary lability of sex chromosomes across the tree of life [26] have challenged the canonical model: The vast majority of known sex chromosomes in lower vertebrates are homomorphic; recombination suppression is involved only in a small fraction of species with known sex chromosomes; recombination suppression does not always cause degradation of sex chromosomes; sex chromosome degeneration and degradation have been demonstrated only in a small fraction of cases in lower vertebrates [27]; sex antagonistic genes have not been widely identified, and even in a few cases of examples, linkage of genes and their related phenotypes cannot automatically interpreted as causations [8,28]; the remarkable turnover of sex chromosomes in many systems, especially in teleost fish, does not support inevitable linearity of sex chromosome evolution [28,29,30]; and dosage compensation was observed in some but not in all species with heteromorphic sex chromosomes. Instead, in some teleost fish species, “selective retention of haploinsufficient genes is apparently an alternative strategy for coping with the imbalanced expression of X- or Z-linked genes [31,32]. All these situations differ from those of human [33] and birds [34] and thus inspires for the proposal of new theories of sex chromosome evolution.
6. The Proposed Cause-Effect Model for the Evolution of Sex Chromosomes
Based on analysis of mechanisms of MSD gene acquisition, we propose a cause-effect model of sex chromosome evolution, which is depicted in Figure 2: How sex chromosomes evolve depends on how the MSD genes were acquired. Sex chromosomes carrying MSD genes acquired from simple mutations such as allelic variations, neofunctionalization, and gene duplications stay homomorphic, without recombination suppression nor degeneration; Sex chromosomes carrying MSD genes acquired from translocations and transpositions carry a small non-homologous region between the sex chromosomes, and therefore, they develop recombination suppression within the transposed segments, but they remain homomorphic; Sex chromosomes carrying MSD genes acquired from chromosomal inversions carry inverted regions between the sex chromosomes. Recombination is reduced in the inverted region, but single crossover in the inverted region causes deletions or duplications, which may be lethal to the gametes, and thus requires double crossovers to survive (Figure 4). Over time, such sex chromosomes degenerate and decay, especially with large inversions. A key factor for the evolution of sex chromosomes is if non-homologous sequences are being created during acquisition of MSD genes. Sex chromosomes with MSD genes acquired through simple mutations do not create non-homologous regions. In contrast, sex chromosomes with MSD genes acquired through transpositions, inversions, fusions and fissions all create non-homologous regions between the sex chromosome pairs, leading to recombination suppression. Key differences of this model from the canonical sex conflict model include: 1) Sex chromosome evolution is to resolve the structural problems created during acquisition of MSD genes, such as the presence of inverted regions on the sex chromosome pairs; 2) Recombination suppression is resultant of inter-chromosomal non-homologous regions between the sex chromosome pairs, not of intrachromosomal linkage of antagonistic alleles; 3) In spite of the reduced recombination due to inversions, recurrent recombination in inverted regions over time leads to deletions on the sex chromosomes. Sequence degeneration and accumulation of transposable elements over time result in evolutionary strata.
6.1. Homomorphic Sex Chromosomes Without Recombination Suppression
The cause-effect model of sex chromosome evolution predicts that sex chromosomes harboring MSD genes acquired from simple mutations are homomorphic, with no recombination suppression or sequence degeneration (Figure 2). These sex chromosomes became sex chromosomes because of a gene they carry had become the MSD gene. Allelic and/or neofunctionalization/subfunctionalization accounted for the largest number of teleost fish species with known MSD genes to date. Teleost fish went through a third round of whole genome duplication [35], and neofunctionalization and subfunctionalization of ohnolog genes is a part of rediploidization processes [14,36]. Similarly, in teleost species with known MSD genes, tandem gene duplications were involved in many species (Table 2). Tandem duplications occur frequently in teleost fish species, especially with cytokines and chemokines [37].
One may argue that these sex chromosomes have not evolved much simply because they are young. However, sex chromosomes included in this category included taxa from cartilaginous fish, teleost fish, amphibians, and reptiles. Among teleost fish, a broad spectrum of teleost orders is involved, ranging from the base of teleost order such as Clupeiformes to the most advanced order of Tetraodontiformes, covering approximately 300 million years of evolutionary time. It is difficult to imagine that all these sex chromosomes are at the very beginning of their evolution. In contrast, in various medaka, both homomorphic and heteromorphic chromosomes were found, where allelic variations were found with homomorphic sex chromosomes (e.g., in O. latipes), but chromosomal inversion was found with heteromorphic sex chromosomes (e.g., O. javanicus), even though their evolutionary time is similar [38].
6.2. Homomorphic Sex Chromosomes with Limited Region of Recombination Suppression
The cause-effect model of sex chromosome evolution predicts that sex chromosomes harboring MSD genes acquired with insertions, translocations or transpositions may develop a sex chromosome-specific region (often referred to as MSY, for male-specific Y under an XY sex system) related to the insertion. In the inserted segments, recombination suppression may be present; sequence degeneration may occur, but the region of recombination suppression or the degenerated sequences may be limited to the insertional segments or slightly larger, while the bordering homologous sequences should have continued homologous recombination. As a result, the sex chromosome-specific region may carry fixed haplotypes, but the sex chromosomes stay homomorphic. As listed in Table 2 and illustrated in Figure 2, good examples of this category are sex chromosomes in species of Silurus, Pangasianodon, and Pangasius genera in the order of Siluriformes. A common insertion was found to have occurred at the Pangasidae base, which carried the MSD gene amhr2 [39,40,41,42]. After acquiring the MSD gene, the sex chromosomes evolve to an extent that sequence degeneration and recombination suppression were observed within the insertions but was limited to the sex chromosome-specific insertion. It should be noted that here the MSD gene is ancient, estimated to have emerged over 100 million years ago [43], directly against the interpretation that limited evolution of the sex chromosomes is because the evolution time has been short. Similarly, sdY gene in all salmonids has evolved approximately 60 million years, but sex chromosomes continue to be homomorphic [25].
6.3. Homomorphic Sex Chromosomes with Extended Recombination Suppression
The cause-effect model of sex chromosome evolution predicts that sex chromosomes involving large chromosomal inversions develop recombination suppression in the region of inversion. If the sex chromosome is young, they are observed as homomorphic sex chromosomes. In several cases in teleost fish, large SDRs were observed (Table 2). Interestingly, levels of sequence degeneration within the SDRs are low in such cases. It is likely that such sex chromosomes are still young; it is also possible that such sex chromosomes stay homomorphic because of other mechanisms of regulation that slow down the evolution of sex chromosomes (see below).
6.4. Heteromorphic Sex Chromosomes
In contrast to the situations of homomorphic sex chromosomes, heteromorphic sex chromosomes evolved from major structural changes of the sex chromosomes. Chromosome inversion is probably the most frequent cause for the evolution of heteromorphic sex chromosomes. With chromosomal inversion, a promoter or enhancer may be directly juxtaposed to a gene close to the inversional junction. If such a gene is a maleness or femaleness gene, the chromosomal inversion may have activated a gene as an MSD gene. In addition to direct juxtaposition of regulatory elements to bordering genes near the inversion junction, changes in chromatin structure and architecture may also have the capacity for activation of inactivation of genes within or near the inversion segment. While genomic and epigenomic regulation of the multi-dimensional chromatin architecture is not well understood, it is increasingly recognized that such architecture and spatial regulation is important for sex chromosome evolution [44]. If inversion is involved in the acquisition of the MSD gene, the “new” chromosome carrying the inversion initially still has the DNA contents, but the major change is the creation of non-homologous sequences between the pair of sex chromosomes in the inverted region. As demonstrated in ninespine stickleback, inversions may have a role in both the evolution of sex determination systems and the differentiation of sex chromosomes [45].
Accurate chromosome segregation during meiosis relies on homology between the maternal and paternal chromosomes. Yet, by definition, heterogametic sex involving heteromorphic sex chromosomes lacks a homologous partner [46]. The presence of chromosomal inversions reduces recombination; however, recombination does occur in inverted regions, which causes unequal crossovers, leading to segmental duplications or deletions in gametes. Once the recombination is suppressed, over time, sequences degenerate, TEs accumulate, and if no regulation, deleterious mutations accumulate and eventually, upon any recombination or rearrangements, deletions occur, leading to decay of sex chromosomes. The sex chromosomes eventually become stable when large, inverted regions are eliminated or are repositioned within heterochromatin regions such as being close to the centromere where there is no recombination.
While such processes have been well documented in the canonical model of sex chromosome evolution, as with those in mammals and birds, the best studied teleost fish under this category is perhaps the sex chromosomes of threespine stickleback (Gasterosteus aculeatus), with a large SDR of 17.5 Mb [32]. The MSD gene amhY is located in the oldest region of the stickleback Y chromosome close to the original inversion junction (stratum one), adjacent to the pseudoautosomal region. The three evolutionary strata suggested additional inversions and rearrangements on the Y chromosome, and such inversions were confirmed by genetic mapping [47]. The Y chromosome is less than 26 million years old, its sequence is degenerated, and it lost the majority of the genes that are present on chromosome X, retaining just 44.1% of the genes [32]. However, gene loss may not be random as many haploinsufficient genes were retained on the Y chromosome [32].
6.5. Multiple and Unequal Sex Chromosome SD Systems
In teleost fish, multiple sex chromosome SD system has been adopted in a sizable number of species. The actual number must be larger, but 75 multiple sex chromosome systems with 60 estimated independent origins have been documented to date [48]. Multiple sex chromosome systems can be viewed as special cases of heteromorphic sex chromosomes. In this context, the cause-effect model of sex chromosome evolution predicts that sex chromosomes derived from chromosome fusion or fission would create heteromorphic sex chromosomes; the extent of recombination suppression depends on the size of fusion or fission segments, as well as on size of additional inversion. Here we present one example to show the cause-effect model also fits the multiple sex chromosome systems. In the spotted Knifejaw (Oplegnathus punctatus), the X1X2X1X2/X1X2Y multiple chromosome SD system is operating. Genome sequencing revealed large genomic regions of recombination suppression, 29.3 Mb of X1 (from 0 to 29.30 Mb) and 17.58 Mb of X2 (from the centromere to 17.58 Mb) were associated with sex, of which a large inversion on X1, and the centromere on X2, as well as the SD locus, accounted for the observed recombination suppression. Sequence degeneration and gene loss were also observed [49].
7. Dosage Compensation and Epigenetic Regulation of Sex Chromosome Evolution
Dosage compensation equalizes the level of expression of X- or W-linked genes between the sexes. While dosage compensation is well understood for mammals [50,51], and somewhat understood for avian species [52,53], it is not well studied with lower vertebrates. Studies with tongue sole and threespine stickleback indicated the lack of dosage compensation, but selective retention of dosage sensitive genes [31,54]. In contrast, complete dosage compensation was observed with guppy species, Poecilia parae and P. picta [55,56]. This appears to be conflicting but may represent alternative strategies of coping with imbalanced expression of sex chromosomes in species with heteromorphic sex chromosomes. In cases where large deletions had occurred from the Y chromosome such as the guppies, complete dosage compensation was needed, and it was so observed. This may be an effective way to balance expression of the lost genes, especially those that are dosage sensitive. Alternatively, with selective retention of dosage sensitive genes, there is no need to develop dosage compensation, as observed with less degenerated sex chromosomes in threespine stickleback and tongue sole [28,29].
Epigenetic regulation is involved in growth, reproduction, disease resistance and stress responses of various fish species [57]. Several isolated studies suggested epigenetic regulation of sex determination and sex chromosome evolution. In channel catfish, an epigenetically marked locus of 8.9 Mb was well aligned with the SDR where recombination is completely suppressed [58,59]. The X chromosome was hypermethylated, leading to silencing of the X-borne hydin gene. In contrast, the Y chromosome was hypomethylated, and the Y-borne hydin gene is expressed, serving as MSD gene [60,65]. Similarly, in threespine stickleback, the sex chromosomes had the majority (65%) of differentially methylated CpG sites (DMS) of the genome, with hypermethylation in females and hypomethylated in males. Most interestingly, the DMS were predominantly located in the SDR, especially in strata 2 and 1, where recombination is suppressed [61]. Similar work was also conducted in two guppies, where differential methylation was observed in testis, with hypermethylation in females and hypomethylation in males. Again, the DMS were found mostly in the SDR, particularly in stratum 2 and stratum 1 where MSD gene is located [62]. These examples demonstrated differential methylation of the sex chromosomes, especially within the SDR. In addition to regulation of gene expression, one possibility is that hypermethylation in the SDR could block recombination, thereby protecting the inverted region from being deleted, possibly allow selective retention of dosage sensitive genes. Inhibition of recombination during meiosis by hypermethylation is well documented in plants [63,64]. If DNA methylation plays a similar role within the SDR, especially with chromosomal inversion, hypermethylation in the SDR would de facto slow down degeneration of sex chromosomes, and perhaps such a mechanism could be used to selectively retain haploinsufficient genes.
8. Conclusions and Perspectives
Recent advances in sequencing technologies allowed rapid discoveries of the enormously diverse MSD genes in vertebrates and the mechanisms of their acquisition. It is apparent that the trajectories of sex chromosomes evolution are resultant of the acquisition modes of MSD genes: sex chromosomes with MSD genes acquired through simple mutations do not involve recombination suppression, nor degeneration; in contrast, sex chromosomes with MSD genes acquired through chromosomal inversions involve recombination suppression that leads to accumulation of transposable elements and sequence degeneration; and when the inversions are large in size, they may lead to the decay of the sex chromosomes. A key factor determining the evolution of sex chromosomes is if inverted regions are being created between the sex chromosome pairs for the acquisition of MSD genes. The creation of long inverted regions between the sex chromosome pairs provided driving force for sex chromosome evolution [17]. Depending on the sizes of the inversions and the nature of the deletions during sex chromosome evolution, dosage compensation evolves when essential or haploinsufficient genes are deleted. In contrast, regulatory mechanisms may have evolved from lower vertebrates with selective retention of haploinsufficient genes [32].
A correct model of sex chromosome evolution is important for future work. For example, large non-recombining SDR likely is predictive of a large inversion. Although allelic variations are the predominant way through which MSD genes were acquired in lower vertebrates, chromosomal inversions are a common way for the acquisition of MSD genes among vertebrates. However, identification of inversion junctions from tens of millions of base pairs are still difficult. Future focus should be given to structural analysis around the borders of SDR, between X and Y or W and Z sex chromosomes and given to epigenetic modification and spatial architecture that may regulate not only the acquisition of MSD genes, but also the evolution of sex chromosomes. Additional efforts with amphibians and reptiles, as well as many taxa of teleost fish, will make up the knowledge gaps for full understanding of the diversity of MSD genes and their chromosome evolution.
Gene knockout is well accepted as the functional analysis of genes. However, with the sex trait, pure reliance on gene knockout could lead to incorrect conclusions. This is because knockout of MSD genes is expected to cause sex reversal, but sex reversal can be achieved with many genes involved in the sex determination pathways (Table 1), not just with the MSD gene under study. For example, knockout of bcar1 gene in channel catfish led to sex reversal of genetic males to neofemales [58], but bcar1 was later demonstrated not to be the MSD gene for channel catfish [60,65]. Similarly, with Japanese strain of Nile tilapia (Oreochromis niloticus), knockouts of maleness genes amhy, gsdf or dmrt1, or femaleness genes foxl2 or cyp19a1a all led to sex reversal [66,67,68]. Thus, caution must be exercised when working with certain candidate MSD genes, especially with transcriptional factors and TGF-β cytokines. Fst mapping, when coupled to demonstrated expression profiles at critical time points of sex differentiation, should provide strong positional and expression evidence for the determination of MSD genes. When the size of SDR is very large, there can still be multiple differentially expressed genes (DEGs) in the mapped SDR. Large differences of DEGs within the SDR, even between closely related species such as channel catfish and blue catfish, are suggestive of distinct MSD genes [128]. Detailed analysis of spatial and temporal expression, as well as epigenetic regulation, is required for the identification of MSD genes.
Author Contributions
ZL conceptualized the project, provided funding, and wrote the manuscript, DG reviewed the literature, prepared for tables and figures, and write and revised the manuscript.
Funding
This project was supported by Agriculture and Food Research Initiative (AFRI) Competitive Grant 2023-67015-41839 from the USDA National Institute of Food and Agriculture (NIFA) Animal Reproduction Program to ZL.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
This project was supported by Agriculture and Food Research Initiative (AFRI) Competitive Grant 2023-67015-41839 from the USDA National Institute of Food and Agriculture (NIFA) Animal Reproduction Program to ZL.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Curzon, A.Y.; Shirak, A.; Ron, M.; Seroussi, E. Master-Key Regulators of Sex Determination in Fish and Other Vertebrates—A Review. Int. J. Mol. Sci. 2023, 24, 2468. [Google Scholar] [CrossRef] [PubMed]
- Kitano, J.; Ansai, S.; Takehana, Y.; Yamamoto, Y. Diversity and Convergence of Sex-Determination Mechanisms in Teleost Fish. Annu. Rev. Anim. Biosci. 2024, 12, 233–259. [Google Scholar] [CrossRef]
- Heule, C.; Salzburger, W.; Böhne, A. Genetics of Sexual Development: An Evolutionary Playground for Fish. Genetics 2014, 196, 579–591. [Google Scholar] [CrossRef] [PubMed]
- Trukhina, A.V.; Lukina, N.A.; Wackerow-Kouzova, N.D.; Smirnov, A.F. The Variety of Vertebrate Mechanisms of Sex Determination. BioMed Res. Int. 2013, 2013, 1–8. [Google Scholar] [CrossRef]
- Adolfi, M.C.; Herpin, A.; Schartl, M. The replaceable master of sex determination: bottom-up hypothesis revisited. Philos. Trans. R. Soc. B: Biol. Sci. 2021, 376, 20200090. [Google Scholar] [CrossRef]
- Parker, G.A. Sexual selection and sexual conflict. In Sexual Selection and Reproductive Competition in Insects; Blum, M.S., Blum, N.A., Eds.; Academic Press: New York, 1979; pp. 123–166. [Google Scholar]
- Charlesworth, D.; Charlesworth, B.; Marais, G. Steps in the evolution of heteromorphic sex chromosomes. Heredity 2005, 95, 118–128. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.H.; Hsiung, K.; Böhne, A. Evaluating the role of sexual antagonism in the evolution of sex chromosomes: new data from fish. Curr. Opin. Genet. Dev. 2023, 81, 102078. [Google Scholar] [CrossRef]
- Zhu, Z.; Younas, L.; Zhou, Q. Evolution and regulation of animal sex chromosomes. Nat. Rev. Genet. 2024, 26, 59–74. [Google Scholar] [CrossRef]
- Sinclair, A.H.; Berta, P.; Palmer, M.S.; Hawkins, J.R.; Griffiths, B.L.; Smith, M.J.; Foster, J.W.; Frischauf, A.-M.; Lovell-Badge, R.; Goodfellow, P.N. A gene from the human sex-determining region encodes a protein with homology to a conserved DNA-binding motif. Nature 1990, 346, 240–244. [Google Scholar] [CrossRef]
- Smith, C.A.; Roeszler, K.N.; Ohnesorg, T.; Cummins, D.M.; Farlie, P.G.; Doran, T.J.; Sinclair, A.H. The avian Z-linked gene DMRT1 is required for male sex determination in the chicken. Nature 2009, 461, 267–271. [Google Scholar] [CrossRef]
- Gamble, T.; Castoe, T.A.; Nielsen, S.V.; Banks, J.L.; Card, D.C.; Schield, D.R.; Schuett, G.W.; Booth, W. The Discovery of XY Sex Chromosomes in a Boa and Python. Curr. Biol. 2017, 27, 2148–2153.e4. [Google Scholar] [CrossRef]
- Kuhl, H.; Tan, W.H.; Klopp, C.; Kleiner, W.; Koyun, B.; Ciorpac, M.; Feron, R.; Knytl, M.; Kloas, W.; Schartl, M.; et al. A candidate sex determination locus in amphibians which evolved by structural variation between X- and Y-chromosomes. Nat. Commun. 2024, 15, 4781. [Google Scholar] [CrossRef] [PubMed]
- Force, A.; Lynch, M.; Pickett, F.B.; Amores, A.; Yan, Y.-L.; Postlethwait, J. Preservation of Duplicate Genes by Complementary, Degenerative Mutations. Genetics 1999, 151, 1531–1545. [Google Scholar] [CrossRef]
- Stevison, L.S.; Hoehn, K.B.; Noor, M.A.F. Effects of Inversions on Within- and Between-Species Recombination and Divergence. Genome Biol. Evol. 2011, 3, 830–841. [Google Scholar] [CrossRef]
- Termolino, P.; Falque, M.; Cigliano, R.A.; Cremona, G.; Paparo, R.; Ederveen, A.; Martin, O.C.; Consiglio, F.M.; Conicella, C. Recombination suppression in heterozygotes for a pericentric inversion induces the interchromosomal effect on crossovers in Arabidopsis. Plant J. 2019, 100, 1163–1175. [Google Scholar] [CrossRef]
- Kirkpatrick, M. How and Why Chromosome Inversions Evolve. PLOS Biol. 2010, 8, e1000501. [Google Scholar] [CrossRef]
- Matsuda, M.; Nagahama, Y.; Shinomiya, A.; Sato, T.; Matsuda, C.; Kobayashi, T.; Morrey, C.E.; Shibata, N.; Asakawa, S.; Shimizu, N.; et al. DMY is a Y-specific DM-domain gene required for male development in the medaka fish. Nature 2002, 417, 559–563. [Google Scholar] [CrossRef] [PubMed]
- Herpin, A.; Schartl, M. Plasticity of gene-regulatory networks controlling sex determination: of masters, slaves, usual suspects, newcomers, and usurpators. Embo Rep. 2015, 16, 1260–1274. [Google Scholar] [CrossRef] [PubMed]
- Curzon, A.Y.; Shirak, A.; Benet-Perlberg, A.; Naor, A.; Low-Tanne, S.I.; Sharkawi, H.; Ron, M.; Seroussi, E. Absence of Figla-like Gene Is Concordant with Femaleness in Cichlids Harboring the LG1 Sex-Determination System. Int. J. Mol. Sci. 2022, 23, 7636. [Google Scholar] [CrossRef]
- Ye, H.; Ruan, R.; Song, X.; Fan, J.; Du, H.; Shao, J.; Wang, Y.; Yue, H.; Zhang, T.; Li, C. Identification of a candidate sex-determining gene, ptf1aY, in the Chinese longsnout catfish (Leiocassis longirostris) through high-throughput sequencing. Aquac. Rep. 2023, 32. [Google Scholar] [CrossRef]
- Bertho, S.; Herpin, A.; Branthonne, A.; Jouanno, E.; Yano, A.; Nicol, B.; Muller, T.; Pannetier, M.; Pailhoux, E.; Miwa, M.; et al. The unusual rainbow trout sex determination gene hijacked the canonical vertebrate gonadal differentiation pathway. Proc. Natl. Acad. Sci. 2018, 115, 12781–12786. [Google Scholar] [CrossRef] [PubMed]
- Graves, J.A.M.; Peichel, C.L. Are homologies in vertebrate sex determination due to shared ancestry or to limited options? Genome Biol. 2010, 11, 205–205. [Google Scholar] [CrossRef]
- Yano, A.; Guyomard, R.; Nicol, B.; Jouanno, E.; Quillet, E.; Klopp, C.; Cabau, C.; Bouchez, O.; Fostier, A.; Guiguen, Y. An Immune-Related Gene Evolved into the Master Sex-Determining Gene in Rainbow Trout, Oncorhynchus mykiss. Curr. Biol. 2012, 22, 1423–1428. [Google Scholar] [CrossRef] [PubMed]
- Yano, A.; Nicol, B.; Jouanno, E.; Quillet, E.; Fostier, A.; Guyomard, R.; Guiguen, Y. The sexually dimorphic on the Y-chromosome gene ( sdY ) is a conserved male-specific Y-chromosome sequence in many salmonids. Evol. Appl. 2013, 6, 486–496. [Google Scholar] [CrossRef]
- Ashman, T.-L.; Bachtrog, D.; Blackmon, H.; E Goldberg, E.; Hahn, M.W.; Kirkpatrick, M.; Kitano, J.; E Mank, J.; Mayrose, I.; et al.; The Tree of Sex Consortium Tree of Sex: A database of sexual systems. Sci. Data 2014, 1, 140015–140015. [Google Scholar] [CrossRef]
- Furman, B.L.S.; Metzger, D.C.H.; Darolti, I.; Wright, A.E.; Sandkam, B.A.; Almeida, P.; Shu, J.J.; Mank, J.E. Sex Chromosome Evolution: So Many Exceptions to the Rules. Genome Biol. Evol. 2020, 12, 750–763. [Google Scholar] [CrossRef]
- van Doorn, G.S.; Kirkpatrick, M. Turnover of sex chromosomes induced by sexual conflict. Nature 2007, 449, 909–912. [Google Scholar] [CrossRef]
- Perrin, N. Sex-chromosome evolution in frogs: what role for sex-antagonistic genes? Philos. Trans. R. Soc. B: Biol. Sci. 2021, 376, 20200094. [Google Scholar] [CrossRef]
- Vicoso, B. Molecular and evolutionary dynamics of animal sex-chromosome turnover. Nat. Ecol. Evol. 2019, 3, 1632–1641. [Google Scholar] [CrossRef]
- Chen, S.; Zhang, G.; Shao, C.; Huang, Q.; Liu, G.; Zhang, P.; Song, W.; An, N.; Chalopin, D.; Volff, J.-N.; et al. Whole-genome sequence of a flatfish provides insights into ZW sex chromosome evolution and adaptation to a benthic lifestyle. Nat. Genet. 2014, 46, 253–260. [Google Scholar] [CrossRef]
- Peichel, C.L.; McCann, S.R.; Ross, J.A.; Naftaly, A.F.S.; Urton, J.R.; Cech, J.N.; Grimwood, J.; Schmutz, J.; Myers, R.M.; Kingsley, D.M.; et al. Assembly of the threespine stickleback Y chromosome reveals convergent signatures of sex chromosome evolution. Genome Biol. 2020, 21, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Lahn, B.T.; Page, D.C. Four Evolutionary Strata on the Human X Chromosome. Science 1999, 286, 964–967. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, J.; Yang, W.; An, N.; Zhang, P.; Zhang, G.; Zhou, Q. Temporal genomic evolution of bird sex chromosomes. BMC Evol. Biol. 2014, 14, 250. [Google Scholar] [CrossRef] [PubMed]
- Glasauer, S.M.K.; Neuhauss, S.C.F. Whole-genome duplication in teleost fishes and its evolutionary consequences. Mol. Genet. Genom. 2014, 289, 1045–1060. [Google Scholar] [CrossRef]
- Sandve, S.R.; Rohlfs, R.V.; Hvidsten, T.R. Subfunctionalization versus neofunctionalization after whole-genome duplication. Nat. Genet. 2018, 50, 908–909. [Google Scholar] [CrossRef]
- Peatman, E.; Bao, B.; Peng, X.; Baoprasertkul, P.; Brady, Y.; Liu, Z. Catfish CC chemokines: genomic clustering, duplications, and expression after bacterial infection with Edwardsiella ictaluri. Mol. Genet. Genom. 2005, 275, 297–309. [Google Scholar] [CrossRef] [PubMed]
- Takehana, Y.; Hamaguchi, S.; Sakaizumi, M. Different origins of ZZ/ZW sex chromosomes in closely related medaka fishes, Oryzias javanicus and O. hubbsi. Chromosom. Res. 2008, 16, 801–811. [Google Scholar] [CrossRef]
- Wen, M.; Pan, Q.; Jouanno, E.; Montfort, J.; Zahm, M.; Cabau, C.; Klopp, C.; Iampietro, C.; Roques, C.; Bouchez, O.; et al. An ancient truncated duplication of the anti-Müllerian hormone receptor type 2 gene is a potential conserved master sex determinant in the Pangasiidae catfish family. Mol. Ecol. Resour. 2022, 22, 2411–2428. [Google Scholar] [CrossRef]
- Zheng, S.; Tao, W.; Yang, H.; Kocher, T.D.; Wang, Z.; Peng, Z.; Jin, L.; Pu, D.; Zhang, Y.; Wang, D. Identification of sex chromosome and sex-determining gene of southern catfish (Silurus meridionalis) based on XX, XY and YY genome sequencing. Proceedings Biol. Sci. 2022, 289, 20212645. [Google Scholar] [CrossRef]
- Zheng, S.; Tao, W.; Tao, H.; Yang, H.; Wu, L.; Shao, F.; Wang, Z.; Jin, L.; Peng, Z.; Wang, D.; et al. Characterization of the male-specific region containing the candidate sex-determining gene in Amur catfish (Silurus asotus) using third-generation- and pool-sequencing data. Int. J. Biol. Macromol. 2023, 248, 125908. [Google Scholar] [CrossRef]
- Wang, T.; Gong, G.; Li, Z.; Niu, J.-S.; Du, W.-X.; Wang, Z.-W.; Wang, Y.; Zhou, L.; Zhang, X.-J.; Lian, Z.-Q.; et al. Genomic Anatomy of Homozygous XX Females and YY Males Reveals Early Evolutionary Trajectory of Sex-determining Gene and Sex Chromosomes inSilurusFishes. Mol. Biol. Evol. 2024, 41, msae169. [Google Scholar] [CrossRef] [PubMed]
- Kappas, I.; Vittas, S.; Pantzartzi, C.N.; Drosopoulou, E.; Scouras, Z.G. A Time-Calibrated Mitogenome Phylogeny of Catfish (Teleostei: Siluriformes). PLOS ONE 2016, 11, e0166988. [Google Scholar] [CrossRef] [PubMed]
- Gong, G.; Xiong, Y.; Xiao, S.; Li, X.-Y.; Huang, P.; Liao, Q.; Han, Q.; Lin, Q.; Dan, C.; Zhou, L.; et al. Origin and chromatin remodeling of young X/Y sex chromosomes in catfish with sexual plasticity. Natl. Sci. Rev. 2022, 10, nwac239. [Google Scholar] [CrossRef] [PubMed]
- Natri, H.M.; Merilä, J.; Shikano, T. The evolution of sex determination associated with a chromosomal inversion. Nat. Commun. 2019, 10, 145. [Google Scholar] [CrossRef]
- Checchi, P.M.; Engebrecht, J. Heteromorphic sex chromosomes: Navigating meiosis without a homologous partner. Mol. Reprod. Dev. 2011, 78, 623–632. [Google Scholar] [CrossRef]
- A Ross, J.; Peichel, C.L. Molecular Cytogenetic Evidence of Rearrangements on the Y Chromosome of the Threespine Stickleback Fish. Genetics 2008, 179, 2173–2182. [Google Scholar] [CrossRef]
- Sember, A.; Nguyen, P.; Perez, M.F.; Altmanová, M.; Ráb, P.; Cioffi, M.d.B. Multiple sex chromosomes in teleost fishes from a cytogenetic perspective: state of the art and future challenges. Philos. Trans. R. Soc. B: Biol. Sci. 2021, 376, 20200098. [Google Scholar] [CrossRef]
- Li, M.; Zhang, R.; Fan, G.; Xu, W.; Zhou, Q.; Wang, L.; Li, W.; Pang, Z.; Yu, M.; Liu, Q.; et al. Reconstruction of the Origin of a Neo-Y Sex Chromosome and Its Evolution in the Spotted Knifejaw,Oplegnathus punctatus. Mol. Biol. Evol. 2021, 38, 2615–2626. [Google Scholar] [CrossRef]
- Payer, B.; Lee, J.T. X Chromosome Dosage Compensation: How Mammals Keep the Balance. Annu. Rev. Genet. 2008, 42, 733–772. [Google Scholar] [CrossRef]
- Brockdorff, N.; Turner, B.M. Dosage Compensation in Mammals. Cold Spring Harb. Perspect. Biol. 2015, 7, a019406–a019406. [Google Scholar] [CrossRef]
- A McQueen, H.; McBride, D.; Miele, G.; Bird, A.P.; Clinton, M. Dosage compensation in birds. Curr. Biol. 2001, 11, 253–257. [Google Scholar] [CrossRef] [PubMed]
- Deviatiiarov, R.; Nagai, H.; Ismagulov, G.; Stupina, A.; Wada, K.; Ide, S.; Toji, N.; Zhang, H.; Sukparangsi, W.; Intarapat, S.; et al. Dosage compensation of Z sex chromosome genes in avian fibroblast cells. Genome Biol. 2023, 24, 213. [Google Scholar] [CrossRef]
- White, M.A.; Kitano, J.; Peichel, C.L. Purifying Selection Maintains Dosage-Sensitive Genes during Degeneration of the Threespine Stickleback Y Chromosome. Mol. Biol. Evol. 2015, 32, 1981–1995. [Google Scholar] [CrossRef]
- Darolti, I.; Wright, A.E.; Sandkam, B.A.; Morris, J.; Bloch, N.I.; Farré, M.; Fuller, R.C.; Bourne, G.R.; Larkin, D.M.; Breden, F.; et al. Extreme heterogeneity in sex chromosome differentiation and dosage compensation in livebearers. Proc. Natl. Acad. Sci. 2019, 116, 19031–19036. [Google Scholar] [CrossRef]
- Metzger, D.C.H.; A Sandkam, B.; Darolti, I.; E Mank, J. Rapid Evolution of Complete Dosage Compensation in Poecilia. Genome Biol. Evol. 2021, 13. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Zhou, T.; Gao, D. Genetic and epigenetic regulation of growth, reproduction, disease resistance and stress responses in aquaculture. Front. Genet. 2022, 13, 994471. [Google Scholar] [CrossRef]
- Bao, L.; Tian, C.; Liu, S.; Zhang, Y.; Elaswad, A.; Yuan, Z.; Khalil, K.; Sun, F.; Yang, Y.; Zhou, T.; et al. The Y chromosome sequence of the channel catfish suggests novel sex determination mechanisms in teleost fish. BMC Biol. 2019, 17, 6. [Google Scholar] [CrossRef]
- Yang, Y.; Zhou, T.; Liu, Y.; Tian, C.; Bao, L.; Wang, W.; Zhang, Y.; Liu, S.; Shi, H.; Tan, S.; et al. Identification of an Epigenetically Marked Locus within the Sex Determination Region of Channel Catfish. Int. J. Mol. Sci. 2022, 23, 5471. [Google Scholar] [CrossRef]
- Wang, W.; Wang, W.; Yang, Y.; Yang, Y.; Tan, S.; Tan, S.; Zhou, T.; Zhou, T.; Liu, Y.; Liu, Y.; et al. Genomic imprinting-like monoallelic paternal expression determines sex of channel catfish. Sci. Adv. 2022, 8, adc8786. [Google Scholar] [CrossRef]
- Metzger, D.C.H.; Schulte, P.M. The DNA Methylation Landscape of Stickleback Reveals Patterns of Sex Chromosome Evolution and Effects of Environmental Salinity. Genome Biol. Evol. 2018, 10, 775–785. [Google Scholar] [CrossRef]
- Metzger, D.; Mank, J.E. Conserved sex-biased DNA methylation patterns target key developmental genes and non-recombining region of the guppy sex chromosome. BioRxiv 2020. [Google Scholar] [CrossRef]
- Melamed-Bessudo, C.; Levy, A.A. Deficiency in DNA methylation increases meiotic crossover rates in euchromatic but not in heterochromatic regions in Arabidopsis. Proc. Natl. Acad. Sci. 2012, 109, E981–E988. [Google Scholar] [CrossRef] [PubMed]
- Yelina, N.E.; Lambing, C.; Hardcastle, T.J.; Zhao, X.; Santos, B.; Henderson, I.R. DNA methylation epigenetically silences crossover hot spots and controls chromosomal domains of meiotic recombination in Arabidopsis. Genes Dev. 2015, 29, 2183–2202. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Gao, D. Hydin as the Candidate Master Sex Determination Gene in Channel Catfish (Ictalurus punctatus) and Its Epigenetic Regulation. Mar. Biotechnol. 2024, 27, 6. [Google Scholar] [CrossRef]
- Jiang, D.; Yang, H.; Li, M.; Shi, H.; Zhang, X.; Wang, D. gsdf is a downstream gene of dmrt1 that functions in the male sex determination pathway of the Nile tilapia. Mol. Reprod. Dev. 2016, 83, 497–508. [Google Scholar] [CrossRef] [PubMed]
- Li, M.-H.; Yang, H.-H.; Li, M.-R.; Sun, Y.-L.; Jiang, X.-L.; Xie, Q.-P.; Wang, T.-R.; Shi, H.-J.; Sun, L.-N.; Zhou, L.-Y.; et al. Antagonistic Roles of Dmrt1 and Foxl2 in Sex Differentiation via Estrogen Production in Tilapia as Demonstrated by TALENs. Endocrinology 2013, 154, 4814–4825. [Google Scholar] [CrossRef]
- Zhang, X.; Li, M.; Ma, H.; Liu, X.; Shi, H.; Li, M.; Wang, D. Mutation of foxl2 or cyp19a1a results in female to male sex reversal in XX Nile tilapia. Endocrinology 2017, 158, 2634–2647. [Google Scholar] [CrossRef]
- Hughes, L.C.; Ortí, G.; Huang, Y.; Sun, Y.; Baldwin, C.C.; Thompson, A.W.; Arcila, D.; Betancur-R, R.; Li, C.; Becker, L.; et al. Comprehensive phylogeny of ray-finned fishes (Actinopterygii) based on transcriptomic and genomic data. Proc. Natl. Acad. Sci. USA 2018, 115, 6249–6254. [Google Scholar] [CrossRef]
- Nanda, I.; Kondo, M.; Hornung, U.; Asakawa, S.; Winkler, C.; Shimizu, A.; Shan, Z.; Haaf, T.; Shimizu, N.; Shima, A.; et al. A duplicated copy of DMRT1 in the sex-determining region of the Y chromosome of the medaka, Oryzias latipes. Proc. Natl. Acad. Sci. 2002, 99, 11778–11783. [Google Scholar] [CrossRef]
- Myosho, T.; Takehana, Y.; Hamaguchi, S.; Sakaizumi, M. Turnover of Sex Chromosomes in Celebensis Group Medaka Fishes. G3 Genes|Genomes|Genetics 2015, 5, 2685–2691. [Google Scholar] [CrossRef]
- Mustapha, U.F.; Jiang, D.-N.; Liang, Z.-H.; Gu, H.-T.; Yang, W.; Chen, H.-P.; Deng, S.-P.; Wu, T.-L.; Tian, C.-X.; Zhu, C.-H.; et al. Male-specific Dmrt1 is a candidate sex determination gene in spotted scat (Scatophagus argus). Aquaculture 2018, 495, 351–358. [Google Scholar] [CrossRef]
- Jiang, D.-N.; Huang, Y.-Q.; Zhang, J.-M.; Mustapha, U.F.; Peng, Y.-X.; Huang, H.; Li, G.-L. Establishment of the Y-linked Dmrt1Y as the candidate sex determination gene in spotbanded scat (Selenotoca multifasciata). Aquaculture 2020, 23, 101085. [Google Scholar] [CrossRef]
- Wang, L.; Sun, F.; Wan, Z.Y.; Yang, Z.; Tay, Y.X.; Lee, M.; Ye, B.; Wen, Y.; Meng, Z.; Fan, B.; et al. Transposon-induced epigenetic silencing in the X chromosome as a novel form of dmrt1 expression regulation during sex determination in the fighting fish. BMC Biol. 2022, 20, 5. [Google Scholar] [CrossRef]
- Sun, S.; Song, C.; Han, F.; He, Q.; Liu, J.; Zhang, S.; Han, W.; Ye, K.; Han, Z.; Wang, Z.; et al. Study on sex-linked region and sex determination candidate gene using a high-quality genome assembly in yellow drum. Aquaculture 2022, 563, 738987. [Google Scholar] [CrossRef]
- Cai, M.; Zou, Y.; Xiao, S.; Li, W.; Han, Z.; Han, F.; Xiao, J.; Liu, F.; Wang, Z. Chromosome assembly of Collichthys lucidus, a fish of Sciaenidae with a multiple sex chromosome system. Sci. Data 2019, 6, 132. [Google Scholar] [CrossRef]
- Cui, Y.; Wang, W.; Ma, L.; Jie, J.; Zhang, Y.; Wang, H.; Li, H. New locus reveals the genetic architecture of sex reversal in the Chinese tongue sole (Cynoglossus semilaevis). Heredity 2018, 121, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Lin, A.; Xiao, S.; Xu, S.; Ye, K.; Lin, X.; Sun, S.; Wang, Z. Identification of a male-specific DNA marker in the large yellow croaker (Larimichthys crocea). Aquaculture 2017, 480, 116–122. [Google Scholar] [CrossRef]
- Martínez, P.; Robledo, D.; Taboada, X.; Blanco, A.; Moser, M.; Maroso, F.; Hermida, M.; Gómez-Tato, A.; Álvarez-Blázquez, B.; Cabaleiro, S.; et al. A genome-wide association study, supported by a new chromosome-level genome assembly, suggests sox2 as a main driver of the undifferentiatiated ZZ/ZW sex determination of turbot (Scophthalmus maximus). Genomics 2021, 113, 1705–1718. [Google Scholar] [CrossRef]
- Takehana, Y.; Matsuda, M.; Myosho, T.; Suster, M.L.; Kawakami, K.; Shin-I, T.; Kohara, Y.; Kuroki, Y.; Toyoda, A.; Fujiyama, A.; et al. Co-option of Sox3 as the male-determining factor on the Y chromosome in the fish Oryzias dancena. Nat. Commun. 2014, 5, 4157. [Google Scholar] [CrossRef]
- Pan, Q.; Feron, R.; Yano, A.; Guyomard, R.; Jouanno, E.; Vigouroux, E.; Wen, M.; Busnel, J.-M.; Bobe, J.; Concordet, J.-P.; et al. Identification of the master sex determining gene in Northern pike (Esox lucius) reveals restricted sex chromosome differentiation. PLOS Genet. 2019, 15, e1008013. [Google Scholar] [CrossRef]
- Hattori, R.S.; Kumazawa, K.; Nakamoto, M.; Nakano, Y.; Yamaguchi, T.; Kitano, T.; Yamamoto, E.; Fuji, K.; Sakamoto, T. Y-specific amh allele, amhy, is the master sex-determining gene in Japanese flounder Paralichthys olivaceus. Front. Genet. 2022, 13, 1007548. [Google Scholar] [CrossRef]
- Hattori, R.S.; Murai, Y.; Oura, M.; Masuda, S.; Majhi, S.K.; Sakamoto, T.; Fernandino, J.I; Somoza, G.M.; Yokota, M.; Strüssmann, C.A. A Y-linked anti-Mullerian hormone duplication takes over a critical role in sex determination. Proc. Natl. Acad. Sci. USA 2012, 109, 2955–2959. [Google Scholar] [CrossRef] [PubMed]
- Hattori, R.S.; Somoza, G.M.; Fernandino, J.I.; Colautti, D.C.; Miyoshi, K.; Gong, Z.; Yamamoto, Y.; Strüssmann, C.A. The Duplicated Y-specific amhy Gene Is Conserved and Linked to Maleness in Silversides of the Genus Odontesthes. Genes 2019, 10, 679. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Zhang, Y.; Sarida, M.; Hattori, R.S.; Strüssmann, C.A. Coexistence of Genotypic and Temperature-Dependent Sex Determination in Pejerrey Odontesthes bonariensis. PLOS ONE 2014, 9, e102574. [Google Scholar] [CrossRef]
- Bej, D.K.; Miyoshi, K.; Hattori, R.S.; A Strüssmann, C.; Yamamoto, Y. A Duplicated, Truncated amh Gene Is Involved in Male Sex Determination in an Old World Silverside. G3 Genes|Genomes|Genetics 2017, 7, 2489–2495. [Google Scholar] [CrossRef]
- Curzon, A.Y.; Shirak, A.; Dor, L.; Zak, T.; Perelberg, A.; Seroussi, E.; Ron, M. A duplication of the Anti-Müllerian hormone gene is associated with genetic sex determination of different Oreochromis niloticus strains. Heredity 2020, 125, 317–327. [Google Scholar] [CrossRef]
- M. Li et al., A Tandem Duplicate of Anti-Müllerian Hormone with a Missense SNP on the Y Chromosome Is Essential for Male Sex Determination in Nile Tilapia, Oreochromis niloticus. PLOS Genetics 2015, 11, e1005678. [CrossRef]
- Liu, X.; Dai, S.; Wu, J.; Wei, X.; Zhou, X.; Chen, M.; Tan, D.; Pu, D.; Li, M.; Wang, D. Roles of anti-Müllerian hormone and its duplicates in sex determination and germ cell proliferation of Nile tilapia. Genetics 2021, 220, iyab237. [Google Scholar] [CrossRef]
- Ansai, S.; Montenegro, J.; Masengi, K.W.A.; Nagano, A.J.; Yamahira, K.; Kitano, J. Diversity of sex chromosomes in Sulawesian medaka fishes. J. Evol. Biol. 2022, 35, 1751–1764. [Google Scholar] [CrossRef]
- Song, W.; Xie, Y.; Sun, M.; Li, X.; Fitzpatrick, C.K.; Vaux, F.; O'Malley, K.G.; Zhang, Q.; Qi, J.; He, Y. A duplicated amh is the master sex-determining gene for Sebastes rockfish in the Northwest Pacific. Open Biol. 2021, 11, 210063. [Google Scholar] [CrossRef]
- Sardell, J.M.; Josephson, M.P.; Dalziel, A.C.; Peichel, C.L.; Kirkpatrick, M. Heterogeneous Histories of Recombination Suppression on Stickleback Sex Chromosomes. Mol. Biol. Evol. 2021, 38, 4403–4418. [Google Scholar] [CrossRef] [PubMed]
- Jeffries, D.L.; Mee, J.A.; Peichel, C.L. Identification of a candidate sex determination gene in Culaea inconstans suggests convergent recruitment of an Amh duplicate in two lineages of stickleback. J. Evol. Biol. 2022, 35, 1683–1695. [Google Scholar] [CrossRef]
- Holborn, M.K.; Einfeldt, A.L.; Kess, T.; Duffy, S.J.; Messmer, A.M.; Langille, B.L.; Brachmann, M.K.; Gauthier, J.; Bentzen, P.; Knutsen, T.M.; et al. Reference genome of lumpfish Cyclopterus lumpus Linnaeus provides evidence of male heterogametic sex determination through the AMH pathway. Mol. Ecol. Resour. 2021, 22, 1427–1439. [Google Scholar] [CrossRef]
- Rondeau, E.B.; Laurie, C.V.; Johnson, S.C.; Koop, B.F. A PCR assay detects a male-specific duplicated copy of Anti-Müllerian hormone (amh) in the lingcod (Ophiodon elongatus). BMC Res. Notes 2016, 9, 230–230. [Google Scholar] [CrossRef]
- Nakamoto, M.; Uchino, T.; Koshimizu, E.; Kuchiishi, Y.; Sekiguchi, R.; Wang, L.; Sudo, R.; Endo, M.; Guiguen, Y.; Schartl, M.; et al. A Y-linked anti-Müllerian hormone type-II receptor is the sex-determining gene in ayu, Plecoglossus altivelis. PLOS Genet. 2021, 17, e1009705. [Google Scholar] [CrossRef]
- Qu, M.; Liu, Y.; Zhang, Y.; Wan, S.; Ravi, V.; Qin, G.; Jiang, H.; Wang, X.; Zhang, H.; Zhang, B.; et al. Seadragon genome analysis provides insights into its phenotype and sex determination locus. Sci. Adv. 2021, 7, abg5196. [Google Scholar] [CrossRef]
- Nacif, C.L.; Kratochwil, C.F.; Kautt, A.F.; Nater, A.; Machado-Schiaffino, G.; Meyer, A.; Henning, F. Molecular parallelism in the evolution of a master sex-determining role for the anti-Mullerian hormone receptor 2 gene (amhr2) in Midas cichlids. Mol. Ecol. 2022, 32, 1398–1410. [Google Scholar] [CrossRef]
- Feron, R.; Zahm, M.; Cabau, C.; Klopp, C.; Roques, C.; Bouchez, O.; Eché, C.; Valière, S.; Donnadieu, C.; Haffray, P.; et al. Characterization of a Y-specific duplication/insertion of the anti-Mullerian hormone type II receptor gene based on a chromosome-scale genome assembly of yellow perch, Perca flavescens. Mol. Ecol. Resour. 2020, 20, 531–543. [Google Scholar] [CrossRef]
- Kuhl, H.; Euclide, P.T.; Klopp, C.; Cabau, C.; Zahm, M.; Lopez-Roques, C.; Iampietro, C.; Kuchly, C.; Donnadieu, C.; Feron, R.; et al. Multi-genome comparisons reveal gain-and-loss evolution of anti-Mullerian hormone receptor type 2 as a candidate master sex-determining gene in Percidae. BMC Biol. 2024, 22, 141. [Google Scholar] [CrossRef]
- Kamiya, T.; Kai, W.; Tasumi, S.; Oka, A.; Matsunaga, T.; Mizuno, N.; Fujita, M.; Suetake, H.; Suzuki, S.; Hosoya, S.; et al. A Trans-Species Missense SNP in Amhr2 Is Associated with Sex Determination in the Tiger Pufferfish, Takifugu rubripes (Fugu). PLOS Genet. 2012, 8, e1002798. [Google Scholar] [CrossRef]
- Kabir, A.; Ieda, R.; Hosoya, S.; Fujikawa, D.; Atsumi, K.; Tajima, S.; Nozawa, A.; Koyama, T.; Hirase, S.; Nakamura, O.; et al. Repeated translocation of a supergene underlying rapid sex chromosome turnover in Takifugu pufferfish. Proc. Natl. Acad. Sci. 2022, 119, e2121469119. [Google Scholar] [CrossRef] [PubMed]
- Imarazene, B.; Du, K.; Beille, S.; Jouanno, E.; Feron, R.; Pan, Q.; Torres-Paz, J.; Lopez-Roques, C.; Castinel, A.; Gil, L.; et al. A supernumerary “B-sex” chromosome drives male sex determination in the Pachón cavefish, Astyanax mexicanus. Curr. Biol. 2021, 31, 4800–4809.e9. [Google Scholar] [CrossRef] [PubMed]
- Richter, *!!! REPLACE !!!*; et al. , The TGF-β family member Gdf6Y determines the male sex in Nothobranchius furzeri by suppressing oogenesis-inducing genes. bioRxiv 2023. [Google Scholar] [CrossRef]
- Myosho, T.; Otake, H.; Masuyama, H.; Matsuda, M.; Kuroki, Y.; Fujiyama, A.; Naruse, K.; Hamaguchi, S.; Sakaizumi, M. Tracing the Emergence of a Novel Sex-Determining Gene in Medaka,Oryzias luzonensis. Genetics 2012, 191, 163–170. [Google Scholar] [CrossRef]
- Kaneko, H.; Ijiri, S.; Kobayashi, T.; Izumi, H.; Kuramochi, Y.; Wang, D.-S.; Mizuno, S.; Nagahama, Y. Gonadal soma-derived factor (gsdf), a TGF-beta superfamily gene, induces testis differentiation in the teleost fish Oreochromis niloticus. Mol. Cell. Endocrinol. 2015, 415, 87–99. [Google Scholar] [CrossRef] [PubMed]
- Herpin, A.; Schartl, M.; Depincé, A.; Guiguen, Y.; Bobe, J.; Hua-Van, A.; Hayman, E.S.; Octavera, A.; Yoshizaki, G.; Nichols, K.M.; et al. Allelic diversification after transposable element exaptation promotedgsdfas the master sex determining gene of sablefish. Genome Res. 2021, 31, 1366–1380. [Google Scholar] [CrossRef]
- Edvardsen, R.B.; Wallerman, O.; Furmanek, T.; Kleppe, L.; Jern, P.; Wallberg, A.; Kjærner-Semb, E.; Mæhle, S.; Olausson, S.K.; Sundström, E.; et al. Heterochiasmy and the establishment of gsdf as a novel sex determining gene in Atlantic halibut. PLOS Genet. 2022, 18, e1010011. [Google Scholar] [CrossRef]
- Jasonowicz, A.J.; Simeon, A.; Zahm, M.; Cabau, C.; Klopp, C.; Roques, C.; Iampietro, C.; Lluch, J.; Donnadieu, C.; Parrinello, H.; et al. Generation of a chromosome-level genome assembly for Pacific halibut (Hippoglossus stenolepis) and characterization of its sex-determining genomic region. Mol. Ecol. Resour. 2022, 22, 2685–2700. [Google Scholar] [CrossRef]
- Rafati, N.; Chen, J.; Herpin, A.; Pettersson, M.E.; Han, F.; Feng, C.; Wallerman, O.; Rubin, C.-J.; Péron, S.; Cocco, A.; et al. Reconstruction of the birth of a male sex chromosome present in Atlantic herring. Proc. Natl. Acad. Sci. 2020, 117, 24359–24368. [Google Scholar] [CrossRef]
- Curzon, A.Y.; Dor, L.; Shirak, A.; Meiri-Ashkenazi, I.; Rosenfeld, H.; Ron, M.; Seroussi, E. A novel c.1759T>G variant in follicle-stimulating hormone-receptor gene is concordant with male determination in the flathead grey mullet (Mugil cephalus). G3 Genes|Genomes|Genetics 2020, 11, jkaa044. [Google Scholar] [CrossRef]
- de la Herrán, R.; Hermida, M.; Rubiolo, J.A.; Gómez-Garrido, J.; Cruz, F.; Robles, F.; Navajas-Pérez, R.; Blanco, A.; Villamayor, P.R.; Torres, D.; et al. A chromosome-level genome assembly enables the identification of the follicule stimulating hormone receptor as the master sex-determining gene in the flatfish Solea senegalensis. Mol. Ecol. Resour. 2023, 23, 886–904. [Google Scholar] [CrossRef] [PubMed]
- Koyama, T.; Nakamoto, M.; Morishima, K.; Yamashita, R.; Yamashita, T.; Sasaki, K.; Kuruma, Y.; Mizuno, N.; Suzuki, M.; Okada, Y.; et al. A SNP in a Steroidogenic Enzyme Is Associated with Phenotypic Sex in Seriola Fishes. Curr. Biol. 2019, 29, 1901–1909.e8. [Google Scholar] [CrossRef]
- Purcell, C.M.; Seetharam, A.S.; Snodgrass, O.; Ortega-García, S.; Hyde, J.R.; Severin, A.J. Insights into teleost sex determination from the Seriola dorsalis genome assembly. BMC Genom. 2018, 19, 31. [Google Scholar] [CrossRef]
- Fan, B.; Xie, D.; Li, Y.; Wang, X.; Qi, X.; Li, S.; Meng, Z.; Chen, X.; Peng, J.; Yang, Y.; et al. A single intronic single nucleotide polymorphism in splicing site of steroidogenic enzyme hsd17b1 is associated with phenotypic sex in oyster pompano, Trachinotus anak. Proc. R. Soc. B: Biol. Sci. 2021, 288, 20212245. [Google Scholar] [CrossRef]
- Catanach, A.; Ruigrok, M.; Bowatte, D.; Davy, M.; Storey, R.; Valenza-Troubat, N.; López-Girona, E.; Hilario, E.; Wylie, M.J.; Chagné, D.; et al. The genome of New Zealand trevally (Carangidae: Pseudocaranx georgianus) uncovers a XY sex determination locus. BMC Genom. 2021, 22, 785. [Google Scholar] [CrossRef]
- Nakamura, Y.; Higuchi, K.; Kumon, K.; Yasuike, M.; Takashi, T.; Gen, K.; Fujiwara, A. Prediction of the Sex-Associated Genomic Region in Tunas (Thunnus Fishes). Int. J. Genom. 2021, 2021, 7226353. [Google Scholar] [CrossRef]
- Shen, X.; Hu, J.; Yáñez, J.M.; Gomes, G.B.; Poon, Z.W.J.; Foster, D.; Alarcon, J.F.; Shao, L.; Guo, X.; Shao, Y.; et al. Exploring the cobia (Rachycentron canadum) genome: unveiling putative male heterogametic regions and identification of sex-specific markers. GigaScience 2024, 13, giae034. [Google Scholar] [CrossRef] [PubMed]
- Tao, W.; Xu, L.; Zhao, L.; Zhu, Z.; Wu, X.; Min, Q.; Wang, D.; Zhou, Q. High-quality chromosome-level genomes of two tilapia species reveal their evolution of repeat sequences and sex chromosomes. Mol. Ecol. Resour. 2020, 21, 543–560. [Google Scholar] [CrossRef]
- Curzon, A.Y.; Shirak, A.; Benet-Perlberg, A.; Naor, A.; Low-Tanne, S.I.; Sharkawi, H.; Ron, M.; Seroussi, E. Gene Variant of Barrier to Autointegration Factor 2 (Banf2w) Is Concordant with Female Determination in Cichlids. Int. J. Mol. Sci. 2021, 22, 7073. [Google Scholar] [CrossRef]
- Behrens, K.A.; Koblmüller, S.; Kocher, T.D. Genome assemblies for Chromidotilapia guntheri (Teleostei: Cichlidae) identify a novel candidate gene for vertebrate sex determination, RIN3. Front. Genet. 2024, 15, 1447628. [Google Scholar] [CrossRef]
- Kirubakaran, T.G.; Andersen, Ø.; De Rosa, M.C.; Andersstuen, T.; Hallan, K.; Kent, M.P.; Lien, S. Characterization of a male specific region containing a candidate sex determining gene in Atlantic cod. Sci. Rep. 2019, 9, 116. [Google Scholar] [CrossRef] [PubMed]
- Dan, C.; Lin, Q.; Gong, G.; Yang, T.; Xiong, S.; Xiong, Y.; Huang, P.; Gui, J.-F.; Mei, J. A novel PDZ domain-containing gene is essential for male sex differentiation and maintenance in yellow catfish (Pelteobagrus fulvidraco). Sci. Bull. 2018, 63, 1420–1430. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Liu, H.; Pan, Z.; Cheng, L.; Sun, Y.; Wang, H.; Chang, G.; Wu, N.; Ding, H.; Zhao, H.; et al. Insights into chromosomal evolution and sex determination of Pseudobagrus ussuriensis (Bagridae, Siluriformes) based on a chromosome-level genome. DNA Res. 2022, 29, dsac028. [Google Scholar] [CrossRef] [PubMed]
- Xue, L.; Gao, Y.; Wu, M.; Tian, T.; Fan, H.; Huang, Y.; Huang, Z.; Li, D.; Xu, L. Telomere-to-telomere assembly of a fish Y chromosome reveals the origin of a young sex chromosome pair. Genome Biol. 2021, 22, 203. [Google Scholar] [CrossRef]
- A Kottler, V.; Feron, R.; Nanda, I.; Klopp, C.; Du, K.; Kneitz, S.; Helmprobst, F.; Lamatsch, D.K.; Lopez-Roques, C.; Lluch, J.; et al. Independent Origin of XY and ZW Sex Determination Mechanisms in Mosquitofish Sister Species. Genetics 2020, 214, 193–209. [Google Scholar] [CrossRef]
- Pan, Q.; Feron, R.; Jouanno, E.; Darras, H.; Herpin, A.; Koop, B.; Rondeau, E.; Goetz, F.W.; A Larson, W.; Bernatchez, L.; et al. The rise and fall of the ancient northern pike master sex-determining gene. eLife 2021, 10, e62858. [Google Scholar] [CrossRef]
- Chen, T.; Gao, D.; Su, B.; Dunham, R.; Liu, Z. Transcriptome analyses suggest distinct master sex determination genes in closely related blue catfish and channel catfish. Aquaculture 2025, 603, 742364. [Google Scholar] [CrossRef]
Figure 1.
Schematic presentation of sex determination systems. Sex chromosomes and master sex determination (MSD) genes and their association with temperature (A). Note a single sex determination system and a single master sex determination gene in homeothermic mammals and birds (upper part), but multiple sex determination systems and master sex determination genes in ectothermic vertebrates (lower part). The groups whose sex are determined by temperature are colored in orange (crocodilians, tuatara, and turtles), and the group in which some species have temperature sex determination while others are controlled by genetic determinants are shown in yellow (lizards). (B) Male and female pathway of sex determination, starting with the master sex determination gene, e.g., Sry in mammals, dmrt1 in birds, and various other genes in lower vertebrates. The male and female pathways are antagonistic each other, but this process is regulated by temperature, which affect sex differentiation through expression of aromatase. Note that SRY transcription factor is an upstream of all other MSD genes.
Figure 1.
Schematic presentation of sex determination systems. Sex chromosomes and master sex determination (MSD) genes and their association with temperature (A). Note a single sex determination system and a single master sex determination gene in homeothermic mammals and birds (upper part), but multiple sex determination systems and master sex determination genes in ectothermic vertebrates (lower part). The groups whose sex are determined by temperature are colored in orange (crocodilians, tuatara, and turtles), and the group in which some species have temperature sex determination while others are controlled by genetic determinants are shown in yellow (lizards). (B) Male and female pathway of sex determination, starting with the master sex determination gene, e.g., Sry in mammals, dmrt1 in birds, and various other genes in lower vertebrates. The male and female pathways are antagonistic each other, but this process is regulated by temperature, which affect sex differentiation through expression of aromatase. Note that SRY transcription factor is an upstream of all other MSD genes.
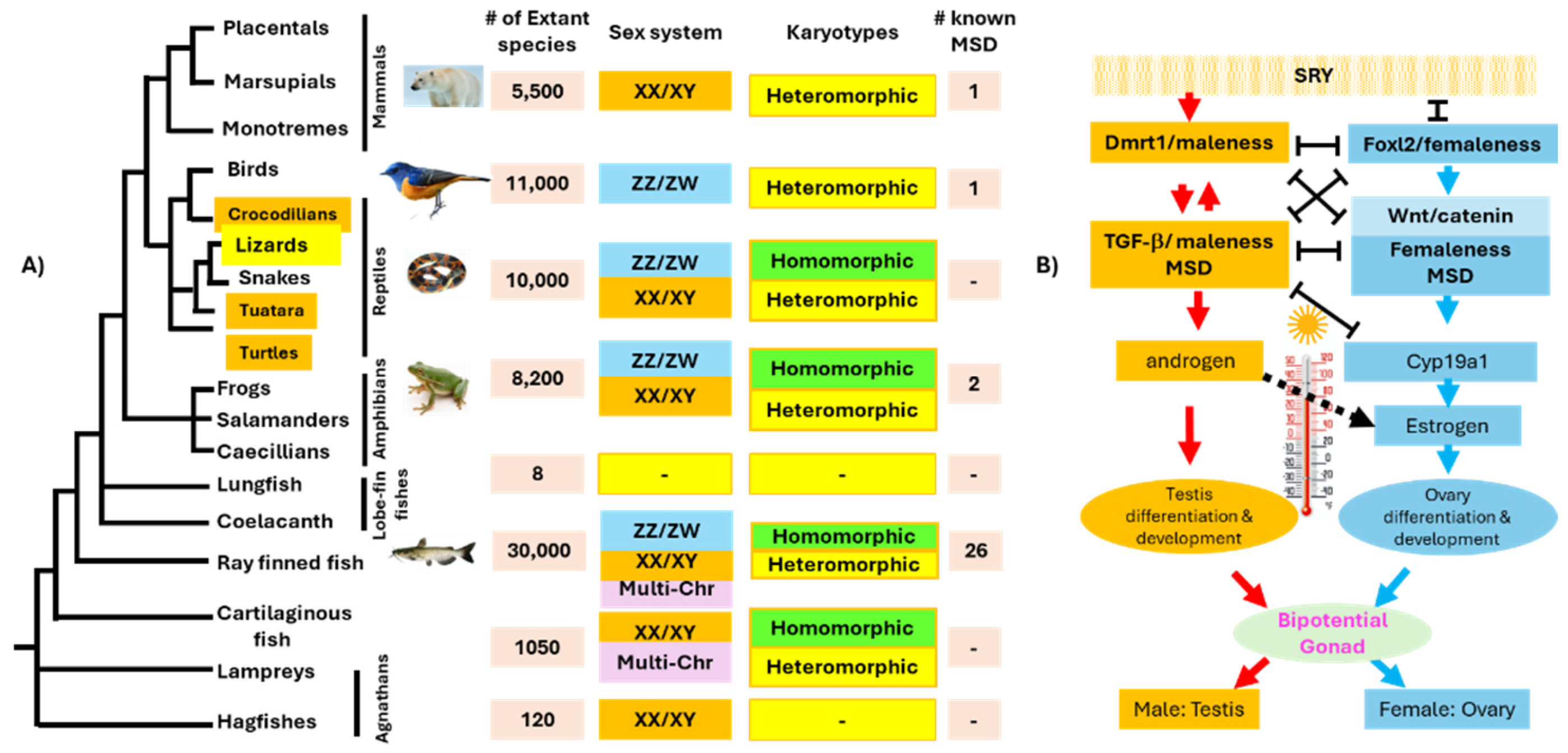
Figure 2.
Schematic presentation of the cause-effect model and predicted paths of sex chromosome evolution. The pathway MSD genes were acquired (the cause) determines the way sex chromosomes evolve (the effect). The way MSD genes were acquired are categorized into two major categories: 1) Simple mutations (left) such as gaining of a new gene, base substitutions, mutations in the promoter region, neofunctionalization, subfunctionalization, tandem gene duplication, and splicing junction mutations, and 2) Structural changes (right) such as large insertions, chromosomal inversion, and fusion and fission. A key element is if non-homologous regions are created during MSD acquisition. With simple mutations, non-homologous regions are not involved, and therefore, no recombination suppression, leading to homomorphic sex chromosomes. Non-homologous sequences reduce recombination, and hence, involve recombination suppression, leading to hoteromorphic sex chromosomes. In the case of major structural change, regions of non-homologous sequences are involved, e.g., in the inserted segment, which lead to recombination suppression. Although recombination suppression causes sequence degeneration and accumulation of transposable elements, recurrent recombination in the reverted regions over time leads to deletions and rearrangements.
Figure 2.
Schematic presentation of the cause-effect model and predicted paths of sex chromosome evolution. The pathway MSD genes were acquired (the cause) determines the way sex chromosomes evolve (the effect). The way MSD genes were acquired are categorized into two major categories: 1) Simple mutations (left) such as gaining of a new gene, base substitutions, mutations in the promoter region, neofunctionalization, subfunctionalization, tandem gene duplication, and splicing junction mutations, and 2) Structural changes (right) such as large insertions, chromosomal inversion, and fusion and fission. A key element is if non-homologous regions are created during MSD acquisition. With simple mutations, non-homologous regions are not involved, and therefore, no recombination suppression, leading to homomorphic sex chromosomes. Non-homologous sequences reduce recombination, and hence, involve recombination suppression, leading to hoteromorphic sex chromosomes. In the case of major structural change, regions of non-homologous sequences are involved, e.g., in the inserted segment, which lead to recombination suppression. Although recombination suppression causes sequence degeneration and accumulation of transposable elements, recurrent recombination in the reverted regions over time leads to deletions and rearrangements.
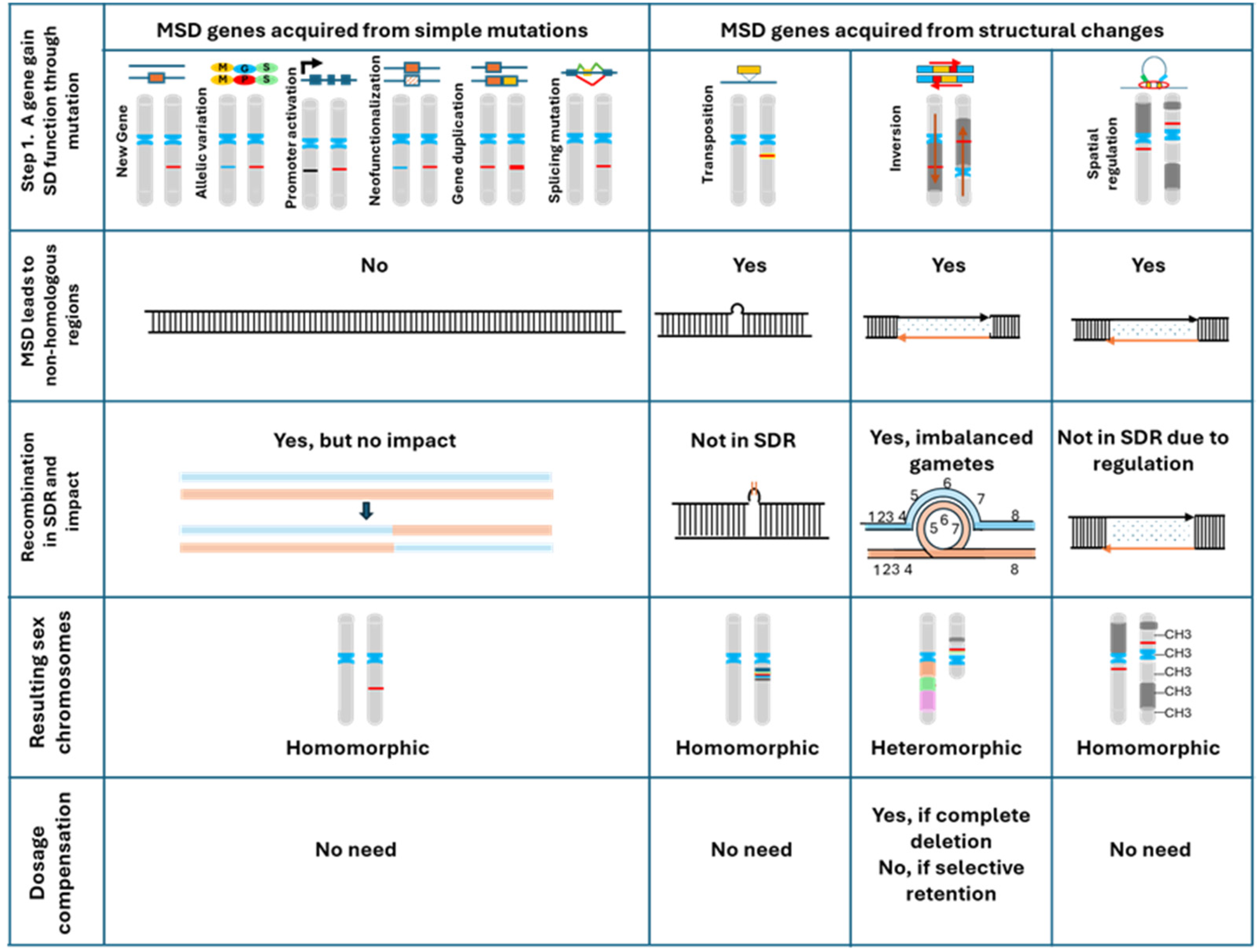
Figure 3.
Divergent evolution of master sex determination (MSD) genes across the teleost fish (Actinopterygii). Shown on the left is the phylogeny of teleost fish, adopted from Hughes et al. [69]. Shown on the right are MSD genes in various categories such as transcription factors (circles), TGF-b (squares), genes involved in steroid synthesis (triangles), and “newcomers” (stars), in various colors and patterns, each representing a specific MSD gene. The symbols are positioned to where they each serve as MSD genes. .
Figure 3.
Divergent evolution of master sex determination (MSD) genes across the teleost fish (Actinopterygii). Shown on the left is the phylogeny of teleost fish, adopted from Hughes et al. [69]. Shown on the right are MSD genes in various categories such as transcription factors (circles), TGF-b (squares), genes involved in steroid synthesis (triangles), and “newcomers” (stars), in various colors and patterns, each representing a specific MSD gene. The symbols are positioned to where they each serve as MSD genes. .
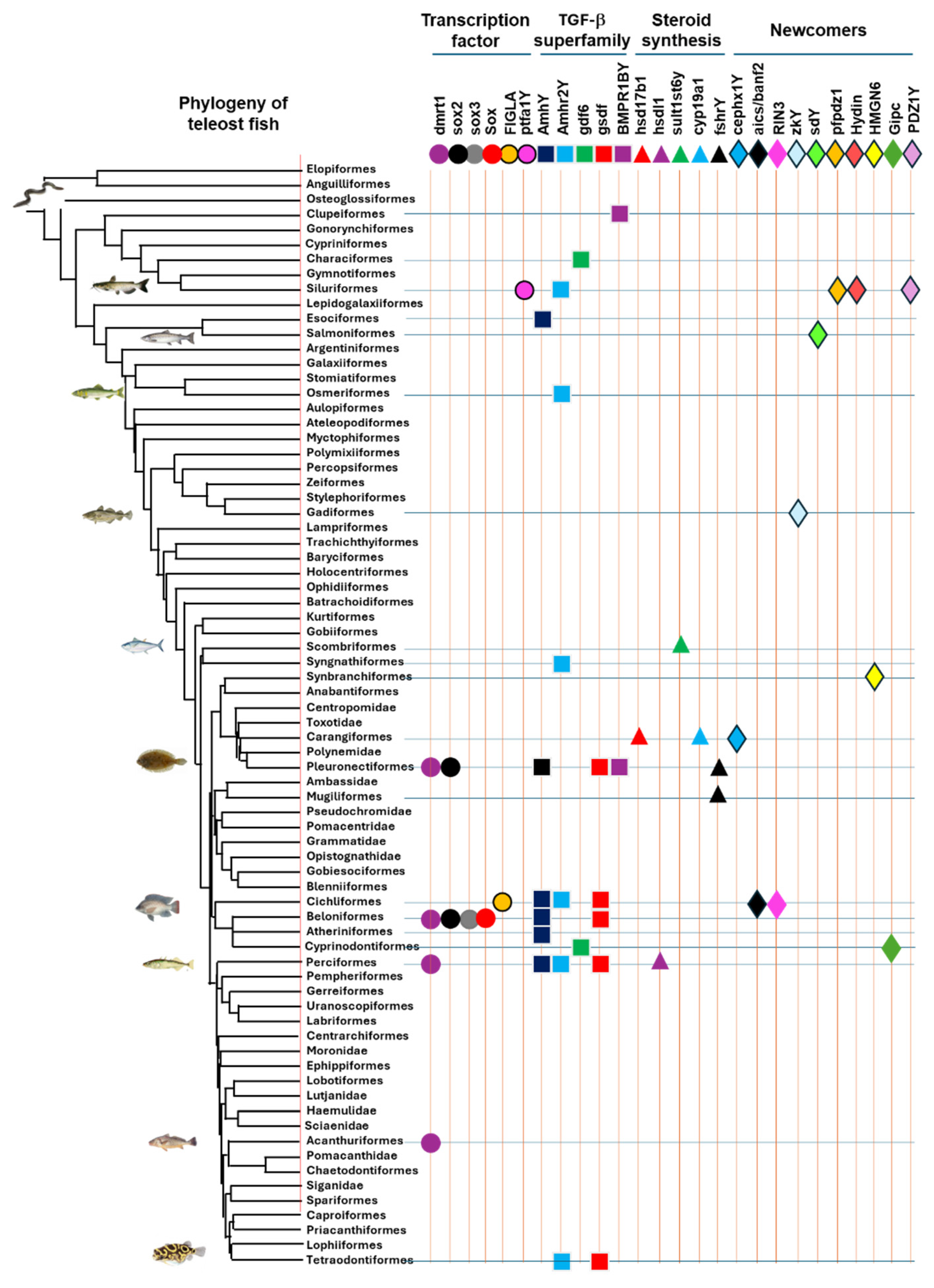
Figure 4.
Recombination involving chromosome inversions. Single crossover of the sex chromosome with a major inversion produces gametes that are not viable because one would contain duplication of one side of the chromosome while the other would contain the other half of the chromosome. Double crossovers produce viable gametes but causing deletions and chromosomal rearrangements. Inversions, therefore, are the driving force of sex chromosome evolution.
Figure 4.
Recombination involving chromosome inversions. Single crossover of the sex chromosome with a major inversion produces gametes that are not viable because one would contain duplication of one side of the chromosome while the other would contain the other half of the chromosome. Double crossovers produce viable gametes but causing deletions and chromosomal rearrangements. Inversions, therefore, are the driving force of sex chromosome evolution.
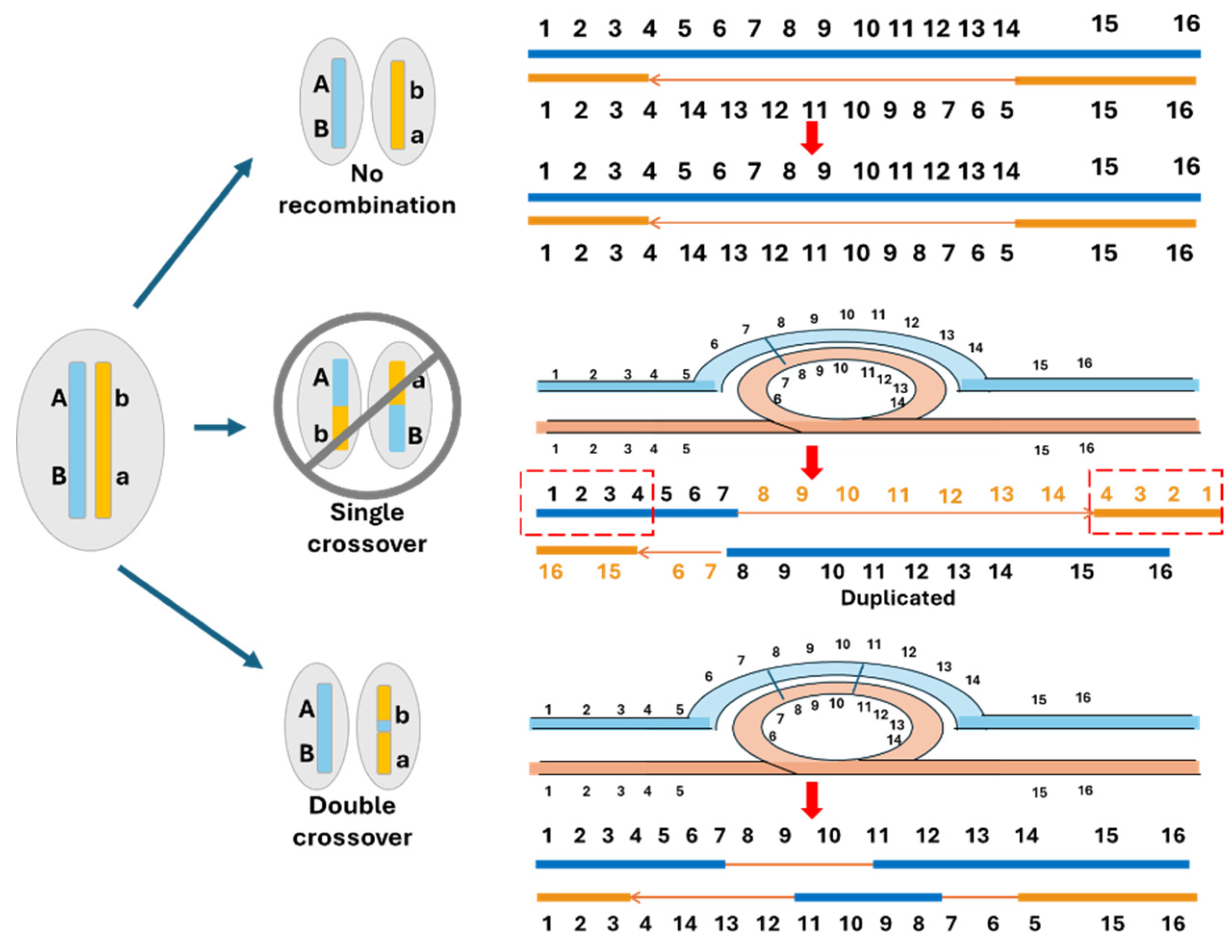

Table 1.
Examples of genes involved in male or female pathway of sex determination, which are antagonistic each other.
Table 1.
Examples of genes involved in male or female pathway of sex determination, which are antagonistic each other.
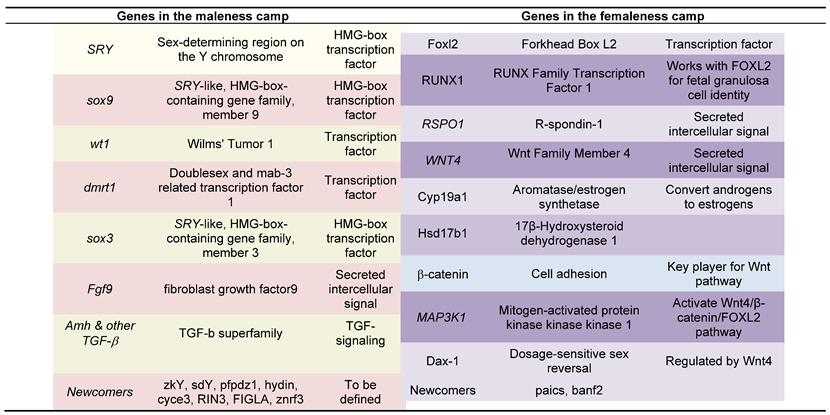 |
Table 2.
Diversity of sex determination (SD) genes and SD systems in vertebrates. ZW sex system is heighted in red. Question mark (?) indicates unknown master sex determination (MSD) genes, but their karyotypes are indicated. *indicates that inversion is likely involved because whole chromosome is non-recombining except a small PAR. The size of sex determination region (SDR) is indicated for those that are well characterized.
Table 2.
Diversity of sex determination (SD) genes and SD systems in vertebrates. ZW sex system is heighted in red. Question mark (?) indicates unknown master sex determination (MSD) genes, but their karyotypes are indicated. *indicates that inversion is likely involved because whole chromosome is non-recombining except a small PAR. The size of sex determination region (SDR) is indicated for those that are well characterized.
| MSD gene | Order or major groups | Common name | Species | MSD gene acquisition | Karyotype | Functional validation | Sex system | References |
|---|---|---|---|---|---|---|---|---|
| Teleost fish | ||||||||
| Dmrt1 | Beloniformes | Japanese medaka | Oryzias latipes | Allelic | Ho | Natural mutation | XY | [17] |
| Dmrt1 | Beloniformes | Hainan medaka | Oryzias curvinotus | Allelic | Ho | - | XY |
[70] |
| Dmrt1 | Beloniformes | Northern medaka | Oryzias Sakaizumii | Allelic | Ho | - | XY |
[71] |
| Dmrt1 | Beloniformes | Hubbs's medaka | Oryzias hubbsi | Inversion |
Hetero | - | ZW | [38] |
| Dmrt1 | Beloniformes | Javanese ricefish | Oryzias javanicus | Inversion |
Hetero | - | ZW | [38] |
| Dmrt1 | Perciformes | Spotted scat | Scatophagus argus | Allelic | Ho | - | XY | [72] |
| Dmrt1 | Perciformes | Spotbanded scat |
Selenotoca multifasciata | Allelic | Ho | - | XY | [73] |
| Dmrt1 | Perciformes | Siamese fighting fish |
Betta splendens |
Allelic | Ho | - | XY | [74] |
| Dmrt1 | Perciformes | Yellow drum |
Nibea albiflora | Allelic | Ho 10 Mb SDR |
- | XY | [75] |
| Dmrt1 | Perciformes | Bighead croaker | Collichthys lucidus | Chromosome fusion generated Y (male 2n=47) female 2n=48 | Hetero | - | X1X1X2X2/X1X2Y | [76] |
| Dmrt1 | Pleuronectiformes |
Chinese tongue sole | Cynoglossus semilaevis | *Whole chromosome non-recombining but a small PAR | Hetero W larger |
knockout |
ZW |
[31] |
| Dmrt1 | Pleuronectiformes | Genko tongue sole | Cynoglossus interruptus | Allelic | Ho | - | ZW | [77] |
| Dmrt1 | Acanthuriformes | Yellow croaker | Larimichthys crocea | Allelic | Ho | - | XY | [78] |
| sox2 | Pleuronectiformes | Turbot | Scophthalmus maximus | Allelic | Ho | - | ZW | [79] |
| sox3Y | Beloniformes | Dwarf medaka | Oryzias minutillus | Allelic | Ho | Transgenic, knockout | XY | [80] |
| sox3Y | Beloniformes | Marmorated ricefish | Oryzias marmoratus | Allelic | Ho | Transgenic, knockout | XY | [80] |
| sox3Y | Beloniformes | Yellow finned medaka | Oryzias profundicola | Allelic | Ho | Transgenic, knockout | XY | [80] |
| sox3Y | Beloniformes |
Indian ricefish | Oryzias dancena | Allelic | Ho | Transgenic, knockout | XY | [80] |
| Sox7 | Beloniformes |
Celebes ricefish |
Oryzias celebensis |
Allelic | Ho | - | XY |
[71] |
| Sox7 | Beloniformes | Matano ricefish | Oryzias matanensis | Allelic | Ho | - | XY | [71] |
| Sox7 | Beloniformes | Wolasi ricefish | Oryzias wolasi | Allelic | Ho | - | XY | [71] |
| Sox7 | Beloniformes | Daisy's ricefish | Oryzias woworae | Allelic | Ho | - | XY | [71] |
| FIGLA | Cichliformes | Tilapia | Oreochromis niloticus LG1 | Allelic | Ho | - | XY | [20] |
| ptfa1 | Siluiriformes | Chinese longsnout catfish | Leiocassis longirostris | Allelic | Ho | - | XY | [21] |
| amhby | Esociformes | Norther pike | Esox Lucius | Tandem duplication | Ho | - | XY | [81,127] |
| amhby | Esociformes | Southern pike | E. cisalpinus | Tandem duplication | Ho | - | XY | [81] |
| amhby | Esociformes | Amur Pike | E. reichertii | Tandem duplication | Ho | - | XY | [81] |
| amhby | Esociformes | Muskellunge | E. masquinongy | Tandem duplication | Ho | - | XY | [81] |
| amhby | Esociformes | Chain pickerel | E. niger | Tandem duplication | Ho | - | XY | [81] |
| amhby | Esociformes | Olympic mudminnow | Novumbra hubbsi | Tandem duplication | Ho | - | XY | [81] |
| amhby | Pleuronectiformes | Olive flounder | Paralichthys olivaceus | Duplication and transposition | Ho | - | XY | [82] |
| amhY | Atheriniformes | Patagonian pejerrey | Odontesthes hatcheri | Duplication and transposition | Ho | Knockdown | XY | [83] |
| amhY | Atheriniformes | Silversides | Odontesthes argentinensis | Duplication and transposition | Ho | - | XY | [84] |
| amhY | Atheriniformes | Silversides | Odontesthes nigricans | Duplication and transposition | Ho | - | XY | [84] |
| amhY | Atheriniformes | Silversides | Odontesthes piquava | Duplication and transposition | Ho | - | XY | [84] |
| amhY | Atheriniformes | Silversides | Odontesthes incisa | Duplication and transposition | Ho | - | XY | [84] |
| amhY | Atheriniformes | Silversides | Odontesthes smitti | Duplication and transposition | Ho | - | XY | [84] |
| amhY | Atheriniformes | Silversides | Odontesthes humensis | Duplication and transposition | Ho | - | XY | [84] |
| amhY | Atheriniformes | Silversides | Odontesthes regia | Duplication and transposition | Ho | - | XY | [84] |
| amhY | Atheriniformes | Silversides | Odontesthes mauleanum | Duplication and transposition | Ho | - | XY | [84] |
| amhY | Atheriniformes | Silversides | Odontesthes perugiae | Duplication and transposition | Ho | - | XY | [84] |
| amhY | Atheriniformes | Pejerrey | Odontesthes bonariensis | Duplication and transposition | Ho | XY | [85] | |
| amhY | Atheriniformes | Cobaltcap silverside | Hypoatherina tsurugae | Truncated duplication on Y | Ho | - | XY | [86] |
| amhY | Cichliformes |
Nile tilapia |
Oreochromis niloticus |
Tandem duplication |
Ho | - |
XY Chr23 | [87,88,89] |
| amhY | Beloniformes | Sulawesian meedaka | Oryzias eversi | Allelic | Ho | - | [90] | |
| amhY | Perciformes |
Black rockfish |
Sebastes schlegelii |
Duplication and transposition | Ho |
Overexpression | XY | [91] |
| amhY | Perciformes | Korean rockfish | Sebastes koreanus | Duplication and transposition | Ho | - | XY | [91] |
| amhY | Perciformes | Rockfish | Sebastes pachycephalus | Duplication and transposition | Ho | - | XY | [91] |
| amhY | Perciformes | Threespine stickleback | Gasterosteus aculeatus | Inversion | Hetero | - | XY | [32] |
| amhY | Perciformes | Japan Sea stickleback | Gasterosteus nipponicus | Inversion followed by chromosome fusion | Hetero |
- | X1X2Y | [92] |
| amhY | Perciformes | Blackspotted stickleback | Gasterosteus wheatlandi | Inversion followed by chromosome fusion | Hetero |
- | X1X2Y | [92] |
| amhY | Perciformes | Brook stickleback | Culaea inconstans | Duplication on Y | Ho | - | XY | [93] |
| amhY | Perciformes | Common lumpfish | Cyclopterus lumpus | Duplication on Y | Ho | - | XY | [94] |
| amhY | Perciformes | Lingcod | Ophiodon elongatus | Duplication on Y | Ho | - | XY | [95] |
| amhY | Siluriformes | Southern catfish | Silurus meridionalis | Transposition | Ho 2.38 Mb SDR |
- | XY | [40] |
| Amhr2Y | Siluriformes | Amur catfish | Silurus asotus | Transposition | Ho 400 Kb SDR |
- | XY | [41] |
| amhr2y | Siluriformes | Lanzhou catfish | Silurus lanzhouensis | Transposition | Ho 400 Kb SDR |
- | XY | [42] |
| Amhr2Y | Siluriformes | Pangasiidae catfishes | Pangasianodon hypophthalmus | Transposition | Ho 320 Kb SDR |
- | XY | [39] |
| Amhr2Y | Siluriformes | Pangasiidae catfishes | Pangasius djambal | Transposition | Ho 320 Kb SDR |
- | XY | [39] |
| Amhr2Y | Siluriformes | Pangasiidae catfishes | Pangasianodon gigas | Transposition | Ho 320 Kb SDR |
- | XY | [39] |
| Amhr2Y | Siluriformes | Pangasiidae catfishes | Pangasius bocourti | Transposition | Ho 320 Kb SDR |
- | XY | [39] |
| Amhr2Y | Siluriformes | Pangasiidae catfishes | Pangasius conchophilus | Transposition | Ho 320 Kb SDR |
- | XY | [39] |
| Amhr2Y | Siluriformes | Pangasiidae catfishes | Pangasius elongatus | Transposition | Ho 320 Kb SDR |
- | XY | [39] |
| Amhr2Y | Siluriformes | Pangasiidae catfishes | Pangasius siamensis | Transposition | Ho 320 Kb SDR |
- | XY | [39] |
| Amhr2Y | Siluriformes | Pangasiidae catfishes | Pangasius macronema | Transposition | Ho 320 Kb SDR |
- | XY | [39] |
| Amhr2Y | Siluriformes | Pangasiidae catfishes | Pangasius larnaudii | Transposition | Ho 320 Kb SDR |
- | XY | [39] |
| Amhr2Y | Siluriformes | Pangasiidae catfishes | Pangasius mekongensis | Transposition | Ho 320 Kb SDR |
- | XY | [39] |
| Amhr2Y | Siluriformes | Pangasiidae catfishes | Pangasius krempfi | Transposition | Ho 320 Kb SDR |
- | XY | [39] |
| Amhr2Y | Siluriformes | Pangasiidae catfishes | Pangasius sanitwongsei | Transposition | Ho 320 Kb SDR |
- | XY | [39] |
| amhr2Y | Osmeriformes | Ayu | Plecoglossus altivelis | Duplication and translocation | Ho 2.03 Mb SDR |
Knockout | XY | [96] |
| amhr2Y | Syngnathiformes | Common seadragon | Phyllopteryx taeniolatus | Truncated duplication and transposition | Ho | - | XY | [97] |
| amhr2Y | Syngnathiformes | Alligator pipefish | Syngnathoides biaculeatus | Truncated duplication and transposition | Ho | - | XY | [97] |
| amhr2Y | Cichliformes | Midas Cichlids | Amphilophus amarillo | Duplication and transposition | Ho | - | XY | [98] |
| amhr2Y | Cichliformes | Midas Cichlids | Amphilophus astorquii | Duplication and transposition | Ho | - | XY | [98] |
| amhr2Y | Cichliformes | Midas Cichlids | Amphilophus chancbo | Duplication and transposition | Ho | - | XY | [98] |
| amhr2Y | Cichliformes | Midas Cichlids | Amphilophus citrinellus | Duplication and transposition | Ho | - | XY | [98] |
| amhr2Y | Cichliformes | Midas Cichlids | Amphilophus flaveolus | Duplication and transposition | Ho | - | XY | [98] |
| amhr2Y | Cichliformes | Midas Cichlids | Amphilophus globosus | Duplication and transposition | Ho | - | XY | [98] |
| amhr2Y | Cichliformes | Midas Cichlids | Amphilophus labiatus | Duplication and transposition | Ho | - | XY | [98] |
| amhr2Y | Cichliformes | Midas Cichlids | Amphilophus sagittae | Duplication and transposition | Ho | - | XY | [98] |
| amhr2Y | Cichliformes | Midas Cichlids | Amphilophus tolteca | Duplication and transposition | Ho | - | XY | [98] |
| amhr2Y | Cichliformes | Midas Cichlids | Amphilophus viridis | Duplication and transposition | Ho | - | XY | [98] |
| amhr2Y | Cichliformes | Midas Cichlids | Amphilophus xiloaensis | Duplication and transposition | Ho | - | XY | [98] |
| amhr2Y | Cichliformes | Midas Cichlids | Amphilophus zaliosus | Duplication and transposition | Ho | - | XY | [98] |
| amhr2Y | Perciformes | Yellow perch | Perca flavescens | Allelic | Ho | - | XY | [99] |
| amhr2Y | Perciformes | Balkhash perch | Perca schrenkii | Allelic | Ho | - | XY | [100] |
| amhr2Y | Perciformes | Walleye | Sander vitreus | Allelic | Ho | - | XY | [100] |
| amhr2Y | Tetraodontiformes | Pufferfish | Takifugu rubripes | Allelic | Ho | - | XY | [101] |
| amhr2Y | Tetraodontiformes | Pufferfish | Takifugu obscurus | Allelic | Ho | - | XY | [102] |
| amhr2Y | Tetraodontiformes | Pufferfish | Takifugu ocellatus | Allelic | Ho | - | XY | [102] |
| amhr2Y | Tetraodontiformes | Pufferfish | Takifugu xanthopterus | Allelic | Ho | - | XY | [102] |
| amhr2Y | Tetraodontiformes | Pufferfish | Takifugu stictonotus | Allelic | Ho | - | XY | [102] |
| amhr2Y | Tetraodontiformes | Pufferfish | Takifugu porphyreus | Allelic | Ho | - | XY | [102] |
| amhr2Y | Tetraodontiformes | Pufferfish | Takifugu poecilonotus | Allelic | Ho | - | XY | [102] |
| amhr2Y | Tetraodontiformes | Pufferfish | Takifugu chrysops | Allelic | Ho | - | XY | [102] |
| amhr2Y | Tetraodontiformes | Pufferfish | Takifugu pardalis | Allelic | Ho | - | XY | [102] |
| amhr2Y | Characiformes | blind cave fish | Astyanax mexicanus | B chromosomes | B chr | Knockout | B chr | [103] |
| gdf6bB | Cyprinodontiformes | African killifish | Nothobranchius furzeri | Allelic | - | Knockout | XY | [104] |
| gdf6bY | Beloniformes | Philippine Medaka | Oryzias luzonensis | Allelic | Ho | Transgenic | XY | [105] |
| gsdfY | Cichliformes | Tilapia | Oreochromis niloticus | Allelic | Ho | Transgenic | XY | [106] |
| gsdfY | Perciformes | Sablefish | Anoplopoma fimbria | Allelic | Ho | - | XY | [107] |
| gsdfY | Pleuronectiformes | Atlantic halibut | Hippoglossus hippoglossus | Allelic | Ho 11.6 Mb SDR |
- | XY Chr12 | [108] |
| gsdfY | Tetraodontiformes | Pufferfish | Takifugu niphobles | Transposition | Ho | XY | [102] | |
| gsdfY | Tetraodontiformes | Pufferfish | Takifugu snyderi | Transposition | Ho | XY | [102] | |
| gsdfY | Tetraodontiformes | Pufferfish | Takifugu vermicularis | Transposition | Ho | XY | [102] | |
| bmpr1ba | Pleuronectiformes | Pacific halibut | Hippoglossus stenolepis | Inversion compared to Chr9 of Atlantic halibut | Ho 12 Mb SDR |
- | ZW Chr9 | [109] |
| bmpr1bbY | Clupeiformes | Atlantic herring | Clupea harengus | Allelic | Ho | - | XY Chr8 | [110] |
| fshrY | Mugiliformes | Flathead grey mullet | Mugil cephalus | Allelic | Ho | - | XY | [111] |
| fshrY | Pleuronectiformes | Senegalese sole | Solea senegalensis | Allelic | Ho | - | XY | [112] |
| Hsd17b1 | Carangiformes | Yellowtail amberjack | Seriola lalandi | Allelic | Ho | - | ZW | [113] |
| Hsd17b1 | Carangiformes | Greater amberjack | Seriola dumerili | Allelic | Ho | - | ZW | [113] |
| Hsd17b1 | Carangiformes | Japanese yellowtail | Seriola quinqueradiata | Allelic | Ho | - | ZW | [113] |
| Hsd17b1 | Carangiformes | California yellowtail | Seriola dorsalis | Allelic | Ho | - | ZW | [114] |
| Hsd17b1 | Carangiformes | Oyster pompano | Trachinotus anak | Allelic | Ho | - | ZW | [115] |
| Cyp19a1 | Carangiformes | Silver trevally | Pseudocaranx georgianus | Allelic | Ho | - | XY | [116] |
| Hsdl1 or Tbc1d32 | Perciformes | European perch | Perca fluviatilis | Allelic | Ho | - | XY | [100] |
| sult1st6y | Scombriformes | Southern bluefin tuna | Thunnus maccoyii | Allelic | Ho | - | XY | [117] |
| sult1st6y | Scombriformes | Pacific bluefin tuna | Thunnus orientalis | Allelic | Ho | - | XY | [117] |
| cephx1Y | Carangiformes | Cobia | Rachycentron canadum | Allelic | Ho 4.04 Mb SDR |
- | XY | [118] |
| Paics/ banf2W | Cichliformes | Blue tilapia | Oreochromis aureus LG3 | Duplication | Ho | - | ZW | [119,120] |
| Paics/ banf2W | Cichliformes | Tanganyika tilapia | Oreochromis tanganicae | Duplication | Ho | - | ZW | [119,120] |
| Paics/ banf2W | Cichliformes | Wami tilapia | Oreochromis hornorum | Duplication | Ho | - | ZW | [119,120] |
| Paics/ banf2W | Cichliformes | Spotted tilapia | Pelmatolapia mariae | Duplication | Ho | - | ZW | [119,120] |
| RIN3 | Cichliformes | - | Chromidotilapia guntheri | Allelic coding region | Ho | - | XY | [121] |
| zkY | Gadiformes | Atlantic cod | Gadus morhua | Allelic | Ho | - | XY | [122] |
| zkY | Gadiformes | Arctic cod | Arctogadus glacialis | Allelic | Ho | - | XY | [122] |
| zkY | Gadiformes | Pacific cod | Gadus macrocephalus | Allelic | Ho | - | XY | [122] |
| sdY | Salmoniformes | Salmonids |
13 species Salmonids |
Transposition/ translocation |
- |
Transgenic and knockout | XY | [24,25] |
| pfpdz1 | Siluiriformes | Yellow catfish | Pelteobagrus fulvidraco | Chromatin architecture, epigenetic regulation | Ho | Overexpression Knockout |
XY | [44,123] |
| Hydin | Siluiriformes | Channel catfish | Ictalurus punctatus | Epigenetic regulation | Ho 8.9 Mb SDR |
Methylation blocker | XY | [59,60] |
| ? (new comer) | Siluiriformes | Ussuri catfish | Pseudobagrus ussuriensis |
Epigenetic regulation |
Ho 16.83 Mb SDR |
- |
XY |
[124] |
| HMGN6/ CYCE3 | Synbranchiformes | The zig-zag eel | Mastacembelus armatus | Allelic | Ho 7.0 Mb SDR |
- | XY | [125] |
| Gipc PDZ1Y? | Cyprinodontiformes | Eastern mosquitofish | Gambusia holbrooki | - | Ho | - | XY | [126] |
| ? | Cyprinodontiformes | Western mosquitofish | Gambusia affinis | W chr larger, fusion? | Hetero | - | ZW | [126] |
| Mammals | ||||||||
| Sry | Mammals | All mammals | Inversion* | Hetero | Knockout Over expression |
XY | [10] | |
| Birds | ||||||||
| Dmrt1 | Birds | All birds | Inversion* | Hetero | Allelic knockout | ZW | [11] | |
| Reptiles | ||||||||
| ? | Python and Boa snakes | - | - | Hetero | - | ZW | [131] | |
| ? | Amazonian puffing snakes and viper snakes | - | - | Ho | - | XY | [12] | |
| Amphibians | ||||||||
| DM-W | African clawed frog | Xenopus laevis | Allelic | Ho | Transgenic | ZW | [66] | |
| Bod1l | European green toad | Bufo viridis | Allelic | Ho | XY | [13] | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



