
Submitted:
14 March 2025
Posted:
17 March 2025
You are already at the latest version
Abstract
Background/Objectives: Chronic hyperglycemia, characteristic of type 2 diabetes mellitus (DM2), promotes the development of complications through oxidative stress. Therefore, it is necessary to strengthen the antioxidant defenses of diabetic patients, even more so if they are elderly. Sechium edule contains high amounts of flavonoids, cucurbitacins, and phenolic acids, which is why it is attributed hypoglycemic and antioxidant properties, so our aim was to determine the antioxidant and hypoglycemic effect of Schium edule in older diabetic patients. Methods: A quasi-experimental study was carried out in a convenience sample of 33 older diabetic adults: (i) placebo group (PG; n = 14); (ii) experimental group (EG; n = 19). Glycosylated hemoglobin (HbA1c), lipoperoxides (LPO), protein carbonylation (PC), Total Oxidant Status (TOS), total antioxidant status (TAS), oxidative stress index (OSI), superoxide dismutase, glutathione peroxidase, and catalase enzyme activity were measured at baseline and three months post-intervention. Results: In the EG, a statistically significant decrease in HbA1c was observed (EG baseline 8.9 2.2 post 7.8 2.0), as well as in the oxidative damage markers LPO, PC, TOS and OSI (LPO baseline 0.243 0.067, post 0.222 0.050; PC baseline 29.4 10, post 19.2 6; TOS baseline 6.0 2.6, post 3.1 1.8; OSI baseline 5.7 3.1 post 2.0 1.1); coupled with a significative increase in TAS (baseline 0.94 0.29 post 1.22 0.28), modifications not observed in the PG. Conclusions: Our results suggest that Sechium edule has a hypoglycemic and antioxidant effect in older adults with DM2.
Keywords:
Sechium edule
; type 2 diabetes mellitus
; hypoglycemic
; antioxidants
; aging
1. Introduction
Type 2 Diabetes Mellitus (DM2) is a metabolic disease characterized by chronic hyperglycemia secondary to defects in the action and production of insulin with alterations in the intermediate metabolism of lipids and proteins. Chronic hyperglycemia favors the development of oxidative stress (OxS), a biochemical alteration that occurs when the production of reactive species exceeds the antioxidant response capacity and oxidative damage to biomolecules occurs with modifications at the cellular, tissue, and systemic levels; this process is linked to the pathophysiology of micro and macroangiopathies, typical complications of this disease [1,3]. Likewise, OxS has been pointed out as a condition present in aging, so older people with DM2 have greater OxS and susceptibility to damage [4].
Given the high prevalence of this disease and the debilitating nature of its complications, it is considered a global health problem due to the high direct and indirect costs associated with its treatment [5]. In México, the prevalence is 18.3% in the adult population and increases to 37% among the population over 60; therefore, finding strategies that contribute to its treatment is a current issue [6]. In this sense, considering that a high percentage of older adults consume alternative natural treatments and based on the background that shows that the bioactive compounds present in the Sechium edule fruit (flavonoids, phenolic acids, and cucurbitacins) have an antioxidant and hypoglycemic effect, we propose the present research, whose objective is to evaluate the effect of consuming the dry extract of Sechium edule in older adults with DM2 [7,8,9,10,11].
2. Materials and Methods
2.1. Experimental Design
The study population was recruited through an open call distributed by printed invitations (leaflets) and digitally (social networks) specifying the objectives of the study and the inclusion criteria, which were to be over 60 years of age, have a clinical diagnosis of DM2, preferably with oral hypoglycemic treatment, without consumption of antioxidants in the three months before the study and physical exercise. The Research Bioethics and Biosafety Committee of the Faculty of Higher Studies Zaragoza approved the study UNAM (FESZ-CE/22-118-22).
2.2. Intervention
A population of n= 60 adults over 60 years were recruited, of which 17 did not meet the inclusion criteria. A sample of n= 43 older adults was studied. After signing the informed consent, they were randomly assigned to two groups: (i) experimental (EG) n= 22, who consumed three 500 mg capsules of Sechium edule per day for three months, and (ii) placebo (PG) n= 21 who consumed three placebo capsules of identical appearance, during the same period. Through the treatment period, three people from the EG and seven from the GP dropped out of the study due to logistical issues, (change of address, medical appointments elsewhere on the same day as the sample collection, lack of a companion to attend appointments) so finally, the data from n=19 in the EG and n=14 in the GP were analyzed. Figure 1 shows the study participants' flow (Figure 1).
The pharmaceutical design of the capsules was carried out at the Faculty of Higher Studies Zaragoza, the fruits of Sechium edule were donated by the Interdisciplinary Research Group on S. edule in Mexico A.C. (GISeM), and the primary treatment (to produce the powder of Sechium edule) was carried out at Faculty of Higher Studies Zaragoza, using food grade equipment. Subsequently, manufacturing and packaging on a larger scale was carried out by a certified pharmaceutical company specialized in nutraceuticals, which delivered the product in bottles containing 90 capsules, which is sufficient treatment for 30 days. The placebo capsules were identical to those of Sechium edule but contained lactose monohydrate and talc, pharmaceutical grade in accordance with the United States Pharmacopeia (USP) (Sigma, St. Louis, MO, USA, EE.UU.).
Phytochemical analysis of Sechium edule capsules revealed that they contained the following amounts of the bioactive compounds 154.8 µg cucurbitacin E, 89.9 µg cucurbitacin B, 6.11 µg cucurbitacin D, and 0.71 µg cucurbitacin I; regarding flavonoids, it contains 48.8 µg of naringenin, 14.2 µg of phlorizin, 0.014 µg of apigenin, 45.5 µg of rutin, 2.38 µg of myricetin, and 1.30 µg of quercetin; finally, it contains phenolic acids such as chlorogenic 1.4 µg, protocatechuic 3.3 µg, caffeic 9.3 µg, ferulic 7.0 µg, syringic 8.7 µg, p-coumaric 1.7 µg, gallic 38.8 µg, and p-hydroxybenzoic acid 0.11 µg [11].
2.3. Biochemical Analysis
2.3.1. Samples
Blood samples were collected by venipuncture after an 8-hour fast into vacutainer/siliconized test tubes without anticoagulant for biochemical determinations (glucose, uric acid, lipid profile, and renal profile) with ethylenediamine tetra acetic acid (EDTA) as anticoagulant for glycated hemoglobin (HbA1c) and heparinized tubes for OxS oxidative stress testing. 600 µL of whole heparinized blood was separated for superoxide dismutase (SOD), 100 µL for gluthatione peroxidase (GPx), 100 µL for catalase (CAT), 100 µL of heparinized plasma for total antioxidant status (TAS), 100 µL of heparinized plasma for total oxidant status (TOS), and 500 µL for lipid peroxidation (LPO). SOD, CAT, TAS, TOS and GPx techniques were performed at microscale in 96-well plates, which were read on a Multiskan Go from Thermo Scientific, Denver, CO, USA. Blood chemistry was performed using colorimetric techniques on a Selectra Junior automated analyzer (Vital Scientific, Dieren, The Netherlands); glycosylated hemoglobin (HbA1c) was determined by an immunoturbidimetry technique in the same analyzer. For all determinations, the intra-assay and inter-assay coefficients of variation were less than 5%.
The measurements were taken from all participants before and after three months of treatment.
2.3.2. Plasma Thiobarbituric Acid Reactive Substances (TBARS)
This test is due to the reaction between thiobarbituric acid (TBA) (0.11 mol/L) (Sigma, St. Louis, MO, USA) and malondialdehyde (MDA) in acid conditions (phosphoric acid, H3PO4, 0.2 mol/L) (Sigma, St. Louis, MO, USA) which products a pink pigment (the adduct TBA-MDA) with absorption at 535 nm. In this assay amplification of the peroxidation is avoided by the addition of butylated hydroxytoluene (BHT) (Sigma, St. Louis, MO, USA) (12.6 mM) [14]. In this assay, 200μL of plasma, 25 μL of BHT, 200μL of H3PO4, and 25 μL of TBA were incubated at 90° for 45 min. The mix was cooled in ice to end the reaction, then 500 μL of butanol (Sigma, St. Louis, MO, USA) and 50 μL of a saturated solution of sodium chloride (NaCl) (Sigma, St. Louis, MO, USA) were added. The absorbance was read at 535 nm and 572 nm to correct for baseline absorption. The quantification was done using a calibration curve. [12]
2.3.3. Proteins Carbonylation
2,4-dinitrophenylhydrazine (DNPH) (Sigma, St. Louis, MO, USA) test was carried out to quantify carbonylated proteins. In this, 20 μL of DNPH (10 mM in 0.5 M H3PO4) and 20μL of plasma were incubated (10 min at room temperature with agitation) and 20 μL of 6 M sodium hydroxide (NaOH) were added. After 10 min of incubation (room temperature and continuous agitation), absorbance was measured at 450 nm against a blank sample [13]. Protein concentration was determined using a commercial Bradford reagent (Bio-Rad, Hercules, CA, USA) and a 1.41 mg/mL standard of bovine serum albumin (BSA). Samples were measured at 595 nm and the results expressed as protein mg/mL.
2.3.4. Whole Blood Superoxide Dismutase
In this technique, the generation of superoxide radicals from xanthine and xanthine Oxidase is performed. The radicals formed react with 2-(4-iodophenyl)-3-(4-nitrophenol)-5-phenyl-tetrazolium chloride to form red formazan, read at 505 nm. The enzyme of the sample inhibits this reaction. It was measured with a commercial kit (Randox Laboratories Ltd., Antrim, UK).
2.3.5. Whole blood Glutathione Peroxidase
The glutathione peroxidase catalyzes the oxidation of glutathione by cumene hydroperoxide in the presence of glutathione reductase and reduced nicotinamide adenine dinucleotide phosphate (NADPH). Oxidized glutathione is converted into the reduced form with the concomitant oxidation of NADPH to nicotinamide adenine dinucleotide phosphate (NADP+). This decrease in absorbance is measured at 340 nm (Randox Laboratories Ltd., Antrim, UK).
The SOD/GPx ratio and the antioxidant gap (AOGAP) were calculated. The AOGAP was calculated using the equation: AOGAP = (TAS mmol- [(albumin (mmol) * 0.69) + uric acid (mmol)] [13].
2.3.6. Catalasa Activity
The activity of the CAT enzyme was determined with an H2O2 inhibition assay using hydrogen peroxide (H2O2) (Sigma, St. Louis, MO, USA) as a substrate, it was a spectrophotometric technique. A working solution was prepared (0.1 M phosphate buffer and 20 mM H2O2), and 10 μl of sample (heparinized total blood) was incorporated into 190μL of working solution. Absorbance detection was performed at 240 nm, inspecting the decay of the H2O2 concentration every 15 s for 2 min [14]. Protein total concentration was determined using a commercial Bradford reagent (Bio-Rad, Hercules, CA, USA) and a 1.41 mg/mL standard of bovine serum albumin (BSA). Samples were measured at 595 nm and the results expressed as protein mg/mL.[14]
The catalase activity was calculated using the equation:
UCAT/g of protein = [(Abs 240 nm/0.0394)*(1000)]/mg/mL of protein].
2.3.7. Plasma Total Antioxidant Status (TAS)
This assay is based on generation of the ABTS+ cation radical, from the reaction of 2,20-azino-bis (3-ethylbenzthiazoline-6-sulfonic acid) (ABTS) with metmyoglobin and H2O2. The bluish-green coloration of this radical is suppressed for the antioxidants in the sample. It was measured at 600 nm. (Randox Laboratories Ltd., Antrim, UK).
2.3.8. Total oxidant status (TOS)
Plasma TOS determination was done using the commercial kit (Rel Assay Diagnostics, Gaziantep, TR), following the supplier’s instructions. This test is based on the oxidation of the ferrous ion (Fe2+)-chelator complex to ferric ion (Fe3+) by the oxidants present in the sample. The Fe3+ forms a complex colored with a chromogen in an acidic medium. The intensity of the color is directly related to the amount of oxidizing molecules present in the simple and is measured spectrophotometrically at 530 nm.
2.4. Statistical Analysis
The results are shown as mean ± standard deviations; after exploratory analysis to corroborate the data distribution, data were analyzed by ANOVA of repeated measures with a significance level of 95%, and paired t test using the IBM SPSS V 20 statistical program (Armonk, NY, USA). Effect size per group was determined using Eta squared (η2). To interpret the results of this test, a partial Eta squared value close to 0.01 is considered low, 0.06 medium, and a value greater than 0.14 large. For all tests, statistical significance was set at a p < 0.05.
3. Results
Regarding biochemical parameters, a statistically significant decrease in HbA1c was observed for the EG compared to placebo group (EG baseline 8.9 ± 2.2 post 7.8 ± 2.0, vs PG baseline 8.1 ± 2.0 post 7.8 ± 1.7). There were no significant differences between the rest of the biochemical markers (Table 1).
A statistically significant decrease in the oxidative damage markers LPO, PC, TOS and OSI was observed in the EG compared to PG. (LPO EG baseline 0.243 ± 0.067, post 0.222 ± 0.050 vs PG baseline 0.338 ± 0.138, post 0.373 ± 0.145; PC, EG baseline 29.4 ± 10, post 19.2 ± 6* vs PG baseline 29.5 ± 10 post 29.4± 15; TOS, EG baseline 6.0 ± 2.6, post 3.1 ± 1.8, vs PG baseline 5.3 ± 2, post 5.4± 3; OSI EG baseline 5.7 ±3.1 post 2.0 ± 1.1 vs PG baseline 6.2 ±3, post 5.5± 4); coupled with a statistically significant increase in TAS (EG baseline 0.94 ± 0.29 post 1.22 ± 0.28 vs PG baseline 1.06 ± 0.28 post 1.09 ± 0.20). As well, the SOD activity decreased in the PG baseline 179 ± 8.6, post 167± 12.9 vs EG baseline 177 ± 13 post 179 ± 17). (Table 2).
The percentage of statistically significant effects for each group on markers of oxidative stress were for EG: LPO -8.6%, PC -34%, TOS -48%, OSI -64%, TAS +30% and AOx GAP +54%, (Figure 2).
And for PG: SOD (-6.7%) and CAT (+51%), (Figure 3).
Finally, regarding effect sizes, the results showed significant improvements with effect sizes from moderate to large in LPO, (η2 = 0.116),TAS (η2 = 0.335), AOX GAP (η2 = 0.306), PC (η2 = 0.628), TOS (η2 = 0.804), and OSI (η2 = 0.656) in EG. While in PG only for SOD (η2 = 0.415) and CAT (η2 = 0.492).
4. Discussion
DM2 is a metabolic disease whose control has become a challenge for health systems worldwide. The hyperglycemia that characterizes it is responsible for the development of sustained alterations that lead to the typical complications of this disease, such as micro and macroangiopathies, which makes this disease a health problem with high direct and indirect costs. Its prevalence is higher among the elderly, who, to maintain health or control their diseases, frequently resort to the use of naturopathic treatments, which is why formal research is important [7,15,16].
In this regard, previous studies conducted in an older population with metabolic syndrome have shown that consuming 1500 mg of Sechium edule fruit powder (three capsules per day) has a hypoglycemic effect; this has already been reported in various preclinical studies and is attributed to the activity of the various compounds present in this fruit [10,17,18,19,20]. In this study, we have observed that in patients with type 2 diabetes mellitus, this hypoglycemic effect also occurs after three months of consumption of the same dose since the percentage of HbA1c was significantly lower in the GE. As mentioned, phytochemical analysis of the capsules has shown that they contain 154.8 µg cucurbitacin E, 89.9 µg cucurbitacin B, 6.11 µg cucurbitacin D, and 0.71 µg cucurbitacin I; Regarding flavonoids, it contains 48.8 µg of naringenin, 14.2 µg of phlorizin, 0.014 µg of apigenin, 45.5 µg of rutin, 2.38 µg of myricetin, and 1.30 µg of quercetin; finally, it contains phenolic acids such as chlorogenic 1.4 µg, protocatechuic 3.3 µg, caffeic 9.3 µg, ferulic 7.0 µg, syringic 8.7 µg, p-coumaric 1.7 µg, gallic 38.8 µg, and p-hydroxybenzoic acid 0.11 µg [11].
Given this varied combination of bioactives, we assume the pharmacological effects result from their synergistic effect. In this sense, regarding the mechanisms involved, there is abundant scientific literature that brings together evidence of the hypoglycemic effect of these biomolecules; the most consistent ones will be described below [18,19,20,21,22]. Regarding flavonoids, it has been reported that quercetin has antidiabetic effects since it promotes insulin secretion, improves insulin resistance and maintains glucose homeostasis [23]. It has been shown to do so by various mechanisms, for example, promotes glucose-stimulated insulin secretion and insulin expression in INS1E (insulin-secreting rat insulinoma) cell; as well as significantly enhance the insulin secretion of INS-1 pancreatic β cells by specifically activating extracellular signal-regulated kinase 1/2 (ERK1/2) [24]. And by activating the intracellular Ca2+ signaling pathway [25,26]. In addition, improves insulin response by upregulating the phosphorylation of insulin receptors (InsRs) and protein kinase (PK). Quercetin treatment restored the impaired protein expression of key insulin signaling molecules, such as IRS-1 and phosphatidyl inositol 3 kinase (PI3K) [27]. In the liver of rats, induced AMPK activation, which reduced glucose production mainly by downregulating key glycogenic isoenzymes, such as glucose-6-phosphate (G6Pase) and phosphoenolpyruvate carboxylase (PEPCK) [28]. The research of Alam et al. shows that quercetin increases glucose uptake by stimulating the translocation of the glucose transporter type 4 (GLUT4) to the plasma membrane in murine skeletal muscle cells. It has been reported that quercetin could promote glucose processing in adipocytes and liver by regulating the activity of sirtuin 1 (SIRT1). It has also been shown to inhibit inflammation and oxidative stress [29,30].
Regarding rutin, proposed mechanisms for its antihyperglycemic effect include a decrease in carbohydrate absorption from the small intestine, inhibition of tissue gluconeogenesis, an increase in tissue glucose uptake, stimulation of insulin secretion from beta cells, and protection of Langerhans islet against degeneration. Rutin also decreases the formation of sorbitol, reactive oxygen species, advanced glycation end-product precursors, and inflammatory cytokines [31,32].
Myricetin has been reported to diminish hyperglycemia by inhibiting carbohydrate digestion by α-glucosidase and α-amylase and acting as a GLP-1R agonist with stimulation of insulin secretion. Additionally, myricetin alleviates the complications through anti-oxidative and anti-inflammatory actions. [33,34]
Phlorizin's fundamental pharmacological action is to cause renal glycosuria and avoid intestinal glucose absorption by inhibiting the sodium–glucose symporters in the proximal renal tubule and mucosa of the small intestine [35].
About cucurbitacins, Kim et al. showed that Cucurbitacin B could significantly reduce the blood glucose level in mice with diabetes. Its hypoglycemic effect was mainly achieved by regulating the intestinal level of AMPK, inducing the release of plasma glucagon-like peptide-1 and insulin, and then affecting mice's appetite and eating behavior [36].
Finally, regarding gallic acid, it has been reported that it facilitates insulin sensitivity and glucose homeostasis in adipocytes. Mechanistically, the activation of PPAR- and C/EBPs simultaneously promoted GLUT4 translocation in adipocytes. Moreover, gallic acid could increase insulin sensitivity by regulating Akt and AMPK signaling pathways, thus showing evidence of dual activation of Akt and AMPK. Additionally, the antidiabetic effects of GA could be mediated via regulation of TNF-α and adipocytokines expression. GA improved the function of the cells by inhibiting caspase-9-related cellular apoptosis [37,38].
Evidently, biomolecules share mechanisms of action, which allow us to assume that a synergistic effect is plausible. Also, in some cases, it is proposed that the hypoglycemic effect is associated with the antioxidant effect since the decrease in the oxidative environment favors a better functioning of the regulatory proteins involved in glucose metabolism. This proposition is consistent with our findings since, as can be seen, the antioxidant effect in those who consumed Sechium edule is evident and coincides with what was observed in previous studies in patients with metabolic syndrome [11,39,40,41].
In this sense, in this investigation, we observed a clear pattern of decreased markers of oxidative damage to biomolecules in the EG, in whom there was a significant decrease in LPO, PC as well as in TOS and OSI, coupled with an increase in TAS. While in the PG, a decrease in SOD activity was observed, accompanied by a striking increase in catalase, which, considering the dynamic nature of the oxidative stress process, allows us to suggest that in this group, the oxidative state associated with hyperglycemia was maintained, which implied a greater demand and wear of antioxidant enzymes.
These effects can be verified by observing the percentage of change in the determined parameters. In the EG, the statistically significant decrease in LPO, PC, TOS, and OSI, together with the increase in TAS and the antioxidant gap, confirmed that there was an antioxidant effect, which was not present in the PG, in whom the decrease in SOD activity was evident, accompanied by an increase in CAT. These data coincide with the intra-group effect sizes, which range from moderate to large, in the same markers, consistently confirming the antioxidant effect of treatment with Sechium edule.
Regarding the mechanisms that explain these findings, abundant evidence from cellular and preclinical studies generally demonstrates that flavonoids can reduce oxidative stress, whether or not it is associated with hyperglycemia. Flavonoids are antioxidants due to the number and arrangement of their phenolic hydroxyl groups attached to ring structures. Their ability to act as antioxidants by donating an electron to an oxidant critically depends on the reduction potentials of their radicals and their accessibility of the radical. The mechanisms determined include direct scavenging of reactive oxygen species, activation of antioxidant enzymes, activation of metal-chelating activity, increasing α-tocopheryl radical levels, inhibiting NADPH oxidases, mitigation of oxidative stress caused by NO, increasing uric acid levels, and increasing the antioxidant properties of low-molecular-weight antioxidants [45,46,47,48,49].
In this regard, concerning the effect observed on LPO, it has been suggested that its decrease may be related to the capacity of biomolecules to activate the Nuclear factor erythroid 2-related factor 2 (Nrf2) pathway, which is a transcriptional factor that regulates the expression of genes that code for enzymes with antioxidant function. Both apigenin and lutein have been shown to activate this pathway. Lutein also provides cytoprotection and reduction of lipid peroxidation by exerting anti-inflammatory effects through the negative regulation of Nuclear factor kappa B (NFκB) and cell death mechanisms in vitro models [50,51].
Besides, quercetin exerts its antioxidant effects by directly inhibiting the production of lipid peroxides, such as malondialdehyde (MDA) and thiobarbituric acid-reactive substances (TBARS) and indirectly promoting the production of endogenous antioxidants. It has been reported that quercetin could ameliorate oxidative stress in STZ-induced diabetic rats by scavenging oxygen free radicals [23,30,52].
It should be noted that previous studies by our research group have shown that the consumption of Sechium edule for six months (dose of 1.5 g/day) decreases the content of LPO and 8-isoprostanes through the activation of the Nrf2 pathway in older adults with metabolic syndrome (MetS). Likewise, this effect coincides with that reported in preclinical studies [11,53,54].
On the other hand, regarding protein carbonylation, which presupposes non-enzymatic oxidative post-translational modification, around 36 carbonylated proteins have been detected in obese diabetic patients, which are involved in molecular signaling, angiogenesis, cell adhesion, and cytoskeletal remodeling [55,56], hence the importance of their oxidation and the relevance of the findings of the present study since we observed a reduction in the content of proteins carbonylated by 34% in EG, which has also been evidenced in shorter intervention times in patients with MetS [17,57]. Regarding the mechanism involved, it has been pointed out that apigenin is a bioactive compound with anti-obesity activity, which improves the activity of antioxidant enzymes and reduces oxidation at the protein level, an effect associated with a decrease in the frequency of the apoptosis-mediated cell death mechanism in β-pancreatic cells in patients with MetS [58]. Likewise, it has been reported that luteolin reduces protein oxidation and improves the intracellular redox state in vitro models [59].
In our study, the decrease in oxidative damage at the lipid and protein levels had a beneficial impact by reducing the total oxidative state (TOS) by 50% and the oxidative stress index (OSI) by 65%. This finding is consistent with the effect observed at the extracellular level where we found a 30% increase in the total antioxidant capacity (TAS) and 54% in the antioxidant gap. These results can be explained by considering the variety of bioactive molecules found in the capsules and coincide with what has been reported by other authors since it has been shown that the different parts that make up the Sechium edule fruit, such as pulp, peel, leaves, seeds, and pedicel, have a high antioxidant potential [11,19,60,61].
Our results show that the DM2 group that consumed Sechium edule for three months presents better antioxidant protection, possibly modulated from transcriptional levels as previously reported, which leads to counteracting oxidative stress. These results have been consistent with previous research reported by our working group [10,11,17,53,57]. These may be due to the bioactive compounds present in chayote, such as cucurbitacin, phenolic acids, and flavonoids, whose antioxidant effects have been widely reported [60,63,64,65,66,67,68,69,70].
In this sense, the antioxidant effect of flavonoids has been consistently reported, so much so that some authors even consider the use of these molecules as alternatives to1 assist in the treatment of diabetes, our results despite the limitations regarding the sample size and duration of the intervention, support the proposal of the consumption of Sechium edule in older adults with type 2 diabetes, since its effectiveness and safety already demonstrated in the treatment of metabolic syndrome is confirmed by our findings, also in patients with diabetes.
5. Conclusions
The results of the present investigation, that is, the decrease in the percentage of HbA1c and the diminution of oxidation markers LPO, CP, TOS, and OSI, coupled with the increase in TAS and the antioxidant gap, suggest that the consumption of Sechium edule powder capsules had a hypoglycemic and antioxidant effect in older adults with DM2.
Author Contributions
V.M.M.-N. designed the study, wrote the manuscript, and analyzed the data. L.D.B.-P. performed the study and wrote the manuscript. J.R.-P. performed the study, wrote the manuscript, and analyzed the data. D.H.-Á., T.L.A.-U., I.A.-S., E.S.-O. and G.G.-G. performed the study and analyzed the data. L.D.B.-P. and J.R.-P contributed equally to the writing of the manuscript. All authors reviewed the final manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by grants from the General Director of Academic Personnel Affairs, National Autonomous University of Mexico (DGAPA-UNAM) (PAPIIT IN223923).
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki and approved by the Ethics Committee of the “National Autonomous University of Mexico (UNAM)—Zaragoza Campus”.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author, [V.M.M.-N.], upon reasonable request.
Conflicts of Interest
No potential conflicts of interest were reported by the authors.
References
- Abel, E.D.; Gloyn AL, Evans-Molina, C.; Joseph, J.J.; Misra, S.; Pajvani, U.V.; Simcox, J.; Susztak, K.; Drucker, D.J. Diabetes mellitus-progress and opportunities in the evolving epidemic. Cell. 2024; 187(15):3789-3820. [CrossRef]
- Surampudi, P.N.; John-Kalarickal, J.; Fonseca, V.A. Emerging concepts in the pathophysiology of type 2 diabetes mellitus. Mt Sinai J Med. 2009; 76(3):216-226. [CrossRef]
- Mahgoub MO, Ali II, Adeghate JO, Tekes K, Kalász H, Adeghate EA. An update on the molecular and cellular basis of pharmacotherapy in type 2 diabetes mellitus. Int J Mol Sci. 2023; 24(11):9328. [CrossRef]
- Mendoza-Núñez, V.M.; Rosado-Pérez, J.; Santiago-Osorio, E.; Ortiz, R.; Sánchez-Rodríguez, M.A.; Galván-Duarte, R.E. Aging linked to type 2 diabetes increases oxidative stress and chronic inflammation. Rejuvenation Res. 2011; 14(1):25-31. [CrossRef]
- GBD 2021 Diabetes Collaborators. Global, regional, and national burden of diabetes from 1990 to 2021, with projections of prevalence to 2050: a systematic analysis for the global burden of disease study 2021. Lancet. 2023; 402(10397):203-234. [CrossRef]
- Basto-Abreu, A.; López-Olmedo, N.; Rojas-Martínez, R.; Aguilar-Salinas, C.A.; Moreno-Banda, G.L.; Carnalla, M.; Rivera, J.A.; Romero-Martínez, M.; Barquera, S.; Barrientos-Gutiérrez, T. Prevalencia de prediabetes y diabetes en México: Ensanut 2022. Salud Publica Mex. 2023; 65(supl1):S163-S168. [CrossRef]
- Agbabiaka, T.B.; Wider, B.; Watson, L.K.; Goodman, C. Concurrent use of prescription drugs and herbal medicinal products in older adults: a systematic review. Drugs Aging. 2017; 34(12):891-905.
- Cadena–Iñiguez, J.; Arévalo-Galarza, L.; Avendaño-Arrazate, C.H.; Soto-Hernández, M.; Ruiz-Posadas, L.M.; Santiago-Osorio, E.; Acosta-Ramos, M.; Cisneros-Solano, V.M.; Aguirre-Medina, J.F.; Ochoa-Martínez, D. Production, genetics, postharvest management and pharmacological characteristics of Sechium edule (Jacq.) Sw. Fresh Produce. 2007; 1(1):41–53.
- Aguiñiga-Sánchez, I.; Cadena-Iñiguez, J.; Santiago-Osorio, E.; Gómez-García, G.; Mendoza-Núñez, V.M.; Rosado-Pérez, J.; Ruíz-Ramos, M.; Cisneros-Solano, V.M.; Ledesma-Martínez, E.; Delgado-Bordonave, A.J.; Soto-Hernández, R.M. Chemical analyses and in vitro and in vivo toxicity of fruit methanol extract of Sechium edule var. nigrum spinosum. Pharm Biol. 2017; 55(1):1638-1645.
- Rosado-Pérez, J.; Aguiñiga-Sánchez, I.; Santiago-OsoriO, E.; Mendoza-Núñez, V.M. Effect of Sechium edule var. nigrum spinosum (chayote) on oxidative stress and pro-inflammatory markers in older adults with metabolic syndrome: an exploratory study. Antioxidants (Basel). 2019; 8(5):146.
- Arista-Ugalde, T.L.; Santiago-Osorio, E.; Monroy-García, A.; Rosado-Pérez, J.; Aguiñiga-Sánchez, I.; Cadena-Iñiguez, J.; Gavia-García, G.; Mendoza-Núñez, V.M. Antioxidant and anti-inflammatory effect of the consumption of powdered concentrate of Sechium edule var. nigrum spinosum in Mexican older adults with metabolic syndrome. Antioxidants (Basel). 2022; 11(6):1076. [CrossRef]
- Jentzsch, A.; Bachmann, H.; Fürst, P.; Bielsaski, H.K. Improved analysis of malondialdehyde in human body fluids. Free Radic Biol Med. 1996; 20:251-256.
- Miller, N.J. Nonvitamin plasma antioxidants. In: Amstrong, D. Free radical and antioxidante protocols. New Jersey: Humana Press; 1998. p.285-297.
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105:121–126.
- Lima, J.E.B.F.; Moreira, N.C.S.; Sakamoto-Hojo, E.T. Mechanisms underlying the pathophysiology of type 2 diabetes: From risk factors to oxidative stress, metabolic dysfunction, and hyperglycemia. Mutat Res Genet Toxicol Environ Mutagen. 2022; 874-875:503437. [CrossRef]
- Lotfy, M.; Adeghate, J.; Kalasz, H.; Singh, J.; Adeghate, E. Chronic complications of diabetes mellitus: a mini review. Curr Diabetes Rev. 2017; 13(1):3-10. [CrossRef]
- Gavia-García, G.; Rosado-Pérez, J.; Aguiñiga-Sánchez, I.; Santiago-Osorio, E.; Mendoza-Núñez, V.M. Effect of Sechium edule var. nigrum spinosum (chayote) on telomerase levels and antioxidant capacity in older adults with metabolic syndrome. Antioxidants (Basel). 2020; 18;9(7):634. [CrossRef]
- Gupta, A.; Jamal, A.; Jamil, D.A.; Al-Aubaidy, H.A. A systematic review exploring the mechanisms by which citrus bioflavonoid supplementation benefits blood glucose levels and metabolic complications in type 2 diabetes mellitus. Diabetes Metab Syndr. 2023; 17(11):102884. [CrossRef]
- Naz, R.; Saqib, F.; Awadallah, S.; Wahid, M.; Latif, M.F.; Iqbal, I.; Mubarak, M.S. Food polyphenols and type II diabetes mellitus: pharmacology and mechanisms. Molecules. 2023; 28(10):3996. [CrossRef]
- Vieira, E.F.; Pinho, O.; Ferreira, I.M.P.L.V.O. Chayote (Sechium edule): a review of nutritional composition, bioactivities and potential applications. Food Chem. 2019 Delerue-Matos C. Chayote (Sechium edule): a review of nutritional composition, bioactivities and potential applications. Food Chem. 2019; 1;275:557-568. [CrossRef]
- Rana, A.; Samtiya, M.; Dhewa, T.; Mishra, V.; Aluko, R.E. Health benefits of polyphenols: a concise review. J Food Biochem. 2022; 46(10):e14264. [CrossRef]
- Munir, K.M.; Chandrasekaran, S.; Gao, F.; Quon, M.J. Mechanisms for food polyphenols to ameliorate insulin resistance and endothelial dysfunction: therapeutic implications for diabetes and its cardiovascular complications. Am J Physiol Endocrinol Metab. 2013; 305(6):E679-E686. [CrossRef]
- Yi, H.; Peng, H.; Wu, X.; Xu, X.; Kuang, T.; Zhang, J.; Du, L.; Fan, G. The therapeutic effects and mechanisms of quercetin on metabolic diseases: pharmacological data and clinical evidence. Oxid Med Cell Longev. 2021; 1:6678662. [CrossRef]
- Youl, E.; Bardy, G.; Magous, R.; Cros, G.; Sejalon, F.; Virsolvy, A.; Richard, S.; Quignard, J.F.; Gross, R.; Petit, P.; Bataille, D.; Oiry, C. Quercetin potentiates insulin secretion and protects INS-1 pancreatic β-cells against oxidative damage via the ERK1/2 pathway. Br J Pharmacol. 2010; 161(4):799-814. [CrossRef]
- Bardy, G.; Virsolvy, A.; Quignard, J.F.; Ravier, M.A.; Bertrand, G.; Dalle, S.; Cros, G.; Magous, R.; Richard, S.; Oiry, C. Quercetin induces insulin secretion by direct activation of L-type calcium channels in pancreatic beta cells. Br J Pharmacol. 2013; 169(5):1102-1113. [CrossRef]
- Hii, C.S.; Howell, S.L. Effects of flavonoids on insulin secretion and 45Ca2+ handling in rat islets of Langerhans. J Endocrinol. 1985; 107(1):1-8. PMID: 3900267. [CrossRef]
- Sandep, M.S.; Nandini, C.D. Influence of quercetin, naringenin and berberine on glucose transporters and insulin signalling molecules in brain of streptozotocin-induced diabetic rats. Biomed Pharmacother. 2017; 94:605-611. [CrossRef]
- Eid, H.M.; Nachar, A.; Thong, F.; Sweeney, G.; Haddad, P.S. The molecular basis of the antidiabetic action of quercetin in cultured skeletal muscle cells and hepatocytes. Pharmacogn Mag. 2015; 11(41):74-81. [CrossRef]
- Peng, J.; Li, Q.; Li, K.; Zhu, L.; Lin, X.; Lin, X.; Shen, Q.; Li, G.; Xie, X. Quercetin improves glucose and lipid metabolism of diabetic rats: involvement of Akt signaling and SIRT1. J Diabetes Res. 2017; 2017:3417306. [CrossRef]
- Alam, M.M.; Meerza, D.; Naseem, I. Protective effect of quercetin on hyperglycemia, oxidative stress and DNA damage in alloxan induced type 2 diabetic mice. Life Sci. 2014; 109(1):8-14. [CrossRef]
- Ghorbani, A. Mechanisms of antidiabetic effects of flavonoid rutin. Biomed Pharmacother. 2017; 96:305-312. [CrossRef]
- Liu, Y.; Sun, Z.; Dong, R.; Liu, P.; Zhang, X.; Li, Y.; Lai, X.; Cheong, H.F.; Wu, Y.; Wang, Y.; Zhou, H.; Gui, D.; Xu, Y. Rutin ameliorated lipid metabolism dysfunction of diabetic NAFLD via AMPK/SREBP1 pathway. Phytomedicine. 2024; 126:155437. [CrossRef]
- Niisato, N.; Marunaka, Y. Therapeutic potential of multifunctional myricetin for treatment of type 2 diabetes mellitus. Front Nutr. 2023; 10:1175660. [CrossRef]
- Li, Y.; Zheng, X.; Yi, X.; Liu, C.; Kong, D.; Zhang, J.; Gong, M. Myricetin: a potent approach for the treatment of type 2 diabetes as a natural class B GPCR agonist. FASEB J. 2017; 31(6):2603-2611. [CrossRef]
- Ehrenkranz, J.R.; Lewis, N.G.; Kahn, C.R.; Roth, J. Phlorizin: a review. Diabetes Metab Res Rev. 2005; 21(1):31-38. [CrossRef]
- Kim, K.H.; Lee, I.S.; Park, J.Y.; Kim, Y.; An, E.J.; Jang, H.J. Cucurbitacin B induces hypoglycemic effect in diabetic mice by regulation of AMP-activated protein kinase alpha and glucagon-like peptide-1 via bitter taste receptor signaling. Front Pharmacol. 2018; 9:1071. [CrossRef]
- Variya, B.C.; Bakrania, A.K.; Patel, S.S. Antidiabetic potential of gallic acid from emblica officinalis: improved glucose transporters and insulin sensitivity through PPAR-γ and Akt signaling. Phytomedicine. 2020; 73:152906. [CrossRef]
- Gandhi, G.R.; Jothi, G.; Antony, P.J.; Balakrishna, K.; Paulraj, M.G.; Ignacimuthu, S.; Stalin, A.; Al-Dhabi, N.A. Gallic acid attenuates high-fat diet fed-streptozotocin-induced insulin resistance via partial agonism of PPARγ in experimental type 2 diabetic rats and enhances glucose uptake through translocation and activation of GLUT4 in PI3K/p-Akt signaling pathway. Eur J Pharmacol. 2014; 745:201-216. [CrossRef]
- Jin, Y.; Arroo, R. The protective effects of flavonoids and carotenoids against diabetic complications- a review of in vivo evidence. Front Nutr. 2023; 10:1020950. [CrossRef]
- Chen, L.; Pu, Y.; Xu, Y.; He, X.; Cao, J.; Ma, Y.; Jiang, W. Anti-diabetic and anti-obesity: Efficacy evaluation and exploitation of polyphenols in fruits and vegetables. Food Res Int. 2022; 157:111202. [CrossRef]
- Caro-Ordieres, T.; Marín-Royo, G.; Opazo-Ríos, L.; Jiménez-Castilla, L.; Moreno, J.A.; Gómez-Guerrero, C.; Egido, J. The coming age of flavonoids in the treatment of diabetic complications. J Clin Med. 2020; 9(2):346. [CrossRef]
- Kawser, H.M.; Abdal, D.A.; Han, J.; Yin, Y.; Kim, K.; Kumar, S.S.; Yang, G.M.; Choi, H.Y.; Cho, S.G. Molecular mechanisms of the anti-obesity and anti-diabetic properties of flavonoids. Int J Mol Sci. 2016; 17(4):569. [CrossRef]
- Jasmin, J.V. A review on molecular mechanism of flavonoids as antidiabetic agents. Mini Rev Med Chem. 2019; 19(9):762-786. [CrossRef]
- Al-Ishaq, R.K.; Abotaleb, M.; Kubatka, P.; Kajo, K.; Büsselberg, D. Flavonoids and their anti-diabetic effects: cellular mechanisms and effects to improve blood sugar levels. Biomolecules. 2019; 9(9):430. [CrossRef]
- Shen, N.; Wang, T.; Gan, Q.; Liu, S.; Wang, L.; Jin, B. Plant flavonoids: classification, distribution, biosynthesis, and antioxidant activity. Food Chem. 2022; 383:132531. Epub 2022 Feb 23. PMID: 35413752. [CrossRef]
- Chagas, M.D.S.S.; Behrens, M.D.; Moragas-Tellis, C.J.; Penedo, G.X.M.; Silva, A.R.; Gonçalves-de-Albuquerque, C.F. Flavonols and flavones as potential anti-inflammatory, antioxidant, and antibacterial compounds. Oxid Med Cell Longev. 2022; 2022:9966750. [CrossRef]
- Pietta, PG. Flavonoids as antioxidants. J Nat Prod. 2000; 63(7):1035-1042. [CrossRef]
- Williams, R,J.; Spencer, J.P.; Rice-Evans, C. Flavonoids: antioxidants or signalling molecules? Free Radic Biol Med. 2004; 36(7):838-849. [CrossRef]
- Rice-Evans, C. Flavonoid antioxidants. Curr Med Chem. 2001; 8(7):797-807. [CrossRef]
- Zhang, B.; Wang, J.; Zhao, G.; Lin, M.; Lang, Y.; Zhang, D.; Feng, D.; Tu, C. Apigenin protects human melanocytes against oxidative damage by activation of the Nrf2 pathway. Cell Stress Chaperones. 2020; 25(2):277-285.
- Qiao, Y.Q.; Jiang, P.F.; Gao, Y.Z. Lutein prevents osteoarthritis through Nrf2 activation and downregulation of inflammation. Arch Med Sci. 2018; 14(3):617-624.
- Iskender, H.; Dokumacioglu, E.; Sen, T.M.; Ince, I.; Kanbay, Y.; Saral, S. The effect of hesperidin and quercetin on oxidative stress, NF-κB and SIRT1 levels in a STZ-induced experimental diabetes model. Biomed Pharmacother. 2017; 90:500-508. [CrossRef]
- Gavia-García, G.; Hernández-Álvarez, D.; Arista-Ugalde, T.L.; Aguiñiga-Sánchez, I.; Santiago-Osorio, E.; Mendoza-Núñez V.M.; Rosado-Pérez, J. The supplementation of Sechium edule var. nigrum spinosum (chayote) promotes Nrf2-mediated antioxidant protection in older adults with metabolic syndrome. Nutrients. 2023; 15(19):4106.
- Wahyuningtyas, A.P.; Putri, D.P.; Maharani, N.; Al-Baarri, A.N.M. Flavonoid fraction from chayote (Sechium edule (Jacq.) Sw) leaves reduced malondialdehyde (MDA) and tumor necrosis factor-α (TNF-α) in hyperuricemic rats. Nutr Food Sci. 2022; 52(2):366-378.
- Hecker, M.; Wagner, A.H. Role of protein carbonylation in diabetes. J Inherit Metab Dis. 2018; 41(1):29-38.
- Bollineni, R.C.; Fedorova, M.; Blüher, M.; Hoffmann, R. Carbonylated plasma proteins as potential biomarkers of obesity induced type 2 diabetes mellitus. J Proteome Res. 2014; 13(11):5081-5093.
- Gavia-García, G.; Rosado-Pérez, J.; Arista-Ugalde, T.L.; Aguiñiga-Sánchez, I.; Santiago-Osorio, E.; Mendoza-Núñez, V.M. The consumption of Sechium edule (chayote) has antioxidant effect and prevents telomere attrition in older adults with metabolic syndrome. Redox Rep. 2023; 28(1):2207323. [CrossRef]
- Javadi, B.; Sobhani, Z. Role of apigenin in targeting metabolic syndrome: A systematic review. Iran J Basic Med Sci. 2024; 27(5):524-534.
- Gao, S.; Qin, T.; Liu, Z.; Caceres, M.A.; Ronchi, C.F.; Chen, C.Y.; Yeum, K.J.; Taylor, A.; Blumberg, J.B.; Liu, Y.; Shang, F. Lutein and zeaxanthin supplementation reduces H2O2-induced oxidative damage in human lens epithelial cells. Mol Vis. 2011; 17:3180-90.
- Ordonez, A.A.L.; Gomez, J.D.; Vattuone, M.A. Antioxidant activities of Sechium edule (Jacq.) Swartz extracts. Food Chem. 2006; 97(3):452-458.
- Fidrianny, I.; Ayu, D.; Hartati, R. Antioxidant capacities, phenolic, flavonoid and carotenoid content of various polarities extracts from three organs of Sechium edule (Jacq.) Swartz. J Chem Pharm Res. 2015; 7(5):914-920.
- Loizzo, M.R.; Bonesi, M.; Menichini, F.; Tenuta, M.C.; Leporini, M.; Tundis, R. Antioxidant and carbohydrate-hydrolysing enzymes potential of Sechium edule (Jacq.) Swartz (Cucurbitaceae) peel, leaves and pulp fresh and processed. Plant Foods Hum Nutr. 2016; 71:381–387.
- Shahwan, M.; Alhumaydhi, F.; Ashraf, G.M.; Hasan, P.M.Z.; Shamsi, A. Role of polyphenols in combating Type 2 Diabetes and insulin resistance. Int J Biol Macromol. 2022; 206:567-579. [CrossRef]
- Dragan, S.; Andrica, F.; Serban, M.C.; Timar, R. Polyphenols-rich natural products for treatment of diabetes. Curr Med Chem. 2015; 22(1):14-22. [CrossRef]
- Xu, Y.; Tang, G.; Zhang, C.; Wang, N.; Feng, Y. Gallic acid and diabetes mellitus: its association with oxidative stress. Molecules. 2021; 26(23):7115. [CrossRef]
- Ganguly, R.; Singh, S.V.; Jaiswal, K.; Kumar, R.; Pandey, A.K. Modulatory effect of caffeic acid in alleviating diabetes and associated complications. World J Diabetes. 2023; 14(2):62-75. [CrossRef]
- Naveed, M.; Hejazi, V.; Abbas, M.; Kamboh, A.A.; Khan, G.J.; Shumzaid, M.; Ahmad, F.; Babazadeh, D.; FangFang, X.; Modarresi-Ghazani, F.; Wen, H.L.; Xiao. H. Z. Chlorogenic acid (CGA): A pharmacological review and call for further research. Biomed Pharmacother. 2018; 97:67-74. [CrossRef]
- Thomas, S.D.; Jha, N.K.; Jha, S.K.; Sadek, B.; Ojha, S. Pharmacological and molecular insight on the cardioprotective role of apigenin. Nutrients. 2023; 15(2):385. [CrossRef]
- Shilpa, V.S.; Shams, R.; Dash, K.K.; Pandey, V.K.; Dar, A.H.; Ayaz, M. S.; Harsányi, E.; Kovács, B. Phytochemical properties, extraction, and pharmacological benefits of naringin: a review. Molecules. 2023; 28(15):5623. [CrossRef]
- Dai, S.; Wang, C.; Zhao, X.; Ma, C.; Fu, K.; Liu, Y.; Peng, C.; Li, Y. Cucurbitacin B: A review of its pharmacology, toxicity, and pharmacokinetics. Pharmacol Res. 2023; 187:106587. [CrossRef]
Figure 1.
Flowchart of the participant monitoring process.
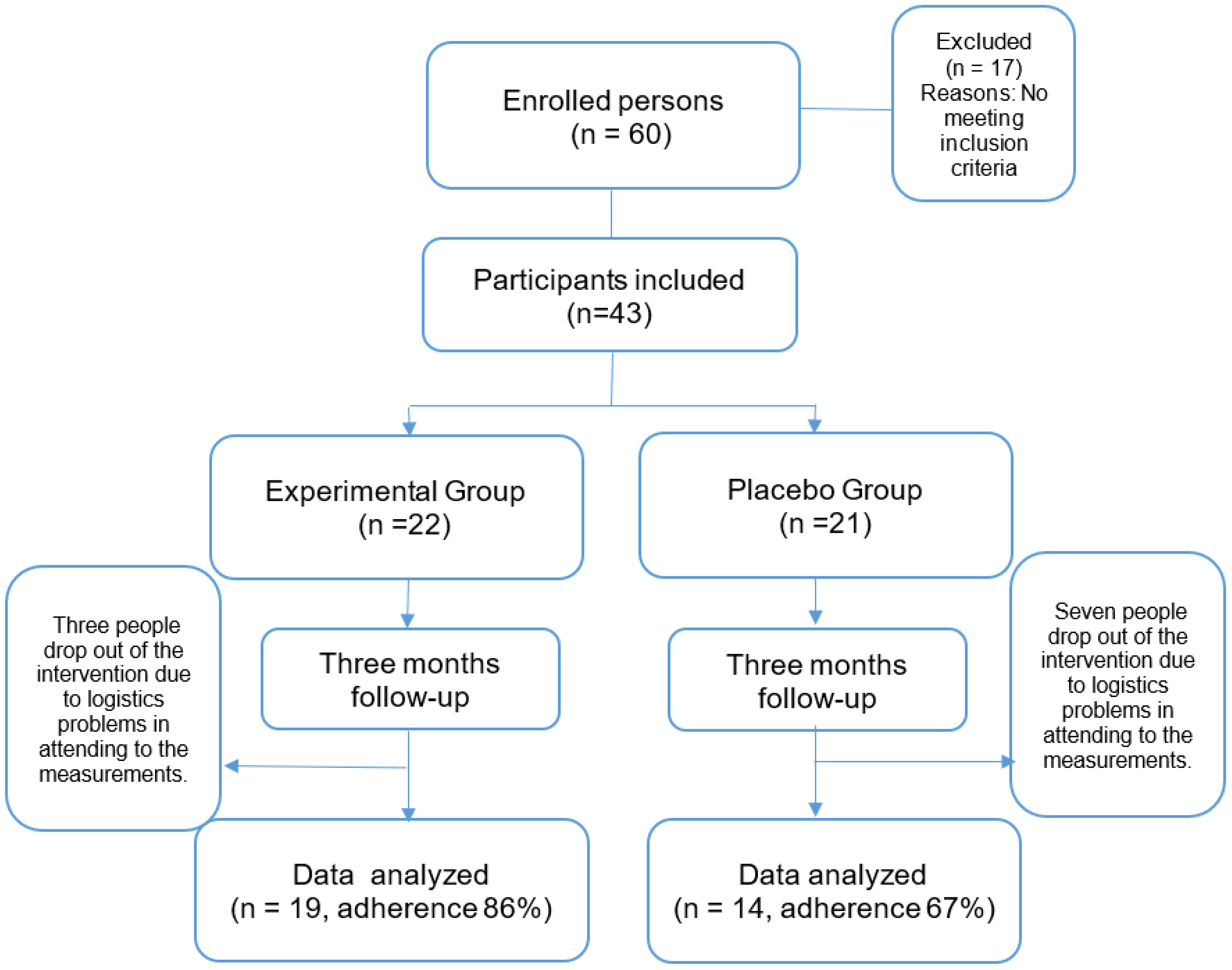
Figure 2.
Percentage changes [Δ% = (Post–Pre)/Pre*100] in parameters in EG following three-month intervention. Significant differences with baseline levels (*p < 0.05). LPO: lipoperoxides, PC: protein carbonylation, TOS: Total oxidant status, OSI: Oxidative stress index, SOD: superoxide dismutase, GPx: gluthatione peroxidase, CAT: catalase, TAS Total antioxidant, AOX GAP: antioxidant gap.
Figure 2.
Percentage changes [Δ% = (Post–Pre)/Pre*100] in parameters in EG following three-month intervention. Significant differences with baseline levels (*p < 0.05). LPO: lipoperoxides, PC: protein carbonylation, TOS: Total oxidant status, OSI: Oxidative stress index, SOD: superoxide dismutase, GPx: gluthatione peroxidase, CAT: catalase, TAS Total antioxidant, AOX GAP: antioxidant gap.
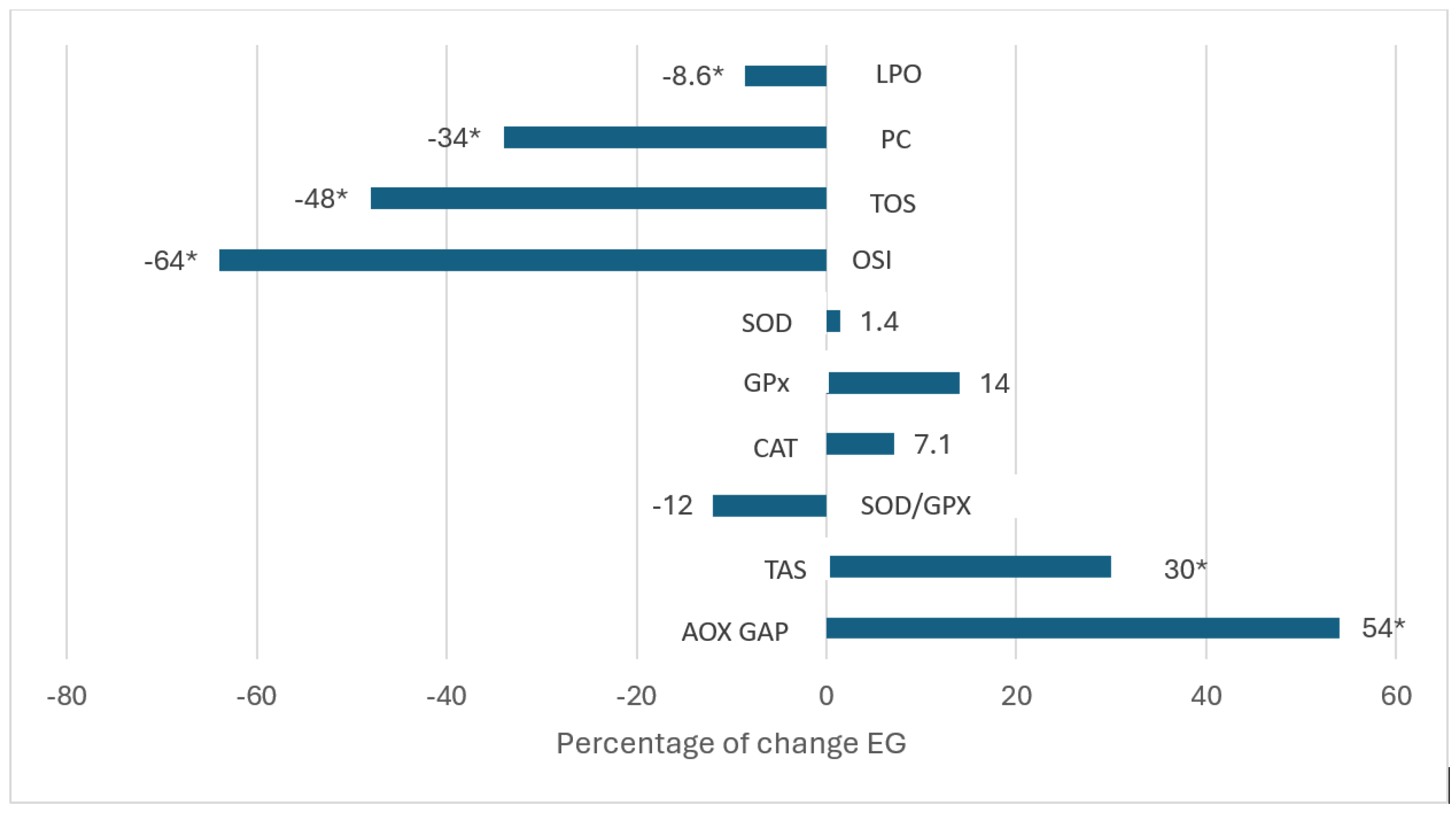
Figure 3.
Percentage changes [Δ% = (Post–Pre)/Pre*100] in parameters in PG following three month intervention. Significant differences with baseline levels (*p < 0.05). LPO: lipoperoxides, PC: protein carbonylation, TOS: Total oxidant status, OSI: Oxidative stress index, SOD: superoxide dismutase, GPx: gluthatione peroxidase, CAT: catalase, TAS Total antioxidant, AOX GAP: antioxidant gap.
Figure 3.
Percentage changes [Δ% = (Post–Pre)/Pre*100] in parameters in PG following three month intervention. Significant differences with baseline levels (*p < 0.05). LPO: lipoperoxides, PC: protein carbonylation, TOS: Total oxidant status, OSI: Oxidative stress index, SOD: superoxide dismutase, GPx: gluthatione peroxidase, CAT: catalase, TAS Total antioxidant, AOX GAP: antioxidant gap.

Table 1.
Biochemical parameters pre- and post-treatment.
|
Placebo Group (n=14) |
Experimental Group (n=19) |
|
| Glucose (mg/dL) Basal Three-months |
169 ± 79 181 ± 80 |
174 ± 76 160 ± 76 |
| Trigycerides (mg/dL) Basal Three-months |
144 ± 61 184 ± 124 |
148 ± 73 170 ± 94 |
| HDL-C (mg/dL) Basal Three-months |
58 ± 12 60 ± 15 |
55 ± 11 54 ± 14 |
| Urea (mg/dL) Basal Three-months |
39 ± 14 34 ± 15 |
36 ± 18 39 ± 17 |
| Uric acid (mg/dL) Basal Three-months |
3.1 ± 0.9 3.3 ± 1.6 |
3.5 ± 1.4 3.7 ± 1.5 |
| Cholesterol (mg/dL) Basal Three-month |
191 ± 46 210 ± 40 |
186 ± 31 197 ± 52 |
| Creatinine (mg/dL) Basal Three-months |
0.94 ± 0.32 0.90 ± 0.42 |
0.97 ± 0.33 0.90 ± 0.42 |
| Albumin (mg/dL) Basal Three-month |
4.0 ± 0.4 4.2 ± 0.5 |
4.0 ± 0.3 4.1 ± 0.4 |
| HbA1c (%) Basal Three-months |
8.1 ± 2.0 7.8 ± 1.7 |
8.9 ± 2.2 7.8 ± 2.0* |
Data are expressed as means ± standard deviation. ANOVA of repeated measures test, significance level 95%. *Baseline vs three months inter group p values p < 0.05 are shown. HbA1c: glycosylated hemoglobin.
Table 2.
Oxidative stress markers and pre- and post-treatment.
|
Placebo Group (n=14) |
Experimental Group (n=19) |
|
|
Lipoperoxides (µmol/L) Basal Three-months |
0.338 ± 0.138 0.373 ± 0.145 |
0.243 ± 0.067 0.222 ± 0.050* |
|
Protein carbonylation Basal Three-months |
29.5 ± 10 29.4± 15 |
29.4 ± 10 19.2 ± 6* |
|
TOS Basal Three-months |
5.3 ± 2 5.4± 3 |
6.0 ± 2.6 3.1 ± 1.8* |
|
OSI Basal Three-months |
6.2 ±3 5.5± 4 |
5.7 ±3.1 2.0 ± 1.1* |
|
SOD (U/L) Basal Three-months |
179 ± 8.6 167 ± 12.9 |
177± 12.8 179 ± 16.8* |
|
GPx (U/L) Basal Three-months |
7327 ± 2234 7473 ± 2192 |
5970 ± 2234 6802 ± 1942 |
|
Catalase activity (U/ml) Basal Three-months |
190 ± 70 280 ± 150 |
280± 150 300 ± 130 |
|
SOD/GPx Basal Three-months |
0.027 ± 0.008 0.024 ± 0.008 |
0.033 ± 0.01 0.029± 0.10 |
|
TAS (mmol/L) Basal Three-months |
1.06 ± 0.28 1.09 ± 0.20 |
0.94 ± 0.29 1.22 ± 0.28 * |
|
AOX GAP (µmol/L) Basal Three-months |
473 ± 270 473 ± 208 |
358 ± 283 551 ± 193 |
Data are expressed as means ± standard deviation. ANOVA of repeated measures test, significance level 95%. *Baseline vs three months inter group p values p < 0.05 are shown. TOS, Total oxidant status; OSI, Oxidative stress index, SOD: superoxide dismutase activity, GPx: glutathione peroxidase activity, TAS, Total antioxidant status; AOX GAP: antioxidant gap.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



