
Submitted:
17 March 2025
Posted:
17 March 2025
You are already at the latest version
Abstract
Undaria pinnatifida fucoidan (UPF), a bioactive sulphated polysaccharide, is widely recognised for its anti-inflammatory, antioxidant, antitumor, anticoagulant, antiviral, and immunomodulatory properties. However, the precise mechanisms by which UPF regulates inflammation and neuronal health remain unclear. This study aimed to investigate the effects of UPF supplementation on pro-inflammatory cytokines in skeletal muscle, small intestine, and the hypothalamus, as well as plasma cytokine levels. Additionally, a brain proteomic investigation in the nucleus accumbens (NAc) was performed to assess UPF’s impact on neuronal protein expression in mice. A total of 64 C57BL/6J mice were administered either a standard chow or high fat diet (HFD) with or without UPF (400 mg/kg/day) for 10 weeks. In HFD-fed mice, UPF significantly reduced the expression of pro-inflammatory cytokines (TNF-α, IL-1β, and IL-6) in skeletal muscle, small intestine, and hypothalamus, while also lowering circulating IL-1α and IL-6 levels. Proteomic analysis of the NAc revealed that UPF modulated proteins involved in oxidative stress, neuroinflammation, neurotransmitter regulation, and endoplasmic reticulum stress. In contrast, in chow-fed mice, UPF had no effect on the neuroinflammatory-oxidative stress markers but influenced the abundance of proteins associated with immune response and innate immunity. These findings highlight diet-dependent effects of UPF, suggesting an adaptive role in modulating stress-response pathways under different metabolic conditions and highlighting its potential neuroprotective support for inflammation-related disorders and brain health.
Keywords:
fucoidan
; Undaria pinnatifida
; high fat diet
; anti-inflammatory agents
; oxidative stress
; neuroprotection
; immunomodulation
; proteomic analysis
; TNF-α
; IL-1β
; IL-6
1. Introduction
Fucoidan is a sulphated polysaccharide found in brown seaweed, such as Undaria pinnatifida, that is widely recognised for its diverse bioactive properties, including anti-inflammatory, antitumor, antioxidant, anticoagulant, antiviral, and immunomodulatory activities [1,2,3,4,5,6,7,8,9,10,11,12].
Recent studies have demonstrated that Undaria pinnatifida fucoidan (UPF) can modulate inflammatory pathways by suppressing pro-inflammatory cytokine expression, reducing oxidative stress, and regulating immune cell infiltration in both in vitro and in vivo investigations [11,13,14,15]. However, the mechanisms by which UPF exerts its anti-inflammatory effects remain an area of active investigation.
Animal models, particularly rodents, serve as valuable tools for studying inflammation due to their ability to closely mimic human dietary patterns and metabolic responses [16]. Specifically, high-fat diet (HFD) consumption provides critical insights into the complex relationship between inflammation and the mechanisms driving obesity-related metabolic disorders. Rodents consuming HFD develop low-grade, chronic inflammation in adipose tissue, characterised by the infiltration of macrophages and other immune cells, and this condition is consistently linked to elevated systemic inflammation [17,18]. Several investigations have demonstrated that adipocyte hypertrophy, a common consequence of HFD feeding, triggers the release of pro-inflammatory cytokines such as TNF-α, IL-1 β and IL-6, and monocyte chemoattractant protein-1 (MCP-1) promoting chronic low-grade inflammation that affects multiple distant organs including muscle, intestine, and brain [19,20].
Little is known about the effects of UPF supplementation in conjunction with long-term HFD consumption, particularly its influence on inflammatory markers in high metabolic organs and tissues such as brain, muscles and the intestine. In our previous investigation we found that ongoing UPF supplementation enhanced exercise performance, improved muscle function, and positively modulated the gut microbiome in mice, regardless of dietary condition [21]. In vitro studies demonstrated that a 4-hour pre-treatment with UPF (10, 50, and 100 µg/ml) significantly suppressed lipopolysaccharide (LPS)-induced upregulation of pro-inflammatory cytokines, including tumour necrosis factor-alpha (TNF-α) and interleukins 1β and 6 (IL-1 β and IL-6), in human macrophages and peripheral blood mononuclear cells [11]. UPF (5-40 μg/ml) also protected hypothalamic neurons from endoplasmic reticulum (ER) stress-induced apoptosis through the Akt/mTOR pathway, highlighting its neuroprotective potential role [22]. In both in vitro and in vivo models of neurodegenerative diseases, UPF exhibited potent antioxidant and neuroprotective effects significantly reducing amyloid-beta (Aβ1-42) and amyloid-beta (Aβ25–35) aggregation and cytotoxicity in neuronal PC-12 cells while enhancing neurite outgrowth [3,23,24].
In humans, clinical studies further support the anti-inflammatory effects of UPF. One clinical trial showed that a single dose of UPF (1 g) modulated microRNA expression related to immune response and inflammation, highlighting its systemic regulatory potential [25]. In another double-blind randomised placebo-controlled clinical trial, 2 weeks of UPF administration (1 g/day) significantly suppressed the upregulation of inflammatory cytokines induced by high-intensity exercise [26]. Similarly, UPF combined with green-lipped mussel mitigated joint pain and prediabetes, demonstrating its antioxidant and anti-inflammatory effects in a clinical setting [27].
Building on UPF’s known bioactive properties, this study aimed to investigate its potential to counteract the negative effects of HFD consumption on peripheral metabolic organs and in the brain. After 10 weeks of UPF administration, plasma levels and gene expression of key pro-inflammatory cytokines including TNF-α, IL-1β, and IL-6 were measured in the skeletal muscle, small intestine, and the hypothalamus of mice consuming either standard chow or HFD. Additionally, the expression of Tjp1 (tight junction protein 1), a marker of intestinal barrier integrity, and the short-chain fatty acid receptors GPR41 and GPR43 to evaluate UPF’s role in gut homeostasis was examined. To further explore UPF’s anti-inflammatory mechanisms and potential neuroprotective activities, a proteomic analysis of the nucleus accumbens (NAc), a brain region involved in reward processing, stress regulation, and neuroinflammation was conducted [28,29]. Collectively, the findings of this study highlight the diet-dependent effects of UPF, demonstrating its ability to mitigate inflammation and modulate stress-related pathways in HFD-fed mice while exerting distinct immune-related effects in chow-fed mice. The observed changes in cytokine expression and proteomic profiles suggest UPF’s potential neuroprotective support for inflammation-related disorders and brain health, particularly under metabolic stress conditions.
2. Results
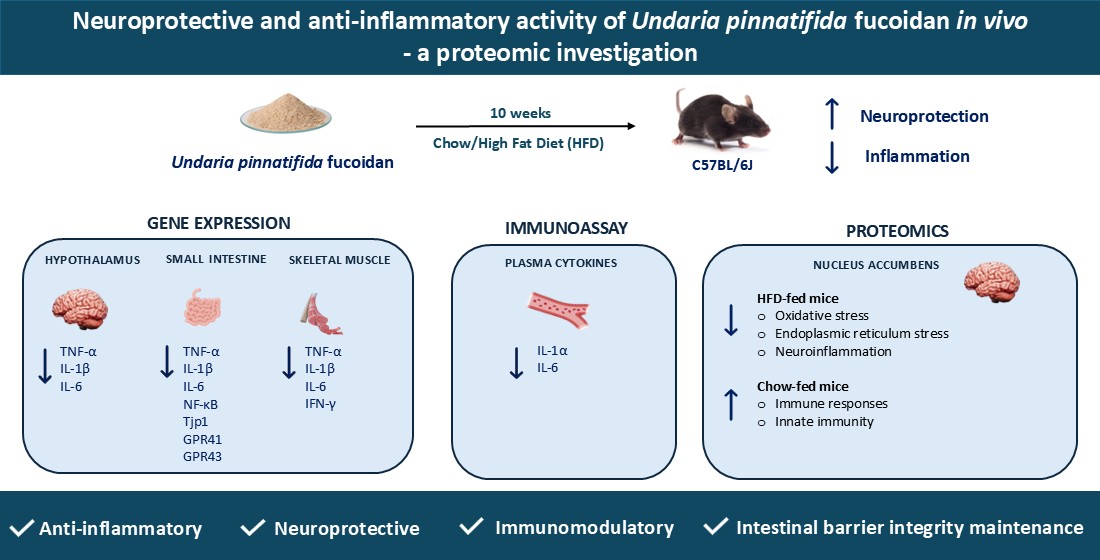
2.1. Effects of HFD and UPF on Skeletal Muscle Gene Expression
HFD consumption increased the muscular mRNA expression of TNF-α (Figure 1A), IL-1β (Figure 1B), and IL-6 (Figure 1C) compared to standard chow diet. However, in mice consuming HFD, UPF supplementation significantly downregulated the mRNA expression of TNF-α (-32.2%; p < 0.05, Figure 1A), IL-1β (-35.6%; p < 0.05, Figure 1B), and IL-6 (-28.9%; p < 0.05, Figure 1C).
No significant effects of UPF supplementation were observed in the mRNA expression of these cytokines in mice consuming a standard chow diet.
2.2. Effects of HFD and UPF on Small Intestine Gene Expression
Overall, HFD increased the intestinal mRNA expression of TNF-α (Figure 2A), IL-1β (Figure 2B), IL-6 (Figure 2C), nuclear factor kappa B (NF-κB, Figure 2D), Tjp1 (Figure 2E), and G-protein coupled receptors 41 and 43 (GPR41 and GPR43, Figure 2F, G) compared to the standard chow diet. However, in mice consuming HFD, UPF supplementation significantly downregulated the mRNA expression of TNF-α (-62.3%; p < 0.05, Figure 2A), IL-1β (-62.8%; p < 0.05, Figure 2B), IL-6 (-58.9%; p < 0.05, Figure 2C), NF-κB (-55.6%; p < 0.05, Figure 2D), Tjp1 (-56.5%; p < 0.05, Figure 2E), GPR41 (-52.8%; p < 0.05, Figure 2F), and GPR43 (-52.9%; p < 0.05, Figure 2G).
No significant effects of UPF supplementation were observed for these genes in mice consuming a standard chow diet.
2.3. Effects of HFD and UPF on Hypothalamic Gene Expression
HFD consumption significantly increased the hypothalamic mRNA expression of TNF-α (Figure 3A) and IL-1β (Figure 3B) compared to standard chow diet.
In HFD-fed mice, UPF supplementation significantly downregulated the mRNA expression of TNF-α (-58.8%; p < 0.05, Figure 3A) and IL-1β (-66.8%; p < 0.05, Figure 3B). Additionally, UPF administration significantly reduced the expression levels of IL-6 (-51.9%; p < 0.05, Figure 3C) and IFN-γ (-41.2%; p < 0.05, Figure 3D) in the hypothalamus. In contrast, UPF supplementation had no statistically significant effect on the mRNA expression of TNF-α (Figure 3A), IL-1β (Figure 3B), IL-6 (Figure 3C), or IFN-γ (Figure 3D) in chow-fed mice.
2.4. Effects of HFD and UPF on Pro-Inflammatory Plasma Cytokine Levels
HFD consumption elevated the plasma levels of IL-1α (Figure 4A) compared to standard chow diet. In HFD-fed mice, UPF supplementation significantly decreased IL-1α by 61.7% (p < 0.05, Figure 4A), with no statistically significant effect observed in the chow group. Additionally, UPF supplementation significantly downregulated the plasma levels of IL-6 (Figure 4B) by 73.9% (p < 0.05, Figure 4B) in mice consuming HFD, while no effect was observed in the chow group.
Although UPF supplementation led to a reduction in TNF-α levels (-45.2%; Figure 4C), this effect was not statistically significant (p = 0.06). No statistically significant changes were observed in mice consuming chow diet.
2.5. Effects of UPF on NAc Protein Abundance
A total of 5423 proteins were identified in the NAc of mice consuming HFD or standard chow diet. In the CHOW group, 23 proteins showed significant differences in expression between UPF-treated and control groups (FDR-adjusted p-value < 0.05, Figure 5A). In the HFD group, UPF significantly regulated the expression of 36 proteins compared to the control group (FDR-adjusted p-value < 0.05, Figure 5B). Volcano plots (Figure 5) highlight these differences, showing proteins with log2 fold changes (FC) greater than ± 0.57 and FDR-adjusted p-values below 0.05.
2.5.1. Expression Profiles of Differentially Expressed Proteins (DEPs) in the NAc
The differentially expressed proteins (DEPs) are detailed in Table 1 (CHOW+UPF vs CHOW) and Table 2 (HFD+UPF vs HFD). Among the 23 DEPs in the CHOW group (Table 1), 3 proteins were upregulated and 20 were downregulated in the NAc of UPF-treated mice. In the HFD group, 4 proteins showed upregulation and 32 downregulation in the NAc of UPF-treated mice (Table 2).
2.5.2. Molecular Functions and Biological Implications of DEPs in the NAc
To further investigate the molecular functions and biological implications of these DEPs, in silico analysis was performed using STRING, a protein-protein interaction (PPI) network tool. This analysis revealed the key biological processes associated with the 20 downregulated DEPs in the CHOW group, including stress response, viral defence, cytokine response, immune response, and bacterial defence (Figure 6).
Similarly, the 32 downregulated DEPs in the HFD group were associated with processes such as stress response, neurotransmitter regulation, neurogenesis, cell death regulation, and synaptic transmission, as shown in Figure 7.
Additionally, in the 3 upregulated DEPs in the NAc of mice consuming a standard chow diet, Cenpv and Nup210 were associated with cellular component biogenesis and organisation, with no PPI detected in Cops2. In contrast, among the 4 upregulated DEPs in mice consuming a HFD, Ppm1j and Mlip were involved in cellular metabolic process. However, no PPI was observed in Fhl2 and Yjefn3.
Furthermore, we carried out a search for biological processes and functions of upregulated and downregulated DEPs by using DAVID. In the CHOW group, the upregulated DEPs were only enriched in cellular component of nuclear membrane and nuclear envelope. In contrast, the downregulated DEPs were linked to type II interferon production, stress response, bacterial defence, and iron-sulphur cluster binding (Figure 8A). In the HFD group, the upregulated DEPs were mainly enriched in the biological process of regulation of transcription by RNA polymerase II, DNA-templated transcription, and RNA biosynthetic process, as well as the molecular function of transcription corepressor activity and coregulator activity, while no cellular component was enriched. However, the downregulated DEPs were principally associated with signaling, neurotransmitter transport regulation, oxidative stress, and synapse-related functions (Figure 8B).
3. Discussion
This study identified potential mechanisms by which UPF exerts its anti-inflammatory effects in vivo. Specifically, we investigated how the compound modulated plasma pro-inflammatory cytokine protein levels and alterations in gene expression across multiple tissues, such as skeletal muscle, the small intestine, and the hypothalamus. Additionally, we conducted a proteomic analysis of the nucleus accumbens to further investigate UPF brain-specific anti-inflammatory effects and potential neuroprotective activities.
Extensive research in animal models indicates that prolonged HFD consumption is typically accompanied by overproduction of pro-inflammatory cytokines including IL-1 and its superfamily members, which results in the activation of the NF-κB pathway, leading to IL-6 and TNF-α secretion [30]. This cascade promotes apoptosis, organ damage, and ultimately increases mortality [31,32,33,34,35]. In this study, and consistent with the literature, HFD mice exhibited increased pro-inflammatory cytokine expression (IL-1β, IL-6, TNF-α) in skeletal muscle (Figure 1), the small intestine (Figure 2), and the hypothalamus (Figure 3), while mice consuming standard chow showed no changes in inflammatory markers. Over the 10-week period, UPF supplementation (400 mg/kg/day), significantly reduced plasma levels of IL-1α and IL-6 (Figure 4) as well as mRNA expression of pro-inflammatory cytokines (IL-1β, IL-6, TNF-α) in skeletal muscle, small intestine, and the hypothalamus of mice consuming HFD (Figure 1, Figure 2, Figure 3).
Chronic inflammation is a major contributor to muscle wasting and reduced athletic function [36]. Previous investigations on UPF supplementation demonstrated that the compound enhanced exercise performance and improved muscle function in mice, regardless of the diet [21]. In addition, in a clinical trial acute treatment of a single dose of UPF (1 g) modulated the expression of serum microRNA and affected several biological pathways including inflammation [25]. Our results are also in line with previous research conducted on other species of low molecular weight fucoidan [37,38]. Specifically, a 3-week supplementation with Sargassum hemiphyllum-derived fucoidan (160 mg/kg/day) reduced NF-κB activation and decreased the expression of pro-inflammatory cytokine (TNF-α, IL-6, and IL-1β) [37], and similarly, a 4-week administration of fucoidan extracted from Laminaria japonica (20, 40, and 80 mg/kg/day) protected gastrocnemius muscles from inflammatory injury in diabetic rats [38].
In the small intestine, several investigations reported that fucoidan extracts showed anti-inflammatory effects by enhancing intestinal barrier function and improving the symptoms of ulcerative colitis while lowering pro-inflammatory cytokine levels [39,40,41,42,43]. In a mouse model of colitis using dextran sodium sulfate (DSS), a chemical colitogen with anticoagulant properties to induce disease, 28 days oral administration of fucoidan derived from Scytosiphon lomentaria (100 and 300 mg/kg/day) inhibited the NF-κB/MAPK pathways, reduced pro-inflammatory cytokine TNF-α and increased anti-inflammatory IL-10 production [42]. Similarly, in our experiment, suppression of NF-κB activity was accompanied by a reduced TNF-α, IL-1β, and IL-6 expression in mice consuming HFD. In mice consuming chow diet and treated with UPF, pro-inflammatory cytokine levels were normal.
Tight junction proteins play a crucial role in maintaining the integrity of the intestinal barrier. When dysregulated, they contribute to increased intestinal permeability and chronic inflammation [44,45]. In our study, we observed an upregulation of Tight Junction Protein 1 (Tjp1) in mice fed HFD. However, UPF supplementation significantly suppressed this dysregulation suggesting that fucoidan may restore tight junction protein levels to a baseline that supports optimal barrier function helping to maintain intestinal integrity [39,42,46]. Additionally, our study found that UPF treatment inhibited HFD-induced upregulation of GPR41 and GPR43 in the small intestine. These receptors interact with short-chain fatty acids (SCFAs) such as acetate, propionate, and butyrate - microbial metabolites with a dual role in gut health [47,48]. While SCFA-mediated activation of GPR41 and GPR43 can enhance gut barrier integrity and immune regulation, excessive activation under inflammatory conditions may worsen cytokine production and immune cell recruitment [47,48,49,50,51]. This suggests that UPF-induced downregulation of these receptors could serve as a protective mechanism, preventing hyperactivation and reducing inflammation.
In the hypothalamus of HFD-fed mice, UPF significantly reduced the mRNA expression of pro-inflammatory cytokines, including TNF-α, IL-1, IL-6, and IFN-γ. The hypothalamus is a key regulator of energy balance and is particularly vulnerable to inflammation induced by HFD [52,53]. Hypothalamic inflammation has been associated with disruption of neuronal function and development of systemic metabolic dysfunction [54,55,56]. By suppressing the expression of hypothalamic pro-inflammatory cytokines, UPF may enhance neuronal resilience, further suggesting potential benefits in mitigating diet-induced metabolic disturbances [57,58]. However, further studies are needed to elucidate the mechanisms through which UPF exerts its systemic anti-inflammatory effects, particularly in plasma, skeletal muscle, the small intestine, and the hypothalamus.
To further investigate the anti-inflammatory role of UPF in the brain, we conducted a proteomic analysis of the NAc, a brain region involved in motivation, reward, and addiction [59]. Overall, our findings revealed that UPF modulated the expression of proteins related to immune response, neuronal stability, oxidative and endoplasmic reticulum stress, neurotransmitter regulation, and neuroinflammation, with effects varying based on diet. Specifically, we identified 23 differentially expressed proteins (DEPs) between the CHOW+UPF and CHOW groups, and 36 DEPs between the HFD+UPF and HFD groups. For clarity, we structured the discussion of the proteomic results based on the effects of UPF in relation to the specific diet consumed.
Proteomics: effects of UPF in mice consuming standard chow
In mice consuming standard chow diet, UPF reduced the abundance of 20 proteins predominantly associated with biological processes involved in immune responses, cellular stress response in the NAc. In relation to immune regulation, the downregulation of two key interferon-stimulated proteins, ubiquitin-like protein interferon-stimulated gene 15 (Isg15) and interferon-induced protein with tetratricopeptide repeats 3 (Ifit3), were observed in the NAc of UPF-treated mice consuming a standard chow diet. Isg15 is a member of the ubiquitin family, while best known for its antiviral and immunomodulatory roles, it can be induced in central nervous system cells (neurons, astrocytes, microglia) in response to viral infection or immune activation [60,61,62]. The expression of Isg15 is associated with type I interferons (IFN-α and IFN-β) signalling and other inflammatory cues (IFN-γ and IL-10) suggesting a possible role for UPF in immune regulation in the brain [60,63] and anti-inflammatory response in cardiac myocytes [64]. In the same way, Ifit3 is also typically induced by type I interferons, acting in concert with other Ifit family members to strengthen antiviral defences and shape inflammatory responses [65]. The reduction in abundance of Isg15 and Ifit3 in mice consuming chow diet and UPF aligns with the known anti-inflammatory and antiviral properties of fucoidan extracts [66,67]. In addition, UPF reduced the abundance of tyrosine-protein kinase (Abl1), a protein involved in synaptic signalling and neuronal plasticity [68,69]. The gene Abl1 encodes a non-receptor tyrosine kinase, a type of enzyme that adds phosphate groups to proteins, influencing their functional function [70]. Previous studies also reported that overactivation of Abl1 has been linked to neurodegenerative diseases, such as Parkinson, Alzheimer, and Tauopathies, by promoting harmful processes like α-synuclein buildup and tau phosphorylation [69,71]. The observed inhibitory effects of UPF on Abl1 suggests that the compound might influence neuronal excitability and synaptic remodelling, potentially promoting neuroprotection by reducing excitotoxic stress.
The suppression of von Willebrand factor (Vwf) and unconventional myosin-Ic (Myo1c) protein highlights the potential role of UPF in maintaining cerebrovascular stability and regulating cytoskeletal dynamics, which is significant in endothelial health and mitigation of vascular inflammation [72,73,74]. Vwf, a large glycoprotein primarily produced by endothelial cells and megakaryocytes, is well-recognised for its essential function in haemostasis [73]. However, emerging evidence suggests that Vwf also actively contributes to leukocyte extravasation and vascular inflammation, extending its influence beyond coagulation [72]. This pro-inflammatory potential of Vwf is attributed to its capacity to activate endothelial cells, augment platelet-leukocyte interactions, enhance vascular permeability, and stabilise neutrophil extracellular traps, collectively facilitating sustained immune cell recruitment and endothelial disruption [72,73]. Notably, Vwf is abundantly expressed in cerebral endothelial cells, with elevated levels correlating with an increased risk of CNS pathologies such as hypoxia, epilepsy, and ischemic stroke [75,76,77]. In a synergistic role with Vwf, Myo1c, a non-muscle myosin influences endothelial cell shape and stability [74,78] reducing unnecessary Vwf release [74]. Therefore, the reduced abundance of Vwf and Myo1c suggests the potential role of UPF in mitigating vascular inflammation and preserving neuronal function.
Conversely, COP9 signalosome complex subunit 2 (Cops2) was the most overexpressed protein in mice consuming chow diet and treated with UPF. This protein is a key component of constitutive photomorphogenesis 9 signalosomes (COPS), a regulatory complex involved in protein degradation, DNA damage repair, and cellular signalling [79]. Evidence indicated that Cops2 downregulated Octamer-binding transcription factor (Oct-3/4) mRNA expression in the rat brain, intensifying neuronal differentiation and development, alongside with maintaining neuronal functions [80]. Other studies have also suggested that Cops2 helps protect cells from stress by stabilizing important proteins, supporting neuronal health, or regulating normal cellular processes [80,81,82]. The increased expression of Cops2 suggests that UPF may support cellular repair mechanisms potentially mitigating the risk of neurodegeneration associated with DNA damage. However, the mechanistic role of Cops2 remains insufficiently understood, highlighting the need for further studies to clarify its function in neuroprotection and its broader impact on cellular processes.
Nuclear pore membrane glycoprotein 210 (Nup210) was the second most overexpressed protein in this study. As a key nuclear pore protein, Nup210 plays a crucial role in maintaining nuclear-cytoplasmic transport and regulating gene expression [83]. Upregulation of Nup210 may indicate enhanced nuclear integrity and transport efficiency, supporting cellular resilience and transcriptional regulation. Its upregulation may indicate enhanced nuclear integrity and transport efficiency, contributing to cellular resilience and transcriptional regulation. This finding suggests that UPF may support neuronal function by promoting efficient nucleocytoplasmic communication, a process that is often impaired during neuroinflammation and aging [84].
Overall, in mice consuming a chow diet, UPF modulated proteins associated with neuroinflammation, synaptic regulation, and cellular stress responses, highlighting its diverse neuroprotective potential. These findings suggest that UPF may contribute to maintaining brain homeostasis and could play a role in reducing the risk of inflammation-driven neurodegeneration.
Proteomics: effects of UPF in mice consuming HFD
In mice consuming HFD, UPF treatment downregulated 32 proteins linked to oxidative stress responses and apoptosis, neurotransmitter regulation, neurogenesis, synaptic transmission, and behaviour. Among those proteins involved in oxidative stress responses, UPF reduced the abundance of leucine-rich repeat serine/threonine-protein kinase 2 (Lrrk2), wolframin (Wfs1), and neuroglobin (Ngb).
Lrrk2, a multifunctional protein kinase, is expressed in immune cells like microglia and macrophages, where it regulates inflammatory responses, including the production of pro-inflammatory cytokines [85]. Lrrk2 kinase activates the MAPK signalling and increases the production of reactive oxygen species (ROS) and pro-inflammatory cytokines, such as TNF-α, to promote neuroinflammation and apoptosis [85,86,87,88]. Other studies also reported that mutations in Lrrk2 are linked to oxidative stress conditions and neurodegeneration in PD [85,86]. Wfs1 is a transmembrane protein, primarily localised to the endoplasmic reticulum (ER) [89]. Its dysregulation induces ER stress, activating inflammation and apoptotic pathways, and has been linked to β-cell dysfunction, resulting in impaired insulin production and secretion [90]. Additionally, Wfs1 malfunctioning has been associated with neurodegenerative disorders [89,90,91,92]. Our findings showed reduction in Ngb protein levels, an oxygen-binding globin protein, which has been associated with protection against oxidative stress and neuroprotection by scavenging ROS and supporting oxygen homeostasis [93]. Ngb expression is upregulated under several oxidative stress conditions, such as chronic inflammation, neurodegenerative diseases, and neurotoxin exposure, with its overexpression shown to reduce oxidative damage and improve mitochondrial function in neurons [94,95,96,97].
Mice fed with HFD alone had significantly lower Ngb levels compared to those receiving UPF alongside HFD. This reduction in Ngb under HFD-induced stress might be attributed to the antioxidant property of UPF to alleviate oxidative stress responses [98,99]. These findings suggest that UPF might restore neuronal homeostasis and offers potential as a therapeutic strategy for HFD-induced neuroinflammatory and neurodegenerative conditions. Further research is needed to elucidate the precise mechanisms and clinical relevance.
Interestingly, a reduction of proteins such as synaptic vesicle glycoprotein 2C (Sv2c), sodium-dependent dopamine transporter (Slc6a3), synaptic vesicular amine transporter (Slc18a2), and proenkephalin-A (Penk) were also observed in HFD-fed mice with UPF administration. These proteins are critical for neurotransmitter storage, release, and receptor interaction [100,101,102,103]. Their dysregulation is often associated with synaptic hyperactivity and neurotransmitter imbalance, which affects cognitive, emotional, and physical functioning [100,101,102,103]. These conditions are usually implicated in a variety of neurological and psychiatric diseases, such as epilepsy, PD, attention-deficit/hyperactivity disorder, schizophrenia, and anxiety [101,102,104,105]. Our results align with a previous study in depressive-like mouse models where 2 weeks of oral administration of fucoidan extracts (25, 50, and 100 mg/kg/day) suggests potential in preventing inflammation-mediated depression [106]. This study reported that fucoidan exerted antidepressant-like effects by reducing caspase-1-induced inflammation, increasing phosphorylation of Ser845 of GluA1, and promoting the stability of surface AMPARs in the hippocampus [106]. Collectively, these findings suggest a potential regulatory role of UPF in mitigating synaptic dysfunction caused by HFD-induced stress. However, the action mechanism of UPF in Sv2c, Slc6a3, Slc18a2, and Penk is still not well-documented, highlighting the need for targeted studies to investigate the pathways and molecular interactions involved in these regulatory processes.
In this study, we found an increase in protein abundance of four and a half LIM domains protein 2 (Fhl2) in the NAc of UPF-treated mice consuming HFD, which suggests significant regulatory effects of UPF on cellular stress responses and inflammatory pathways. Fhl2, a multifunctional protein, is associated with cellular signalling, cytoskeletal organization, and inflammatory responses [107,108,109,110]. For example, Fhl2 plays a significant anti-inflammatory role across various tissues by modulating key pathways like NF-κB and p38 MAPK, balancing immune responses, and promoting tissue repair [109,111,112]. Fhl2 also acts as a scaffolding protein by interacting with various structural and signalling molecules, facilitating cellular processes such as gene transcription, cytoskeletal organisation, and signal transduction [107,113]. In addition, Fhl2 has been reported to modulate neurogenesis and brain function, because its deficiency in mice leads to delayed neuroblast migration and premature astrocyte differentiation [110]. These findings suggest that the ability of UPF to increase Fhl2 protein abundance may underlie its neuroprotective and anti-inflammatory properties, highlighting UPF’s potential to modulate cellular and molecular pathways involved in stress adaptation, neurogenesis, and immune regulation.
Similarly, UPF also increased the levels of protein phosphatase 1J (Ppm1j), a member of the metal-dependent Ppm phosphatase family, which possesses key cellular functions such as cell metabolism, oxidative stress response, and immune signalling [114]. Dysregulation of these phosphatases can lead to abnormal stress responses and metabolic disorders [114]. Although there is a lack of comprehensive research specifically on Ppm1j, previous studies have shown that several members of the Ppm family, such as Ppm1D, Ppm1F, and Ppm1G, regulate synaptic plasticity, maintain neuronal morphology, regulate cellular stress response, and promote neuroprotection in the brain neurons [114,115,116,117]. These findings highlight the potential significance of UPF-induced Ppm1j upregulation in neuronal resilience and stress response; however, further studies are needed to elucidate the specific mechanisms and roles of Ppm1j in brain neurons.
In this study, our results aim to provide a descriptive mechanism by which 10 weeks of UPF administration reduced systemic inflammation and promoted neuronal health in mice. Specifically, UPF significantly downregulated pro-inflammatory cytokines (TNF-α, IL-1β, and IL-6) in the skeletal muscle, small intestine, and the hypothalamus, while also reducing circulating IL-1α and IL-6 levels in mice fed HFD. Furthermore, our findings highlight the context-dependent effects of UPF on stress-related biological processes in the brain, as evidenced by the modulation of distinct protein sets in the chow and HFD groups. While no single protein was differentially expressed across both dietary conditions, UPF consistently influenced proteins involved in immune response, neuronal stability, oxidative stress response pathways and neuroinflammation. In HFD-fed mice, UPF influenced the abundance of stress-response proteins, including Lrrk2, Wfs1, Ngb, Slc6a3, and Penk, which are implicated in oxidative stress, neuroinflammation, and metabolic dysfunction. These findings suggest that UPF exerts adaptive effects, modulating stress-response pathways in a context-dependent manner based on metabolic and inflammatory conditions. In contrast, in mice consuming standard chow, UPF had no effect on metabolic markers, but influenced the abundance of proteins such as Isg15, Ifit3, Abl1, Vwf, and Myo1c that are associated with inflammatory, immune response as well as innate immunity. Collectively, our findings suggest that UPF supplementation can mitigate systemic inflammation, and enhance neuronal function, highlighting its potential neuroprotective support for inflammation-related disorders and brain health.
Future studies should aim to delineate the specific signalling pathways involved in UPF-mediated cytokine modulation, with a particular focus on identifying any direct interactions between UPF and cytokine gene promoters or other elements of the transcriptional machinery. Additionally, extending these investigations to models of chronic inflammation, neurodegeneration, and autoimmune diseases could elucidate the therapeutic scope of UPF in clinical settings.
4. Materials and Methods
4.1. Ethics Statement
This investigation was authorised by the Animal Ethics Committee of the University of Tasmania (A0027164). Animal work and procedures in this study were executed in strict adherence to the provisions delineated in the Tasmanian Animal Welfare Act (1993/63) and the Australian Code of Practice for the Care and Use of Animals for Scientific Purposes 8 edition 2013 [118].
4.2. Animals and Diet
Four-week-old male and female C57BL/6J mice (n=32 males and n=32 females, Animal Services, University of Tasmania) were housed under controlled conditions at a temperature of 20 ± 2°C and maintained on a standard 12:12h light/dark cycle. Following a one-week acclimatization period, 32 mice were provided with ad libitum access to a commercial high-fat pelleted diet (HFD) (19.4 MJ/kg, 23.5% fat, 23% protein, 5.4% crude fibre, SF16-059, Specialty Feeds, Perth, Australia), while the control group (32 mice) received a standard chow diet (12.8 MJ/kg, 6% fat, 20% protein, 3.2% crude fibre, product code 102108, Ridly Agri-Products) for ten weeks. During this time, mice were singularly housed with unrestricted access to food and drinking water ad libitum.
4.3. Fucoidan Administration and Experimental Groups
A fucoidan extract (≥85% fucoidan) derived from Undaria pinnatifida (UPF, Batch No.: UPF2022532) was provided by Marinova Pty Ltd. (Tasmania, Australia) and stored at ambient temperature (21-29°C). The chemical composition of fucoidan used in this study is described in Supplementary Information. Within each dietary cohort, half of the mice received UPF (400 mg/kg/day) in a form incorporated into artificially flavoured and sweetened jelly for voluntary oral administration, while the control group received vehicle (non-UPF) jelly. Prior to commencing the study, mice were trained to consume the jelly using a previously reported method to ensure they were accustomed to its taste and texture [119]. The successful use of this method has already been described elsewhere [21].This method yielded four experimental groups (n=16 per group): CHOW, CHOW+UPF, HFD, and HFD+UPF. Dietary and UPF intervention continued for ten weeks.
4.4. Sample Collection
Mice were fasted overnight (12 h) and euthanised via carbon dioxide inhalation for plasma and tissue collection. Blood samples were collected in a 1.5 mL Eppendorf tube containing 7.5 µL of Heparin (1000 U/mL) by cardiac puncture and kept on ice. Plasma supernatants were collected by centrifugation (12,000 rpm/10 min, R.T.). Plasma was aliquoted and stored at −80°C until use. Plasma levels of inflammatory cytokines IL-1α, IL-6, and TNF- α were measure with a commercial MILLIPLEX® mouse high sensitivity T cell magnetic bead panel (MHSTCMAG-70K, Millipore, Australia) according to the manufacturer’s instructions. Mice small intestines, skeletal muscles (gastrocnemius), hypothalamus, and NAc were dissected, snap-frozen in liquid nitrogen, and stored at −80°C for RNA extraction in gene expression studies and for protein extraction in proteomic assays.
4.5. Real-Time Quantitative PCR (RT-qPCR) Assay
Total mRNA was extracted from mice small intestine, skeletal muscle, and hypothalamus tissues using RNeasy Mini Kit (Cat# 74106, Qiagen, Chuo-ku, Japan) according to the manufacturer’s recommendations. A NanodropTM 8000 Spectrophotometer (NanoDrop Technologies Inc., Minato-ku, Japan) was used to measure the RNA concentration and purity ratios (A260/280 and A260/230) and only samples with absorbance ratio of ~2.0 were used for complementary DNA (cDNA) synthesis. One microgram of RNA template from each sample was reverse transcribed into cDNA using High-Capacity cDNA Reverse Transcription Kit (Cat# 4368814, Applied Biosystem, Waltham, MA, USA). All primers were purchased from Kicqstart SYBR Green Primers (Sigma-Aldrich, St. Louis, MO, USA). The sequence of the primers used for quantification of gene expression of TNF-α, IL-1β, IL-6, IL-10, NF-κB, Tjp1, GPR41, GPR43, TLR4, β-actin, and GAPDH can be found in Supplementary Information.
RT-qPCR reactions were performed using PowerUp SYBR green master mix (Cat# A25918, Applied Biosystems, Waltham, MA, USA) in the CFX Connect TM Real-Time PCR Detection system (Bio-Rad) following the manufacturer’s instructions. The amplification program included an initial denaturation step at 95 °C for 5 min, followed by 40 cycles of 10 s at 95 °C, 10 s at 60 °C, and 20 s at 72 °C. After each amplification, a melting curve study was conducted to validate the reliability of the results and product specificity. Target gene expression was normalised against β-actin and GAPDH housekeeping genes using the average of control group as a calibrator. Analysis was performed using the comparative 2-Δ Δ Ct method [120].
4.6. Proteomic Analysis
Frozen NAc tissues were homogenised using a tissue protein extraction reagent (TPER, Thermo Fisher Scientific, USA) to efficiently lyse cells and solubilise proteins. Protein concentrations in the extracts were quantified using a Nanodrop Spectrophotometer (NanoDrop Technologies Inc., Minato-ku, Japan) at an absorbance at A660nm, to ensure uniform protein (30 μg/90 μL) quantity for downstream analysis.
Proteins were reduced with 10 mM dithiothreitol (DTT) overnight at 4°C, alkylated with 55 mM iodoacetamide (IAA) for 2 hrs in the dark at room temperature then digested with 1.2 μg trypsin/rLysC (Promega) after sample clean-up according to the SP3 method [121]. Peptides were then desalted using C18 ZipTips (Millipore, USA) and analysed by nano-flow HPLC and data-independent acquisition mass spectrometry using a Q-Exactive HF and Ultimate 3000 RSLCnano, as previously described [122].
4.7. Proteomics Data Processing and Statistical Analysis
Raw DIA-MS files were imported into Spectronaut software version 18 (Biognosys) and searched against the Mus musculus UniProt proteome UP00000589 comprising 44,456 entries (accessed 06/04/2021) using the Pulsar search engine according to Biognosys (BGS) factory settings. The resulting library of 37,622 proteotypic peptides, mapping to 6,298 protein groups, was then used for targeted data re-extraction and cross-run protein quantitation/normalisation using BGS factory settings, with the exception that single-hit protein IDs were excluded. The Perseus software platform (version 2.0.11) was used for imputation of missing intensity values, statistical analysis, and data visualization.
Differentially abundant proteins were identified using pair-wise t-test comparisons of all replicates (n = 10) of both CHOW and HFD (CHOW vs. CHOW+UPF and HFD vs. HFD+UPF), with a false-discovery rate (FDR) of 0.05 and an s0 value of 0.1 used to define significant proteins. Functional analyses of the differentially abundant proteins were performed using DAVID (Database for Annotation Visualization and Integrated Discovery) version 2024 (https://david.ncifcrf.gov/ accessed on 12 September 2024) [123,124] to analyse biological processes, molecular functions, and cellular components. STRING: Functional protein association network (https://string-db.org/ accessed on 12 September 2024) [125] was used to develop interaction maps.
Other statistical analyses were performed using GraphPad Prism version 8.3.0 for Windows (GraphPad Software, San Diego, California USA, www.graphpad.com, accessed on 12 September 2024) and results were expressed as mean ± SEM. The effects of UPF on mRNA gene expression in mice muscle, intestine, and hypothalamus, and plasma inflammatory cytokine levels were analysed by One-way ANOVA. ANOVA results were then assessed by a post hoc analysis using Fisher’s least significant difference test (LSD) as appropriate. When no overall diet effect was observed between groups [(CHOW and CHOW+UPF) vs (HFD and HFD+UPF)], data were analysed by Unpaired t test. Results were considered statistically significant when p < 0.05.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1: Absolute mass percentages of Undaria pinnatifida fucoidan (UPF) extract; Table S2: Carbohydrate breakdown (mass %) of neutral carbohydrates in Undaria pinnatifida fucoidan (UPF) extract; Table S3: Sequence of primers used for bacterial profiling by quantitative PCR; Table S4: Sequence of primers used for RT-qPCR assay in mouse skeletal muscle, small intestine, and hypothalamus.
Author Contributions
Conceptualization, C.Y. and V.C.; methodology, C.Y. and V.C.; software, C.Y., M.R., R.W., and V.C.; validation, C.Y., C.D., B.W., M.R., R.W., and V.C.; formal analysis, C.Y., C.D., B.W., M.R., R.W., L.J. and V.C.; investigation, C.Y., C.D., B.W. and V.C.; resources, C.D., B.W. and V.C.; data curation, C.Y., C.D., B.W., M.R., R.W., and V.C.; writing original draft preparation, C.Y. and V.C.; writing review and editing, C.Y., C.D., B.W., M.R., R.W., L.J. and V.C.; visualization, C.Y. and V.C.; supervision, R.W, L.J. and V.C.; project administration, C.D., B.W. and V.C.; funding acquisition, V.C., C.D. and B.W. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Marinova Pty. Ltd.
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki, and approved by the Institutional Review Board (or Ethics Committee) of Washington Biotechnology IACUC (Approval number A0027164 (A-94788) on 19 April 2023).
Data Availability
The mass spectrometry proteomics data have been deposited to the ProteomeXchange Consortium via the PRIDE partner repository [126] with the dataset identifier PXD06086 [accessible via login reviewer_pxd060861@ebi.ac.uk; password 7Mwe5q83hJPo].
Acknowledgments
This study was funded by the Marine Bioproducts CRC – Grant. We would like to thank Mr. Harmanjeet Harmanjeet and Ms. Devika Gaire for providing technical support during the study, and the Animal Service Team of UTAS for assisting with animal care and monitoring.
Conflicts of Interest
The current study was sponsored by Marinova Pty. Ltd. (Cambridge, TAS, Australia) C.D. and B.W. are employees of Marinova Pty. Ltd.
References
- Zhao, Y.; Zheng, Y.; Wang, J.; Ma, S.; Yu, Y.; White, W.L.; Yang, S.; Yang, F.; Lu, J. Fucoidan Extracted from Undaria pinnatifida: Source for Nutraceuticals/Functional Foods. Marine Drugs 2018, 16. [Google Scholar] [CrossRef]
- Apostolova, E.; Lukova, P.; Baldzhieva, A.; Katsarov, P.; Nikolova, M.; Iliev, I.; Peychev, L.; Trica, B.; Oancea, F.; Delattre, C.; et al. Immunomodulatory and Anti-Inflammatory Effects of Fucoidan: A Review. Polymers 2020, 12, 2338. [Google Scholar] [CrossRef]
- Wei, H.; Gao, Z.; Zheng, L.; Zhang, C.; Liu, Z.; Yang, Y.; Teng, H.; Hou, L.; Yin, Y.; Zou, X. Protective Effects of Fucoidan on Aβ25-35 and d-Gal-Induced Neurotoxicity in PC12 Cells and d-Gal-Induced Cognitive Dysfunction in Mice. Marine Drugs 2017, 15. [Google Scholar] [CrossRef]
- Fitton, H.J.; Stringer, D.S.; Park, A.Y.; Karpiniec, S.N. Therapies from Fucoidan: New Developments. Mar Drugs 2019, 17. [Google Scholar] [CrossRef]
- Fitton, J.H.; Stringer, D.N.; Karpiniec, S.S. Therapies from Fucoidan: An Update. Marine Drugs 2015, 13, 5920–5946. [Google Scholar] [CrossRef]
- Fitton, J.H.; Dell'Acqua, G.; Gardiner, V.-A.; Karpiniec, S.S.; Stringer, D.N.; Davis, E. Topical Benefits of Two Fucoidan-Rich Extracts from Marine Macroalgae. Cosmetics 2015, 2, 66–81. [Google Scholar] [CrossRef]
- Park, A.Y.; Bourtembourg, M.; Chrétien, A.; Hubaux, R.; Lancelot, C.; Salmon, M.; Fitton, J.H. Modulation of Gene Expression in a Sterile Atopic Dermatitis Model and Inhibition of Staphylococcus aureus Adhesion by Fucoidan. Dermatopathology (Basel) 2021, 8, 69–83. [Google Scholar] [CrossRef]
- Myers, S.P.; O'Connor, J.; Fitton, J.H.; Brooks, L.; Rolfe, M.; Connellan, P.; Wohlmuth, H.; Cheras, P.A.; Morris, C. A Combined Phase I and II Open Label Study on the Effects of A Seaweed Extract Nutrient Complex on Osteoarthritis. Biologics 2010, 4, 33–44. [Google Scholar] [CrossRef]
- Carvalho, A.C.S.; Sousa, R.B.; Franco, Á.X.; Costa, J.V.G.; Neves, L.M.; Ribeiro, R.A.; Sutton, R.; Criddle, D.N.; Soares, P.M.G.; de Souza, M.H.L.P. Protective Effects of Fucoidan, a P- and L-Selectin Inhibitor, in Murine Acute Pancreatitis. Pancreas 2014, 43, 82–87. [Google Scholar] [CrossRef]
- Lim, J.D.; Lee, S.R.; Kim, T.; Jang, S.A.; Kang, S.C.; Koo, H.J.; Sohn, E.; Bak, J.P.; Namkoong, S.; Kim, H.K.; et al. Fucoidan from Fucus vesiculosus Protects Against Alcohol-Induced Liver Damage by Modulating Inflammatory Mediators in Mice and HepG2 Cells. Marine Drugs 2015, 13, 1051–1067. [Google Scholar] [CrossRef]
- Ahmad, T.; Eapen, M.S.; Ishaq, M.; Park, A.Y.; Karpiniec, S.S.; Stringer, D.N.; Sohal, S.S.; Fitton, J.H.; Guven, N.; Caruso, V.; et al. Anti-Inflammatory Activity of Fucoidan Extracts In Vitro. Marine Drugs 2021, 19, 702. [Google Scholar] [CrossRef] [PubMed]
- Cooper, R.; Dragar, C.; Elliot, K.; Fitton, J.H.; Godwin, J.; Thompson, K. GFS, A Preparation of Tasmanian Undaria pinnatifida Is Associated with Healing and Inhibition of Reactivation of Herpes. BMC Complement Altern Med 2002, 2, 11. [Google Scholar] [CrossRef]
- Phull, A.-R.; Majid, M.; Haq, I.-u.; Khan, M.R.; Kim, S.J. In Vitro and In Vivo Evaluation of Anti-arthritic, Antioxidant Efficacy of Fucoidan from Undaria pinnatifida (Harvey) Suringar. International Journal of Biological Macromolecules 2017, 97, 468–480. [Google Scholar] [CrossRef] [PubMed]
- Herath, K.H.I.N.M.; Kim, H.J.; Kim, A.; Sook, C.E.; Lee, B.-Y.; Jee, Y. The Role of Fucoidans Isolated from the Sporophylls of Undaria pinnatifida against Particulate-Matter-Induced Allergic Airway Inflammation: Evidence of the Attenuation of Oxidative Stress and Inflammatory Responses. Molecules 2020, 25, 2869. [Google Scholar] [CrossRef] [PubMed]
- Shanmugasundaram, D.; Dwan, C.; Wimmer, B.C.; Srivastava, S. Fucoidan Ameliorates Testosterone-Induced Benign Prostatic Hyperplasia (BPH) in Rats. Res Rep Urol 2024, 16, 283–297. [Google Scholar] [CrossRef]
- Duan, Y.; Zeng, L.; Zheng, C.; Song, B.; Li, F.; Kong, X.; Xu, K. Inflammatory Links Between High Fat Diets and Diseases. Front Immunol 2018, 9, 2649. [Google Scholar] [CrossRef]
- Bae, H.R.; Shin, S.-K.; Yoo, J.-H.; Kim, S.; Young, H.A.; Kwon, E.-Y. Chronic inflammation in high-fat diet-fed mice: Unveiling the early pathogenic connection between liver and adipose tissue. Journal of Autoimmunity 2023, 139, 103091. [Google Scholar] [CrossRef]
- Wu, Y.; Wu, T.; Wu, J.; Zhao, L.; Li, Q.; Varghese, Z.; Moorhead, J.F.; Powis, S.H.; Chen, Y.; Ruan, X.Z. Chronic inflammation exacerbates glucose metabolism disorders in C57BL/6J mice fed with high-fat diet. J Endocrinol 2013, 219, 195–204. [Google Scholar] [CrossRef]
- Hotamisligil, G.S. Foundations of Immunometabolism and Implications for Metabolic Health and Disease. Immunity 2017, 47, 406–420. [Google Scholar] [CrossRef]
- Heydemann, A. An Overview of Murine High Fat Diet as a Model for Type 2 Diabetes Mellitus. J Diabetes Res 2016, 2016, 2902351. [Google Scholar] [CrossRef]
- Yang, C.; Dwan, C.; Wimmer, B.C.; Wilson, R.; Johnson, L.; Caruso, V. Fucoidan from Undaria pinnatifida Enhances Exercise Performance and Increases the Abundance of Beneficial Gut Bacteria in Mice. Marine Drugs 2024, 22, 485. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Moon, I.S.; Goo, T.W.; Moon, S.S.; Seo, M. Algae Undaria pinnatifida Protects Hypothalamic Neurons against Endoplasmic Reticulum Stress through Akt/mTOR Signaling. Molecules 2015, 20, 20998–21009. [Google Scholar] [CrossRef] [PubMed]
- Azzini, E.; Peña-Corona, S.I.; Hernández-Parra, H.; Chandran, D.; Saleena, L.A.K.; Sawikr, Y.; Peluso, I.; Dhumal, S.; Kumar, M.; Leyva-Gómez, G.; et al. Neuroprotective and anti-inflammatory effects of curcumin in Alzheimer's disease: Targeting neuroinflammation strategies. Phytotherapy Research 2024, 38, 3169–3189. [Google Scholar] [CrossRef]
- Alghazwi, M.; Smid, S.; Karpiniec, S.; Zhang, W. Comparative study on neuroprotective activities of fucoidans from Fucus vesiculosus and Undaria pinnatifida. Int J Biol Macromol 2019, 122, 255–264. [Google Scholar] [CrossRef]
- Gueven, N.; Spring, K.J.; Holmes, S.; Ahuja, K.; Eri, R.; Park, A.Y.; Fitton, J.H. Micro RNA Expression after Ingestion of Fucoidan; A Clinical Study. Mar Drugs 2020, 18. [Google Scholar] [CrossRef]
- McFadden, B.A.; Vincenty, C.S.; Chandler, A.J.; Cintineo, H.P.; Lints, B.S.; Mastrofini, G.F.; Arent, S.M. Effects of fucoidan supplementation on inflammatory and immune response after high-intensity exercise. Journal of the International Society of Sports Nutrition 2023, 20, 2224751. [Google Scholar] [CrossRef]
- Tay, A.; Jiang, Y.; Signal, N.; O'Brien, D.; Chen, J.; Murphy, R.; Lu, J. Combining mussel with fucoidan as a supplement for joint pain and prediabetes: Study protocol for a randomized, double-blinded, placebo-controlled trial. Frontiers in Nutrition 2022, 9. [Google Scholar] [CrossRef]
- Xu, Y.; Lin, Y.; Yu, M.; Zhou, K. The nucleus accumbens in reward and aversion processing: Insights and implications. Frontiers in Behavioral Neuroscience 2024, 18. [Google Scholar] [CrossRef]
- Dudek, K.A.; Dion-Albert, L.; Lebel, M.; LeClair, K.; Labrecque, S.; Tuck, E.; Ferrer Perez, C.; Golden, S.A.; Tamminga, C.; Turecki, G.; et al. Molecular adaptations of the blood-brain barrier promote stress resilience vs. depression. Proc Natl Acad Sci U S A 2020, 117, 3326–3336. [Google Scholar] [CrossRef]
- Barbier, L.; Ferhat, M.; Salamé, E.; Robin, A.; Herbelin, A.; Gombert, J.-M.; Silvain, C.; Barbarin, A. Interleukin-1 Family Cytokines: Keystones in Liver Inflammatory Diseases. Frontiers in Immunology 2019, 10. [Google Scholar] [CrossRef]
- Bleau, C.; Karelis, A.D.; St-Pierre, D.H.; Lamontagne, L. Crosstalk between intestinal microbiota, adipose tissue and skeletal muscle as an early event in systemic low-grade inflammation and the development of obesity and diabetes. Diabetes/Metabolism Research and Reviews 2015, 31, 545–561. [Google Scholar] [CrossRef] [PubMed]
- Andersen, K.; Kesper, M.S.; Marschner, J.A.; Konrad, L.; Ryu, M.; Kumar VR, S.; Kulkarni, O.P.; Mulay, S.R.; Romoli, S.; Demleitner, J.; et al. Intestinal Dysbiosis, Barrier Dysfunction, and Bacterial Translocation Account for CKD–Related Systemic Inflammation. Journal of the American Society of Nephrology 2017, 28, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Cole, C.L.; Kleckner, I.R.; Jatoi, A.; Schwarz, E.M.; Dunne, R.F. The Role of Systemic Inflammation in Cancer-Associated Muscle Wasting and Rationale for Exercise as a Therapeutic Intervention. JCSM Clin Rep 2018, 3. [Google Scholar] [CrossRef]
- Burfeind, K.G.; Michaelis, K.A.; Marks, D.L. The central role of hypothalamic inflammation in the acute illness response and cachexia. Seminars in Cell & Developmental Biology 2016, 54, 42–52. [Google Scholar] [CrossRef]
- Perry, V.H. The influence of systemic inflammation on inflammation in the brain: Implications for chronic neurodegenerative disease. Brain, Behavior, and Immunity 2004, 18, 407–413. [Google Scholar] [CrossRef]
- Tuttle, C.S.L.; Thang, L.A.N.; Maier, A.B. Markers of inflammation and their association with muscle strength and mass: A systematic review and meta-analysis. Ageing Research Reviews 2020, 64, 101185. [Google Scholar] [CrossRef]
- Chen, M.C.; Hsu, W.L.; Hwang, P.A.; Chen, Y.L.; Chou, T.C. Combined administration of fucoidan ameliorates tumor and chemotherapy-induced skeletal muscle atrophy in bladder cancer-bearing mice. Oncotarget 2016, 7, 51608–51618. [Google Scholar] [CrossRef]
- Wang, Z.; Liu, T.; Chen, X.; You, H.; Zhang, Q.; Xue, J.; Zheng, Y.; Luo, D. Low molecular weight fucoidan ameliorates hindlimb ischemic injury in type 2 diabetic rats. Journal of Ethnopharmacology 2018, 210, 434–442. [Google Scholar] [CrossRef]
- Iraha, A.; Chinen, H.; Hokama, A.; Yonashiro, T.; Kinjo, T.; Kishimoto, K.; Nakamoto, M.; Hirata, T.; Kinjo, N.; Higa, F.; et al. Fucoidan enhances intestinal barrier function by upregulating the expression of claudin-1. World J Gastroenterol 2013, 19, 5500–5507. [Google Scholar] [CrossRef]
- Lean, Q.Y.; Eri, R.D.; Fitton, J.H.; Patel, R.P.; Gueven, N. Fucoidan Extracts Ameliorate Acute Colitis. PLoS ONE 2015, 10, e0128453. [Google Scholar] [CrossRef]
- Choi, J.-i.; Raghavendran, H.R.B.; Sung, N.-Y.; Kim, J.-H.; Chun, B.S.; Ahn, D.H.; Choi, H.-S.; Kang, K.-W.; Lee, J.-W. Effect of fucoidan on aspirin-induced stomach ulceration in rats. Chemico-Biological Interactions 2010, 183, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.; Wang, L.; Ren, X.; Song, S.; Ai, C. Fucoidan Alleviates Colitis and Metabolic Disorder by Protecting the Intestinal Barrier, Suppressing the MAPK/NF-κB Pathways, and Regulating the Gut Microbiota. Journal of Food Biochemistry 2024, 2024, 7955190. [Google Scholar] [CrossRef]
- Selim, H.M.; Negm, W.A.; Hawwal, M.F.; Hussein, I.A.; Elekhnawy, E.; Ulber, R.; Zayed, A. Fucoidan mitigates gastric ulcer injury through managing inflammation, oxidative stress, and NLRP3-mediated pyroptosis. International Immunopharmacology 2023, 120, 110335. [Google Scholar] [CrossRef] [PubMed]
- Zong, X.; Cao, X.; Wang, H.; Xiao, X.; Wang, Y.; Lu, Z. Cathelicidin-WA Facilitated Intestinal Fatty Acid Absorption Through Enhancing PPAR-γ Dependent Barrier Function. Frontiers in Immunology 2019, 10. [Google Scholar] [CrossRef]
- Landy, J.; Ronde, E.; English, N.; Clark, S.K.; Hart, A.L.; Knight, S.C.; Ciclitira, P.J.; Al-Hassi, H.O. Tight junctions in inflammatory bowel diseases and inflammatory bowel disease associated colorectal cancer. World J Gastroenterol 2016, 22, 3117–3126. [Google Scholar] [CrossRef]
- Sun, T.; Liang, H.; Xue, M.; Liu, Y.; Gong, A.; Jiang, Y.; Qin, Y.; Yang, J.; Meng, D. Protective effect and mechanism of fucoidan on intestinal mucosal barrier function in NOD mice. Food and Agricultural Immunology 2020, 31, 939–953. [Google Scholar] [CrossRef]
- Kim, M.H.; Kang, S.G.; Park, J.H.; Yanagisawa, M.; Kim, C.H. Short-Chain Fatty Acids Activate GPR41 and GPR43 on Intestinal Epithelial Cells to Promote Inflammatory Responses in Mice. Gastroenterology 2013, 145, 396–406.e310. [Google Scholar] [CrossRef]
- Ang, Z.; Ding, J.L. GPR41 and GPR43 in Obesity and Inflammation - Protective or Causative? Front Immunol 2016, 7, 28. [Google Scholar] [CrossRef]
- Maslowski, K.M.; Vieira, A.T.; Ng, A.; Kranich, J.; Sierro, F.; Di, Y.; Schilter, H.C.; Rolph, M.S.; Mackay, F.; Artis, D.; et al. Regulation of inflammatory responses by gut microbiota and chemoattractant receptor GPR43. Nature 2009, 461, 1282–1286. [Google Scholar] [CrossRef]
- Chang, G.; Zhang, H.; Wang, Y.; Ma, N.; Chandra, R.A.; Ye, G.; Zhuang, S.; Zhu, W.; Shen, X. Microbial community shifts elicit inflammation in the caecal mucosa via the GPR41/43 signalling pathway during subacute ruminal acidosis. BMC Veterinary Research 2019, 15, 298. [Google Scholar] [CrossRef]
- Wang, J.J.; Zhang, Q.M.; Ni, W.W.; Zhang, X.; Li, Y.; Li, A.L.; Du, P.; Li, C.; Yu, S.S. Modulatory effect of Lactobacillus acidophilus KLDS 1.0738 on intestinal short-chain fatty acids metabolism and GPR41/43 expression in β-lactoglobulin-sensitized mice. Microbiol Immunol 2019, 63, 303–315. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Suk, K.; Yu, R.; Kim, M.S. Cellular Contributors to Hypothalamic Inflammation in Obesity. Mol Cells 2020, 43, 431–437. [Google Scholar] [CrossRef] [PubMed]
- Cavaliere, G.; Viggiano, E.; Trinchese, G.; De Filippo, C.; Messina, A.; Monda, V.; Valenzano, A.; Cincione, R.I.; Zammit, C.; Cimmino, F.; et al. Long Feeding High-Fat Diet Induces Hypothalamic Oxidative Stress and Inflammation, and Prolonged Hypothalamic AMPK Activation in Rat Animal Model. Front Physiol 2018, 9, 818. [Google Scholar] [CrossRef]
- Ishaq, M.; Tran, D.; Yang, C.; Ng, M.J.; Kackanattil, A.; Tata, K.; Deans, B.J.; Bleasel, M.; Vicenzi, S.; Randall, C.; et al. The Anti-Obesity Compound Asperuloside Reduces Inflammation in the Liver and Hypothalamus of High-Fat-Fed Mice. Endocrines 2022, 3, 641–653. [Google Scholar] [CrossRef]
- Jais, A.; Brüning, J.C. Hypothalamic inflammation in obesity and metabolic disease. J Clin Invest 2017, 127, 24–32. [Google Scholar] [CrossRef]
- Le Thuc, O.; Stobbe, K.; Cansell, C.; Nahon, J.-L.; Blondeau, N.; Rovère, C. Hypothalamic Inflammation and Energy Balance Disruptions: Spotlight on Chemokines. Frontiers in Endocrinology 2017, 8. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, K.; Ding, X.; Wang, Y.; Bai, H.; Yang, Q.; Ben, J.; Zhang, H.; Li, X.; Chen, Q.; et al. Fucoidan antagonizes diet-induced obesity and inflammation in mice. J Biomed Res 2020, 35, 197–205. [Google Scholar] [CrossRef]
- Huang, J.; Huang, J.; Li, Y.; Lv, H.; Yin, T.; Fan, S.; Zhang, C.; Li, H. Fucoidan Protects Against High-Fat Diet-Induced Obesity and Modulates Gut Microbiota in Institute of Cancer Research Mice. J Med Food 2021, 24, 1058–1067. [Google Scholar] [CrossRef]
- Ambroggi, F.; Ghazizadeh, A.; Nicola, S.M.; Fields, H.L. Roles of Nucleus Accumbens Core and Shell in Incentive-Cue Responding and Behavioral Inhibition. The Journal of Neuroscience 2011, 31, 6820–6830. [Google Scholar] [CrossRef]
- Perng, Y.-C.; Lenschow, D.J. ISG15 in antiviral immunity and beyond. Nature Reviews Microbiology 2018, 16, 423–439. [Google Scholar] [CrossRef]
- Przanowski, P.; Loska, S.; Cysewski, D.; Dabrowski, M.; Kaminska, B. ISG'ylation increases stability of numerous proteins including Stat1, which prevents premature termination of immune response in LPS-stimulated microglia. Neurochemistry International 2018, 112, 227–233. [Google Scholar] [CrossRef] [PubMed]
- Hwang, M.; Bergmann, C.C. Alpha/Beta Interferon (IFN-α/β) Signaling in Astrocytes Mediates Protection against Viral Encephalomyelitis and Regulates IFN-γ-Dependent Responses. J Virol 2018, 92. [Google Scholar] [CrossRef] [PubMed]
- Swaim, C.D.; Scott, A.F.; Canadeo, L.A.; Huibregtse, J.M. Extracellular ISG15 Signals Cytokine Secretion through the LFA-1 Integrin Receptor. Mol Cell 2017, 68, 581–590.e585. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Zhang, Q.; Liu, X.; Shang, X. Downregulation of interferon-induced protein with tetratricopeptide repeats 3 relieves the inflammatory response and myocardial fibrosis of mice with myocardial infarction and improves their cardiac function. Exp Anim 2021, 70, 522–531. [Google Scholar] [CrossRef]
- Mears, H.V.; Sweeney, T.R. Better together: The role of IFIT protein–protein interactions in the antiviral response. Journal of General Virology 2018, 99, 1463–1477. [Google Scholar] [CrossRef]
- Park, H.Y.; Han, M.H.; Park, C.; Jin, C.-Y.; Kim, G.-Y.; Choi, I.-W.; Kim, N.D.; Nam, T.-J.; Kwon, T.K.; Choi, Y.H. Anti-inflammatory effects of fucoidan through inhibition of NF-κB, MAPK and Akt activation in lipopolysaccharide-induced BV2 microglia cells. Food and Chemical Toxicology 2011, 49, 1745–1752. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Q.; Han, X.; Ma, Y.; Zhang, Z.; Zhao, L.; Guan, F.; Ma, S. Fucoidan: A promising agent for brain injury and neurodegenerative disease intervention. Food & Function 2021, 12, 3820–3830. [Google Scholar] [CrossRef]
- Gutiérrez, D.A.; Chandía-Cristi, A.; Yáñez, M.J.; Zanlungo, S.; Álvarez, A.R. c-Abl kinase at the crossroads of healthy synaptic remodeling and synaptic dysfunction in neurodegenerative diseases. Neural Regen Res 2023, 18, 237–243. [Google Scholar] [CrossRef]
- Hebron, M.L.; Lonskaya, I.; Olopade, P.; Selby, S.T.; Pagan, F.; Moussa, C.E. Tyrosine Kinase Inhibition Regulates Early Systemic Immune Changes and Modulates the Neuroimmune Response in α-Synucleinopathy. J Clin Cell Immunol 2014, 5, 259. [Google Scholar] [CrossRef]
- Wang, J.Y. The capable ABL: What is its biological function? Mol Cell Biol 2014, 34, 1188–1197. [Google Scholar] [CrossRef]
- Brahmachari, S.; Ge, P.; Lee, S.H.; Kim, D.; Karuppagounder, S.S.; Kumar, M.; Mao, X.; Shin, J.H.; Lee, Y.; Pletnikova, O.; et al. Activation of tyrosine kinase c-Abl contributes to α-synuclein–induced neurodegeneration. The Journal of Clinical Investigation 2016, 126, 2970–2988. [Google Scholar] [CrossRef] [PubMed]
- Petri, B.; Broermann, A.; Li, H.; Khandoga, A.G.; Zarbock, A.; Krombach, F.; Goerge, T.; Schneider, S.W.; Jones, C.; Nieswandt, B.; et al. von Willebrand factor promotes leukocyte extravasation. Blood 2010, 116, 4712–4719. [Google Scholar] [CrossRef] [PubMed]
- Gragnano, F.; Sperlongano, S.; Golia, E.; Natale, F.; Bianchi, R.; Crisci, M.; Fimiani, F.; Pariggiano, I.; Diana, V.; Carbone, A.; et al. The Role of von Willebrand Factor in Vascular Inflammation: From Pathogenesis to Targeted Therapy. Mediators Inflamm 2017, 2017, 5620314. [Google Scholar] [CrossRef]
- El-Mansi, S.; Mitchell, T.P.; Mobayen, G.; McKinnon, T.A.J.; Miklavc, P.; Frick, M.; Nightingale, T.D. Myosin-1C augments endothelial secretion of von Willebrand factor by linking contractile actomyosin machinery to the plasma membrane. Blood Advances 2024, 8, 4714–4726. [Google Scholar] [CrossRef]
- Suidan, G.L.; Brill, A.; De Meyer, S.F.; Voorhees, J.R.; Cifuni, S.M.; Cabral, J.E.; Wagner, D.D. Endothelial Von Willebrand factor promotes blood-brain barrier flexibility and provides protection from hypoxia and seizures in mice. Arterioscler Thromb Vasc Biol 2013, 33, 2112–2120. [Google Scholar] [CrossRef]
- Bongers, T.N.; de Maat, M.P.M.; van Goor, M.-L.P.J.; Bhagwanbali, V.; van Vliet, H.H.D.M.; Gómez García, E.B.; Dippel, D.W.J.; Leebeek, F.W.G. High von Willebrand Factor Levels Increase the Risk of First Ischemic Stroke. Stroke 2006, 37, 2672–2677. [Google Scholar] [CrossRef]
- Zhu, X.; Cao, Y.; Wei, L.; Cai, P.; Xu, H.; Luo, H.; Bai, X.; Lu, L.; Liu, J.-R.; Fan, W.; et al. von Willebrand factor contributes to poor outcome in a mouse model of intracerebral haemorrhage. Scientific Reports 2016, 6, 35901. [Google Scholar] [CrossRef]
- Maravillas-Montero, J.L.; Gillespie, P.G.; Patiño-López, G.; Shaw, S.; Santos-Argumedo, L. Myosin 1c Participates in B Cell Cytoskeleton Rearrangements, Is Recruited to the Immunologic Synapse, and Contributes to Antigen Presentation. The Journal of Immunology 2011, 187, 3053–3063. [Google Scholar] [CrossRef]
- Liu, J.; Han, D.; Xuan, J.; Xie, J.; Wang, W.; Zhou, Q.; Chen, K. COP9 signalosome complex is a prognostic biomarker and corresponds with immune infiltration in hepatocellular carcinoma. Aging (Albany NY) 2024, 16, 5264–5287. [Google Scholar] [CrossRef]
- Akiyama, H.; Sugiyama, A.; Uzawa, K.; Fujisawa, N.; Tashiro, Y.; Tashiro, F. Implication of Trip15/CSN2 in early stage of neuronal differentiation of P19 embryonal carcinoma cells. Developmental Brain Research 2003, 140, 45–56. [Google Scholar] [CrossRef]
- Zhang, W.; Ni, P.; Mou, C.; Zhang, Y.; Guo, H.; Zhao, T.; Loh, Y.-H.; Chen, L. Cops2 promotes pluripotency maintenance by Stabilizing Nanog Protein and Repressing Transcription. Scientific Reports 2016, 6, 26804. [Google Scholar] [CrossRef]
- Lykke-Andersen, K.; Schaefer, L.; Menon, S.; Deng, X.W.; Miller, J.B.; Wei, N. Disruption of the COP9 signalosome Csn2 subunit in mice causes deficient cell proliferation, accumulation of p53 and cyclin E, and early embryonic death. Mol Cell Biol 2003, 23, 6790–6797. [Google Scholar] [CrossRef] [PubMed]
- D'Angelo, M.A.; Gomez-Cavazos, J.S.; Mei, A.; Lackner, D.H.; Hetzer, M.W. A change in nuclear pore complex composition regulates cell differentiation. Dev Cell 2012, 22, 446–458. [Google Scholar] [CrossRef] [PubMed]
- Ding, B.; Sepehrimanesh, M. Nucleocytoplasmic Transport: Regulatory Mechanisms and the Implications in Neurodegeneration. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef]
- Moehle, M.S.; Webber, P.J.; Tse, T.; Sukar, N.; Standaert, D.G.; DeSilva, T.M.; Cowell, R.M.; West, A.B. LRRK2 Inhibition Attenuates Microglial Inflammatory Responses. The Journal of Neuroscience 2012, 32, 1602–1611. [Google Scholar] [CrossRef]
- Mendivil-Perez, M.; Velez-Pardo, C.; Jimenez-Del-Rio, M. Neuroprotective Effect of the LRRK2 Kinase Inhibitor PF-06447475 in Human Nerve-Like Differentiated Cells Exposed to Oxidative Stress Stimuli: Implications for Parkinson’s Disease. Neurochemical Research 2016, 41, 2675–2692. [Google Scholar] [CrossRef]
- Kim, J.; Pajarillo, E.; Rizor, A.; Son, D.S.; Lee, J.; Aschner, M.; Lee, E. LRRK2 kinase plays a critical role in manganese-induced inflammation and apoptosis in microglia. PLoS ONE 2019, 14, e0210248. [Google Scholar] [CrossRef]
- Wallings, R.L.; Tansey, M.G. LRRK2 regulation of immune-pathways and inflammatory disease. Biochem Soc Trans 2019, 47, 1581–1595. [Google Scholar] [CrossRef]
- Takeda, K.; Inoue, H.; Tanizawa, Y.; Matsuzaki, Y.; Oba, J.; Watanabe, Y.; Shinoda, K.; Oka, Y. WFS1 (Wolfram syndrome 1) gene product: Predominant subcellular localization to endoplasmic reticulum in cultured cells and neuronal expression in rat brain. Human Molecular Genetics 2001, 10, 477–484. [Google Scholar] [CrossRef]
- Fonseca, S.G.; Fukuma, M.; Lipson, K.L.; Nguyen, L.X.; Allen, J.R.; Oka, Y.; Urano, F. WFS1 Is a Novel Component of the Unfolded Protein Response and Maintains Homeostasis of the Endoplasmic Reticulum in Pancreatic β-Cells*. Journal of Biological Chemistry 2005, 280, 39609–39615. [Google Scholar] [CrossRef]
- Rigoli, L.; Bramanti, P.; Di Bella, C.; De Luca, F. Genetic and clinical aspects of Wolfram syndrome 1, a severe neurodegenerative disease. Pediatric Research 2018, 83, 921–929. [Google Scholar] [CrossRef] [PubMed]
- Hotamisligil, G.S. Endoplasmic Reticulum Stress and the Inflammatory Basis of Metabolic Disease. Cell 2010, 140, 900–917. [Google Scholar] [CrossRef] [PubMed]
- Li, R.C.; Morris, M.W.; Lee, S.K.; Pouranfar, F.; Wang, Y.; Gozal, D. Neuroglobin protects PC12 cells against oxidative stress. Brain Research 2008, 1190, 159–166. [Google Scholar] [CrossRef]
- Fiocchetti, M.; Fernandez, V.S.; Montalesi, E.; Marino, M. Neuroglobin: A Novel Player in the Oxidative Stress Response of Cancer Cells. Oxid Med Cell Longev 2019, 2019, 6315034. [Google Scholar] [CrossRef]
- Watanabe, S.; Takahashi, N.; Uchida, H.; Wakasugi, K. Human Neuroglobin Functions as an Oxidative Stress-responsive Sensor for Neuroprotection*. Journal of Biological Chemistry 2012, 287, 30128–30138. [Google Scholar] [CrossRef]
- Wei, X.; Yu, Z.; Cho, K.-S.; Chen, H.; Malik, M.T.A.; Chen, X.; Lo, E.H.; Wang, X.; Chen, D.F. Neuroglobin Is an Endogenous Neuroprotectant for Retinal Ganglion Cells against Glaucomatous Damage. The American Journal of Pathology 2011, 179, 2788–2797. [Google Scholar] [CrossRef]
- Liu, J.; Yu, Z.; Guo, S.; Lee, S.R.; Xing, C.; Zhang, C.; Gao, Y.; Nicholls, D.G.; Lo, E.H.; Wang, X. Effects of neuroglobin overexpression on mitochondrial function and oxidative stress following hypoxia/reoxygenation in cultured neurons. J Neurosci Res 2009, 87, 164–170. [Google Scholar] [CrossRef]
- Alrafiah, A. Thymoquinone Protects Neurons in the Cerebellum of Rats through Mitigating Oxidative Stress and Inflammation Following High-Fat Diet Supplementation. Biomolecules 2021, 11. [Google Scholar] [CrossRef]
- Koh, H.S.A.; Lu, J.; Zhou, W. Structure characterization and antioxidant activity of fucoidan isolated from Undaria pinnatifida grown in New Zealand. Carbohydrate Polymers 2019, 212, 178–185. [Google Scholar] [CrossRef]
- Tokudome, K.; Okumura, T.; Shimizu, S.; Mashimo, T.; Takizawa, A.; Serikawa, T.; Terada, R.; Ishihara, S.; Kunisawa, N.; Sasa, M.; et al. Synaptic vesicle glycoprotein 2A (SV2A) regulates kindling epileptogenesis via GABAergic neurotransmission. Scientific Reports 2016, 6, 27420. [Google Scholar] [CrossRef]
- Kristensen, A.S.; Andersen, J.; Jørgensen, T.N.; Sørensen, L.; Eriksen, J.; Loland, C.J.; Strømgaard, K.; Gether, U.; Simonsen, U. SLC6 Neurotransmitter Transporters: Structure, Function, and Regulation. Pharmacological Reviews 2011, 63, 585–640. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Zhao, Y.; Chung, C.Y.; Zhou, Y.; Xiong, N.; Glatt, C.E.; Isacson, O. High regulatability favors genetic selection in SLC18A2, a vesicular monoamine transporter essential for life. Faseb j 2010, 24, 2191–2200. [Google Scholar] [CrossRef] [PubMed]
- Nikoshkov, A.; Drakenberg, K.; Wang, X.; Horvath, M.C.; Keller, E.; Hurd, Y.L. Opioid neuropeptide genotypes in relation to heroin abuse: Dopamine tone contributes to reversed mesolimbic proenkephalin expression. Proc Natl Acad Sci U S A 2008, 105, 786–791. [Google Scholar] [CrossRef]
- Lynch, B.A.; Lambeng, N.; Nocka, K.; Kensel-Hammes, P.; Bajjalieh, S.M.; Matagne, A.; Fuks, B. The synaptic vesicle protein SV2A is the binding site for the antiepileptic drug levetiracetam. Proceedings of the National Academy of Sciences 2004, 101, 9861–9866. [Google Scholar] [CrossRef]
- Kennedy, J.L.; Xiong, N.; Yu, J.; Zai, C.C.; Pouget, J.G.; Li, J.; Liu, K.; Qing, H.; Wang, T.; Martin, E.; et al. Increased Nigral SLC6A3 Activity in Schizophrenia Patients: Findings From the Toronto-McLean Cohorts. Schizophr Bull 2016, 42, 772–781. [Google Scholar] [CrossRef]
- Li, M.; Sun, X.; Li, Q.; Li, Y.; Luo, C.; Huang, H.; Chen, J.; Gong, C.; Li, Y.; Zheng, Y.; et al. Fucoidan exerts antidepressant-like effects in mice via regulating the stability of surface AMPARs. Biochemical and Biophysical Research Communications 2020, 521, 318–325. [Google Scholar] [CrossRef]
- Johannessen, M.; Møller, S.; Hansen, T.; Moens, U.; Van Ghelue, M. The multifunctional roles of the four-and-a-half-LIM only protein FHL2. Cell Mol Life Sci 2006, 63, 268–284. [Google Scholar] [CrossRef]
- Wixler, V. The role of FHL2 in wound healing and inflammation. The FASEB Journal 2019, 33, 7799–7809. [Google Scholar] [CrossRef]
- Wixler, V.; Cromme, C.; Retser, E.; Meyer, L.-H.; Smyth, N.; Mühlenberg, K.; Korb-Pap, A.; Koers-Wunrau, C.; Sotsios, Y.; Bassel-Duby, R.; et al. FHL2 regulates the resolution of tissue damage in chronic inflammatory arthritis. Annals of the Rheumatic Diseases 2015, 74, 2216–2223. [Google Scholar] [CrossRef]
- Kim, S.Y.; Völkl, S.; Ludwig, S.; Schneider, H.; Wixler, V.; Park, J. Deficiency of Fhl2 leads to delayed neuronal cell migration and premature astrocyte differentiation. Journal of Cell Science 2019, 132. [Google Scholar] [CrossRef]
- van de Pol, V.; Vos, M.; DeRuiter, M.C.; Goumans, M.J.; de Vries, C.J.M.; Kurakula, K. LIM-only protein FHL2 attenuates inflammation in vascular smooth muscle cells through inhibition of the NFκB pathway. Vascular Pharmacology 2020, 125–126, 106634. [Google Scholar] [CrossRef]
- Alnajar, A.; Nordhoff, C.; Schied, T.; Chiquet-Ehrismann, R.; Loser, K.; Vogl, T.; Ludwig, S.; Wixler, V. The LIM-only protein FHL2 attenuates lung inflammation during bleomycin-induced fibrosis. PLoS ONE 2013, 8, e81356. [Google Scholar] [CrossRef]
- Verset, L.; Feys, L.; Trépant, A.L.; De Wever, O.; Demetter, P. FHL2: A scaffold protein of carcinogenesis, tumour-stroma interactions and treatment response. Histol Histopathol 2016, 31, 469–478. [Google Scholar] [CrossRef] [PubMed]
- Kamada, R.; Kudoh, F.; Ito, S.; Tani, I.; Janairo, J.I.B.; Omichinski, J.G.; Sakaguchi, K. Metal-dependent Ser/Thr protein phosphatase PPM family: Evolution, structures, diseases and inhibitors. Pharmacology & Therapeutics 2020, 215, 107622. [Google Scholar] [CrossRef]
- Liu, J.; Meng, F.; Wang, W.; Cui, M.; Wu, M.; Jiang, S.; Dai, J.; Lian, H.; Li, Q.; Xu, Z.; et al. PPM1F in hippocampal dentate gyrus regulates the depression-related behaviors by modulating neuronal excitability. Experimental Neurology 2021, 340, 113657. [Google Scholar] [CrossRef]
- Ozaki, H.; Katoh, T.; Nakagawa, R.; Ishihara, Y.; Sueyoshi, N.; Kameshita, I.; Taniguchi, T.; Hirano, T.; Yamazaki, T.; Ishida, A. Ca2+/calmodulin-dependent protein kinase phosphatase (CaMKP/PPM1F) interacts with neurofilament L and inhibits its filament association. Biochemical and Biophysical Research Communications 2016, 477, 820–825. [Google Scholar] [CrossRef]
- Reitman, Z.J. Smaller Protein, Larger Therapeutic Potential: PPM1D as a New Therapeutic Target in Brainstem Glioma. Pharmacogenomics 2014, 15, 1639–1641. [Google Scholar] [CrossRef]
- Health, N.; Council, M.R. Australian Code of Practice for the Care and Use of Animals for Scientific Purposes; Australian Government Publishing Service: 1997.
- Zhang, L. Method for Voluntary Oral Administration of Drugs in Mice. STAR Protoc 2021, 2, 100330. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A New Mathematical Model for Relative Quantification in Real-Time RT-PCR. Nucleic Acids Res 2001, 29, e45. [Google Scholar] [CrossRef]
- Hughes, C.S.; Moggridge, S.; Müller, T.; Sorensen, P.H.; Morin, G.B.; Krijgsveld, J. Single-pot, solid-phase-enhanced sample preparation for proteomics experiments. Nature Protocols 2019, 14, 68–85. [Google Scholar] [CrossRef]
- Chear, S.; Perry, S.; Wilson, R.; Bindoff, A.; Talbot, J.; Ware, T.L.; Grubman, A.; Vickers, J.C.; Pébay, A.; Ruddle, J.B.; et al. Lysosomal alterations and decreased electrophysiological activity in CLN3 disease patient-derived cortical neurons. Dis Model Mech 2022, 15. [Google Scholar] [CrossRef]
- Huang da, W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat Protoc 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Sherman, B.T.; Hao, M.; Qiu, J.; Jiao, X.; Baseler, M.W.; Lane, H.C.; Imamichi, T.; Chang, W. DAVID: A web server for functional enrichment analysis and functional annotation of gene lists (2021 update). Nucleic Acids Research 2022, 50, W216–W221. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Morris, J.H.; Cook, H.; Kuhn, M.; Wyder, S.; Simonovic, M.; Santos, A.; Doncheva, N.T.; Roth, A.; Bork, P.; et al. The STRING database in 2017: Quality-controlled protein-protein association networks, made broadly accessible. Nucleic Acids Res 2017, 45, D362–D368. [Google Scholar] [CrossRef]
- Perez-Riverol, Y.; Bandla, C.; Kundu, D.J.; Kamatchinathan, S.; Bai, J.; Hewapathirana, S.; John, N.S.; Prakash, A.; Walzer, M.; Wang, S.; et al. The PRIDE database at 20 years: 2025 update. Nucleic Acids Res 2025, 53, D543–D553. [Google Scholar] [CrossRef]
Figure 1.
Effects of UPF on skeletal muscle gene expression. mRNA levels of (A) TNF-α, (B) IL-1β, and (C) IL-6. n = 11-16. Results are expressed as mean ± SEM. Data were analysed by one-way ANOVA, followed by post hoc LSD tests. #, p < 0.05, overall difference between (CHOW and CHOW+UPF) vs (HFD and HFD+UPF). *, p < 0.05, difference between CHOW vs CHOW+UPF and HFD vs HFD+UPF.
Figure 1.
Effects of UPF on skeletal muscle gene expression. mRNA levels of (A) TNF-α, (B) IL-1β, and (C) IL-6. n = 11-16. Results are expressed as mean ± SEM. Data were analysed by one-way ANOVA, followed by post hoc LSD tests. #, p < 0.05, overall difference between (CHOW and CHOW+UPF) vs (HFD and HFD+UPF). *, p < 0.05, difference between CHOW vs CHOW+UPF and HFD vs HFD+UPF.
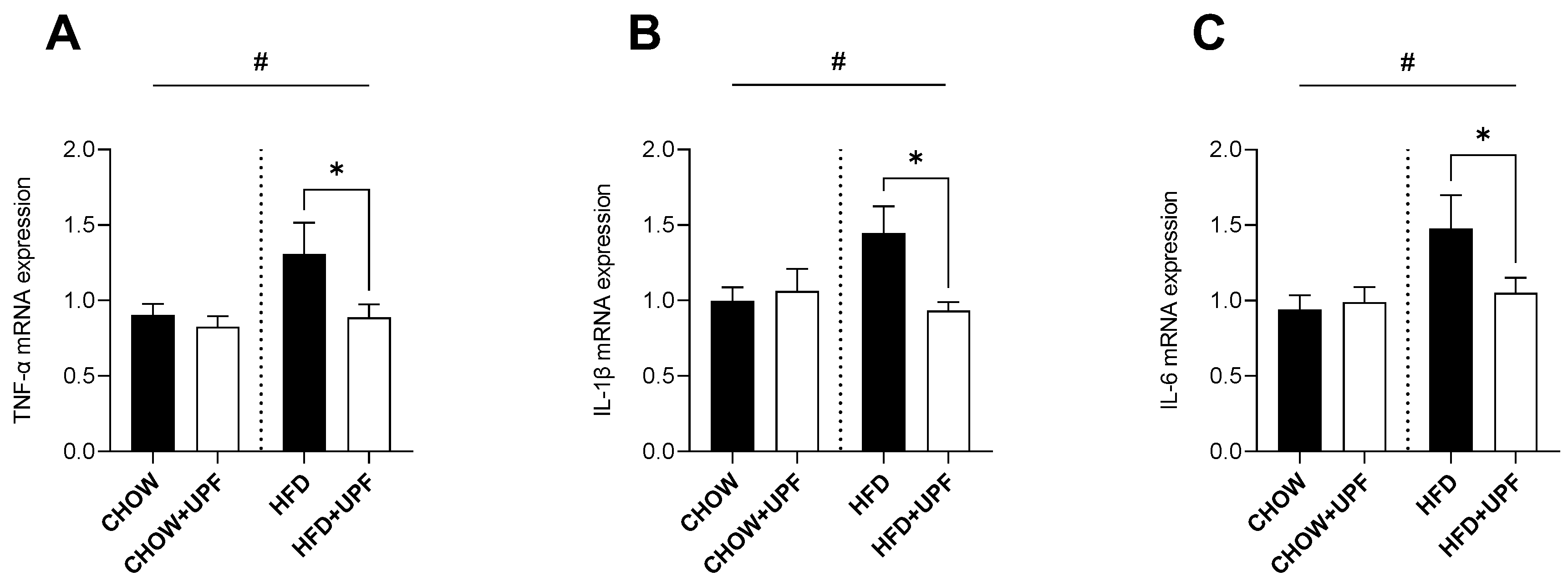
Figure 2.
Effects of UPF on small intestine gene expression. mRNA levels of (A) TNF-α, (B) IL-1β, (C) IL-6, (D) NF-κB, (E) Tjp1, (F) GPR41, and (G) GPR43. n = 7-15. Results are expressed as mean ± SEM. Data were analysed by one-way ANOVA, followed by post hoc LSD tests. #, p < 0.05, overall difference between (CHOW and CHOW+UPF) vs (HFD and HFD+UPF). *, p < 0.05, difference between CHOW vs CHOW+UPF and HFD vs HFD+UPF.
Figure 2.
Effects of UPF on small intestine gene expression. mRNA levels of (A) TNF-α, (B) IL-1β, (C) IL-6, (D) NF-κB, (E) Tjp1, (F) GPR41, and (G) GPR43. n = 7-15. Results are expressed as mean ± SEM. Data were analysed by one-way ANOVA, followed by post hoc LSD tests. #, p < 0.05, overall difference between (CHOW and CHOW+UPF) vs (HFD and HFD+UPF). *, p < 0.05, difference between CHOW vs CHOW+UPF and HFD vs HFD+UPF.
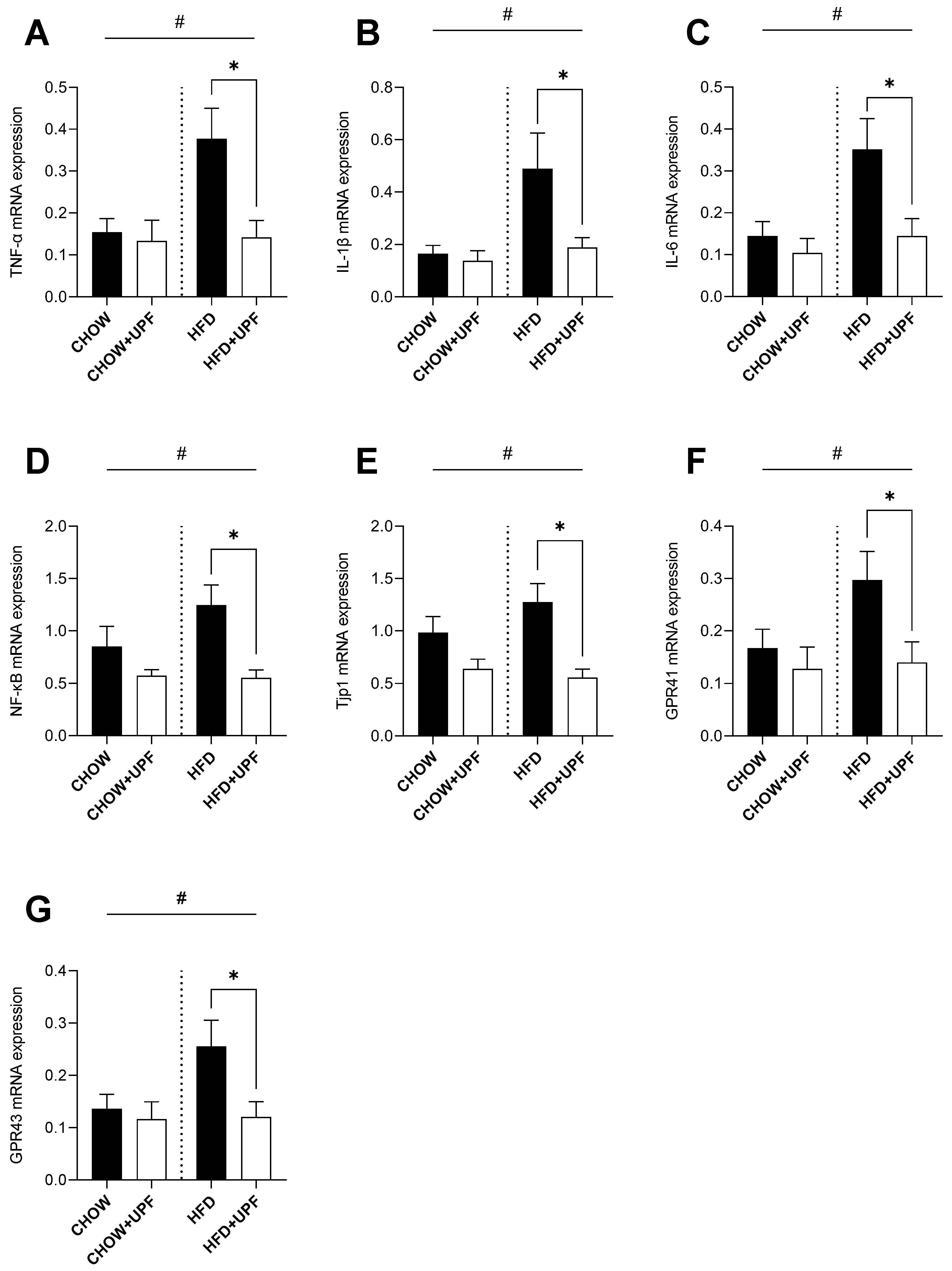
Figure 3.
Effects of UPF on hypothalamic gene expression. mRNA levels of (A) TNF-α, and (B) IL-1β. n = 7-10. Results are expressed as mean ± SEM. Data were analysed by one-way ANOVA, followed by post hoc LSD tests. #, p < 0.05, overall difference between (CHOW and CHOW+UPF) vs (HFD and HFD+UPF). *, p < 0.05, difference between CHOW vs CHOW+UPF and HFD vs HFD+UPF. (C) IL-6, and (D) IFN-γ. n = 7-10. Results are expressed as mean ± SEM. Data were analysed by Unpaired t test. *, p < 0.05, difference between HFD vs HFD+UPF.
Figure 3.
Effects of UPF on hypothalamic gene expression. mRNA levels of (A) TNF-α, and (B) IL-1β. n = 7-10. Results are expressed as mean ± SEM. Data were analysed by one-way ANOVA, followed by post hoc LSD tests. #, p < 0.05, overall difference between (CHOW and CHOW+UPF) vs (HFD and HFD+UPF). *, p < 0.05, difference between CHOW vs CHOW+UPF and HFD vs HFD+UPF. (C) IL-6, and (D) IFN-γ. n = 7-10. Results are expressed as mean ± SEM. Data were analysed by Unpaired t test. *, p < 0.05, difference between HFD vs HFD+UPF.
Figure 4.
Effects of UPF on plasma cytokines. (A) IL-1α. n = 6-10. Results are expressed as mean ± SEM. Data were analysed by one-way ANOVA, followed by post hoc LSD tests. #, p < 0.05, overall difference between (CHOW and CHOW+UPF) vs (HFD and HFD+UPF). *, p < 0.05, difference between HFD vs HFD+UPF. (B) IL-6, n=4-6. Results are expressed as mean ± SEM. Data were analysed by Unpaired t test. *, p < 0.05, difference between HFD vs HFD+UPF. (C) TNF-α, n=6-8. Results are expressed as mean ± SEM. Data were analysed by Unpaired t test.
Figure 4.
Effects of UPF on plasma cytokines. (A) IL-1α. n = 6-10. Results are expressed as mean ± SEM. Data were analysed by one-way ANOVA, followed by post hoc LSD tests. #, p < 0.05, overall difference between (CHOW and CHOW+UPF) vs (HFD and HFD+UPF). *, p < 0.05, difference between HFD vs HFD+UPF. (B) IL-6, n=4-6. Results are expressed as mean ± SEM. Data were analysed by Unpaired t test. *, p < 0.05, difference between HFD vs HFD+UPF. (C) TNF-α, n=6-8. Results are expressed as mean ± SEM. Data were analysed by Unpaired t test.
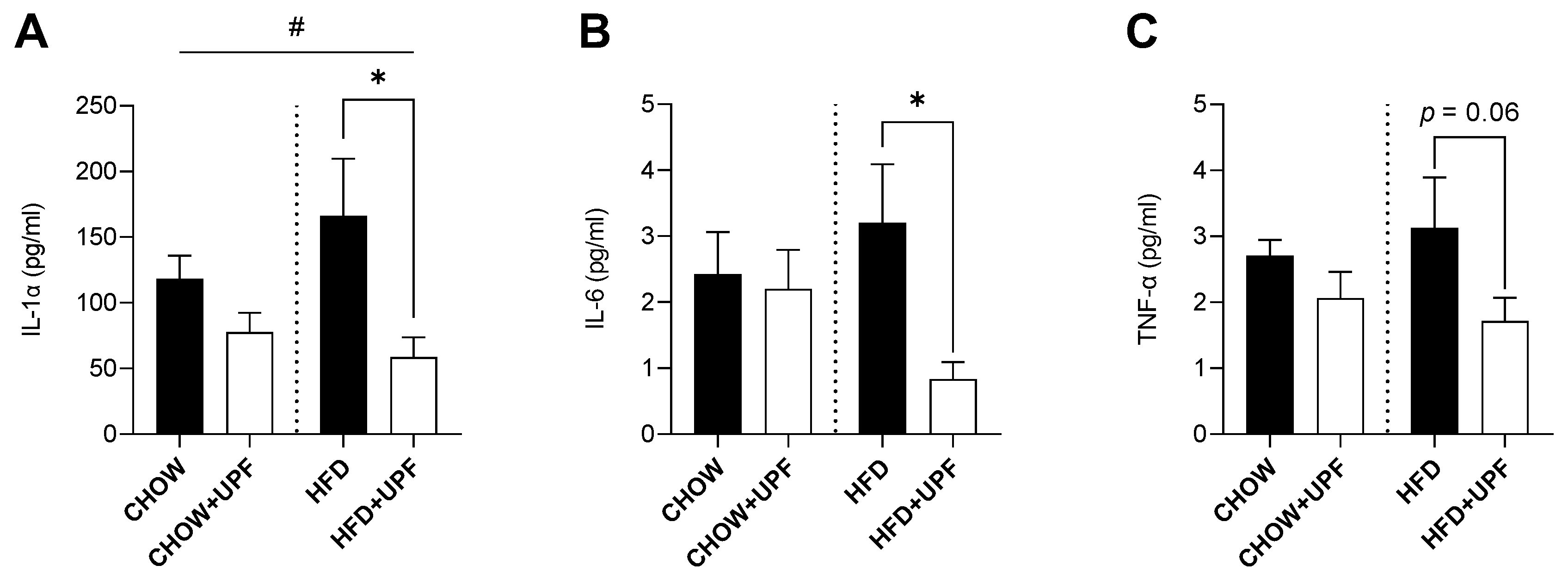
Figure 5.
Volcano plot of proteomic changes in mice NAc. (A) CHOW+UPF vs CHOW, and (B) HFD+UPF vs HFD. Proteins that were statistically significantly different relative to the control groups for their respective diets are shown above the horizontal line in red (reduced abundance in the UPF group) or blue (increased abundance in the UPF group).
Figure 5.
Volcano plot of proteomic changes in mice NAc. (A) CHOW+UPF vs CHOW, and (B) HFD+UPF vs HFD. Proteins that were statistically significantly different relative to the control groups for their respective diets are shown above the horizontal line in red (reduced abundance in the UPF group) or blue (increased abundance in the UPF group).
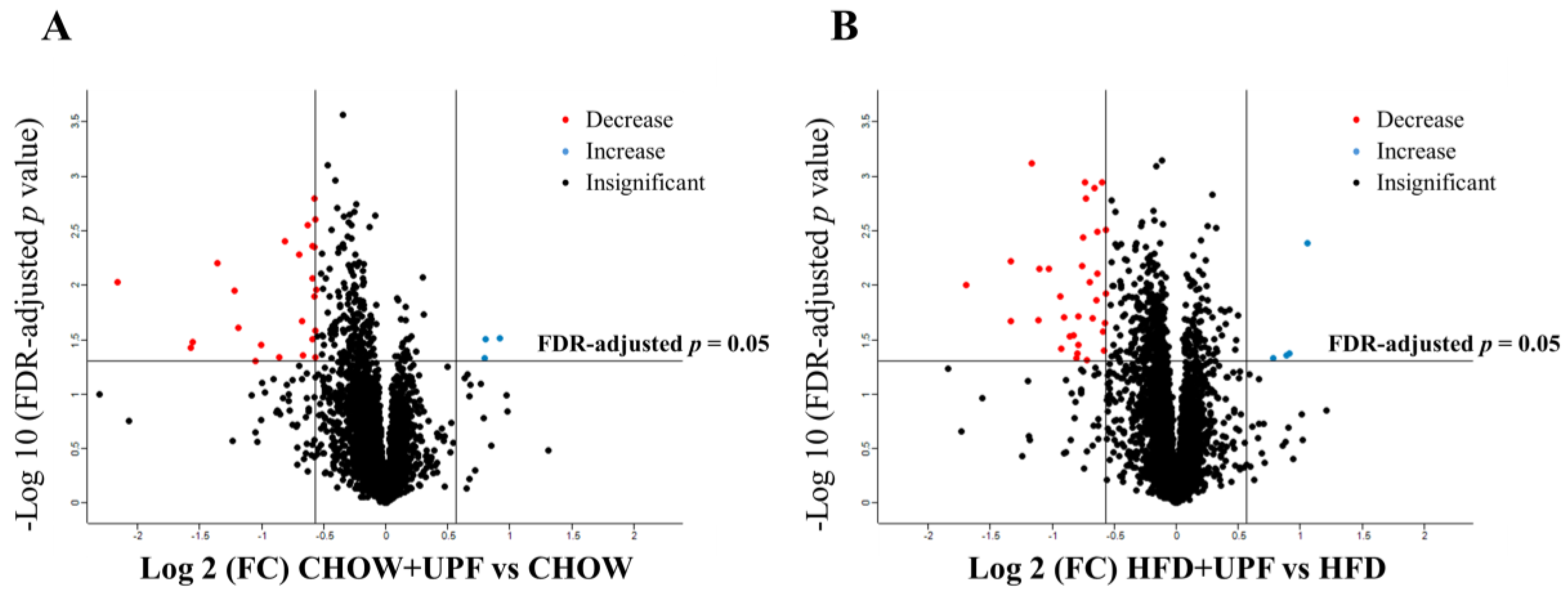
Figure 6.
Main biological processes of downregulated DEPs in CHOW groups. PPIs were visualised using the STRING interaction network. Downregulated proteins are represented as a cluster. The coloured proteins in the clusters are involved in the indicated biological process. Classification of proteins was based on Gene Ontology (GO) biological processes. All the biological processes that are referred to present a false-discovery rate less than 0.05.
Figure 6.
Main biological processes of downregulated DEPs in CHOW groups. PPIs were visualised using the STRING interaction network. Downregulated proteins are represented as a cluster. The coloured proteins in the clusters are involved in the indicated biological process. Classification of proteins was based on Gene Ontology (GO) biological processes. All the biological processes that are referred to present a false-discovery rate less than 0.05.
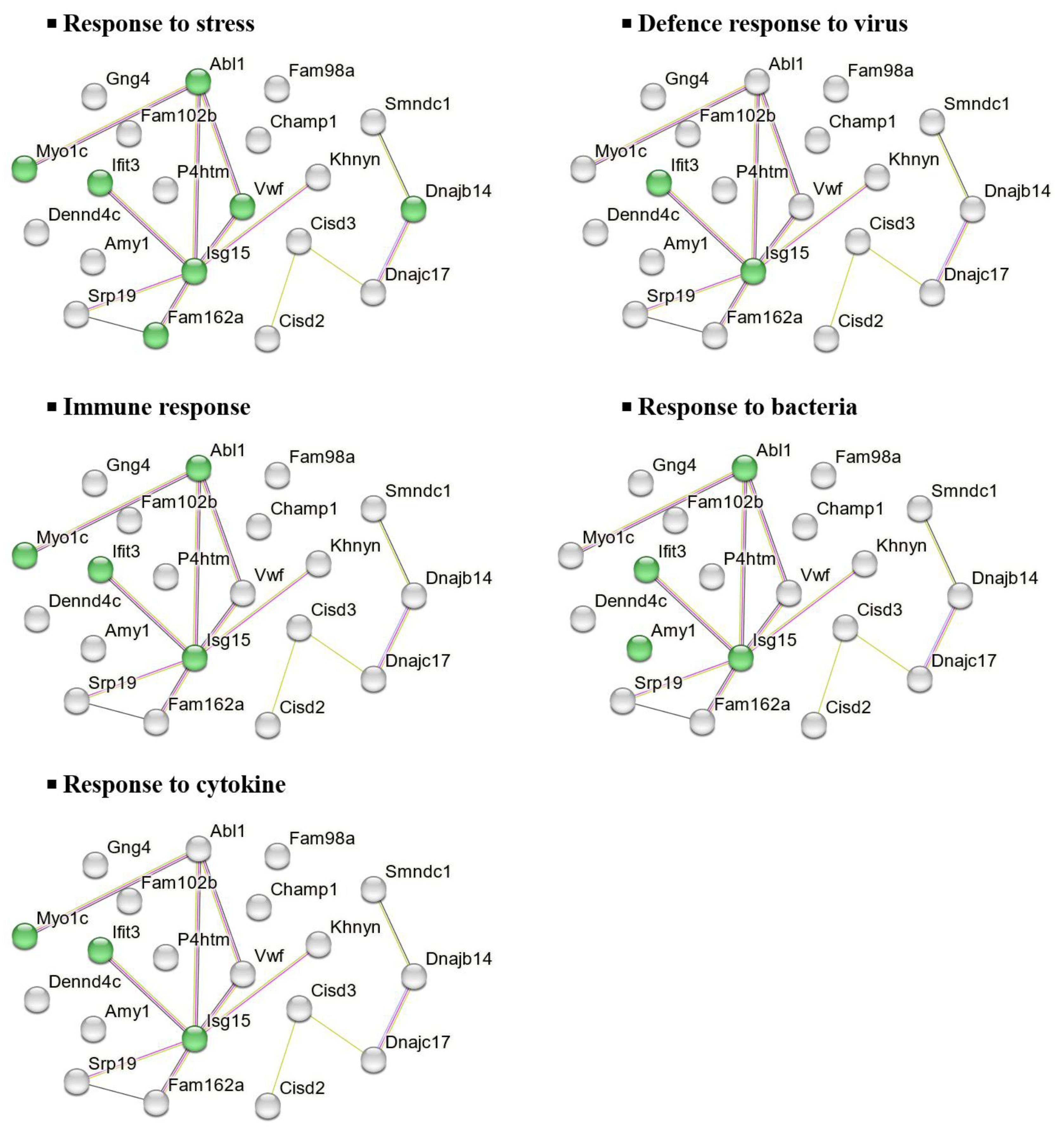
Figure 7.
Main biological processes of downregulated DEPs in HFD groups. PPIs were visualised using the STRING interaction network. Downregulated proteins are represented as a cluster. The coloured proteins in the clusters are involved in the indicated biological process. Classification of proteins was based on GO biological processes. All the biological processes that are referred to present a false-discovery rate less than 0.05.
Figure 7.
Main biological processes of downregulated DEPs in HFD groups. PPIs were visualised using the STRING interaction network. Downregulated proteins are represented as a cluster. The coloured proteins in the clusters are involved in the indicated biological process. Classification of proteins was based on GO biological processes. All the biological processes that are referred to present a false-discovery rate less than 0.05.
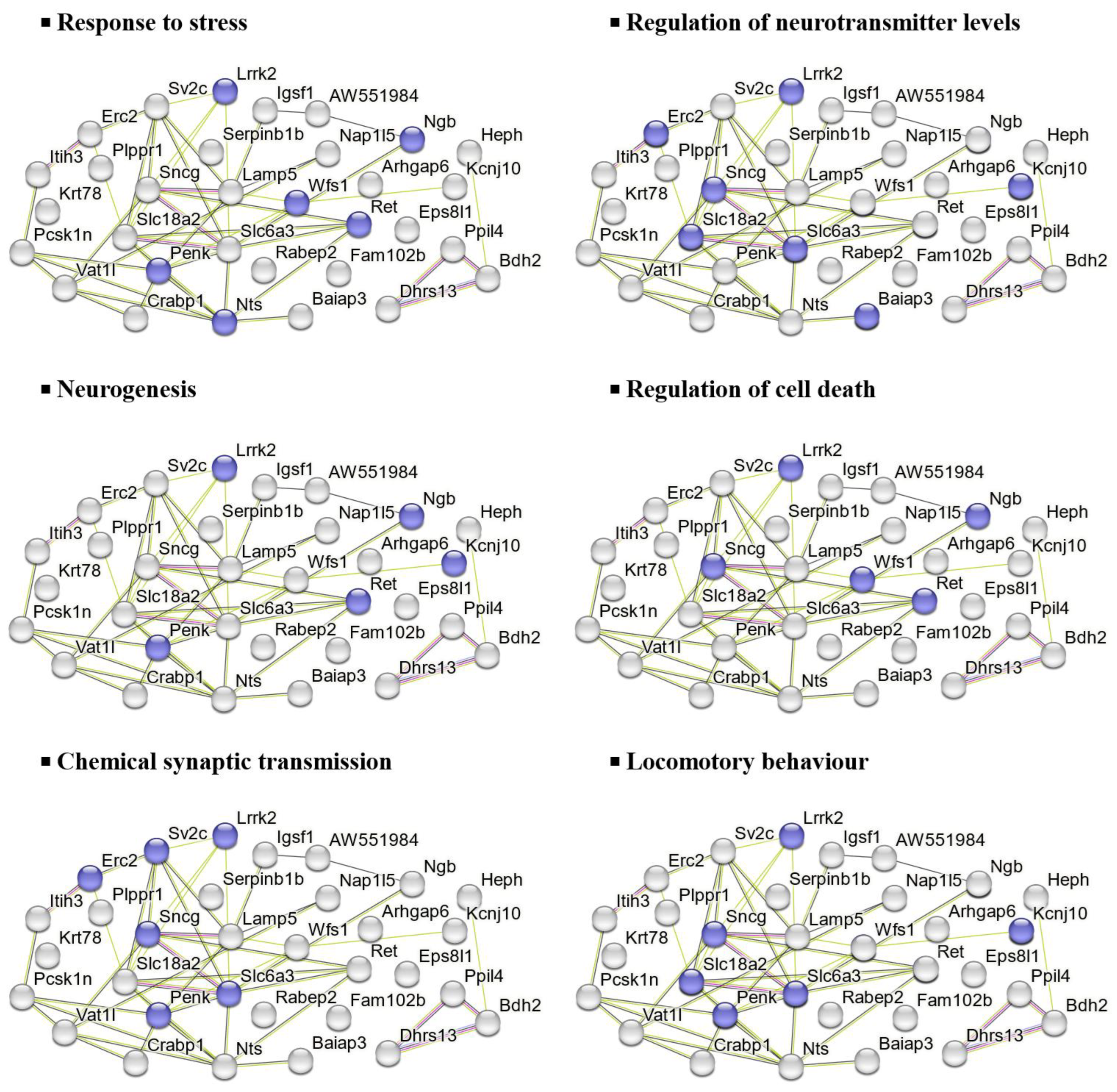
Figure 8.
Functional annotation histogram of DEPs. (A) CHOW+UPF vs CHOW, and (B) HFD+UPF vs HFD. Biological process (BP), molecular function (MF), and cellular component (CC) mainly related to the DEPs. The histograms represent the main categories for each GO term in which the upregulated and downregulated DEPs were involved (FDR-adjusted p-value < 0.05). The x-axis shows the biological activity, and the y-axis shows the percentage and count of proteins involved in this process among the total proteins in the database.
Figure 8.
Functional annotation histogram of DEPs. (A) CHOW+UPF vs CHOW, and (B) HFD+UPF vs HFD. Biological process (BP), molecular function (MF), and cellular component (CC) mainly related to the DEPs. The histograms represent the main categories for each GO term in which the upregulated and downregulated DEPs were involved (FDR-adjusted p-value < 0.05). The x-axis shows the biological activity, and the y-axis shows the percentage and count of proteins involved in this process among the total proteins in the database.
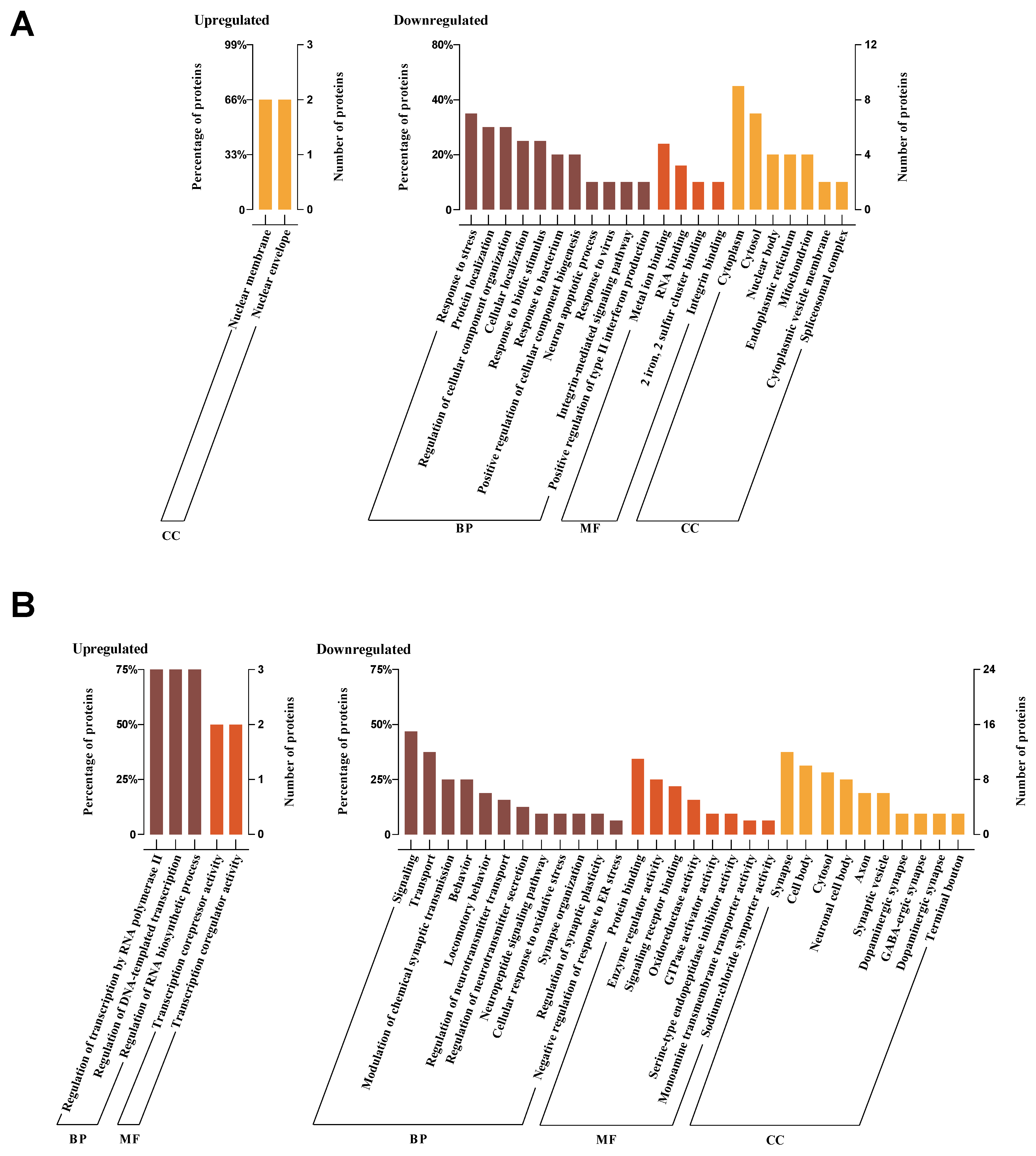

Table 1.
Proteins with significantly differential expression between CHOW+UPF and CHOW group using Perseus software platform.
Table 1.
Proteins with significantly differential expression between CHOW+UPF and CHOW group using Perseus software platform.
| Protein | UniProt Accession | Gene Name | FDR | FC |
|---|---|---|---|---|
| COP9 signalosome complex subunit 2 | A2AQE4 | Cops2 | 0.030 | 1.9 |
| Nuclear pore membrane glycoprotein 210 | Q9QY81 | Nup210 | 0.031 | 1.8 |
| Centromere protein V | Q9CXS4 | Cenpv | 0.047 | 1.7 |
| Interferon-induced protein with tetratricopeptide repeats 3 | Q64345 | Ifit3 | 0.004 | 0.7 |
| DnaJ homolog subfamily C member 17 | Q91WT4 | Dnajc17 | 0.002 | 0.7 |
| Signal recognition particle 19 kDa protein | Q9D104 | Srp19 | 0.013 | 0.7 |
| DENN domain-containing protein 4C | A6H8H2 | Dennd4c | 0.032 | 0.7 |
| Survival of motor neuron-related-splicing factor 30 | Q8BGT7 | Smndc1 | 0.009 | 0.7 |
| CDGSH iron-sulfur domain-containing protein 2 | Q9CQB5 | Cisd2 | 0.004 | 0.7 |
| Protein FAM102B | Q8BQS4 | Fam102b | 0.003 | 0.6 |
| CDGSH iron-sulfur domain-containing protein 3, mitochondrial | B1AR13 | Cisd3 | 0.044 | 0.6 |
| Guanine nucleotide-binding protein G(I)/G(S)/G(O) subunit gamma-4 | P50153 | Gng4 | 0.021 | 0.6 |
| Protein KHNYN | Q80U38 | Khnyn | 0.005 | 0.6 |
| Alpha-amylase 1 | P00687 | Amy1 | 0.046 | 0.6 |
| Ubiquitin-like protein ISG15 | Q64339 | Isg15 | 0.036 | 0.5 |
| DnaJ homolog subfamily B member 14 | Q149L6 | Dnajb14 | 0.050 | 0.5 |
| Chromosome alignment-maintaining phosphoprotein 1 | Q8K327 | Champ1 | 0.025 | 0.4 |
| Protein FAM162A | Q9D6U8 | Fam162a | 0.011 | 0.4 |
| Tyrosine-protein kinase ABL1 | P00520 | Abl1 | 0.006 | 0.4 |
| Transmembrane prolyl 4-hydroxylase | Q8BG58 | P4htm | 0.034 | 0.3 |
| Unconventional myosin-Ic | Q9WTI7 | Myo1c | 0.037 | 0.3 |
| von Willebrand factor | Q8CIZ8 | Vwf | 0.009 | 0.2 |
| Protein FAM98A | Q3TJZ6 | Fam98a | 0.015 | 0.2 |
Fold changes (FC) shown in upregulated proteins if FC > 1 or in downregulated proteins if FC < 1. Sample size: n = 10 in both CHOW+UPF and CHOW Groups.
Table 2.
Proteins with significantly differential expression between HFD+UPF and HFD group using Perseus software platform.
Table 2.
Proteins with significantly differential expression between HFD+UPF and HFD group using Perseus software platform.
| Protein | UniProt Accession | Gene Name | FDR | FC |
|---|---|---|---|---|
| Protein phosphatase 1J | Q149T7 | Ppm1j | 0.004 | 2.1 |
| Four and a half LIM domains protein 2 | O70433 | Fhl2 | 0.043 | 1.9 |
| YjeF N-terminal domain-containing protein 3 | F6W8I0 | Yjefn3 | 0.044 | 1.9 |
| Muscular LMNA-interacting protein | V9GWW6 | Mlip | 0.047 | 1.7 |
| Rho GTPase activating protein 6, isoform CRA_b | G3UZI7 | Arhgap6 | 0.012 | 0.7 |
| Synaptic vesicle glycoprotein 2C | Q69ZS6 | Sv2c | 0.022 | 0.7 |
| Sodium-dependent dopamine transporter | Q61327 | Slc6a3 | 0.040 | 0.7 |
| Dehydrogenase/reductase SDR family member 13 | Q5SS80 | Dhrs13 | 0.026 | 0.7 |
| Gamma-synuclein | Q9Z0F7 | Sncg | 0.001 | 0.7 |
| Hephaestin | Q9Z0Z4 | Heph | 0.003 | 0.6 |
| Wolframin | Q3UN10 | Wfs1 | 0.008 | 0.6 |
| ATP-sensitive inward rectifier potassium channel 10 | Q9JM63 | Kcnj10 | 0.014 | 0.6 |
| Lipid phosphate phosphatase-related protein type 1 | Q8BFZ2 | Lppr1 | 0.001 | 0.6 |
| Epidermal growth factor receptor kinase substrate 8-like protein 1 | Q8R5F8 | Eps8l1 | 0.020 | 0.6 |
| Lysosome-associated membrane glycoprotein 5 | Q9D387 | Lamp5 | 0.009 | 0.6 |
| Synaptic vesicular amine transporter | Q8BRU6 | Slc18a2 | 0.049 | 0.6 |
| Inter-alpha-trypsin inhibitor heavy chain H3 | Q61704 | Itih3 | 0.002 | 0.6 |
| Nucleosome assembly protein 1-like 5 | Q9JJF0 | Nap1l5 | 0.001 | 0.6 |
| ProSAAS | Q9QXV0 | Pcsk1n | 0.004 | 0.6 |
| Synaptic vesicle membrane protein VAT-1 homolog-like | Q80TB8 | Vat1l | 0.007 | 0.6 |
| Leucine-rich repeat serine/threonine-protein kinase 2 | Q5S006 | Lrrk2 | 0.036 | 0.6 |
| Proto-oncogene tyrosine-protein kinase receptor Ret | P35546 | Ret | 0.019 | 0.6 |
| Peptidyl-prolyl cis-trans isomerase-like 4 | Q9CXG3 | Ppil4 | 0.042 | 0.6 |
| ERC protein 2 | Q6PH08 | Erc2 | 0.029 | 0.6 |
| Proenkephalin-A | P22005 | Penk | 0.029 | 0.6 |
| Cellular retinoic acid-binding protein 1 | P62965 | Crabp1 | 0.020 | 0.5 |
| Protein FAM102B | Q8BQS4 | Fam102b | 0.038 | 0.5 |
| Protein AW551984 | Q8BGF0 | AW551984 | 0.013 | 0.5 |
| 3-hydroxybutyrate dehydrogenase type 2 | Q8JZV9 | Bdh2 | 0.007 | 0.5 |
| Leukocyte elastase inhibitor B | Q8VHP7 | Serpinb1b | 0.007 | 0.5 |
| BAI1-associated protein 3 | S4R1E7 | Baiap3 | 0.021 | 0.5 |
| Neuroglobin | Q9ER97 | Ngb | 0.001 | 0.4 |
| Protein Krt78 | E9Q0F0 | Krt78 | 0.021 | 0.4 |
| Neurotensin/neuromedin N | Q9D3P9 | Nts | 0.006 | 0.4 |
| Immunoglobulin superfamily member 1 | Q7TQA1 | Igsf1 | 0.010 | 0.3 |
| Rab GTPase-binding effector protein 2 | Q91WG2 | Rabep2 | 0.028 | 0.1 |
Fold changes (FC) shown in upregulated proteins if FC > 1 or in downregulated proteins if FC < 1. Sample size: n = 10 in both HFD+UPF and HFD Groups.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



