
Submitted:
13 March 2025
Posted:
17 March 2025
You are already at the latest version
Abstract
The application of whole-exome sequencing (WES) for diagnostic purposes has the potential to unravel secondary findings unrelated with the primary reason of testing. Some of those might be of high clinical utility and comprise disease-causing variants in genes, related to lifethreatening and clinically actionable diseases. Clarifying the allelic frequencies of such variants in specific populations is a crucial step for the large-scale deployment of genomic medicine. We analysed medically relevant variants in the 81 genes from the American College of Medical Genetics and Genomics (ACMG) v3.2 list of actionable loci, using WES data from a diagnostic laboratory cohort of 3,972 persons, tentatively resampled to represent the Portuguese population geographic distribution. We identified medically actionable variants in 6.2% of our cohort, distributed across several disease domains: cardiovascular disorders (3.0%), cancer predisposition (2.0%), miscellaneous disorders (1.1%), and metabolic disorders (0.1%). Additionally, we estimated a frequency of heterozygotes for recessive disease alleles of 11.1%. Overall, our results suggest that medically actionable findings can be identified in approximately 6.2% of persons from our population. This is the first study estimating medically actionable findings in Portugal. These results provide valuable insight for patients, healthcare providers, and policymakers involved in advancing genomic medicine at the national and international level.
Keywords:
Whole-exome sequencing
; actionable
; secondary findings
; genomic medicine
1. Introduction
Whole-exome sequencing (WES) has become widely used in clinical practice to diagnose hereditary diseases. In addition to this primary purpose, WES holds a strong potential to uncover findings with significant health implications that may be unrelated to the original indication for testing. These findings can be unexpectedly discovered (incidental findings) or intentionally sought (secondary findings).
The American College of Medical Genetics and Genomics (ACMG) published guidelines for reporting secondary findings that could influence medical management of both adults and minors undergoing genomic testing. These guidelines allow for opting-out from receiving such findings. Over time, the ACMG has progressively expanded its list of genes recommended to identify secondary findings. In 2013, the list included 57 genes; these grew to 59 in 2017, 73 in 2021, 78 in 2022, and 81 genes by 2023. They were selected with specific criteria: association with highly penetrant diseases, long asymptomatic or prodromal stages, significant contribution to morbidity and/or mortality, availability of preventive measures or treatments, and potential for early detection to reduce long-term health risks effectively [1,2,3,4,5,6,7].
In contrast, the European Society of Human Genetics (ESHG) recommended a more cautious approach, aligned with the concept of opportunistic screening. The ESHG suggests further research, and does not recommend a specific list of genes [8,9].
Since the introduction of these guidelines, various research groups have reported secondary findings identified in diverse populations. These efforts have contributed to understand the clinical significance and implications of such findings in different genetic groups; they resorted to different sampling strategies [10,11,12,13,14,15,16,17,18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39].
In Portugal, however, frequency of pathogenic (PAT) or likely-pathogenic (L-PAT) variants in ACMG-actionable genes remain unknown, and no national policies for reporting secondary findings have been issued. Access to this information is crucial to support informed decision-making by patients, genetics laboratories, healthcare providers, and policymakers. Ultimately, it might support the development of health policies for genomic screening, in each country.
In this study, we aimed at determining the frequency of PAT/L-PAT variants at loci listed as medically actionable by the ACMG (n=81), and estimating the overall frequency of secondary findings in a diagnostic cohort at the Centre for Predictive and Preventive Genetics of the Institute for Molecular and Cellular Biology (CGPP-IBMC), which was resampled to be potentially representative of the Portuguese population.
2. Results
2.1. Data Set
A subset of 3,972 persons was selected from a global dataset of 12,167 WES sequenced, as described in the Material and Methods section, and data were anonymized.
2.2. Variant Filtering and Curation
In the selected cohort, there were 12,336 variants in the 81 ACMG medically actionable genes (Figure 1). To select variants of sufficient quality, we limited the analysis to those with a minimum read depth of 10x and genotype quality of 60. In general, each of the remaining 11,207 variants was then annotated with information available from (i) our internal laboratory database, (ii) ClinVar and HGMD, (iii) gnomAD, and (iv) molecular consequence predictors, by local bioinformatic analysis. After the filtering steps, 164 PAT/L-PAT variants internally validated or present in specific disease databases were included. Then, 5,289 variants required further revision and manual curation using the ACMG/AMP guidelines and gene specific recommendations, when available; of these, additional 95 variants met the inclusion criteria.
Thus, a total of 259 variants were classified as PAT/L-PAT, across 50 genes associated to autosomal dominant (n=42), recessive (n=7) and X-linked (n=1) phenotypes (Supplementary Table S1).
2.3. Medically Actionable Findings and Carriers
After interpretation and classification of each of the 259 variants, we estimated the number of mutated alleles for each variant and corresponding gene. As expected, mutated alleles were more frequent in genes related to recessive (n=449), then dominant (n=238), and X-linked (n=4) diseases. The ACMG recommendations specify reporting only di-allelic (homozygous or compound heterozygous variants at the same locus) PAT/L-PAT variants in recessive disease genes. Frequency of potential medically actionable findings was estimated separately from carriers’ frequency, based on the zygosity of the variant, number of mutated alleles, and total number of alleles analysed.
2.3.1. Frequency of Medically Actionable Findings
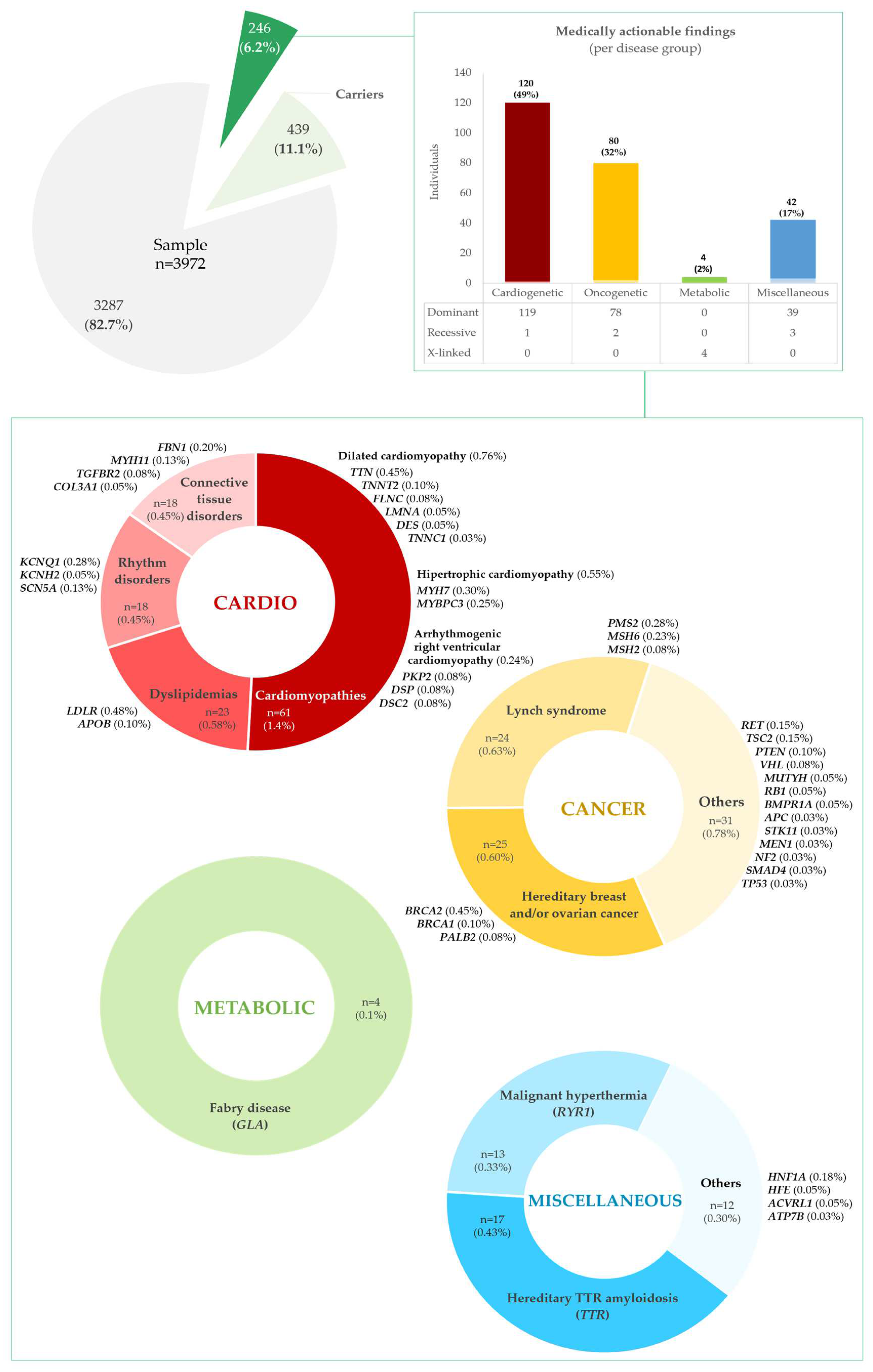
A total of 246 persons (6.19%) was estimated to have a potential medically actionable finding (Figure 2, Supplementary Tables S2A, S2B, S2C and S2D, Supplementary Tables S3A and S3B) in the cohort, distributed across four disease groups: cardiovascular, cancer, metabolic and miscellaneous disorders. Table 1 provides a summary of the total number of variants per gene and their combined allele counts, distributed across the respective disease group.
Actionable variants in genes for cardiovascular disease were the most frequent, present in 120 individuals, 48.78% of all persons with medically actionable findings and 3.02% of our cohort; these were subdivided in cardiomyopathies, CMP, (n=61, 1.54%); dyslipidaemias (n=23, 0.58%); rhythm disorders (n=18, 0.45%); and hereditary connective tissue disorders, HCTD, (n=18, 0.45%), as detailed in Supplementary Table S2A. Among the CMP, variants related to dilated CMP (DCM) were the most frequent (n=30, 0.76%), and found in TTN, TNNT2, LMNA, FLNC, DES and TNNC1; followed by hypertrophic CMP (HCM) variants (n=22, 0.55%), in MYH7 and MYBPC3. Arrhythmogenic right ventricular CMP (ARVC) variants were the least common (n=9, 0.23%), and identified in PKP2, DSP and DSC2. Among the dyslipidaemias, variants in LDLR (n=19, 0.48%) and APOB (n=4, 0.10%) were found, associated to familial hypercholesterolemia (FH). Arrhythmia predisposition variants were found in KCNQ1 (n=11, 0.28%), KCNH2 (n=2, 0.05%), and SCN5A (n=5, 0.13%) genes. Finally, HCTD findings were reported to FBN1 (n=8, 0.20%), MYH11 (n=5, 0.13%), TGFBR2 (n=3, 0.08%), and COL3A1 (n=2, 0.05%).
Pathogenic variants in genes predisposing to hereditary cancer were present in 80 persons, or 32.52% of all with medically actionable findings, and 2.01% of the cohort (see Supplementary Table S2B for details). The most frequent were associated to hereditary breast and ovarian cancer (HBOC) and Lynch syndrome (HNPCC), in 25 (0.63%) and 24 (0.60%) persons, respectively. HBOC-related variants were present in BRCA2 (n=18, 0.45%), PALB2 (n=4, 0.10%), and BRCA1 (n=3, 0.08%); and HNPCC variants in PMS2 (n=11, 0.28%), MSH6 (n=9, 0.23%), and MSH2 (n=4, 0.10%). Less frequent cancer actionable variants were seen in RET (n=6, 0.15%), TSC2 (n=6, 0.15%), PTEN (n=4, 0.10%), VHL (n=3, 0.08%), BMPR1A (n=2, 0.05%), MUTYH (n=2, 0.05%), RB1 (N=2, 0.05%), APC (n=1, 0.03%), MEN1 (n=1, 0.03%), NF2 (n=1, 0.03%), SMAD4 (n=1, 0.03%), STK11 (n=1, 0.03%), TP53 (n=1, 0.03%), genes.
Concerning metabolic disorders, PAT/L-PAT variants were identified in 4 persons (3 females, 1 male), in GLA, associated with Fabry disease, 1.63% of all with medically actionable findings and 0.10% of the cohort (refer to Supplementary Table S2C).
In the group of miscellaneous disorders, PAT/L-PAT variants were identified in 42 persons, 17.07% of all with medically actionable findings and 1.03% of the cohort (outlined in Supplementary Table S2D). Most represented variants for TTR-related hereditary amyloidosis (n=17, 0.43%) and RYR1-related malignant hyperthermia (n=13, 0.33%). Other, less frequent variants related to HNF1A - maturity onset of diabetes of the young, MODY, (n=7, 0.18%); HFE - hereditary hemochromatosis (n=2, 0.05%), ACVRL1 - hereditary haemorrhagic telangiectasia (n=2, 0.05%), and Wilson disease (n=1, 0.03%).

Table 1.
Summary of genes by phenotype group with the number of pathogenic and likely-pathogenic variants identified in our cohort according to ACMG/AMP criteria.
Table 1.
Summary of genes by phenotype group with the number of pathogenic and likely-pathogenic variants identified in our cohort according to ACMG/AMP criteria.
| Disease | Gene |
Disease Inheritance |
Nr. of variants per gene | Allele count | Allelic frequency (%) | ||
|---|---|---|---|---|---|---|---|
| Total | Hom. | Het. | |||||
| Cancer phenotype group | |||||||
| Familial adenomatous polyposis (FAP) | APC | AD | 1 | 1 | 0 | 1 | 0.013 |
| Familial medullary thyroid cancer | RET | AD | 3 | 6 | 0 | 6 | 0.076 |
| Hereditary breast and/or ovarian cancer | BRCA1 | AD | 3 | 3 | 0 | 3 | 0.038 |
| BRCA2 | AD | 16 | 18 | 0 | 18 | 0.227 | |
| PALB2 | AD | 2 | 4 | 0 | 4 | 0.050 | |
| Hereditary paraganglioma-pheochromocytoma syndrome | SDHD | AD | 0 | 0 | 0 | 0 | 0.000 |
| SDHAF2 | AD | 0 | 0 | 0 | 0 | 0.000 | |
| SDHC | AD | 0 | 0 | 0 | 0 | 0.000 | |
| SDHB | AD | 0 | 0 | 0 | 0 | 0.000 | |
| MAX | AD | 0 | 0 | 0 | 0 | 0.000 | |
| TMEM127 | AD | 0 | 0 | 0 | 0 | 0.000 | |
| Juvenile polyposis syndrome (JPS) | BMPR1A | AD | 2 | 2 | 0 | 2 | 0.025 |
| Juvenile polyposis syndrome / hereditary hemorrhagic telangiectasia syndrome | SMAD4 | AD | 1 | 1 | 0 | 1 | 0.013 |
| Li–Fraumeni syndrome | TP53 | AD | 1 | 1 | 0 | 1 | 0.013 |
Footnote: Hom.- Homozygous; Het.- Heterozygous. (end of table)
| Disease | Gene |
Disease Inheritance |
Nr. of variants per gene | Allele count | Allelic frequency (%) | |||||||
| Total | Hom. | Het. | ||||||||||
| Lynch syndrome (HNPCC) | MLH1 | AD | 0 | 0 | 0 | 0 | 0.000 | |||||
| MSH2 | AD | 3 | 4 | 0 | 4 | 0.050 | ||||||
| MSH6 | AD | 8 | 9 | 0 | 9 | 0.113 | ||||||
| PMS2 | AD | 6 | 11 | 0 | 11 | 0.138 | ||||||
| Multiple endocrine neoplasia type 1 | MEN1 | AD | 1 | 1 | 0 | 1 | 0.013 | |||||
| MUTYH-associated polyposis (MAP) | MUTYH | AR | 17 | 95 | 4 | 91 | 1.196 | |||||
| Neurofibromatosis type 2 | NF2 | AD | 1 | 1 | 0 | 1 | 0.013 | |||||
| Peutz-Jeghers syndrome (PJS) | STK11 | AD | 1 | 1 | 0 | 1 | 0.013 | |||||
| PTEN hamartoma tumor syndrome | PTEN | AD | 4 | 4 | 0 | 4 | 0.050 | |||||
| Retinoblastoma | RB1 | AD | 2 | 2 | 0 | 2 | 0.025 | |||||
| Tuberous sclerosis complex | TSC1 | AD | 0 | 0 | 0 | 0 | 0.000 | |||||
| TSC2 | AD | 6 | 6 | 0 | 6 | 0.076 | ||||||
| von Hippel-Lindau syndrome | VHL | AD | 1 | 3 | 0 | 3 | 0.038 | |||||
| WT1-related Wilms tumor | WT1 | AD | 0 | 0 | 0 | 0 | 0.000 | |||||
| Cardiovascular phenotype group | ||||||||||||
| Aortopathies | FBN1 | AD | 8 | 8 | 0 | 8 | 0.101 | |||||
| TGFBR1 | AD | 0 | 0 | 0 | 0 | 0.000 | ||||||
| TGFBR2 | AD | 2 | 3 | 0 | 3 | 0.038 | ||||||
| SMAD3 | AD | 0 | 0 | 0 | 0 | 0.000 | ||||||
| ACTA2 | AD | 0 | 0 | 0 | 0 | 0.000 | ||||||
| MYH11 | AD | 3 | 5 | 0 | 5 | 0.063 | ||||||
| Arrhythmogenic right ventricular cardiomyopathy (a subcategory of ACM) | PKP2 | AD | 3 | 3 | 0 | 3 | 0.038 | |||||
| DSP | AD | 3 | 3 | 0 | 3 | 0.038 | ||||||
| DSC2 | AD | 3 | 3 | 0 | 3 | 0.038 | ||||||
| TMEM43 | AD | 0 | 0 | 0 | 0 | 0.000 | ||||||
| DSG2 | AD | 0 | 0 | 0 | 0 | 0.000 | ||||||
| Catecholaminergic polymorphic ventricular tachycardia | RYR2 | AD | 0 | 0 | 0 | 0 | 0.000 | |||||
| CASQ2 | AR | 0 | 0 | 0 | 0 | 0.000 | ||||||
| TRDN | AR | 6 | 12 | 0 | 12 | 0.151 | ||||||
| Dilated cardiomyopathy | TNNT2 | AD | 3 | 4 | 0 | 4 | 0.050 | |||||
| LMNA | AD | 2 | 2 | 0 | 2 | 0.025 | ||||||
| FLNC | AD | 3 | 3 | 0 | 3 | 0.038 | ||||||
| TTN | AD | 18 | 18 | 0 | 18 | 0.227 | ||||||
| BAG3 | AD | 0 | 0 | 0 | 0 | 0.000 | ||||||
| DES | AD | 2 | 2 | 0 | 2 | 0.025 | ||||||
| RBM20 | AD | 0 | 0 | 0 | 0 | 0.000 | ||||||
| TNNC1 | AD | 1 | 1 | 0 | 1 | 0.013 | ||||||
| Disease | Gene |
Disease Inheritance |
Nr. of variants per gene | Allele count | Allelic frequency (%) | |||||||
| Total | Hom. | Het. | ||||||||||
| Ehlers-Danlos syndrome. vascular type | COL3A1 | AD | 2 | 2 | 0 | 2 | 0.025 | |||||
| Familial hypercholesterolemia | LDLR | AD | 15 | 19 | 0 | 19 | 0.239 | |||||
| APOB | AD | 4 | 4 | 0 | 4 | 0.050 | ||||||
| PCSK9 | AD | 0 | 0 | 0 | 0 | 0.000 | ||||||
| Hypertrophic cardiomyopathy | MYH7 | AD | 8 | 12 | 0 | 12 | 0.151 | |||||
| MYBPC3 | AD | 10 | 10 | 0 | 10 | 0.126 | ||||||
| TNNI3 | AD | 0 | 0 | 0 | 0 | 0.000 | ||||||
| TPM1 | AD | 0 | 0 | 0 | 0 | 0.000 | ||||||
| MYL3 | AD | 0 | 0 | 0 | 0 | 0.000 | ||||||
| ACTC1 | AD | 0 | 0 | 0 | 0 | 0.000 | ||||||
| PRKAG2 | AD | 0 | 0 | 0 | 0 | 0.000 | ||||||
| MYL2 | AD | 0 | 0 | 0 | 0 | 0.000 | ||||||
| Long QT syndrome types 1 and 2 | KCNQ1 | AD | 6 | 12 | 2 | 10 | 0.151 | |||||
| KCNH2 | AD | 2 | 2 | 0 | 2 | 0.025 | ||||||
| Long QT syndrome 3. Brugada syndrome | SCN5A | AD | 4 | 5 | 0 | 5 | 0.063 | |||||
| Long QT syndrome types 14-16 | CALM1 | AD | 0 | 0 | 0 | 0 | 0.000 | |||||
| CALM2 | AD | 0 | 0 | 0 | 0 | 0.000 | ||||||
| CALM3 | AD | 0 | 0 | 0 | 0 | 0.000 | ||||||
| Inborn errors of metabolism phenotype group | ||||||||||||
| Biotinidase deficiency | BTD | AR | 10 | 15 | 0 | 15 | 0.189 | |||||
| Fabry disease | GLA | XL | 3 | 4 | 1 | 3 | 0.063 | |||||
| Pompe disease | GAA | AR | 9 | 20 | 0 | 20 | 0.252 | |||||
| Ornithine transcarbamylase deficiency | OTC | XL | 0 | 0 | 0 | 0 | 0.000 | |||||
| Miscellaneous phenotype group | ||||||||||||
| Hereditary hemochromatosis | HFE | AR | 1 | 211 | 4 | 207 | 2.656 | |||||
| Hereditary hemorrhagic telangiectasia | ACVRL1 | AD | 2 | 2 | 0 | 2 | 0.025 | |||||
| ENG | AD | 0 | 0 | 0 | 0 | 0.000 | ||||||
| Malignant hyperthermia | RYR1 | AD | 7 | 13 | 0 | 13 | 0.164 | |||||
| CACNA1S | AD | 0 | 0 | 0 | 0 | 0.000 | ||||||
| Maturity-onset of diabetes of the young | HNF1A | AD | 3 | 7 | 0 | 7 | 0.088 | |||||
| RPE65-related retinopathy | RPE65 | AR | 11 | 21 | 0 | 21 | 0.264 | |||||
| Wilson disease | ATP7B | AR | 24 | 75 | 2 | 73 | 0.944 | |||||
| Hereditary TTR-related amyloidosis | TTR | AD | 2 | 17 | 0 | 17 | 0.214 | |||||
2.3.2. Carrier Frequencies
In addition to medically actionable alleles, we have estimated that 439 persons (11.05%) in our cohort were carriers of PAT/L-PAT disease-causing alleles in recessive actionable disease genes (Figure 2, Supplementary Tables S3C). These variants were most frequent in HFE (n=207, 5.21%), MUTYH (n=91, 2.29%) and ATP7B (n=73, 1.84%), associated with hereditary hemochromatosis, MUTYH-associated polyposis (MAP), and Wilson disease. Less frequently, variants in heterozygosity were found in RPE65 (n=21, 0.53%), GAA (n=20, 0.50%), BTD (n=15, 0.38%), and TRDN (n=12, 0.30%), associated with RPE65-related retinopathy, Pompe disease, biotinidase deficiency, and catecholaminergic polymorphic ventricular tachycardia. Further details are provided for each disorder group—cardiovascular, cancer, metabolic, and miscellaneous—in Supplementary Tables S4A, S4B, S4C and S4D, respectively.
2.3.3. Novel Variant Findings
Out of the 259 variants identified, we found 68 novel PAT/L-PAT variants (~26%) that have not been previously reported in the literature (as ascertained in the HGMD2024.4 Pro database and literature review). Detailed information on these novel variants is presented in Table 2. Their proportion varied across phenotype groups, with 30% in cancer-related genes, 38% in cardiovascular genes, and 6% in the miscellaneous group, while no novel findings were identified in the metabolic group.
Table 2.
Novel pathogenic and likely-pathogenic variants identified in this study that have not been previously reported in the literature.
Table 2.
Novel pathogenic and likely-pathogenic variants identified in this study that have not been previously reported in the literature.
| Gene |
cDNA (HGVS) |
Predicted Splicing Impact (Y / N) |
Protein change (HGVS) |
Freq. (gnomAD 4.1) (%) |
ClinVar ID (2025-01-02) |
||
| BMPR1A | NM_004329.3:c.231-2A>T | Y | - | - | 2866138 | ||
| NM_004329.3:c.231-1G>T | Y | - | - | 567998 | |||
| BRCA1 | NM_007294.4:c.109A>G | N | NP_009225.1:p.Thr37Ala | - | 868146 | ||
| BRCA2 | NM_000059.4:c.2974A>T | N | NP_000050.3:p.Lys992* | - | - | ||
| NM_000059.4:c.4933A>T | N | NP_000050.3:p.Lys1645* | - | 51744 | |||
| NM_000059.4:c.7258delG | N | NP_000050.3:p.Glu2420Asnfs*47 | - | - | |||
| MEN1 | NM_130799.3:c.467G>C | N | NP_570711.2:p.Gly156Ala | - | - | ||
| MSH2 | NM_000251.3:c.2084T>G | N | NP_000242.1:p.Val695Gly | - | - | ||
| MSH6 | NM_000179.3:c.195_199delACCGC | N | NP_000170.1:p.Pro66Glnfs*22 | 0.0001 | - | ||
| NM_000179.3:c.198_199insTT | N | NP_000170.1:p.Pro67Phefs*15 | - | - | |||
| NM_000179.3:c.841G>T | N | NP_000170.1:p.Gly281* | - | 2673649 | |||
| NM_000179.3:c.2437A>T | N | NP_000170.1:p.Lys813* | 0.0001 | 1791241 | |||
| NM_000179.3:c.3682_3698del | N | NP_000170.1:p.Ala1228Argfs*4 | - | - | |||
| MUTYH | NM_001128425.2:c.788+2_788+4delTAG | Y | - | - | - | ||
| NM_001128425.2:c.785_786insG | N | NP_001121897.1:p.Trp263Leufs*66 | - | - | |||
| NM_001128425.2:c.781delC | N | NP_001121897.1:p.Gln261Serfs*5 | - | - | |||
| Gene |
cDNA (HGVS) |
Predicted Splicing Impact (Y / N |
Protein change (HGVS) |
Freq. (gnomAD 4.1) (%) |
ClinVar ID (2025-01-02) |
| PTEN | NM_000314.8:c.802-1_805delGGACA | N | NP_000305.3:p.? | - | - |
| NM_000314.8:c.804_805insTTTTT | N | NP_000305.3:p.Lys269Phefs*9 | - | - | |
| RB1 | NM_000321.3:c.1422-2A>T | Y | - | 0.0004 | - |
| SMAD4 | NM_005359.6:c.904+1_904+2ins(45) | Y | - | 0.0053 | - |
| TSC2 | NM_000548.5:c.264_265delGT | N | NP_000539.2:p.Leu89Alafs*36 | 0.0160 | 45485999 |
| NM_000548.5:c.340G>T | N | NP_000539.2:p.Glu114* | - | 65033 | |
| NM_000548.5:c.775-2A>C | Y | - | - | - | |
| NM_000548.5:c.2340_2341ins(37) | N | NP_000539.2:p.Asp781Phefs*12 | - | - | |
| APOB | NM_000384.3:c.9743_9744insG | N | NP_000375.3:p.Ile3248Metfs*12 | - | - |
| NM_000384.3:c.9735delC | N | NP_000375.3:p.Gln3247Lysfs*19 | - | - | |
| NM_000384.3:c.2297_2298delAA | N | NP_000375.3:p.Lys766Ilefs*25 | - | 1553385715 | |
| COL3A1 | NM_000090.4:c.1429G>A | N | NP_000081.2:p.Gly477Arg | - | - |
| NM_000090.4:c.2229+1G>A | Y | - | - | 640856 | |
| DES | NM_001927.4:c.75_76insAG | N | NP_001918.3:p.Leu26Serfs*6 | - | - |
| DSC2 | NM_024422.6:c.1044_1047dupAAAT | N | NP_077740.1:p.Asp350Lysfs*2 | - | - |
| NM_024422.6:c.631-1G>A | Y | - | 0.0001 | 2775190 | |
| DSP | NM_004415.4:c.107delG | N | NP_004406.2:p.Gly36Alafs*12 | - | - |
| NM_004415.4:c.1258G>T | N | NP_004406.2:p.Glu420* | - | - | |
| NM_004415.4:c.2572delG | N | NP_004406.2:p.Glu858Lysfs*6 | - | - | |
| FBN1 | NM_000138.5:c.4282C>T | N | NP_000129.3:p.Arg1428Cys | 0.0007 | - |
| NM_000138.5:c.4015_4016insTG | N | NP_000129.3:p.Cys1339Leufs*75 | - | - | |
| FLNC | NM_001458.5:c.502delT | N | NP_001449.3:p.Trp168Glyfs*84 | - | - |
| NM_001458.5:c.2550+1G>A | Y | - | - | - | |
| KCNH2 | NM_000238.4:c.1621C>T | N | NP_000229.1:p.Arg541Cys | 0.0004 | 937094 |
| LDLR | NM_000527.5:c.1315A>T | N | NP_000518.1:p.Asn439Tyr | - | 375813 |
| MYBPC3 | NM_000256.3:c.2995-2A>G | Y | - | 0.0001 | - |
| MYH7 | NM_000257.4:c.1756G>A | N | NP_000248.2:p.Val586Met | 0.0004 | 1172186 |
| PKP2 | NM_004572.4:c.1489C>T | N | NP_004563.2:p.Arg497* | 0.0003 | 78974 |
| NM_004572.4:c.328delA | N | NP_004563.2:p.Met110Cysfs*2 | - | - | |
| SCN5A | NM_198056.3:c.5306C>T | N | NP_932173.1:p.Ala1769Val | 0.0001 | - |
| TGFBR2 | NM_003242.6:c.760C>T | N | NP_001020018.1:p.Arg279Cys | 0.0001 | 213942 |
| TNNT2 | NM_001276345.2:c.87_88delGG | N | NP_001263274.1:p.Asp30Argfs*13 | - | - |
| NM_001276345.2:c.80G>A | N | NP_001263274.1:p.Trp27* | - | - | |
| Gene |
cDNA (HGVS) |
Predicted Splicing Impact (Y / N |
Protein change (HGVS) |
Freq. (gnomAD 4.1) (%) |
ClinVar ID (2025-01-02) |
| TRDN | NM_006073.4:c.1831+1G>A | Y | - | - | - |
| NM_006073.4:c.1155delA | N | NP_006064.2:p.Lys385Asnfs*5 | 0.0013 | - | |
| NM_001256021.2:c.601_610delCTGGCGAAAG | N | NP_001242950.1:p.Leu201Asnfs*19 | 0.0029 | - | |
| NM_001256021.2:c.439_440delAA | N | NP_001242950.1:p.Lys147Aspfs*2 | 0.0001 | 2114339116 | |
| TTN | NM_001267550.2:c.107409_107410insCC | N | NP_001254479.2:p.Leu35804Profs*2 | - | - |
| NM_001267550.2:c.97573_97574insTC | N | NP_001254479.2:p.Asp32525Valfs*8 | - | - | |
| NM_001267550.2:c.95576_95577delAA | N | NP_001254479.2:p.Lys31859Argfs*6 | - | - | |
| NM_001267550.2:c.93623_93626dupAGCC | N | NP_001254479.2:p.Gln31210Alafs*8 | - | - | |
| NM_001267550.2:c.84525G>A | N | NP_001254479.2:p.Trp28175* | - | - | |
| NM_001267550.2:c.79811dupT | N | NP_001254479.2:p.Arg26605Lysfs*19 | - | - | |
| NM_001267550.2:c.70971_70972insT | N | NP_001254479.2:p.Leu23658Serfs*18 | - | - | |
| NM_001267550.2:c.64266delA | N | NP_001254479.2:p.Asp21423Ilefs*2 | 0.0001 | - | |
| NM_001267550.2:c.58709C>G | N | NP_001254479.2:p.Ser19570* | - | - | |
| NM_001267550.2:c.52975_52976delCA | N | NP_001254479.2:p.Gln17659Thrfs*6 | 0.0001 | - | |
| NM_001267550.2:c.41845dupA | N | NP_001254479.2:p.Ile13949Asnfs*2 | - | - | |
| NM_001267550.2:c.13184delT | N | NP_001254479.2:p.Leu4395Argfs*25 | - | - | |
| ACVRL1 | NM_000020.3:c.830C>T | N | NP_000011.2:p.Thr277Met | 0.0004 | 2731545 |
| ATP7B | NM_000053.4:c.3959G>C | N | NP_000044.2:p.Arg1320Thr | 0.0010 | 1479012 |
| RPE65 | NM_000329.3:c.1544G>A | N | NP_000320.1:p.Arg515Gln | 0.0015 | 1052287 |
Footnote: Freq.- Frequency; Y- Yes; N- No. (end of table).
3. Discussion
This research developed a strategy to obtain a potentially representative sample of the Portuguese population using WES-derived data from a diagnostic setting. The final purpose was to determine the frequency of PAT/L-PAT variants at ACMG´s medically actionable loci. Such data, however, were previous unavailable for our Portuguese population.
We could infer that 6.19% of healthy persons had potentially actionable findings. This is in line with previous reports, ranging 0.59% to 12.64% [10,11,12,13,14,15,16,17,18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39]. Several factors may explain this variation, including differences in populations, sample size, study design, number of genes screened, sequencing technology, and variant filtering, interpretation and classification; in addition to actionability criteria, and the increasing number of validated PAT/L-PAT variants in disease databases over time.
This study may also stand out from others for several reasons: (i) unlike most other, our cohort was resampled to be as representative as possible of the general population; (ii) sample size was larger and more significant considering our population (of about 10.5 million people); (iii) the most recent ACMG list (v. 3.2, 2023) of 81 genes was analysed; and (iv) variant filtering and interpretation followed the most recent guidelines and gene specifications, and included a manual curation step.
In line with previous studies, PAT/L-PAT variants identified associated with “silent” but life-threatening diseases within four groups: cardiovascular, cancer, miscellaneous and metabolic disorders. Variants associated to cardiovascular diseases were the most frequent, and included cardiomyopathy, arrhythmias, dyslipidaemia, and connective tissue disorders.
CMP-related variant frequencies aligned with similar studies or disease prevalence (DCM: 1:500, HCM: 1:500, ARVC: 1:1,000–5,000), considering penetrance (DCM: 45%, HCM: 40%, ARVC: 30–75%) [40,41,42]. Likewise, primary arrhythmia variants, notably LQTS (0.45%), matched reported prevalence (1:2,500) and penetrance (75%) [43]. Both CMP and LQTS pose sudden death risks, warranting regular clinical surveillance, pharmacotherapy, and potentially implantable cardioverter defibrillators.
Concerning dyslipidaemia, frequency of FH variants (0.58%) is in accordance with previously estimated disease frequency (1:200-250) in a meta-analysis [44]. FH leads to coronary heart disease events and death, if untreated. It has been estimated that only less than 10% of persons with FH are diagnosed, and even less receive treatment. Thus, early detection and possible treatment with lipid-lowering drugs are crucial.
On the other hand, HCTD related findings, including Marfan syndrome (MS, 0.20%), familial thoracic aortic aneurism and dissection (FTAAD, 0.13%), Loeys-Dietz syndrome (LDS, 0.08%), and vascular type of Ehlers-Danlos syndrome (vEDS, 0.05%) were more frequent than expected, according to disease frequency in the literature (1:3,300-1:20,000 for MS; 1:5,000-1:4,000,000 for FTAAD; unknown for LDS; and 1:50,000 for vEDS) [45,46,47,48]. Due to an increased risk of rupture of the aorta, early diagnosis is life-saving, in order to proceed with surveillance, therapy, and eventually prosthetic surgery.
The second most frequent group of findings is related with cancer, being HBOC and HNPCC the most common. Frequency of reportable HBOC-related variants was estimated as 0.63%. This is 2-5-fold higher, when compared to the described frequency of BRCA pathogenic variants (1:400 to 1:800) [49]. However, this rate may be underestimate in our cohort, given the limitation to detect the NM_000059.4(BRCA2):c.156_157insAlu (a known founder variant in the Portuguese population) using WES data [50]. Mismatch-repair variants showed a frequency of 0.60%, similarly to reported prevalence of HNPCC of 1:279 in the general population, and incomplete penetrance (>90% for colonic adenomas) [51]. All these findings support the relevance of early diagnosis of hereditary cancer syndromes, since these persons may be offered clinical surveillance, following family history and knowing genetic status.
In the metabolic disorders group, we highlight the findings related to Fabry disease. Our screening showed a frequency of 0.10% for variants in GLA, corroborating recent data on prevalence of Fabry disease (1:1,250 to 1:5,732) [58,59,60,61,62]. Specifically, NM_000169.3:c.337T>C, p.(Phe113Leu), a founder variant in the region of Guimarães, Portugal and associated to the late-onset form, was found in 0.03% [63]. This higher prevalence of Fabry disease is related to knowledge of a broader phenotypic spectrum for GLA, encompasses the classical and a late-onset form. Persons with this late-onset phenotype may have significant diagnostic delay [64]. Genetic testing may allow for earlier diagnosis, critical for efficiency of enzyme replacement or chaperone therapy, recommended to be initiated as early as possible.
TTR-related hereditary amyloidosis has a significant cluster in northern Portugal, the largest worldwide [52,53]. In our data the frequency of TTR pathogenic variants was 0.43%. The p.Val50Met variant (NM_000371.4:c.148G>A) showed a frequency of 0.28%, comparable to that reported in northern Portugal (1:538) [54]. This variant is predominantly associated to TTR-related amyloid neuropathy [55]. Surprisingly, the p.Val142Ile variant (NM_000371.4:c.424G>A), known to be associated to the TTR-related cardiac amyloidosis, particularly prevalent in persons of African ancestry, was also present in 0.16% [55]. The higher prevalence of TTR variants identified supports the greater awareness for TTR amyloidosis in Portugal, allowing for early diagnosis, follow-up and treatment, including liver transplant, pharmacotherapy, and ICD implant when indicated.
We also identified variants in the RYR1 gene with an overall frequency of 0.33%. The exact worldwide prevalence of malignant hyperthermia susceptibility (MHS) has been difficult to clarify given disparities in clinical diagnosis methods and criteria. Using population genomics data, estimated prevalence of an MHS-related pathogenic variants was 1:1450-1:1556 [56,57]. Here, we report a frequency 10-times higher, after having restricted to variants functionally validated and curated by the European Malignant Hyperthermia Group. Assessment of risk for MHS is relevant so that effective measures, such as avoidance of certain anaesthetics upon surgery.
In addition, we identified heterozygous variants in genes associated with autosomal recessive diseases. In total, 11.05% of persons were carriers for any high-risk ACMG actionable genes linked to recessive diseases, including HFE, MUTYH, ATP7B, RPE65, GAA, BTD and TRDN. Frequency, in our cohort, of heterozygotes at HFE (5.21%), MUTYH (2.29%), ATP7B (1.83%), GAA (0.50%) and BTD (0.38%) was comparable to previous estimates in the general population (1:10 for HFE, 1:50 for MUTYH, 1:50-90 for ATP7B, 1:70 for GAA, and 1:120 for BTD) [51,65,66,67,68,69,70,71]. For comparison, the American College of Obstetricians and Gynecologists endorsed carrier screening for cystic fibrosis and spinal muscular atrophy, as well as other diseases with a carrier frequency ≥1/100 [72]. Our data may provide support to which genes should be selected for preconception carrier screening based on population carrier frequency, to address together with considerations on penetrance, severity, and predictable genotype-phenotype correlation.
Our work shows some limitations, most of which are transversal to similar published studies:
- Lab cohort bias. Our sample was derived from cases ascertained for genetic diagnosis of various Mendelian disorders; therefore, a few persons in the cohort may already be affected by a disease attributable to one of the genes in the ACMG list. Despite this, when excluding L-PAT/PAT variants listed as primary diagnosis in the genetic test reports of these patients, the overall frequency did not differ significantly (only 0.6%);
- Gene list. We limited our analysis to the current set of the ACMG genes. We did not consider other clinically relevant genes, as those curated by the ClinGen Actionability Working Group, for instance. Inclusion of additional conditions, some of specific impact in the Portuguese population, should be considered in future studies, what might increase the overall frequency of actionable findings;
- Study design. In order to minimize impact of data used, our project protocol prevented us from including individual-level information regarding ethnic background, age, gender, reason for referral for WES, or phenotype. Additionally, genotypes obtained were related to the whole cohort, not the patient. Consequently, we were not able to estimate compound heterozygosity, or the number of findings per individual;
- Technical limitations. Methodologies used may have led to missed variants due to: (i) intrinsic WES limitation to detect deep intronic, triplet repeats expansion, and structural variants; (ii) use of different capture kits along time within this cohort; (iii) incomplete coverage in some regions; (iv) not considering structural variants, including copy number variants (CNVs); and (v) MAF cut-off;
- Potential for false-positive interpretation of variants. Variants accurately classified as PAT/L-PAT, based on available evidence, may not be in fact disease causing, due to incomplete penetrance or variable expressivity. This is exacerbated when genetic testing is performed in the context of population screening;
- Actionability. The term “actionable” is highly subjective and its application may fluctuate. The ClinGen Actionability Working Group is addressing this issue by curating the actionability of several gene-disease groups, including those listed by the ACMG. We took this into consideration; however, some gene-disease groups are not yet curated and others are classified as actionable depending on individual-level information, such as age and sex, which were not considered due to our study protocol.
Future studies, including more persons, are needed to determine more accurately the frequency of ultra-rare disease-causing alleles in our study population. Our methodology could also be used to evaluate the allelic frequency for non-ACMG actionable loci. This could include other diseases known to have impact in Portugal, such as haemoglobinopathies, congenital adrenal hyperplasia, and cystic fibrosis; some of those genes may be of interest for designing of preconceptional screening programmes. Optimizing guidelines for variant interpretation and reporting criteria in asymptomatic persons is also suggested, as well as research on long-term phenotypic effects of presumed PAT/L-PAT variants in the general population. Finally, it is recommended to perform responsible research on genomic screening, before considering its eventual implementation within healthcare services.
4. Materials and Methods
4.1. Study Design and Data Set of Exomes
This is a cross-sectional observational study with a quantitative approach. We analysed the WES data obtained from patients tested at CGPP-IBMC (12,167 samples, retrieved in 19-05-2023), in an anonymized and aggregated form. All patients had been clinically diagnosed or had a diagnostic suspicion of a disease for which the molecular test requested by their physician was either a virtual gene-panel based on WES or a complete WES analysis.
4.2. Resampling from CGPP-IBMC Clinical Database
The approach used to generate a sample approaching representativeness of the Portuguese population is detailed elsewhere [74]. Briefly, to remove possible population biases, the CGPP-IBMC cohort was resampled to i) include only one sample per family; ii) include one partner per consanguineous couple; iii) remove pre-natal or foetal samples. The final subset used was corrected for geographic distribution of cases and population size, by using data from Census 2021 [75]; also, distinct datasets were made according to the known distribution of the population, by municipalities both in mainland Portugal, and Madeira and the Azores archipelagos.
4.3. Selection of Genes for Which Reporting of Secondary Findings Are Recommended
The list of genes in the present analysis covered the 81 genes that are part of the revised guidelines on secondary findings from the ACMG, version 3.2 [7]. This list encompasses genes related to cancer (n=28), cardiovascular (n=40), inborn errors of metabolism (n=4) and miscellaneous (n=7) phenotypes (Supplementary Table S5).
4.4. Data Processing
BED files, containing genomic coordinates corresponding to all 81 ACMG genes (exonic coding regions plus 20bp intronic flanks), were intersected with the full WES VCF files from each sample, to select variants in those genes. Variants with <10x coverage, <30% heterozygous allele ratio and base-call quality score ≤Q20 were excluded. For each gene, the obtained individual VCF files, containing the gene’s variants, were processed into aggregated, multi-sample VCF files. These multi-sample VCFs were then further processed by calculating allele count, allele number, allele frequency, number of homozygotes, heterozygotes and hemizygotes, and removing individual genotype information to ensure irreversible anonymization of patients’ data. This aggregation and anonymization were performed for (i) all samples and (ii) resampled data, grouped by municipalities, as described previously.
4.5. Variant Annotation
Variants were systematically annotated with information available from (i) our in-house database, (ii) disease databases, namely ClinVar (retrieved 2023-09-07) and HGMD (version 2023.2), (iii) population database gnomAD (versions 2.1.1, 3.1.2 and 4.1), and (iv) molecular consequence predictors.
4.6. Variant Filtering
The overall filtering and manual curation workflow are explained in Figure 1 and Supplementary Tables S6A, 6B and 6C. First, variants were filtered based on confidence metrics including genotype quality (GQ, ≥60) and read depth (RD, minimum 10x). Second, variants were prioritized according to prior classification in our in-house laboratory database, being automatically included if classified as PAT/L-PAT. Third, variants were filtered based in previous ClinVar classification, being included if classified as PAT/L-PAT 2 stars and excluded if BEN/L-BEN 2 stars. Fourth, variants were filtered based on their minor allele frequency (MAF), with exclusion of variants with MAF≥0.5% in gnomAD, except for the HFE variant NM_000410.4:c.845G>A (p.Cys282Tyr). Finally, variants were filtered according to effect prediction (VEP); variants with low predicted impact were excluded.
4.7. Manual Variant Curation, Classification, and Actionability
We further curated manually the remaining variants, following the guidelines for interpretation and classification of sequence variants from the ACMG/AMP [76]. Additionally, specifications for variant classification of the Association for Clinical Genomic Science (ACGS), Clinical Genome Resource (ClinGen), Cancer Variant Interpretation Group UK (CanVIG-UK), and the European Molecular Genetics Quality Network (EMQN) were considered, when applicable (Supplementary Tables S6A, S6B and S6C) [77,78,79,80]. All variants classified as PAT/L-PAT, referring to the potential of the variant to cause a relevant phenotype according to the ACMG v3.2 recommendations, were considered to be medically actionable.
4.8. Frequency of Actionable Findings Calculation
The overall frequency of actionable findings was then calculated considering the total of size of the WES cohort and the number of genes.
5. Conclusions
To the best of our knowledge, this is the first evaluation of medically actionable findings in the Portuguese population, making this study a pioneering effort. We demonstrate that this population, as others, is expected to harbour medically actionable variants that can be identified through WES in routine diagnosis; however, while our study provides important insight, additional evidence is needed to take its findings to the population level.
This foundational research establishes a basis for future investigations into genomic screening and highlights the potential for data from WES-based genetic testing to significantly impact public health.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1- Medically ACMG actionable findings and carrier status identified in our study. Tables S2: Frequency of medically actionable findings per gene [S2A- Cardiogenetic; S2B- Oncogenetic; S2C- Metabolic; S2D- Miscellaneous]. Tables S3: Overall frequencies, actionable findings and carrier frequencies per disease [S3A- Overall frequency of ACMG medically actionable findings and non-medically actionable findings (carriers); S3B- Frequency of ACMG medically actionable findings (per disease group); S3C- Frequency of carrier findings (per disease group)]. Tables S4: Carrier frequencies for recessive disease findings per gene [S4A- Cardiogenetic carrier; S4B- Oncogenetic carrier; S4C- Metabolic carrier; S4D- Miscellaneous carrier]. Table S5- List of genes for which reporting incidental findings are recommended by the ACMG. Tables S6: Manual curation and variant classification [S6A- Gene specifications; S6B- Standard manual curation - evidence analysis; S6C- Standard manual curation - variant class calculation]. Supplementary data S1. References for Table S6A Gene specifications - variant interpretation details.
Author Contributions
Conceptualization and supervision: J.P.F., D.A. and J.O.; Methodology: M.M., P.S., M.R., S.V., F.A., J.P.F, J.O.; Formal Analysis: M.M., M.R., S.V, P.S.; Investigation, M.M., M.R., S.V.; Writing – Original Draft Preparation, M.M.; Writing – Review & Editing, M.R., S.V., P.S., J.P.F., D.A., J.S., J.O.; Critical commenting and improvement: M.M., P.S., M.R., S.V., F.A., M.V., J.S., J.P.F., D.A., J.O.
Funding
This research received no external funding.
Institutional Review Board Statement
The research protocol was reviewed and approved by the Committee for Ethical and Responsible Conduct of Research of i3S (Appraisal N 10/CECRI/2023).
Informed Consent Statement
In accordance with national law, informed consent was waived, due to the use of an irreversibly anonymized WES dataset and analysis in an aggregated manner, with no individual-level data available.
Data Availability Statement
NGS datasets used for this article resulted from a clinical diagnostic setting and are not publicly available. Main data needed to assess the conclusions in the paper are presented in the main text or in the Supplementary files.
Acknowledgments
All resources were provided by the Centre for Predictive and Preventive Genetics, of the Institute for Cellular and Molecular Biology (CGPP-IBMC). We thank Ana Coutinho for the collaboration on variant curation procedures.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Green, R.C.; Berg, J.S.; Grody, W.W.; Kalia, S.S.; Korf, B.R.; Martin, C.L.; McGuire, A.L.; Nussbaum, R.L.; O’Daniel, J.M.; Ormond, K.E.; et al. ACMG Recommendations for Reporting of Incidental Findings in Clinical Exome and Genome Sequencing. Genet. Med. 2013, 15, 565–574. [Google Scholar] [CrossRef]
- Kalia, S.S.; Adelman, K.; Bale, S.J.; Chung, W.K.; Eng, C.; Evans, J.P.; Herman, G.E.; Hufnagel, S.B.; Klein, T.E.; Korf, B.R.; et al. Recommendations for Reporting of Secondary Findings in Clinical Exome and Genome Sequencing, 2016 Update (ACMG SF v2.0): A Policy Statement of the American College of Medical Genetics and Genomics. Genet. Med. 2017, 19, 249–255. [Google Scholar] [CrossRef] [PubMed]
- Biesecker, L.G. ACMG Secondary Findings 2.0. Genet. Med. 2017, 19, 604. [Google Scholar] [CrossRef]
- Miller, D.T.; Lee, K.; Gordon, A.S.; Amendola, L.M.; Adelman, K.; Bale, S.J.; Chung, W.K.; Gollob, M.H.; Harrison, S.M.; Herman, G.E.; et al. Recommendations for Reporting of Secondary Findings in Clinical Exome and Genome Sequencing, 2021 Update: A Policy Statement of the American College of Medical Genetics and Genomics (ACMG). Genet. Med. 2021, 23, 1391–1398. [Google Scholar] [CrossRef]
- Miller, D.T.; Lee, K.; Chung, W.K.; Gordon, A.S.; Herman, G.E.; Klein, T.E.; Stewart, D.R.; Amendola, L.M.; Adelman, K.; Bale, S.J.; et al. ACMG SF v3.0 List for Reporting of Secondary Findings in Clinical Exome and Genome Sequencing: A Policy Statement of the American College of Medical Genetics and Genomics (ACMG). Genet. Med. 2021, 23, 1381–1390. [Google Scholar] [CrossRef]
- Miller, D.T.; Lee, K.; Abul-Husn, N.S.; Amendola, L.M.; Brothers, K.; Chung, W.K.; Gollob, M.H.; Gordon, A.S.; Harrison, S.M.; Hershberger, R.E.; et al. ACMG SF v3.1 List for Reporting of Secondary Findings in Clinical Exome and Genome Sequencing: A Policy Statement of the American College of Medical Genetics and Genomics (ACMG). Genet. Med. 2022, 24, 1407–1414. [Google Scholar] [CrossRef] [PubMed]
- Miller, D.T.; Lee, K.; Abul-Husn, N.S.; Amendola, L.M.; Brothers, K.; Chung, W.K.; Gollob, M.H.; Gordon, A.S.; Harrison, S.M.; Hershberger, R.E.; et al. ACMG SF v3.2 List for Reporting of Secondary Findings in Clinical Exome and Genome Sequencing: A Policy Statement of the American College of Medical Genetics and Genomics (ACMG). Genetics in Medicine 2023, 25. [Google Scholar] [CrossRef] [PubMed]
- Van El, C.G.; Cornel, M.C.; Borry, P.; Hastings, R.J.; Fellmann, F.; Hodgson, S.V.; Howard, H.C.; Cambon-Thomsen, A.; Knoppers, B.M.; Meijers-Heijboer, H.; et al. Whole-Genome Sequencing in Health Care. Eur. J. Hum. Genet. 2013, 21, 580–584. [Google Scholar] [CrossRef]
- de Wert, G.; Dondorp, W.; Clarke, A.; Dequeker, E.M.C.; Cordier, C.; Deans, Z.; van El, C.G.; Fellmann, F.; Hastings, R.; Hentze, S.; et al. Opportunistic Genomic Screening. Recommendations of the European Society of Human Genetics. Eur. J. Hum. Genet. 2021, 29, 365–377. [Google Scholar] [CrossRef]
- Elfatih, A.; Mohammed, I.; Abdelrahman, D.; Mifsud, B. Frequency and Management of Medically Actionable Incidental Findings from Genome and Exome Sequencing Data: A Systematic Review. Physiol Genom. 2021, 53, 373–384. [Google Scholar] [CrossRef]
- Dorschner, M.O.; Amendola, L.M.; Turner, E.H.; Robertson, P.D.; Shirts, B.H.; Gallego, C.J.; Bennett, R.L.; Jones, K.L.; Tokita, M.J.; Bennett, J.T.; et al. Actionable, Pathogenic Incidental Findings in 1,000 Participants’ Exomes. Am J Hum Genet 2013, 93, 631–640. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, L.; Sincan, M.; Markello, T.; Adams, D.R.; Gill, F.; Godfrey, R.; Golas, G.; Groden, C.; Landis, D.; Nehrebecky, M.; et al. The Implications of Familial Incidental Findings from Exome Sequencing: The NIH Undiagnosed Diseases Program Experience. Genet. Med. 2014, 16, 741–750. [Google Scholar] [CrossRef]
- Amendola, L.M.; Dorschner, M.O.; Robertson, P.D.; Salama, J.S.; Hart, R.; Shirts, B.H.; Murray, M.L.; Tokita, M.J.; Gallego, C.J.; Kim, D.S.; et al. Actionable Exomic Incidental Findings in 6503 Participants: Challenges of Variant Classification. Genome Res 2015, 25, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Olfson, E.; Cottrell, C.E.; Davidson, N.O.; Gurnett, C.A.; Heusel, J.W.; Stitziel, N.O.; Chen, L.S.; Hartz, S.; Nagarajan, R.; Saccone, N.L.; et al. Identification of Medically Actionable Secondary Findings in the 1000 Genomes. PLoS One 2015, 10. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Muzny, D.M.; Xia, F.; Niu, Z.; Person, R.; Ding, Y.; Ward, P.; Braxton, A.; Wang, M.; Buhay, C.; et al. Molecular Findings among Patients Referred for Clinical Whole-Exome Sequencing. JAMA - J. Am. Med. Assoc. 2014, 312, 1870–1879. [Google Scholar] [CrossRef]
- Dewey, F.E.; Murray, M.F.; Overton, J.D.; Habegger, L.; Leader, J.B.; Fetterolf, S.N.; O’Dushlaine, C.; Van Hout, C.V.; Staples, J.; Gonzaga-Jauregui, C.; et al. Distribution and Clinical Impact of Functional Variants in 50,726 Whole-Exome Sequences from the DiscovEHR Study. Science (1979) 2016, 354. [Google Scholar] [CrossRef]
- Jurgens, J.; Ling, H.; Hetrick, K.; Pugh, E.; Schiettecatte, F.; Doheny, K.; Hamosh, A.; Avramopoulos, D.; Valle, D.; Sobreira, N. Assessment of Incidental Findings in 232 Whole-Exome Sequences from the Baylor-Hopkins Center for Mendelian Genomics. Genet. Med. 2015, 17, 782–788. [Google Scholar] [CrossRef]
- Strauss, K.A.; Gonzaga-Jauregui, C.; Brigatti, K.W.; Williams, K.B.; King, A.K.; Van Hout, C.; Robinson, D.L.; Young, M.; Praveen, K.; Heaps, A.D.; et al. Genomic Diagnostics within a Medically Underserved Population: Efficacy and Implications. Genet. Med. 2018, 20, 31–41. [Google Scholar] [CrossRef]
- Jain, A.; Gandhi, S.; Koshy, R.; Scaria, V. Incidental and Clinically Actionable Genetic Variants in 1005 Whole Exomes and Genomes from Qatar. Mol. Genet. Genom. 2018, 293, 919–929. [Google Scholar] [CrossRef]
- Rego, S.; Dagan-Rosenfeld, O.; Zhou, W.; Reza Sailani, M.; Limcaoco, P.; Colbert, E.; Avina, M.; Wheeler, J.; Craig, C.; Salins, D.; et al. High-Frequency Actionable Pathogenic Exome Variants in an Average-Risk Cohort. Cold Spring Harb Mol Case Stud 2018, 4. [Google Scholar] [CrossRef]
- Tang, C.S. man; Dattani, S.; So, M. ting; Cherny, S.S.; Tam, P.K.H.; Sham, P.C.; Garcia-Barcelo, M.M. Actionable Secondary Findings from Whole-Genome Sequencing of 954 East Asians. Hum Genet 2018, 137, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Thompson, M.L.; Finnila, C.R.; Bowling, K.M.; Brothers, K.B.; Neu, M.B.; Amaral, M.D.; Hiatt, S.M.; East, K.M.; Gray, D.E.; Lawlor, J.M.J.; et al. Genomic Sequencing Identifies Secondary Findings in a Cohort of Parent Study Participants. Genet. Med. 2018, 20, 1635–1643. [Google Scholar] [CrossRef] [PubMed]
- Yehia, L.; Ni, Y.; Sesock, K.; Niazi, F.; Fletcher, B.; Chen, H.J.L.; LaFramboise, T.; Eng, C. Unexpected Cancer-Predisposition Gene Variants in Cowden Syndrome and Bannayan-Riley-Ruvalcaba Syndrome Patients without Underlying Germline PTEN Mutations. PLoS Genet 2018, 14. [Google Scholar] [CrossRef]
- Haer-Wigman, L.; van der Schoot, V.; Feenstra, I.; Vulto-van Silfhout, A.T.; Gilissen, C.; Brunner, H.G.; Vissers, L.E.L.M.; Yntema, H.G. 1 in 38 Individuals at Risk of a Dominant Medically Actionable Disease. Eur. J. Hum. Genet. 2019, 27, 325–330. [Google Scholar] [CrossRef]
- Ragan Hart, M.; Biesecker, B.B.; Blout, C.L.; Christensen, K.D.; Amendola, L.M.; Bergstrom, K.L.; Biswas, S.; Bowling, K.M.; Brothers, K.B.; Conlin, L.K.; et al. Secondary Findings from Clinical Genomic Sequencing: Prevalence, Patient Perspectives, Family History Assessment, and Health-Care Costs from a Multisite Study. Genet. Med. 2019, 21, 1100–1110. [Google Scholar] [CrossRef]
- Benson, K.A.; White, M.; Allen, N.M.; Byrne, S.; Carton, R.; Comerford, E.; Costello, D.; Doherty, C.; Dunleavey, B.; El-Naggar, H.; et al. A Comparison of Genomic Diagnostics in Adults and Children with Epilepsy and Comorbid Intellectual Disability. Eur. J. Hum. Genet. 2020, 28, 1066–1077. [Google Scholar] [CrossRef]
- Jalkh, N.; Mehawej, C.; Chouery, E. Actionable Exomic Secondary Findings in 280 Lebanese Participants. Front Genet 2020, 11. [Google Scholar] [CrossRef]
- Kuo, C.W.; Hwu, W.L.; Chien, Y.H.; Hsu, C.; Hung, M.Z.; Lin, I.L.; Lai, F.; Lee, N.C. Frequency and Spectrum of Actionable Pathogenic Secondary Findings in Taiwanese Exomes. Mol Genet Genomic Med 2020, 8. [Google Scholar] [CrossRef]
- Van Rooij, J.; Arp, P.; Broer, L.; Verlouw, J.; Van Rooij, F.; Kraaij, R.; Uitterlinden, A.; Verkerk, A.J.M.H. Reduced Penetrance of Pathogenic ACMG Variants in a Deeply Phenotyped Cohort Study and Evaluation of ClinVar Classification over Time. Genetics in Medicine 2020, 22. [Google Scholar] [CrossRef]
- Kwak, S.H.; Chae, J.; Choi, S.; Kim, M.J.; Choi, M.; Chae, J.H.; Cho, E.H.; Hwang, T.J.; Jang, S.S.; Kim, J. Il; et al. Findings of a 1303 Korean Whole-Exome Sequencing Study. Exp Mol Med 2017, 49. [Google Scholar] [CrossRef]
- Yamaguchi-Kabata, Y.; Yasuda, J.; Tanabe, O.; Suzuki, Y.; Kawame, H.; Fuse, N.; Nagasaki, M.; Kawai, Y.; Kojima, K.; Katsuoka, F.; et al. Evaluation of Reported Pathogenic Variants and Their Frequencies in a Japanese Population Based on a Whole-Genome Reference Panel of 2049 Individuals Article. J Hum Genet 2018, 63, 213–230. [Google Scholar] [CrossRef] [PubMed]
- Gordon, A.S.; Zouk, H.; Venner, E.; Eng, C.M.; Funke, B.H.; Amendola, L.M.; Carrell, D.S.; Chisholm, R.L.; Chung, W.K.; Denny, J.C.; et al. Frequency of Genomic Secondary Findings among 21,915 EMERGE Network Participants. Genet. Med. 2020, 22, 1470–1477. [Google Scholar] [CrossRef]
- Arslan Ateş, E.; Türkyilmaz, A.; Yıldırım, Ö.; Alavanda, C.; Polat, H.; Demir, Ş.; Çebi, A.H.; Geçkinli, B.B.; Güney, A.İ.; Ata, P.; et al. Secondary Findings in 622 Turkish Clinical Exome Sequencing Data. J Hum Genet 2021, 66, 1113–1119. [Google Scholar] [CrossRef]
- Elfatih, A.; Mifsud, B.; Syed, N.; Badii, R.; Mbarek, H.; Abbaszadeh, F.; Estivill, X.; Ismail, S.; Al-Muftah, W.; Badji, R.; et al. Actionable Genomic Variants in 6045 Participants from the Qatar Genome Program. Hum Mutat 2021, 42, 1584–1601. [Google Scholar] [CrossRef]
- van der Schoot, V.; Haer-Wigman, L.; Feenstra, I.; Tammer, F.; Oerlemans, A.J.M.; van Koolwijk, M.P.A.; van Agt, F.; Arens, Y.H.J.M.; Brunner, H.G.; Vissers, L.E.L.M.; et al. Lessons Learned from Unsolicited Findings in Clinical Exome Sequencing of 16,482 Individuals. Eur. J. Hum. Genet. 2022, 30, 170–177. [Google Scholar] [CrossRef]
- Rodríguez-Salgado, L.E.; Silva-Aldana, C.T.; Medina-Méndez, E.; Bareño-Silva, J.; Arcos-Burgos, M.; Silgado-Guzmán, D.F.; Restrepo, C.M. Frequency of Actionable Exomic Secondary Findings in 160 Colombian Patients: Impact in the Healthcare System. Gene 2022, 838. [Google Scholar] [CrossRef]
- Huang, Y.; Liu, B.; Shi, J.; Zhao, S.; Xu, K.; Sun, L.; Chen, N.; Tian, W.; Zhang, J.; Wu, N. Landscape of Secondary Findings in Chinese Population: A Practice of ACMG SF v3.0 List. 0 List. J Pers Med 2022, 12. [Google Scholar] [CrossRef]
- Martone, S.; Buonagura, A.T.; Marra, R.; Rosato, B.E.; Del Giudice, F.; Bonfiglio, F.; Capasso, M.; Iolascon, A.; Andolfo, I.; Russo, R. Clinical Exome-Based Panel Testing for Medically Actionable Secondary Findings in a Cohort of 383 Italian Participants. Front Genet 2022, 13. [Google Scholar] [CrossRef]
- Kasak, L.; Lillepea, K.; Nagirnaja, L.; Aston, K.I.; Schlegel, P.N.; Gonçalves, J.; Carvalho, F.; Moreno-Mendoza, D.; Almstrup, K.; Eisenberg, M.L.; et al. Actionable Secondary Findings Following Exome Sequencing of 836 Non-Obstructive Azoospermia Cases and Their Value in Patient Management. Hum. Reprod. 2022, 37, 1652–1663. [Google Scholar] [CrossRef]
- Hershberger, R.E.; Morales, A.; Siegfried, J.D. Clinical and Genetic Issues in Dilated Cardiomyopathy: A Review for Genetics Professionals. Genet. Med. 2010, 12, 655–667. [Google Scholar] [CrossRef]
- Maron, B.; Gardin, J.; Flack, J.; Gidding, SS.; Kurosaki, TT.; Bild DE Prevalence of Hypertrophic Cardiomyopathy in a General Population of Young Adults. Echocardiographic Analysis of 4111 Subjects in the CARDIA Study. Coronary Artery Risk Development in (Young) Adults. Circulation 1995, 92, 785–789. [Google Scholar] [CrossRef] [PubMed]
- Tabib, A.; Loire, R.; Chalabreysse, L.; Meyronnet, D.; Miras, A.; Malicier, D.; Thivolet, F.; Chevalier, P.; Bouvagnet, P. Circumstances of Death and Gross and Microscopic Observations in a Series of 200 Cases of Sudden Death Associated with Arrhythmogenic Right Ventricular Cardiomyopathy and/or Dysplasia. Circulation 2003, 108, 3000–3005. [Google Scholar] [CrossRef]
- Schwartz, P.J.; Stramba-Badiale, M.; Crotti, L.; Pedrazzini, M.; Besana, A.; Bosi, G.; Gabbarini, F.; Goulene, K.; Insolia, R.; Mannarino, S.; et al. Prevalence of the Congenital Long-QT Syndrome. Circulation 2009, 120, 1761–1767. [Google Scholar] [CrossRef]
- Akioyamen, L.E.; Genest, J.; Shan, S.D.; Reel, R.L.; Albaum, J.M.; Chu, A.; Tu, J.V. Estimating the Prevalence of Heterozygous Familial Hypercholesterolaemia: A Systematic Review and Meta-Analysis. BMJ Open 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Dietz, H. FBN1-Related Marfan Syndrome - GeneReviews® - NCBI Bookshelf Available online:. Available online: https://www.ncbi.nlm.nih.gov/books/NBK1335/ (accessed on 21 October 2024).
- Milewicz, D. Heritable Thoracic Aortic Disease Overview - GeneReviews® - NCBI Bookshelf Available online:. Available online: https://www.ncbi.nlm.nih.gov/books/NBK1120/ (accessed on 21 October 2024).
- Loeys-Dietz Syndrome - GeneReviews® - NCBI Bookshelf Available online:. Available online: https://www.ncbi.nlm.nih.gov/books/NBK1133/ (accessed on 21 October 2024).
- Byers, P. Vascular Ehlers-Danlos Syndrome - GeneReviews® - NCBI Bookshelf Available online:. Available online: https://www.ncbi.nlm.nih.gov/books/NBK1494/ (accessed on 21 October 2024).
- Ponder, B.; Pharoah, P.D.P.; Ponder, B.A.J.; Lipscombe, J.M.; Basham, V.; Gregory, J.; Gayther, S.; Dunning, A. Prevalence and Penetrance of BRCA1 and BRCA2 Mutations in a Population-Based Series of Breast Cancer Cases. Br J Cancer 2000, 83, 1301–1308. [Google Scholar] [CrossRef]
- Teugels, E.; De Brakeleer, S.; Goelen, G.; Lissens, W.; Sermijn, E.; De Grève, J. De Novo Alu Element Insertions Targeted to a Sequence Common to the BRCA1 and BRCA2 Genes. Hum Mutat 2005, 26, 284. [Google Scholar] [CrossRef]
- Win, A.K.; Jenkins, M.A.; Dowty, J.G.; Antoniou, A.C.; Lee, A.; Giles, G.G.; Buchanan, D.D.; Clendenning, M.; Rosty, C.; Ahnen, D.J.; et al. Prevalence and Penetrance of Major Genes and Polygenes for Colorectal Cancer. Cancer Epidemiol. Biomark. Prev. 2017, 26, 404–412. [Google Scholar] [CrossRef]
- Andrade, C. A Peculiar Form of Peripheral Neuropathy; Familiar Atypical Generalized Amyloidosis with Special Involvement of the Peripheral Nerves. Brain 1952, 75, 408–427. [Google Scholar] [CrossRef]
- Sousa, A.; Coelho, T.; Barros, J.; Sequeiros, J. Genetic Epidemiology of Familial Amyloidotic Polyneuropathy (FAP)-Type I in Póvoa Do Varzim and Vila Do Conde (North of Portugal). Am. J. Med. Genet. (Neuropsychiatr. Genet. ) 1995, 60, 512–521. [Google Scholar] [CrossRef]
- Inês, M.; Coelho, T.; Conceição, I.; Duarte-Ramos, F.; de Carvalho, M.; Costa, J. Epidemiology of Transthyretin Familial Amyloid Polyneuropathy in Portugal: A Nationwide Study. Neuroendocrinology 2018, 51, 177–182. [Google Scholar] [CrossRef]
- Coelho, T.; Maurer, M.S.; Suhr, O.B. THAOS - The Transthyretin Amyloidosis Outcomes Survey: Initial Report on Clinical Manifestations in Patients with Hereditary and Wild-Type Transthyretin Amyloidosis. Curr Med Res Opin 2013, 29, 63–76. [Google Scholar] [CrossRef]
- Mungunsukh, O.; Deuster, P.; Muldoon, S.; O’Connor, F.; Sambuughin, N. Estimating Prevalence of Malignant Hyperthermia Susceptibility through Population Genomics Data. Br J Anaesth 2019, 123, e461–e463. [Google Scholar] [PubMed]
- Fridriksdottir, R.; Jonsson, A.J.; Jensson, B.O.; Sverrisson, K.O.; Arnadottir, G.A.; Skarphedinsdottir, S.J.; Katrinardottir, H.; Snaebjornsdottir, S.; Jonsson, H.; Eiriksson, O.; et al. Sequence Variants in Malignant Hyperthermia Genes in Iceland: Classification and Actionable Findings in a Population Database. Eur. J. Hum. Genet. 2021, 29, 1819–1824. [Google Scholar] [CrossRef]
- Spada, M.; Pagliardini, S.; Yasuda, M.; Tukel, T.; Thiagarajan, G.; Sakuraba, H.; Ponzone, A.; Desnick, R.J. High Incidence of Later-Onset Fabry Disease Revealed by Newborn Screening. Am J Hum Genet 2006, 79, 31–40. [Google Scholar] [CrossRef]
- Sawada, T.; Kido, J.; Yoshida, S.; Sugawara, K.; Momosaki, K.; Inoue, T.; Tajima, G.; Sawada, H.; Mastumoto, S.; Endo, F.; et al. Newborn Screening for Fabry Disease in the Western Region of Japan. Mol Genet Metab Rep 2020, 22. [Google Scholar] [CrossRef]
- Burton, B.K.; Charrow, J.; Hoganson, G.E.; Waggoner, D.; Tinkle, B.; Braddock, S.R.; Schneider, M.; Grange, D.K.; Nash, C.; Shryock, H.; et al. Newborn Screening for Lysosomal Storage Disorders in Illinois: The Initial 15-Month Experience. J. Pediatr. 2017, 190, 130–135. [Google Scholar] [CrossRef] [PubMed]
- Hwu, W.L.; Chien, Y.H.; Lee, N.C.; Chiang, S.C.; Dobrovolny, R.; Huang, A.C.; Yeh, H.Y.; Chao, M.C.; Lin, S.J.; Kitagawa, T.; et al. Newborn Screening for Fabry Disease in Taiwan Reveals a High Incidence of the Later-Onset GLA Mutation c.936+919G>A (IVS4+919G>A). Hum Mutat 2009, 30, 1397–1405. [Google Scholar] [CrossRef] [PubMed]
- Gilchrist, M.; Casanova, F.; Tyrrell, J.S.; Cannon, S.; Wood, A.R.; Fife, N.; Young, K.; Oram, R.A.; Weedon, M.N. Prevalence of Fabry Disease-Causing Variants in the UK Biobank. J Med Genet 2023, 60, 391–396. [Google Scholar] [CrossRef]
- Azevedo, O.; Gal, A.; Faria, R.; Gaspar, P.; Miltenberger-Miltenyi, G.; Gago, M.F.; Dias, F.; Martins, A.; Rodrigues, J.; Reimão, P.; et al. Founder Effect of Fabry Disease Due to p.F113L Mutation: Clinical Profile of a Late-Onset Phenotype. Mol Genet Metab 2020, 129, 150–160. [Google Scholar] [CrossRef]
- Mehta, A.; Ricci, R.; Widmer, U.; Dehout, F.; Garcia De Lorenzo, A.; Kampmann, C.; Linhart, A.; Sunder-Plassmann, G.; Ries, M.; Beck, M. Fabry Disease Defined: Baseline Clinical Manifestations of 366 Patients in the Fabry Outcome Survey. Eur J Clin Invest 2004, 34, 236–242. [Google Scholar] [CrossRef]
- Hanson, E.H.; Imperatore, G.; Burke, W. HFE Gene and Hereditary Hemochromatosis: A HuGE Review. Human Genome Epidemiology. Am J Epidemiol 2001, 154, 193–206. [Google Scholar] [CrossRef] [PubMed]
- Phatak, P.D.; Sham, R.L.; Raubertas, R.F.; Dunnigan, K.; O’leary, T.; Braggins, C.; Cappuccio, J.D. Prevalence of Hereditary Hemochromatosis in 16031 Primary Care Patients. 1998, 129, 954–961. [CrossRef]
- Adams, P.C.; Reboussin, D.M.; Barton, J.C.; Mclaren, C.E.; Eckfeldt, J.H.; Mclaren, G.D.; Dawkins, F.W.; Acton, R.T.; Harris, E.L.; Gordeuk, V.R.; et al. Hemochromatosis and Iron-Overload Screening in a Racially Diverse Population. New Engl. J. Med. 2005, 352, 1769–1778. [Google Scholar] [CrossRef] [PubMed]
- Park, R.; McCabe, P.; Fell, G.; Russell, R. Wilson’s Disease in Scotland. Gut 1991, 32, 1541–1545. [Google Scholar] [CrossRef]
- Coffey, A.J.; Durkie, M.; Hague, S.; McLay, K.; Emmerson, J.; Lo, C.; Klaffke, S.; Joyce, C.J.; Dhawan, A.; Hadzic, N.; et al. A Genetic Study of Wilson’s Disease in the United Kingdom. Brain 2013, 136, 1476–1487. [Google Scholar] [CrossRef] [PubMed]
- Pinto, R.; Caseiro, C.; Lemos, M.; Lopes, L.; Fontes, A.; Ribeiro, H.; Pinto, E.; Silva, E.; Rocha, S.; Marcão, A.; et al. Prevalence of Lysosomal Storage Diseases in Portugal. Eur. J. Hum. Genet. 2004, 12, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Wolf, B. Worldwide Survey of Neonatal Screening for Biotinidase Deficiency. J Inherit Metab Dis 1991, 14, 923–927. [Google Scholar] [CrossRef] [PubMed]
- Romero, S.; Biggio, J.R.; Saller, D.N.; Giardine, R. Committee Opinion Number 432; 2017; Vol. 129;
- Board of Directors, A. The Use of ACMG Secondary Findings Recommendations for General Population Screening: A Policy Statement of the American College of Medical Genetics and Genomics (ACMG). Genet. Med. 2019, 21, 1467–1468. [Google Scholar] [CrossRef]
- Valente, S.; Ribeiro, M.; Schnur, J.; Alves, F.; Moniz, N.; Seelow, D.; Freixo, J.P.; Silva, P.F.; Oliveira, J. Analysis of Regions of Homozygosity: Revisited Through New Bioinformatic Approaches. BioMedInformatics 2024, 4, 2374–2399. [Google Scholar] [CrossRef]
- População Residente Por Município Segundo Os Censos | Pordata. Available online: Https://Www.Pordata.Pt/Municipios/Populacao+residente+segundo+os+censos+total+e+por+sexo-17 (accessed on 13 June 2023).
- Richards, S.; Aziz, N.; Bale, S.; Bick, D.; Das, S.; Gastier-Foster, J.; Grody, W.W.; Hegde, M.; Lyon, E.; Spector, E.; et al. Standards and Guidelines for the Interpretation of Sequence Variants: A Joint Consensus Recommendation of the American College of Medical Genetics and Genomics and the Association for Molecular Pathology. Genet. Med. 2015, 17, 405–424. [Google Scholar] [CrossRef]
- Best Practice Guidelines - The Association for Clinical Genomic Science Available online:. Available online: https://www.acgs.uk.com/quality/best-practice-guidelines/ (accessed on 21 October 2024).
- Criteria Specification Registry Available online:. Available online: https://cspec.genome.network/cspec/ui/svi/ (accessed on 21 October 2024).
- CanVIG-UK Consensus Specifications | CanGene-CanVar Available online:. Available online: https://www.cangene-canvaruk.org/canvig-uk-guidance (accessed on 21 October 2024).
- Hayesmoore, J.B.; Bhuiyan, Z.A.; Coviello, D.A.; du Sart, D.; Edwards, M.; Iascone, M.; Morris-Rosendahl, D.J.; Sheils, K.; van Slegtenhorst, M.; Thomson, K.L. EMQN: Recommendations for Genetic Testing in Inherited Cardiomyopathies and Arrhythmias. Eur. J. Hum. Genet. 2023, 31, 1003–1009. [Google Scholar] [PubMed]
Figure 1.
Flowchart of the variant filtering and curation process. RD: read depth; GQ: genotype quality; MAF: minor allele frequency; PAT: pathogenic; L-PAT: likely-pathogenic; BEN: benign; L-BEN: likely-benign.
Figure 1.
Flowchart of the variant filtering and curation process. RD: read depth; GQ: genotype quality; MAF: minor allele frequency; PAT: pathogenic; L-PAT: likely-pathogenic; BEN: benign; L-BEN: likely-benign.
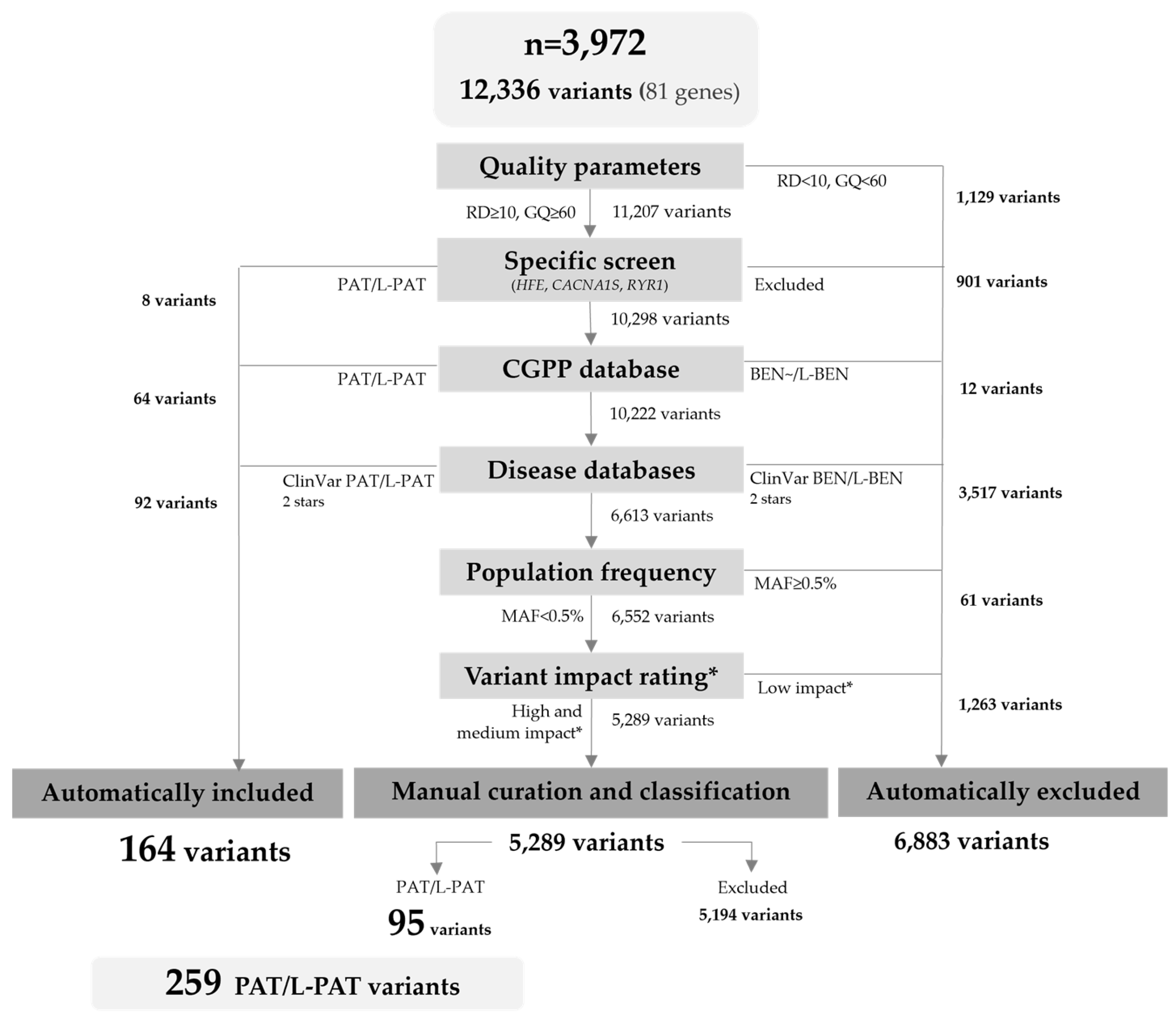
Figure 2.
Total medically actionable findings and non-medically actionable findings (carrier status) present in our cohort of 3,972 persons, presumed to be representative of the Portuguese population.
Figure 2.
Total medically actionable findings and non-medically actionable findings (carrier status) present in our cohort of 3,972 persons, presumed to be representative of the Portuguese population.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



