
Submitted:
13 March 2025
Posted:
14 March 2025
You are already at the latest version
Abstract
Insects demonstrate remarkable adaptability to extremely cold environments, a critical survival trait contributing to their evolutionary success and ecological distribution. This manuscript explores the mechanisms of insect cold hardiness, focusing on cryoprotectants—biochemical substances that mitigate damage caused by freezing temperatures. Important cryoprotectants, such as glycerol, trehalose, and amino acids, are discussed in detail, highlighting their mechanisms of action, which include colligative and non-colligative effects, membrane stabilization, ice nucleation inhibition, and osmotic regulation. Case studies of species-specific adaptations illustrate the diversity of strategies that insects employ, ranging from the accumulation of a single cryoprotectant to complex combinations that enhance resilience. The manuscript also addresses the relationship between cryoprotectants and other adaptations, such as antifreeze proteins and ice-nucleating proteins, emphasizing the integrated nature of insect survival strategies. By linking these findings to broader ecological and agricultural contexts, the manuscript underscores the importance of understanding insect cold hardiness in the face of climate change. Trehalose and glycerol are the predominant cryoprotectants. Among insect orders, Coleoptera exhibits the highest diversity of both cryoprotectants and species, reflecting their broad range of ecological strategies. Lepidoptera demonstrates notable diversity in cryoprotectants, which align with their adaptations for overwintering at different life stages, including larvae and pupae. Future research directions are proposed to uncover regulatory pathways, explore lesser-known cryoprotectants, and apply these insights to manage insect populations in dynamic environments. This review provides a comprehensive framework for understanding insect cold hardiness's biochemical, physiological, and ecological dimensions, with implications for both fundamental biology and applied sciences.
Keywords:
cold tolerance
; cold hardiness
; diapause
; Cryobiology
; cryoprotectants
1. Introduction
Insects are one of the most diverse groups of organisms on Earth, with over a million species described. Their success across a wide range of habitats, including extreme environments, is largely due to their exceptional adaptability. Insects are highly responsive to ambient temperature across multiple biological levels, from cellular processes to population dynamics. Temperature fundamentally influences cellular activities by regulating chemical reactions and other biological functions critical to insect survival and behavior. The effects of ambient temperature on insects are multifaceted and can range from immediate impacts on individual survival, such as cold-shock injuries caused by sudden drops in temperature, to longer-term effects that shape the distribution, population dynamics, and evolutionary history of species [1,2,3]. To fully understand how temperature affects insect survival at both physiological and ecological scales, it is essential to examine their capacity to withstand low temperatures, commonly referred to as cold hardiness [4]. Cold tolerance is a crucial survival trait that enables insects to tolerate freezing or sub-zero temperatures, a resilience often achieved through a combination of physiological and biochemical adaptations. These adaptations include the production of cryoprotectants, which protect insect tissues from damage during freezing and thawing cycles. Consequently, the development of cold tolerance is a key factor influencing not only individual insect survival but also broader patterns of reproduction and population viability in response to environmental temperature fluctuations [5,6,7,8]. Cold tolerance and cold hardiness are two related but distinct concepts in animal adaptations to cold environments. While cold tolerance and cold hardiness are often used interchangeably, there are subtle differences between the two. Cold tolerance is a common adaptation that refers to an organism's overall ability to survive and function within a specific range of low temperatures. This concept encompasses a wide array of adaptations, including behavioral, physiological, and biochemical mechanisms, and does not necessarily imply the capacity to endure freezing conditions. In contrast, cold hardiness is a more precise term that describes an organism's ability to survive prolonged periods of freezing temperatures. This often involves specialized adaptations that either protect against ice formation or mitigate its damaging effects (Table 1). For example, in the pistachio fruit hull borer, Arimania comaroffi cold tolerance (survival rate) increased from October onward. In January with the mean supercooling point (SCP) of -16.0 °C, approximately 39% of the pupae survived -15.0 °C/24h. This indicates that only a portion of the population is cold-hardy and can survive near their supercooling point (SCP) [9]. I would like to say that while this insect can tolerate cold temperatures during winter, cold hardiness is achieved in January when the ambient temperature and the SCP are at the lowest levels whereas, the highest level of cryoprotectants is recorded [10]. Similarly, in the grapevine moth, Lobesia botrana, while the overwintering pupae can tolerate winter conditions from July onward, it is only in February that the pupae reach their highest level of cold hardiness. In this month, at least a portion of the population survived temperatures close to their extended SCP. Interestingly, in this species, cold hardiness was achieved only through SCP depression and natural cold acclimation, rather than cryoprotectant accumulation [11]. However, the capacity for cold tolerance is influenced by various biotic factors such as the insect species, developmental stage, age, sex, nutritional and physiological status, diapause development, and genetic potential, as well as abiotic factors such as the season of the year, the extent of cold, and the duration of cold exposure [4,12,13,14,15]. Enhancing cold tolerance is a crucial strategy for insects to survive in harsh ambient temperatures and have a well-established physiological adaptation to enhance their cold tolerance. Cryoprotectant accumulation in insects is a complex process involving synthesizing and regulating various substances essential for freezing tolerance. The specific cryoprotectants and mechanisms can differ based on species, geography, and environmental conditions [16,17,18,19,20]. Understanding the biochemical pathways and regulatory mechanisms governing cryoprotectant production is essential for advancing our knowledge of insect physiology and ecology. Future research should focus on elucidating how environmental factors influence these adaptations and exploring other potential cryoprotective agents beyond polyols. The insights gained could also have applications in fields such as agriculture and conservation, where managing insect populations under changing climate conditions is increasingly critical. Cryoprotectants are vital for insect survival in cold environments, enabling them to withstand freezing temperatures through a combination of biochemical strategies that protect cellular integrity and enhance overall cold hardiness [21,22,23]. In this article, I explore the fascinating world of cryoprotectants and their indispensable role in insect cold hardiness. I delve into the mechanisms of insect cold hardiness, the types and functions of cryoprotectants, the adaptations and strategies employed by insects for cold tolerance, the environmental factors influencing cold hardiness, and the practical applications of cryoprotectants in insect management. Additionally, I discuss future research directions that hold immense potential for advancing our understanding of cryoprotectants and their interactions with insect cold hardiness.
2. Mode of Action of Cryoprotectants
Winter can be challenging for humans, but spare a thought for insects. These tiny creatures somehow survive freezing temperatures thanks to their secret weapon: cryoprotectants. Cryoprotectants are substances that play a vital role in protecting biological tissues from damage caused by ice crystal formation during freezing and thawing. By lowering the freezing point of water, these compounds prevent the formation of ice crystals within an organism's cells, a critical mechanism that enables insects to survive subzero temperatures. Without cryoprotectants, ice crystals can disrupt cellular structures, dehydrate cells, and cause cell lysis, leading to severe tissue damage or cell death [21,24,25]. Surviving freezing environments can be achieved through various mechanisms (Figure 1):
Colligative Effects (Concentration-Dependent) and Non-Colligative Cryoprotective Function
Colligative properties are characteristics of solutions that depend on the number of dissolved solute particles, regardless of their identity. Cryoprotectants use these properties to lower the freezing point of solutions, aiding in the preservation of biological tissues. By increasing solute concentration, cryoprotectants alter water's phase transitions, reducing ice formation at typical freezing temperatures, and binding water molecules limits the availability of free water for crystallization. These colligative effects lead to a lowered vapor pressure, elevated boiling point, depressed freezing point, and increased osmotic pressure. However, research shows that cryoprotectants also contribute to freeze tolerance through non-colligative mechanisms. They interact with biological structures, stabilizing cell membranes and proteins, and maintaining osmotic balance under cold stress. While colligative properties are important for cryoprotection, non-colligative effects, which arise from the chemical nature of the solute and its interactions with the solvent, also play a significant role in enhancing cellular stability and freeze resistance (Figure 2A,B). As the solute concentration increases, colder temperatures are required for water to freeze into ice. Additionally, cryoprotectants can interact with water molecules, diminishing their ability to form organized ice crystals [26,27,28,29]. Glycerol, a well-known cryoprotectant, works mainly through its colligative properties. Pytho depressus reduces body fluids' freezing point in freeze-tolerant beetles, enabling survival in colder temperatures. Its protective effectiveness increases with concentration, as higher levels lower the freezing point further [30]. However, key findings show that different cryoprotectants play specific roles in freeze tolerance through mechanisms beyond simple colligative effects. These compounds act in complex ways to protect cell membranes, and proteins, and maintain osmotic balance during cold stress [28,31]. For example, cryoprotectants such as trehalose and proline provide non-colligative functions like antioxidant activity and membrane stabilization. Still, their essential colligative roles, freezing point depression, and osmotic regulation are crucial for insect cold tolerance [31,32]. In the Arctic springtail, Megaphorura arctica, trehalose accumulation raises osmotic pressure, extracting water from cells and stabilizing membranes during extreme desiccation. This illustrates the relationship between colligative solute concentration and non-colligative membrane protection [32]. Small cryoprotectants like proline, trehalose, and myo-inositol accumulate in freeze-tolerant insects, such as Gryllus veletis, and improve survival during freezing by stabilizing cell components rather than solely lowering the freezing point [31]. Cryoprotectants play a vital role in enabling insects to survive sub-zero temperatures by protecting their cells and tissues from damage caused by ice crystal formation. They not only lower the freezing point of water but also preserve cellular integrity and functionality under freezing conditions [33,34,35]. For example, antifreeze proteins found in some insects bind to ice nuclei and inhibit the growth of ice crystals [36,37,38]. Additionally, cryoprotectants help adjust the osmotic balance, allowing cells to retain water and avoid dehydration, a common issue under cold stress. By managing internal water content, these compounds enable insects to maintain physiological processes even in extreme cold [39,40]. Another critical function of cryoprotectants is their role in mitigating oxidative damage caused by reactive oxygen species (ROS), which can accumulate during stress conditions such as freezing. Many cryoprotectants enhance antioxidant enzyme activity, reducing the harmful effects of ROS and further protecting insect cells [20,41]. Cryoprotectants also lower the supercooling point (SCP), allowing insects to remain in a liquid state at temperatures below freezing. This mechanism is especially significant for freeze-avoiding species that must prevent internal ice formation to survive. Insects employ additional strategies to enhance their supercooling ability, such as reducing their water content and increasing the concentration of cryoprotectants in their body fluids. These adaptations not only enhance their ability to supercool but also minimize potential damage from ice formation, ensuring survival in harsh, frozen environments [16,19,31,42,43,44].
3. Cryoprotectant Classification
Cryoprotectants can be classified into several categories based on their chemical composition, permeability, cellular situation, and mode of action (Figure 3).
3.1. Chemical Composition
Cryoprotectants can be classified into three main structural groups: polyols or polyhydric alcohols, low molecular-weight carbohydrates (such as glucose and trehalose), and free amino acids (such as proline) [45,46,47,48]. Polyols (such as glycerol, sorbitol, and mannitol), organic compounds containing multiple hydroxyl groups (−OH), are highly soluble and non-toxic sugar alcohols that can stabilize the functional state of proteins. This helps to prevent the denaturation of the native structures of proteins that may occur as a result of harsh ambient temperature or freezing [25,49].
3.2. Permeability and Cellular Situation
Intracellular cryoprotectants (penetrating cryoprotectants) are specifically designed to protect the intracellular components of cells during freezing. They can penetrate the cell membrane and protect the cellular structures from damage. Examples include dimethyl sulfoxide (DMSO), ethylene glycol, and glycerol. The anti-freeze properties of these substances are achieved by forming strong hydrogen bonds with water. These bonds compete with the hydrogen bonds that typically connect water molecules, contributing to the substance's ability to lower the freezing point of water [50]. Extracellular cryoprotectants (non-penetrating cryoprotectants) primarily protect the extracellular environment and are not capable of penetrating the cell membrane. Non-permeating cryoprotectants affect the osmolality of the extracellular environment, which regulates osmosis during both dehydration and rehydration processes [51,52]. They work by reducing the freezing point of the solution, preventing ice crystal formation. Examples include sugars like trehalose, sucrose, and glucose [53,54,55].
3.3. Solution Property
The impact of cryoprotectants may be attributed to their colligative or non-colligative properties. Low-molecular-weight carbohydrates, sugar alcohol, and amino acids are hypothesized to act as cryoprotectants colligatively or non-colligatively [31]. Most of the studies indicate the colligative effects of the cryoprotectants but a few studies show the non-colligative effects of cryoprotectants on cold tolerance enhancement of certain insect species. For example, the cold-acclimated spring field cricket, Gryllus veletisare is freeze-tolerant and can accumulate myo-inositol, proline, and trehalose in their hemolymph and fat body but no cryoprotectant, either alone or in combination, can provide freeze tolerance to non-acclimated, freeze-intolerant specimens. The differential impact of each cryoprotectant on survival in the frozen state suggests that small cryoprotectants are not interchangeable and likely function non-colligatively in this freeze-tolerant species [31]. The red firebug, Pyrrhocoris apterus (L.) (Heteroptera: Pyrrhocoridae) exhibits relatively low concentrations of polyols such as ribitol, suggesting that they may not serve as colligative cryoprotectants in this species [56,57]. It's crucial to understand that these classifications are not mutually exclusive. A single cryoprotectant can exhibit properties from multiple categories. For instance, glycerol is both extracellular and contributes to colligative properties. Trehalose is intracellular, contributes to colligative properties, and is an LMWC.
4. Predominant Cryoprotectants of Insects
The predominant classes of cryoprotectants include polyols, low-molecular-weight carbohydrates, and amino acids (Figure 4).
4.1. Polyols
Polyols are the most common and abundant cryoprotectants in insects. Glycerol, sorbitol, mannitol, and myo-inositol are identified as putative cryoprotectants in some insects
4.1.1. Mode of Action of Glycerol as a Cryoprotectant
Glycerol, a triol, is a hygroscopic, non-toxic, and viscous liquid with moderate solubility in water. As a non-ionic polyol, it is critical in protecting cells from freezing damage by inhibiting intracellular ice formation. This is achieved through its ability to form strong hydrogen bonds with water molecules, thereby disrupting water-water hydrogen bonding that would otherwise facilitate ice formation [23,58]. Glycerol's permeability across cellular membranes further enhances its cryoprotective properties. As a permeating cryoprotectant, it can regulate the rate and extent of cellular dehydration during freezing-induced membrane phase transitions, stabilizing biomolecules in their native states [59]. This stabilization is crucial for mitigating the structural and functional damage caused by freeze-thaw cycles (Figure 5). The cryoprotective effects of glycerol are also attributed to its colligative properties, which lower the freezing point of solutions. In freeze-tolerant organisms, the accumulation of high glycerol concentrations allows survival at lower temperatures [30]. Additionally, glycerol modulates the diffusivity of water near lipid bilayers, maintaining the hydration layer around cellular membranes and preserving membrane integrity during freezing [60]. It also influences lipid membrane phase behavior, stabilizing the membranes to prevent phase transitions that could result in cellular damage [61]. The accumulation of glycerol as a cold-tolerance strategy is widespread among insect species, highlighting its importance in enhancing survival under extreme temperatures (Table 2).
4.1.2. Mode of Action of Trehalose as a Cryoprotectant
Trehalose, a multifunctional cryoprotectant, is a non-reducing disaccharide composed of two glycolytic intermediates: glucose-1-phosphate and glucose-6-phosphate. It is the primary circulating sugar in insect hemolymph, with its levels influenced by ambient temperature, physiological status, and nutrition. Trehalose plays several vital roles in insect physiology, including: serving as the primary energy source for insects, acting as a cryoprotectant to enhance cold tolerance, stabilizing proteins and membranes under stress conditions, and playing a central role in feedback mechanisms that regulate energy demands [62,63,64,65]. Trehalose protects cells from freezing and dehydration through multiple biochemical mechanisms (Figure 6). It stabilizes proteins and cellular membranes by forming a protective layer around them, which is particularly critical during freezing. This protective layer prevents protein denaturation and helps maintain protein functionality. Trehalose effectively slows the movement of surrounding water molecules, forming a "cage" that inhibits the crystallization of hydration water—essential for maintaining protein stability [66,67]. In addition to stabilizing proteins, trehalose disrupts the hydrogen bonding network among water molecules, inhibiting ice formation in body fluids. This action lowers the freezing point and reduces ice nucleation activity. At higher concentrations, trehalose significantly suppresses ice growth compared to other sugars, such as sucrose, enhancing its effectiveness as a cryoprotectant. Trehalose also plays a critical role in protecting cells from dehydration. By retaining water within cells, it prevents cellular dehydration and osmotic stress. This property is particularly important for freeze-tolerant insects, which face the dual challenges of freezing temperatures and dehydration [68,69]. In freeze-tolerant insects, trehalose complements antifreeze proteins (AFPs) to enhance cold survival. While AFPs lower the freezing temperature and prevent ice growth, trehalose inhibits ice crystallization within the hemolymph. This synergistic effect allows insects to withstand extreme sub-zero temperatures more effectively [70,71]. During cold acclimation, insects upregulate their metabolism to synthesize trehalose, increasing the activity of trehalose-synthesizing enzymes. This leads to elevated trehalose levels in hemolymph and tissues, providing both an immediate energy source and enhanced cold tolerance in response to lower temperatures [71,72]. Table 2 presents insects that accumulate trehalose under cold conditions.
4.1.3. Mode of Action of Glucose as a Cryoprotectant
Glucose, the most abundant monosaccharide, serves as the primary fuel source for cells and plays a vital role in maintaining cellular function and stability. Its metabolism is tightly regulated through multiple enzymatic pathways, ensuring a balance between energy production and cellular homeostasis. Beyond its role as an energy source, glucose contributes to enhancing cell membrane stability and integrity, particularly under freezing conditions (Figure 7) [73,74]. During freezing, glucose aids in maintaining osmotic balance within cells by increasing the concentration of solutes in the hemolymph (insect blood). This elevation of solute concentration lowers the freezing point of body fluids via colligative properties, a mechanism especially critical for freeze-avoiding insects. By allowing body fluids to remain supercooled, glucose helps prevent ice formation and the associated cellular damage [31,34,75,76]. Glucose also stabilizes cell membranes against the physical stresses of freezing. By preventing the formation of disruptive ice crystals, glucose helps maintain membrane integrity and preserves cellular structure and function. This stabilization is essential for cell viability under sub-zero temperatures. Furthermore, glucose protects proteins from denaturation during freezing. It forms a protective hydration shell around proteins, helping to maintain their structure and function under stress. This effect is particularly important for enzymes and other proteins crucial to metabolic processes in cold environments [31,34]. Insects frequently rely on glucose as an energy source during cold stress. When other energy reserves, such as glycogen, are depleted due to low temperatures, the metabolism of glucose provides the energy needed to sustain vital cellular processes. In some insect species, glucose works synergistically with antifreeze proteins (AFPs) to enhance cold tolerance. While AFPs inhibit ice crystal growth and lower freezing points, glucose supports overall cellular health and function, further increasing the insect's ability to survive freezing conditions [76,77]. However, glucose plays a less prominent role as a direct cryoprotectant in insects compared to trehalose or glycerol. This limitation arises from several factors. Glucose’s lower molecular weight results in a higher number of molecules at a given concentration, but each glucose molecule exerts a smaller impact on colligative properties and macromolecular stabilization than larger cryoprotectants. Additionally, glucose contains fewer hydroxyl (-OH) groups than trehalose or polyols like glycerol. These hydroxyl groups are crucial for forming hydrogen bonds with water, membranes, and proteins, and their reduced number in glucose limits its ability to replace water and stabilize biological structures. Although glucose can accumulate during cold acclimation and provides some cryoprotection, its effectiveness is constrained by its limited structural properties. Excessive glucose concentrations can disrupt cellular metabolism and lead to adverse effects, which is why insects do not accumulate glucose to the same high levels as glycerol or trehalose. Nevertheless, glucose remains indispensable as a primary energy source and a secondary contributor to cold tolerance mechanisms in insects [78].
4.1.4. Mode of Action of Myo-Inositol as a Cryoprotectant
Myo-inositol, a cyclic sugar-alcohol with six hydroxyl groups (cyclohexanehexol), is used to cope with low-temperature stress through various physiological and biochemical mechanisms. An enzyme called L-myo-inositol-1-phosphate synthase (MIPS) plays a crucial role in this process by catalyzing the conversion of D-glucose 6 phosphate to L-myo-inositol-1-phosphate. Myo-inositol functions as a cryoprotectant through several key mechanisms. It stabilizes the osmotic balance between intracellular and extracellular environments, preventing excessive dehydration and intracellular ice formation. It also inhibits ice nucleation and growth, fostering a vitrified state during cooling to preserve cellular structure. Additionally, myo-inositol protects membranes by maintaining their fluidity and functionality at lower temperatures, essential for sustaining cellular processes. [79,80]. These mechanisms work together to improve the ability of insects to survive in extremely cold conditions, emphasizing the importance of myo-inositol for their physiological adaptations to winter environments [31,79]. In Drosophila montana, a dramatic increase in myo-inositol concentrations surged up to 400-fold during winter, highlighting its potential role in the insect's overwintering strategy [79]. Overwintering adults of the lady beetle, Harmonia axyridis, accumulate significant myo-inositol to enhance cold tolerance [81].
4.1.5. Mode of Action of Sorbitol as a Cryoprotectant
Sorbitol is a type of polyol or sugar alcohol that is synthesized from glucose 6-phosphate through the action of two enzymes, sorbitol-6-phosphate dehydrogenase (S6PDH) and sorbitol-6-phosphate phosphatase (S6PP). The synthesis of sorbitol in insects, such as the Silverleaf whitefly, Bemisia argentifolii, involves reducing fructose with the aid of a novel NADPH-dependent ketose reductase. This enzyme plays a vital role in facilitating sorbitol accumulation, particularly under stress conditions [82]. Sorbitol enhances the cold tolerance of insects through several physiological mechanisms. It acts as a cryoprotectant, stabilizing cellular structures during freezing conditions. Additionally, sorbitol helps maintain osmotic balance and ion homeostasis when insects are exposed to cold temperatures. These combined effects are crucial for the physiological adaptation of insects to winter environments [25,83,84]. Cold-acclimated insects, such as Drosophila melanogaster, exhibit improved cold tolerance associated with the accumulation of organic osmolytes like sorbitol. [85] This accumulation helps mitigate the effects of hemolymph hyperkalemia, which is characterized by elevated potassium levels that can disrupt cellular function [85]. Furthermore, sorbitol levels increase during the cold acclimation process as part of a broader biochemical response that involves changes in other cryoprotectants and proteins, ultimately enhancing the overall cold tolerance of the insect. Thus, sorbitol plays a crucial role in providing direct cellular protection and facilitating broader physiological adaptations to cold stress [19,86]. Adult Upis ceramboides beetles can withstand extreme cold, tolerating freezing temperatures as low as -60 °C in winter, aided by the accumulation of cryoprotective polyols sorbitol and threitol. Threitol, in particular, is unique to these beetles and is synthesized from erythrose 4-phosphate through a series of enzymatic processes, ultimately producing high concentrations of threitol without significant levels of erythritol [83]. In the red flour beetle, Tribolium castaneum, sorbitol levels increase under cold acclimation, which correlates with improved cold hardiness and survival rates [87]. The Chinese white pine beetle, Dendroctonus armandi, demonstrates that higher levels of sorbitol are associated with lower supercooling points, suggesting that sorbitol plays a crucial role in reducing the freezing point of the beetle's body fluids [88]. However, the differences in sorbitol accumulation among various species and geographic locations indicate its important role in enhancing cold tolerance in insects.
4.1.6. Mode of Action of Fructose as a Cryoprotectant
Fructose is a hexose monosaccharide with multiple hydroxyl groups. It is primarily found in fruits. Fructose acts as a precursor to cryoprotectants and contributes to metabolic pathways that enable survival in low temperatures. While fructose aids in insect cold tolerance through colligative functions, it is less important as a primary cryoprotectant compared to other substances like glycerol or trehalose. It acts as an effective cryoprotectant by stabilizing cellular structures, regulating osmotic pressure, and working in conjunction with other protective molecules [25,31].
For example, diapausing eggs of Gomphocerus sibiricus exhibited significantly higher levels of fructose, glycerol, and sorbitol compared to both the early embryogenesis stage and the post-diapause development stage [89].
4.2. Amino Acids as Cryoprotectants
Amino acids serve an important role as insect’s cryoprotectants, particularly in improving their cold tolerance and survival during freezing conditions. They can help by lowering the freezing point, stabilizing cellular structures, and working together with other cryoprotectants to enhance survival in extremely low temperatures. The primary amino acids that function as cryoprotectants are glutamine, proline, and asparagine [34,90,91]. Two key protein classes that aid insect adaptation to cold environments are ice nucleating proteins (INPs) and antifreeze proteins (AFPs). Both are essential for cold tolerance but operate through different mechanisms.
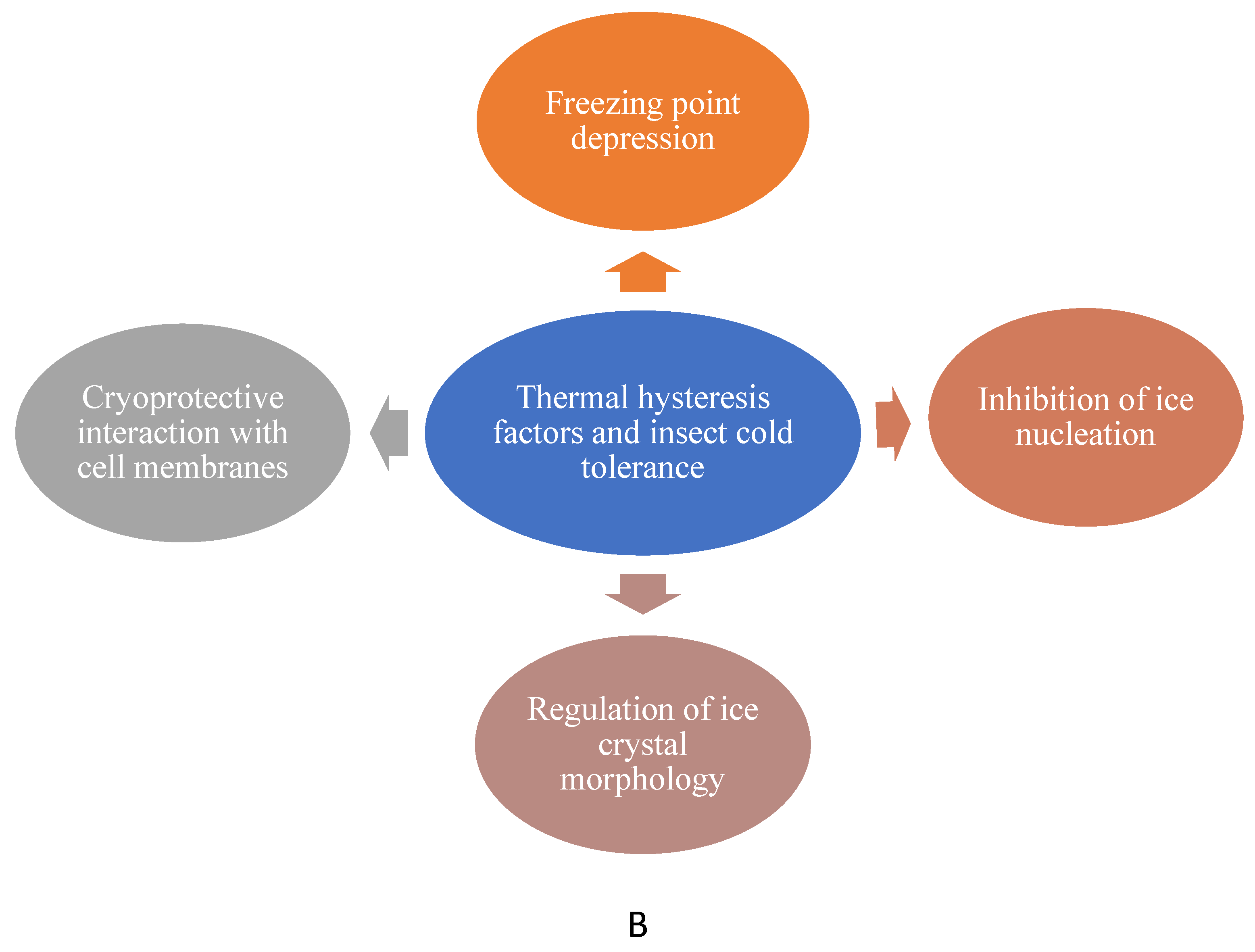
4.3. Thermal Hysteresis Factors
Thermal hysteresis factors, also known as antifreeze proteins (AFPs) or antifreeze glycoproteins (AFGPs), are molecules that non-colligatively lower the freezing point of solutions. Their impact on freezing point depression exceeds expectations based solely on concentration. This unique characteristic is referred to as "thermal hysteresis," highlighting the difference between the freezing and melting points in solutions containing these factors (Figure 3) [92,93]). Thermal hysteresis factors play a critical role in stabilizing supercooled states and preventing ice crystal growth during freezing and thawing processes, thereby protecting cellular integrity [20]. These proteins bind to the surface of ice crystals and alter their growth patterns, effectively halting further crystallization [94,95]. This function is essential for stabilizing cellular structures, as the formation of large ice crystals can rupture cell membranes, leading to irreparable damage [96,97]. By suppressing ice crystal growth, THFs not only preserve cellular architecture but also maintain the functionality of membranes under freezing conditions. Additionally, THFs prevent recrystallization—a harmful process where larger ice crystals grow at the expense of smaller ones. By inhibiting this phenomenon, they further reduce the risk of mechanical damage to cells during prolonged exposure to freezing temperatures. This dual role of stabilizing supercooled states and preventing ice crystal formation makes thermal hysteresis factors indispensable for cold-tolerant organisms [45,98]. For example, in the darkling beetle, Meracantha contracta, a protein improves survival at low temperatures by lowering the freezing and supercooling points of the larval hemolymph [99]. Additionally, winter spiders (Philodromus sp. and Clubiona sp.) have thermal hysteresis factors that may be lost if they acclimate to warmer temperatures [100].
4.4. Ice Nucleating Proteins (INPs)
Ice Nucleating Proteins (INPs) are specialized proteins that possess repetitive structural motifs that mimic ice crystal lattices, providing nucleation sites for ice formation to facilitate the controlled formation of ice crystals at relatively high subzero temperatures, often as high as -5°C to -10°C. This controlled extracellular ice formation is a critical adaptation for freeze-tolerant insects, enabling them to survive in freezing conditions by managing ice growth in a way that protects cellular integrity and minimizes mechanical damage to membranes and organelle [16,20]. The formation of ice in extracellular compartments concentrates cryoprotectants within cells, lowering the freezing point of the remaining fluids and stabilizing cellular structures [95,98]. This mechanism is particularly crucial for freeze-tolerant species that rely on the freezing of bodily fluids as a survival strategy. INPs not only regulate the location of ice formation but also reduce the risk of large, damaging ice crystals by promoting controlled and uniform ice growth. In this way, INPs act as both facilitators of extracellular freezing and as indirect cryoprotectants, ensuring the survival of insects in extremely cold environments (Table 2)[45,98]. For example, using ice nucleating agents, the Goldenrod gall fly, Eurosta solidaginis overwinters as a freeze-tolerant larva to avoid freezing damage in cold weather. By converting glycogen into antifreeze substances like sorbitol and glycerol, the larvae reduce body water content and, consequently, the formation of ice. Ice nucleating substances are essential because they help the larvae generate regulated ice, which protects their cells from harm and enables them to withstand extremely cold temperatures [101].
5. Insect Strategies for Accumulating Cryoprotectants
Based on the reviewed results, insects may employ one of three primary strategies for accumulating cryoprotectants to enhance their cold tolerance. The majority of species produce only a single cryoprotectant. However, dual or even multiple-component systems are also observed, with glycerol and trehalose being the most common combination (Figure 8) [31,102,103]. However, the effectiveness of cryoprotectant strategies in insects is influenced by various factors, including species-specific evolutionary adaptations, environmental conditions of their habitats, and the physiological state of the insects, such as their developmental stage and nutritional or dormancy status. Different species may deploy unique strategies to cope with cold environments, necessitating varied cryoprotectant types or concentrations for survival.
Figure 8.
Primary strategies for accumulating cryoprotectants to enhance their cold tolerance.
Figure 9.
Time-dependent cryoprotectant accumulation strategies of insects.
Figure 10.
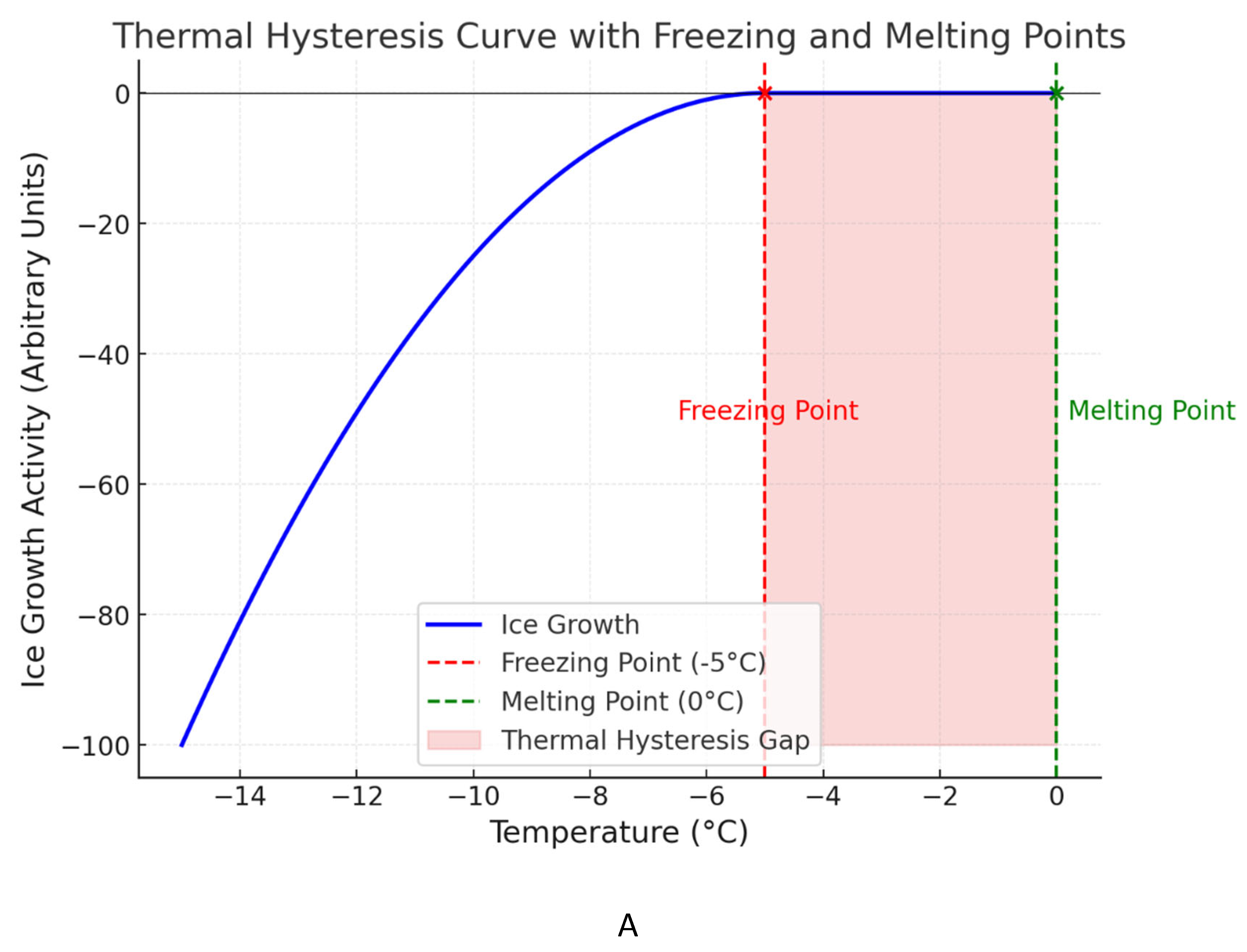
A. A graph depicts a thermal hysteresis curve, with the red dashed line representing the gap between the freezing and melting points of water due to antifreeze proteins or cryoprotectants. This gap illustrates the suppression of ice formation below 0°C, a crucial adaptation for insects surviving sub-zero temperatures. OpenAI. (2025). ChatGPT [Large language model]. https://chatgpt.com. B. Mode of action of thermal hysteresis factors in insect cold tolerance.
Figure 10.
A. A graph depicts a thermal hysteresis curve, with the red dashed line representing the gap between the freezing and melting points of water due to antifreeze proteins or cryoprotectants. This gap illustrates the suppression of ice formation below 0°C, a crucial adaptation for insects surviving sub-zero temperatures. OpenAI. (2025). ChatGPT [Large language model]. https://chatgpt.com. B. Mode of action of thermal hysteresis factors in insect cold tolerance.
5.1. Accumulation of a Single Cryoprotectant in Large Quantities
In this strategy, insects focus on synthesizing and storing a high concentration of a single type of cryoprotectant. This approach allows for maximized efficacy. A large quantity of one effective cryoprotectant, such as trehalose or glycerol, can significantly lower the freezing point of body fluids and stabilize cellular structures. Moreover, by relying on a single compound, metabolic pathways may be streamlined, allowing for efficient synthesis and utilization during cold stress. For instance, a 45-fold increase in myo-inositol level has been observed in overwintering adults of Hippodamia variegata (Goeze) [104,105]. Most insects that use a single cryoprotectant rely on glycerol. Of the 56 species studied, 19 species (34%) use glycerol exclusively (Table 2).
5.2. Accumulation of Two Primary Cryoprotectants at High Levels
This strategy involves the balanced accumulation of two key cryoprotectants, which can work synergistically to enhance cold tolerance. For example, trehalose and glycerol may be accumulated in similar amounts to provide both osmotic protection and membrane stabilization. The presence of two different cryoprotectants can offer a broader range of protective effects, improving overall resilience against freezing damage. Trehalose and myo-inositol in the overwintering larvae of the carob moth, Ectomyelois ceratoniae [106], and in overwintering adults of Acrosternum millierei [107], trehalose and glycerol in the variegated ladybug, Hippodamia variegat [108]
5.3. Combination of Different Cryoprotectants
Insects employing this strategy accumulate a diverse array of cryoprotectants, each contributing unique protective properties (Table 2). For example, Solenopsis invicta accumulates four cryoprotectants (glycerol, trehalose, glucose, myo-inositol), Acanthococcus lagerstroemiae utilizes five (mannitol, trehalose, myo-inositol, sorbitol, glucose), and Sesamia inferens also has five different cryoprotectants (trehalose, glycerol, glucose, fructose, myo-inositol). These species probably reside in severe or fluctuating settings necessitating various metabolic adaptations. Combining different cryoprotectants can offer several advantages. A mixture of low molecular weight sugars, polyols, and amino acids can create a more robust defense against cold stress by targeting various cellular vulnerabilities (synergistic effects). For instance, in the subarctic fly Chymomyza costata, a combination of cryoprotectants like proline and trehalose synergistically reduces ice formation and protects cells from freezing damage [90]. Moreover, this strategy allows insects to adapt their cryoprotectant profiles based on environmental conditions, potentially optimizing their survival strategies in fluctuating temperatures (flexibility in response). Multiple cryoprotectants offer non-colligative protection, providing specific roles that do not depend solely on their concentration. In the spring field cricket Gryllus veletis, different cryoprotectants such as myo-inositol, proline, and trehalose uniquely influence survival during freezing, showing they function in distinct ways [31]. The combination of cryoprotectants helps insects adapt to varying environmental conditions, as demonstrated in the aphid Cinara tujafilina, where a complex sugar-polyol system supports survival at low temperatures [109]. Temperature changes can trigger metabolic adjustments that produce specific cryoprotectants. In Drosophila suzukii, cold acclimation leads to the accumulation of sugars and amino acids, suggesting a physiological adaptation to cold stress [102]. By either focusing on a single cryoprotectant or employing a combination of multiple compounds, insects can effectively enhance their cold tolerance and improve their chances of survival in harsh environments. Understanding these strategies not only provides insights into insect physiology but also has broader implications for ecology and climate change research.
6. Accumulation of Cryoprotectants Over Time
Based on time, insects use various strategies to accumulate cryoprotectants. These strategies generally fall into three primary categories (Table 3). Each strategy is shaped by the insect's evolutionary history, habitat, and environmental pressures, highlighting the remarkable adaptability of these organisms to survive extreme conditions.
6.1. Rapid Accumulation Before Cold Exposure
Insects can detect environmental cues, such as shorter days and temperature drops, and quickly produce cryoprotectants in preparation for winter. This process often involves transcriptional upregulation of genes related to cryoprotectant synthesis (e.g., trehalose-6-phosphate synthase) and converting stored glycogen into cryoprotective compounds. This adaptive mechanism allows insects to prepare for sudden cold snaps, which are typical in temperate regions with predictable seasonal changes, enabling them to effectively prepare for imminent cold conditions [110,111]. The larvae of the leopard moth, Zeuzera pyrina, show marked increases in trehalose and sorbitol levels as winter nears, with trehalose concentrations rising about 11-fold and sorbitol levels increasing 7.5-fold in preparation for cold conditions [112]. Rapid cold-hardening enables insects to quickly enhance their cold tolerance within minutes to hours. While it was initially believed that RCH shared mechanisms with seasonal cold-hardening, recent research challenges this notion, suggesting that the two processes may operate via distinct mechanisms rather than overlapping ones [113,114]. For example, rapid cold hardening may improve the cold tolerance of Bactrocera dorsalis by elevating levels of proteins, glycerol, and trehalose while reducing water content [86]. Glycerol acts as a low molecular weight cryoprotectant in the overwintering eggs of the bamboo grasshopper, Ceracris kiangsu. Treatment with rapid cold-hardening and cold acclimation promotes its accumulation, thereby enhancing cold tolerance [115]. Research has shown that RCH occurs in non-diapause larvae and pharate adults of flesh flies (Sarcophaga crassipalpis), as well as in adult elm leaf beetles (Xanthogaleruca luteola) and milkweed bugs (Oncopeltus fasciatus). This mechanism protects against cold shock at temperatures above the supercooling point. The effectiveness of this response is associated with the accumulation of glycerol [116]. Adults of Leafminer, Liriomyza trifolii undergo RCH by adjusting fatty acid composition (e.g., increasing unsaturated fatty acids like palmitoleic acid) and accumulating glucose and trehalose within 4 hours of cold exposure. These biochemical shifts enhance membrane fluidity and energy reserves, enabling survival during sudden cold snaps [110].
6.2. Incremental Accumulation During Cold Exposure
Certain insects gradually increase their cryoprotectant levels as they experience prolonged exposure to colder temperatures by triggering metabolic pathways (e.g., upregulating enzymes like glycogen phosphorylase). This dynamic process allows them to adapt as conditions worsen. For instance, cold exposure may trigger enzymatic changes that convert stored sugars into protective compounds over time. Cold acclimation over several days can significantly boost cold tolerance in some species by increasing proteins, glycerol, and trehalose [102,113,115]. In the diapausing pupae of the pistachio fruit hull borer, Arimania comaroffi [9], and the diapausing adults of the Sunn pest, Eurygaster integriceps [91], the levels of cryoprotectants were lowest at the onset of diapause and increased steadily as diapause progressed. Cold acclimation of the red flour beetle, Tribolium castaneum, led to a significant increase in the contents of trehalose and myo-inositol, subsequently enhancing the cold tolerance of the beetles [87]. The arctic springtail (Megaphorura arctica) uses cryoprotective dehydration, losing up to 40% of its body water to avoid freezing. During this process, trehalose accumulates incrementally as osmotically active water is lost. Trehalose stabilizes membranes and proteins, acting synergistically with antioxidants like glutathione-S-transferase to protect cells during prolonged cold exposure [32,68].
6.3. Basal Accumulation Throughout Life
Some species maintain a constant level of cryoprotectants, regardless of environmental changes. This approach offers consistent protection against unexpected cold events but may come at the cost of energy and metabolic resources. These insects are often found in habitats with unpredictable or harsh climatic conditions. In the larvae of the pistachio twig borer, Kermania pistaciella, the high cold tolerance observed during overwintering is not linked to the accumulation of any low-molecular-mass cryoprotective substances [117]. Basal accumulation is often genetically programmed, involving constitutive expression of cryoprotectant-related genes.
The cold-adapted species of Drosophila montana exhibits inherent cold tolerance linked to the circadian clock gene vrille. Silencing vrille via RNAi disrupts both short- and long-term cold acclimation, suggesting that basal molecular mechanisms tied to circadian rhythms pre-adapt these flies to cold stress [118]. The fall armyworm (Spodoptera frugiperda) relies on basal levels of glucose and trehalose in its hemolymph. RNAi knockdown of trehalase (Treh) genes reduces cryoprotectant hydrolysis, impairing cold survival. This highlights the importance of constitutive trehalose metabolism in maintaining baseline cold tolerance [76]. However, some species may employ multiple strategies to cope with unfavorable environmental conditions. For instance, the oriental fruit fly (Bactrocera dorsalis) enhances its cold tolerance through rapid cold hardening and acclimation. Exposure to mild low temperatures (5–10°C) for several hours (rapid cold hardening) or days (cold acclimation) increases survival rates at extreme temperatures (-6.5°C). This process is accompanied by a rapid accumulation of trehalose, glycerol, and proteins and a reduction in body water content. These physiological changes stabilize cell membranes and prevent ice formation [111].
Table 3 summarizes the predominant cryoprotectants identified in various insect species across different orders and families. Data presented in this table demonstrates that trehalose (approximately 57%) and glycerol (approximately 46%) are the primary cryoprotectants that contribute to the improvement of cold tolerance in insect species. Together, they account for nearly 80% of the documented cases. Myo-inositol, though less common, ranks third, underscoring its significance in cold tolerance across various taxa. The research highlights the considerable diversity in cryoprotectant usage among insects. While some species use a combination of cryoprotectants to further enhance their cold tolerance, in others, a single cryoprotectant plays a dominant role, often at much higher concentrations. Glycerol, in particular, emerges as the leading cryoprotectant, found in 60% of species that rely on just one compound for cold tolerance. The table lists 56 species across multiple insect orders, including Coleoptera, Diptera, Lepidoptera, and Hymenoptera, among others. However, the distribution of these compounds is not uniform, certain insect orders, like Coleoptera and Lepidoptera, are more commonly studied, while others have limited data. This reflects both the focus of research, particularly on agricultural pests, and the varying mechanisms of cryoprotection across species.
Data presented in Table 3 show that many studied species employ combinations of cryoprotectants, which might offer synergistic effects for thermal tolerance and osmotic stress management. For example: Anoplophora glabripennis uses glycerol, glucose, sorbitol, and trehalose. Sesamia inferens utilizes trehalose, glycerol, glucose, fructose, and myo-inositol. Certain species exhibit rare or unique cryoprotectants like D-mannitol in Acanthococcus lagerstroemiae (Hemiptera: Eriococcidae) or specific amino acids in Lipoptena cervi (Diptera: Hippoboscidae). In Trogoderma granarium, the accumulation of trehalose, sorbitol, and myo-inositol suggests a highly effective mechanism for water retention and cold tolerance. This biochemical strategy is likely linked to the insect's pest behavior in stored grains. In Dendroctonus ponderosae, glycerol plays a crucial role in its adaptation to the boreal forest climate, characterized by frequent freezing temperatures. Similarly, both Pytho depressus and P. americanus rely solely on glycerol for their overwintering strategies, which aligns with their survival in colder environments.
The widespread use of glycerol, trehalose, and myo-inositol is common among Coleoptera, while glycerol and trehalose are predominant in Diptera. Myo-inositol and proline accumulation in D. melanogaster indicate roles beyond cold tolerance, including desiccation resistance. Amino acids like proline and glutamine may signal a demand for nitrogen-rich metabolites during the overwintering of Lipoptena cervi. Meanwhile, a combination of proteins, glycerol, and trehalose likely aids in the metabolic adjustments during diapause in Bactrocera dorsalis. High variability has been observed in Lepidoptera, where trehalose and glycerol are frequently utilized. In this order, cryoprotectants help larvae and pupae survive winter and resist desiccation. For instance, trehalose, sorbitol, and myo-inositol indicate significant adaptations for temperate climates in Pieris brassicae, while trehalose and glycerol support Ostrinia nubilalis in agricultural environments with variable temperatures. Glycerol is the main cryoprotectant among species, likely due to the social structures and overwintering behaviors of ants and wasps. For example, Solenopsis invicta relies on glycerol to sustain colony metabolism in colder months. In Orthoptera, A diverse set of glycerol, sorbitol, and amino acids is highlighted.
7. Cryoprotectant Accumulation Processes
Insects rely on environmental signals, such as cooler temperatures and shorter daylight hours, to trigger the production of cryoprotectants—molecules that protect them from freezing. These cues activate complex signaling pathways that regulate gene expression and control the synthesis of cryoprotectants. For insects in temperate regions, glycerol plays a critical role in preventing ice formation within their bodies during winter. This process mainly utilizes glycogen reserves through the pentose phosphate pathway (PPP), although insects can also produce glycerol quickly through glycolysis in response to sudden cold events. The PPP generates NADPH, which is crucial for reducing intermediates in the production of polyols like glycerol. Glycerol synthesis is regulated by cold exposure, which upregulates the enzyme glycerol-3-phosphate dehydrogenase (GPDH), converting dihydroxyacetone phosphate (DHAP) to glycerol-3-phosphate, which is then dephosphorylated to form glycerol. Another important cryoprotectant, trehalose, is synthesized from glucose-6-phosphate and UDP-glucose in a two-step process involving the enzymes trehalose-6-phosphate synthase (TPS) and trehalose-6-phosphate phosphatase (TPP). Cold stress can also induce the expression of these enzymes to increase trehalose production. Producing these cryoprotectants requires considerable energy. As a result, insects often shift their metabolism to prioritize cryoprotectant synthesis, sometimes diverting resources away from growth and reproduction to cope with the stresses of cold temperatures. For example, in Choristoneura fumiferana, enzymes such as glucose-6-phosphate dehydrogenase play a critical role in this redirection [119,120]. In the mountain pine beetle, Dendroctonus ponderosae, mRNA transcript analyses indicated that glycerol is primarily produced through glycogenolytic and gluconeogenic pathways, rather than lipid metabolism [121]. In rapid cold-hardening (RCH) or short-term responses to cold, glycerol can also be synthesized via glycolysis. Here, dihydroxyacetone phosphate (DHAP) is converted into glycerol-3-phosphate, which is subsequently reduced to glycerol. This pathway is faster than the PPP but consumes ATP, which is limited during winter dormancy [119]. Cold exposure induces differential expression of genes encoding enzymes in cryoprotectant pathways. For example, genes involved in PPP are upregulated during seasonal acclimation, while glycolytic enzymes are modulated during rapid cold responses. It was found that RCH enhances cold tolerance in the leafminer Liriomyza trifolii, by altering fatty acid composition and increasing levels of cryoprotectants such as glucose and trehalose. RNA sequencing analysis after a brief acclimation period reveals significant transcriptional regulation, with differentially expressed genes involved in various metabolic pathways [110]. Insects rely on internal macromolecular reserves like glycogen for cryoprotectant synthesis during winter dormancy when food intake is restricted. Glycogen degradation is triggered by low temperatures, activating glycogen phosphorylase to release glucose for subsequent conversion into polyols or sugars like trehalose [120,122]. During sudden cold exposure, insects rapidly increase cryoprotectant levels through transcriptional regulation and metabolic shifts. For instance, L. trifolii shows increased expression of genes involved in amino acid and sugar metabolism within hours of cold exposure [110]. Cryoprotectants are stored in the hemolymph or intracellular compartments to stabilize proteins and membranes against freezing damage. This ensures their availability at critical sites where freezing stress occurs [31,113]. Insects employ a combination of metabolic reprogramming, enzymatic regulation, and gene expression changes to synthesize and accumulate cryoprotectants such as glycerol, trehalose, and proline. These mechanisms ensure survival under both gradual seasonal cooling and sudden temperature drops through the efficient use of internal reserves and dynamic metabolic flexibility.
8. Cryoprotectant Accumulation in Other Ectotherms
Insects demonstrate a greater diversity and adaptability in producing cryoprotectants, which enables them to develop complex mechanisms for freeze tolerance, including the ability to tolerate intracellular freezing. In contrast, other ectotherms generally rely on simpler strategies that focus on extracellular freezing and have limited biochemical diversity (Table 4). These differences underscore the evolutionary specialization of insects to survive in extremely cold environments. For instance, ectotherms like amphibians and fish typically accumulate fewer cryoprotectants, with glycerol and glucose being the primary compounds they use for freeze tolerance. Wood frogs, a type of amphibian, depend heavily on glucose for cold tolerance during freezing [25]. Both amphibians and reptiles largely rely on external temperature cues to trigger the synthesis of glycerol or glucose, often derived from the breakdown of glycogen [123]. Unlike insects, many other ectotherms cannot withstand intracellular freezing. Their survival strategies primarily focus on extracellular freezing or supercooling, which require less biochemical diversity overall [124].
9. Conclusions
This manuscript explores the biochemical, physiological, and ecological adaptations that enable insects to survive extreme cold. Cold hardiness is a key factor for insect survival, influencing species distribution, reproductive success, and evolutionary fitness. The synthesis and accumulation of cryoprotectants, such as glycerol, trehalose, and amino acids, highlight the sophistication of these adaptations. These substances play critical roles in membrane stabilization, ice nucleation inhibition, osmotic regulation, and antioxidant activity. The variety of cryoprotectant strategies among species, ranging from the accumulation of a single predominant compound to the synergistic use of multiple cryoprotectants, demonstrates the evolutionary flexibility of insects in response to environmental pressures. This adaptability ensures their survival not only during seasonal cold stress but also amid the broader challenges posed by climate change. Additionally, the interaction of cryoprotectants with factors like antifreeze proteins, ice-nucleating proteins, and environmental cues enhances our understanding of the mechanisms underlying cold hardiness. These insights not only advance the field of insect physiology but also offer practical applications in pest management, conservation, and ecological research. Future studies should prioritize uncovering the regulatory pathways that govern cryoprotectant synthesis and exploring the potential roles of less common cryoprotectants. Such efforts could deepen our understanding of insect resilience and inform strategies to mitigate the impacts of climate change on both pest and beneficial insect populations. Ultimately, studying insect cold hardiness reveals the remarkable adaptability of life and its ability to thrive under even the harshest conditions.
Author Contributions
Data collection and manuscript writing were carried out by Hamzeh Izadi.
Funding
No funding.
Data Availability Statement
This is a review manuscript, and no original data is available.
References
- Logan, J.D.; Wolesensky, W.; Joern, A. Temperature-Dependent Phenology and Predation in Arthropod Systems. Ecol Modell 2006, 196, 471–482. [Google Scholar] [CrossRef]
- Yang, L.H.; Rudolf, V.H.W. Phenology, Ontogeny and the Effects of Climate Change on the Timing of Species Interactions. Ecol Lett 2010, 13, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Régnière, J.; St-Amant, R.; Duval, P. Predicting Insect Distributions under Climate Change from Physiological Responses: Spruce Budworm as an Example. Biol Invasions 2012, 14, 1571–1586. [Google Scholar] [CrossRef]
- Sinclair, B.J.; Coello Alvarado, L.E.; Ferguson, L. V. An Invitation to Measure Insect Cold Tolerance: Methods, Approaches, and Workflow. J Therm Biol 2015, 53, 180–197. [Google Scholar] [CrossRef]
- Feng, Y.; Zhang, L.; Li, W.; Yang, X.; Zong, S. Cold Hardiness of Overwintering Larvae of Sphenoptera Sp. (Coleoptera: Buprestidae) in Western China. J Econ Entomol 2018, 111, 247–251. [Google Scholar] [CrossRef]
- Sinclair, B.J.; Vernon, P.; Klok, C.J.; Chown, S.L. Insects at Low Temperatures: An Ecological Perspective. Trends Ecol Evol 2003, 18, 257–262. [Google Scholar] [CrossRef]
- Watanabe, M.; Kikawada, T.; Minagawa, N.; Yukuhiro, F.; Okuda, T. Mechanism Allowing an Insect to Survive Complete Dehydration and Extreme Temperatures. J Exp Biol 2002, 205, 2799–2802. [Google Scholar] [CrossRef]
- Bale, J.S.; Gerday, C.; Parker, A.; Marahiel, M.A.; Shanks, I.A.; Davies, P.L.; Warren, G. Insects and Low Temperatures: From Molecular Biology to Distributions and Abundance. Philosophical Transactions of the Royal Society B: Biological Sciences 2002, 357, 849–862. [Google Scholar] [CrossRef]
- Bemani, M.; Izadi, H.; Mahdian, K.; Khani, A.; Amin samih, M. Study on the Physiology of Diapause, Cold Hardiness and Supercooling Point of Overwintering Pupae of the Pistachio Fruit Hull Borer, Arimania Comaroffi. J Insect Physiol 2012, 58, 897–902. [Google Scholar] [CrossRef]
- Bemani, M.; Izadi, H.; Mahdian, K.; Khani, A.; Amin samih, M. Study on the Physiology of Diapause, Cold Hardiness and Supercooling Point of Overwintering Pupae of the Pistachio Fruit Hull Borer, Arimania Comaroffi. J Insect Physiol 2012, 58, 897–902. [Google Scholar] [CrossRef]
- Masoudmagham, A.; Izadi, H.; Mohammadzadeh, M. Expanded Supercooling Capacity With No Cryoprotectant Accumulation Underlies Cold Tolerance of the European Grapevine Moth. J Econ Entomol 2021, 114, 828–838. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.E.; Lee Richard E, Jr.; Lee, R.E.; Lee Jr., R. E. Insect Cold-Hardiness: To Freeze or Not to Freeze. Bioscience 1989, 39, 308–313. [Google Scholar] [CrossRef]
- Milonas, P.G.; Savopoulou-Soultani, M. Cold Hardiness in Diapause and Non-Diapause Larvae of the Summer Fruit Tortrix, Adoxophyes Orana (Lepidoptera: Tortricidae). Eur J Entomol 1999, 96, 183–187. [Google Scholar]
- Mohammadzadeh, M.; Izadi, H. Cooling Rate and Starvation Affect Supercooling Point and Cold Tolerance of the Khapra Beetle, Trogoderma Granarium Everts Fourth Instar Larvae (Coleoptera: Dermestidae). J Therm Biol 2018, 71, 24–31. [Google Scholar] [CrossRef]
- Stockton, D.; Wallingford, A.; Rendon, D.; Fanning, P.; Green, C.K.; Diepenbrock, L.; Ballman, E.; Walton, V.M.; Isaacs, R.; Leach, H.; et al. Interactions between Biotic and Abiotic Factors Affect Survival in Overwintering Drosophila Suzukii (Diptera: Drosophilidae). Environ Entomol 2019, 48, 454–464. [Google Scholar] [CrossRef]
- Zachariassen, K.E. Physiology of Cold Tolerance in Insects. Physiol. Rev. 1985, 65, 799–832. [Google Scholar] [CrossRef]
- Marshall, K.E.; Sinclair, B.J. The Impacts of Repeated Cold Exposure on Insects. Journal of Experimental Biology 2012, 215, 1607–1613. [Google Scholar] [CrossRef]
- Colinet, H.; Sinclair, B.J.; Vernon, P.; Renault, D. Insects in Fluctuating Thermal Environments. Annu Rev Entomol 2015, 60, 123–140. [Google Scholar] [CrossRef]
- Chen, S.-Y.; Zhao, R.-N.; Li, Y.; Li, H.-P.; Xie, M.-H.; Liu, J.-F.; Yang, M.-F.; Wu, C.-X. Cold Tolerance Strategy and Cryoprotectants of Megabruchidius Dorsalis in Different Temperature and Time Stresses. Front Physiol 2023, 13. [Google Scholar] [CrossRef]
- Toxopeus, J.; Sinclair, B.J. Mechanisms Underlying Insect Freeze Tolerance. Biological Reviews 2018, 93. [Google Scholar] [CrossRef]
- Ankit, J. Joshi A Review and Application of Cryoprotectant: The Science of Cryonics. PharmaTutor 2016, 4, 12–18. [Google Scholar]
- Marcantonini, G.; Zatini, L.; Costa, S.; Passerini, M.; Rende, M.; Luca, G.; Basta, G.; Murdolo, G.; Calafiore, R.; Galli, F. Natural Cryoprotective and Cytoprotective Agents in Cryopreservation: A Focus on Melatonin. Molecules 2022, 27, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.-Q.; Tan, P.-C.; Gao, Y.-M.; Zhang, X.-J.; Xie, Y.; Zheng, D.-N.; Zhou, S.-B.; Li, Q.-F. The Effect of Glycerol as a Cryoprotective Agent in the Cryopreservation of Adipose Tissue. Stem Cell Res Ther 2022, 13, 152. [Google Scholar] [CrossRef]
- Fuller, B.J. Cryoprotectants: The Essential Antifreezes to Protect Life in the Frozen State. Cryo-Letters 2004, 25, 375–388. [Google Scholar]
- Storey, K.B.; Storey, J.M. Biochemistry of Cryoprotectants. Insects at Low Temperature 1991, 64–93. [Google Scholar] [CrossRef]
- Hincha, D.K.; Schmidt, J.E.; Heber, U.; Schmitt, J.M. Colligative and Non-Colligative Freezing Damage to Thylakoid Membranes. Biochimica et Biophysica Acta (BBA) - Biomembranes 1984, 769, 8–14. [Google Scholar] [CrossRef]
- Benson, E.E. Cryopreservation Theory. In Plant Cryopreservation: A Practical Guide; Springer New York: New York, NY, 2008; pp. 15–32. [Google Scholar]
- Lee, R.E. A Primer on Insect Cold-Tolerance. In Low Temperature Biology of Insects; 2010 ISBN 9780511675997.
- Marcantonini, G.; Bartolini, D.; Zatini, L.; Costa, S.; Passerini, M.; Rende, M.; Luca, G.; Basta, G.; Murdolo, G.; Calafiore, R.; et al. Natural Cryoprotective and Cytoprotective Agents in Cryopreservation: A Focus on Melatonin. Molecules 2022, 27, 3254. [Google Scholar] [CrossRef]
- Zachariassen, K.E. The Mechanism of the Cryoprotective Effect of Glycerol in Beetles Tolerant to Freezing. J Insect Physiol 1979, 25, 29–32. [Google Scholar] [CrossRef]
- Toxopeus, J.; Koštál, V.; Sinclair, B.J. Evidence for Non-Colligative Function of Small Cryoprotectants in a Freeze-Tolerant Insect. proceedings of the royal society B 2019. [Google Scholar] [CrossRef]
- Worland, M.R.; Grubor-Lajšić, G.; Purać, J.; Thorne, M.A.S.; Clark, M.S. Cryoprotective Dehydration: Clues from an Insect. In Topics in Current Genetics; 2010; Vol. 21, pp. 147–163 ISBN 9783642124211.
- Olsson, T.; MacMillan, H.A.; Nyberg, N.; Staerk, D.; Malmendal, A.; Overgaard, J. Hemolymph Metabolites and Osmolality Are Tightly Linked to Cold Tolerance of Drosophila Species: A Comparative Study. Journal of Experimental Biology 2016, 219, 2504–2513. [Google Scholar] [CrossRef]
- Grgac, R.; Rozsypal, J.; Des Marteaux, L.; Štětina, T.; Koštál, V. Stabilization of Insect Cell Membranes and Soluble Enzymes by Accumulated Cryoprotectants during Freezing Stress. Proceedings of the National Academy of Sciences 2022, 119. [Google Scholar] [CrossRef] [PubMed]
- Rudolph, A.S.; Crowe, J.H. Membrane Stabilization during Freezing: The Role of Two Natural Cryoprotectants, Trehalose and Proline. Cryobiology 1985, 22, 367–377. [Google Scholar] [CrossRef] [PubMed]
- K. E., Z.; E., K. Ice Nucleation and Antinucleation in Nature. Cryobiology 2000, 41, 257–279. [Google Scholar] [CrossRef]
- Lee, R.E.; Costanzo, J.P.; Mugnano, J.A. Regulation of Supercooling and Ice Nucleation in Insects. Eur J Entomol 1996, 93, 405–418. [Google Scholar]
- Lee, R.E.; Costanzo, J.P. Biological Ice Nucleation and Ice Distribution in Cold-Hardy Ectothermic Animals. Annu Rev Physiol 1998, 60, 55–72. [Google Scholar] [CrossRef]
- Halberg, K.V.; Denholm, B. Mechanisms of Systemic Osmoregulation in Insects. Annu Rev Entomol 2024, 69, 415–438. [Google Scholar] [CrossRef]
- Chown, S.L.; Nicolson, S.W. Water Balance Physiology. In Insect Physiological Ecology; Oxford University Press, 2004; pp. 87–114.
- Miao, Z.Q.; Tu, Y.Q.; Guo, P.Y.; He, W.; Jing, T.X.; Wang, J.J.; Wei, D.D. Antioxidant Enzymes and Heat Shock Protein Genes from Liposcelis Bostrychophila Are Involved in Stress Defense upon Heat Shock. Insects 2020, 11, 839. [Google Scholar] [CrossRef]
- Sformo, T.; Walters, K.; Jeannet, K.; Wowk, B.; Fahy, G.M.; Barnes, B.M.; Duman, J.G. Deep Supercooling, Vitrification and Limited Survival to -100 C in the Alaskan Beetle Cucujus Clavipes Puniceus (Coleoptera: Cucujidae) Larvae. Journal of Experimental Biology 2010, 213, 502–509. [Google Scholar] [CrossRef]
- Wharton, D.A. Supercooling and Freezing Tolerant Animals. Supercooling 2012. [Google Scholar] [CrossRef]
- Sømme, L. Supercooling and Winter Survival in Terrestrial Arthropods. Comp Biochem Physiol A Physiol 1982, 73, 519–543. [Google Scholar] [CrossRef]
- Duman, J.G. Animal Ice-Binding (Antifreeze) Proteins and Glycolipids: An Overview with Emphasis on Physiological Function. Journal of Experimental Biology 2015, 218, 1846–1855. [Google Scholar] [CrossRef] [PubMed]
- Danks, H.V. Insect Adaptations to Cold and Changing Environments. Can Entomol 2009, 138, 1–23. [Google Scholar] [CrossRef]
- Ishiguro, S.; Li, Y.; Nakano, K.; Tsumuki, H.; Goto, M. Seasonal Changes in Glycerol Content and Cold Hardiness in Two Ecotypes of the Rice Stem Borer, Chilo Suppressalis, Exposed to the Environment in the Shonai District, Japan. J Insect Physiol 2007, 53, 392–397. [Google Scholar] [CrossRef]
- Gerber, L.; Overgaard, J. Cold Tolerance Is Linked to Osmoregulatory Function of the Hindgut in Locusta Migratoria. Journal of Experimental Biology 2018, 221. [Google Scholar] [CrossRef]
- Storey, K.B.; Storey, J.M. Freeze Tolerance: Constraining Forces, Adaptive Mechanisms. Can J Zool 1988, 66, 11–22. [Google Scholar] [CrossRef]
- Elliott, G.D.; Wang, S.; Fuller, B.J. Cryoprotectants: A Review of the Actions and Applications of Cryoprotective Solutes That Modulate Cell Recovery from Ultra-Low Temperatures. Cryobiology 2017, 76, 74–91. [Google Scholar] [CrossRef]
- Bartolac, L.K.; Lowe, J.L.; Koustas, G.; Grupen, C.G.; Sjöblom, C. Effect of Different Penetrating and Non-penetrating Cryoprotectants and Media Temperature on the Cryosurvival of Vitrified in Vitro Produced Porcine Blastocysts. Animal Science Journal 2018, 89, 1230–1239. [Google Scholar] [CrossRef]
- Upton, R.; Clulow, S.; Colyvas, K.; Mahony, M.; Clulow, J. Paradigm Shift in Frog Sperm Cryopreservation: Reduced Role for Non-Penetrating Cryoprotectants. Reproduction 2023, 165, 583–592. [Google Scholar] [CrossRef]
- Tao, D.; Li, P.H. Classification of Plant Cell Cryoprotectants. J Theor Biol 1986, 123, 305–310. [Google Scholar] [CrossRef]
- Jaiswal, A.N.; Vagga, A. Cryopreservation: A Review Article. Cureus 2022. [Google Scholar] [CrossRef]
- Pegg, D.E. Principles of Cryopreservation. In; 2007; pp. 39–57.
- Košťál, V.; Šimek, P. Overwintering Strategy in Pyrrhocoris Apterus (Heteroptera): The Relations between Life-Cycle, Chill Tolerance and Physiological Adjustments. J Insect Physiol 2000, 46, 1321–1329. [Google Scholar] [CrossRef] [PubMed]
- Košt’ál, V.; Šlachta, M.; Šimek, P. Cryoprotective Role of Polyols Independent of the Increase in Supercooling Capacity in Diapausing Adults of Pyrrhocoris Apterus (Heteroptera: Insecta). Comparative Biochemistry and Physiology - B Biochemistry and Molecular Biology 2001, 130, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Towey, J.J.; Dougan, L. Structural Examination of the Impact of Glycerol on Water Structure. J Phys Chem B 2012, 116, 1633–1641. [Google Scholar] [CrossRef] [PubMed]
- Sieme, H.; Oldenhof, H.; Wolkers, W.F. Mode of Action of Cryoprotectants for Sperm Preservation. Anim Reprod Sci 2016, 169, 2–5. [Google Scholar] [CrossRef]
- Schrader, A.M.; Cheng, C.-Y.; Israelachvili, J.N.; Han, S. Communication: Contrasting Effects of Glycerol and DMSO on Lipid Membrane Surface Hydration Dynamics and Forces. J Chem Phys 2016, 145. [Google Scholar] [CrossRef]
- Patrick Williams, W.; Quinn, P.J.; Tsonev, L.I.; Koynova, R.D. The Effects of Glycerol on the Phase Behaviour of Hydrated Distearoylphosphatidylethanolamine and Its Possible Relation to the Mode of Action of Cryoprotectants. Biochimica et Biophysica Acta (BBA) - Biomembranes 1991, 1062, 123–132. [Google Scholar] [CrossRef]
- Jain, N.K.; Roy, I. Trehalose and Protein Stability. Curr Protoc Protein Sci 2010. [Google Scholar] [CrossRef]
- Tang, B.; Wang, S.; Wang, S.G.; Wang, H.J.; Zhang, J.Y.; Cui, S.Y. Invertebrate Trehalose-6-Phosphate Synthase Gene: Genetic Architecture, Biochemistry, Physiological Function, and Potential Applications. Front Physiol 2018, 9. [Google Scholar] [CrossRef]
- Thompson, S.N. Trehalose - The Insect “Blood” Sugar. Adv In Insect Phys 2003, 31, 205–285. [Google Scholar] [CrossRef]
- Tellis, M.B.; Kotkar, H.M.; Joshi, R.S. Regulation of Trehalose Metabolism in Insects: From Genes to the Metabolite Window. Glycobiology 2023, 33, 262–273. [Google Scholar] [CrossRef]
- Lee, H.-J.; Yoon, Y.-S.; Lee, S.-J. Mechanism of Neuroprotection by Trehalose: Controversy Surrounding Autophagy Induction. Cell Death Dis 2018, 9, 712. [Google Scholar] [CrossRef] [PubMed]
- Corradini, D.; Strekalova, E.G.; Stanley, H.E.; Gallo, P. Microscopic Mechanism of Protein Cryopreservation in an Aqueous Solution with Trehalose. Sci Rep 2013, 3, 1218. [Google Scholar] [CrossRef] [PubMed]
- Clark, M.S.; Thorne, M.A.S.; Purać, J.; Burns, G.; Hillyard, G.; Popović, Ž.D.; Grubor-Lajšić, G.; Worland, M.R. Surviving the Cold: Molecular Analyses of Insect Cryoprotective Dehydration in the Arctic Springtail Megaphorura Arctica (Tullberg). BMC Genomics 2009, 10, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Campbell, L.H.; Brockbank, K.G.M. Comparison of Electroporation and ChariotTM for Delivery of β-Galactosidase into Mammalian Cells: Strategies to Use Trehalose in Cell Preservation. In Vitro Cell Dev Biol Anim 2011, 47, 195–199. [Google Scholar] [CrossRef]
- Wen, X.; Wang, S.; Duman, J.G.; Arifin, J.F.; Juwita, V.; Goddard, W.A.; Rios, A.; Liu, F.; Kim, S.-K.; Abrol, R.; et al. Antifreeze Proteins Govern the Precipitation of Trehalose in a Freezing-Avoiding Insect at Low Temperature. Proceedings of the National Academy of Sciences 2016, 113, 6683–6688. [Google Scholar] [CrossRef]
- Alyssa, R. Stephens Biochemistry of Trehalose Accumulation in the Spring Field Cricket, Gryllus Veletis, The University of Western Ontario, 2022.
- Yang, H.-J.; Cui, M.-Y.; Zhao, X.-H.; Zhang, C.-Y.; Hu, Y.-S.; Fan, D. Trehalose-6-Phosphate Synthase Regulates Chitin Synthesis in Mythimna Separata. Front Physiol 2023, 14. [Google Scholar] [CrossRef]
- Chen, Y.; Pan, Y.; Feng, Y.; Li, D.; Man, J.; Feng, L.; Zhang, D.; Chen, H.; Chen, H. Role of Glucose in the Repair of Cell Membrane Damage during Squeeze Distortion of Erythrocytes in Microfluidic Capillaries. Lab Chip 2021, 21, 896–903. [Google Scholar] [CrossRef]
- Burton, K.P.; Templeton, G.H.; Hagler, H.K.; Willerson, J.T.; Buja, L.M. Effect of Glucose Availability on Functional Membrane Integrity, Ultrastructure and Contractile Performance Following Hypoxia and Reoxygenation in Isolated Feline Cardiac Muscle. J Mol Cell Cardiol 1980, 12, 109–133. [Google Scholar] [CrossRef]
- Amornwittawat, N.; Wang, S.; Banatlao, J.; Chung, M.; Velasco, E.; Duman, J.G.; Wen, X. Effects of Polyhydroxy Compounds on Beetle Antifreeze Protein Activity. Biochimica et Biophysica Acta (BBA) - Proteins and Proteomics 2009, 1794, 341–346. [Google Scholar] [CrossRef]
- Gokulanathan, A.; Mo, H.; Park, Y. Glucose Influence Cold Tolerance in the Fall Armyworm, Spodoptera Frugiperda via Trehalase Gene Expression. Sci Rep 2024, 14, 27334. [Google Scholar] [CrossRef]
- STEINER, A.A.; PETENUSCI, S.O.; BRENTEGANI, L.G.; BRANCO, L.G.S. The Importance of Glucose for the Freezing Tolerance/Intolerance of the Anuran Amphibians Rana Catesbeiana and Bufo Paracnemis. Rev Bras Biol 2000, 60, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Bakó, I.; Pusztai, L.; Pothoczki, S. Outstanding Properties of the Hydration Shell around β- <scp>d</Scp> -Glucose: A Computational Study. ACS Omega 2024, 9, 20331–20337. [Google Scholar] [CrossRef] [PubMed]
- Vesala, L.; Salminen, T.S.; Kostál, V.; Zahradníĉková, H.; Hoikkala, A. Myo-Inositol as a Main Metabolite in Overwintering Flies: Seasonal Metabolomic Profiles and Cold Stress Tolerance in a Northern Drosophilid Fly. Journal of Experimental Biology 2012, 215, 2891–2897. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, S. Cryopretectants and Their Usage in Cryopreservation Process. In Cryopreservation Biotechnology in Biomedical and Biological Sciences; IntechOpen, 2018.
- Watanabe, M. Cold Tolerance and Myo-Inositol Accumulation in Overwintering Adults of a Lady Beetle, Harmonia Axyridis (Coleoptera: Coccinellidae). Eur J Entomol 2002, 99, 5–9. [Google Scholar] [CrossRef]
- Wolfe, G.R.; Hendrix, D.L.; Salvucci, M.E. A Thermoprotective Role for Sorbitol in the Silverleaf Whitefly, Bemisia Argentifolii. J Insect Physiol 1998, 44, 597–603. [Google Scholar] [CrossRef]
- Walters, K.R.; Pan, Q.; Serianni, A.S.; Duman, J.G. Cryoprotectant Biosynthesis and the Selective Accumulation of Threitol in the Freeze-Tolerant Alaskan Beetle, Upis Ceramboides. Journal of Biological Chemistry 2009, 284, 16822–16831. [Google Scholar] [CrossRef]
- Miller, L.K.; Smith, J.S. Production of Threitol and Sorbitol by an Adult Insect: Association with Freezing Tolerance. Nature 1975, 258, 519–520. [Google Scholar] [CrossRef]
- Helou, B.; Ritchie, M.W.; MacMillan, H.A.; Andersen, M.K. Dietary Potassium and Cold Acclimation Additively Increase Cold Tolerance in Drosophila Melanogaster. J Insect Physiol 2024, 159, 104701. [Google Scholar] [CrossRef]
- Xie, Z.; Xu, L.; Zhao, J.; Li, N.; Qin, D.; Xiao, C.; Lu, Y.; Guo, Z. Rapid Cold Hardening and Cold Acclimation Promote Cold Tolerance of Oriental Fruit Fly, Bactrocera Dorsalis (Hendel) by Physiological Substances Transformation and Cryoprotectants Accumulation. Bull Entomol Res 2023, 113, 574–586. [Google Scholar] [CrossRef]
- Izadi, H.; Mohammadzadeh, M.; Mehrabian, M. Cold Tolerance of the Tribolium Castaneum (Coleoptera: Tenebrionidae), under Different Thermal Regimes: Impact of Cold Acclimation. J Econ Entomol 2019, 112, 1983–1988. [Google Scholar] [CrossRef]
- Wang, J.; Gao, G.; Zhang, R.; Dai, L.; Chen, H. Metabolism and Cold Tolerance of Chinese White Pine Beetle Dendroctonus Armandi (Coleoptera: Curculionidae: Scolytinae) during the Overwintering Period. Agric For Entomol 2017, 19, 10–22. [Google Scholar] [CrossRef]
- Song, Y.; Huang, W.; Zhou, Y.; Li, Z.; Ji, R.; Ye, X. Physiological Characteristics and Cold Tolerance of Overwintering Eggs in Gomphocerus Sibiricus L. (Orthoptera: Acrididae). Arch Insect Biochem Physiol 2021, 108. [Google Scholar] [CrossRef] [PubMed]
- Kučera, L.; Moos, M.; Štětina, T.; Korbelová, J.; Vodrážka, P.; Des Marteaux, L.; Grgac, R.; Hůla, P.; Rozsypal, J.; Faltus, M.; et al. A Mixture of Innate Cryoprotectants Is Key for Freeze Tolerance and Cryopreservation of a Drosophilid Fly Larva. Journal of Experimental Biology 2022, 225. [Google Scholar] [CrossRef] [PubMed]
- Hasanvand, H.; Izadi, H.; Mohammadzadeh, M. Overwintering Physiology and Cold Tolerance of the Sunn Pest, Eurygaster Integriceps, an Emphasis on the Role of Cryoprotectants. Front Physiol 2020, 11. [Google Scholar] [CrossRef]
- Duman, J.G. Thermal-Hysteresis-Factors in Overwintering Insects. J Insect Physiol 1979, 25, 805–810. [Google Scholar] [CrossRef]
- Patterson, J.L.; Duman, J.G. The Role of the Thermal Hysteresis Factor in Tenebrio Molitor Larvae. Journal of Experimental Biology 1978, 74, 37–45. [Google Scholar] [CrossRef]
- Lorv, J.S.H.; Rose, D.R.; Glick, B.R. Bacterial Ice Crystal Controlling Proteins. Scientifica (Cairo) 2014, 2014, 1–20. [Google Scholar] [CrossRef]
- Melnik, B.S.; Glukhova, K.A.; Sokolova (Voronova), E.A.; Balalaeva, I. V.; Garbuzynskiy, S.O.; Finkelstein, A. V. Physics of Ice Nucleation and Antinucleation: Action of Ice-Binding Proteins. Biomolecules 2023, 14, 54. [Google Scholar] [CrossRef]
- Roeters, S.J.; Golbek, T.W.; Bregnhøj, M.; Drace, T.; Alamdari, S.; Roseboom, W.; Kramer, G.; Šantl-Temkiv, T.; Finster, K.; Pfaendtner, J.; et al. Ice-Nucleating Proteins Are Activated by Low Temperatures to Control the Structure of Interfacial Water. Nat Commun 2021, 12, 1183. [Google Scholar] [CrossRef]
- Białkowska, A.; Majewska, E.; Olczak, A.; Twarda-Clapa, A. Ice Binding Proteins: Diverse Biological Roles and Applications in Different Types of Industry. Biomolecules 2020, 10, 274. [Google Scholar] [CrossRef]
- John G Duman Antifreeze and Ice Nucleator Proteins in Terrestrial Arthropods. Annu Rev Physiol 2001, 63, 327–357. [CrossRef] [PubMed]
- Duman, J.G. The Role of Macromolecular Antifreeze in the Darkling Beetle,Meracantha Contracta. Journal of Comparative Physiology ? B 1977, 115, 279–286. [Google Scholar] [CrossRef]
- Duman, J.G. Subzero Temperature Tolerance in Spiders: The Role of Thermal-Hysteresis-Factors. Journal of Comparative Physiology ? B 1979, 131, 347–352. [Google Scholar] [CrossRef]
- Baust, J.G.; Nishino, M. Freezing Tolerance in the Goldenrod Gall Fly (Eurosta Solidaginis). In Insects at Low Temperature; Springer US: Boston, MA, 1991; pp. 260–275. [Google Scholar]
- Enriquez, T.; Colinet, H. Cold Acclimation Triggers Lipidomic and Metabolic Adjustments in the Spotted Wing Drosophila Drosophila Suzukii (Matsumara). Am J Physiol Regul Integr Comp Physiol 2019, 316. [Google Scholar] [CrossRef] [PubMed]
- Block, W. Cold Tolerance of Insects and Other Arthropods. Philosophical Transactions - Royal Society of London, B 1990, 326, 613–633. [Google Scholar] [CrossRef]
- Hamedi, N.; Moharramipour, S. Long-Term Cold Response in Overwintering Adults of Ladybird Hippodamia Variegata (Coleoptera: Coccinellidae). J Crop Prot 2013, 2, 119–126. [Google Scholar]
- Hamedi, N.; Moharramipour, S.; Barzegar, M. Temperature-Dependent Chemical Components Accumulation in Hippodamia Variegata (Coleoptera: Coccinellidae) during Overwintering. Environ Entomol 2013, 42, 375–380. [Google Scholar] [CrossRef]
- Heydari, M.; Izadi, H. Effects of Seasonal Acclimation on Cold Tolerance and Biochemical Status of the Carob Moth, Ectomyelois Ceratoniae Zeller, Last Instar Larvae. Bull Entomol Res 2014, 104, 592–600. [Google Scholar] [CrossRef]
- Khajehoseini Saleh Abad, M.; Khani, A.; Izadi, H.; Sahebzadeh, N.; Samih, M.-A. Seasonal Changes in Supercooling Capacity and Main Polyols in Overwintering Adults of Acrosternum Millierei (Hemiptera: Pentatomidae). Int J Pest Manag 2023, 1–8. [Google Scholar] [CrossRef]
- Khabir, M.; Izadi, H.; Mahdian, K. The Supercooling Point Depression Is the Leading Cold Tolerance Strategy for the Variegated Ladybug, [Hippodamia Variegata (Goezel)]. Front Physiol 2023, 14. [Google Scholar] [CrossRef]
- Durak, R.; Depciuch, J.; Kapusta, I.; Kisała, J.; Durak, T. Changes in Chemical Composition and Accumulation of Cryoprotectants as the Adaptation of Anholocyclic Aphid Cinara Tujafilina to Overwintering. Int J Mol Sci 2021, 22, 511. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.-X.; Iqbal, J.; Wang, Y.-C.; Chang, Y.-W.; Hu, J.; Du, Y.-Z. Integrated Transcriptional and Biochemical Profiling Suggests Mechanisms Associated with Rapid Cold Hardening in Adult Liriomyza Trifolii (Burgess). Sci Rep 2024, 14, 24033. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Xu, L.; Zhao, J.; Li, N.; Qin, D.; Xiao, C.; Lu, Y.; Guo, Z. Rapid Cold Hardening and Cold Acclimation Promote Cold Tolerance of Oriental Fruit Fly, Bactrocera Dorsalis (Hendel) by Physiological Substances Transformation and Cryoprotectants Accumulation. Bull Entomol Res 2023, 113, 574–586. [Google Scholar] [CrossRef]
- Khodayari, S. Cryoprotectants in Overwintering Larvae of Leopard Moth, Zeuzera Pyrina (Lepidoptera: Cossidae), Collected from Northwestern Iran. J Crop Prot 2022, 11, 301–313. [Google Scholar]
- Teets, N.M.; Denlinger, D.L. Physiological Mechanisms of Seasonal and Rapid Cold-Hardening in Insects. Physiol Entomol 2013, 38, 105–116. [Google Scholar] [CrossRef]
- Teets, N.M.; Gantz, J.D.; Kawarasaki, Y. Rapid Cold Hardening: Ecological Relevance, Physiological Mechanisms and New Perspectives. J Exp Biol 2020, 223. [Google Scholar] [CrossRef]
- Zhao, Q.; Zhu, D.-H. Effects of Rapid Cold-Hardening and Cold Acclimation on Egg Survival and Cryoprotectant Contents in Ceracris Kiangsu (Orthoptera: Arcypteridae). Ann Entomol Soc Am 2023, 116, 404–411. [Google Scholar] [CrossRef]
- Lee, R.E.; Chen, C.-P.; Denlinger, D.L. A Rapid Cold-Hardening Process in Insects. Science (1979) 1987, 238, 1415–1417. [Google Scholar] [CrossRef]
- Mollaei, M.; Izadi, H.; Šimek, P.; Koštál, V. Overwintering Biology and Limits of Cold Tolerance in Larvae of Pistachio Twig Borer, Kermania Pistaciella. Bull Entomol Res 2016, 106, 538–545. [Google Scholar] [CrossRef]
- Poikela, N.; Tyukmaeva, V.; Hoikkala, A.; Kankare, M. Multiple Paths to Cold Tolerance: The Role of Environmental Cues, Morphological Traits and the Circadian Clock Gene Vrille. BMC Ecol Evol 2021, 21, 117. [Google Scholar] [CrossRef]
- van der Burg, K.R.L.; Bozorgi, Y.; Gyte, K.; Roe, A.D.; Marshall, K.E. Plasticity in Cryoprotectant Synthesis Involves Coordinated Shunting Away from Pyruvate Production 2024.
- Moos, M.; Korbelová, J.; Štětina, T.; Opekar, S.; Šimek, P.; Grgac, R.; Koštál, V. Cryoprotective Metabolites Are Sourced from Both External Diet and Internal Macromolecular Reserves during Metabolic Reprogramming for Freeze Tolerance in Drosophilid Fly. Metabolites 2022, 12, 163. [Google Scholar] [CrossRef] [PubMed]
- Fraser, J.D.; Bonnett, T.R.; Keeling, C.I.; Huber, D.P.W. Seasonal Shifts in Accumulation of Glycerol Biosynthetic Gene Transcripts in Mountain Pine Beetle, Dendroctonus Ponderosae Hopkins (Coleoptera: Curculionidae), Larvae. PeerJ 2017, 2017. [Google Scholar] [CrossRef]
- Li, N.G. Cryoprotectant Systems and Cold Tolerance of Insects Inhabiting Central Yakutia (Russian Far East). Eur J Entomol 2016, 113, 537–541. [Google Scholar] [CrossRef]
- Ali, F.; Wharton, D.A. Infective Juveniles of the Entomopathogenic Nematode, Steinernema Feltiae Produce Cryoprotectants in Response to Freezing and Cold Acclimation. PLoS One 2015. [Google Scholar] [CrossRef]
- Nieminen, P.; Paakkonen, T.; Eerilä, H.; Puukka, K.; Riikonen, J.; Lehto, V.; Mustonen, A. Freezing Tolerance and Low Molecular Weight Cryoprotectants in an Invasive Parasitic Fly, the Deer Ked ( Lipoptena Cervi ). J Exp Zool A Ecol Genet Physiol 2012, 317A, 1–8. [Google Scholar] [CrossRef]
- Mohammadzadeh, M.; Izadi, H. Cold Acclimation of Trogoderma Granarium Everts Is Tightly Linked to Regulation of Enzyme Activity, Energy Content and Ion Concentration. bioRxiv 2018. [CrossRef]
- Soudi, Sh.; Moharramipour, S. Cold Tolerance and Supercooling Capacity in Overwintering Adults of Elm Leaf Beetle Xanthogaleruca Luteola (Coleoptera: Chrysomelidae). Environ Entomol 2012, 40, 1546–1553. [Google Scholar] [CrossRef]
- Soudi, S. , Moharramipour, S., Barzegar, M., Atapour, M. Cryoprotectant Contents of Overwintering Elm Leaf Beetle Xanthogaleruca Luteola. J. Entomol. Soc. Iran 2011, 31, 17–32. [Google Scholar]
- Soudi, S.; Moharramipour, S. Seasonal Patterns of the Thermal Response in Relation to Sugar and Polyol Accumulation in Overwintering Adults of Elm Leaf Beetle, Xanthogaleruca Luteola (Coleoptera: Chrysomelidae). J Therm Biol 2012, 37, 384–391. [Google Scholar] [CrossRef]
- Zhou, Z.S.; Rasmann, S.; Li, M.; Guo, J.Y.; Chen, H.S.; Wan, F.H. Cold Temperatures Increase Cold Hardiness in the Next Generation Ophraella Communa Beetles. PLoS One 2013, 8. [Google Scholar] [CrossRef]
- Boychuk, E.C.; Smiley, J.T.; Dahlhoff, E.P.; Bernards, M.A.; Rank, N.E.; Sinclair, B.J. Cold Tolerance of the Montane Sierra Leaf Beetle, Chrysomela Aeneicollis. J Insect Physiol 2015, 81, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Tanaka, K. Cold Tolerance Strategy of the Freeze-Intolerant Chrysomelid, Aulacophora Nigripennis (Coleoptera: Chrysomelidae), in Warm-Temperate Regions. Eur J Entomol 1999, 96, 175–181. [Google Scholar]
- Saeidi, M.; Moharramipour, S. Physiology of Cold Hardiness, Seasonal Fluctuations, and Cryoprotectant Contents in Overwintering Adults of Hypera Postica (Coleoptera: Curculionidae). Environ Entomol 2017, 46, 960–966. [Google Scholar] [CrossRef] [PubMed]
- Koštál, V.; Miklas, B.; Doležal, P.; Rozsypal, J.; Zahradníčková, H. Physiology of Cold Tolerance in the Bark Beetle, Pityogenes Chalcographus and Its Overwintering in Spruce Stands. J Insect Physiol 2014, 63, 62–70. [Google Scholar] [CrossRef]
- Lee, K.Y.; Chang, Y.D.; Kim, Y.G. Trehalose, a Major Sugar Cryoprotectant of the Overwintering Rice Water Weevil, Lissorhoptrus Oryzophilus (Coleoptera: Curculionidae). J Asia Pac Entomol 2002, 5, 35–41. [Google Scholar] [CrossRef]
- Bonnot, G.; Peypelut, L.; Gérard Febvay; Lavenseau, L. ; Fleurat-Lessard, F.; Fields, P.G. The Effect of Cold Acclimation and Deacclimation on Cold Tolerance, Trehalose and Free Amino Acid Levels in Sitophilus Granarius and Cryptolestes Ferrugineus (Coleoptera). J Insect Physiol 1998, 44, 955–965. [Google Scholar]
- Robert, J.A.; Bonnett, T.; Pitt, C.; Spooner, L.J.; Fraser, J.; Yuen, M.M.S.; Keeling, C.I.; Bohlmann, J.; Huber, D.P.W. Gene Expression Analysis of Overwintering Mountain Pine Beetle Larvae Suggests Multiple Systems Involved in Overwintering Stress, Cold Hardiness, and Preparation for Spring Development. PeerJ 2016, 4, e2109. [Google Scholar] [CrossRef]
- Cheng, L.; Pei, J.; Chen, X.; Shi, F.; Bao, Z.; Hou, Q.; Zhi, L.; Zong, S.; Tao, J. Cold Tolerance and Metabolism of Red-Haired Pine Bark Beetle Hylurgus Ligniperda (Coleoptera: Curculionidae) during the Overwintering Period. J Econ Entomol 2024, 117, 1553–1563. [Google Scholar] [CrossRef]
- Ring, R.A.; Tesar, D. Cold-Hardiness of the Arctic Beetle, Pytho Americanus Kirby Coleoptera, Pythidae (Salpingidae). J Insect Physiol 1980, 26, 763–774. [Google Scholar] [CrossRef]
- Crosthwaite, J.C.; Sobek, S.; Lyons, D.B.; Bernards, M.A.; Sinclair, B.J. The Overwintering Physiology of the Emerald Ash Borer, Agrilus Planipennis Fairmaire (Coleoptera: Buprestidae). J Insect Physiol 2011, 57, 166–173. [Google Scholar] [CrossRef]
- Feng, Y.; Xu, L.; Tian, B.; Tao, J.; Wang, J.; Zong, S. Cold Hardiness of Asian Longhorned Beetle (Coleoptera: Cerambycidae) Larvae in Different Populations. Environ Entomol 2014, 43, 1419–1426. [Google Scholar] [CrossRef] [PubMed]
- Košťál, V.; Šimek, P. Dynamics of Cold Hardiness, Supercooling and Cryoprotectants in Diapausing and Non-Diapausing Pupae of the Cabbage Root Fly, Delia Radicum L. J Insect Physiol 1995. [Google Scholar] [CrossRef]
- Pruitt, N.L.; Moqueet, N.; Shapiro, C.A. Evidence for a Novel Cryoprotective Protein from Freeze-Tolerant Larvae of the Goldenrod Gall Fly Eurosta Solidaginis. Cryobiology 2007, 54, 125–128. [Google Scholar] [CrossRef] [PubMed]
- Marshall, K.E.; Sinclair, B.J. Repeated Freezing Induces a Trade-off between Cryoprotection and Egg Production in the Goldenrod Gall Fly, Eurosta Solidaginis. Journal of Experimental Biology 2018, 221. [Google Scholar] [CrossRef]
- Yoder, J.A.; Benoit, J.B.; Denlinger, D.L.; Rivers, D.B. Stress-Induced Accumulation of Glycerol in the Flesh Fly, Sarcophaga Bullata: Evidence Indicating Anti-Desiccant and Cryoprotectant Functions of This Polyol and a Role for the Brain in Coordinating the Response. J Insect Physiol 2006, 52, 202–214. [Google Scholar] [CrossRef]
- Vanin, S.; Bubacco, L.; Beltramini, M. SEASONAL VARIATION OF TREHALOSE AND GLYCEROL CONCENTRATIONS IN WINTER SNOW-ACTIVE INSECTS; 2008; Vol. 29;
- BLOCK, W.; ERZINCLIOGLU, Y.Z.; WORLAND, M.R. Cold Resistance in All Life Stages of Two Blowfly Species (Diptera, Calliphoridae). Med Vet Entomol 1990, 4, 213–219. [Google Scholar] [CrossRef]
- Vatanparast, M.; Park, Y. Cold Tolerance Strategies of the Fall Armyworm, Spodoptera Frugiperda (Smith) (Lepidoptera: Noctuidae). Sci Rep 2022, 12. [Google Scholar] [CrossRef]
- Vatanparast, M.; Sajjadian, S.M.; Park, Y. Glycerol Biosynthesis Plays an Essential Role in Mediating Cold Tolerance the Red Imported Fire Ant, Solenopsis Invicta. Arch Insect Biochem Physiol 2022, 109. [Google Scholar] [CrossRef]
- Cha, W.H.; Lee, D. Suppression of Glycerol Biosynthesis-related Genes Decreases the Effect of Rapid Cold Hardening in <scp> Helicoverpa Assulta </Scp>. Entomol Res 2022, 52, 24–32. [Google Scholar] [CrossRef]
- Atapour, M.; Moharramipour, S. Cold Hardiness Process of Beet Armyworm Larvae, Spodoptera Exigua ( Lepidoptera : Noctuidae ). 2014, 3, 147–158.
- Sun, M.; Tang, X.-T.; Lu, M.-X.; Yan, W.-F.; Du, Y.-Z. Cold Tolerance Characteristics and Overwintering Strategy of Sesamia Inferens (Lepidoptera: Noctuidae). Florida Entomologist 2014, 97, 1544–1553. [Google Scholar] [CrossRef]
- Gillyboeuf, N.; Anglade, P.; Lavenseau, L.; Peypelut, L. Cold Hardiness and Overwintering Strategy of the Pink Maize Stalk Borer, Sesamia Nonagrioides Lef (Lepidoptera, Noctuidae). Oecologia 1994, 99, 366–373. [Google Scholar] [CrossRef] [PubMed]
- Andreadis, S.S.; Vryzas, Z.; Papadopoulou-Mourkidou, E.; Savopoulou-Soultani, M. Cold Tolerance of Field-Collected and Laboratory Reared Larvae of Sesamia Nonagrioides (Lepidoptera: Noctuidae). Cryo-Letters 2011, 32, 297–307. [Google Scholar] [PubMed]
- PULLIN, A.S.; BALE, J.S.; FONTAINE, X.L.R. Physiological Aspects of Diapause and Cold Tolerance during Overwintering in Pieris Brassicae. Physiol Entomol 1991, 16, 447–456. [Google Scholar] [CrossRef]
- Stanic, B.; Jovanovic-Galovic, A.; Blagojevic, D.P.; Grubor-Lajsic, G.; Worland, R.; Spasic, M.B. Cold Hardiness in Ostrinia Nubilalis (Lepidoptera: Pyralidae): Glycerol Content, Hexose Monophosphate Shunt Activity, and Antioxidative Defense System. Eur J Entomol 2004, 101, 459–466. [Google Scholar] [CrossRef]
- Andreadis, S.S.; Vryzas, Z.; Papadopoulou-Mourkidou, E.; Savopoulou-Soultani, M. Age-Dependent Changes in Tolerance to Cold and Accumulation of Cryoprotectants in Overwintering and Non-Overwintering Larvae of European Corn Borer Ostrinia Nubilalis. Physiol Entomol 2008, 33, 365–371. [Google Scholar] [CrossRef]
- Kojić, D.; Popović, Ž.D.; Orčić, D.; Purać, J.; Orčić, S.; Vukašinović, E.L.; Nikolić, T. V.; Blagojević, D.P. The Influence of Low Temperature and Diapause Phase on Sugar and Polyol Content in the European Corn Borer Ostrinia Nubilalis (Hbn.). J Insect Physiol 2018, 109, 107–113. [Google Scholar] [CrossRef]
- Khani, A.; Moharramipour, S.; Barzegar, M. Cold Tolerance and Trehalose Accumulation in Overwintering Larvae of the Codling Moth, Cydia Pomonella (Lepidoptera: Tortricidae). Eur J Entomol 2007, 104, 385–392. [Google Scholar] [CrossRef]
- Rozsypal, J.; Koštál, V.; Zahradníčková, H.; Šimek, P. Overwintering Strategy and Mechanisms of Cold Tolerance in the Codling Moth (Cydia Pomonella). PLoS One 2013, 8. [Google Scholar] [CrossRef]
- Li, N.G. Relationships between Cold Hardiness, and Ice Nucleating Activity, Glycerol and Protein Contents in the Hemolymph of Caterpillars, Aporia Crataegi L. Cryo-Letters 2012, 33, 135–143. [Google Scholar]
- Marshall, K.E.; Sinclair, B.J. The Relative Importance of Number, Duration and Intensity of Cold Stress Events in Determining Survival and Energetics of an Overwintering Insect. Funct Ecol 2015, 29, 357–366. [Google Scholar] [CrossRef]
- Strassmann, J.E.; Lee, R.E.; Rojas, R.R.; Baust, J.G. Caste and Sex Differences in Cold-Hardiness in the Social Wasps,Polistes Annularis AndP. Exclamans (Hymenoptera: Vespidae). Insectes Soc 1984, 31, 291–301. [Google Scholar] [CrossRef]
- Sømme, L. EFFECTS OF GLYCEROL ON COLD-HARDINESS IN INSECTS. Can J Zool 1964, 42, 87–101. [Google Scholar] [CrossRef]
- Song, Y.; Huang, W.; Zhou, Y.; Li, Z.; Ji, R.; Ye, X. Physiological Characteristics and Cold Tolerance of Overwintering Eggs in Gomphocerus Sibiricus L. (Orthoptera: Acrididae). Arch Insect Biochem Physiol 2021, 108. [Google Scholar] [CrossRef] [PubMed]
- Toxopeus, J.; Lebenzon, J.E.; McKinnon, A.H.; Sinclair, B.J. Freeze Tolerance of Cyphoderris Monstrosa (Orthoptera: Prophalangopsidae). Canadian Entomologist 2016, 148, 668–672. [Google Scholar] [CrossRef]
- Saeidi, F.; Moharramipour, S.; Barzegar, M. Seasonal Patterns of Cold Hardiness and Cryoprotectant Profiles in <I>Brevicoryne Brassicae</I> (Hemiptera: Aphididae). Environ Entomol 2012, 41, 1638–1643. [Google Scholar] [CrossRef]
- Wang, Z.; Chen, Y.; Diaz, R.; Laine, R.A. Physiology of Crapemyrtle Bark Scale, Acanthococcus Lagerstroemiae (Kuwana), Associated with Seasonally Altered Cold Tolerance. J Insect Physiol 2019, 112, 1–8. [Google Scholar] [CrossRef]
- Sadeghi, R.; Izadi, H.; Mahdian, K. Energy Allocation Changes in Overwintering Adults of the Common Pistachio Psylla, Agonoscena Pistaciae Burckhardt & Lauterer (Hemiptera: Psyllidae). Neotrop Entomol 2012, 41, 493–498. [Google Scholar] [CrossRef]
- Fuller, B.J. Cryoprotectants: The Essential Antifreezes to Protect Life in the Frozen State. Cryo-Letters 2004, 25, 375–388. [Google Scholar]
- Elliott, G.D.; Wang, S.; Fuller, B.J. Cryoprotectants: A Review of the Actions and Applications of Cryoprotective Solutes That Modulate Cell Recovery from Ultra-Low Temperatures. Cryobiology 2017, 76, 74–91. [Google Scholar] [CrossRef]
Figure 1.
Mode of action of cryoprotectants.
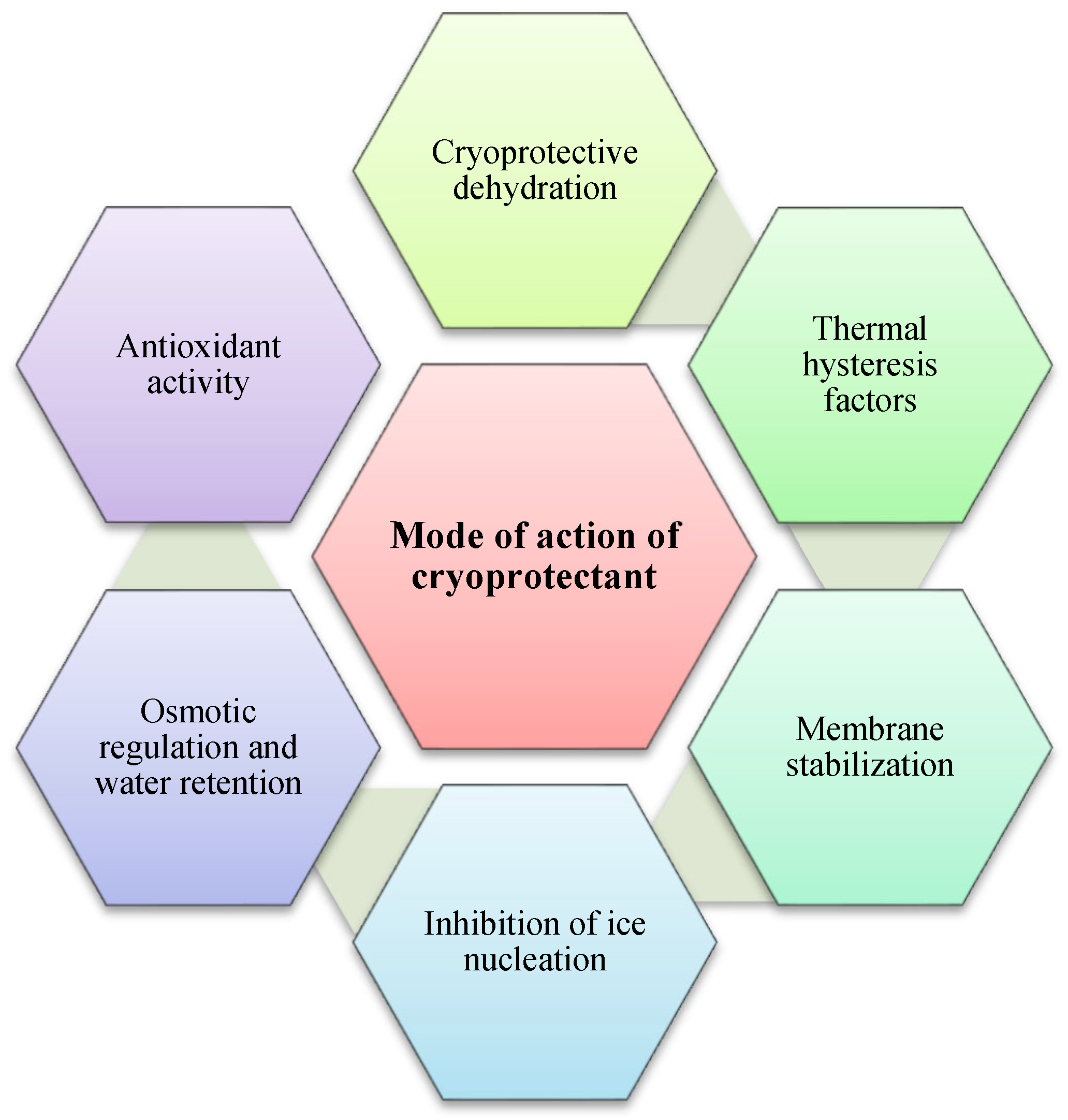
Figure 2.
A. Colligative effects of cryoprotectants. B. Non-colligative effects of cryoprotectants.
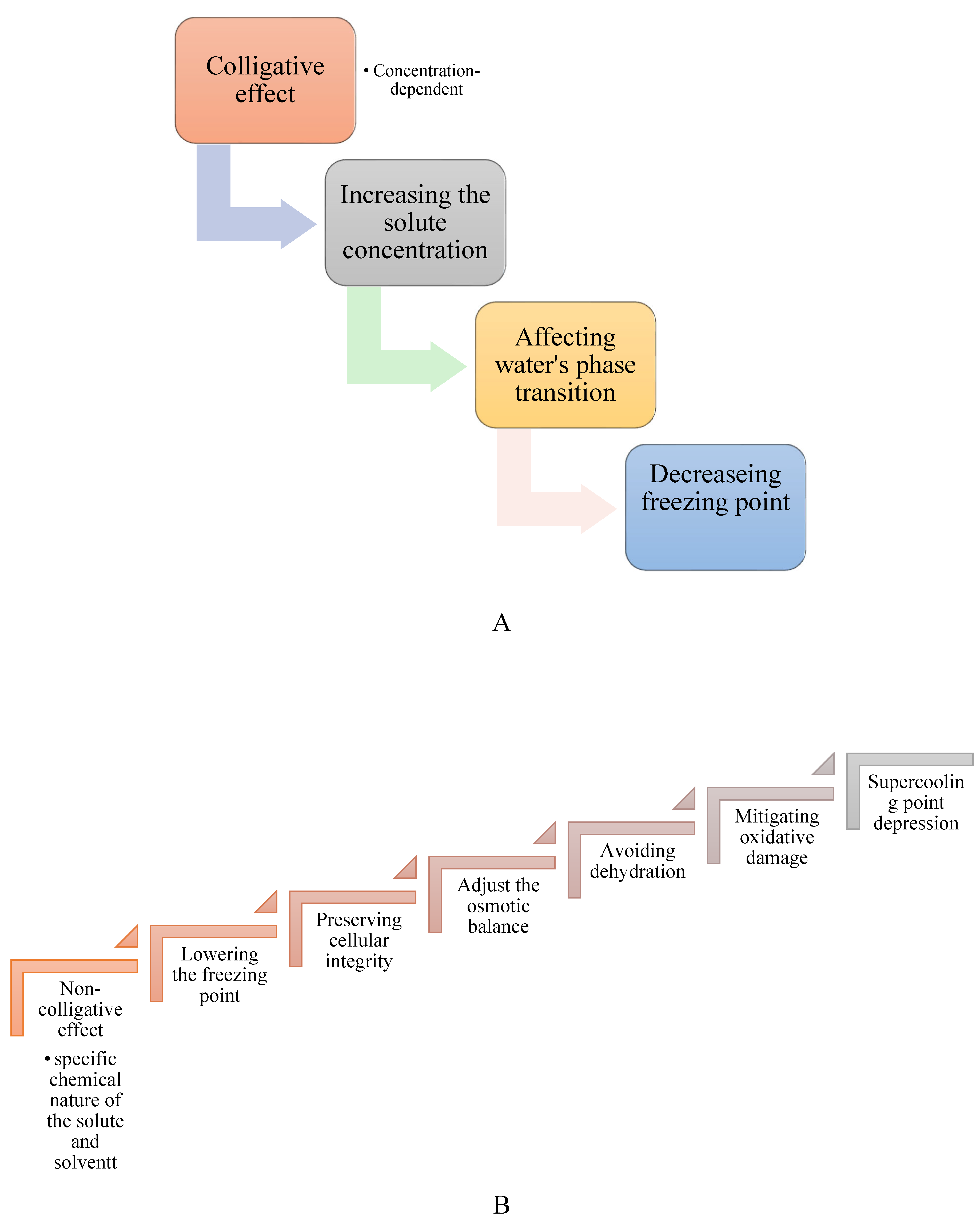
Figure 3.
The primary classification of insects' cryoprotectants. In this figure, cryoprotectants have been classified: 1- based on their structure, 2- based on their location in the cell (extracellular and intracellular), and 3- based on their behavior in a solution (colligative and non-colligative). LMWC: Low molecular-weight carbohydrates [25,31,169,170].
Figure 3.
The primary classification of insects' cryoprotectants. In this figure, cryoprotectants have been classified: 1- based on their structure, 2- based on their location in the cell (extracellular and intracellular), and 3- based on their behavior in a solution (colligative and non-colligative). LMWC: Low molecular-weight carbohydrates [25,31,169,170].

Figure 4.
Predominant cryoprotectants of insects.
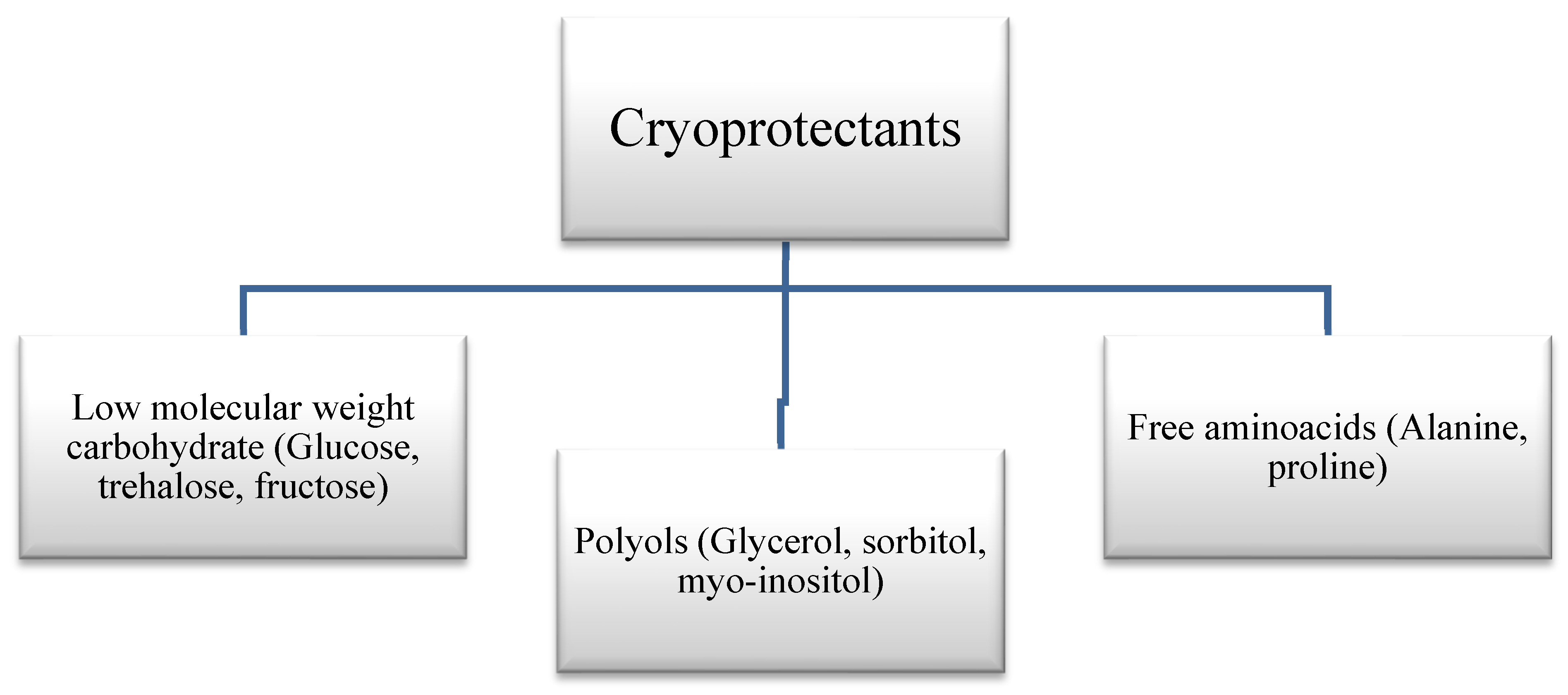
Figure 5.
Mode of action of glycerol as a cryoprotectant.
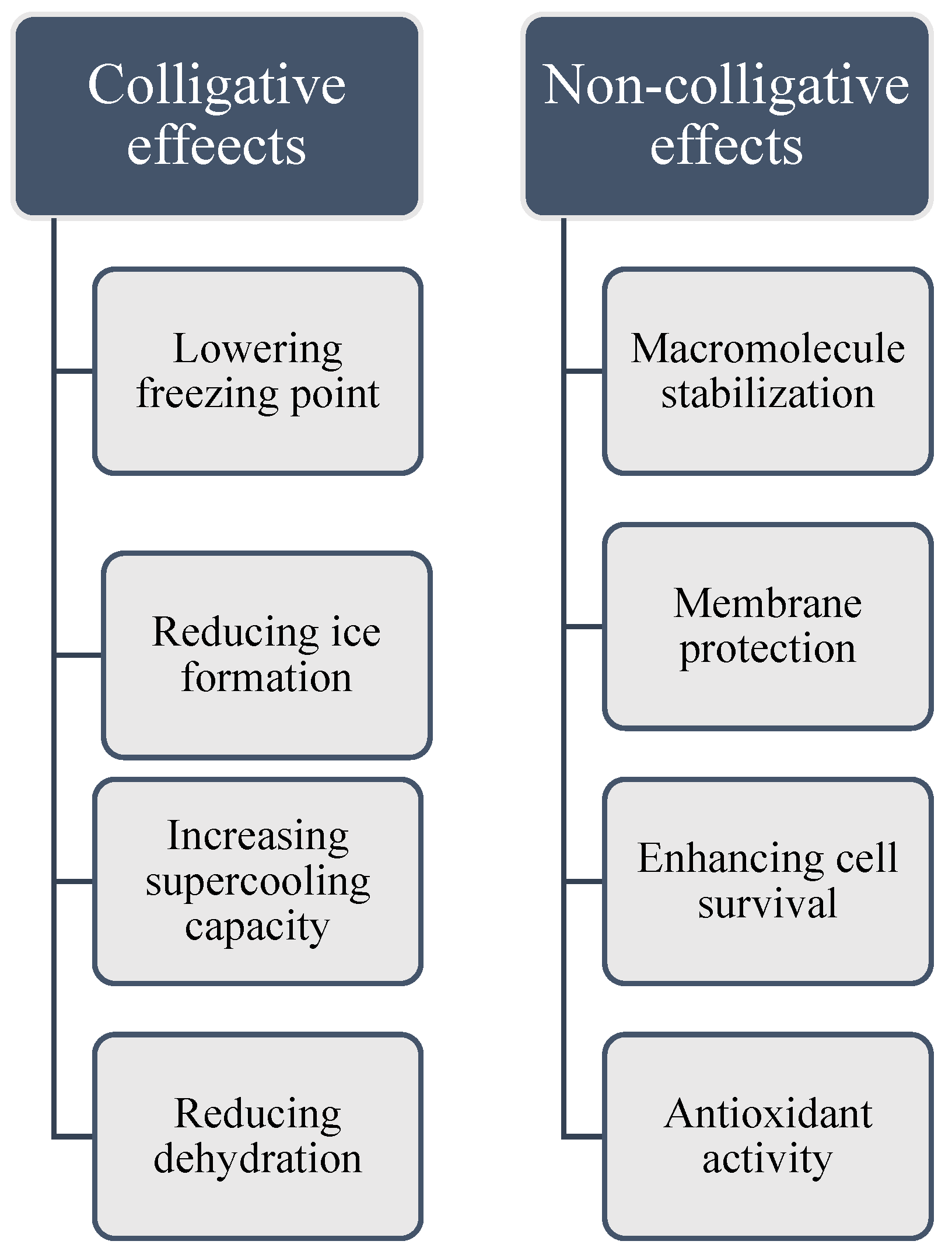
Figure 6.
Mode of action of trehalose as a cryoprotectant. Trehalose is an extraordinary molecule with a range of cryoprotective functions that enable many insects to endure extreme freezing conditions. Its capacity to safeguard cellular components from various forms of cryoinjury is essential for insect survival during harsh winters.
Figure 6.
Mode of action of trehalose as a cryoprotectant. Trehalose is an extraordinary molecule with a range of cryoprotective functions that enable many insects to endure extreme freezing conditions. Its capacity to safeguard cellular components from various forms of cryoinjury is essential for insect survival during harsh winters.
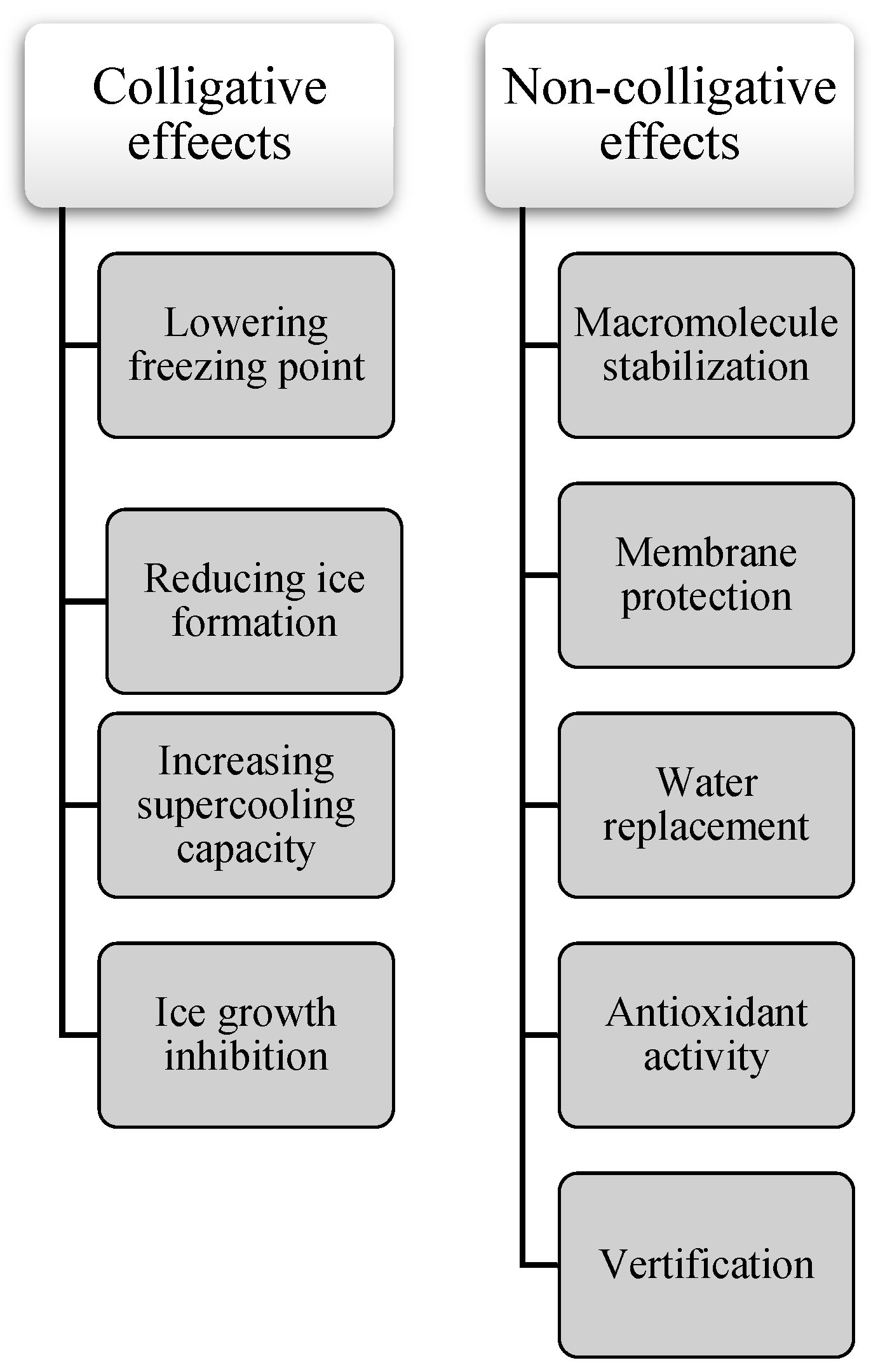
Figure 7.
Mode of action of glucose as a cryoprotectant.
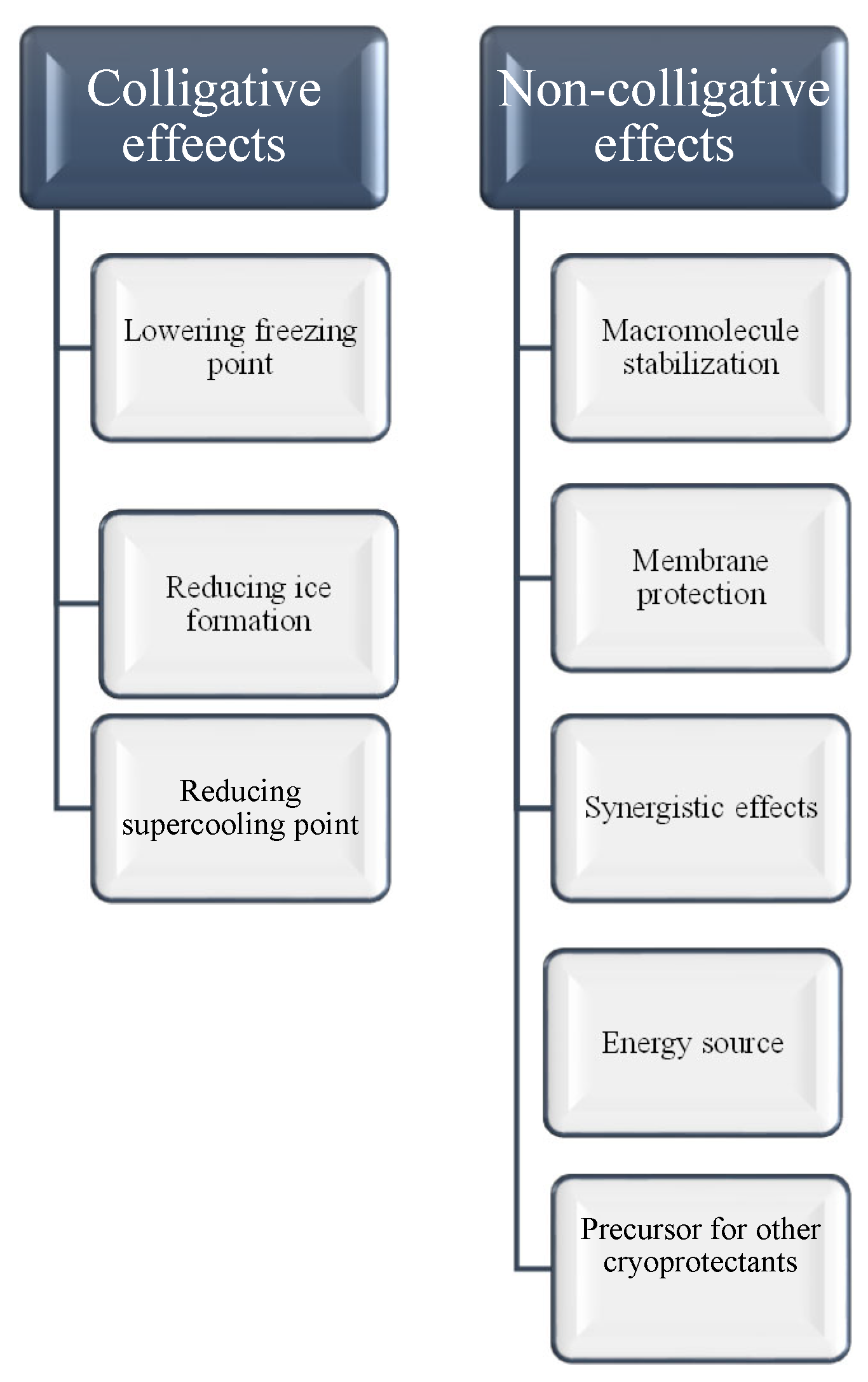

Table 1.
A comparison between cold tolerance and cold hardiness.
| Feature | Cold tolerance | Cold hardiness |
|---|---|---|
| Definition | The ability of insects to survive and function at low temperatures in the short term. | The ability to resist long-term exposure to low temperatures and survive freezing temperatures |
| Freezing | May or may not involve freezing | Necessarily involves dealing with freezing |
| Mechanisms | Behavioral and physiological (supercooling and cryoprotectants) adaptations. | Involves specialized adaptations such as cold acclimation, rapid cold hardening freeze avoidance, and freeze tolerance |
Table 2.
Mode of action of ice nucleating proteins and antifreeze proteins as a cryoprotectant.
| Feature | Ice nucleating proteins (INPs) | Antifreeze proteins (AFPs) |
|---|---|---|
| Primary function | Induce ice formation | Prevent ice formation |
| Mechanism of action | Promote nucleation at high subzero temperatures | Bind to existing ice crystals to inhibit growth |
| Role in cold tolerance | Control location and timing of ice formation | Stabilize cells and prevent recrystallization |
| Adaptation strategy | Essential for freeze-tolerant species | Common in freeze-avoiding species |
| Seasonal variation | May be removed or masked during winter | Often expressed year-round to prevent freezing |
Table 3.
Predominant cryoprotectants of different insect species.
| Order | Family | Species | Predominant cryoprotectant | Reference |
|---|---|---|---|---|
| Coleoptera | Dermestidae | Trogoderma granarium | Trehalose, sorbitol, myo-inositol, glucose | [125] |
| Chrysomelidae | Xanthogaleruca luteola | Myo-inositol, trehalose, glucose | [126,127,128] | |
| Ophraella communa | Glycerol | [129] | ||
| Chrysomela aeneicollis | Glycerol | [130] | ||
| Aulacophora nigripennis | Myo-inositol | [131] | ||
| Galerucella nymphaea | Glycerol | [122] | ||
| Chrysolina graminis | Glycerol | |||
| Curculionidae | Hypera postica | Sorbitol, glycerol, glucose, trehalose | [132] | |
| Pityogenes chalcographus | Glycerol | [133] | ||
| Lissorhoptrus oryzophilus | Trehalose | [134] | ||
| Sitophilus granarius | Trehalose | [135] | ||
| Dendroctonus ponderosae | Glycerol | [136] | ||
| Upis ceramboides | Sorbitol, threitol | [83] | ||
| Hylurgus ligniperda | Sorbitol, trehalose, glycerol | [137] | ||
| Tenebrionidae | Tribolium castaneum | Trehalose, myo-inositol | [87] | |
| Pythidae | Pytho depressus | Glycerol | [30] | |
| P. americanus | Glycerol | [138] | ||
| Buprestidae | Agrilus planipennis | Glycerol | [139] | |
| Laemophloeidae | Cryptolestes ferrugineus | Trehalose | [135] | |
| Cerambycidae | Anoplophora glabripennis | Glycerol, glucose, sorbitol, trehalose | [140] | |
| Coccinellidae | Hippodamia variegata | Trehalose, glucose, myo-inositol | [104,105] | |
| Harmonia axyridis | Myo-inositol | [81] | ||
| Diptera | Anthomyiidae | Delia antiqua | Trehalose, glucose, myo-inositol | [141] |
| Drosophilidae | Drosophila melanogaster | Myo-inositol, proline | [79] | |
| Tephritidae | Eurosta solidagnis | Carbohydrates and proteins | [142,143] | |
| Sarchophagidae | Sarcophaga bullata | Glycerol | [144] | |
| Limoniidae | Chionea sp. | Trehalose | [145] | |
| Tephritidae | Bactrocera dorsalis | Proteins, glycerol, trehalose | [86] | |
| Hippoboscidae | Lipoptena cervi | Proline, arginine, asparagine, cystine, glutamate, glutamine | [124] | |
| Sarcophagidae | Sarcophaga bullata | Glycerol | [144] | |
| Calliphoridae | Calliphora vicina | Glucose | [146] | |
| C. vomitoria | ||||
| Lepidoptera | Noctuidae | Spodoptera frugiperda | Glycerol | [147,148] |
| Chilo suppressalis | Glycerol | [31] | ||
| Helicoverpa assulta | Glycerol | [149] | ||
| Ectomyelois ceratoniae | Trehalose, myo-inositol | [106] | ||
| Arimania comaroffi | Trehalose, myo-inositol, sorbitol | [9] | ||
| Spodoptera exigua | Trehalose, glycerol, myo-inositol | [150] | ||
| Sesamia inferens | Trehalose, glycerol, glucose, fructose, myo-inositol | [151] | ||
| S. nonagrioides | Trehalose, glycerol | [152,153] | ||
| Pieris brassicae | Trehalose, myo-inositol, sorbitol | [154] | ||
| Ostrinia nubilalis | Trehalose, glycerol | [155,156,157] | ||
| Cydia pomonella | Trehalose, glucose, glycerol, | [158,159] | ||
| Aporia crataegi | Glycerol | [160] | ||
| Choristoneura fumiferana | Glycerol | [161] | ||
| Hymenoptera | Formicidae | Solenopsis invicta | Glycerol | [147,148] |
| Polistes exclamans | Glycerol | [31,162] | ||
| Braconidae | Bracon cephi | Glycerol | [163] | |
| Orthoptera | Gryllidae | Gryllus veletis | Glycerol7 | [31,162] |
| Acrididae | Gomphocerus sibiricus | Glycerol, fructose, sorbitol, amino acids | [164] | |
| Prophalangopsidae | Cyphoderris monstrosa | Trehalose, proline | [165] | |
| Heteroptera | Aphididae | Schizaphis graminum | Trehalose, glucose | [166] |
| Eriococcidae | Acanthococcus lagerstroemiae | D-mannitol | [167] | |
| Scutelleridae | Eurygaster integriceps | Glycerol, trehalose | [91] | |
| Psyllidae | Agonoscena pistaciae | Trehalose, myoinositol, sorbitol | [168] | |
| Aphididae | Cinara tujafilina | Glucose, trehalose, mannitol, myo-inositol, glycerol | [109] |
Table 4.
Time-dependent cryoprotectant accumulation strategies of insects.
| Strategy | Trigger | Example Species | Environment |
|---|---|---|---|
| Rapid (Anticipatory) | Photoperiod/temperature | Goldenrod gall fly, Eurosta solidaginis, Fire-colored beetle, Dendroides canadensis | Temperate seasonal |
| Incremental (Acclimatory) | Gradual cooling | Woolly bear caterpillar, Pyrrharctia isabella, Lady beetle, Coccinella septempunctata | Unpredictable cold |
| Basal (Constitutive) | Genetic adaptation | Antarctic midge, Belgica antarctica, Arctic springtail, Megaphorura arctica | Polar/alpine |
Table 5.
A comparison between insects and other ectotherms in cryoprotectant accumulation.
| Feature | Insects | Other ectotherms |
|---|---|---|
| Cryoprotectant diversity | High (polyols, amino acids, sugars) | Moderate (mainly glycerol, glucose) |
| Freeze tolerance mechanism | Intracellular freezing (select species) | Extracellular freezing only |
| Cryoprotectant sources | Internal reserves and dietary intake | Predominantly glycogen-derived |
| Adaptation | Rapid accumulation, diapause-linked | Gradual accumulation, seasonal |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



