Submitted:
10 March 2025
Posted:
11 March 2025
You are already at the latest version
Abstract
Background/Objectives: The risk of introduction of highly pathogenic avian influenza virus (HPAIV) in geese breeding and fattening flocks is heightened due to the necessity of free-range access to grazing grounds. This study aimed to evaluate the safety, immunogenicity, and protective efficacy of five commercial vaccines against HPAIV subtype H5N1 (clade 2.3.4.4b) in subadult fattening geese. Methods: A prime-boost vaccination trial was conducted using five commercial vaccines, including H5 expressing vaccines of novel technology (subunit, vector, RNA) and whole inactivated virus (WIV) vaccines. Based on serological results, one RNA and one WIV vaccine were selected for a homologous challenge experiment. Results: Two vaccines of novel technology (vector, RNA) required a booster dose to raise specific antibodies titers above a threshold of 4 log2 by hemagglutination inhibition (HI) assay, whereas a subunit vaccine and two WIV vaccines induced seroconversion after primary vaccination. In the challenge experiment, all non-vaccinated control geese succumbed to infection by day 4. In contrast, all vaccinated geese that had seroconverted exhibited full clinical protection. Although sterile immunity was not achieved, viral excretion was significantly reduced in the vaccinated groups compared to controls. Conclusions: Vaccination substantially mitigated the impact of HPAIV H5N1, clade 2.3.4.4b infection in geese, greatly improving animal welfare by preventing severe disease. Additionally, there was a significant reduction in viral burden. Further studies are necessary to verify the potential of these vaccines to reduce susceptibility to infection and virus excretion in order to achieve suppression of the between-flock reproduction number to below 1 in geese flocks at high risk of infection.
Keywords:
vaccination
; free range
; poultry
; protection
; HPAIV
; H5N1
; goose
1. Introduction
The ongoing pandemic of a highly pathogenic avian influenza virus (HPAIV) of subtype H5N1 (goose/Guangdong lineage [gs/GD]), clade 2.3.4.4b, is posing an unprecedented threat, impacting both industrial and small scale poultry operations [1,2]. Similarly, HPAIV infections in wild birds, particularly those leading to mass mortalities among colony-breeding species, are exerting significant pressure on avian biodiversity [3,4]. The recent increase in spillover infections of HPAIV H5N1 clade 2.3.4.4b to wild mammals and livestock (e.g. dairy cows) further expands the interface of exposure between animals and humans [5]. The increased risk of zoonotic transmission to humans has prompted reconsideration of preventive measures against HPAIV infections in animals.
In Europe, the control of HPAI in poultry and other captive birds has traditionally relied on enhanced biosecurity measures to prevent virus introduction from the environment or wild bird populations [6]. These measures also aim to limit virus spread from infected poultry enterprises and are supported by highly effective diagnostic systems and surveillance strategies for early disease detection [7]. Upon confirmation of infection, immediate restriction measures are enforced, including the culling of affected and suspect flocks, holding closures, restocking bans, and regional trade restrictions. These principles have successfully prevented HPAIV from becoming enzootic in captive birds in Europe for nearly 15 years. However, the current panzootic spread of the clade 2.3.4.4b virus and its established year-round presence in wild birds in Europe have let to EU legislation establishing a legal basis for HPAIV vaccination as an additional preventing measure (EU 2023/361) [8,9].
According to fundamental virological knowledge and experience gained from vaccination campaigns in non-European countries, the success of poultry vaccination programs depends on two critical factors: (i) the antigenic similarity between the vaccine virus strains and the HPAIV strains currently circulating in the field, and (ii) the rigorous monitoring of the vaccinated herds [10,11,12]. Employing vaccines that antigenically match circulating field viruses provides superior efficacy and should limit the risk of emerging virus escape variants [13]. Close-meshed surveillance activities aim at early detection of HPAIV circulating stealthily in vaccinated herds that appear clinically healthy.
In Europe, practical experience with HPAI vaccinations is limited and further insight is needed into the appropriate use of diagnostics that are fit-for-purpose in vaccinated flocks. Vaccination has been conducted across Europe in various poultry species using both laboratory-based approaches (Italy – turkeys, Netherlands – laying hens) and field studies (France – fattening ducks) [14,15,16]. Although geese are a minor poultry species economically, their production can be locally significant. Breeding geese requires sufficient free-range access, increasing the risk of HPAIV introduction. Despite the relatively small number of geese farms, they have been disproportionally affected by HPAIV outbreaks in Europe [11]. Therefore, additional protection through vaccination could have a major impact in geese as a particularly exposed sector of poultry production.
A laboratory-based pilot trial was conducted to assess the immunogenicity and efficacy of selected commercial, though not yet licensed, vaccines in geese. Additionally, various diagnostic tools were evaluated for the surveillance of vaccinated flocks.
2. Materials and Methods
2.1. Ethics Statement
All animal experiments were approved by an independent ethics committee of the German Federal State of Mecklenburg-Vorpommerania (LALLF 7221.3-1-037/23-1). All animal trials were conducted in biosafety level 3 (BSL3) animal facilities of the Friedrich-Loeffler-Institute, the Isle of Riems, Greifswald.
2.2. Animal Experiments
Three-week-old goslings, seronegative for AIV-specific antibodies and negative in combined oropharyngeal and cloacal swabs by generic M-gene-specific real time RT-PCR (RT-qPCR) of a commercial breed of German laying geese were obtained from a commercial hatchery and housed in stables at Friedrich-Loeffler-Institute (FLI) with continuous access to dry bedding material and bathing facilities. Feed and water were provided ad libitum. After a three-week acclimatization, the animals were randomly separated into six groups of ten birds each (Table 1). Throughout the experiments, the animals were supervised at least twice daily by veterinary staff and animal caretakers. Continuous video supervision and a remote-controlled light monitoring regime were also enabled. Any clinical signs of disease occurring before, during, or after the vaccination, as well as those resulting from the challenge infection in two selected groups, were fully documented. Humane endpoints were defined for significant clinical deterioration, including severe neurological deficits and/or loss of vital functions. Animals that reached these endpoints were first anesthetized with combination of Ketamine-Xylazin (4.4 mg/kg Ketanest®, and 2.2 mg/kg Xylavet®) injected into the caudal femoral muscle. Unconscious animals were then euthanized by exsanguination through opening the Sinus occipitalis and subsequent dislocation of the neck.
2.3. Vaccines
None of the vaccines employed had been licensed for use in Europe in geese. The vaccines were provided by the producers under Material Transfer Agreement (MTA) and Non-Disclosure Agreements (NDA) for exclusive use at FLI in the trial. The vaccines were kept at FLI under the recommended storage conditions. Vaccines 1-4 expressed a hemagglutinin (HA) H5 of clade 2.3.4.4, while vaccine 5 consisted of baculovirus-expressed H5 with an altered amino acid sequence for broader protection [17].
2.4. Experimental Design
After a three-week acclimatization, the animals were randomly separated into six groups of ten birds each (five vaccine groups + one non-immunized control group) (Table 1). Geese were immunized at six weeks of age with one of the vaccines (Table 1; Figure 1). Due to a delayed disposability, geese of the #BI group were vaccinated in the tenth week of age. A booster vaccination was administered four weeks later at ten weeks of age and at 14 wekks of age for the #BI group, respectively. The HPAIV H5N1 challenge was conducted three weeks following booster immunization.
2.5. Virus
For challenge experiments, a clade 2.3.4.4b duck pathogenic HPAIV strain (HPAIV H5N1 A/chicken/Germany-NI/AI 4286/2022) was used (see Table 2A) [18]. The viral stock was derived from the amnion allantoic fluid of inoculated specific pathogen-free (SPF) embryonated chicken eggs. The virus titer was determined in cell culture (immortalized chicken hepatocyte cell line LMH [ATCC: CRL-2117]) as well as in embryonated SPF chicken eggs, both in triplicates and calculated by Reed and Muench’s method [19]. Additional viruses used for serological purposes in this study are listed in Table 2.
2.6. Serology
Heparinized blood samples were collected from all geese at indicated timepoints (Figure 1) i.e. before prime immunization, before booster vaccination, three weeks after booster vaccination (prior to challenge) and 14 days after the challenge infection at the end of the observation period. Plasma samples were heat inactivated (56°C, 30 min) and tested by two commercial ELISA test kits for Influenza A-specific antibodies against the NP or H5 protein (ID screen® Influenza A Antibody Competition Multi-species ELISA; ID screen® Influenza competitive subtype-specific H5 kit) according to the manufacturer’s instructions.
Plasma samples were also tested by hemagglutination inhibition (HI) assay, serum neutralization test (SNT), using antigens listed in Table 2. For HI assays, geese plasma was pre-absorbed with 10% (v/v) chicken erythrocytes (37°C, 30 min) and subsequently tested according to standard procedures using chicken red blood cells [20,21].
The SNT was performed in a 96-well plate format, producing a serial twofold dilution of the plasma samples. Each 100 µl of plasma dilution was mixed with 100 µl of prediluted virus containing 400 TCID50. After incubation for 30 minutes at 37°C, a triplicate of 50 µl from each dilution series was transferred to a 96 well plate with a preformed monolayer of the LMH cells. After an 72-hour incubation period at 37°C and 5% CO2, the plates were analyzed by light microscopy for viral cytopathic effects.
Arithmetic mean titers and standard deviations (stdv) were calculated from all individuals with positive HI results (>3 [log2]) according to the vaccination schedule. Arithmetic means were calculated using Microsoft Excel (Microsoft Corporation. (2018). Microsoft Excel. Retrieved from https://office.microsoft.com/excel) formula (=MITTELWERT(range)), where 'range' referred to the selected data set. The standard deviation was calculated using (=STABW.N(range)) for the sample standard deviation, where 'range' represents the corresponding data range.
2.7. Challenge Experiment
Protection was assessed by challenge experiments using 10 geese per vaccine group. To minimize the number of animals in potentially strenuous challenge trials, two vaccine groups were preselected based on their average serological response by HI against the challenge virus after booster immunization (Figure 2). Additionally, a mock-vaccinated control group of 10 geese was established. A total of 30 geese were inoculated oculo-nasally with 0.5 ml/nostril HPAIV H5N1 A/chicken/Germany-NI/AI 4286/2022 with a titer of 106 TCID50/0.5 ml (Table 2A). Daily observations were conducted for up to two weeks. A clinical scoring system (as calculated according to the established IVPI for pathotyping of HPAIV [21]) was used, and humane endpoints were pre-defined for the development of severe disease signs expected to inevitably lead to the death of the animals. These included considerable apathy and lethargy in affected individuals, as well as disorders of the locomotive and central nervous system, disorders of the respiratory tract, manifesting as gasping for breath or repeated (<2 following days) minor clinical disturbances, including mild depression and delayed feed intake. Each animal on trial had swab samples collected from the oropharynx and cloaca at days 2, 3, 4, 5, 7, 10 and 14 post-challenge. Swabs were transferred into tubes containing 1 ml cell culture medium supplemented with Enrofloxacin (Baytril®; 20µg/ml) and stored at -70°C until further analysis. Tissue samples of brain, lung, duodenum/pancreas, kidney, liver and heart obtained at post mortems of geese that died within the observation period were collected for virological examination into tubes containing 1 ml of cell culture medium with Enrofloxacin (Baytril®; 20µg/ml) or 10% buffered paraformaldehyde.
2.8. Real-Time RT-PCR
RNA was extracted from swab samples and tissues as described elsewhere [18,22] using the Macherey-Nagel NucleoMag® VET-Kit on a Biosprint 96 extraction robot (Qiagen, Hilden, Germany) and eluted in 100 µl elution buffer. Subsequently, RNA extracts were tested by a generic real-time RT-PCR (RT-qPCR) for the presence of influenza A virus matrix gene fragment using the AgPath ID One-Step RT-PCR Kit (Ambion-Applied Biosystems) [7]. PCR tests were carried out in a CFX96™Real-Time-System C1000™ thermal cycler (BioRad, Munich, Germany). Estimation of virus genome equivalents (VE) in each sample was based on correlation of the individual Cq values to an intra-assay calibration curve of a defined HPAIV H5N1 virus stock with known TCID50 infectivity titers.
2.9. Virus Isolation from Clinical Materials
To test for infectivity in swab samples, LMH cells were seeded in 24-well cell culture plates with 10% fetal calf serum (FCS) and 10% Baytril® (Enroflocaxine). Examination of oropharyngeal swabs only was chosen because of the reduced load of bacterial contamination compared to cloacal swabs. Following a 24-hour incubation period at 37°C and 5% CO₂, 100 µl of each of the 1:10 pre-diluted supernatants of oropharyngeal swabs, filtered through a 0.22 µm filter (Millex® Syringe-driven Filter Unit-0.22 µm) (SigmaAldrich), were infected in duplicates onto a confluent monolayer of LMH cells. Microscopic examination of cytopathic effect (CPE), manifested as cell necrosis and plaque formation, was conducted after 72 hours incubation at 37°C and 5% CO₂. Subsequently, the supernatants from cells showing CPE were examined for virus-induced hemagglutination ability in a hemagglutination assay (HA). A CPE associated with an HA titer >3 log2 in the supernatant was considered an indication for residual infectivity. In the case of inconclusive CPE and HA in the first cell passage, a second cell passage on LMH cells was conducted, followed by further HA activity investigation.
2.10. Statistics
Differences between antibody titers within the groups and assay methods at various time points were determined using two-tailed Mann-Whitney-U tests with the statistical program GraphPad Prism in version 10.3.1 for Windows, GraphPad Software, Boston, Massachusetts USA, www.graphpad.com. Differences on oropharyngeal virus excretion between the groups subjected to challenge infection were statistically evaluated using an unpaired two-tailed Mann-Whitney-U test and Kruskal-Wallis test, respectively, with help of the statistical analysis tool SigmaPlot™ version 11 (SigmaPlot Software, Grafiti LLC, Palo Alto, CA, USA). The total excretion of virus per animal was determined by calculating the area under the curve (AUC) for each animal individually and as a sum of all animals per group. Comparisons between vaccinated and control groups were limited to 1-4 days post infection (dpi). The AUC of excretion data was determined using the software package R, version 4.3.2 (RStudio Team. (2023). RStudio: Integrated Development Environment for R. RStudio, PBC). Results were analyzed and visualized using the R packages “pracma”, “dplyr” and “ggplot2”.
3. Results
3.1. Safety and Immunogenicity
Prior to vaccination, all geese tested seronegative for AI virus-specific antibodies by ELISA, HI and SNT. Combined oropharyngeal/ cloacal swab samples were negative for AI virus-specific RNA by RT-qPCR (data not shown). Vaccination did not induce adverse local or systemic signs in any of the geese.
Seroconversion in vaccinated groups depended on type of vaccine and diagnostic assay. All geese of the non-vaccinated control group remained seronegative in all assays applied (for details of serological tests see Supplementary Tables S1–S4).
3.1.1. AI Virus H5-ELISA:
The H5 antibody response was assessed using a commercial H5 competition ELISA, where a positive response was defined as having residual reactivity below 50%. Seroconversion after prime immunization was detected only in the group #BI, with nine out of ten geese showing a positive response (Figure 2A). Booster immunization further increased H5-ELISA titers in all tested geese within this group. In the groups #Zoe and #Avi-2, only two geese in each group tested positive for H5-antibodies following first vaccine dose. Booster vaccination increased the number of geese with positive H5-antibody response to seven in group #Zoe and eight in group #Avi-2. The H5-antibody response in groups #Ceva and #Avi-1 was undetectable after primary immunization, however it increased after booster vaccination, with half of the animals (n=5) of each group showing a positive response. One and two geese in the #Ceva and #Avi-1 groups, respectively, did not seroconvert even after the booster vaccination.
3.1.2. AI Virus NP-ELISA:
Geese immunized with novel technology vaccines from groups #Ceva, #Avi-2 and #BI, that lacked AI virus nucleoprotein (NP) and other AI virus proteins except HA, did not develop detectable NP-specific antibodies after the primary and booster vaccination. In contrast, all geese vaccinated with whole virus antigens (groups #Zoe and #Avi-1) showed NP-specific reactivity already after the primary vaccination (Figure 2B).
3.1.3. HI Assay
Plasma samples were tested by HI assay using an antigen homologous to the challenge virus HPAIV H5N1 A/chicken/Germany-NI/AI 4286/2022. Seroconversion after prime immunization was detected only in the groups #Zoe and #BI with HI arithmetic mean titers of 5.1±0.7(stdv)[log2] and 4.3±1.4 [log2] (for details see Supplementary Table S3). Booster vaccination further increased HI titers to 8.1±2.8 [log2] in #Ceva and 7.0±1.5 [log2] in the #Zoe group, developing the highest mean HI-titers. Primary vaccination induced antibody titers exceeding HI titers of 3 [log2] in three out of ten geese in both groups #Ceva and #Avi-1 (Figure 2C; Supplementary Figure S1). Booster vaccination increased the number of geese with positive H5-antibody response to eight in group #Ceva (8.1±2.8 [log2]) and ten in group #Zoe (7.0±1.5 [log2]). However, two animals from the #Ceva group remained seronegative by HI. Vaccines #Avi-1 and #BI induced HI arithmetic mean titers of 5.3±1.7(stdv)[log2] (see Supplementary Table S3 for statistical data) and 4.9±1.2 [log2], respectively. No HI titers were detected in the group #Avi-2 after either prime or booster vaccination. Antibody cross-reactivity was further investigated, using another H5N1 clade 2.3.4.4b antigen from 2021, as well as an antigen from a H5N1 virus clade 1.0 heterologous to the challenge virus (see Table 2B; Supplementary Table S3; Supplementary Figure S1). This analysis only included groups that reacted positive (>3 [log2]) to homologous HI assay antigen, thereby excluding #Avi-2. This reactivity was notable already after prime immunization in individual birds and only in geese that were reactive with the clade 2.3.4.4 antigen (see Supplementary Table S3). The reactivity to clade 1.0 virus was most abundant for the #BI vaccinated group (n=8) and increased after booster immunization from 6.8±1.3 [log2] to 8.1±1.5 [log2], clearly exceeding titers against clade 2.3.4.4b antigens from 2022 and 2021, (deviation from other groups tested: p<0.001-0.0001). In the #Ceva group, number of geese reactive with the clade 1.0 antigen increased from three after prime to six after booster vaccination. In group #Zoe, all ten geese initially reacted but showed no booster-induced increase in antibody titers. In group #Avi-1, only 3 geese had an initial reactivity, with no booster-associated increase. In these latter three groups, reactivity was superior against clade 2.3.4.4b antigen (Figure 3; Supplementary Figure S1; Supplementary Table S3).
HI-reactivity was also tested by employing an NDV antigen. All vaccines that contained NDV components, i.e. group #Avi-2 and #BI, induced NDV-specific antibodies already after prime immunization (HI-titer: #Avi-2 8.3±1.7 [log2]; #BI 8.2±2.7 [log2]). These titers increased after booster vaccination (HI-titer: #Avi-2 9.9±1.6 [log2]; #BI 9.3±1.6 [log2]) (deviation to other groups tested: p<0.0001) (Figure 2D; Supplementary Table S3). In contrast, #Ceva, #Zoe and Avi-1 remained HI-negative (<2 [log2]) in NDV HI since they did not contain this antigen (Figure 2D; Figure 3; Supplementary Table S3).
3.1.4. SNT Assay
All groups were tested for their seroneutralization activity against the 2.3.4.4b antigen, homologous to the challenge virus. The results corroborate those of the HI assay. Vaccines #Zoe (SNT arithmetic mean 3.4±1.2 [log2], #Avi-1 and #BI (both 2.3±0.6 [log2])) induced measurable SNT-titers after first dose vaccination. In contrast, no SNT reactivity was observed after priming with the #Ceva vaccine (deviation #Ceva to other groups tested: p<0.0001). Booster immunization induced an increase of SNT-titers across all tested groups except #Avi-2: After the booster, the highest SNT-titers were observed in groups #Ceva (6.6±2.3 [log2]) and #Zoe (6± 0.8 [log2]), followed by groups #Avi-1 and #BI with 4.1±1.2 [log2] and 3.8±0.9 [log2] (deviation to #Zoe: p=0.0002).
HI-and SNT assay detect antibodies that interact with native viral components, therefore they have biologically relevant read out. Based on these results, vaccine candidates #Ceva and #Zoe were selected for a subsequent challenge experiment.
3.2. Protective Efficacy Against Homologous Challenge
3.2.1. Full Clinical Protection in Seropositive Vaccinees
Geese in the mock-vaccinated control group started to show clinical signs 3 days post infection (dpi) onwards. Initially, one animal exhibited a reduced general condition, and a few others had light green, watery feces. By 4 dpi, the health of all animals had deteriorated, with four geese in very poor condition. Additional further 5 geese presented severe central nervous clinical signs, including torticollis and loss of flight reactions. These nine birds were euthanized on 4 dpi, including the last goose, which although mildly affected at that timepoint, was expected to die during the night and was euthanized to prevent suffering from isolation. For clinical score (CS) estimation, this last animal was considered dead the following day, resulting in a CS of 2.13 for the control group. In vaccine group #Ceva, 9 out of 10 challenged birds showed no signs of disease during the 14-day observation period (CS = 0.24). However, the one goose that remained seronegative in all tests including the SNT, died peracutely on 4 dpi. In the challenged group #Zoe, none of the 10 test animals showed any clinical reactions (CS = 0).
3.2.2. Vaccination Reduced Virus Shedding
Despite the initial absence of clinical signs, viral shedding was detected by RT-qPCR in swabs of all non-vaccinated geese at 2 dpi. The virus load in the oropharyngeal swabs ranged from 3.75 x 105 to 6.23 x 107(VE/ml). The virus load increased to 3.61 to 5 x 108 (VE/ml) in individual animals until 3 dpi and remained high on 4 dpi, when animals were euthanized (Figure 4, Supplementary Table S5). A similar excretion pattern was observed in cloacal swabs but with generally lower viral loads, averaging between 4.31 x 103 to 4.11 x 106(VE/ml) (Figure 4; Supplementary Table S5). In vaccine group #Ceva, oropharyngeal swab samples of six individual animals were positive on 2 dpi with virus loads between 4.93 x 103 and 1.07 x 104 (VE/ml). By 3 and 4 dpi, all nine clinical healthy geese shed virus, with one bird reaching a viral load as high as 1.94 x 107 (VE/ml) on 3 dpi. The single seronegative goose in this group, which succumbed to infection on 4 dpi, shed virus to levels comparable to non-vaccinated control group with 2.39 x 108 VE/ml detected in oropharyngeal swab samples early as 2 dpi. In the remaining clinically healthy geese, virus excretion gradually subsided until 10 dpi, when seven out of nine surviving birds tested positive for the last time with very low viral loads (Figure 4; Supplementary Table S5). Cloacal swabs of this group showed excretion kinetics similar to the oropharyngeal results, with a gradual increase in number of positive birds and viral load until 4 dpi, returning to negative PCR results by 14 dpi. Shedding patterns in the #Zoe vaccine group were similar to those in the #Ceva vaccine group, with virus shedding observed between 2 to 10 dpi. Statistical comparison of excretion patterns, analyzed by estimating the area under the curve (AUC), revealed that both vaccines significantly reduced excretion (#Ceva p=0.0003; #Zoe p<0.0001) compared to unvaccinated controls, excluding the serological non-responder in group #Ceva (see Supplementary Table S8, time period 1-4 dpi). No significant differences were found between the two vaccine groups when comparing the time period until 4 dpi or until 14 dpi (Figure 4 A; Supplementary Table S8).
3.2.3. Failure to Isolate Virus in Cell Culture from Swabs of Vaccinated Geese
In addition to RT-qPCR, positive oropharyngeal swab samples were analyzed for infectivity by cultivation on LMH cells. Infectious virus was recovered from numerous swab samples from non-vaccinated control geese between 2 and 4 dpi (8/10, 5/10 and 3/10, respectively) (Figure 4B and Supplementary Table S7). In contrast, no infectious virus was recovered from any swabs obtained from clinically healthy vaccinated geese, except the seronegative goose in the #Ceva group that died on 4 dpi, where infectious virus was recovered from swab samples from 2 and 3 dpi (Figure 4B; Supplementary Table S7).
3.2.4. AIV Infection Was Cleared in Vaccinated, Seropositive Geese
At the end of the observation period, organ samples of euthanized geese were analyzed. This included geese that succumbed to infection by 4 dpi and vaccinated geese that survived challenge infection without clinical signs until 14 dpi. The results confirmed systemic infection, with the highest viral load detected in the brain tissues of clinically diseased geese (3.26 x 108 to 2.79 x 1010VE/ml), regardless of whether they showed clinical CNS disorders (see Supplementary Table S6). Interestingly, fat tissues from individual animals also yielded high viral RNA loads (1.12 x 106 to 7.02 x 107VE/ml). In contrast, none of the organ samples from the vaccinated animals sacrificed on 14 dpi yielded any PCR-positive results in the organs, indicating full clearance of the infection.
3.2.5. NP-Specific Seroconversion as a Serological DIVA Surveillance Tool
Animals receiving marker vaccines that lacked viral NP (groups #Ceva, #Avi-2, #BI) did not seroconvert following primary and booster immunizations in an NP-specific ELISA, while all geese receiving whole virus vaccines seroconverted after primary vaccination (see Figure 2B, Supplementary Table S2). Post challenge plasma samples taken on 14 dpi revealed that all nine geese in the #Ceva vaccine group seroconverted in the NP-ELISA, whereas reactivity in the #Zoe vaccine group, which already tested positive before challenge, remained stable. Antibody reactivity increased in both vaccinated groups as measured by H5 ELISA and by HI with the antigen homologous to the challenge virus (Figure 5).
4. Discussion
The study revealed that primary antibody response to five commercial AI virus H5 vaccines in subadult fattening geese was weak after one vaccine dose administration. However, this response was significantly boosted by a second application of the same vaccine.
To assess vaccine immunogenicity, comparison of four serological assays with different test principles proved valuable. The competitive subtype-specific H5 ELISA was an effective, non-species-specific tool for detecting vaccine-induced antibodies. Its rapid test format made it suitable for tracking antibody formation at various stages of the vaccination process. However, biological relevance of the target epitope in the competitive H5 ELISA is not defined. Therefore, complementary diagnostic methods were used: the HI assay, which indicates antibody-mediated interference of virus-receptor binding, is considered a better surrogate marker for protection, particularly when the homologous antigen is used [12]. A possible discrepancy between H5-ELISA reactivity and HI becomes evident when comparing the data in the #BI group. By H5-ELISA, the #BI-vaccine was considered the most immunogenetic (Figure 2A; Supplementary Table S3) (deviation to all other groups: p<0.01-0.0001). However, using the HI assay with an antigen homologous to the challenge virus, reactivity was considerably lower than in group #Ceva and significantly lower than in group #Zoe (p=0.002). Similarly, #Avi-2 group showed the second best reactivity by H5-ELISA, but plasma from this group failed to show any HI-reactivity with AI virus H5 clade 2.3.4.4b antigen. This might in part to an antigenic mismatch of the HA-antigen in the vaccine and in the test procedure. Using clade 1.0 antigen, revealed that the #BI vaccine induced a stronger reactivity toward this historical antigen. This broad cross-reactivity induced by the #BI vaccine is remarkable and might be beneficial for broad application against H5 clades of the gs/GD lineage. For intervention to the currently circulating HPAIV H5 clade 2.3.4.4b, a clade-specific, tailored antigen seems more efficient. This conclusion is further supported by the SNT, that corroborated the HI-results with the clade 2.3.4.4b antigen.
Our results also highlight the limitations to using the HI as a prognostic marker for the protection status of a bird, as previously attempted [23]. Although the HI assay provided valuable insights into strain/clade specific immune responses when different antigens were used (Table 2), it is important to recognize that antibody levels alone do not necessarily correlate with clinical protective efficacy. In our study, at least 3 out of 10 animals of group #Ceva showed homologous HI titers of log2 <4, which are often interpreted as insufficient protection. However, only one of these geese ultimately proved not to be protected after challenge infection. This is similar to what was observed with the #AVI-2 vaccine, which, despite the absence of HI titers, protected chickens against challenge with the HPAI virus A/turkey/Italy/21VIR 9520-3/2021 strain, belonging to clade 2.3.4.4b (Ramirez-Martínez, unpublished 2024). Therefore, HI results can be used to estimate the overall protective efficacy of vaccines in a flock and to facilitate comparisons between them. At the individual level, however, the correlation of HI in the lower titer ranges with clinical protection remains weak. In particular, this might be true for the #Ceva vaccine, as this vaccine is based on a new RNA technology, expressing the antigen intracellularly whereby enabling MHC-1 presentation and induction of cellular, i.e. CD8+, immune response [24,25,26,27].
The lack of sufficient quantification methods for avian cellular immune responses applicable to routine diagnostics, apart from IFN-α staining [28,29], impedes developing alternative immunological marker systems as correlates of protection [12]. Meanwhile, even though technically intricate and labor-intensive, the HI assay can be considered the method of choice to test antigenic match of a vaccine. Plasma sample processing with pre-incubation of plasma with erythrocytes proved to reduce non-specific reaction often observed with serum from waterfowl species. In this context, testing sera in parallel for NDV antibodies was a good indication of specificity of the processed plasma samples, as three vaccines did not contain NDV components and remained seronegative. For vaccines #Avi-2 and #BI on the other hand, it was another indicator of proper vaccine administration. Altogether, considering the practical limitations of the HI assays, the H5-ELISA appears more appropriate for large-scale surveillance, once correlation of immunogenicity of certain vaccines have been established.
The NP-ELISA as fourth serological assay, was applied to verify the concept of Differentiating Infected from Vaccinated Animals (DIVA). None of the geese immunized with any of the three vaccines that lacked the AIV-NP protein (#Ceva, # AVI2, #BI) were reactive within the NP-ELISA. Furthermore, following challenge infection, all geese immunized with the #Ceva vaccine seroconverted by the NP-ELISA. This finding aligns with shedding data, demonstrating productive infection in vaccinated geese after challenge, and corroborates the concept of the NP-ELISA as a DIVA tool for retrospective surveillance.
For risk-based surveillance, however, direct detection of the virus is mandatory to differentiate notifiable AI virus H5/H7-infections from infection with other subtypes like AI virus H6 or H9, that regularly circulate in poultry. Using the generic RT-qPCR, targeting the AI virus matrix (M)-gene, viral RNA was detected in swab samples collected from the oropharynx and to a lesser extent from cloacal swabs (Supplementary Table S5), identifying infected geese as early as 2 dpi, i.e. 2 days before onset of the disease in unvaccinated control geese. The test provides evidence that vaccinated geese of both groups shed virus between 2 and 10 dpi. Yet, virus shedding was significantly reduced compared to naïve geese (Figure 4). The detected residual virus shedding in clinically healthy birds confirms previous results in other poultry species and highlights a key challenge of AI virus vaccination regimes, which might facilitate silent virus spread within and between flocks [31,32]. Identifying field virus-infected individual birds that might act as effective transmitters and spreaders of the virus within a flock is considered of utmost importance for any surveillance strategy of vaccinated flocks [33]. Concerning the efficacy of the vaccine, a key factor is the decrease in susceptibility and transmissibility affecting virus spread within and between farms. In this respect, our results might hold some promising perspectives: Even though RT-qPCR results point to viral RNA shedding in vaccinated geese, no infectious virus was recovered in oropharyngeal swabs by LMH cell culture from infected vaccinated geese, while virus isolation was successful from swabs of non-vaccinated control animals (Figure 4B, Supplementary Table S7). This might be in part due to virus neutralization by mucosal antibodies [34]. Else, this might reflect an insufficient detection limit of the LMH cell culture system used here. Ultimately, further studies focusing on the basic reproduction number, R0 are needed to investigate whether this reduced virus shedding has an impact on virus transmission between geese. This index indicates the average number of secondary cases caused by one individual throughout its infectious phase [35]. An effective vaccine should aim at reducing transmission to a level of R0 < 1. Based on estimates of R0, the anticipated protection efficacy of candidate vaccines can be compared to develop more effective disease control strategies. The data obtained here will be used to select candidates for further transmission experiments. Moreover, the protective efficacy of the vaccines that were not challenged remains to be determined. Future studies should include challenge trials for these vaccines that demonstrated immunogenicity to achieve a more complete evaluation of their potential role in disease control.
In conclusion, four out of five tested commercial AIV H5 vaccines were immunogenic in geese after booster immunization, and two candidate vaccines inducing highest antibody titers by HI and SNT protected animals against clinical disease. Although vaccinated geese remained susceptible to HPAIV H5 challenge infection, virus excretion was significantly reduced and no infectivity was recoverable in a cell culture-based system from oropharyngeal swab samples of these birds. Studies on the reduction of virus transmissibility to and between vaccinated geese by estimating R0 are recommended for a full characterization of the epidemiological efficacy of such vaccines.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Plasma samples from 4 vaccine groups after prime immunization with positive HI results; Table S1: H5 ELISA; Table S2 NP ELISA; Table S3 HI assay; Table S4 SNT; Table S5 Virus excretion after challenge infection; Table S6 Virla organ loads of all challenged groups; Table S7 Infectivity of swab samples on LMH cells; Table S8 AUC excretion data.
Author Contributions
Conceptualization and methodology by C.G. and T.C.H.; investigation and data curation by R.P., C.G., and T.C.H.; formal analysis by R.P. and C.G.; writing—original draft preparation by R.P., T.C.H., and C.G.; writing—review and editing by all co-authors; visualization by R.P.; allocation of vaccines: C.C., L.FdS.; L.R-M.; H.H.; project administration by C.G.; funding acquisition by T.C.H. and M.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research was financed by a grant issued by a consortium headed by the Tierseuchenkasse Niedersachsen, Hannover, Germany.
Institutional Review Board Statement
The animal study protocol has been granted permission by an independent ethics commission of the German Federal State of Mecklenburg-Vorpommerania (LALLF 7221.3-1-037/23-1).
Data Availability Statement
All curated data are included in the supplementary materials accompanying this manuscript. Additional data are available upon request from the corresponding authors.
Acknowledgments
Thanks to all co-authors, including Stephane Lemiere (Boehringer-Ingelheim) for evaluating the concept of the study and editing the manuscript. A special thanks to the technical staff, Cornelia Illing, Kristin Trippler, and Diana Parlow, for their invaluable assistance in the laboratory. We also extend our heartfelt gratitude to the animal caretakers for their dedicated and meticulous support during the animal experiments.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Abbreviations
The following abbreviations are used in this manuscript:
| BSL 3 | Biosafety Level 3 |
| CPE | cytopathic effect |
| DIVA | Differentiation of infected from vaccinated animals |
| FCS | Fetal calf serum |
| FLI | Friedrich-Loeffler-Institute |
| gs/GD | Goose/Guandong |
| HA | Haemagglutinin |
| HA | Hemagglutination assays |
| HI | Hemagglutination inhibition assay |
| HPAI | Highly patogenic avian influenza |
| HPAIV | Highly patogenic avian influenza virus |
| IFN | Interferon |
| LMH | Leghorn male hepatoma |
| MTA/NDA | Material Transfer Agreement/ Non-Disclosure Agreement |
| NDV | Newcastle Disease Virus |
| SNT | Serum Neutralization test |
| SPF | Specific-pathogen-free |
| TCID | Tissue-culture-infection dose |
| VE | Viral equivalents |
References
- Alders, R., Awuni, J. A., Bagnol, B., Farrell, P. & Haan, N. de. Impact of avian influenza on village poultry production globally. EcoHealth 2014, 11, 63–72. [CrossRef]
- Hautefeuille, C. Hautefeuille, C., Dauphin, G. & Peyre, M. Knowledge and remaining gaps on the role of animal and human movements in the poultry production and trade networks in the global spread of avian influenza viruses - A scoping review. PloS one 2020, 15, e0230567. [CrossRef]
- Verhagen, J. H., Fouchier, R. A. M. & Lewis, N. Highly Pathogenic Avian Influenza Viruses at the Wild-Domestic Bird Interface in Europe: Future Directions for Research and Surveillance. Viruses 2021, 13. [CrossRef]
- Klaassen, M. & Wille, M. The plight and role of wild birds in the current bird flu panzootic. Nature ecology & evolution 2023, 7, 1541–1542. [CrossRef]
- Koopmans, M. P. G. et al. The panzootic spread of highly pathogenic avian influenza H5N1 sublineage 2.3.4.4b: a critical appraisal of One Health preparedness and prevention. The Lancet. Infectious diseases 2024. [CrossRef]
- Delpont, M. et al. Monitoring biosecurity in poultry production: an overview of databases reporting biosecurity compliance from seven European countries. Frontiers in veterinary science 2023, 10, 1231377. [CrossRef]
- Hassan, K. E. et al. Improved Subtyping of Avian Influenza Viruses Using an RT-qPCR-Based Low Density Array: 'Riems Influenza a Typing Array', Version 2 (RITA-2). Viruses 2022, 14. [CrossRef]
- EFSA Scientific Report. Avian influenza overview December 2022 - March 2023. EFSA Journal 2023, 21, e07917. [CrossRef]
- EFSA Scientific Report. Drivers for a pandemic due to avian influenza and options for One Health mitigation measures. EFSA Journal 2024, 22, e8735. [CrossRef]
- Lewis, N. S. et al. Antigenic evolution of contemporary clade 2.3.4.4 HPAI H5 influenza A viruses and impact on vaccine use for mitigation and control. Vaccine 2021, 39, 3794–3798. [CrossRef]
- EFSA Panel on Animal Health and Animal Welfare (AHAW), European Union Reference Laboratory for Avian Influenza. Vaccination of poultry against highly pathogenic avian influenza - part 1. Available vaccines and vaccination strategies. EFSA journal. European Food Safety Authority 2023, 21, e08271. [CrossRef]
- EFSA Panel on Animal Health and Animal Welfare (AHAW), European Union Reference Laboratory for Avian Influenza. Vaccination of poultry against highly pathogenic avian influenza - Part 2. Surveillance and mitigation measures (2024).
- Harder, T. et al. Epidemiology-driven approaches to surveillance in HPAI-vaccinated poultry flocks aiming to demonstrate freedom from circulating HPAIV. Biologicals: journal of the International Association of Biological Standardization 2023, 83, 101694. [CrossRef]
- French Ministry of Agriculture and Food Sovereignty. Arrêté Ministériel du 25 septembre 2023 relatif aux mesures de surveillance, de prévention, de lutte et de vaccination contre l’influenza aviaire hautement pathogène (IHAP). [Ministerial decree of 25 September 2023 as regards surveillance, prevention, intervention and vaccination measures against highly pathogenic avian influenza (HPAI)] (2023).
- French Ministry of Agriculture and Food Sovereignty. Influenza aviaire: le plan de vaccination de la France. [Avian influenza: France’s vaccination plan. 2024. Available online: https://agriculture.gouv.fr/tout-ce-quil-faut-savoir-sur-le-plan-daction-vaccination-iahp-en-france.
- Guinat, C. et al. Promising effects of duck vaccination against highly pathogenic avian influenza, France 2023-24 (2024).
- Oliveira Cavalcanti, M. et al. A genetically engineered H5 protein expressed in insect cells confers protection against different clades of H5N1 highly pathogenic avian influenza viruses in chickens. Avian pathology: journal of the W.V.P.A 2017, 46, 224–233. [CrossRef]
- Piesche, R. et al. Dominant HPAIV H5N1 genotypes of Germany 2021/2022 are linked to high virulence in Pekin ducklings. npj Viruses 2024, 2. [CrossRef]
- Reed, L.J.; Muench, H. A simple method of estimating fifty percent endpoints. Am. J. Hyg. 1938, 27, 493–497. [Google Scholar]
- Alexander, D.J. Highly pathogenic avian influenza. In Manual of Standards for Diagnostic Tests and Vaccines, 4th ed.; Office International des Epizooties: Paris, France, 2000. [Google Scholar]
- WOAH. Terrestrial Manual on Avian Influenza (Chapter 3.3.4) (2023).
- Ahrens, A. K., Selinka, H.-C., Mettenleiter, T. C., Beer, M. & Harder, T. C. Exploring surface water as a transmission medium of avian influenza viruses - systematic infection studies in mallards. Emerging microbes & infections 2022, 11, 1250–1261. [CrossRef]
- Spackman, E. & Swayne, D. E. Vaccination of gallinaceous poultry for H5N1 highly pathogenic avian influenza: current questions and new technology. Virus research 2013, 178, 121–132. [CrossRef]
- Hirai, T. & Yoshioka, Y. Considerations of CD8+ T Cells for Optimized Vaccine Strategies Against Respiratory Viruses. Frontiers in immunology 2022, 13, 918611. [CrossRef]
- Roth, G. A. et al. Designing spatial and temporal control of vaccine responses. Nature reviews. Materials 2022, 7, 174–195. [CrossRef]
- Borriello, F. et al. An adjuvant strategy enabled by modulation of the physical properties of microbial ligands expands antigen immunogenicity. Cell 2022, 185, 614-629.e21. [CrossRef]
- Li, C. et al. Mechanisms of innate and adaptive immunity to the Pfizer-BioNTech BNT162b2 vaccine. Nature immunology 2022, 23, 543–555. [CrossRef]
- Reemers, S. S. N., van Haarlem, D. A., Sijts, A. J. A. M., Vervelde, L. & Jansen, C. A. Identification of novel avian influenza virus derived CD8+ T-cell epitopes. PloS one 2012, 7, e31953. [CrossRef]
- Reemers, S., Verstegen, I., Basten, S., Hubers, W. & van de Zande, S. A broad spectrum HVT-H5 avian influenza vector vaccine which induces a rapid onset of immunity. Vaccine 2021, 39, 1072–1079. [CrossRef]
- Germeraad, E. A. et al. Transmissiestudie met vier vaccins tegen H5N1 hoogpathogeen vogelgriepvirus (clade 2.3.4.4b). CVI Virology, 2023. [CrossRef]
- Cha, R. M. et al. Suboptimal protection against H5N1 highly pathogenic avian influenza viruses from Vietnam in ducks vaccinated with commercial poultry vaccines. Vaccine 2013, 31, 4953–4960. [CrossRef]
- Poetri, O. N. et al. Silent spread of highly pathogenic Avian Influenza H5N1 virus amongst vaccinated commercial layers. Research in veterinary science 2014, 97, 637–641. [CrossRef]
- Elbers, A. R. W. & Gonzales, J. L. Mortality Levels and Production Indicators for Suspicion of Highly Pathogenic Avian Influenza Virus Infection in Commercially Farmed Ducks. Pathogens (Basel, Switzerland) 2021, 10. [CrossRef]
- Suarez, D.L.; Schultz-Cherry, S. Immunology of avian inuenza virus: a review. Dev. Comp. Immunol. 1999, 269–283. [Google Scholar] [CrossRef]
- Halloran, M.E. Concepts of transmission and dynamics. In Epidemiologic Methods for the Study of Infectious Diseases.; 2001; pp. 63–64. [Google Scholar]
Figure 1.
Vaccination scheme. After an acclimatization phase of three weeks, six groups of ten goslings each at six weeks of age received primary vaccination followed by a booster vaccination in week 10. One group received a mock vaccine preparation without H5 antigen. Plasma samples taken at each vaccination and three weeks after booster immunization (week 13) were used for determining antibody response. Two vaccine groups showing highest antibody titers by HI were chosen for a challenge infection experiment at week 13, that included a group of non-vaccinated control geese. After an observation time of up to two weeks or when animals reached a human endpoint of disease, geese were euthanized and organ samples taken for virological investigation.
Figure 1.
Vaccination scheme. After an acclimatization phase of three weeks, six groups of ten goslings each at six weeks of age received primary vaccination followed by a booster vaccination in week 10. One group received a mock vaccine preparation without H5 antigen. Plasma samples taken at each vaccination and three weeks after booster immunization (week 13) were used for determining antibody response. Two vaccine groups showing highest antibody titers by HI were chosen for a challenge infection experiment at week 13, that included a group of non-vaccinated control geese. After an observation time of up to two weeks or when animals reached a human endpoint of disease, geese were euthanized and organ samples taken for virological investigation.
Figure 2.
Comparative analysis of the antibody response after prime and booster vaccination with one of five commercial vaccines in five different serological assays. Plasma samples were taken after prime vaccination at week (wk) 10* and three weeks after booster immunization (week 13)* and tested for the presence of AI virus antibodies through H5-ELISA (A), NP-ELISA (B), HI against H5 antigen (C), HI against Newcastle virus antigen (D) and SNT (E). Dotted lines on graphs A-D mark the respective cut-off, separating negative (below dotted line) from positive titers (above dotted line) (“indeterminate” area between dotted lines for ELISAs). Box-plots showing 25-75th percentiles and median titers. *Plasma samples of geese of the #BI group were drawn and tested at wk 14 and wk 17.
Figure 2.
Comparative analysis of the antibody response after prime and booster vaccination with one of five commercial vaccines in five different serological assays. Plasma samples were taken after prime vaccination at week (wk) 10* and three weeks after booster immunization (week 13)* and tested for the presence of AI virus antibodies through H5-ELISA (A), NP-ELISA (B), HI against H5 antigen (C), HI against Newcastle virus antigen (D) and SNT (E). Dotted lines on graphs A-D mark the respective cut-off, separating negative (below dotted line) from positive titers (above dotted line) (“indeterminate” area between dotted lines for ELISAs). Box-plots showing 25-75th percentiles and median titers. *Plasma samples of geese of the #BI group were drawn and tested at wk 14 and wk 17.
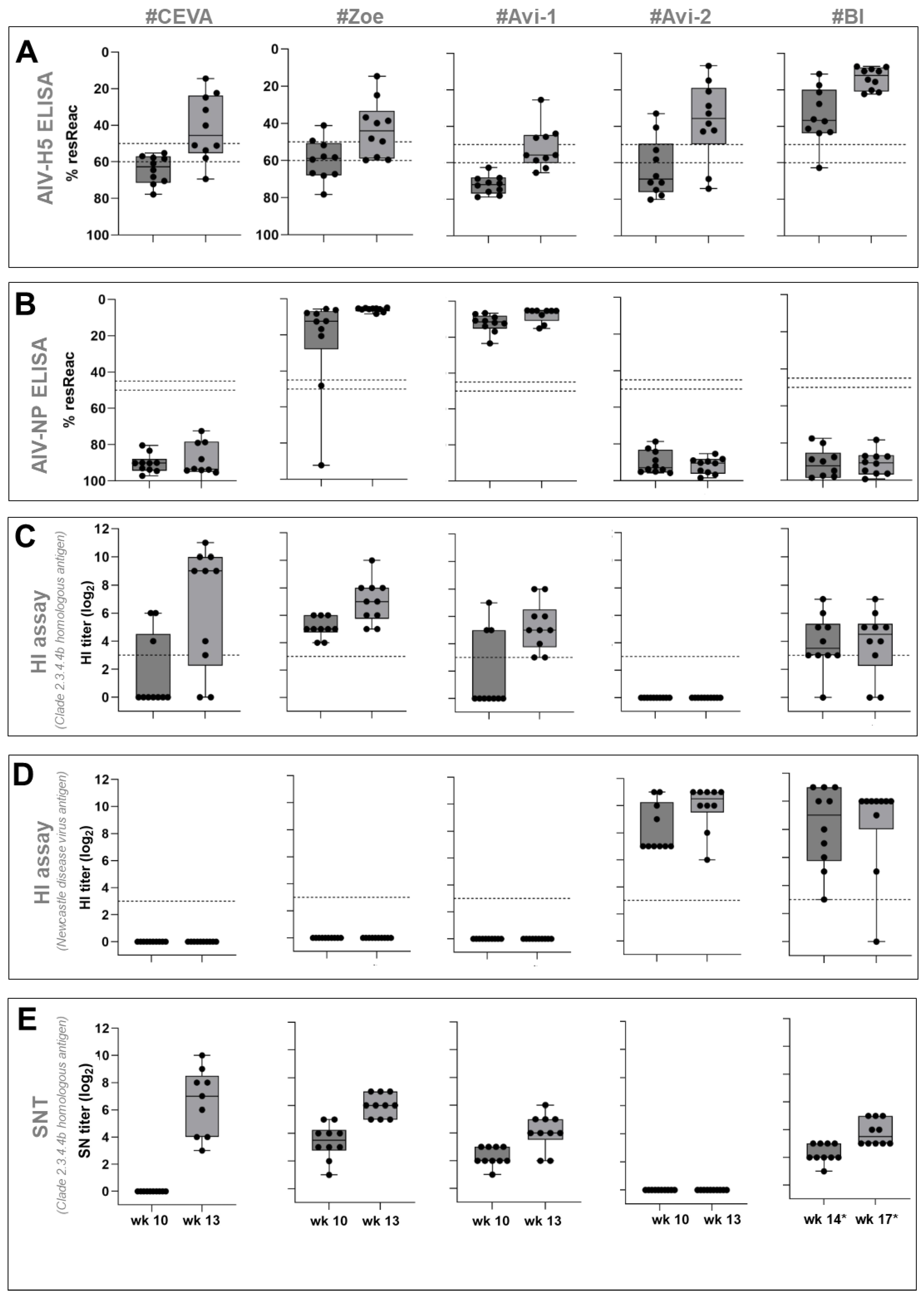
Figure 3.
Overview of plasma reactivity in HI assays against different antigens. Plasma samples from 4 vaccine groups were tested after booster immunization by HI assay against antigens of clade 2.3.4.4b: (A) A/chicken/Germany-NI/AI 4286/2022 and (B) A/chicken/Germany-SH/AI08298/2021. Heterologous antigen originated from clade 1.0 (C) A/chicken/Vietnam/P41-05 (R75/05). In addition, an NDV antigen was used: (D) Genotype 2.II (NDV Clone 30).
Figure 3.
Overview of plasma reactivity in HI assays against different antigens. Plasma samples from 4 vaccine groups were tested after booster immunization by HI assay against antigens of clade 2.3.4.4b: (A) A/chicken/Germany-NI/AI 4286/2022 and (B) A/chicken/Germany-SH/AI08298/2021. Heterologous antigen originated from clade 1.0 (C) A/chicken/Vietnam/P41-05 (R75/05). In addition, an NDV antigen was used: (D) Genotype 2.II (NDV Clone 30).
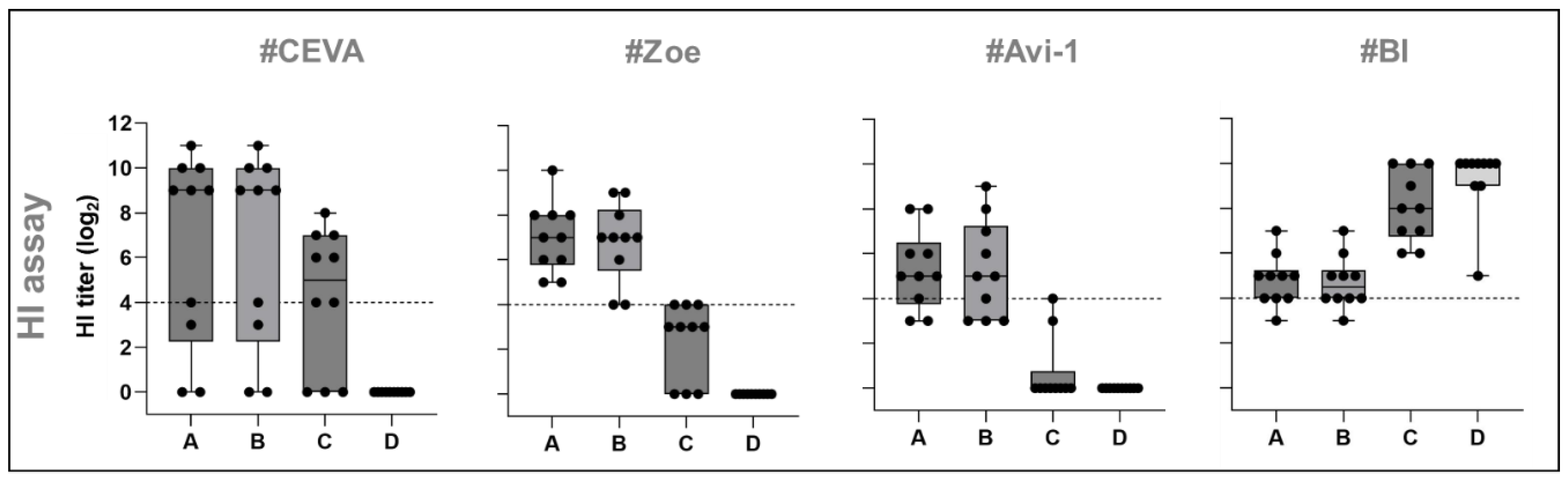
Figure 4.
Comparison of total (A) and individual daily (B) oropharyngeal viral RNA shedding of infected groups. Total oropharyngeal shedding per goose (VE/ml [log10]) was compared up to 4 days post infection, measured by calculating the area under the curve (AUC) values of all animals of the three challenged groups (A). The viral load in VE/ml [log2] per oropharyngeal swab of each individual animal per day was plotted in (B). Using LMH cell cultures, infectious virus (B, indicated by red symbols) could be recovered from swabs derived from the control group only as well as from one non-responder of the group. The levels of significance are indicated by *** (p<0.001), **** (p<0.0001), and ns (not significant).
Figure 4.
Comparison of total (A) and individual daily (B) oropharyngeal viral RNA shedding of infected groups. Total oropharyngeal shedding per goose (VE/ml [log10]) was compared up to 4 days post infection, measured by calculating the area under the curve (AUC) values of all animals of the three challenged groups (A). The viral load in VE/ml [log2] per oropharyngeal swab of each individual animal per day was plotted in (B). Using LMH cell cultures, infectious virus (B, indicated by red symbols) could be recovered from swabs derived from the control group only as well as from one non-responder of the group. The levels of significance are indicated by *** (p<0.001), **** (p<0.0001), and ns (not significant).
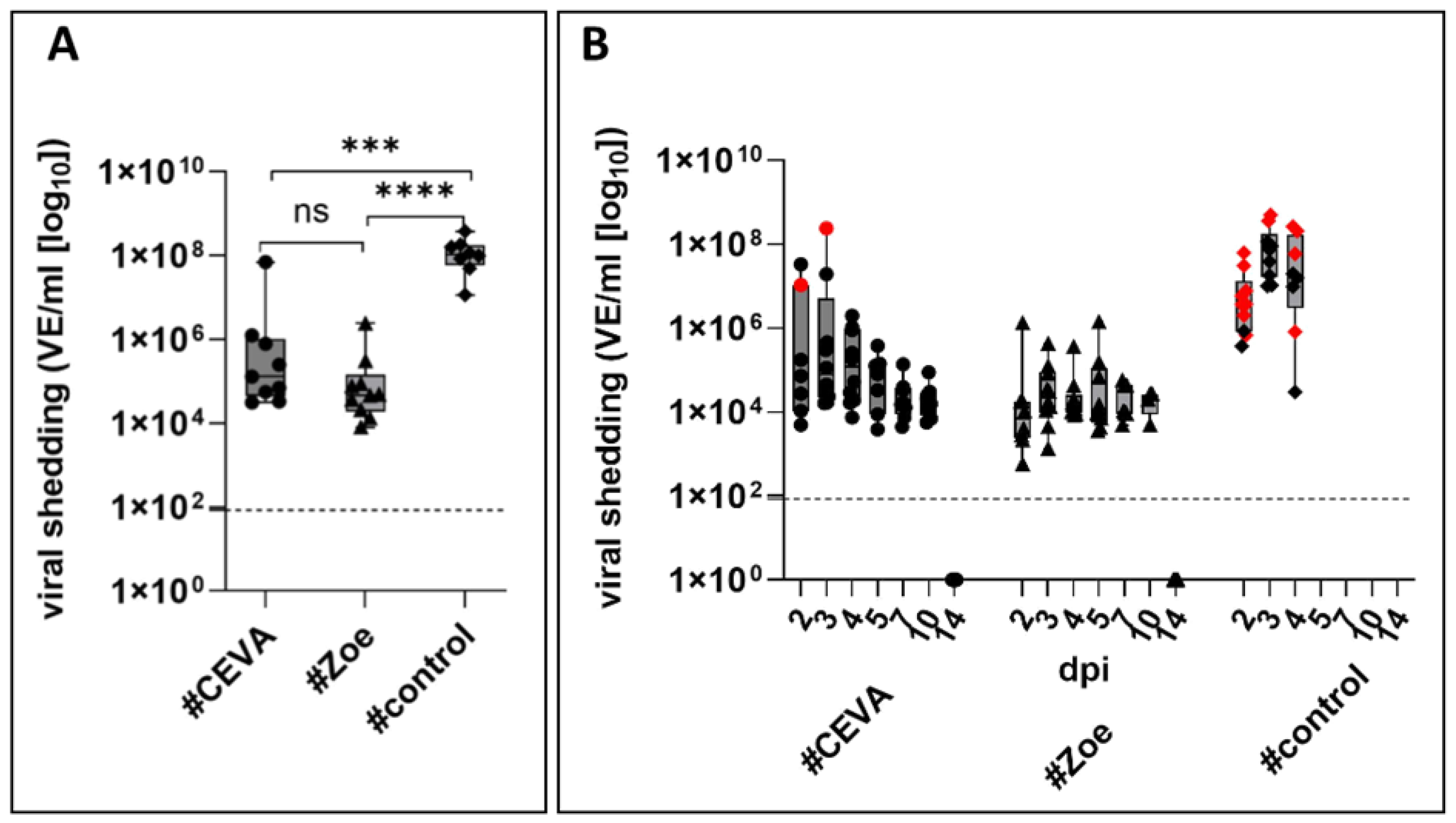
Figure 5.
Antibody responses in vaccinated geese following HPAIV H5N1 challenge infection. Sero-reactivity was tested by NP-(A) and H5-specific ELISA (B) as well as by HI (C). Significance levels are indicated by ** (p<0.01) and **** (p<0.0001).
Figure 5.
Antibody responses in vaccinated geese following HPAIV H5N1 challenge infection. Sero-reactivity was tested by NP-(A) and H5-specific ELISA (B) as well as by HI (C). Significance levels are indicated by ** (p<0.01) and **** (p<0.0001).
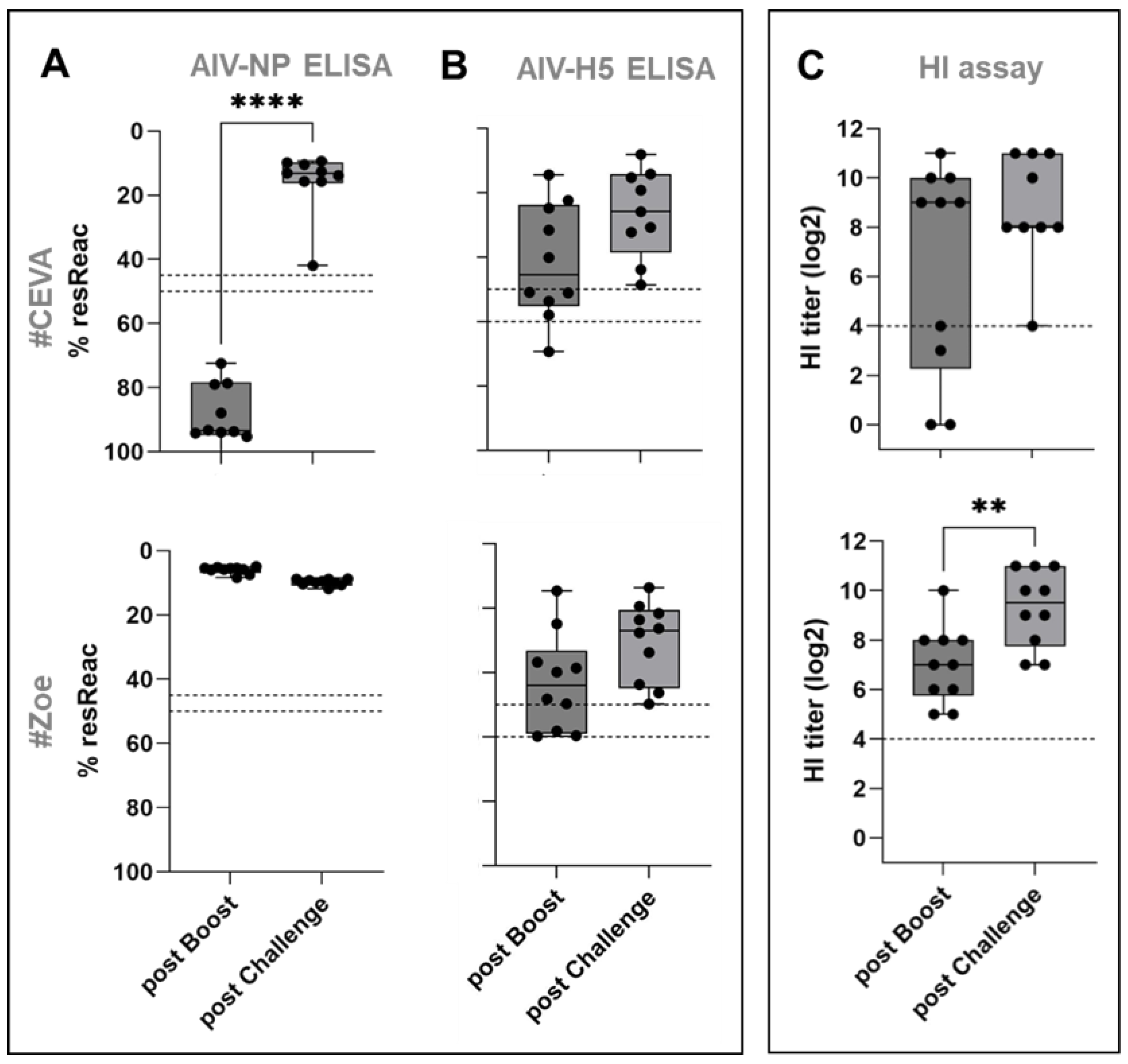

Table 1.
Characteristics of vaccines used.
| Group | Vaccine producer | Vaccine name | Batch number | Type [gs/GD clade] | Dose and application | |
|---|---|---|---|---|---|---|
| #Ceva | CEVA | RESPONS AI H5 | Lot 0409LF | Amplicon (H5) [2.3.4.4b] |
0.2 ml | i.m. caudal femoral muscle |
| #Zoe | Zoetis | NE 69521 LO523AS03 | WIV (H5N2) [2.3.4.4b] |
0.5 ml | s.c. neck fold | |
| #Avi-1 | Avimex | Vaxigen Flu H5N8 clado 2.3.4.4 | Reg B-0258-131 | WIV (H5N8) [2.3.4.4b] |
0.5 ml | s.c. neck fold |
| #Avi-2 | Avimex | KNewH5 | Lote E.PM-2304 | recNDV (H5) [2.3.4.4b] |
0.5 ml | s.c. neck fold |
| #BI | Boehringer | Volvac B.E.S.T. AI+ND | 2307011A | Baculovirus-based expression system of an inserted optimized H5 sequence for antigen formulation | 0.5 ml | s.c. neck fold |
Table 2.
List of viruses used in challenge trials and in serology.
| Sub-and phenotype | Clade | Isolate | Sequence accession | |
|---|---|---|---|---|
| A | HP H5N1 | 2.3.4.4b | A/chicken/Germany-NI/AI 4286/2022 | Epi16096050 |
| B | HP H5N1 | 2.3.4.4b | A/chicken/Germany-SH/AI08298/2021 | Epi18006820 |
| C | HP H5N1 | 1.0 | A/chicken/Vietnam/P41-05/2005 (R75/05) | Epi13970 |
| D | NDV | Genotype 2.II | NDV LaSota | ON713864 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



