
Submitted:
28 February 2025
Posted:
28 February 2025
You are already at the latest version
Abstract
Rhamnan sulfate (RS), extracted from the seaweed Monostroma nitidum, suppresses vascular endothelial inflammation and arteriosclerosis, decreases blood glucose levels, and improves blood lipid metabolism and the intestinal environment. We investigated whether RS improves diabetic brain dysfunction using a hyperglycemic mouse model induced by pretreatment with nicotinamide and streptozotocin followed by oral administration of a high-fat diet and maltodextrin (MD) for 4 months. RS was administered in a 1% MD solution at doses of 100, 300, and 1,000 mg/kg of mouse body weight. Administration of RS to hyperglycemic mice significantly reduced blood glucose levels and showed a tendency to improve memory function in behavioral pharmacological tests using spontaneous locomotor activity, the rotarod test, and eight-arm maze test. Brain histopathological analysis showed that RS significantly reduced the atrophy of the neuronal layers in each region of the hippocampus compared to the untreated hyperglycemic controls. RS also significantly suppressed TNF-α expression and microglial activation in the brain, and showed a tendency to reduce the accumulation of advanced glycation end products in the brain. These results suggest that RS intake suppresses inflammation in the brain and alleviates the cognitive impairment associated with hyperglycemic diabetes.
Keywords:
diabetic cognitive impairment
; hippocampal atrophy
; AGEs
; rhamnan sulfate
; Monostroma nitidum
1. Introduction
In recent years, the incidence of diabetes, which is closely related to lifestyle habits, has been increasing year by year. Diabetes is not limited to middle-aged and elderly people, but is also increasing in younger people. The ACCORD-MIND study has demonstrated that diabetes is an important risk factor for dementia, including Alzheimer’s disease [1]. Meta-analyses [2,3] and cohort studies [4,5], including the Hisayama-cho Study [6], have shown that patients with diabetes are at higher risk of developing Alzheimer’s disease than people without diabetes. Recent studies have revealed that some causes of dementia in hyperglycemia and diabetes are caused by advanced glycation end products (AGEs) and brain inflammation leading to cognitive impairment [7], increased oxidative stress, and insufficient insulin action in the brain due to insulin resistance [8,9].
Our previous studies have shown that long-term hyperglycemia causes cognitive impairment and promotes the formation of AGEs in the brain, leading to hippocampal atrophy [10,11,12]. This suggests that there may be a relationship between AGEs formed by hyperglycemia and cognitive impairment and hippocampal atrophy. However, studies conducted to date have not yet provided conclusive evidence on the causal relationship between elevated blood glucose levels and cognitive decline.
Therefore, in order to clarify the relationship between diabetes and dementia and its underlying causes, many studies have been conducted to develop preventive measures such as treatments for diabetic dementia using the PPAR-γ agonist pioglitazone [13], dipeptidyl peptidase-4 inhibitors [14], sulfated seaweed polysaccharides [15], and strict control of postprandial blood glucose levels [16]. However, most studies to date have not conclusively proven that these approaches are effective in preventing or treating cognitive decline.
Seaweeds, which are traditionally consumed in East Asia, including Japan, are rich in essential nutrients, including iodine, iron, vitamin C, vitamin K, and vitamin B12, and are a natural source of antioxidants, soluble and insoluble dietary fiber, and polysaccharides [17]. Previous research has revealed that seaweed is a functional food with health-promoting properties, and among the physiologically active components of seaweed, rhamnan sulfate (RS), a polysaccharide extracted from the green algae Monostroma nitidum, has been found to have various health-promoting properties. RS is a soluble fiber found in the intercellular substance of M. nitidum and is composed mainly of linear rhamnose chains with side chains and approximately 25% sulfate groups [18,19]. We have previously demonstrated that RS has numerous beneficial effects, including suppression of blood glucose levels [20,21,22], anti-obesity effects [20,21], improvement of intestinal environment [21], anti-inflammatory effects on vascular endothelial cells [23] and the whole body [24], and anti-arteriosclerosis effects through improvement of lipid metabolism and suppression of vascular inflammation [25], suggesting its potential to prevent various lifestyle-related diseases.
In this study, we investigated whether RS, which has been shown in previous studies to suppress blood glucose levels and have anti-inflammatory effects on vascular endothelial cells, is effective in improving cognitive impairment and brain tissue damage in a glucose-loaded mouse model.
2. Results
2.1. Body Weight and Blood Glucose Levels
The results of body weight and blood glucose level measurements for each group are shown in Figure 1A and 1 B, respectively. Body weight gradually increased during the study period (1.4 to 1.8 times higher than that of the control), and the MD group showed significant weight gain compared with the control group (p < 0.0001). These results suggest that RS administration partially suppresses weight gain associated with glucose loading.
After 4 months, blood glucose levels in the MD group were significantly higher than those in the control group (p < 0.00001). In contrast, all RS-treated groups (MD + RS100, p < 0.01; MD + RS300, p < 0.001; MD + RS1000, p < 0.001) showed a significant decrease in blood glucose levels compared with the MD group. These results suggested that RS effectively suppressed the increase in blood glucose levels in this model.
2.1.1. Effect of RS on Spontaneous Locomotor Activity
To evaluate the effect of RS on locomotor activity, we measured the number of spontaneous movements. The results are shown in Figure 2. There was no statistically significant difference in locomotive activity between the control group and MD-fed group, and all conditions showed similar activity levels (0.75 to 0.86 times higher than the control group).
2.1.2. Effect of RS on Rotarod Test
The fall latency was measured using the rotarod test to investigate the effects of RS administration on locomotor function. The fall latency values are shown in Figure 3. After 4 months of glucose loading, the MD group showed a significant decrease in fall latency compared to the control group (p < 0.001). The RS group tended to have a longer fall latency than the MD group, although this difference was not statistically significant.
2.1.3. Effect of RS on Eight-Way Maze Test
To investigate the effects of RS administration on motor function and memory in cognitive function, task completion time and number of errors were measured using the eight-way maze test. The time required to complete the task is shown in Figure 4A, and the number of errors is shown in Figure 4B. The MD group showed a significant increase in task completion time compared to the control group (p < 0.0001), suggesting that glucose loading impairs motor and cognitive functions.
The error rate was significantly higher in the MD group than in the control group (p < 0.0001). The MD + RS100 group showed a trend toward fewer errors than the MD group (p < 0.1), but this did not reach statistical significance. No significant difference in task completion time was observed between the RS and MD groups. Overall, these results suggest that RS had a limited effect on memory improvement in this model.
2.2. Analysis of Hippocampal Pathology Specimens
2.2.1. Effect of RS on Hippocampal Neuronal Layers (CA1, CA2, CA3, and Dentate Gyrus)
To investigate the effect of RS administration on hippocampal structure, the thicknesses of the CA1, CA2, CA3, and dentate gyrus regions of the hippocampal neuronal layer were compared. As shown in Figure 5, the thicknesses of the CA1, CA2, CA3, and dentate gyrus regions were significantly decreased in the MD group compared to those in the control group (CA1, p < 0.0001; CA2, p < 0.0001; CA3, p <0.0001; dentate gyrus, p < 0.0001). In contrast, the RS treatment significantly improved the thickness of these regions. The MD + RS100 and MD + RS300 groups showed statistically significant improvements compared to the control group for all thicknesses of the CA1, CA2, CA3, and dentate gyrus regions (MD + RS100/ CA1: p < 0.0001; CA2, p < 0.0001; CA3, p < 0.01; dentate gyrus, p < 0.0001) (MD + RS300/ CA1: p < 0.0001, CA2: p < 0.0001, CA3: p < 0.05, dentate gyrus: p < 0.0001). In the MD + RS1000 group, the thickness of the CA2 region improved significantly (CA2: p < 0.05), and the thicknesses of the CA1, CA3, and dentate gyrus regions showed a trend toward improvement; however, the difference did not reach statistical significance (p < 0.1 each). These results indicate that RS treatment suppresses hippocampal atrophy induced by glucose loading and partially restores the thickness of the hippocampal neuronal layer.
2.2.2. Effect of RS on TNF-α Expression in the Brain
To investigate the effect of RS on brain inflammation, the expression of the inflammatory cytokine TNF-α was assessed by immunostaining with anti-TNF-α antibody, and the expression levels were compared between groups, and the results are shown in Figure 6. The expression levels of TNF-α were scored based on staining intensity, and the results were shown in Table 1: 0 (no staining), +1 (weakly positive), +2 (moderately positive), and +3 (strongly positive). Although no TNF-α expression was detected in the control group, the expression of TNF-α was markedly significantly increased in the MD group compared to the control group (p < 0.01). RS treatment effectively suppressed the increase in TNF-α expression observed in the MD group. Statistical analysis showed that TNF-α expression was significantly suppressed in the MD + RS100, MD + RS300, and MD + RS1000 groups compared to the MD group (all p < 0.01). These results indicate that RS suppresses TNF-α expression and reduces inflammation-associated brain injury.
2.2.3. Effect of RS on Iba1 Expression in the Brain
To examine the effect of RS on brain inflammation, microglia cells were stained with an antibody against ionized calcium binding adapter molecule 1 (Iba1), a marker of microglial activation [26], and the expression levels were compared between groups. The results are shown in Figure 7. The expression level of Iba1 was scored based on staining intensity, and the results are shown in Table 2: 0 (no staining), +1 (weak staining), +2 (moderate staining), and +3 (strong staining). In the control group, all the samples showed weak Iba1 expression. In the MD group, Iba1 expression was significantly higher than that in the control group (p < 0.01). RS effectively suppressed the increase in Iba1 expression observed in the MD group. In the MD + RS100 group, all samples decreased to the same level as that in the control group. The MD + RS300 and MD + RS1000 groups also showed a decrease in Iba to levels similar to the control group, as did the MD + RS100. Statistical analysis showed that Iba1 expression was significantly suppressed in both MD + RS groups compared to the MD group (p < 0.01). These results indicated that RS suppressed microglial cells in the brain by inhibiting glucose loading-associated inflammation.
2.2.4. Effect of RS on AGEs Accumulation in the Brain
To evaluate the effect of RS administration on hippocampal glycation, brain tissues were stained with an anti-AGE antibody and AGE accumulation levels were compared between the groups, and the results are shown in Figure 8. The expression levels of AGEs were scored based on staining intensity, and the results are shown in Table 3: 0 (no staining), +1 (weakly positive), and +2 (strongly positive). In the control group, the AGE accumulation scores were mainly negative. In contrast, AGE accumulation was significantly elevated in the MD group (p < 0.05 compared to the control group). There was no statistically significant difference between the RS-administered and MD groups (p < 0.1); however, AGE accumulation in the brain tended to decrease.
3. Discussion
In this study, we investigated the effects of 4 months of RS administration on cognitive function and brain tissue in a 1% MD-induced hyperglycemic diabetic mouse model.
In the behavioral pharmacology test, no significant differences were observed between the MD and control groups regardless of whether RS was administered, indicating that neither MD nor RS had a direct effect on motor function. MD administration does not impair motor function [10,12]; therefore, the results of this study are considered valid. In the rotarod test, the MD group showed a significant decrease in fall latency compared with the control group. Although not statistically significant, the RS group tended to have a longer fall latency than the MD group. Although no significant differences were observed in the 8-way maze test, the RS group tended to have a lower number of errors compared to the MD group, suggesting that RS administration may improve memory and cognitive function in tasks.
Histopathological studies of the brain suggested that significant atrophy was observed in all regions of the hippocampus (CA1, CA2, CA3, and dentate gyrus) in the MD group, whereas the RS group (100 mg/kg or 300 mg/kg) showed significantly inhibited glucose loading-induced hippocampal atrophy and partially restored the thickness of the hippocampal neuronal layer.
In the hyperglycemic diabetic model mice used in this study, it is presumed that hyperglycemia induces pancreatic β cell death, and the inflammatory cytokines TNF-α and IL-1β secreted from pancreatic β cells migrate to the brain [27]. In fact, hyperglycemia and significant TNF-α production were observed in the brain of the MD group, whereas the TNF-α levels in the brain were significantly reduced in the RS-administered group, likely cause by the suppression of pancreatic inflammation. We previously reported that RS directly suppresses TNF-α-induced inflammation in cultured vascular endothelial cells [21,23]. Therefore, our findings suggest that RS suppresses inflammation in vascular endothelial cells in organs including the pancreas, suppresses TNF-α-induced pancreatic β cell damage, improves insulin secretion from β cells, and suppresses the increase in blood glucose levels. Additionally, because RS also has an α-amylase activity inhibitory effect [20,21], RS intake could suppress the absorption of MD-glucose, thereby suppressing the increase in postprandial blood glucose levels.
Iba1, a microglial activation marker that increases during inflammatory responses and the exacerbation of neurological diseases [26,28] was specifically expressed in the MD group. This indicates that inflammation occurs in the brain under hyperglycemia and is thought to induce cognitive dysfunction. Furthermore, this study showed that RS administration significantly reduced Iba1 expression in the brain, suggesting that RS suppressed microglial cell activation during glucose loading-induced brain inflammation.
AGE accumulation in the brain was significantly increased in the MD group. It has been suggested that AGEs are formed and accumulated in a chronic hyperglycemic state [29], and that accumulated AGEs are associated with hippocampal atrophy [12]. The formation and accumulation of AGEs is related to the increased production of carbonyl compounds produced from sugars by the non-enzymatic Maillard reaction and the breakdown of the detoxification system for carbonyl compounds [30]. Diseases such as diabetes are thought to promote this reaction and are considered to be one of the causes of diabetic complications [30]. Therefore, reducing sugar in the body reduces the production of carbonyl compounds and suppresses the formation of AGEs. The receptor for AGE (RAGE) is known to play an important role in inflammation development in the brain [30]. AGEs generated by chronic hyperglycemia are recognized by RAGE, and AGEs themselves increase the expression of RAGE, promote the production of intracellular oxidative stress, and activate NF-κB [8,31,32]. Furthermore, hyperglycemia is known to induces RAGE-dependent hippocampal spatial memory impairment [33]. In this study, moderate RS intake (300 mg/kg or 1000 mg/kg) tended to suppress AGE accumulation in the brain hippocampus tissue of hyperglycemic mice. It has been reported that systemic inflammation changes the site of RAGE expression from endothelial cells in various parts of the brain to neurons [34]. As RS is known to suppress inflammation in vascular endothelial cells [23] and throughout the body [24], it is speculated that the ingestion of RS may suppress RAGE expression in organs, including the brain, and thus suppress AGE accumulation in the brain.
Another important anti-inflammatory effect of RS is to protect the glycocalyx layer on the vascular endothelium [24]. The glycocalyx layer is very fragile and is destroyed by chronic inflammatory diseases such as arteriosclerosis, hyperglycemia, and diabetes [35,36,37,38]. In a hyperglycemic state, the synthesis of heparan sulfate and hyaluronic acid from glycosaminoglycans bound to the glycocalyx layer is inhibited and their detachment is promoted [35]. Mice administered MD are in a state of chronic inflammation [39]. Therefore, it is thought that the synthesis of heparan sulfate and hyaluronic acid is inhibited under inflammation, and that hyaluronidase secreted from inflamed endothelial cells degrades the glycocalyx layer of the pancreatic vascular endothelium [40,41]. We reported that RS administered to mice with LPS-induced inflammation protected the glycocalyx by inhibiting the degradation of syndecan-4, a component of glycosaminoglycans [24].
Regarding the mechanism of RS absorption in the body, because RS is a sulfated rhamnose polymer with a molecular weight of 50–100 kDa, it is difficult to transport directly into the brain. Instead, it is absorbed into the bloodstream via intestinal M cells and exerts anti-inflammatory effects on the vascular endothelium [21].
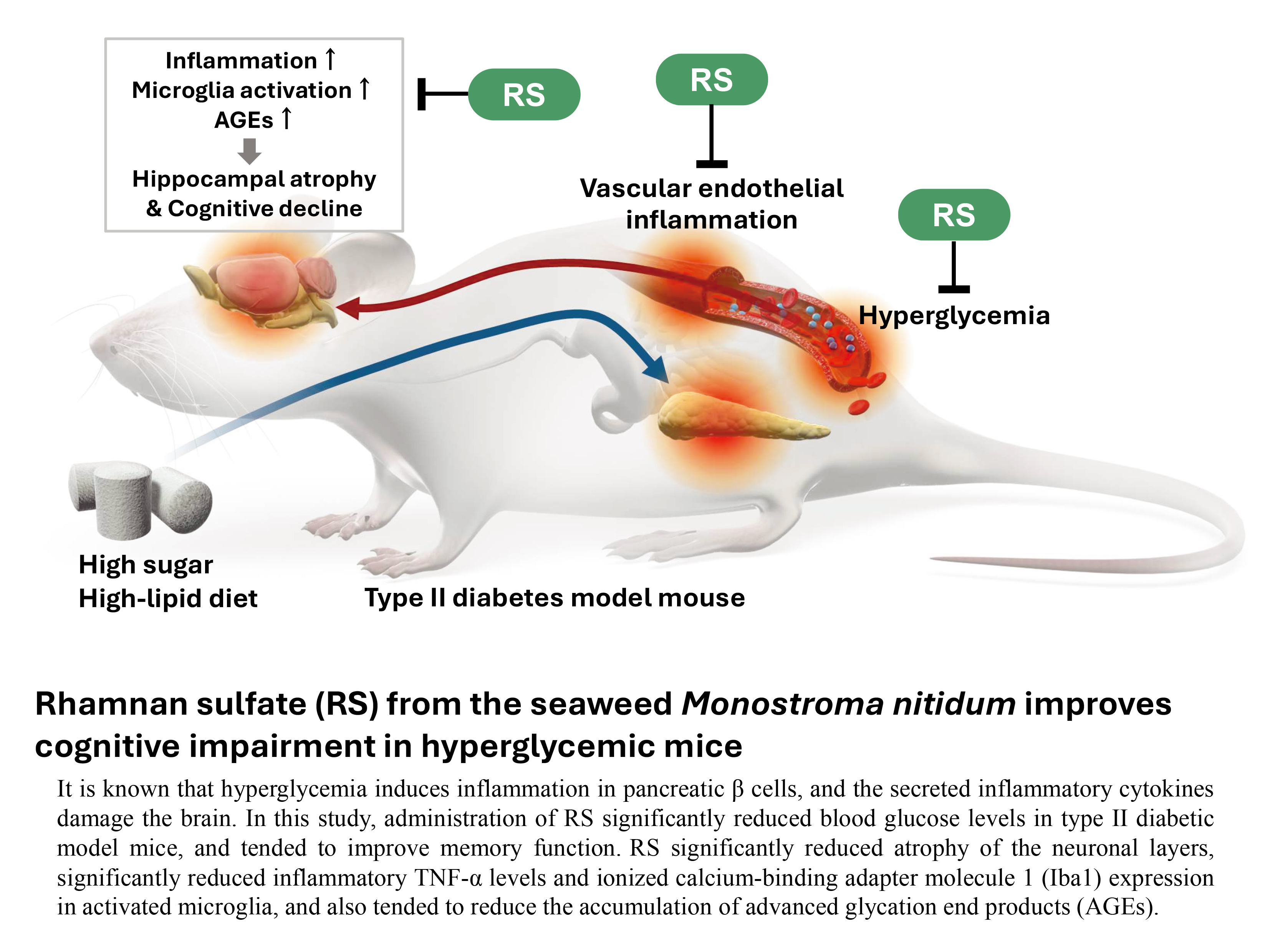
In this study, RS did not show a clear improvement in behavioral and pharmacological tests, such as cognitive function, but showed a weak tendency for improvement in memory impairment. However, a clear improvement with RS was observed in physiological tests, such as the effect on blood glucose levels, and in pathological tests, such as hippocampal atrophy and changes in the expression of inflammation-related molecules. These results suggest that continuous long-term intake of RS may improve cognitive function. Based on these results, a putative diagram of the process of brain inflammation and cognitive impairment caused by hyperglycemia and the mechanism by which RS administration suppresses brain inflammation and improves cognitive impairment are shown in Figure 9.
Epidemiological studies have shown that the incidence of Alzheimer’s disease is more than twice as high in people with type 2 diabetes than in those without diabetes [3,6]; however, it is unclear why patients with diabetes are more likely to develop Alzheimer’s disease. Insulin resistance observed in patients with diabetes leads to the accumulation of amyloid-β, the causative agent of Alzheimer’s disease [12]. In response to this, Wakabayashi et al. [42] revealed that when type 2 diabetes develops due to ingestion of a high-fat diet, the action of insulin in the blood is reduced, but the transfer of insulin to the brain is reduced, resulting in insulin resistance in the brain. They also showed that amyloid-β clearance rate is reduced and its accumulation is increased in patients with diabetes. In fact, it has been suggested that various anti-diabetic drugs suppress the accumulation of amyloid-β [43], and a study by the American Academy of Neurology reported that people taking anti-diabetic drugs have a lower risk of Alzheimer’s disease than those not taking them [44]. In relation to this study, it is unclear whether hyperglycemia-induced brain inflammation leads to insulin resistance in the brain, slowing the removal rate of amyloid-β and causing amyloid-β to accumulate in the brain. Furthermore, it is unclear whether administration of RS, which has the effect of improving hyperlipidemia [25], reduces the amount of amyloid-β accumulation in the brain. These questions require further investigation.
In this study, we were able to reveal that RS improves blood glucose levels in hyperglycemic mice, shows a tendency to improve memory function in behavioral pharmacological tests, inhibits atrophy of the neural layers in each region of the hippocampus, suppresses inflammation in the brain, and tends to suppress the accumulation of AGEs in the brain. However, there are limitations to clarifying the causal relationship between these individual results, and they remain in the realm of speculation, so further investigation is required to determine the causal relationships between the results.
4. Materials and Methods
4.1. Experimental Animals
A mouse model of type 2 diabetes was created by administrating nicotinamide (NA) (Wako Pure Chemicals, Tokyo, Japan) and streptozotocin (STZ) (Wako Pure Chemicals, Tokyo, Japan) to C57BL/6J mice (7 weeks old) (CLEA Japan, Tokyo, Japan) [45,46,47]. Mice were fasted for 24 h, after which 1.2% w/v NA was administered intraperitoneally at 240 mg/kg mouse body weight, followed 15 min later by 1% w/v STZ administered intraperitoneally at 100 mg/kg mouse body weight, followed by treatment with 1% maltodextrin (MD) (SLC, Shizuoka, Japan), which is a polysaccharide with 2-20 D-glucose sugar chains, for 4 months [12]. This procedure was repeated twice every other day to create a mouse model of type 2 diabetes. After this procedure, the MD-loaded group was fed a high-fat diet, DIO Rodent Purified Diet w/60% Energy From Fat-Blue (SLC, Shizuoka, Japan), and 1% MD solution as drinking water. The DIO Rodent Purified Diet with w/60% Energy from Fat-Blue is a high-fat diet for rodents (60% of the energy comes from fat). The RS solution was prepared using Rhamnox 100 (Konan Chemical Manufacturing, Mie, Japan) containing 70% RS, diluted with tap water, or 1% MD solution to contain 100, 300, or 1,000 mg RS mouse body weight. Five groups of mice were created, with six mice in each group: a control group fed ad libitum with normal food and tap water, 1% MD group, 1% MD + RS (100 mg/kg) group (MD + RS100 group), 1% MD + RS (300 mg/kg) group (MD + RS300 group), and 1% MD + RS (1000 mg/kg) group (MD + RS1000 group). Body weights and blood glucose levels were measured monthly during the rearing period. Blood glucose levels were measured by drawing blood from the tail vein of the mice and measuring blood glucose levels using a human self-test glucose meter (Nipro Freestyle Freedom Lite) and a blood glucose-measuring electrode (Nipro FS blood glucose-measuring electrode Lite). The blood glucose level was set at an upper limit of 500 mg/dL because the upper limit of the measuring device was 500 mg/dL. The breeding method and schedule for the type 2 diabetes model used in this study are shown in Figure 10.
4.2. Behavioral Pharmacological Test
After a 4-month glucose tolerance test, spontaneous locomotor activity was measured, followed by a rotarod test and 8-way maze test. Spontaneous locomotor activity was measured using a locomotor activity measuring device (MERCQUEST Y. K, Toyama, Japan), which quantifies activity using an infrared sensor attached to the bottom of the cage. The cage used was the same as a normal breeding cage; one animal from each concentration group was placed in a new environment (without instruction), and the spontaneous movement of each mouse was measured for 20 min. In the rotarod test (Shin Factory, Fukuoka, Japan), the rotation speed of the rod was set to 10 rpm, and the mouse was placed on the roller lane of the rotarod. The time taken for the mouse to fall into the lane was recorded as fall latency with a cutoff value of 360 s. One forced run was performed until the mouse fell or until 360 s elapsed. The eight-way maze test (BioResearch, Nagoya, Japan) was performed using food as a reward for the fasting mice from the previous day. The MD solution administered to the glucose-loaded group was replaced with drinking water during fasting. The task completion time was defined as the time taken to finish eating the three foods, and the number of errors was counted as the number of times the mouse entered the room without eating the food, with a cut-off time of 10 min. The rotarod and eight-way maze tests were performed three times after 4 months of glucose loading; the first and second tests were used as training tests, and the third test was used as data.
4.3. Brain Sample Preparation
After performing the behavioral tests, the mice were anesthetized with barbital sodium salt solution (120 mg/kg). Brain tissue samples were fixed in 4% formaldehyde in phosphate-buffered saline (PBS) for 1 d, followed by dehydration and paraffin infiltration, after which the brains were embedded in paraffin blocks and sectioned to a thickness of 6 μm using Leica Microsystems (Wetzlar, Germany).
4.4. Histochemical and Immunohistochemical Analysis of Brain Samples
Histopathological changes in the brain were analyzed using hematoxylin-eosin (HE) staining. Immunohistochemical staining was performed to measure brain inflammation. Levels of inflammatory factor TNF-α was measured using anti-TNF-α antibody (Abcam ab1793, USA). Microglial activation was measured using an anti-Iba1antibody (FUJIFILM 013-27691, Tokyo, JAPAN). The levels of glycation products were measured using an anti-AGE antibody (Abcam ab23722, USA). The paraffin-embedded mouse brain sections were deparaffinized with xylene, hydrophilized with 100, 90, 80, 70, or 50% (v/v) ethanol, and immersed in PBS for 30 min. To further activate antigens, sections were immersed in 5% (w/v) urea and heated in a microwave oven. Endogenous peroxidase activity was blocked in 1% H2O2 in PBS, followed by immunohistochemical staining for 30 min in PBS containing 3% (v/v) normal goat serum antibody buffer. Rabbit polyclonal anti-TNF-α antibody (2 mg/ml, 1:400), anti-Iba1 antibody (2 mg/ml, 1:400) and anti-AGE antibody (2 mg/ml, 1:400) were used as primary antibodies and incubated overnight at room temperature. Cells were washed with PBS and detected using a biotinylated horse anti-rabbit immunoglobulin kit (Vector Laboratories, Vectastain Elite ABC-HRP Kit, PK6101, and PK6102). Immune cells were detected using a 3,3’-diaminobenzidine peroxidase staining DAB kit (Nacalai Tesque, Kyoto, Japan). Nuclear counterstaining of AGE-stained samples was observed on glass slides after hematoxylin staining and sealing with marinol (Muto Chemicals, Tokyo, Japan). Morphological observations were performed using a microscope and a digital camera (BX53, DP74, Olympus, Tokyo, Japan). For the H&E-stained specimens, the thickness of the neuronal layer in the CA1, CA2, and CA3 regions and the dentate gyrus was measured at 10 points using ImageJ (National Institutes of Health, Bethesda, MD, USA), and the average of the 10 measured points was quantified.
4.5. Statistical Analysis
Dunnet’s test and one-way analysis of variance were performed for body weight, blood glucose level, neuronal layer width in each part of the hippocampus, and behavioral pharmacological test (locomotor activity, rotarod test, and eight-way maze test), and chi-square test was performed for AGE1, Iba1, and TNF-α. The statistical analysis software SPSS version 28 (IBM) was used for all data, and a significance level of less than 5% (p < 0.05) was considered significant.
5. Conclusions
The results of this study showed that the administration of RS may prevent cognitive decline in a hyperglycemic mouse model. The mechanism of action was suggested to be that PS suppresses the rise in blood glucose levels, inhibits the production of inflammatory factors such as TNF-α, inhibits the activation of glia cells, inhibits hippocampal atrophy, and improves hyperglycemia-induced cognitive impairment, including the repair of brain organs, in response to inflammatory cognitive impairment caused by hyperglycemia. These findings suggest that RS intake is useful in preventing brain decline and alleviating symptoms.
Author Contributions
Conceptualization, T. M. and K. S.; methodology, T. M., M. S., N. M., and M. T.; validation, T. M., M. S., N. M., M. T., and K. S.; resources, T. M., M. T., and K. S.; writing—original draft preparation, M. S., T. M., and M. T.; writing, review, and editing, T. M. and K. S.; visualization, N. M., M. T., and K. S.; supervision, T. M. and K. S.; project administration, K. S.; funding acquisition, K. S. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported in part by the Japan Society for the Promotion of Science (Grants to K. S., numbers 19K08850), and the fund of the collaborative research (principal investigator: Koji Suzuki) between Suzuka University of Medical Science and Konan Chemical Manufacturing Co., Ltd.
Institutional Review Board Statement
This study was approved by the Institutional Animal Care and Use Committee and was carried out according to the Suzuka University of Medical Science Animal Experimentation Regulations (principal investigator: Koji Suzuki, Approval Number: 84, Approval Date: March 22, 2022).
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
We would like to thank Editage (www.editage.com) for the English language editing.
Conflicts of Interest
Koji Suzuki conducted joint research on the anti-inflammatory effects of RS with Konan Chemical Manufacturing Co., Ltd. and received research funding from the company. Masahiro Terasawa is affiliated with Konan Chemical Manufacturing Co., Ltd. The authors declare no conflicts of interest.
References 10, 11, and 12 are currently unavailable to anyone other than members of the Japanese Society of Psychiatric Pharmacology. This is because the journal in which these articles are published, “Jpn. Soc. Psychiatr. Pharm.”, is currently in the process of registering the DOI. Until registration is complete, please contact Dr. Takaichi Miwa (e-mail address: t-miwa@suzuka-u.ac.jp), the person responsible for these articles, and request a PDF file of the article.
Additional notice
References 10, 11, and 12 are currently unavailable to anyone other than members of the Japanese Society of Psychiatric Pharmacology. This is because the journal in which these articles are published, “Jpn. Soc. Psychiatr. Pharm.”, is currently in the process of registering the DOI. Until registration is complete, please contact Dr. Takaichi Miwa (e-mail address: t-miwa@suzuka-u.ac.jp), the person responsible for these articles, and request a PDF file of the article.
Abbreviations
The following abbreviations are used in this manuscript:
| AGE | advanced glycation end products |
| HE | hematoxylin-eosin |
| Iba1 | ionized calcium binding adapter molecule 1 |
| MD | maltodextrin |
| NA | nicotinamide |
| PBS | phosphate-buffered saline |
| PPAR | peroxisome proliferator-activated receptor |
| RAGE | receptor of AGE |
| RS | rhamnan sulfate |
| SD | standard deviation |
| STZ | streptozotocin |
| TNF | tumor necrosis factor |
References
- Launer, L.J.; Miller, M.E.; Williamson, J.D.; Lazar, R.M.; Gerstein, H.C.; Murray, A.M.; Sullivan, M.; Horowitz, K.R.; Ding, J.; Marcovina, S.; et, al. Effects of intensive glucose lowering on brain structure and function in people with type 2 diabetes (ACCORD MIND): a randomized open-label substudy. Lancet Neurol. 2011, 10, 969–977. [Google Scholar] [CrossRef] [PubMed]
- Mukai, N.; Doi, Y.; Ninomiya, T.; Hirakawa, Y.; Nagata, M.; Yoshida, D.; Hata, J.; Fukuoka, M.; Nakamura, U.; Kitazono, T.; Kiyohara, Y. Trends in the prevalence of type 2 diabetes and prediabetes in community-dwelling Japanese subjects: The Hisayama Study. J. Diabetes Invest. 2014, 5, 162–169. [Google Scholar] [CrossRef]
- Kopf, D.; Frolich, L. Risk of incident Alzheimer’s disease in diabetic patients: a systematic review of prospective trials. J. Alzheimers Dis. 2009, 16, 677–685. [Google Scholar] [CrossRef] [PubMed]
- Peila, R.; Rodriguez, B.L.; Launer, L.J. Type2 diabetes, APOE Gene, and the Risk for Dementia and Related Pathologies: The Honolulu-Asia Aging Study. Diabetes 2002, 51, 1256–1262. [Google Scholar] [CrossRef]
- Ott, A.; Stolk, R.P.; van Harskamp, F.; Pols, H.A.; Hofman, A.; Breteler, M.M. Diabetes mellitus and the risk of dementia: The Rotterdam Study. Neurology 1999, 53, 1937–1942. [Google Scholar] [CrossRef] [PubMed]
- Ohara, T.; Doi, Y.; Ninomiya, T.; Hata, J.; Iwaki, T.; Kanda, S.; Kiyohara, Y. Glucose tolerance status and risk of dementia in the community: the Hisayama study. Neurology 2011, 7, 1126–1134. [Google Scholar] [CrossRef] [PubMed]
- Adav, S.S.; Sze, S.K. Insight of brain degenerative protein modifications in the pathology of neurodegeneration and dementia by proteomic profiling. Mol. Brain 2016, 9, 92. [Google Scholar] [CrossRef]
- Yamagishi, S.; Imaizumi, T. Diabetic vascular complications: pathophysiology, biochemical basis and potential therapeutic strategy. Curr. Pharm. Des. 2005, 11, 2279–2299. [Google Scholar] [CrossRef]
- Baglietto-Vargas, D.; Shi, J.; Yager, D.M.; Ager, R.; LaFerla, F.M. Diabetes and Alzheimer’s disease crosstalk. Neurosci. Biobehav. Rev. 2016, 64, 272–287. [Google Scholar] [CrossRef]
- Sato, M.; Wada, T.; Ban, S.; Fujita, K.; Miwa, T. Cognitive function investigation in patients with Alzheimer’s disease with diabetes in a psychiatric hospital. Jpn. Soc. Psychiatr. Pharm. 2022, 6, 52–59. [Google Scholar]
- Sato, M.; Wada, T.; Furukawa, A.; Koriyama, Y.; Miwa, T. Effects of loading glucose on cognitive function in mice. Jpn. Soc. Psychiatr. Pharm. 2023, 6, 87–95. [Google Scholar]
- Sato, M.; Ning, Ma.; Miwa, T. Effects of glucose loading on cognitive function in streptozotocin-induced hyperglycemic mouse models. Jpn. Soc. Psychiatr. Pharm. 2023, 6, 96–106. [Google Scholar]
- Sato, T.; Hanyu, H.; Hirao, K.; Kanetaka, H.; Sakurai, H.; Iwamoto, T. Efficacy of PPAR-γ agonist pioglitazone in mild Alzheimer disease. Neurobiol. Aging 2011, 32, 1626–1633. [Google Scholar] [CrossRef]
- Rizzo, M.R.; Barbieri, M.; Boccardi, V.; Angellotti, E.; Marfella, R.; Paolisso, G. Dipeptidyl peptidase-4 inhibitors have protective effect on cognitive impairment in aged diabetic patients with mild cognitive impairment. J. Gerontol. A Biol. Sci. Med. Sci. 2014, 69, 1122–1131. [Google Scholar] [CrossRef] [PubMed]
- Cunha, L.; Grenha, A. Sulfated seaweed polysaccharides as multifunctional materials in Drug Delivery Applications. Mar. Drugs 2016, 14, 42. [Google Scholar] [CrossRef]
- Ye, F.; Xiao, J.; Yu, N.; Yi, G. Impact of Insulin Sensitizers on the Incidence of Dementia:A Meta-Analysis. Dement. Geriatr. Cogn. Disord. 2016, 41, 251–260. [Google Scholar] [CrossRef]
- Abbatecola, A.M.; Rizzo, M.; Grella, R.; Arciello, A.; Laieta, M.T.; Acampora, R.; Passariello, N.; Cacciapuoti, F.; Paolisso, G. Postprandial plasma glucose excursions and cognitive functioning in aged type 2 diabetics. Neurology, 2006, 67, 235–240. [Google Scholar] [CrossRef]
- Li, H.; Mao, W.; Hou, Y.; Gao, Y.; Qi, X.; Zhao, C.; Chen, Y.; Chen, Y.; Li, N.; Wang, C. Preparation, structure and anticoagulant activity of a low molecular weight fraction produced by mild acid hydrolysis of sulfated rhamnan from Monostroma latissimum. Bioresour. Technol. 2012, 114, 414–418. [Google Scholar] [CrossRef]
- Harada, N.; Maeda, M. Chemical structure of antithrombin-active Rhamnan sulfate from Monostroma nitidum. Biotechnol. Biochem. 1998, 62, 1647–1652. [Google Scholar] [CrossRef]
- Zang, L.; Shimada, Y.; Tanaka, T.; Nishimura, N. Rhamnan sulphate from Monostroma nitidum attenuates hepatic steatosis by suppressing lipogenesis in a diet-induced obesity zebrafish model. J. Funct. Foods. [CrossRef]
- Shimada, Y.; Terasawa, M.; Okazaki, F.; Nakayama, H.; Zang, L.; Nishiura, K.; Matsuda, K.; Nishimura, N. Rhamnan sulphate from green algae Monostroma nitidum improves constipation with gut microbiome alteration in double-blind placebo-controlled trial. Sci. Rep. 2021, 11, 13384. [Google Scholar] [CrossRef]
- Shimada, Y.; Zang, L.; Ishimaru, T.; Nishiura, K.; Matsuda, K.; Uchida, R.; Nakayama, H.; Matsuoka, I.; Terasawa, M.; Nishimura, N. Lipid-and glucose-lowering effects of Rhamnan sulphate from Monostroma nitidum with altered gut microbiota in mice. doi: 10.1002/fsn3.4100. Food Sci. Nutr. 2024, 12, 4342–4352. [CrossRef]
- Okamoto, T.; Akita, N.; Terasawa, M.; Hayashi, T.; Suzuki, K. Rhamnan sulfate extracted from Monostroma nitidum attenuates blood coagulation and inflammation of vascular endothelial cells. J. Nat. Med. 2019, 73, 614–619. [Google Scholar] [CrossRef]
- Terasawa, M.; Hiramoto, K.; Uchida, R.; Suzuki, K. Anti-Inflammatory activity of orally administered Monostroma nitidum rhamnan sulfate against lipopolysaccharide-induced damage to mouse organs and vascular endothelium. Mar. Drugs 2022, 20, 121. [Google Scholar] [CrossRef] [PubMed]
- Terasawa, M.; Zang, L.; Hiramoto, K.; Shimada, Y.; Mitsunaka, M.; Uchida, R.; Nishiura, K.; Matsuda, K.; Nishimura, N.; Suzuki, K. Oral administration of rhamnan sulfate from Monostroma nitidum suppresses atherosclerosis in ApoE-deficient mice fed a high-fat diet. Cells 2023, 12, 2666. [Google Scholar] [CrossRef]
- Ito, D. : Imai.; Y., Ohsawa, K..; Nakajima, K., Fukuuchi, Y.; Kohsaka, S. Microglia-specific localization of a novel calcium binding protein, Iba1. Brain Res. Mol. Brain Res. 1998, 57, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Donath, M.Y.; Shoelson, S.E. Type 2 diabetes as an inflammatory disease. Nat. Rev. Immunol. 2011, 11, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Graeber, M.B.; Streit, W.J. Microglia:biology and pathology. Acta Neuropathol. 2010, 119, 89–105. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Yamagishi, S.; Yonekura, H.; Doi, T.; Tsuji, H.; Kato, I.; Takasawa, S.; Okamoto, H.; Abedin, J.; Tanaka, N.; et. al. Role of AGE-RAGE system in Vascular injury in diabetes. Ann. N.Y. Acad. Sci. 2000, 902, 163–170. [Google Scholar] [CrossRef]
- Miyata, T.; van Ypersele de Strihou, C.; Kurokawa, K.; Baynes, J.W. Alterations in nonenzymatic biochemistry in uremia: origin and significance of “carbonyl stress” in long-term uremic complications. Kidney Int. 1999, 55, 389–399. [Google Scholar] [CrossRef]
- Goldin, A.; Beckman, J.A.; Schmidt, A.M.; Creager, M.A. Advanced glycation end products: sparking the development of diabetic vascular injury. Circulation 2006, 114, 597–605. [Google Scholar] [CrossRef]
- Yamagishi, S.; Fukami, K.; Matsui, T. Crosstalk between advanced glycation end products (AGEs)-receptor RAGE axis and dipeptidyl peptidase-4-incretin system in diabetic vascular complications. Cardiovasc. Diabetol. 2015, 14, 2. [Google Scholar] [CrossRef]
- Momeni, Z.; Neapetung, J.; Pacholko, A.; Kiir, T.A.B.; Yamamoto, Y.; Bekar, L.K.; Campanucci, V.A. Hyperglycemia induces RAGE-dependent hippocampal spatial memory impairments. Physiol. Behav. 2021, 229, 113287. [Google Scholar] [CrossRef] [PubMed]
- Gasparotto, J.; Ribeiro, C.T.; da Rosa-Silva, H.T.; Bortolin, R.C.; Rabelo, T.K.; Peixoto, D.O.; Moreira, J.C.F.; Gelain, D.P. Systemic inflammation changes the site of RAGE expression from endothelial cells to neurons in different brain areas. Mol. Neurobiol. 2019, 56, 3079–3089. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Quintero, S.V.; Cancel, L.M.; Pierides, A.; Antonetti, D.; Spray, D.C.; Tarbell, J.M. High Glucose Attenuates Shear-Induced Changes in Endothelial Hydraulic Conductivity by Degrading the Glycocalyx. PLoS ONE 2013, 8. [Google Scholar] [CrossRef]
- Yeo, T.W.; Bush, P.A.; Chen, Y.; Young, S.P.; Zhang, H.; Millington, D.S.; Granger, D.L.; Mwaikambo, E.D.; Anstey, N.M.; Weinberg, J.B. Glycocalyx breakdown is increased in African children with cerebral and uncomplicated falciparum malaria. FASEB J. 2019, 33, 14185–14193. [Google Scholar] [CrossRef]
- Bar, A.; Targosz-Korecka, M.; Suraj, J.; Proniewski, B.; Jasztal, A.; Marczyk, B.; Sternak, M.; Przybylo, M.; Kurpinska, A.; Walczak, M.; et. al. Degradation of Glycocalyx and Multiple Manifestations of Endothelial Dysfunction Coincide in the Early Phase of Endothelial Dysfunction Before Atherosclerotic Plaque Development in Apolipoprotein E/Low-Density Lipoprotein Receptor-Deficient Mice. J. Am. Heart Assoc. 2019, 8. [Google Scholar] [CrossRef]
- Chelazzi, C.; Villa, G.; Mancinelli, P.; De Gaudio, A.R.; Adembri, C. Glycocalyx and sepsis-induced alterations in vascular permeability. Crit. Care 2015, 19, 26. [Google Scholar] [CrossRef]
- Kim, K.T.; Rioux, L.E.; Turgeon, S.L. Alpha-amylase and alpha-glucosidase inhibition is differentially modulated by fucoidan obtained from Fucus vesiculosus and Ascophyllum nodosum. Phytochemistry 2014, 98, 27–33. [Google Scholar] [CrossRef] [PubMed]
- van Golen, R.F.; Reiniers, M.J.; Vrisekoop, N.; Zuurbier, C.J.; Olthof, P.B.; van Rheenen, J.; van Gulik, T.M.; Parsons, B.J.; Heger, M.M. The mechanisms and physiological relevance of glycocalyx degradation in hepatic ischemia/reperfusion injury. Antioxid. Redox Signal. 2014, 21, 1098–1118. [Google Scholar] [CrossRef]
- Mulivor, A.W.; Lipowsky, H.H. Inflammation- and ischemia-induced shedding of venular glycocalyx. Am. J. Physiol. Heart Circ. Physiol. 2004, 286, 1672–1680. [Google Scholar] [CrossRef]
- Wakabayashi, T.; Yamaguchi, K.; Matsui, K.; Sano, T.; Kubota, T.; Hashimoto, T.; Mano, A.; Yamada, K.; Matsuo, Y.; Kubota, N.; et. al. Differential effects of diet- and genetically induced brain insulin resistance on amyloid pathology in a mouse model of Alzheimer’s disease. Mol. Neurodegener 2019, 14, 15. [Google Scholar] [CrossRef]
- Ztotek, M.; Kurowska, A.; Herbet, M.; Piatkowska-Chmiel, I. GLP-1 Analogs, SGLT-2, and DPP-4 Inhibitors: A Triad of Hope for Alzheimer’s Disease Therapy. Biomedicines 2023, 11, 3035. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.H.; Kim, H.R.; Kim, J.; Kim, H.; Hong, N.; Jung, J.H.; Baik, K.; Cho, H.; Lyoo, C.H.; Ye, B.S.; Sohn, Y.H.; Seong, J.-K.; Lee, PH. Association of Dipeptidyl Peptidase-4 Inhibitor Use and Amyloid Burden in Patients With Diabetes and AD-Related Cognitive Impairment. Neurology 2021, 97, 1110–1122. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, R. ; Sakazaki, F,; Okuno, T.; Nakamuro, K,; Ueno, H. Difference in glucose intolerance between C57BL/6J and ICR strain mice with streptozotocin/nicotinamide-induced diabetes. Biomed. Res, 33. [CrossRef]
- Szkudelski, T. The mechanism of alloxan and streptozotocin action in B cells of the rat pancreas. Physiol. Res. 2001, 50, 537–546. [Google Scholar] [CrossRef] [PubMed]
- Uchigata, Y, Yamamoto, H, Kawamura, A, Okamoto, H. Protection by superoxide dismutase, catalase, and poly (ADP-ribose) synthetase inhibitors against alloxan- and streptozotocin-induced islet DNA strand breaks and against the inhibition of proinsulin synthesis. J. Biol. Chem. 6: 6084–6088. PMID, 6084.
Figure 1.
Effect of rhamnan sulfate (RS) administration on body weight and blood glucose level. (A) Body weight and (B) blood glucose level after 4 months in each group are shown. RS administration did not change body weight, but blood glucose level improved significantly. Each value is expressed as mean ± SD. (*p < 0.01, **p < 0.001, ***p < 0.0001, ****p < 0.00001).
Figure 1.
Effect of rhamnan sulfate (RS) administration on body weight and blood glucose level. (A) Body weight and (B) blood glucose level after 4 months in each group are shown. RS administration did not change body weight, but blood glucose level improved significantly. Each value is expressed as mean ± SD. (*p < 0.01, **p < 0.001, ***p < 0.0001, ****p < 0.00001).
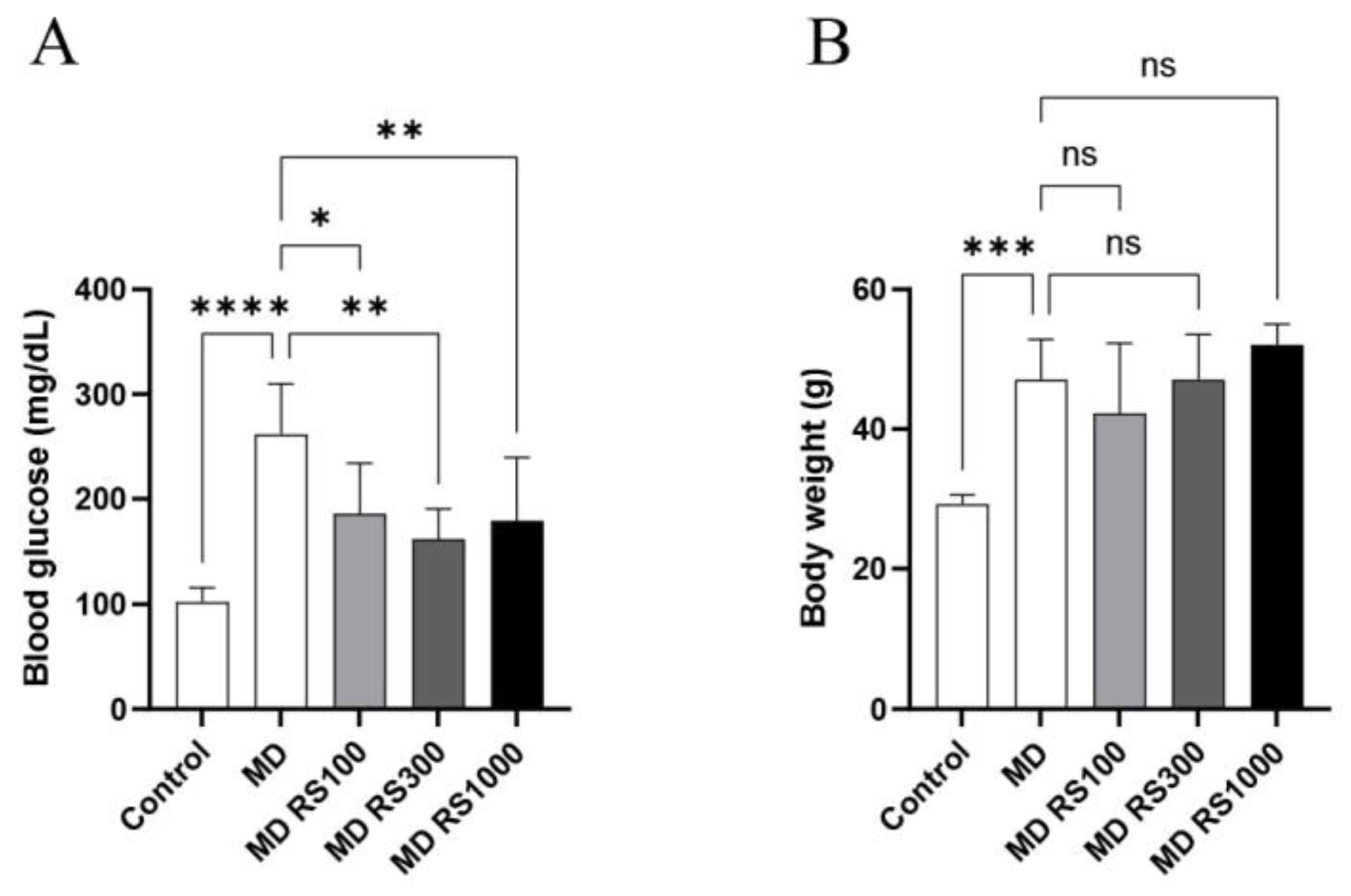
Figure 2.
Effect of Rhamnan sulfate (RS) administration on locomotion. After 4 months of MD loading, each mouse group was placed in a new environment and 20 min of spontaneous locomotion was measured to compare the differences in locomotion. RS administration had no effect on locomotion. Each value is expressed as the mean ± SD.
Figure 2.
Effect of Rhamnan sulfate (RS) administration on locomotion. After 4 months of MD loading, each mouse group was placed in a new environment and 20 min of spontaneous locomotion was measured to compare the differences in locomotion. RS administration had no effect on locomotion. Each value is expressed as the mean ± SD.
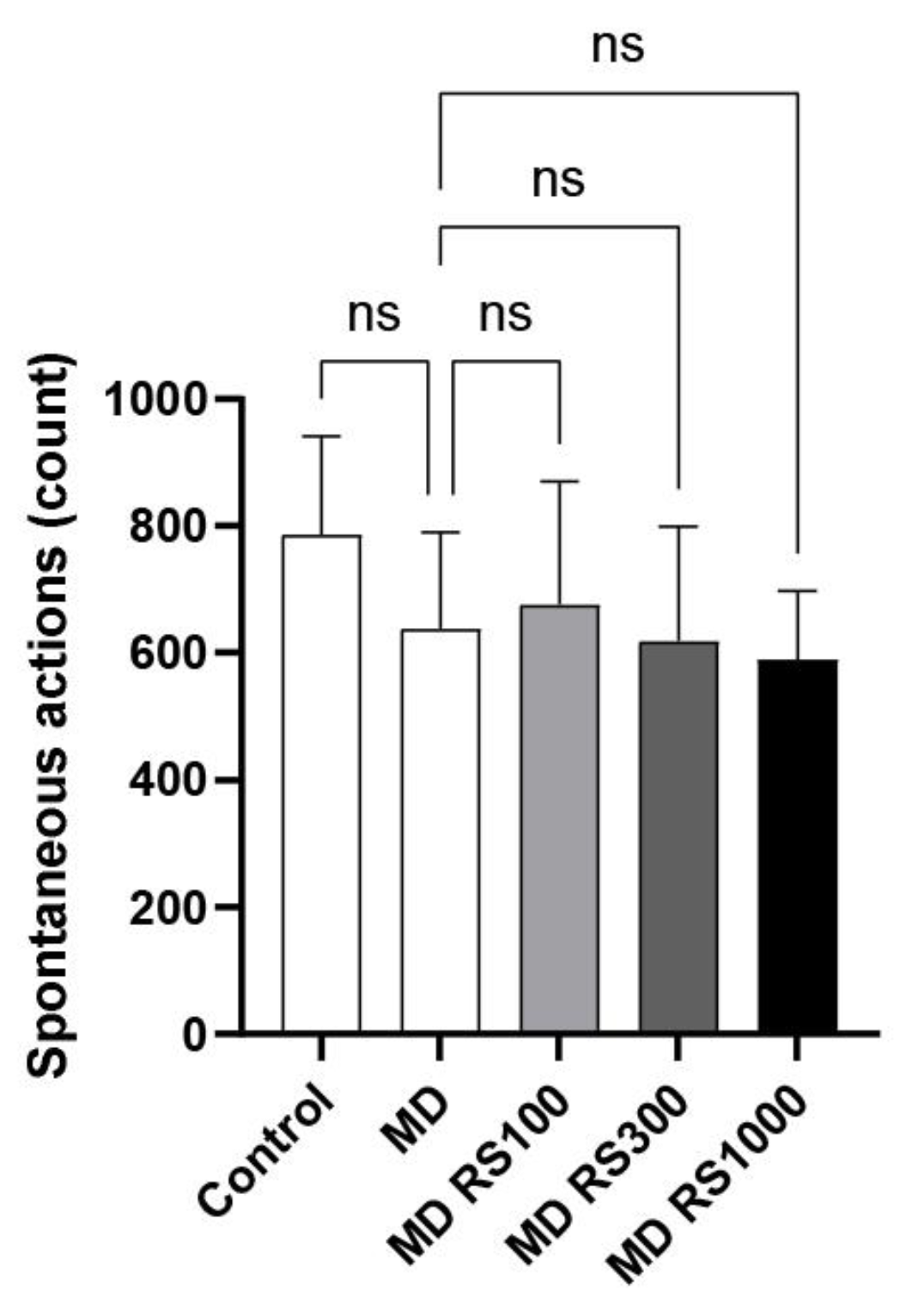
Figure 3.
Effect of rhamnan sulfate (RS) administration on motor coordination (motor function). The upper limit was set at 6 min, and the time until the mice fell was measured to evaluate and compare motor and cognitive functions. Motor function, which decreased owing to glucose loading, tended to improve with RS administration. Each value is expressed as the mean ± S.D. (***p < 0.0001).
Figure 3.
Effect of rhamnan sulfate (RS) administration on motor coordination (motor function). The upper limit was set at 6 min, and the time until the mice fell was measured to evaluate and compare motor and cognitive functions. Motor function, which decreased owing to glucose loading, tended to improve with RS administration. Each value is expressed as the mean ± S.D. (***p < 0.0001).
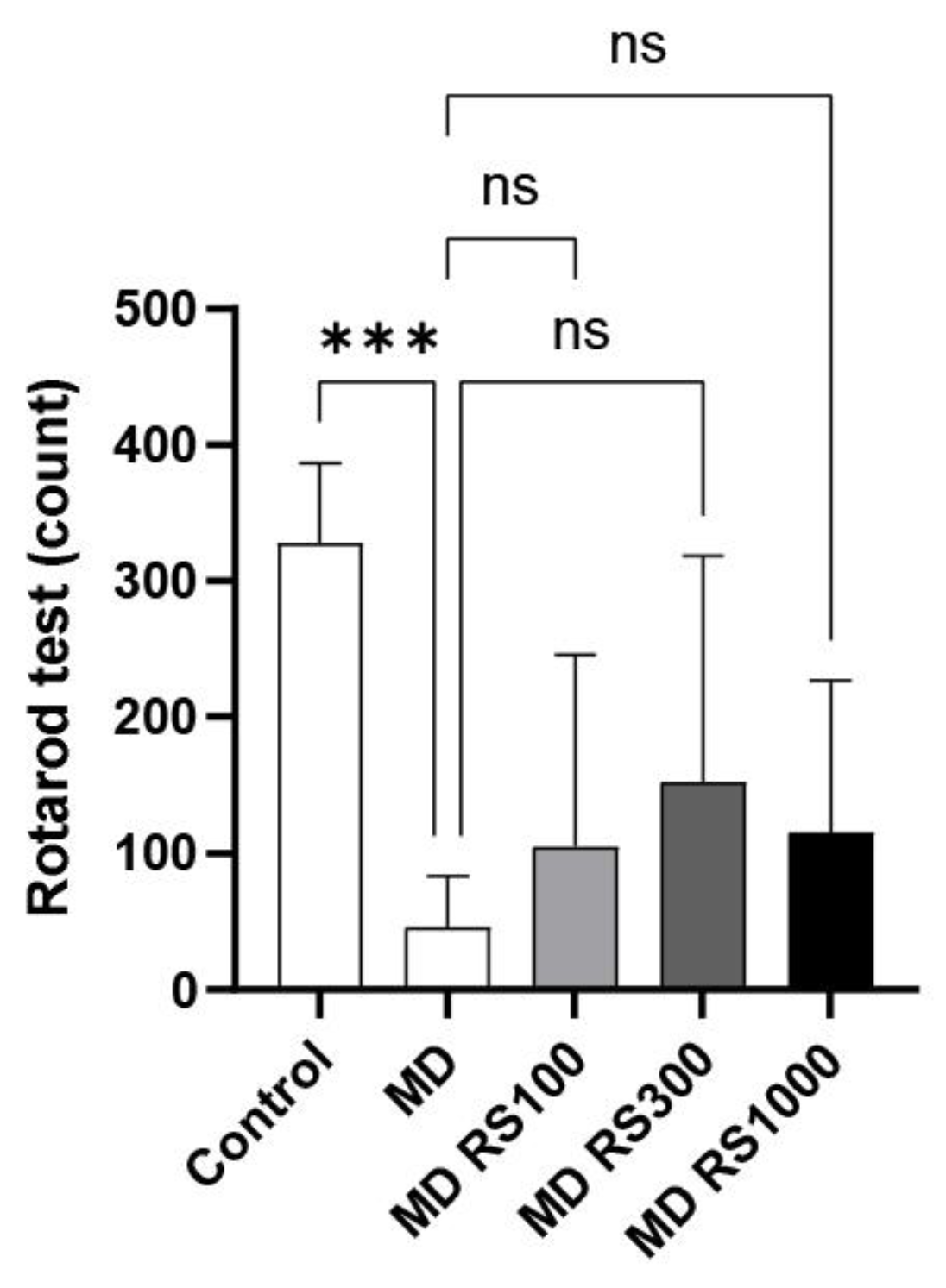
Figure 4.
Effect of rhamnan sulfate (RS) administration on memory function. Cognitive memory function was assessed and compared by measuring (A) the time it took the mice to complete a task and (B) the number of errors they made while trying three rewards in 10 min. RS tended to improve cognitive function. Each value is expressed as mean ± S.D. (****p < 0.00001 vs. control).
Figure 4.
Effect of rhamnan sulfate (RS) administration on memory function. Cognitive memory function was assessed and compared by measuring (A) the time it took the mice to complete a task and (B) the number of errors they made while trying three rewards in 10 min. RS tended to improve cognitive function. Each value is expressed as mean ± S.D. (****p < 0.00001 vs. control).
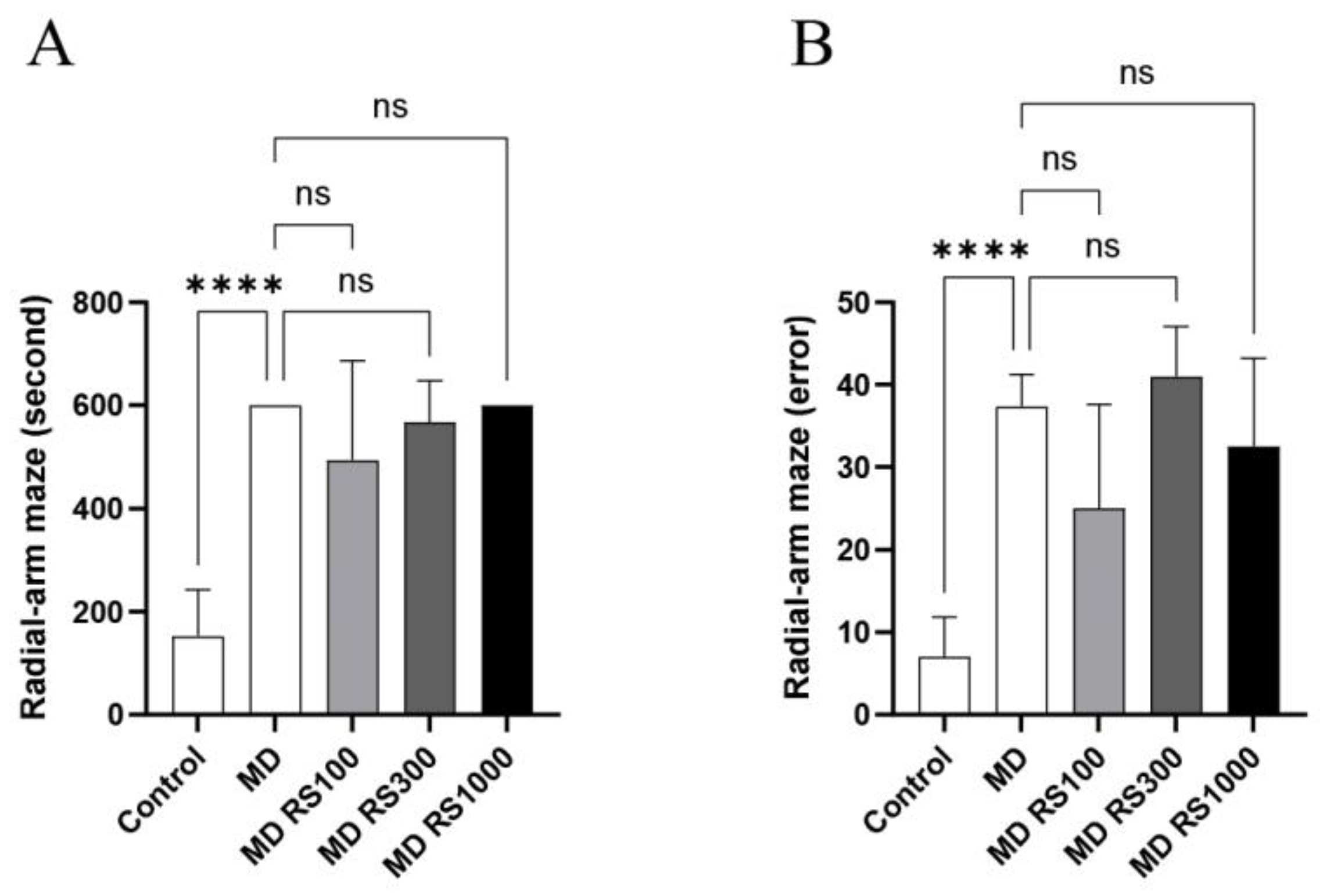
Figure 5.
Effect of rhamnan sulfate (RS) administration on the hippocampal region (neuronal layer). RS was administered to a hyperglycemic mouse model, and its effects on the hippocampus (neuronal layer) were compared. ImageJ software was used to calculate the average of 10 areas in the CA1, CA2, CA3, and dentate gyrus regions, and the results were quantified. RS improved hippocampal neuronal layer atrophy A: CA1 region, B: CA2 region, C: CA3 region, D: dentate gyrus region. Each value is expressed as the mean ± SD. ( *p < 0.01, ** p < 0.001, ***p < 0.0001, ****p < 0.00001).
Figure 5.
Effect of rhamnan sulfate (RS) administration on the hippocampal region (neuronal layer). RS was administered to a hyperglycemic mouse model, and its effects on the hippocampus (neuronal layer) were compared. ImageJ software was used to calculate the average of 10 areas in the CA1, CA2, CA3, and dentate gyrus regions, and the results were quantified. RS improved hippocampal neuronal layer atrophy A: CA1 region, B: CA2 region, C: CA3 region, D: dentate gyrus region. Each value is expressed as the mean ± SD. ( *p < 0.01, ** p < 0.001, ***p < 0.0001, ****p < 0.00001).
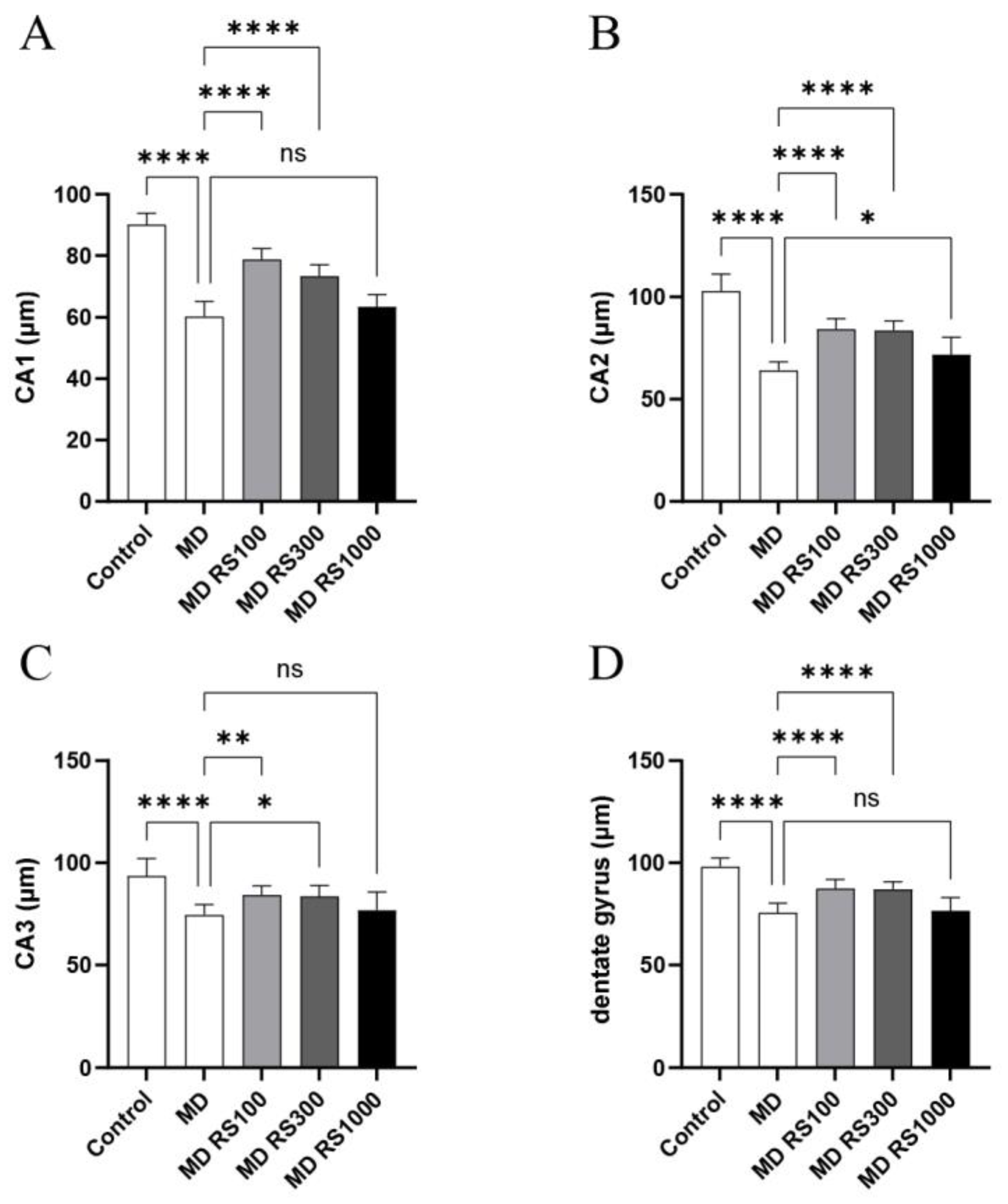
Figure 6.
Effect of rhamnan sulfate (RS) on TNF-α expression in hyperglycemic mice. Stained images of TNF-α in each group (A: control group, B: MD group, C: MD + RS100 group, D: MD + RS300 group). Scale bar: 100 μm.
Figure 6.
Effect of rhamnan sulfate (RS) on TNF-α expression in hyperglycemic mice. Stained images of TNF-α in each group (A: control group, B: MD group, C: MD + RS100 group, D: MD + RS300 group). Scale bar: 100 μm.
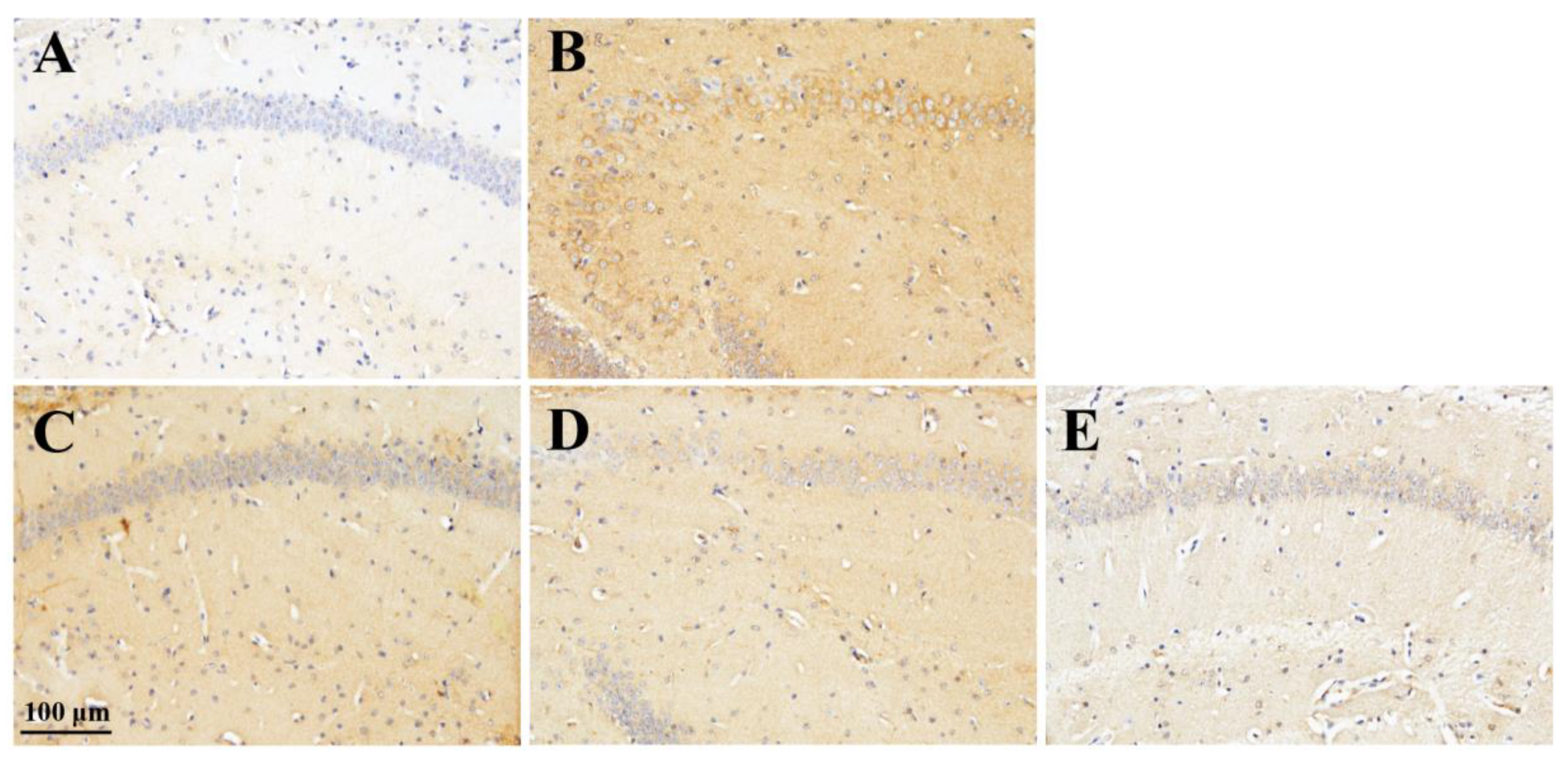
Figure 7.
Effect of rhamnan sulfate (RS) on microglial activation in hyperglycemic mice. Stained images of Iba1 in each group (A, control group; B, MD group; C, MD + RS100 group; D, MD + RS300 group; E, MD + RS1000 group). Scale bar: 100 μm.
Figure 7.
Effect of rhamnan sulfate (RS) on microglial activation in hyperglycemic mice. Stained images of Iba1 in each group (A, control group; B, MD group; C, MD + RS100 group; D, MD + RS300 group; E, MD + RS1000 group). Scale bar: 100 μm.
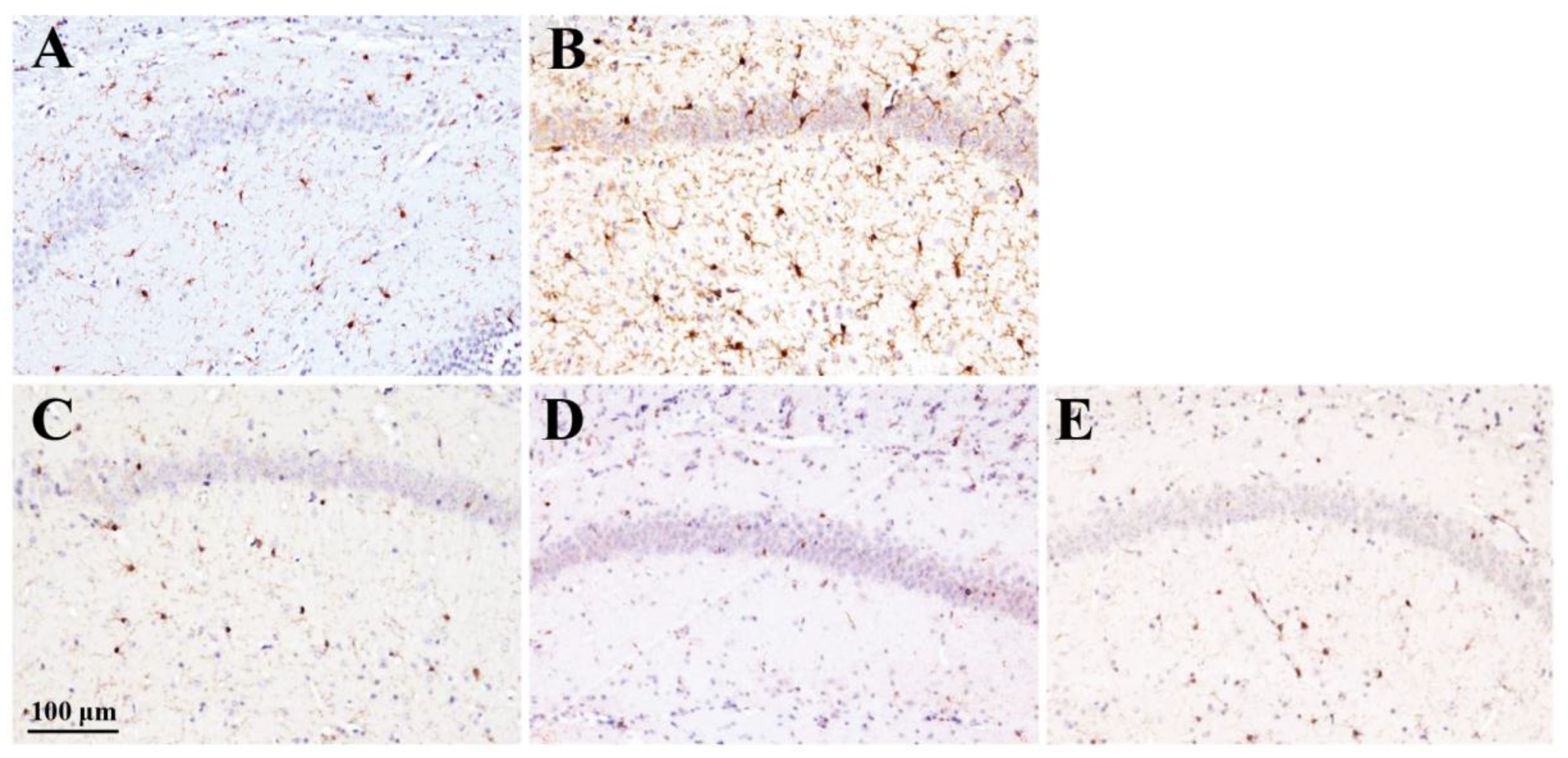
Figure 8.
Effect of rhamnan sulfate (RS) administration on AGEs accumulation levels in hyperglycemic mice. AGEs staining images of each group (A, control group; B, MD group; C, MD + RS100 group; D, MD + RS300 group; and E, MD + RS1000 group). Scale bar: 100 μm.
Figure 8.
Effect of rhamnan sulfate (RS) administration on AGEs accumulation levels in hyperglycemic mice. AGEs staining images of each group (A, control group; B, MD group; C, MD + RS100 group; D, MD + RS300 group; and E, MD + RS1000 group). Scale bar: 100 μm.
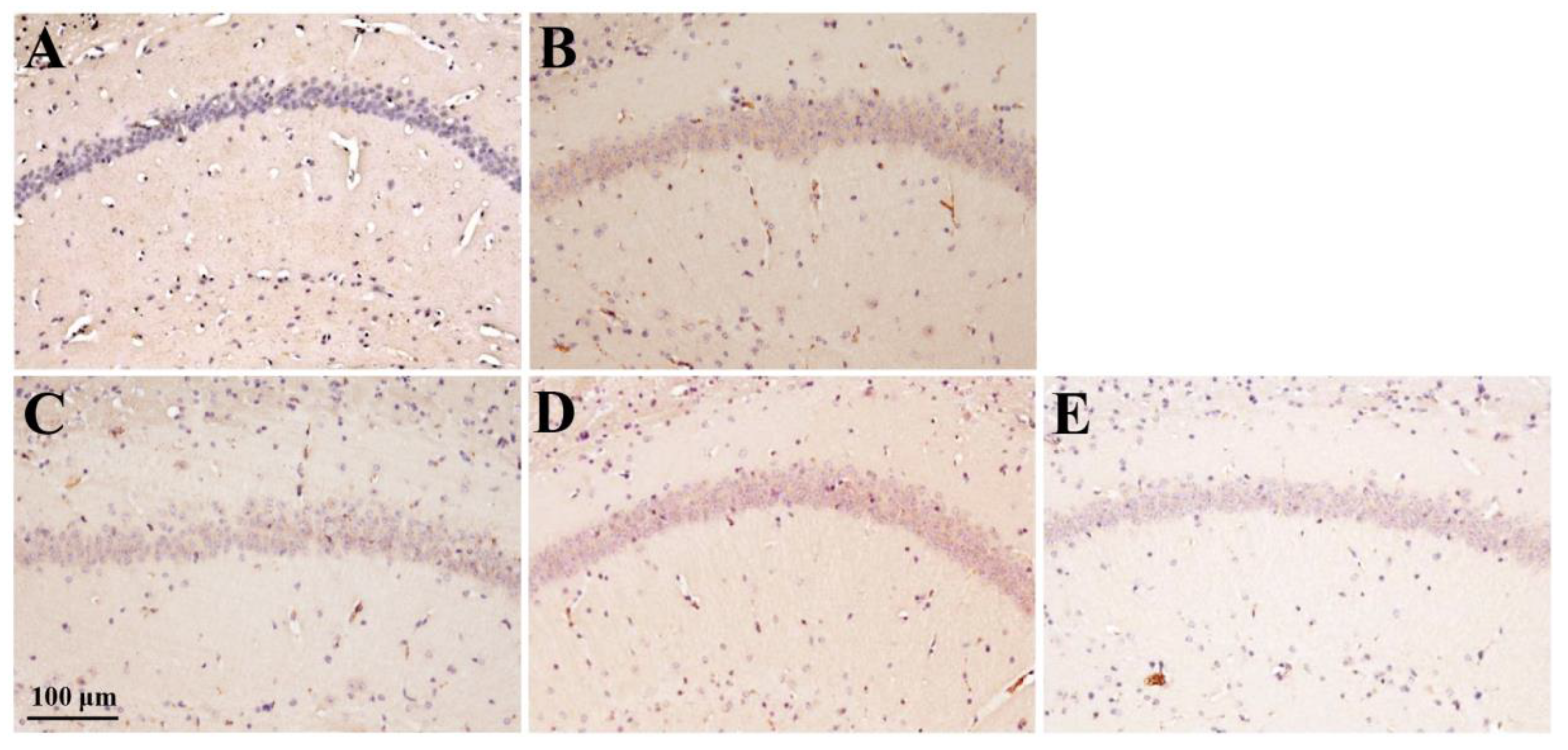
Figure 9.
Hypothesized mechanism of the process of hyperglycemia-induced brain inflammation and cognitive decline, and mechanism by which rhamnan sulfate (RS) suppresses brain inflammation and improves cognitive decline. AGE: advanced glycation products.
Figure 9.
Hypothesized mechanism of the process of hyperglycemia-induced brain inflammation and cognitive decline, and mechanism by which rhamnan sulfate (RS) suppresses brain inflammation and improves cognitive decline. AGE: advanced glycation products.
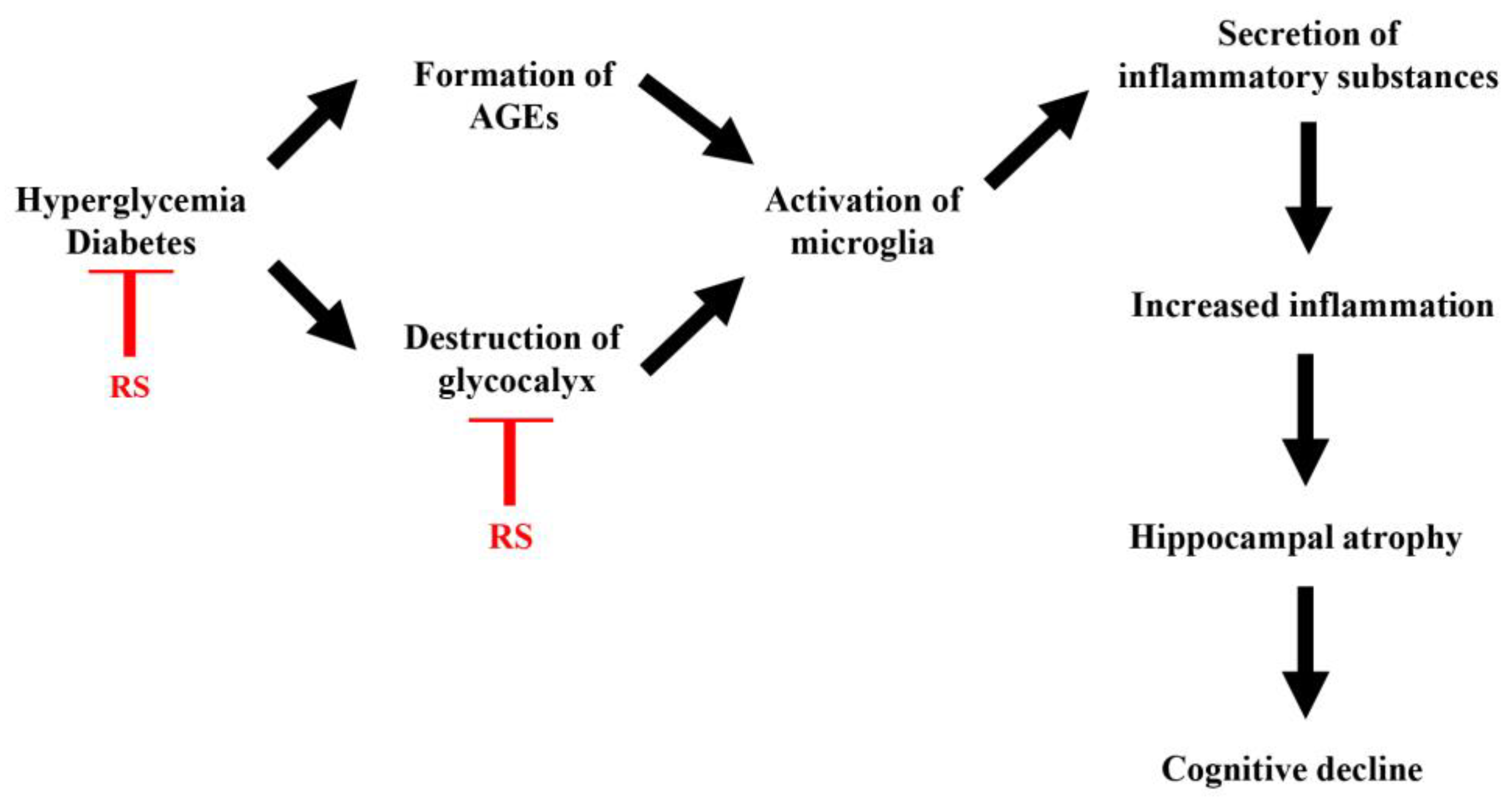
Figure 10.
A protocol for creating a mouse model of type 2 diabetes and a four-month feeding schedule. (A) A hyperglycemic mouse model was established by pretreating mice with nicotinamide (NA) and streptozotocin (STZ) followed by glucose loading for 4 months. (B)Three behavioral pharmacological tests were performed after the 4-month glucose tolerance test: spontaneous locomotor activity, rotarod test, and 8-way maze test.
Figure 10.
A protocol for creating a mouse model of type 2 diabetes and a four-month feeding schedule. (A) A hyperglycemic mouse model was established by pretreating mice with nicotinamide (NA) and streptozotocin (STZ) followed by glucose loading for 4 months. (B)Three behavioral pharmacological tests were performed after the 4-month glucose tolerance test: spontaneous locomotor activity, rotarod test, and 8-way maze test.
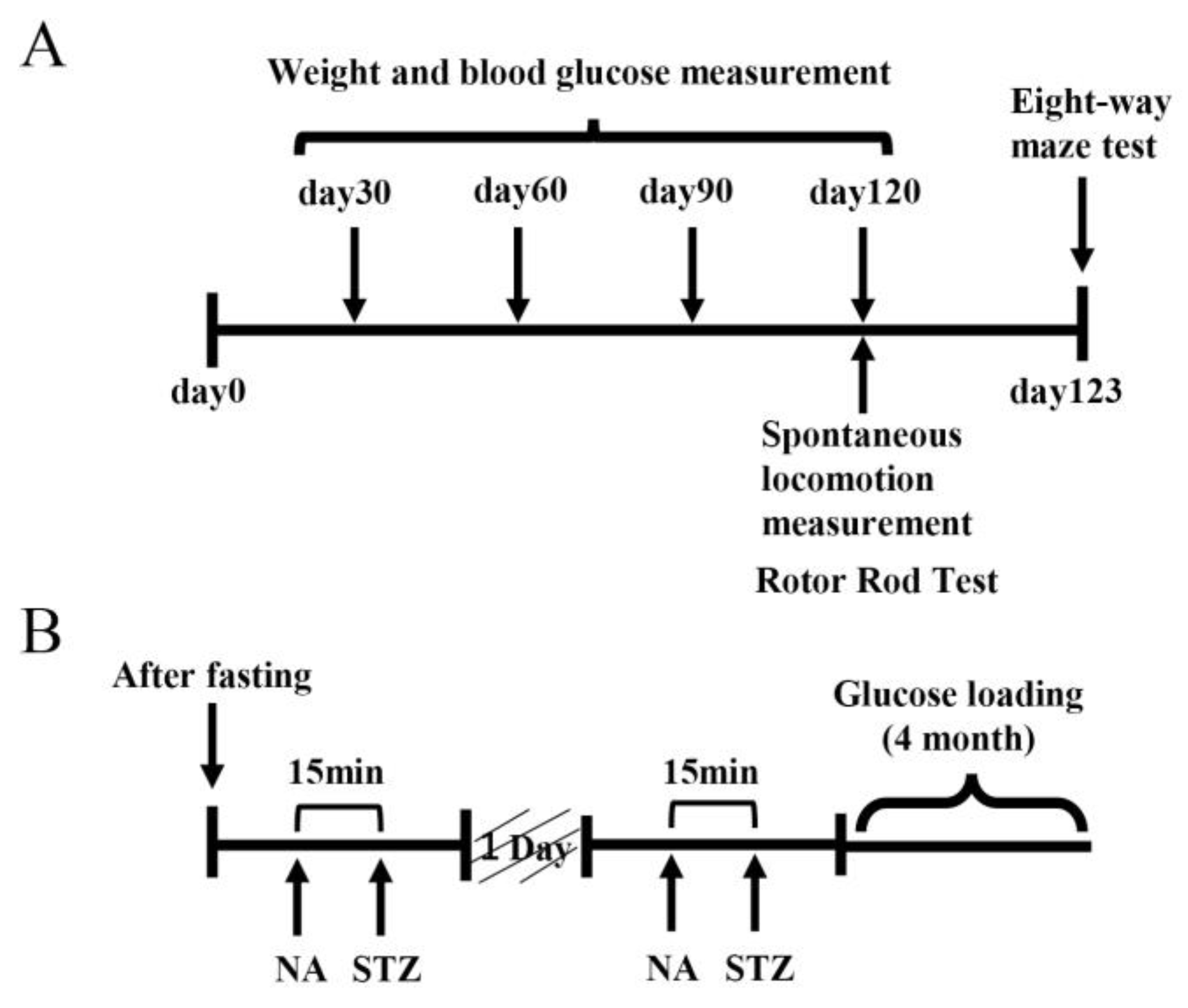

Table 1.
Effect of rhamnan sulfate (RS) intake on TNF-α expression level score (n=35).
| TNF-α | 0 | +1 | +2 | +3 | p value |
| A Control | 6 | 0 | 0 | 0 | |
| B MD | 0 | 0 | 1 | 5 | 0.001 a * |
| C MD + RS100 | 0 | 3 | 3 | 0 | 0.005 b * |
| D MD + RS300 | 0 | 6 | 0 | 0 | 0.001 b * |
| E MD + RS1000 | 1 | 5 | 0 | 0 | 0.001 b * |
|
a vs. Control b vs. MD |
1 The score of TNF-α expression level for each group was obtained by comparing the effects of RS on Iba1 expression level shown in Figure 6. The TNF-α expression levels are indicated as 0 (no staining), +1 (weakly positive), +2 (moderately positive), and +3 (strongly positive). Each value is indicated as mean ± S.D. (*p < 0.01).
Table 2.
Effect of rhamnan sulfate (RS) intake on Iba1 expression level score (n=35).
| lba1 | +1 | +2 | +3 | p value |
| A Control | 6 | 0 | 0 | |
| B MD | 0 | 1 | 5 | 0.003 a * |
| C MD + RS100 | 6 | 0 | 0 | 0.002 b * |
| D MD + RS300 | 5 | 0 | 0 | 0.004 b * |
| E MD + RS1000 | 5 | 1 | 0 | 0.007 b * |
|
a vs. Control b vs. MD |
1 The Iba1 expression score for each group was obtained by comparing the effects of RS on Iba1 expression levels, as shown in Figure 7. Iba1 expression levels were indicated as +1 (weak staining), +2 (moderate staining), and +3 (strong staining). Each value is indicated as the mean ± S.D. (*p < 0.01).
Table 3.
Effect of rhamnan sulfate(RS)intake on AGEs accumulation level score (n=35).
| AGE | 0 | +1 | +2 | p value |
| A Control | 4 | 2 | 0 | |
| B MD | 0 | 4 | 2 | 0.036 a * |
| C MD + RS100 | 3 | 3 | 0 | 0.076 b |
| D MD + RS300 | 1 | 3 | 1 | 0.497 b |
| E MD + RS1000 | 2 | 3 | 1 | 0.289 b |
|
a vs. Control b vs. MD |
1 The AGE accumulation level score for each group was obtained by comparing the effects of RS on AGE accumulation levels, as shown in Figure 8. AGEs accumulation levels were indicated as 0 (no staining), +1 (weakly positive), and +2 (strongly positive). Each value is expressed as the mean ± SD (*p < 0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



