
Submitted:
20 February 2025
Posted:
21 February 2025
You are already at the latest version
Abstract
Two of the most studied species: monophage Trioza magnisetoza and narrow oligophage Altica ballasogloi are offered for biological control of Russian Olive (Elaeagnus angustifolia) in the USA. The data obtained over 15 years and presented in this publication on biological and ecological features, host-plants, damage to the host plants, and suggested approaches for establishing new populations of both species are sufficient for their use in biological control in North America. Currently, 72 species of insects from 58 genera, 33 families, and six orders are known in Central Asia as pests of Russian Olive. The most numerous are insects from the order Coleoptera (36 species, 29 genera, 10 families), Homoptera (14 species, eight genera, six families), and Lepidoptera (13 species from 12 genera, 10 families). The most suitable for biological control of Russian Olive from this list are 24 species, including 8 species of monophages and 16 species of oligophages. In addition, other six species of insect pests with unexplored food specialization but associated with Russian Olive might also be used.
Keywords:
Russian Olive
; Kazakhstan
; biological control
; Trioza magnisetoza
; Altica balassogloi
1. Introduction
The genus Elaeagnus (common names: - silver berry, Russian Olive (oleaster), Persian olive in English; - Lokh in Russian, - Zhide, Zhiyde, Dzhida or Dzhigida in Kazakh, Uzbek, Kyrgyz and other Turkic local languages) consists of about 80 species native to Southern Europe, Western, Central and Southeast Asia, as well as North America; the center of origin of the genus is considered to be Southeast Asia [1]. Currently, there is an information on the growth of 3 species of Elaeagnus in Kazakhstan [2]: 2 indigenous E. oxycarpa Schlecht. and E. angustifolia L. and E. orientalis L. introduced probably from Transcaucasia and Turkey. All three species are widely cultivated by the local population as hedges, for fixing sand, riverbanks and canals, as well as for landscaping arid areas and creating shelterbelts. One of these 3 species, Russian Olive E. angustifolia has a very wide range, it is known from the countries of the European Mediterranean, Western, Central and Eastern Europe, Western Siberia, Asia Minor, the Caucasus, Iran, Northern India, Central Asia, Western China (Xinjiang) and Mongolia. In addition, Russian Olive was introduced into the North America. Currently, it has become the subject of biological control in the United States. The 6th volume of “Flora of Kazakhstan”, published in 1963, provides information on the growth in Kazakhstan of only 2 species of Elaeagnus: E. oxycarpa и E. turcomanica. Moreover, in the essay on E. oxycarpa the Russian Olive (E. angustifolia) is mentioned only as a cultivated plant, known mainly in Europe. In this regard, apparently, it is necessary to conduct thorough studies on the systematics of all species of Elaeagnus observed in Kazakhstan, including molecular studies of Russian Olive and other species biotypes from Kazakhstan and the United States. This applies, first of all, to biotypes of closely related species of E. angustifolia and E. oxycarpa.
The aim of this publication is to present the results of our research on insect species that damage Russian Olive in the natural areas of its growth in Kazakhstan and other parts of Central Asia. Our work in Russian Olive biocontrol studies began in 2006 with two research goals: (1) to find effective biological control agents of Russian Olive in Kazakhstan and (2) to study their biological features under natural conditions. Conducted research shows that Altica balassogloi (Coleoptera, Chrysomelidae) and Trioza magnisetosa (Homoptera, Psylloidea) are the most preferable potential Russian Olive biocontrol agents for introduction into the USA. The analyzed information and original observations and experiments on distribution, habitats, host-plants, biological and phenological characteristics, parasites, effectiveness for biological control and testing in natural conditions for both species are presented in this publication
The first publications on insect species that damage Russian Olive in its native habitat of Central Asia appeared in the middle of the 20th century. The reference book “Harmful animals of Central Asia” [3] (1949) lists 20 species of insects living on Elaeagnus. The biological and environmental features of cerambycid, Chlorophorus varius - which damages Elaeagnus in Western Kazakhstan, have been studied fairly well [4,5,6]. Eulecanium (Parthenolecanium) persicae and Diaspidiotus elaeagni (Diaspididae), Xylotrechus grumi (asellus), X. namanganensis, Turanium scabrum (Cerambycidae) and others [7,8] were also found as pests of Russian Olive, which grows in protective forest belts of Uzbekistan and Southern Kazakhstan. Around the same time, in Naryn Sands in Western Kazakhstan [9] and in South and Southeast Kazakhstan [10,11,12,13,14,15] other pests of Russian Olive were also found: Trioza magnisetosa (Triozidae), Capitophorus hippophaes (Aphididae), Lepidosaphes turanica (Diaspididae), Tropinota (Epicometis) hirta (Scarabaeidae), Molorchus kiesenwetteri, Tetrops elaeagni, Chlorophorus elaeagni (Cerambycidae), Scolytus jaroschewskii (Scolytidae), Argyroploce lutozana (Torticidae), Ananarsia elaeagnella (Gelechiidae) and others. Quite complete reviews of the pests of the Elaeagnus were given by Mityaev I.D. [14] for Kazakhstan and Sinadsky Yu.V. [16] for Central Asia. Based on these and some other publications [10, 16–46] we have compiled a preliminary list of 72 species of insects that damage Russian Olive and live in the Central Asian region. Brief characteristics of these species are given below in a separate section.
2. Materials and Methods
The research material is the results of observations, collections and testing, which took place in the field seasons of 2006 - 2013 in the Southern Balkhash region (the valleys of the Ili, Karatal, Lepsy rivers), as well as in Almaty on the grounds of the Institute of Zoology of the Republic of Kazakhstan and the adjacent area of the Main Botanical Garden. In subsequent years (2013 - 2022), only phenological observations on Russian Olive psyllid Triosa magnisetosa and cerambycid Altica balassogloi were carried out and the state of populations of both species was assessed at previously selected monitoring sites. From mid-June 2006 to 2013, special exploratory studies were conducted in the Ili Valley and the lower reaches of the Lepsy River in order to detect modern habitats of Russian Olive psyllid, Triosa magnisetosa, and the cerambycid, Altica balassogloi. In 2007, for the same purposes, the Karatal River valley and the lower reaches of the Ili River were examined, as well as monitoring observations of the state of already discovered populations in the Ili River Valley were carried out. In 2008 and subsequent years, observations continued at monitoring sites of the Ili and Karatal river valleys in order to study the biological and phenological characteristics of these insects, and from 2008 to 2014, insects were tested on experimental Elaeagnus trees in the vicinity of Almaty and in the territories adjacent to the Institute of Zoology RK. In total, six Altica balassogloi populations and one large Trioza magnisetosa population were selected for research, the location of them is shown in Figure 1. During the studies, classical entomological techniques were used for searching, collecting, fixing, observing and testing insects for host plant specialization [47]. To test both species to assess their nutrition on other plants, a certain number of eggs, larvae or adult insects were planted on other potential host plants in natural habitats. In addition, in 2008-2012, we worked out a methodology for establishing new colonies of both species by releasing adult insects on other Russian Olive plants in the vicinity of Almaty.
This research on various noxious weeds from central Asia (often considered native there) and their natural enemies was collected in the field to determine host range, then tested in cages overseas and then shipped to labs in the U.S. for final testing.
3. Results
As a result of our research in Kazakhstan, we suggest 2 species as the preferential potential Russian Olive biocontrol agents for introduction into U.S. First, Altica balassogloi (Coleoptera, Chrysomelidae), which damages foliage and shoot and is distributed in south and southeastern Kazakhstan (Arys, Ili, Karatal, Charyn rivers riparian forests). Second, Trioza magnisetosa (Homoptera, Triozidae), which usually damages foliage on young trees and is distributed in south, central and west Kazakhstan. It should be noted that over the past 15 years, in the territory of Southeast Kazakhstan, general warming of the climate and uneven seasonal precipitation have been observed. Winters were not snowy, and early springs, sometimes with April frosts, leading to the death of buds, especially on young plants. For example, in 2007, a sharp warming occurred already in the third decade of March with heavy rainfall until mid-June and with hot, arid summers and autumn. The years 2008 and 2015 turned out to be especially arid, when drought almost throughout the entire growing season covered not only deserts and piedmont steppes, but also the lower zones of the mountains, and the burnout of the vegetation reached its maximum in July and early August. Such unusual weather conditions determined phenological shifts in the development of insects not only by year, but even among different populations during one growing season.
3.1. Trioza Magnisetosa Loginova, 1964
Oleaster jumping plant-lice or Russian Olive psyllid is one of 23 species of the genus Trioza known in Kazakhstan (there are more than 300 species in the world).
3.1.1. Distribution
Species is known in Romania; Ukraine [48]; Russia: Central and East-European area [49,50], South-European area (environs of Baskunchak Lake, Astrakhan region) [51]; Georgia [48,52] and Armenia [49,51]; Turkey [53,54,55]; Iran [57,58]; Kyrgyzstan; Uzbekistan, Tajikistan [51]; China: Shaanxi, Xinjiang, Gansu, Hebei [58]. In Kazakhstan, these jumping plant-lice are widely distributed in the southern half of West Kazakhstan as well as in Central and South Kazakhstan [14,15,23,59,60], some small populations were found also in the southern portion of East Kazakhstan [61] and in the vicinity of Almaty City. At first time in the South Balkhash Lake region, we discovered some large populations of these jumping plant-lice in June 2006 in the environs of Zhelturanga Town and Topar Town in the lower reaches of the Ili River.
3.1.2. Habitats
This species usually inhabits the lower reaches of the rivers at an altitude of 400-500 m above sea level, although in Tajikistan sometimes it rises along river channels to 2500 m above sea level [51]. It prefers to settle on Russian Olive, which grows in dry areas of the floodplain or in sandy, desert and dry steppe floodplain terraces. An indication of the occurrence of Trioza magnisetoza on the needles of Picea shrenkiana in the mid-mountain belt of the Ketmen ridge (Southeastern Kazakhstan) at an altitude of 1920-2220 m above sea level [61] is erroneous, since earlier we observed some phenomena of wind introducing flying adults (during swarming) into the mountains on various random plants. In the Southern Balkhash region, the populations we discovered lived in the lower reaches of the Ili River in a grove dominated by Russian Olive and Turanga (Populus diversifolia) inside a small inter-dune basin, where psyllids were found in large numbers on young and middle-aged trees of Russian Olive (near the Zhelturanga Town) and in the grove along the edges sand dunes (surroundings Topar Town).
3.1.3. Host-Plants
Trioza magnisetosa is a monophagous species on Elaeagnus angustifolia and therefore appears to be one of the most promising biological agents of Russian Olive in US. The results of our many years of field observations and repeated attempts in 2008-2012 to transplant insects to other plant species in natural habitats for feeding showed a strict connection between this species with Russian Olive as a host plant. The host range tests in 2012-2013 carried out in laboratory conditions of the Institute of Zoology of the Republic of Kazakhstan also confirmed the monophagy of this psyllid species. Information about feeding of this psyllid on Rhamnus L. (Rhamnaceae) [62], apparently refers to the introduction of insects on alien random plants and requires clarification.
3.1.4. Biological and Phenological Features
Wintering. Winged adults of oleaster jumping plant-lice overwinter in large clusters in the litter under leaves of Elaeagnus angustifolia, a small part of these insects is located in the upper layers of the soil. Often some concentrations of overwintering psyllids are observed near trunks.
Moving from the wintering places and copulation. Moving the winged adults from the wintering places in conditions of South Balkhash Lake region occurs in the beginning or the end of April – beginning of May, this depends on the weather conditions of spring and the presence of developed buds on the host plant. In more southern regions, moving from wintering places was observed in early/late March – early April [14] and it is associated with the appearance of buds on the Russian Olive. Immediately after leaving the wintering sites, the psyllids move to the branches of oleaster and begin to feed first on the buds, and then on the leaves and young shoots. The females opened at this time had no eggs at all in the oviducts. Only after a period of additional nutrition, that is necessary for the accumulation of fat reserves and maturation of eggs, the imagoes begin to copulate, and then females begin to lay eggs. Additional nutrition occurs first on the buds (the end of April – May), and then on young shoots and leaves (the end of May – June).
Egg-laying. Females begin laying their eggs in the mid-May and through the end of mid-June, the bulk of egg laying occurs in late May – early June. The eggs are laid by jumping plant-lice on the underside of the leaf. Eggs are about 0.40–0.45 mm long, smooth and oval, but pointed at one end and blunt on the other end. The pointed half of the egg becomes transparent 4–6 days after laying and faces the upper side of the leaf. On the contrary, the blunted half of the egg is covered with radiating leaf scales. Egg clutches are usually single, rarely double. The number of eggs laid on a single leaf varies greatly and depends on the abundance of females on the tree. In the South Balkhash Lake region, 20 to 30 eggs were found on one leaf, although up to 40 eggs were observed on some individual leaves. At the same time, in more Northern populations with a small number of insects, clutches of 2–3 or 6–11 eggs are often laid on the leaves, depending on the population density [14]. The fertility of a single female can reach 450 eggs as it also was shown by other researchers [23]. At the place where the egg is laid, small bumps appear on the upper side of the leaf, which can easily be used to calculate the population of the leaves with eggs. The embryonic period lasts 14 to 17 days.
The development cycle then passes through 5 larval instars, which have a light gray with a silvery tinge that is not distinguishable in color from the leaves of Russian Olive. Wing covers and complex facet eyes appear already in first instar larvae and persist in all other instars; the larvae simply grow in size from age to age, so often the larval stages are referred to as nymphs of the 1st–5th age. Emerging of first instar larvae from eggs was observed in the third decade of May. First, they stick to the lower side of the leaf, and then migrate to the upper surface, spreading along the central and lateral veins of the leaf, or sometimes stick to the leaf petioles. With a high population density, there were an average of about 60 larvae per leaf. With a low population, the larvae are concentrated in groups of 6–9 individuals, and with mass reproduction, the psyllid larvae cover the entire leaf with a continuous layer on both sides.
Emerging and flight of the first-generation adults in the southern Balkhash region are observed at the end of the second decade of June. After emerging, adults almost immediately start feeding on leaves and copulating. During mass flying and with a large number of insects in the population, adults sometimes form flying clusters (swarms) that can be carried a considerable distance by the wind. Apparently, this is how the population of jumping plant-lice came to settle on uninfested Russian Olive trees. Oviposition occurs from the end of the second decade to the beginning of the third decade of June until about the middle of July. The eggs are laid singly or in groups on the lower side of the leaf. After an embryonic period lasting about 2 weeks, the first instar larvae emerge; these larvae first start feeding on the lower side of the leaves and then also move to the upper side of the leaf and on the leaf petioles. The larvae grow and increase in size, passing through 5 instars.
Emerging and swarming of second-generation imagoes is observed in the southern Balkhash region in late July – early August. After additional feeding, the adult psyllids begin to copulate and then the females start laying eggs on the leaves. The full cycle of development of the second generation ends by mid-September, when the third-generation imagoes appear, adults begin to feed sluggishly and when the temperature drops, they gather in groups on the surface and in the upper layers of the soil for subsequent overwintering.
During the year, 3 generations of psyllids usually develop in the southern Balkhash region, as well as in the more southern regions of Central Asia [51]. Although in 2008, due to severe drought and in 2009, due to relatively cold weather conditions in the summer, we noted the development of only two generations of insects here (see Table 1).
3.1.5. Parasites
Tetrastichus sp. (Chalcidoidea, Eulophidae) and mites of the family Trombidiidae are parasitic on the larvae of psyllids.
3.1.6. Efficiency for Biological Control
Larvae of Russian Olive psyllid (especially larvae of the last instars) cause the formation of chlorotic spots, bending and twisting of leaves. Such leaves turn yellow before time and fall off. Adults do not cause any externally noticeable harm, even with a significant population. The greatest harm occurs during mass reproduction of psyllids, when the nymphs cover not only the leaves with a continuous layer, but also their petioles and even the tips of young shoots. In this case, most Russian Olive trees lose more than 50% of their foliage, and some lose it completely, which has been repeatedly described in the literature [9,14,23,63]. We observed such mass reproduction of the Russian Olive psyllid’s population only in the southern Balkhash Lake region only once in 2006. Unfortunately, in the following years, during our observations in 2007–2018, we did not find mass population outbreaks, although changes of the normal development of Russian Olive in these years were noted. Partial or complete loss of leaves in mid-summer and intense poisoning by saliva released by nymphs when sucking, disrupts the normal development of trees, leads to severe oppression and slowing of growth rate. All this, together with other adverse factors (progressive salinization, excessive waterlogging of the soil, or conversely, a lack of ground moisture) usually leads to poor overwintering of plants, even to their death in the winter or spring periods. Thus, the Russian Olive psyllid, without causing any noticeable harm to the plant in small populations, turns out to be an effective control agent over the course of years in places of high population.
3.1.7. Testing in Nature
To develop methods of establishing new colonies of Russian Olive psyllids, 5 tests were conducted in natural areas of Kazakhstan. Tests were made in 2007 and 2008, none of which established.
Test 1 (Mid–June 2007). More than a hundred adult individuals from the vicinity of Topar town were tested on a medium-aged Russian Olive bush in the vicinity of Almaty in mid-June 2007 (Failed to establish).
Test 2 (Mid–June 2007). 300 adult individuals were released on a young and relatively small Russian Olive tree in the vicinity of the Avat Town (50 km East of Almaty) on the bank of an irrigation channel. However, for unclear reasons, no psyllids survived in either location (Test 1 and 2).
Test 3 (2008). A new attempt was made to plant large groups (more than 300 individuals in each group) of Russian Olive psyllids from the Topar population on two Russian Olive trees (young and middle-aged) in the vicinity of Avat Town, but it also failed.
Test 4 (2009). Attempts were made to plant more than 300 psyllids of the Topar population on a healthy and young oleaster tree in the vicinity of Avat Town which was initially successful.
Test 5 (2010). 300 more psyllids were released on a medium-aged tree in the Ili river floodplain in the vicinity of the Bakanas City were initially successful.
In tests 4 and 5, where newly emerged adults were collected in one location and transferred to young, healthy Russian Olive trees in a different location, adults were feeding and copulating and showed signs of successful establishment. However, in 2014 these populations disappeared due to a fire in the floodplain of the Ili river and construction work in the vicinity of the Avat Town.
Unfortunately, our monitoring studies in 2019 showed that both oleaster jumping plant-lice populations found in 2006 in the lower reaches of the Ili river also disappeared as a result of anthropogenic influence: in 2018, the population died in the vicinity of Zhelturanga and in 2019 in the vicinity of Topar towns. The main reasons for the death of populations are strong overgrazing and artificial fires, which are arranged by local residents. Overgrazing completely destroyed the layer of fallen leaves and mixed it with sand so that the psyllids ‘ wintering sites were completely destroyed, so that adult psyllids had no place to winter. Currently, to continue research, it is necessary to find new large populations of Russian olive psyllids both in the southern Balkhash region and in southern Kazakhstan.
In general, our experiments have shown the possibility of establishing artificial populations of psyllids on new and healthy young Russian Olives by transferring feeding and copulating jumping plant-lices to lay eggs on a new plant with the main threats to population establishment and maintenance being anthropogenic disruption of the site.
3.2. Altica Balassogloi (Jakobson, 1892) Lepidoptera, Chrysomelidae (=Haltica suvorovi Ogl., H. lopatini Pal.)
The Russian Olive (oleaster) leaf beetle is a representative of a large genus consisting of about 250 species and having a worldwide distribution. There are 17 known species in Kazakhstan and Central Asia.
3.2.1. Distribution
South and South-East Kazakhstan, Uzbekistan, Kyrgyzstan, Tajikistan [29].
3.2.2. Habitats
It is a floodplain forest species throughout its range. In the southern Balkhash region, this species lives in floodplain forests of the Ili, Charyn, Karatal and Lepsy rivers and sometimes enters the mountain floodplain forest at an altitude of 2000-2500 m above sea level.
3.2.3. Host-Plants
It is the oligophage of Elaeagnus spp. and sea-buckthorn (Hippophae rhamnoides), most commonly found on oleasters growing along rivers.
3.2.4. Biological and Phenological Features
There are 2 generations per year (see Table 2).
Adults of the second generation are over-wintering stage, beetles are located singly or in groups under the peeling bark of Russian Olive, as well as in deep cracks in the bark and in the litter. There have been cases of beetles wintering under the bark of old large-stemmed willow trees (Salix). In the spring, with the onset of warm days, the beetles leave the wintering areas and are located on the trunk and branches of Russian Olive, usually in mid-April or early May, although there are cases of both early, at the end of the first decade of April (Kulenova, 1968), and very late appear on new buds and young leaves, and with a decrease in temperature in the evening, they go to the litter under the oleaster or hide under the bark and in its cracks.
Egg laying in normal weather conditions in South-Eastern Kazakhstan begins at the end of April. Eggs are laid in groups on the leaves (usually on the underside), and on the petioles and young branches. The ovipositor usually has from 3 to 30 eggs, although it is not uncommon to come across leaves and twigs completely covered with egg masses (Figure 2a).
Larvae of the first instar usually appear in the second decade of May. The newly emerged larva has a dark yellow color, which darkens to a grayish-black color within half an hour and the larvae begin to feed actively.
The development cycle of all three larval instars together takes about one month. By the end of the first decade of June, the third-instar larvae stop feeding, fall to the ground and burrow under fallen leaves and in the litter near the roots of oleaster trees; after 2-3 days, these larvae turn into pupae.
First-generation adults usually emerge in large numbers at the beginning of the third decade of June. Emerged beetles immediately begin additional feeding and later copulation. Females begin laying eggs in early July, first-instar larvae appear from the second decade of July, and second-instar larvae are observed in the second half of July. Emerged larvae are first found on the lower surface of the leaves and finely skeletonize them (Figure 2b). Later, they crawl to the upper side of leaves and skeletonize them more closely, sometimes gnawing through. Pupation of third-instar larvae occurs in the second decade of August also in the litter or in the upper layer of soil.
Beetles of the second wintering generation appear from mid-August, at which time they begin to migrate to the litter or under the bark of the oleaster for wintering. In a cold spring with frosts and a cool beginning of summer, there are usually noticeable shifts in the timing of the development of the phases of the life cycle in the overwintered generation of leaf beetles. But, despite this, by mid-summer, phenological shifts usually stop and the second generation appears in the second half of August.
3.2.5. Monitoring of Modern Populations
During route surveys of floodplains of the Ili, Karatal and Lepsy rivers in 2006, we found several populations of oleaster leaf beetles, from which 6 populations were selected for monitoring observations in 2006-2018:
1) Ili population (40 km North of Kapchagai, Russian Olive thickets in the floodplain of the Ili river) were found on 13 June 2006. The population size for all the time of observations was very low, every year in the second decade of June there were only about 15–20 beetles and 25–30 larvae of 1–2 ages. Near the place where the leaf beetle was found, a fairly large young grove grows on the site of a previous fire, but it was not possible to find a single individual in it.
2) Tasmurun population (70 km North of Kapchagai, near Miyaly village, Russian Olive trees grew between the beginning of the Tasmurun canal and the Ili river) was found in 14 June 2006. Unburned bushes were still preserved in this part of the riparian forest, where a small number (27 individuals) of beetles, 2 oviposits, and several larvae of 1–3 ages were found. The number of insects of this population also remained very low in 2006-2018.
3) Topar population (10–12 km South-West of the village of Topar, in the lower reaches of the Ili river, a small oleaster forest along a moist roadside depression on the edge of sand dunes), was discovered on 16 June 2006, in mid-June, 2nd–3rd instar larvae were already predominant here, and egg-laying and larvae of the 1st instar were rare. Despite the relatively high number of insects (there were an average of about 100 individuals per tree), this population did not cause much damage to its host plants in 2006. But observations in 2007 showed that beetles and larvae of the overwintering generation had severely damaged individual trees growing along roadside ditches and on some young bushes, all the leaves were completely eaten. Compared to 2006, the number of leaf beetles in 2007 increased to an average of 120 individuals per tree. In 2008, the number of oleaster leaf beetle became even higher and reached an average of 160 individuals per 1 tree, although the number of leaf beetles in the adjacent barkhan (sand dune) area did not change and remained at the level of 2006 – 100 individuals per tree. The damage to host plants in 2008 was noticeably greater compared to the previous year: devoid of leaves, which caused the drying and death of individual branches.
The increase in the number of beetles in 2008, in our opinion, was due to the early and hot spring, which also caused an earlier appearance of overwintered beetles and, accordingly, a phenological shift in the development of this (overwintered) leaf beetle generation. In 2008, beetles after wintering came to the crown of trees in the first decade of April. In late April and early may of the same year, copulation and egg laying took place, and larvae developed in May – the first decade of June. At the beginning of June, massive leaf damage was already observed on young plants, and on old trees, such damage was noted only in the lower part of the crown. At the end of May – beginning of June, there were no more beetles and ovipositors. the larvae of the 3rd age found on the branches were ready for pupation. Only pupae were found in the litter under the crown of trees, they were located among the plant remains or on the border with the soil at a depth of 1-2 cm; the pupae do not form cradles and lie freely in different poses, mostly on their sides. The first emergent adults in nature were found in 4 June. In artificial cages of 34 pupae collected for observation, all adults were born within 4 days (from 4 to 7 June, 2008). In natural conditions, on trees growing in a wet depression along a roadside ditch, mass concentrations of beetles on tree crowns were observed as early as June 10. They actively fed; preferring shaded areas, avoiding direct sunlight, and constantly moving around the crown in choosing the most favorable leaves for food.
The mass emerging of first-generation beetles in 2008 was observed in the middle – early third decade of June, copulating beetles – in the third decade of June, females started oviposition at the end of June, mass oviposition was noted in early July. The first-instar larvae appeared at the end of the first decade of July, and the second instar larvae appeared in the second half of July. The first third instar larvae were recorded at the end of July, and their pupation in the tree litter began at the end of the first decade of August. Adults of the second generation of leaf beetles appeared in mid-August, at this time there were some cases of their additional feeding with leaves and gradual departure for wintering in the litter and in the cracks of the bark of host trees.
Hereinafter the population density decreased in 2009–2014 and remained at the same level – 120 individuals per tree, and in 2015–2018, the number of insects in this population gradually decreased to an average of 95-100 individuals per tree, accordingly, the damage to host trees has also decreased.
4) Karatal population 1 (15 km North of Ushtobe City, a grove of sparse old trees) was found at first time in 15 May 2007. At this time, in the lower part of the crown of one of the Russian Olive trees, on the upper side of the leaf, only 5 larvae of the 2nd age were found, 1 larva that had just transformed to the 3rd age, 1 ovipositor and 1 female. This is all that could be found in this fairly extensive grove. The appearance of beetles, the laying of eggs and the development of larvae here took place in the early spring cycle.
5) Karatal population 2 (22 km North of the Ushtobe City, a vast stretch of floodplain forest consisting of old and medium-aged Russian Olive trees mixed with willows) was discovered in 15 May 2007. This leaf beetle population was represented by small scattered spots, but the number of leaf beetles in individual spot was very high. In mid-May 2007, egg-laying, larvae of 1–3 instars (mostly 1–2), males and egg-laying females were recorded here. The leaves were usually damaged mainly in the lower half of the crown. Most of the eggs were laid on the leaves; and in one ovipositor the number of eggs located on the upper side of the leaf was usually less (5–15 eggs) than in the ovipositor on the lower side of the leaves (30–43 eggs). Large oviposits were also found on young branches, in the forks of branches and on the petioles of leaves. Such egg-laying was sometimes observed in the 1950s in powerful centers of mass reproduction of the species on overpopulated bushes. An average of 5–10 adults and 15-20 oviposits were usually observed on the same branch length 1.5 m. In this population, emerging larvae of the 1st age of the wintering generation were usually observed in the third decade of May. In general, this leaf beetle population developed almost a month earlier than the Topar population over the entire period of observations in 2007–2018. This is due to the peculiarities of the microclimate in this place.
6) Karatal population 3 (55 km North of the Ushtobe City, moistened depression in the floodplain on the border with sand dunes along the drainage channel) was found at first time in 15 May 2007, when this population lived only on a few low young bushes, the number of leaf beetles here was quite high about 430 beetles per plant. At this time, only copulating beetles were observed, and no egg-laying was found. The appearance of beetles from wintering sites on the oleaster crown here occurred 10–15 days later than in the Karatal population 2, despite the fact that they are located relatively close to each other (23 km). Here, apparently, local features of the microclimate and wintering places of this population also affected. Unfortunately, observations in 2008 showed that this population has completely disappeared – no damage or beetles were found on the bushes. The main causes of population death are drying up of the coastal lowlands, fire and livestock that completely destroyed the litter under the Russian Olive bushes.
Further searches for the leaf beetle in 2009–2018 in the Ili and Karatal valleys did not yield positive results. The main reason for the decline in the number of oleaster leaf beetle in the southern Balkhash region is frequent fires. For example, repeated surveys of areas of the Russian Olive forest in North of Bakanas that were densely populated by leaf beetles showed that even 25 years after the fire, the oleaster leaf beetle was not able to populate the apparently safe forest.
In general, in the last few decades, the floodplain forests of the southern Balkhash region have been severely degraded due to negative anthropogenic impacts (fires, land withdrawal for agriculture, construction, etc.). Full-fledged floodplain forests have been preserved only in a few protected natural areas.
3.2.6. Efficiency for Biological Control
Since the mid-20th century, this species has been known as a dangerous pest of Russian Olive [10,14,16,18,19,20,63,64]. The harm caused by the leaf beetle to Russian Olive is especially evident during the mass reproduction of the beetle. Damaging trees for 2–3 years, the leaf beetle causes their severe oppression and subsequent drying. Weakened trees that grow in less favorable conditions are particularly affected. Cases of mass reproduction of the leaf beetle during our observations in the southern Balkhash region in 2006–2018 were not observed. Although in the second half of the 20th century, such cases were not uncommon in South-Eastern Kazakhstan. For example, in 1965, in the floodplain of the Charyn river in the Sarytogay forest (now the Charyn state national natural park), the oleaster leaf beetle caused huge damage to the Russian Olive population in the time of mass reproduction. At this time, 432 leaves were damaged on a single branch of 600 leaves. At the end of summer, trees on large areas were completely stripped [64]. In the mid-20th century, there was a mass reproduction of the leaf beetle in the lower reaches of the Karatal river, which caused great damage to the populations of Russian Olive; the area covered by the mass reproduction of the leaf beetle was about 10 hectares. Eating the leaves of trees by first-generation larvae reached 60% of the entire crown, and some trees lost their leaves completely[14]. Numerous mass breeding of beetles was also observed in southern Kazakhstan in the riparian forests of the Syr Darya river basin [10,16,18,19,20,62].
3.2.7. Testing in Nature Control
Altica balassogloi was tested on Elaeagnus angustifolia in the vicinity of the Avat Town (floodplain of the Besagach river, 50 km East of Almaty) to work out the method of establishing new beetle colonies (photo). On the afternoon of June 17, 2006, 55 first generation beetles were planted on a young tree, beetles were collected in the vicinity of the Topar Town in the lower reaches of the Ili river basin. The tested group of beetles did not establish; after 5 days, we did not find a single beetle on this or on the neighboring bushes.
The next attempt was made on the evening of May 16, 2007 in the same place with beetles from the second overwintered generation, selected from the Karatal population 3. As there was a very strong North-easterly wind at this time, we placed 76 beetles on the leeward side of the lower part of the crown. The beetles spread along the lower branches and began to eat the leaves; they managed to establish and start laying eggs on June 1. Only 3 oviposits were found on the upper side of the leaves on the branch on which they were planted. In the future, the entire subsequent life of this population was associated only with this branch; no attempt was observed in distribution of beetles to attack adjacent neighboring or remote branches. On the morning of June 10, one ovipositor female, a pair of copulating beetles, one ovipositor, nine larvae of the first instar, and many leaves gnawed by beetles were recorded on this branch. In the evening of the same day, three larvae on one of the leaves disappeared; they may have been eaten by a ladybird larva crawling here. On July 4, the beetles were no longer found, but there were many damaged leaves; at this time, there were small numbers of third instar larvae feeding singly or in pairs on the upper side of the leaves. On July 10, all the third instar larvae disappeared, and on July 25, only one beetle was found. Since July 31, no beetles, ovipositors, or larvae have been found, despite our search for the leaf beetle in August and October. The population may have completely disappeared again or migrated elsewhere.
In 2008, oleaster leaf beetles from the Topar population were planted on a medium-aged Russian Olive tree in the vicinity of the Institute of Zoology in Almaty. For testing on experimental bushes in Almaty, 224 beetles were taken, which were transplanted to the lower branches on June 12, 2008. In the future, the transferred beetles were kept only on one tree and did not fly away. Immediately after landing on the lower branches, the beetles started feeding on the leaves. In the third decade of June, they laid eggs; in late June – early July first larvae were born, they strongly ate the lower part of the crown. Some of the leaves were almost completely eaten. According to observations on July 11, only six third instar large and two second instar larvae remained on the crown, actively feeding on the leaves, although numerous exuvia of first, second and third instar larvae were found on the branches. At this time, most of the larvae migrated to the litter and have already started pupating. In September, several second-generation beetles that went to winter were found under the bark of the trunk and in the litter under the tree crown. Unfortunately, in the spring of 2009, overwintered beetles were not found.
In our opinion, the low survival rate of transplanted beetles is primarily due to the food and geographical conservatism of the transplanted population (the beetles did not fly away and remained only on the original branches), caused, apparently, by the small number of “colonists”, as well as the large influence of predators attacking both adults (birds) and leaf-eating larvae (predatory bugs, ladybug larvae, photo). For example, when beetles were released in the vicinity of the Institute of Zoology, there was a noticeable concentration of insectivorous birds (mainly, Indian Myna Acridotheres tristis) on the surrounding trees for an hour. Unfortunately, given the low number of leaf beetles in 2006-2022, we have not yet been able to transplant a large number of beetles (2-3 or more thousand) without causing damage to the natural population.
3.3. Preliminary Annotated List of Insect Species Damaging to Russian Olive in Central Asia
Order HOMOPTERA
Suborder Cicadinea
Family Cicadellidae
Macropsis elaeagni Emeljanov, 1964 (= M. unicolor). Monophagous insect of Elaeagnus angustifolia; floodplain forest’s species that settled in Kazakhstan with Russian Olive plantings [14,15,34]. Emerging first instar larvae occurs in the second half of April – early May and coincides with the appearance and budding of the oleaster. The larvae suck on the buds, then on the leaves and tops of shoots and young twigs. Imagoes are found from June to August, and individuals are found until October. The species is distributed in southern Russia, Kazakhstan, Uzbekistan, Tajikistan, and Kyrgyzstan.
Macropsis elaeagnicola Dubovsky, 1966. Monophagous insect of Elaeagnus angustifolia. The species lives in the mountains, settlements, and woodlands of Russian Olive. Larvae begin emerging in May, and adults appear in June and July. The larvae suck the leaves and tops of the shoots. The species is widely distributed in Northern Kazakhstan along with oleaster plantings. It is known from Uzbekistan, Kyrgyzstan, and southern and Northern Kazakhstan [34].
Suborder Psyllinea
Family Triozidae
Trioza magnisetosa Loginova, 1964 (see chapter above).
Trioza elaeagni Scott, 1880 (= furcata). Monophagous species e of Elaeagnus angustifolia, floodplain forest’s species. It is also found on oleaster landings in settlements in Western, Central and southern Kazakhstan [14,37]. Winter adults, two generations per year. Second generation adults were recorded in October, leaving for the winter – in early November. It is known from Uzbekistan, Tajikistan and the Caucasus region, and is found in Central Asia together with T. magnisetosa.
Suborder Aphidinea
Family Aphididae
Capitophorus archangelskii Nevsky, 1928. Monophagous insect of Elaeagnus angustifolia, non-migratory species. Overwinter eggs in the forks of branches, bases of buds, petioles of last year’s leaves, in the crevices of the bark on the trunk and branches. Emerging of the first larvae is observed in early April, and their mass appearance is observed in mid-April. They feed on the buds and then on the tops of young shoots. The damage to Russian Olive trees is significant: the leaves turn yellow and fall off, the shoots bend and slow down in growth, with strong infestation, the tree loses up to half of the entire foliage. It is distributed in Western, Central and South-Eastern Kazakhstan.
Capitophorus hippophaes (Walker, 1852) (=gillettei). Oligophagous of Russian Olive and Hippophae rhamnoides. The eggs overwinter in the buds ‘ sinuses and branch forks. With a large population, it causes the fall of leaves of Russian Olive and Hippophae rhamnoides. The species is known from Northern, Central and southern Kazakhstan.
Capitophorus elaeagni (Del Guercio, 1894). Oligophagous of Russian Olive and Hippophae rhamnoides, it lives on the underside of leaves, optionally migrates on Cirsium incanum in summer, and lives in willow-oleaster floodplain forests. Known all over the world.
Aphis craccivora Koch, 1854. A broad polyphagous insect that damages numerous cultivated and wild plants from the family Ranunculaceae, Rosaceae, Fabaceae, Caryophyllaceae, Chenopodiaceae, Malvaceae, etc. It is observed also on oleaster, aphids settle in numerous colonies in the lower part of the crown at the ends of young shoots, which later bend and wither. It is distributed in Europe, the Anterior and Central Asia, southern Siberia and the Far East of Russia, as well as North and South America, and Africa. It is the most harmful in Central Asia, Azerbaijan and southern Kazakhstan [14,15 – information about Aphis medicaginis and A. fabae belong to Aphis craccivora].
Suborder Coccinea
Family Monophlebidae
Drosicha turkestanica Archangelskaya, 1931. A wide polyphagus, the main host plants are willow, poplar, and oleaster. One generation develops per year. Eggs and first instar larvae overwinter under the bark of trees, often near the ground surface or on the roots of plants. In early spring, the larvae come out of hiding places and spread over various plants. In late March – April, emerging second instar larvae appear, they begin to move along tree trunks, feeding on leaves. In mid-to late May, after emerging, the third instar larvae begin to gather in colonies on twigs, bark cracks, or exposed areas. At the end of June – the first half of July, imagos are born, and mating occurs. Females after fertilization are driven into the cracks of the bark on the trunk near the surface of the earth, during oviposition they are covered with a white wax fluff. Within females, Cryptochetum turanicum (Diptera, Cryptochaetidae) flies develop as an endoparasite. The larvae and adults of the coccid are also eaten by the larvae of the ladybeetle Rodolia fisheri (Coleoptera, Coccinellidae), which are similar in body shape and color to the victim.
Family Coccidae
Parthenolecanium corni (Bouche, 1844). A wide polyphage, feeding also on Russian Olive. The species is known worldwide.
Parthenolecanium persicae (Fabricius, 1776). A wide polyphage, it feeds also on Russian Olive. The species is known from Europe, Transcaucasia and Central Asia.
Family Diaspididae
Lepidosaphes turanica Archangelskaya, 1937. Monophagous species of Elaeagnus angustifolia. It lives on branches and trunk, very harmful, causing drying of both branches and entire trees. It was described for the first time from Uzbekistan, it is now also known from Iran, Kazakhstan, and Western China.
Lepidosaphes ulmi (Linnaeus, 1758). It is a broad polyphagous species, a serious pest of Apple trees, is also noted on oleaster. Causes drying of Russian Olive branches. It is widely distributed around the world.
Diaspidiotus elaeagni (Borchsenius, 1939). The oligophagous species, it lives on Elaeagnus angustifolia and Hippophae rhamnoides. The species lives in floodplain forests and floodplains of mountain rivers, develops two generations per year, and second instar overwinter. Egg laying occurs in May and August, and the fertility of one female is up to 40 eggs. It is known from Turkey, Armenia, Georgia, Iran, Central Asia and China.
Order HETEROPTERA
Family Miridae
Orthotylus elaeagni Jakovlev, 1881. Monophagous insect of Elaeagnus angustifolia, when mass reproduction causes the death of shoots. In the year 2-4 generations develop, eggs overwinter, larvae appear in early May. It is known from Western, Southern and South-Eastern Kazakhstan.
Deraeocoris pilipes (Reuter, 1879). Polyphagous species living on oleaster (Elaeagnus spp.), willows (Salix spp.) and elm (Ulmus spp.). It is known from Eastern and South-Eastern Kazakhstan, as well as Uzbekistan and Tajikistan.
Glaucopterum kareli Wagner, 1963. A narrow oligophagous species, it feeds on Elaeagnus angustifolia and E. orientalis. The species is known from Turkey, Kazakhstan, Turkmenistan, Tajikistan, and Mongolia [65,66].
Family Pentatomidae
Brachynema germarii (Kolenati, 1846). Polyphagous insect living on the oleaster. It is widely distributed from the Canary Islands and the entire Mediterranean through Transcaucasia, the deserts of the Near and Middle East, Central and Southern Kazakhstan, Central Asia to Mongolia and Northern China [67].
Family Lygaeidae
Spilostethus pandurus Scopoli, 1763 (= Lygaeus pandurus). Polyphagous species, but prefers plants of Lamiaceae, especially fallen seeds of many plants and juice of green parts. In Kazakhstan it usually lives on Calligonum, Atraphaxis and sometimes Elaeagnus angustifolia. One generation per year, adults overwinter. This species is widespread worldwide, as it is found in South Africa, southern Europe, Kazakhstan and China.
Order COLEOPTERA
Family Scolytidae
Scolytus jaroschewskii Schevyrew, 1893. It is known as a pest of Russian Olive in the floodplain forests of Central Asia. Larvae overwinter, pupae in May, and adults in late May. The species inhabits the trunks and branches on the north side and leads to weakened and dried out trees.
Scolytus schevyrewi Semenov Tjan-Shansky, 1902. Wide polyphagous species, it lives mainly on a variety of native elm species, as well as occasionally on willows (Salix spp.), apricot, cherry, and peach (Prunus spp.), and Russian olive. Despite its Asian native distribution (Russia, Kazakhstan, Kyrgyzstan, Tajikistan, Turkmenistan, and Uzbekistan, China, Mongolia and Korea) this species appeared in USA in 2003 as a pest in Colorado and Utah. Later, it has since been found to occur in 10 western and central states. This insect attacks elms stressed by drought. Infested trees are killed by such attack.
Family Bostrichidae
Enneadesmus scopini Fursov, 1936. Usually inhabits weakened oleaster trees, in Tajikistan it is also known on Gleditsia triacanthos (Fabacea). One generation develops per year. The larvae overwinter at the end of passages gnawed through the wood; in late March or early April, the larvae awaken and continue to sharpen the passages. In the first decade of May, the larvae finish feeding and prepare chambers 1.5–2 mm wide and 5 mm long for pupation. The pupal stage lasts 10–15 days, the beetles emerge from mid-May to early June. In June, the first larvae appear, and the larvae feed until the beginning of winter. Developing on the branches, this species circles them with its moves, causing the branches above the settlement to shrink. The species is known from the floodplain forests of Central Asia (Amu Darya basin) and Kazakhstan (Ili, Karatal and Syr Darya river basins).
Lyctus turkestanicus Lesne, 1935. This species develops on thin (up to 5 mm in diameter) shrinking shoots or branches of oleaster and inhabits the floodplain forests of Uzbekistan, Turkmenistan, and Western China.
Family Scarabaeidae
Tropinota hirta (Poda von Neuhaus, 1761). Beetles eat the flowers of many fruit trees, after blossomimg of fruit trees the imagoes usually move to the oleaster flowers. Beetles damage mainly unopened flowers of Russian Olive. The range of the species extends from Central and southern Europe to Central Kazakhstan.
Tropinota turanica (Reitter, 1889.). It feeds on the flowers of many plants. In southern Kazakhstan, it is listed as an oleaster pest [15]. The species is known from Central Asia, South-Eastern Kazakhstan, and Western China.
Oxythyrea cinctella (Schaum, 1841). Among other flowering plants, this scarab also damages the flowers of Russian Olive, eating their ovary. The species is known from Portugal, Spain, southern Italy, the Balkan Peninsula, the Caucasus and Transcaucasia, Iran, Afghanistan, Central Asia, southern, South-Eastern and Eastern Kazakhstan, and Western China.
Adoretus nigrifrons (Steven, 1809). It damages the flowers of Russian Olive and other plants and inhabits floodplain forests, plains and foothills. Adults fly from April to mid-September. It is known from the Caucasus, Transcaucasia, Central Asia and Kazakhstan.
Protaetia marginicollis (Ballion, 1870). It feeds on the flowers of Russian Olive and other flowering plants and lives in floodplain forests. It is distributed in the Eastern part of Central Asia, in Kazakhstan – the upper reaches of the Ili river and the Syrdarya valley.
Family Cerambycidae
Aeolesthes sarta (Solsky, 1871). Polyphagous species. The preferred hosts of this insect are: Ulmus minor, U. pumila, Populus alba, P. diversifolia, P. euphratica, P. talassica, P. canadensis, Salix acmophylla, S. songarica, S. turanica, Platanus orientalis, P. hispanica, Malus domestica and Juglans regia. In general, it may damage species of Elaeagnus, Acer, Betula, Fraxinus, Gleditsia, Juglans, Malus, Morus, Platanus, Populus, Prunus, Pyrus, Quercus, Robinia, Salix, Ulmus, and other hardwood and fruit trees. This species is an important pest of many ornamental and deciduous fruit trees [25,26,27,68]. It attacks both stressed and healthy trees of different ages. Successive generations remain on the same tree for several consecutive years, eventually causing its death. Sometimes, young larvae encircle a tree feeding on the cambium, which leads to the rapid death of the tree. Young trees with thin bark are most susceptible to the beetle and 1–3 larvae may be enough to kill a tree [69]. It is distributed in south Kyrgyzstan, Uzbekistan, Tajikistan, Turkmenistan, Afghanistan, India (Western Himalayas), Iran and north Pakistan.
Anoplistes halodendri halodendri (Pallas, 1776) (=Asias halodendri). Polyphagous species. It can feed on Russian Olive. Distribution: Bulgaria; Ukraine; Albania; Romania, Kazakhstan; Siberia and European Russia.
Anoplophora glabripennis (Motchulsky, 1853). Polyphagous species, but usually prefers Acer, Populus, Salix, Morus, Malus, Prunus, sometimes on Elaeagnus. Adults feed on the bark and leaves of trees, preferring to attack young shoots. They overwinter in the egg, larval, or pupal stages. It originates from China, but in recent years it has become widely distributed and found in Central Asia, Europe and North America.
Chlorophorus elaeagni Plavilstshikov, 1956. Polyphagous species, it lives on tamarisk, oleaster and Halimodendron halodendron. The larvae live in the trunks and thick branches of weakened Russian Olive trees, causing the death of the plant. Mass emerging of imagoes happens from mid-June. The species is known from Russia (Dagestan), Western and South-Eastern Kazakhstan (floodplain forests of the Ili and Karatal rivers).
Chlorophorus faldermanni (Faldermann, 1837). Polyphagous species, it lives on young trees of Russian Olive, Halimodendron halodendron (Syr Darya river floodplain) and poplar (Northern Iran). The appearance of the imago begins in mid-May and continues until the end of September. The larvae grind holes up to 10-30 cm in the wood and cause great damage to Russian Olive trees. This species is distributed in Uzbekistan, South Kazakhstan, Afghanistan and China.
Molorchus kiesenwetteri (Mulsant et Rey, 1861). It is associated with Russian Olive, inhabiting thin branches with a diameter of 0.3–1.5 cm. It causes drying of Russian Olive branches. Species is known from Uzbekistan, Turkmenistan and Western Kazakhstan.
Tetrops elaeagni Plavilstshikov, 1954. Larvae lives only on Elaeagnus spp. The species is known from Kazakhstan, Uzbekistan, Turkmenistan and also South of European Russia.
Tetrops formosus Baeckmann, 1903. The larvae live mainly in weakened and shrinking Russian Olive trees, as well as on Apple trees (Kyrgyzstan). The passages are drilled under the bark in the upper layer of wood. Pupae appear in early May, and the imago emerges in the second half of May. The species is quite rare and is known from Kazakhstan, Kyrgyzstan, and Western China.
Turanium scabrum (Kraatz, 1882). This species lives on Russian Olive, although it is also recorded on apple trees [14]. Larvae of the last and pre-last instar overwinter at the end of passages located in the wood of trunks and branches. Pupation in South-Eastern Kazakhstan occurs in late April-early May, appearance of adults begins in the middle of the first decade of May; swarming of adults occur in the second half of May-early June. This species is very harmful and is known from Uzbekistan and Kazakhstan, as well as from Russia (Orenburg).
Xylotrechus asellus (Thieme, 1881) (= grummi). It lives on the Russian Olive. Larvae overwinter, pupae are formed in late May and mid-June, imago flights begin in the first half of June and continue until the end of June. Larvae populate the trunks of usually old or severely weakened trees, they cause very severe damage, leading to the death of the tree. The species shows a very large conservatism in choosing a tree, if the larvae have populated the tree, they will live on it until it is completely destroyed. It inhabits floodplain forests throughout Central Asia, southern and South-Eastern Kazakhstan.
Xylotrechus namanganensis Heyden, 1885. Wide polyphagous species, it lives on Juglans, Prunus, Malus, Morus, Crataegus, Elaeagnus, Populus, Ulmus, Celtis, Salix, Betula, Alnus, Platanus, as well as other deciduous trees in Kyrgyzstan, Kazakhstan, Tajikistan, Turkmenistan, Uzbekistan, Afghanistan and China.
Family Buprestidae
Anthaxia elaeagni (Richter, 1945). (=Cratomerus elaeagni). It lives only on Elaeagnus spp., inhabits the branches and trunks of weakened trees in the floodplain forests of Central Asia. Larvae overwinter, in early May; larvae of the last age and pupae are recorded, imago in May-June. It causes significant damage to Russian Olive.
Capnodis miliaris (Klug, 1829). Polyphagous species, it is an important pest of poplar, willow and occasionally lives on oleaster. The adults emerge between the end of May and August, and feed on leaves and young shoot, then overwinter in the soil or under the dried leaves. One generation develops in two years. It is known from Turkmenistan, Tajikistan, as well as Israel, Turkey, Northern Iran, and Armenia.
Family Bruchidae
Spermophagus sericeus (Geoffroy, 1785). Beetles during flowering of many plants feed on their flowers, gnawing out generative organs. It is also found on Russian Olive in southern Kazakhstan [14]. It is known from Europe, Kazakhstan, Siberia and Mongolia.
Family Curculionidae
Rhyncolus culinaris Germar, 1824. It lives in the wood of many coniferous and deciduous trees, as well as on oleaster in the floodplain forests of the Amu Darya. Larvae overwinter, feeding begins in early spring. It causes significant damage to decayed wood, as well as to live Russian Olive trees [16]. It is known from many parts of Eurasia.
Platymycterus trapezicollis Ballion, 1878. Polyphagous species. Beetles are very harmful to foliage of Russian Olive, poplars (especially Populus diversifolia), Halimodendron halodendron, Robinia pseudoacacia, Alhagi spp. When feeding, the beetles gnaw out the edges of the leaf blade or skeletonize the leaves. Adults appear by mid-May, and are observed in mass in June-July. It is known from Central Asia.
Polydrusus obliquatus Faust, 1884 (= ferghanensis). Polyphagous species. In Kazakhstan, it is found on oleaster, willow, peach, apple and pear. They are very harmful, beetles in May gnaw the bark of young shoots, leaf petioles and the edges of the leaf blade, sometimes trees lose up to half of their foliage [15]. It is known from Central Asia.
Polydrusus pilifer Hochhuth, 1847. Polyphagous species, often found on strawberries (Fragaria sp.). On oleaster in southern Kazakhstan [15], beetles eat the edges and petioles of leaves, buds, and tops of young shoots. It is known from Central Asia.
Chloebius immeritus (Schoenherr, 1826). Polyphagous species, beetles damage leaves and flower buds mainly on Russian Olive and licorice, in floodplain forests this species prefers to feed on oleaster. Adults appear in late April, and copulation occurs in the first half of May. It is known from Central Asia and Kazakhstan.
Chlorophanus caudatus Fahraeus, 1840. Polyphagous species, in floodplain forests, it damages the leaves of oleaster, willow, turanga, and tamarisk. Adults appear in early May in the ground in June. It is known from Central Asia and the Caucasus.
Megamecus cinctus (Faust, 1887). It is observed on Elaeagnus spp., including Russian Olive as a harmful species, and known from Central Asia.
Sitona macularius (Marsham, 1802). Polyphagous species. It is known as a pest of lentils and almonds, and has also been noted on Russian Olive (Schaffner et al., 2007). Widely distributed in the Palearctic, the species is also known in the United States.
Prisistus caucasicus (Kirsch, 1878). It is found Elaeagnus [40], and known from Russia (Moscow and Ryazan regions, Caucasus), Georgia, Armenia, Azerbaijan, Kazakhstan, Uzbekistan, Turkmenistan, Tajikistan, Iran and Afghanistan.
Family Rhynchitidae
Temnocerus elaeagni (Korotyaev et Legalov, 2006.) Oligophagous species, it lives on Elaeagnus and found also on Salix. Species is described from Central and South-East Kazakhstan [71].
Family Chrysomelidae
Altica balassogloi (Jakobson, 1892) (see chapter above)
Altica deserticola (Weise, 1889). Polyphagous species, it feeds on the leaves of Russian Olive, willow, tamarisk and Halimodendron halodendron. In the floodplain forests of Central Asia, the leaf beetle mainly damages the foliage of Russian olive, as a result of which the trees lose their growth, shrink and do not bear fruit [16]. There are 2 generations per year. It is known from Central Asia.
Family Meloidae
Epicauta erythrocephala (Pallas, 1776). It is a widespread polyphagous species, a pest of potatoes, beets, and other crops. In the floodplain forests of Central Asia, it damages foliage on Russian Olive, ash, Ammodendron sp., Zygophyllum sp. and other plants.
Order HYMENOPTERA
Family Formicidae
Tetramorium caespitum (Linnaues, 1758). In southern Kazakhstan, this species of ant was noted as a pest of young oleaster shoots in the lower part of the crown in May-July. Ants in groups of up to several dozen individuals chewed or nibbled the tops of young shoots along with the leaves, causing the tops of the shoots to wither [15]. The species is known in Northern Eurasia.
Family Megachilidae
Megachile dohrandti Morrawitz, 1880. In the floodplain forests of Central Asia, this leaf-cutter bee cuts out pieces of leaves and uses them to build its nests in the old passages of the cerambycid beetle Xylotrechus asellus. In the floodplain forests of the Amu Darya river, more than one-third of the leaves on individual trees are affected by this bee [16]. It is known from Central Asia, Kazakhstan and southern China.
Order LEPIDOPTERA
Family Torticidae
Apotomis lutosana (Kennel, 1901). This oligophagous species, that lives on various species of Elaeagnus, is noted as a pest on European olive plantations Olea europaea [71]. Larvae damage oleaster trees, feeding between spun leaves of oleaster and skeletonizing them. Parasitoid is Nemorilla maculosa Meig. (Tachinidae) [72]. It is distributed in Romania, Trans-Caucasus, South Russia (Caspian area), Kazakhstan, Turkmenistan, Uzbekistan, Tajikistan [73].
Pandemis chondrillana (Herrich-Schäffer, 1860). Wide polyphagous species, its larvae prefer apple, pear, apricot, plum, almond, peach and cherry, quince, poplar, willow, as well as rose, pomegranate, cotton, pistachio, lilac, and oak. It was recorded also on Elaeagnus. There are 2 to 3 generations yearly in Southern Kazakhstan, Uzbekistan, and Tajikistan. Second and third instar larvae (sometimes also fourth instars) hibernate in dense silken cocoons. In wintertime they are usually in bark cracks on the root neck or under leaf remains webbed to bark, as well as on the trunks and branches. The larvae start feeding in April in the beginning of blossoming, they make shelters by webbing two to four leaves together, or by rolling the leaf around its main vein. The pupae of the hibernated generation appear in the first decade of May, the mass appearance – in mid of May, this stage lasts 8-15 days. Adults emerged in mid of May – beginning of June and fly until the late June and again from mid-August until mid-September. There are two known parasitoids: Sympiesis albiventris Storozheva (Eulophidae) and Actia pamirica Richter (Tachinidae). Most damage is caused by the second and third instar larvae. It is distributed in Southern and Eastern Europe, Russia (from European part to South West Siberia), Asia Minor, Iran, Pakistan, Kazakhstan, Mongolia, North-Western China [74].
Archips rosana (Linnaeus, 1758). This widely polyphagous species occurs on apple, pear, plum, hop, raspberry, hawthorn, privet as well as on Abies, Acer, Alnus, Betula, Cistus, Corylus, Cydonia, Fraxinus, Hippophae, Laurus, Lonicera, Medicago, Morus, Myrtus, Plantanus, Populus, Prunus, Quercus, Rhododendron, Rosa, Salix, Syringa, Tamarix, Tilia, Ulmus, Urtica, Vaccinium and Viburnum. In Kazakhstan it is observed also on Elaeagnus angustifolia: larvae in April-May, pupae in the second half of May, adults from the beginning of June. There is only one generation yearly. It damages leaves, flowers, and fruitlets; it is distributed and abundant throughout Europe, Asia Minor, Trans-Caucasus, Kazakhstan, and North America.
Family Gelechiidae
Ananarsia eleagnella (Kuznetsov, 1957). It is a widely polyphagous species associated with Elaeagnus spp. and Hippophae spp., it was noted also feeding of moth larvae on Crataegus oxyacantha and Malus domestica in experimental conditions [39,42]. It feeds on leaves and develops in stems, buds and fruits. In Kazakhstan, young caterpillars appear in the second half of May on shoots of Russian Olive (this year) between newly unfolded leaves of apical buds. They glue several leaves together with a web and, having settled between them, scrape off the epidermis or chew through a hole in it. Grown caterpillars bite into the apical parts of young shoots and make heart-shaped moves in them. Affected shoots at first do not stand out among healthy shoots, but later their ends shrink and bend hook-like, sometimes bursting along the course. This species can cause serious damage to the growth of Ruissian Olive, affecting up to 50–70% of all shoots in large trees and almost 100% in young bushes, as it was observed in Central Kazakhstan [14]. The species is known from Central Asia and Kazakhstan.
Ananarsia lineatella Zeller, 1839. A polyphagous species, but it lives mainly on Prunus spinosa, Malus spp., Armeniaca spp., Persica spp., Cerasus spp., Amygdalus spp., Acer tataricum, where it damages leaves, shoots and fruits. Widely distributed in Central and South Europe, Russia (European part), Caucasus; Transcaucasia, Central Asia, China, North Africa, Asia Minor, Nearest East, Iran, Afghanistan, India, Australia and North America [32].
Family Geometridae
Lycia hirtarius (Clerck, 1759). Wide polyphagous species, feeding on birch, oak, poplar, willow, blackthorn, raspberry, rose and oleaster. Pupae overwinter in the soil. The caterpillars appear in June and are observed until October, and the imago flies from late May to August. They eat the leaves of oleaster but the damage is insignificant. Widespread species in the Palearctic: Europe, Asia Minor, Central Asia, Kazakhstan, and the far East [75].
Family Noctuidae
Acronicta rumicis (Linnaeus, 1758). A wide polyphagous species, caterpillar in southern Kazakhstan that eats the leaves of Russian Olive, the harm to oleaster is insignificant. It is a widespread species in the Palearctic.
Family Cossidae
Cossus cossus (Linnaeus, 1758). A broad polyphagous species, it lives on ash, willow, many fruit and other deciduous trees. In the floodplain forests of Central Asia and Kazakhstan, this species is common on willow, but also lives on Russian Olive. The species is distributed throughout Europe, the Caucasus, Central Asia and Kazakhstan, Siberia and the Far East of Russia, as well as in Western and Northern China.
Family Sphingidae
Hyles hippophaes (Esper, 1793). Oligophagous species, in Central Asia and Kazakhstan; subspecies of H. h. bienerti Stgr. It feeds on Russian Olive and sea buckthorn, and has been noted as a pest of Russian Olive leaves [16]. The caterpillars completely ate the leaves on some trees. There are two generations per year. Pupae winter, flights of adults of the overwintered generation occurs in May, after that females lay their eggs singly. Emerging caterpillars have developing period within 20–30 days, pupation takes place in the litter, and the mass flight of first-generation butterflies occurs in July. It is found in Afghanistan, Armenia, Azerbaijan, China, France, Georgia, Germany, Greece, Iran, Iraq, Kazakhstan, Kyrgyzstan, Mongolia, Pakistan, Romania, Serbia and Montenegro, Spain, Switzerland, Syria, Tajikistan, Turkey, Turkmenistan, and Uzbekistan.
Family Pyralidae
Euzophera alpherakyella Ragonot, 1887. It damages shoots of Elaeagnus in Tajikistan and Kazakhstan. Distribution: South Russia, Tajikistan, South East Kazakhstan and West China.
Family Nolidae
Meganola elaeagni (Shchetkin, 1980). A narrow oligophage, noted as an endemic pest of Elaeagnus orientalis and E. angustifolia in Tajikistan and Turkmenistan.
Family Lymantriidae
Lymantria dispar (Linnaeus, 1758). Wide polyphagous and quarantine species. Feeding on leaves of Quercus, fruit trees, Populus, Betula, Larix, Elaeagnus, etc. Adults of Asian populations are capable of flight, but adults of European populations cannot fly. Distributed in Europe (widely) and Mediterranean region; Russia (widespread); Baltic countries; Belarus; Moldova; Ukraine; Transcaucasus; Kazakhstan; Central Asia, India; Afghanistan; Iran; Iraq; China; Japan; Korea; Taiwan as well as introduced in Canada and USA.
Family Lasiocampidae
Malacosoma paralella Staudinger, 1887. Wide polyphagous and quarantine species. It damages leaves on Atraphaxis pyrifolia, Berberis integerrima, Cerasus verrucosa, Chaenomeles japonica, Cotoneaster acutiuscula, C. insignis, C. suavis, Crataegus hissarica, C. pontica, C. turkestanica, Cydonia oblonga, Fraxinus sogdiana, Hippophae rhamnoides, Juglans regia, Lonicera korolkowii, L. nummulariifolia, Malus domestica, M. sieversii, Myricaria bracteata, Populus alba, P. tremula, Prunus mahaleb, P. avium, P. armeniaca, P. bucharica, P. cerasus, P. divaricata, P. dulcis, P. padus var. pubescens, P. persica, Pyrus communis, Quercus boissieri, Q. macranthera, Q. robur subsp. robur, Ribes nigrum, R. rubrum, Rosa canina, R. corymbifera, R. kokanica, R. maracandica, Rubus idaeus, R. turkestanicus, Salix excelsa, S. tenuijulis, Sorbus persica, S. turkestanica and Ulmus sp. Important damage also occurs on Berberis, Chaenomeles, Cotoneaster, Crataegus, Cydonia, Malus, Prunus, Pyrus, Rosa, Salix and Sorbus species. Leaves of other plants are damaged occasionally. It is distributed in Turkey, Syria, Armenia, eastern Kazakhstan, Kyrgyzstan, Uzbekistan, Tajikistan, Turkmenistan and North Iran [76].
Order DIPTERA
Family Agromyzidae
Amauromyza elaeagni E.Rohdendorf-Holmanova, 1959. A common species for Central Asia and South-East Kazakhstan, the oleaster fly-miner develops in several generations per year. The appearance of the first mines on the leaves of Russian Olive was noted in the second half of May, and mass-in June. Mines of round or irregular shape from 0.5 to 2 cm in diameter are usually located on the edge of the leaf, on one leaf was observed from 1–2 to 15 mines. The larvae that have settled in the leaves feed continuously for 7–10 days, then chew through a hole in the epidermis and leave the mine, after which they burrow into the litter and pupate. Damage caused by the fly-miner leads to drying up to 10 % of the foliage.
Family Cecidomyiidae
Cantarinia elaeagniflorae Fedotova, 1988. Oligophagous species, according to Z. A. Fedotova [33] this species lives on Elaeagnus oxycarpa, E. angustifolia, forming flower galls on it. The species was described from South-Eastern Kazakhstan (Southern Balkhash Lake region).
4. Discussion
Currently, in Central Asia, 72 species of insects from 58 genera, 33 families, and six orders have been detected as pests of Russian Olive. The most numerous are insects from the order Coleoptera (36 species, 29 genera, 10 families), Homoptera (14 species, eight genera, six families), and Lepidoptera (13 species from 12 genera, 10 families). For the purposes of biological control of Russian Olive in the United States, 24 species from this species list are suitable:
Monophages (8 species): Cicadellidae: Macropsis elaeagni, M. elaeagnicola, Triozidae: Trioza magnisetosa, T. elaeagni, Aphididae: Capitophorus archangelskii, Diaspididae: Lepidosaphes turanica, Miridae: Orthotylus elaeagni, Cerambycidae: Molorchus kiesenwetteri.
Oligophages (16 species): Aphididae: Capitophorus hippophaes, Capitophorus elaeagni, Diaspididae: Diaspidiotus elaeagni, Miridae: Glaucopterum kareli, Bostrichidae: Lyctus turkestanicus, Cerambycidae: Tetrops elaeagni, Xylotrechus asellus, Buprestidae: Anthaxia elaeagni, Curculionidae: Megamecus cinctus, Prisistus caucasicus, Rhynchitidae: Temnocerus elaeagni, Chrysomelidae: Altica balassogloi, Torticidae: Apotomis lutosana, Sphingidae: Hyles hippophaes, Nolidae: Meganola elaeagni, Cecidomyiidae: Cantarinia elaeagniflorae.
In addition, it is possible to use another 6 species of insect pests that have been noted on Elaeagnus, but whose feeding specialization has not yet been clarified: Bostrichidae: Enneadesmus scopini, Cerambycidae: Tetrops formosus, Turanium scabrum, Formicidae: Tetramorium caespitum, Megachilidae: Megachile dohrandti, Pyralidae: Euzophera alpherakyella.
5. Conclusions
The most studied species of fauna of Kazakhstan for the purposes of biological control of Russian Olive in the United States are the monophagous Russian Olive psyllid Trioza magnisetoza and the narrow oligophagous species Russian Olive leaf beetle Altica ballasogloi. Information on the distribution, biological and environmental characteristics, feeding, and damage to the host plant is sufficient for the use of these species in biological control in North America. However, due to the degradation of the natural ecosystems of the southern Balkhash region due to anthropogenic influence, the state of the populations of both species has deteriorated in the last 30 years; some populations have disappeared, while others have declined in number. For this reason, it is necessary to continue the search for this species and work out methods for establishing new artificial populations both in nature and in the laboratory.
Author Contributions
Conceptualization and methodology, R.J. and C.J.D; investigation, R.J., and V.I.; writing—original draft preparation, R.J., and V.I.; writing—review and editing, C.J.D.; visualization, R.J. All authors have read and agreed to the published version of the manuscript.
Funding
This research was initiated by a research program for the biological control of weeds and was funded by the U.S. Department of Agriculture (USDA). Funding from this program was also distributed to the Agricultural Research Service (USDA – ARS), part of which specifically covered overseas research, along with additional funding primarily provided by the U.S. Bureau of Reclamation, managed by Dr. Fred Nibling. Overseas research was directed by Dr. Lloyd Andres, USDA-ARS Albany, CA. This research was partially funded by the Kazakhstan scientific-technical program BR21882199, “Cadastre of wild animals arid territories of Balkhash-Alakol basin with an assessment of threats for their conservation and sustainable use,” and by the Chinese Academy of Science President’s International Fellowship Initiative (PIFI) for 2019.
Data Availability Statement
Data will be made available on request.
Acknowledgments
The authors cordially thank Prof. Ivan Mitjaev (Institute of Zoology RK, Almaty), who shared his broad entomological research experience in Central Asia and took part in the field research on potential agents for Russian Olive biocontrol in 2006-2012. Unfortunately, Prof. Mitjaev passed away in January 2015 and could not continue his work together with the authors of this publication. The authors also acknowledge the technical support provided by Ms. Ekaterina Roschektayeva on the laboratory observation and collection of field material.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Baitenov, M.S. Flora Kazakhstana, rodovoy kompleks flory; Gylym: Almaty, Kazakhstan, 2001; Volume 2, pp. 1—280. (=Flora of Kazakhstan, generic complex of flora).
- Abdulina, S.A. Spisok sosydistykh rasteniy Kazakhstana; Editor Kamelin, R.V.; Gylym: Almaty, Kazakhstan, 1999; pp. 1—187. (List of vascular plants in Kazakhstan).
- Vrednye zhivotnye Sredney Azii, Editor Pavlovskiy, E.N.; USSR Academy of Sciences: Moscow—Leningrad, USSR, 1949; pp. 1—404. (=Harmful animals of Central Asia).
- Vorontsov, A.I. Vrediteli lesomeliorativnykh posadok zapadnogo Kazakhstana i Zavolzhiya. Itogi raboty VIZR za 1936 g.; VIZR: Leningrad, USSR, part 1, pp. 202—-205. (=Pests of forest landings in Western Kazakhstan and the Volga region. Results of the VIZR’s work for 1936).
- Grechkin, V.P. Ocherki po biologii vrediteley lesa; Moscow, USSR, 1951; pp. 75-90, 128-135. (=Essays on the biology of forest pests).
- Vorontsov, A.I.; Zakharchenko, I.S. Lokhoviy izmenchiviy usach i mery boriby s nim. In Sbornik rabot po lesozaschite; Moscow Forestry Institute: Moscow, USSR, 1957; Part 1, pp. 46–54. (=The grape wood borer (Chlorophorus varius) and measures to combat it).
- Makhnovskiy, I.K. Vrediteli zaschitnykh lesonasazhdeniy Sredney Azii i boriba s nimi; Tashkent, 1955; pp. 43—166. (=Pests of protective forest plantations in Central Asia and their control).
- Makhnovskiy, I.K. Vrediteli drevesno-kustarnikovoy rastitelnosti Chirchik-Angrenskogo gorno-lesnogo massiva i boriba s nimi. In Trudy of the Central Asian Institute of Forestry of the Uzbek Academy of Agricultural Sciences; Tashkent, 1959; Issue 5, pp. 13—56.
- (=Pests of wood and shrub vegetation of the Chirchik-Angren mountain forest massif and their control).
- Rafes, M.P. Vrednye nasekomye lokha, dzhuzguna i tamariska, proizrastayuschikh na Narynskikh peskakh polupustynnogo Zavolzhiya Entomologicheskoe Obozrenie 1956, 35(4), 805-817. (=Pest insects of oleaster, Calligonum and tamarisk, that grow on the Naryn Sands of the semi-desert Zavolzhye).
- Parfentiev, V.Ya. Lokhoviy listed Haltica suvorovi Ogl. (Coleoptera, Chrysomelidae) v tugainykh lesakh Kazakhstana. Entomologicheskoe Obozrenie 1957, 36(1), 96—97. (=Oleaster leaf beetle Haltica suvorovi Ogl. (Coleoptera, Chrysomelidae) in the tugai forests of Kazakhstan).
- Kuznetsov, A.V. Lokhovoya moli Anarsia eleagnella W. Kuzn. sp. n. (Lepidoptera, Gelechiidae) – novyi vreditel lokha v SSSR. Zoologicheskiy zhurnal 1957, 36(7), 1096—1098. (=Oleaster moth Anarsia eleagnella W. Kuzn. sp. n. (Lepidoptera, Gelechiidae) – new pest of oleaster in the USSR).
- Matesova, G.Ya., 1958. Заметки пo биoлoгии червецoв и щитoвoк (Hom. Coccoidea) Югo-Вoстoчнoгo Казахстана. In Trudy of Instituta Zoologii Akademii Nauk Kazakhskoy SSR; Alma-Ata,1958; Issue 8, pp. 130—137. (=Notes on the biology of mealybugs and scale insects (Homoptera, Coccoidea) of South-Eastern Kazakhstan).
- Kostin, I.A. Materialy po faune koroedov Kazakhstana (Coleoptera, Ipidae). In Trudy of Instituta Zoologii Akademii Nauk Kazakhskoy SSR; Alma-Ata,1960; Issue 11, pp. 129—136. (=Materials on bark beetle fauna in Kazakhstan (Coleoptera, Ipidae).
- Mityaev, I.D. K faune nasekomykh – vrediteley lokha v Kazakhstane. In Trudy of Instituta Zoologii Akademii Nauk Kazakhskoy SSR; Alma-Ata,1960; Issue 11, pp. 108—128. (=To the fauna of oleaster insect pests in Kazakhstan).
- Mityaev, I.D. Nasekomye, vredyaschie lokhu (Eleagnus angustifolia L.) v kulturnoy zone yuzhnykh oblastey Kazakhstana. In Trudy of Instituta Zoologii Akademii Nauk Kazakhskoy SSR; Alma-Ata,1962; Issue 18, pp. 61—68. (=Insects that harm Russian Olive (Elaeagnus angustifolia L.) in the cultural zone of the southern regions of Kazakhstan).
- Sinadskiy, Yu.V. Vrediteli tugainykh lesov Sredney Azii i mery boriby s nimi; Nauka: Moscow-Leningrad, USSR, 1963; pp. 1—150. (=Pests of Central Asian tugai forests and measures to control them).
- Skopin, N.G. Nasekomye – vrediteli lesoposadok v peskakh Bolishie Barsuki i puti boriby s nimi. Uchenye zapiski KazGU 1955, 27, 84—101. (=Insects - pests of forest plantations in the sand Big Barsuki and ways to fight them).
- Sinadskiy, Yu.V., 1958. Siniy listoed – Haltica deserticola Parf., vreditel lokha uzkolistnogo v tugayakh reki Syrdariya. In Nauchnye doklady vyshey shkoly. Lesoinzhenernoye delo; Nauka: Moskva-Leningrad, USSR, 1958; Issue 3, pp. 47—50. (=Blue leaf beetle – Haltica deserticola Parf., Russian Olive pest in the riparian forests of the Syr Darya river).
- Sinadskiy, Yu.V. Vrednaya entomofauna lokha (dzhidy) v tugainykh lesakh Sredney Azii i Kazakhstana. Zoologicheskiy zhurnal 1961, 40(7), 1019—1029. (=Harmful entomofauna of oleaster (dzhida) in the tugai forests of Central Asia and Kazakhstan).
- Sinadskiy, Yu.V. Dendrofilinye nasekomye pustyn’ Sredney Azii i Kazakhstana I mery boriby s nimi; Nauka: Moscow, USSR, 1968; pp. 1—126. (=Dendrophilous insects of the deserts of Central Asia and Kazakhstan and measures to combat them).
- Emeljanov, A.F., 1964. Noviy vid roda Macropsis Low. (Homoptera, Cicadellidae) s lokha. Doklady Academy of Sciences of Tajik SSR 1964, 7(1); pp. 47—48. (=New species of the genus Macropsis Low. (Homoptera, Cicadellidae) from the Elaeagnus).
- Pripisnova, M.G., 1965. Vrednaya entomofauna tugaynoi drevesnoo-kustarnikovoy rastitelnosti Yuzhnogo Tadzhikistana; Tajik University: Dushanbe, USSR, 1965; pp. 1—118. (=Harmful entomofauna of tugai tree and shrub vegetation of southern Tajikistan).
- Kuznetsov, A.V. Nasaekomye i kleschi, povrzhdayuschie plodovye i dekorativnye nasazhdeniya Severnogo Pribalkhashiya, i biologocheskie osnovy meropriyatiy po boribe s nimi. Abstract of the dissertation on competition of a scientific degree of candidate of agricultural sciences, Kazakh State University, Alma-Ata, 1971, pp. 1—22. (=Insects and mites that damage fruit and ornamental plantings of the Northern Balkhash region, and the biological basis of measures to combat them).
- Kostin, I.A. Zhiki-dendrofagi Kazakhstana (koroedy, drovoseki, zlatki); Nauka: Alma-Ata, USSR, pp. 1—286. (=Dendrophagous beetles of Kazakhstan (bark beetles, Longhorn, buprestids).
- Yagdyev, A. Stvolovye vrediteli turangi v Turkmenii. Izvestiya Akademii Nauk Turkmenskoi SSR, seria biologicheskie nauki 1975, 6; pp. 60—64. (=Trunk pests of Turanga in Turkmenia).
- Yagdyev, A. Obzor nasekomykh-ksilofagov lesov Centralnogo Kopetdaga. Entomologicheskoe Obozrenie 1979, 58(4), pp. 776–781. (=A review of the xylophagous insects of the forests of the Central Kopetdag).
- Yagdyev, A. Vrediteli dekorativnykh rasteniy v gorodakh Turkmenistana. Izvestiya Akademii Nauk Turkmenskoi SSR, seria biologicheskie nauki 1987, 1; pp. 47–50. (=Pests of ornamental plants in towns of Turkmenistan). (in Russian).
- Asanova, R.B.; Iskakov, B.V. Vrednye i poleznye poluzhestkokrylye (Heteroptera) Kazakhstann (opredelitel); Kainar: Alma-Ata, USSR, 1977; pp. 1—204. (=Harmful and useful hemipterans (Heteroptera) of Kazakhstan (key table).
- Lopatin, I.K.; Kulenova, K.Z. Zhuki-listoedy (Coleoptera, Chrysomelidae) Kazakhstana: opredelitel; Nauka: Alma-Ata, 1986; pp. 1—199. (=Leaf beetles (Coleoptera, Chrysomelidae) of Kazakhstan: key table).
- Danzig, E.M., 1993. Fauna of Russia and adjoining states, suborder coccids (Coccinea), families Phoenicoccidae and Diaspididae; Nauka: Sanct-Petersburg, 1993; pp. 1—453.
- Ponomarenko, M.G. Trophic relationships of caterpillars of subfamily Dichomeridinae (Lepidoptera, Gelechiidae) of the fauna of Russia and adjacent countries. In Lecturing for the Memory of A.I. Kurenzov, Nauka: Leningrad, Russia, 1993; Issue 4, pp. 41—48.
- Ponomarenko, M.G., 1997. Catalogue of the subfamily Dichomeridinae (Lepidoptera, Gelechiidae) of the Asia. Far Eastern Entomologist 1997, 50, 1—67.
- Fedotova, Z.A. Gallitsy-fitofagi (Diptera, Cecidomyiidae) pustyn i gor Kazakhstana: morphologiya, biologiya, rasprostranenie, filogeniya i sistematika; Samara State Agricultiral Academy: Samara, Russia, 2000; pp. 1-803. (=Midges – phytophages (Diptera, Cecidomyiidae) of deserts and mountains of Kazakhstan: morphology, biology, distribution, phylogeny and systematics).
- Mityaev I.D. Fauna, ecology, and zoogeography of cicadas (Homoptera, Cicadinea) in Kazakhstan. Tethys Entomological Research 2002, 5, 3—168.
- Aitzhanova, M.O.; Kadyrbekov, R.Kh., 2006. To the fauna of aphids (Homoptera, Aphididae) of the tugai forests of the Karatal river basin. Vestnik KazNU,seria biological 2006, 2(28), 82—87.
- Kadyrbekov, R.Kh.; Aitzhanova, M.O. To the fauna of aphids (Homoptera, Aphididae) of the tugai forests between the rivers Aksu and Lepsy. Izvestiya NAS RK, series biological and medical 2006, 3, 7—13.
- Jashenko, R.; Mityaev, I.; DeLoach C.J. 2007 Potential agents for Russian biocontrol in USA. In Proceedings of the XII International Symposium on Biological Control of Weeds, Le Grand Motte, France, 22-27 April 2007, pp. 249.
- Jashenko, R.V. Annotated list of species of the family Margarodidae (Homoptera, Coccinea) of Central Asia and Kazakhstan. Izvestiya NAS RK, series biological and medical 2007, 2, 3—10.
- Schaffner, U. Update on CABI‘s weed biocontrol program; CABI: Delémont, Switzerland, 2009; pp.1—27.
- Schaffner, U.; Hinz, H.L.; Cristofaro, M. Biological control of Russian olive, Elaeagnus angustifolia. Annual report 2007; CABI: Delémont, Switzerland, 2007; pp. 1—20.
- Schaffner, U.; Grözinger, F.; Cristofaro, M. Biological control of Russian olive, Elaeagnus angustifolia. Annual report 2007. CABI: Delémont, Switzerland, 2008; pp. 1—20.
- Schaffner, U.; Dingle, K.; Swart, C.; Cristofaro, M. Biological control of Russian olive, Elaeagnus angustifolia. Annual report 2011. CABI: Delémont, Switzerland, 2012; pp. 1—20.
- Schaffner, U.; Asadi G.; Chetverikov P.; Ghorbani R;, Khamraev A.; Petanović R.; Rajabov T.; Scott T.; Vidović B.; Cristofaro, M. Biological control of Russian olive, Elaeagnus angustifolia. Annual report 2013. CABI: Delémont, Switzerland, 2014; pp. 1—20.
- Bean, D.; Norton, A.; Jashenko, R.; Cristofaro, M.; Shaffner, U. Status of Russian olive biological control in North America. Ecological Restoration 2008, 26 (2), 105—107. [CrossRef]
- Weyl, P.; Schaffner, U.; Asadi, G.; Klötzli, J.; Vidović, B.; Petanović, R; Cristofaro, M. Biological control of Russian olive, Elaeagnus angustifolia. Annual Report 2016. CABI: Delémont, Switzerland, 2017, pp. 1—20.
- Weyl, P.; Closça, C.; Asad, G.; Vidović, B.; Petanović, R.; Marini, F.; Cristofaro, M. Biological control of Russian olive, Elaeagnus angustifolia. Annual Report 2018. CABI: Delémont, Switzerland, 2019, pp. 1—20.
- Fasulati, K.K. Polevoe izuchenie nazemnykh bespozvonochnykh; Vyshaya shkola: Moscow, USSR, 1961; pp. 1—304 (=Field study of terrestrial invertebrates); Higher School: Moscow, USSR, 1971; pp. 1—424.
- Loginova, M.M. New data on the fauna and biology of the Caucasian Psylloidea (Homoptera). In Trudy vsesoyuznogo entomologicheskogo obshchestva; Akademiya Nauk SSSR: Moscow, USSR, 1968, Issue 52; pp. 275—328.
- Loginova, M.M. Podotrjad Psyllinea. In: Opredelitel Nasekomykh Evropeiskoi Tchasti SSSR, part 1; Bei-Bienko, G. Ya.; Nauka: Moscow, Russia, 1964; pp. 437—482.
- Loginova M. M., 1978. New species of psyllids (Homoptera, Psylloidea). In Trudy Zoologicheskogo Instituta AN SSSR, Nauka: Leningrad, USSR, 1978; Issue 61, pp. 30—123.
- Baeva, V.G. 1985. Fauna of the Tajik SSR, Psyllids or jumping plant-lice (Homoptera, Psylloidea); Donish: Dushanbe, USSR, 1985; Issue 8, pp. 1—329.
- Gegechkori, F.M. Psyllids (Homoptera, Psуlloidea) of Caucasia; Metsniereba: Tbilisi, USSR, 1984; pp. 1—294.
- Klimaszewski, S. M.; Lodos, N. New informations about Jumping Plant Lice of Turkey (Homoptera: Psylloidea). Ege Üniversitesi Ziraat Fakültesi Dergisi [Turkish Journal of Agriculture and Forestry] 1977, 14(1), pp. 1—9.
- Burckhardt, D.; Önuçar, A. A review of Turkish jumping plant-lice (Homoptera, Psylloidea). Revue Suisse de Zoologie 1993, 100(3); pp. 547—574.
- Drohojowska, J.; Burckhardt, D. The jumping plant-lice (Hemiptera: Psylloidea) of Turkey: a checklist and new records. Turkish Journal of Zoology 2014, 38, pp. 1—10. [CrossRef]
- Burckhardt, D.; Lauterer, P. The jumping plant-lice of Iran (Homoptera, Psylloidea). Revue Suisse de Zoologie 1993, 100(4), pp. 829—898. [CrossRef]
- Ahmadi, Z.; Mehrvar, A.; Lotfalizadeh, H.; Gharekhani, Gh. Comparative study of the superfamily Psylloidea in Iran and East-Azarbaijan. Iranian Journal of Entomological Research 2013, 5(3), pp. 183—193.
- Li, Fasheng. Psyllidomorpha of China (Insecta: Hemiptera). Huayu Nature Book Trade Co.Ltd: Beijing, China, 2011; part 1-2, pp. 1—1976.
- Kuznetsov, A.V. Lokhovoya listobloshka Trioza magnisetosa Log. (Homoptera, Triozidae) v Severnom Pribalkhashiye. Trudy of Kazakh State Agricultural Institute 1970, 13(1), pp. 62—70. (=Oleaster jumping plant-lice Trioza magnisetosa Log. (Homoptera, Triozidae) in North Balkhash Lake area).
- Kuznetsov, A.V. Morfologiya lokhovoy listobloshki (Trioza magnisetosa Log.), povrezhdayuschey lokh uzkolistniy v Severnom Pribalkhashie. In Aktualnye voprosy ozeleneniya i ustoichivosti derevesnykh i kustarnikovykh porod v Centralnom Kazakhstane [Actual issues of greening and sustainability of tree and shrub species in Central Kazakhstan]; Kainar: Alma-Ata, USSR, 1975; pp. 148—157. (=Morphology of the oleaster jumping plant-lice (Trioza magnisetosa Log.), damaged Russian Olive in North Balkhash region).
- Khlebutina, L.G., 1982. Insects of the South and East of Kazakhstan, part 2, section 1, Psyllids of East Kazakhstan. In Report of the entomological laboratory of the Institute of Zoology of the Academy of Sciences of the Kazakh SSR, Institute of Zoology: Alma-Ata, 1982; pp. 4—45.
- Hodkinson, I. D. Life cycle variation and adaptation in jumping plant lice (Insecta: Hemiptera: Psylloidea): a global synthesis. Journal of Natural History 2009, 43(1), pp. 65—179. [CrossRef]
- Parfentiev, V.Ya. Vredityeli Urdinskikh lesnykh nasazhdeniy. In Trudy of Republican Station of Plant Protection (Kazakh branch of All-Union Lenin Academy of Agricultural Sciences), Academy of Sciences of Kazakh SSR: Alma-Ata, USSR, 1953; Issue I, pp. 59—61. (=Pests of Urda forest plantations).
- Kulenova, K.Z., 1968. Fauna i ekologicheskie osobennosti Zhukov-listoedov (Coleoptera, Chrysomelidae) yugo-vostoka Kazakhstana. In Trudy of the Institute of Zoology of the Academy of Sciences of the Kazakh SSR, Nauka: Alma-Ata, USSR, 1968; Issue 30, pp. 158—183. (=Fauna and ecological features of leaf beetles (Coleoptera, Chrysomelidae) South-East of Kazakhstan).
- Putshkov, V. G. Species of the genus Glaucopterum Wagner 1963 (Heteroptera, Miridae) of the Soviet Union fauna. Doklady Akademii Nauk Ukrainskoi SSR, Ser. B 1975, 11, pp. 1037—1042.
- Kerzhner, I.M. Novye i maloizvestnye vidy Heteroptera iz Mongolii i. Sopredelnikh rayonov SSSR. IV. Miridae, 1. In Nasekomye Mongolii [Insects of Mongolia); Nauka: Leningrad, USSR, 1984; Issue 9, pp. 35-72. (=New and little-known species of Heteroptera from Mongolia and adjacent regions of the USSR. IV. Miridae, 1).
- Vinokurov, N.N.; Dubatolov, V.V. Desert shield bug Brachynema germarii (Heteroptera: Pentatomidae) is found in the south of Eastern Siberia, Russia. Zoosystematica Rossica 2018, 27(1), pp. 146–149. [CrossRef]
- Yagdyev, A.; Tashlieva, A.O. Zhuki-vrediteli orekha i lokha v Turkmenii. In Ekologicheskoe i khozyaistvennoe znachenie nasekomykh Turkmenii; Ilim: Ashghabad, USSR, 1976; pp. 83–92. (=Beetle pests of walnut and oleaster in Turkmenia).
- Data sheets on quarantine pests. Aeolesthes sarta. In OEPP/EPPO Bulletin, 2005, Issue 35, pp. 387–389.
- Legalov, A.A.; Korotyaev, B.A. A new species of the genus Temnocerus Thunb. (Coleoptera: Rhynchitidae) from Kazakhstan. Baltic J. Coleopterol. 2006, 6 (2), pp. 125–127.
- Popescu-Gorj, A. Apotomis lutosana Kennel (Lepidoptera, Tortricidae) en Roumanie, espèce nouvelle pour la faune d’Europe. Travaux du Muséum d’Histoire Naturelle ’Grigore Antipa’ 1984, 25, pp. 237–238.
- Rikhter, V.A.; Durdyev, S.K. Tachinids (Diptera, Tachinidae) - parasites of Lepidoptera - orchard pests in Turkmenia. Vestnik Zoologii 1988, 1, pp. 62.
- Kuznetsov, V.I. Tortricidae. In: Keys to the Insects of the European Part of the USSR, IV Lepidoptera part 1, G.S. Medvedev, 1987; pp. 279-967.
- Bulyginskaya, M.A.; Akhmedov, A.M.; Dusmanov, I.E. Biological characteristics of the green tortricid Pandemis chondrillana H.-S. (Lepidoptera, Tortricidae) in the Penjikent district of Tajikistan. Entomologicheskoe Obozrenie 1994, 73(2), pp. 234–237.
- Skou, P. The geometroid moths of North Europe (Lepidoptera, Drepanidae and Geometridae). E.J.Brill/Scandinavian Science Press: Leiden-Copenhagen, Entomonograph, 1986, Issue 6, pp. 1–348.
- Forest pests on the territories of the former USSR. In: European and Mediterranean Plant Protection Organization, 2000, pp. 1–117. Available online: http://www.eppo.org/QUARANTINE/forestry_project/EPPOforestry_project.pdf (accessed on 12 February 2025).
Figure 1.
Map-scheme of the discovered populations of Altica balossogloi (yellow circles) and Trioza magnisetosa (blue circles) in nature and in experimental sites (greenish circles) in environs of Almaty and Avat Town in South East Kazakhstan.
Figure 1.
Map-scheme of the discovered populations of Altica balossogloi (yellow circles) and Trioza magnisetosa (blue circles) in nature and in experimental sites (greenish circles) in environs of Almaty and Avat Town in South East Kazakhstan.
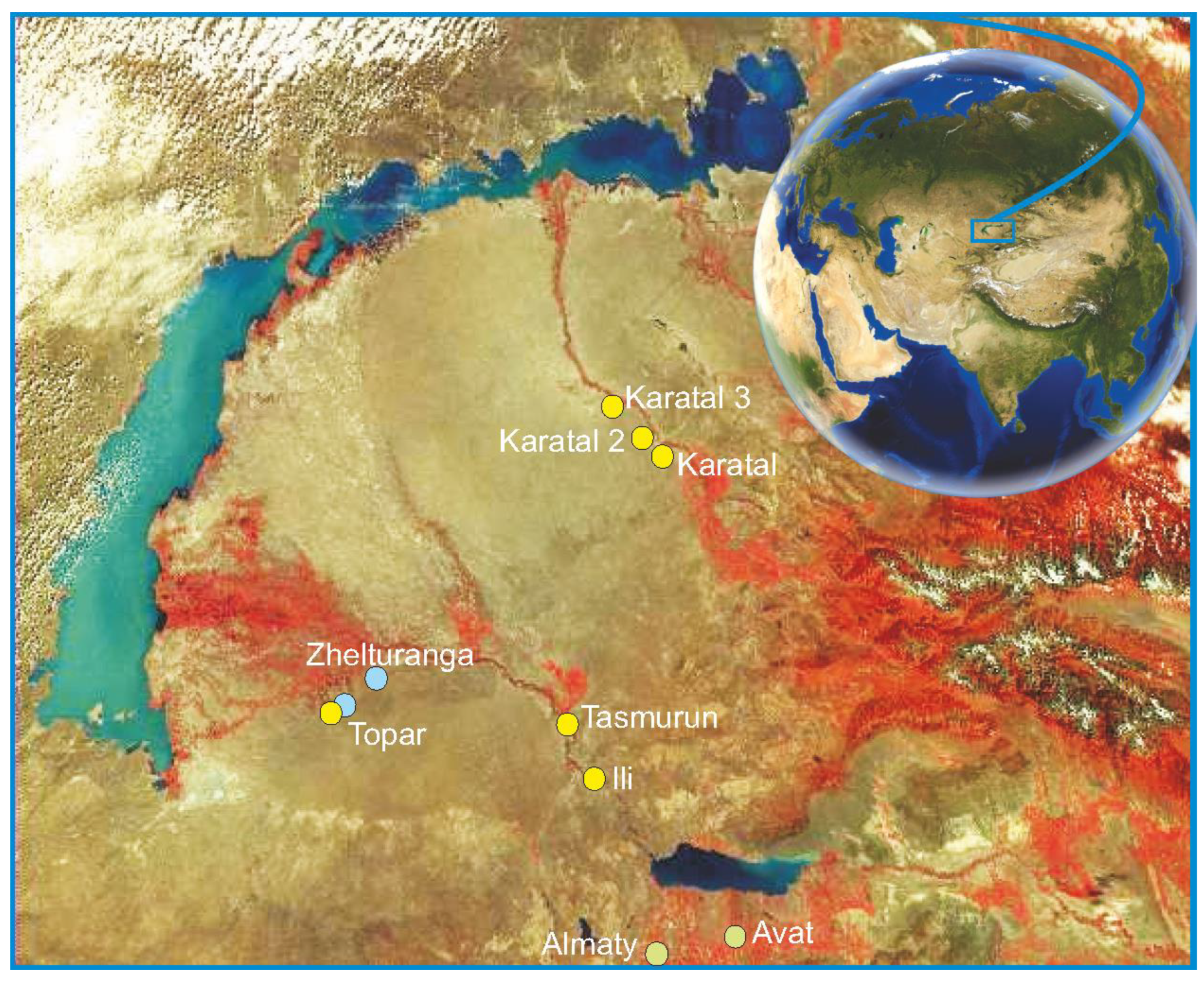
Figure 2.
a) Eggs of Altica balassogloi are laid in groups on the leaves and on the petioles and young branches; b) skeletonized leaves.
Figure 2.
a) Eggs of Altica balassogloi are laid in groups on the leaves and on the petioles and young branches; b) skeletonized leaves.


Table 1.
Phenogram of Triosa magnisetosa development in the southern Balkhash region.
| Months | ||||||||||||||||||
| IV | V | VI | VII | VIII | IX | X-XII-I-III | ||||||||||||
| Decades | Decades | Decades | Decades | Decades | Decades | |||||||||||||
| 1 | 2 | 3 | 1 | 2 | 3 | 1 | 2 | 3 | 1 | 2 | 3 | 1 | 2 | 3 | 1 | 2 | 3 | |
| Normal Spring | ||||||||||||||||||
|
3 Generation (Overwintered Adults) |
1 Generation (Adults) |
2 Generation (Adults) |
3 Generation (Overwintered Adults) | |||||||||||||||
| W | W | W | W | |||||||||||||||
| ♂ | ♂ | ♂ | ♂ | ♂ | ♂ | ♂ | ♂ | ♂ | ♂ | ♂ | ♂ | ♂ | ♂ | ♂ | ♂ | ♂ | ||
| ♀ | ♀ | ♀ | ♀ | ♀ | ♀ | ♀ | ♀ | ♀ | ♀ | ♀ | ♀ | ♀ | ♀ | ♀ | ♀ | ♀ | ||
| ◎ | ◎ | ◎ | ◎ | ◎ | ◎ | ◎ | ◎ | ◎ | ◎ | ◎ | ◎ | |||||||
| oo | oo | oo | oo | oo | oo | oo | oo | oo | ||||||||||
| L1 | L1 | L1 | L1 | L1 | L1 | |||||||||||||
| L2 | L2 | L2 | L2 | L2 | L2 | |||||||||||||
| L3 | L3 | L3 | L3 | L3 | L3 | |||||||||||||
| L4 | L4 | L4 | L4 | L4 | L4 | |||||||||||||
| L5 | L5 | L5 | L5 | L5 | L5 | |||||||||||||
* Notes: W – wintering adults on the ground; ♂ – male; ♀ – female (adults moving and, on a tree,); ◎ – copulation; oo – eggs; L1 - first instar larva, L2 – second instar larva, L3 – third instar larva, L4 – fourth instar larva, L5 – fifth instar larva.
Table 2.
Phenogram of Altica balassogloi development in the southern Balkhash region.
| Months | ||||||||||||||||||
| IV | V | VI | VII | VIII | IX | X-XII-I-III | ||||||||||||
| Decades | Decades | Decades | Decades | Decades | Decades | |||||||||||||
| 1 | 2 | 3 | 1 | 2 | 3 | 1 | 2 | 3 | 1 | 2 | 3 | 1 | 2 | 3 | 1 | 2 | 3 | |
| Normal Spring | ||||||||||||||||||
| W | W | W | W | W | W | W | ||||||||||||
| ♂ | ♂ | ♂ | ♂ | ♂ | ♂ | ♂ | ||||||||||||
| ♀ | ♀ | ♀ | ♀ | ♀ | ♀ | ♀ | ||||||||||||
| ◎ | ◎ | ◎ | ◎ | |||||||||||||||
| oo | oo | oo | oo | |||||||||||||||
| L1 | L1 | L1 | L1 | |||||||||||||||
| L2 | L2 | L2 | L2 | |||||||||||||||
| L3 | L3 | L3 | L3 | |||||||||||||||
| P | P | P | P | |||||||||||||||
| Earlier Warm Spring | ||||||||||||||||||
| W | W | W | W | W | ||||||||||||||
| ♂ | ♂ | ♂ | ♂ | ♂ | ♂ | ♂ | ♂ | ♂ | ♂ | ♂ | ||||||||
| ♀ | ♀ | ♀ | ♀ | ♀ | ♀ | ♀ | ♀ | ♀ | ♀ | ♀ | ||||||||
| ◎ | ◎ | ◎ | ◎ | |||||||||||||||
| oo | oo | oo | oo | oo | ||||||||||||||
| L1 | L1 | L1 | L1 | L1 | L1 | |||||||||||||
| L2 | L2 | L2 | L2 | L2 | L2 | |||||||||||||
| L3 | L3 | L3 | L3 | L3 | L3 | |||||||||||||
| P | P | P | P | P | ||||||||||||||
* Notes: W - wintering adults on the ground/under bark; ♂ - male; ♀ - female (adults moving and on a tree); ◎ – copulation; oo – eggs; L1 - first instar larva, L2 - second instar larva, L3 - third instar larva; P – pupa.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



