
Submitted:
20 February 2025
Posted:
21 February 2025
You are already at the latest version
Abstract
Agmatine is a naturally occurring biogenic amine, acts primarily as an inhibitor of neuronal nitric oxide synthase (nNOS). Previous studies have shown that both acute and chronic agmatine administration induced anxiolytic and antidepressant-like effects in rodents. In the dorsal raphe nucleus (DRN), nitric oxide (NO) donors inhibit serotonergic (5-HT) neuronal activity, with nNOS expressing 5-HT neurons showing lower baseline firing rates than non-nNOS expressing neurons. Our study aimed to test the hypothesis that the psychoactive effects of agmatine are mediated, at least in part, via a mechanism involving the stimulation of the DRN 5-HT neurons, and to assess the molecular pathway allowing agmatine to modulate the excitability of 5-HT neurons. Using extracellular in vivo electrophysiology, we demonstrated that both acute (1-3 mg/kg, i.v.) and chronic (40 mg/kg/day, i.p., 14 days) agmatine administration significantly increased the firing rate of DRN 5-HT neurons. Quantitative PCR (qPCR) analysis revealed that chronic agmatine treatment selectively upregulated the expression of serotonin-1B (5-HT1B) and serotonin-2A (5-HT2A) receptor mRNA in the DRN. Since previous studies have shown that DRN 5-HT2A receptor activation stimulates 5-HT neurons and produces antidepressant-like effects, out findings suggest that agmatine’s excitatory effect on DRN 5-HT neurons may be partially 5-HT2A receptor-dependent. . Given that modulation of the 5-HT neuronal firing activity is critical for the proper antidepressant efficacy, nNOS inhibitors can be potential antidepressants by their own and/or effective adjuncts to other antidepressant drugs.
Keywords:
neuronal nitric oxide (NO) synthase (nNOS)
; serotonin (5-HT) transporter (SERT)
; serotonin-1A/AB receptors (5-HT1A/1B)
; serotonin-2A/2B/2C receptors (5-HT2A/2B/2C)
; antidepressant drug response
1. Introduction
Agmatine, a naturally occurring biogenic amine, is synthesized through the amino acid L-arginine by the enzyme arginine decarboxylase. Since its discovery in mammalian brains in the 20th century, agmatine has garnered considerable scientific attention due to its diverse biological activities and therapeutic potential [1,2]. One of the primary biological functions of agmatine is inhibition of the nitric oxide (NO) synthase (NOS) [3,4], particularly, of the neuronal form (nNOS) of this enzyme [5]. Since NO is an important neuromodulator, and nNOS is its primary source within the central nervous system (CNS), this enzyme modulates multiple brain neurotransmitter systems [6]. Among the CNS neurotransmitter systems modulated by nNOS, there is a serotonergic (5-HT) system, playing a key role in pathophysiology and treatment of depression and related stress-related anxiety and mood disorders [7].
It was reported that nNOS molecularly binds to and creates a heterodimer with the 5-HT transporter (SERT) [8]. The latter plays a key role in the regulation of the extracellular 5-HT concentrations in the dorsal raphe nucleus (DRN), a brain area containing the cell bodies of 5-HT-secreing neurons. Extracellular 5-HT negatively regulates the firing activity of 5-HT neurons, via a mechanism primarily involving somatodendritic serotonin-1A (5-HT1A) autoreceptors. The 5-HT1A autoreceptors-mediated inhibition of 5-HT neurons of the DRN is responsible, at least in part, for the delayed behavioral response and/or for the lack of adequate therapeutic response to antidepressant drugs, such as the selected serotonin (5-HT) reuptake inhibitors [7]. Consistently, molecules dissociating the nNOS-SERT dimerization were reported to have robust and rapid antidepressant-like effect [9] This molecular interaction between nNOS and SERT represents a potential therapeutic target, as disruption of this complex has been shown to produce rapid antidepressant effects.
Agmatine, as a natural inhibitor of the NOS/nNOS, exhibits significant psychoactive effects through modulation of 5-HT neurotransmission. Chronic agmatine treatment produces robust antidepressant-like behavioral effect in mice, accompanied by increased extracellular brain concentrations of 5-HT and glutamate along with elevated expression of neuroplasticity markers such as brain derived neurotrophic factor (BDNF) and synaptotagmin I [10]. Additional studies have confirmed both anxiolytic and antidepressant-like effects following chronic agmatine treatment [11,12]. Notably, even acute agmatine treatment demonstrates antidepressant-like effect and increases BDNF levels [13] with similar anxiolytic effects reported in mice [14]. The mechanism underlying these behavioral effects may be linked to agmatine’s influence on 5-HT neuronal activity, as previous studies have shown that DRN 5-HT neurons expressing nNOS exhibit lower spontaneous firing rates compared to non-NOS expressing neurons. This is further supported by the observation that the nitric oxide donor diethylamineNONOate directly inhibits 5-HT neuronal firing in vitro electrophysiology [15]. These findings suggest that agmatine's psychoactive effects might be mediated through disinhibition of serotonergic neurotransmission via nNOS inhibition.
Bulding on previous findings suggesting NO’s role in serotonergic signaling, weaimed to investigate, using in vivo electrophysiology, both acute and chronic effects of agmatine on 5-HT neuronal firing activity. We further investigated potential molecular adaptations following chronic agmatine treatment by analyzing the expression of nNOS, serotonin transporter (SERT), and key 5-HT receptor subtypes (serotonin-1A/AB receptors (5-HT1A/1B) and serotonin-2A/2B/2C (5-HT2A/2B/2C)) in the rat dorsal raphe nucleus (DRN).
2. Results
2.1. Effects Acute and Chronic Agmatine Treatment on the Firing Activity of 5-HT Neurons in the DRN
Figure 1 illustrates the effect of acute intravenous (i.v.) administration of agmatine, at the cumulative doses of 1-3 mg/kg, on the firing activity of 5-HT neurons of the DRN. Agmatine significantly and dose-dependently increased the firing rate of 5-HT neurons, comparing to their own basal firing activity (F3,22 = 4.45, analysis of variance for repeated measures: RM ANOVA). Bonferroni post-hoc test confirmed a significant difference between the basal firing activity of 5-HT neurons and their firing activity after the administration of 2 and 3 mg/kg of agmatine (p<0.05). We showed that the neuronal firing rate demonstrated a progressive increase correlated with increasing agmatine doses.
2.2. Effects of Chronic Agmatine Treatment on the Expression of 5-HT Neurons in the DRN
Figure 2 illustrates the effect of chronic intraperitoneal (i.p.) administration of agmatine on the firing activity of 5-HT neurons in the DRN. In this experiment, rats received daily agmatine injections (40 mg/kg) for 14 consecutive days.
Chronic agmatine treatment led to the significant increase in the mean spontaneous firing rate of 5-HT neurons, compared to the vehicle-treated control animals (p<0.01, two-tailed Student’s t-test). To further characterize the effects of chronic agmatine treatment, we examined the expression of nNOS, SERT, and serotonin receptor subtypes in the dorsal raphe nucleus (DRN) using quantitative polymerase chain reaction (qPCR).
2.3. Effects of Chronic Agmatine Treatment on the nNOS and SERT mRNA Expression in the DRN
Figure 3 presents the results of RT-qPCR analysis examining the expression of mRNA encoding nNOS and SERT in the DRN. Contrary to the initial hypothesis, chronic i.p. administration of agmatine (40 mg/kg/day for 14 days) did not induce statistically significant changes in the mRNA expression levels of either nNOS or SERT in the DRN (p>0.05, two-tailed Student’s t-test).
2.4. Effects of Chronic Agmatine Treatment on the Expression of the DRN 5-HT Receptors Belonging to the 5-HT1 Subfamily
Figure 4 illustrates the effect of chronic agmatine treatment on the mRNA expression of 5-HT receptors belonging to the 5-HT1 subfamily in the DRN.
For the mRNA encoding 5-HT1A receptor in the agmatine-treated rats showed no statistical difference compared to the control group. In contrast, chronic agmatine treatment led to a robust and statistically significant increase in the expression of 5-HT1B receptor encoding mRNA in the DRN (**p<0.01, two-tailed Student’s t-test).
2.5. Effects of Chronic Agmatine Treatment on the Expression of the DRN 5-HT Receptors Belonging to the 5-HT2 Subfamily
Figure 5 illustrates the effect of chronic agmatine treatment on the mRNA expression of serotonin-2A (5-HT2A), serotonin-2B (5-HT2B), and serotonin-2C (5-HT2C) receptors in the DRN.
Chronic agmatine led to the statistically significant increase in the expression of mRNA encoding 5-HT2A receptor in the DRN, compared to the vehicle-treated controls (*p<0.05, two-tailed Student’s t-test). The mRNA expression of 5-HT2B and 5-HT2C receptors in the agmatine-treated rats was not statistically different from that in the vehicle-treated controls (p>0.05, two-tailed Student’s t-test).
3. Discussion
We found that both acute and chronic agmatine stimulated 5-HT neurons of the DRN. Chronic agmatine also enhances the expression of 5-HT2A receptors in the DRN. The DRN expressions of other subtypes of 5-HT receptors, belonging to the 5-HT1 and 5-HT2 subfamilies, as well as the expression of nNOS and SERT, were not affected by chronic agmatine treatment.
It was found that acute administration of agmatine led to the activation of 5-HT neurons of the DRN (Fig 1). It is consistent with the inhibitory effect of the NO donor on 5-HT neurons, reported in the previous study [15]. Since 5-HT neurotransmission is fundamental in antidepressant drug response [7], our findings may explain the antidepressant-like behavioral effect of acute agmatine, reported in a previous study [13]. Since 5-HT stimulates BDNF expression [16], excitation of 5-HT neurons by acute agmatine may explain its ability to enhance BDNF levels, reported in the same study. Our findings on agmatine’s bi-directional effects on 5-HT neuronal firing is consistent with the previous work showing that 5-HT1B autoreceptor expression in the DRN can either decrease or increase anxiety-like behaviors depending on stress context, suggesting that modulation of serotonergic signaling has complex state-dependent effects on emotional behavior [17].
The acute effect of agmatine on the excitability of the DRN 5-HT neurons was observed within seconds after the drug injection. It is therefore unlikely that any alteration in a protein and/or mRNA expression underlines the acute agmatine effect of the excitability of 5-HT neurons. It is however possible that the rapid dissociation of the nNOS/SERT dimmer and subsequent 5-HT1A-autoreceptor-mediated disinhibition of 5-HT neurons is involved [9]. Further experiments should be performed to test this hypothesis.
The mean spontaneous firing activity of 5-HT neurons in the agmatine-treated rats was significantly higher than in the vehicle-treated control (Fig 2). This finding is consistent with the previously observed antidepressant, anxiolytic, neuroprotective, and BDNF expression-enhancing effects of chronic agmatine [10,11,14]. It is also consistent with the previous finding that 5-HT neurons expressing the nNOS have lower spontaneous firing activity than the non-nNOS expressing ones [15].
It is known that the certain psychoactive drugs have distinct acute and chronic effects on the excitability of 5-HT neurons. Thus, acute selective serotonin reuptake inhibitor (SSRI) escitalopram [18] or trace amine associated receptor 1 (TAAR1) agonist RO5256390 [19] inhibit 5-HT neurons, when the chronic administration of these drugs have no effect on the firing activity of 5-HT neurons. It is also well established that the adaptive changes in the expression and/or activity of the SERT and 5-HT1A receptors are responsible for the different response of 5-HT neurons to the acute and chronic administration of the same psychoactive drugs [7]. In the present study, the persistence of the ability of agmatine to stimulate 5-HT neurons after its chronic administration (Fig 2) is consistent with the lack of adaptive changes in the expression of the SERT and 5-HT1A receptors after chronic treatment with this drug (Figs 3 and 4).
It was found that chronic agmatine treatment led to the increased expression of 5-HT1B receptor-coding mRNA in the DRN (Fig 4). The 5-HT1B receptor-coding mRNA, detected in the DRN, is likely to be expressed within the cell bodies of 5-HT neurons. This mRNA is latterly translated into the HT1B receptor protein, expressed in the nerve terminals of 5-HT neurons in various brain areas, such as hippocampus. These nerve terminal 5-HT1B autoreceptors negatively regulate 5-HT neurotransmission [20]. It was however reported that the 5-HT1B autoreceptor-mediated suppression of 5-HT release from the nerve terminal did not necessarily involve suppression of firing of 5-HT neurons. Furthermore, a 5-HT1B agonist even increased, and a 5-HT1B antagonist decreased the firing rate of 5-HT neurons [21,22]. The increased expression of HT1B autoreceptors, observed after chronic agmatine, does not therefore contradict the increase in the tonic firing activity of 5-HT neurons.
The observed upregulation of 5-HT1B receptor expression following chronic agmatine treatment warrants careful interpretation. While this effect could be directly related to agmatine’s action, several alternative mechanisms should be considered. First, the chronic intravenous administration protocol itself may have introduced a stress component that could influence 5-HT1B receptor expression, as previous studies have shown stress-dependent modulation of serotonergic systems [17]. Additionally, the increased 5-HT1B receptor expression may represent a compensatory homeostatic response to enhanced serotonergic transmission induced by agmatine. Since 5-HT1B receptor function as inhibitory autoreceptors on serotonergic terminals, their upregulation could serve as a negative feedback mechanism to maintain appropriate serotonin release. Furthermore, given that DRN serotonergic neurons receive substantial dopaminergic and glutamatergic inputs, agmatine’s effects on 5-HT1B expression might be mediated indirectly through these neurotransmitter systems. To complex interplay between these various mechanisms deserves further investigation, particularly using selective antagonists and region-specific manipulations to dissect the relative contributions of direct and indirect pathways.
We found that the sustained treatment with agmatine led to increased expression of mRNA coding for 5-HT2A receptors in the DRN (Fig 5). Since 5-HT2A receptors are not known to act as autoreceptors, the 5-HT2A receptor-coding mRNA, detected in the DRN, is likely to be expressed in the non-5-HT neurons, such as GABAergic and/or opioidergic interneurons [23,24]. It was reported that the local administration of 5-HT2A/2C receptor agonist (+)-DOI hydrochloride (DOI) into the DRN activated local 5-HT neurons. Another study suggested that the 5-HT2 receptor-modulated stimulation of the DRN 5-HT neurons involves the excitation of opioidergic interneurons [23]. It is therefore possible that the chronic agmatine-induced stimulation of 5-HT neurons involves a 5-HT2A receptor-based mechanism. The involvement of 5-HT2A receptors in the beneficial CNS effect of agmatine has been previously reported as well. Santos and colleagues [25] found that the agmatine-induced antinociception in mice was significantly attenuated by 5-HT2A receptor antagonist ketanserin. Freitas and co-authors [26] reported that ketanserin abolished the neuroprotective effect of agmatine in the hippocampal neuronal cell culture. The ability of chronic agmatine to upregulate both 5-HT1B and 5-HT2A receptor expression while stimulating 5-HT neuronal firing aligns with previous evidence that baseline serotonergic signaling may play a protective role against anxiety under normal conditions, though these relationships become more complex following stress exposure.
The primary limitation of this study is that the expression of 5-HT receptors was assessed in the DRN only. It is however known that 5-HT1A/1B receptors expressed in other brain areas, such as prefrontal cortex [27], are also involved in the regulation of excitability of the DRN 5-HT neurons. The effect of chronic agmatine on the nNOS, SERT, and 5-HT receptors in brain areas other than the DRN should be assessed in future studies.
4. Materials and Methods
4.1. Animals
Male Wistar rats (initial weight 250–350 g; 2–3 months old) were obtained from the Department of Toxicology and Laboratory Animals Breeding, Centre of Experimental Medicine of the Slovak Academy of Sciences, Dobra Voda, Slovak Republic. All the animals were housed (38 × 59 × 25 cm large cages) under standard laboratory conditions (temperature: 22 ± 2°C, humidity: 55 ± 10%) with a 12 h light/12 h dark cycle (lights on at 7.00 a.m.). Pelleted food and tap water were available ad libitum. The State Veterinary and Food Administration of the Slovak Republic approved all the experimental procedures. The rats were handled according to the Guide for the Care and Use of Laboratory Animals (N.R.C., 1996) and the European Communities Council Directive of September 22, 2010 (2010/63/EU, 74).
4.2. Chemicals
Agmatine sulfate salt (A7127-5G, ≥97% purity) was purchased from Sigma-Aldrich. DNA/RNA Shield (R1100-50, 50 ml) and Quick-RNA Microprep Kit (R1050), obtained from Zymo Research. SOLIScript® 1-step Multiplex Probe Kit purchased from Solis Biodyne. Primers and probes were synthesized by MultiplexDX s.r.o. (Bratislava, Slovakia). All chemicals were of analytical grade and stored per manufacturer guidelines.
4.3. Electrophysiology
In vivo electrophysiological experiments were performed as previously described [19,28,29]. The rats were anesthetized with chloral hydrate (0.4 g/kg, i. p) and mounted in the stereotaxic frame (David Kopf Instruments, Tujunga, CA). Their scalp was opened, and a 3 mm hole was drilled in the skull for insertion of electrodes. Glass electrodes pulled with a DMZ-Universal Puller (Zeitz-Instruments GmbH, Martinsried, Germany) to a fine tip of ∼1 µM and filled with 2M sodium chloride (NaCl). The impedance of the electrodes was 4–6 MΩ. The electrodes were lowered through the DRN using the hydraulic micro-positioner (David Kopf Instruments, Tujunga, CA). The action potentials generated by the neurons were recorded using the AD Instruments Extracellular Recording System (Dunedin, New Zealand). The 5-HT neurons of the DRN were identified according to the waveform of their action potentials and the pattern of their generation, as explained in our previous works [19,28,29]. During the experiment, the rats’ body temperature was maintained at 37°C with a heating pad (Gaymor Instruments, Orchard Park, NY, United States).
4.4. Assessment of the Acute Agmatine Effect on 5-HT Neuronal Firing Activity
In experiments aiming to assess the effect of acute agmatine on the firing activity of 5-HT neurons, after a first DRN 5-HT neuron with a stable firing activity was found, its basal firing was recorded for two minutes. Thereafter, 1 mg/kg of agmatine was administered via a catheter placed in the femoral vein. The neuronal firing activity was recorded for another two minutes. Subsequently, additional dose of 1 mg/kg of agmatine was administered (the cumulative dose 2 mg/kg) and the neuronal firing activity was recorded for another two minutes. Finally, the last administration of 1 mg/kg of agmatine was performed (the cumulative dose 2 mg/kg), and the neuron was recorded for the last two minutes. Thereafter, the animal was euthanized by overdose of chloralhydrate.
4.6. Assessment of the Chronic Agmatine Effect on 5-HT Neuronal Firing Activity
In experiments aiming to assess the effect of chronic agmatine on the firing activity of 5-HT neurons, rats were randomly divided into two groups. One group received daily injections of agmatine (40 mg/kg/day, i.p.). The control animals received daily i.p. injections of the vehicle (saline: 0.9% sodium chloride: NaCl in water). On the 15th day, electrophysiological experiments were performed. The electrode was lowered through the DRN 3-4 times and the spontaneously active 5-HT neurons detected during each electrode descend were recorded. After the completion of the electrophysiological recording, the rats were euthanized by overdose of chloralhydrate.
4.7. Assessment of Target Gene Expression in DRN Using Multiplex RT-qPCR
Following electrophysiological recordings, rats were euthanized by decapitation, and their brains were rapidly removed. Fresh brains were placed on a cold plate and sectioned coronally with razor blade. The DRN region was precisely isolated from these coronal sections using a punch tool (1.5 mm diameter) and transferred to 1.5 ml tubes. The samples were immediately flash-frozen in liquid nitrogen and stored at -80°C until further analysis. On the day of PCR experiments, DNA/RNA Shield solution (300 µL; Zymo Research) was added to the samples immediately upon removal from -80°C storage to prevent RNA degradation. Total RNA was extracted using the Quick-RNA Miniprep Kit with DNase I treatment (Zymo Research) according to the manufacturer's protocol.
RT-qPCR primer and probe sequences were designed for multiplex analysis. Primers and dual-labeled hydrolysis probes were obtained from MultiplexDX s.r.o. (Bratislava, Slovakia). The sequences are detailed in Table 1. Each probe was labeled at the 5' end with a fluorescent reporter dye (FAM, HEX, ROX and Cy5) and at the 3' end with a quencher dye (BHQ1, BHQ2). BLAST analysis confirmed the specificity of the primer/probe sets to their target sequences.
First multiplex reaction contains 5-HT1A, 5-HT2A, nNOS, and GAPDH. Second multiplex reaction contain 5-HT1B and GAPDH. Third multiplex reaction contains 5-HT2B, 5-HT2C, SERT, and GAPDH. For first and third multiplex reaction, the thermal cycling conditions were: cDNA synthesis at 50°C for 30 minutes, initial denaturation at 95°C for 10 minutes, followed by 40 cycles of denaturation at 95°C for 15 seconds and combined annealing/extension at 60°C for 1 minute with plate read. For second multiplex reaction, the protocol was modified with an annealing temperature of 56°C for 1 minute during the cycling steps, while maintaining the same initial cDNA synthesis and denaturation conditions. Gene expression was analyzed using CFX Maestro Software (Bio-Rad) using the 2^-ΔΔCt method, with GAPDH serving as the reference gene. Relative expression values were calculated and normalized to control samples.
4.8. Statistical Analysis
To examine the effect of different doses of acute agmatine on 5-HT neuronal firing activity, one-way RM ANOVA with a time as a factor of comparison (basal and after the administration of 1, 2, and 3 mg/kg of agmatine) , followed by the Bonferroni post hoc test, was used. To examine the effect of chronic agmatine on the firing rate of 5-HT neurons and DRN expression of the NOS, SERT, 5-HT1A/1B, and 5-HT2A/2B/2C mRNA, two-tailed Student’s t-test was applied to assess the difference between the neuronal firing rate or mRNA expression in agmatine- and vehicle-treated animals. The results are expressed as the mean ± standard error of the mean (SEM), and a value of p ≤ 0.05 was considered statistically significant.
5. Conclusions
We found that acute and chronic treatment with agmatine led to the stimulation of 5-HT neurons of the DRN. The ability to stimulate central 5-HT neurons might explain the anxiolytic and antidepressant-like effects of agmatine observed in previous studies. While the acute effect of agmatine is likely to be based on its direct effect on the nNOS-SERT complex, the chronic effect of this drug putatively involves upregulation of the 5-HT2A receptor. Since the lack of the timely and adequate response to the antidepressant drugs is frequently resulting from the auto-inhibition 5-HT neurotransmission, the ability of the nNOS inhibitors to stimulate 5-HT neurotransmission may make them potential antidepressants by their own and/or adjuncts to other antidepressants, such as SSRIs and/or TAAR1 agonists. On the other hand, chronic agmatine-induced increase in the expression of 5-HT1B autoreceptors might have a diminishing effect of the net 5-HT transmission. The exact effect of the nNOS inhibition on the nerve-terminal 5-HT release should be examined in the future studies.
Author Contributions
Conceptualization H.Ö. and E.D.; investigation H.Ö., R.P., D.G., and E.D.; methodology H.Ö., R.P., R.D., D. G. and E.D.; data curation and visualization H.Ö., D. G., and E.D.; writing—original draft preparation H.Ö. and E.D.; supervision, D.G. and E.D.; project administration, D.G. and E.D.; funding acquisition, D.G. and E.D. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the Slovak Research and Development Agency under Contracts No. APVV-20-0202 and by the Research Grant Agency of the Slovak Academy of Sciences and Ministry of Education, Science, Research and Sport of the Slovak Republic via the grants No. VEGA 2/0057/22 and VEGA 2/0133/23.
Institutional Review Board Statement
The State Veterinary and Food Administration of the Slovak Republic approved all experimental procedures with animals (No. 3334-5/2020-220).
Data Availability Statement
The data that support the findings of this study are available from the corresponding authors upon reasonable request.
Acknowledgments
The authors thank Katarína Ondáčová for her outstanding technical assistance and sincerely appreciate Halil Önder Özbaşak for his support in qPCR analyses and proof-reading.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| 5-HT | Serotonin |
| ANOVA | Analysis of variance |
| MDPI | Multidisciplinary Digital Publishing Institute |
| NO | Nitric oxide |
| NOS | Nitric oxide synthase |
| nNOS | Neuronal nitric oxide synthase |
| MDPI | Multidisciplinary Digital Publishing Institute |
| SERT | 5-HT transporter |
| SSRIsTAAR1 | Selective serotonin reuptake inhibitorsTrace amine associated receptor 1 |
References
- Molderings, G. J.; Haenisch, B., Agmatine (decarboxylated l-arginine): Physiological role and therapeutic potential. Pharmacology & Therapeutics 2012, 133, (3), 351-365. [CrossRef]
- Piletz, J. E.; Aricioglu, F.; Cheng, J.-T.; Fairbanks, C. A.; Gilad, V. H.; Haenisch, B.; Halaris, A.; Hong, S.; Lee, J. E.; Li, J.; Liu, P.; Molderings, G. J.; Rodrigues, A. L. S.; Satriano, J.; Seong, G. J.; Wilcox, G.; Wu, N.; Gilad, G. M., Agmatine: clinical applications after 100 years in translation. Drug Discovery Today 2013, 18, (17), 880-893. [CrossRef]
- Galea, E.; Regunathan, S.; Eliopoulos, V.; Feinstein, D. L.; Reis, D. J., Inhibition of mammalian nitric oxide synthases by agmatine, an endogenous polyamine formed by decarboxylation of arginine. Biochem J 1996, 316 ( Pt 1), (Pt 1), 247-9. [CrossRef]
- Gadkari, T. V.; Cortes, N.; Madrasi, K.; Tsoukias, N. M.; Joshi, M. S., Agmatine induced NO dependent rat mesenteric artery relaxation and its impairment in salt-sensitive hypertension. Nitric Oxide 2013, 35, 65-71. [CrossRef]
- Demady, D. R.; Jianmongkol, S.; Vuletich, J. L.; Bender, A. T.; Osawa, Y., Agmatine enhances the NADPH oxidase activity of neuronal NO synthase and leads to oxidative inactivation of the enzyme. Molecular pharmacology 2001, 59, (1), 24-29.
- Zhu, L. J.; Li, F.; Zhu, D. Y., nNOS and Neurological, Neuropsychiatric Disorders: A 20-Year Story. Neurosci Bull 2023, 39, (9), 1439-1453. [CrossRef]
- Grinchii, D.; Dremencov, E., Mechanism of Action of Atypical Antipsychotic Drugs in Mood Disorders. Int J Mol Sci 2020, 21, (24).
- Garthwaite, J., Neuronal nitric oxide synthase and the serotonin transporter get harmonious. Proceedings of the National Academy of Sciences 2007, 104, (19), 7739-7740.
- Sun, N.; Qin, Y.-J.; Xu, C.; Xia, T.; Du, Z.-W.; Zheng, L.-P.; Li, A.-a.; Meng, F.; Zhang, Y.; Zhang, J., Design of fast-onset antidepressant by dissociating SERT from nNOS in the DRN. Science 2022, 378, (6618), 390-398.
- Freitas, A. E.; Egea, J.; Buendia, I.; Gómez-Rangel, V.; Parada, E.; Navarro, E.; Casas, A. I.; Wojnicz, A.; Ortiz, J. A.; Cuadrado, A.; Ruiz-Nuño, A.; Rodrigues, A. L. S.; Lopez, M. G., Agmatine, by Improving Neuroplasticity Markers and Inducing Nrf2, Prevents Corticosterone-Induced Depressive-Like Behavior in Mice. Mol Neurobiol 2016, 53, (5), 3030-3045.
- Aglawe, M. M.; Kale, M. B.; Rahangdale, S. R.; Kotagale, N. R.; Umekar, M. J.; Taksande, B. G., Agmatine improves the behavioral and cognitive impairments associated with chronic gestational ethanol exposure in rats. Brain Res Bull 2021, 167, 37-47.
- Sahin Ozkartal, C.; Tuzun, E.; Kucukali, C. I.; Ulusoy, C.; Giris, M.; Aricioglu, F., Antidepressant-like effects of agmatine and NOS inhibitors in chronic unpredictable mild stress model of depression in rats: The involvement of NLRP inflammasomes. Brain Res 2019, 1725, 146438. [CrossRef]
- Neis, V. B.; Moretti, M.; Bettio, L. E.; Ribeiro, C. M.; Rosa, P. B.; Gonçalves, F. M.; Lopes, M. W.; Leal, R. B.; Rodrigues, A. L., Agmatine produces antidepressant-like effects by activating AMPA receptors and mTOR signaling. Eur Neuropsychopharmacol 2016, 26, (6), 959-71.
- Jiang, X. Z.; Li, Y. F.; Zhang, Y. Z.; Chen, H. X.; Li, J.; Wang, N. P., [5-HT1A/1B receptors, alpha2-adrenoceptors and the post-receptor adenylate cyclase activation in the mice brain are involved in the antidepressant-like action of agmatine]. Yao Xue Xue Bao 2008, 43, (5), 467-73.
- Gartside, S. E.; Yurttaser, A. E.; Burns, A. L.; Jovanović, N.; Smith, K. J.; Amegashiti, N. S.; Olthof, B. M. J., A role for nitric oxide in serotonin neurons of the midbrain raphe nuclei. Eur J Neurosci 2020, 51, (9), 1881-1899. [CrossRef]
- Dremencov, E.; Jezova, D.; Barak, S.; Gaburjakova, J.; Gaburjakova, M.; Kutna, V.; Ovsepian, S. V., Trophic factors as potential therapies for treatment of major mental disorders. Neurosci Lett 2021, 764, 136194. [CrossRef]
- Clark, M. S.; Vincow, E. S.; Sexton, T. J.; Neumaier, J. F., Increased expression of 5-HT1B receptor in dorsal raphe nucleus decreases fear-potentiated startle in a stress dependent manner. Brain Res 2004, 1007, (1-2), 86-97.
- Dremencov, E.; El Mansari, M.; Blier, P., Distinct electrophysiological effects of paliperidone and risperidone on the firing activity of rat serotonin and norepinephrine neurons. Psychopharmacology (Berl) 2007, 194, (1), 63-72.
- Grinchii, D.; Hoener, M. C.; Khoury, T.; Dekhtiarenko, R.; Nejati Bervanlou, R.; Jezova, D.; Dremencov, E., Effects of acute and chronic administration of trace amine-associated receptor 1 (TAAR1) ligands on in vivo excitability of central monoamine-secreting neurons in rats. Mol Psychiatry 2022, 27, (12), 4861-4868.
- Tiger, M.; Varnäs, K.; Okubo, Y.; Lundberg, J., The 5-HT(1B) receptor - a potential target for antidepressant treatment. Psychopharmacology (Berl) 2018, 235, (5), 1317-1334.
- Sinton, C. M.; Fallon, S. L., Electrophysiological evidence for a functional differentiation between subtypes of the 5-HT1 receptor. Eur J Pharmacol 1988, 157, (2-3), 173-81. [CrossRef]
- Hertel, P.; Lindblom, N.; Nomikos, G. G.; Svensson, T. H., Receptor-mediated regulation of serotonin output in the rat dorsal raphe nucleus: effects of risperidone. Psychopharmacology (Berl) 2001, 153, (3), 307-14.
- Ferreira, M. D.; Menescal-de-Oliveira, L., Nociceptive vocalization response in guinea pigs modulated by opioidergic, GABAergic and serotonergic neurotransmission in the dorsal raphe nucleus. Brain Res Bull 2014, 106, 21-9.
- Hernández-Vázquez, F.; Garduño, J.; Hernández-López, S., GABAergic modulation of serotonergic neurons in the dorsal raphe nucleus. Rev Neurosci 2019, 30, (3), 289-303.
- Santos, A. R.; Gadotti, V. M.; Oliveira, G. L.; Tibola, D.; Paszcuk, A. F.; Neto, A.; Spindola, H. M.; Souza, M. M.; Rodrigues, A. L.; Calixto, J. B., Mechanisms involved in the antinociception caused by agmatine in mice. Neuropharmacology 2005, 48, (7), 1021-34.
- Freitas, A. E.; Egea, J.; Buendía, I.; Navarro, E.; Rada, P.; Cuadrado, A.; Rodrigues, A. L.; López, M. G., Agmatine induces Nrf2 and protects against corticosterone effects in hippocampal neuronal cell line. Mol Neurobiol 2015, 51, (3), 1504-19. [CrossRef]
- Hajós, M.; Hajós-Korcsok, E.; Sharp, T., Role of the medial prefrontal cortex in 5-HT1A receptor-induced inhibition of 5-HT neuronal activity in the rat. Br J Pharmacol 1999, 126, (8), 1741-50.
- Grinchii, D.; Janáková Csatlósová, K.; Viñas-Noguera, M.; Dekhtiarenko, R.; Paliokha, R.; Lacinová, Ľ.; Dremencov, E.; Dubovický, M., Effects of pre-gestational exposure to the stressors and perinatal bupropion administration on the firing activity of serotonergic neurons and anxiety-like behavior in rats. Behav Brain Res 2024, 459, 114796.
- Dremencov, E.; Oravcova, H.; Grinchii, D.; Romanova, Z.; Dekhtiarenko, R.; Lacinova, L.; Jezova, D., Maternal treatment with a selective delta-opioid receptor agonist during gestation has a sex-specific pro-cognitive action in offspring: mechanisms involved. Front Pharmacol 2024, 15, 1357575. [CrossRef]
Figure 1.
Effect of acute administration of agmatine on the firing activity of 5-HT neurons. A: Representative recording from a selected DRN 5-HT neuron. B: Summary effect from 6 neurons from 6 rats. *p<0.05, Bonferroni post-hoc test. Data are presented as mean ± SEM.
Figure 1.
Effect of acute administration of agmatine on the firing activity of 5-HT neurons. A: Representative recording from a selected DRN 5-HT neuron. B: Summary effect from 6 neurons from 6 rats. *p<0.05, Bonferroni post-hoc test. Data are presented as mean ± SEM.
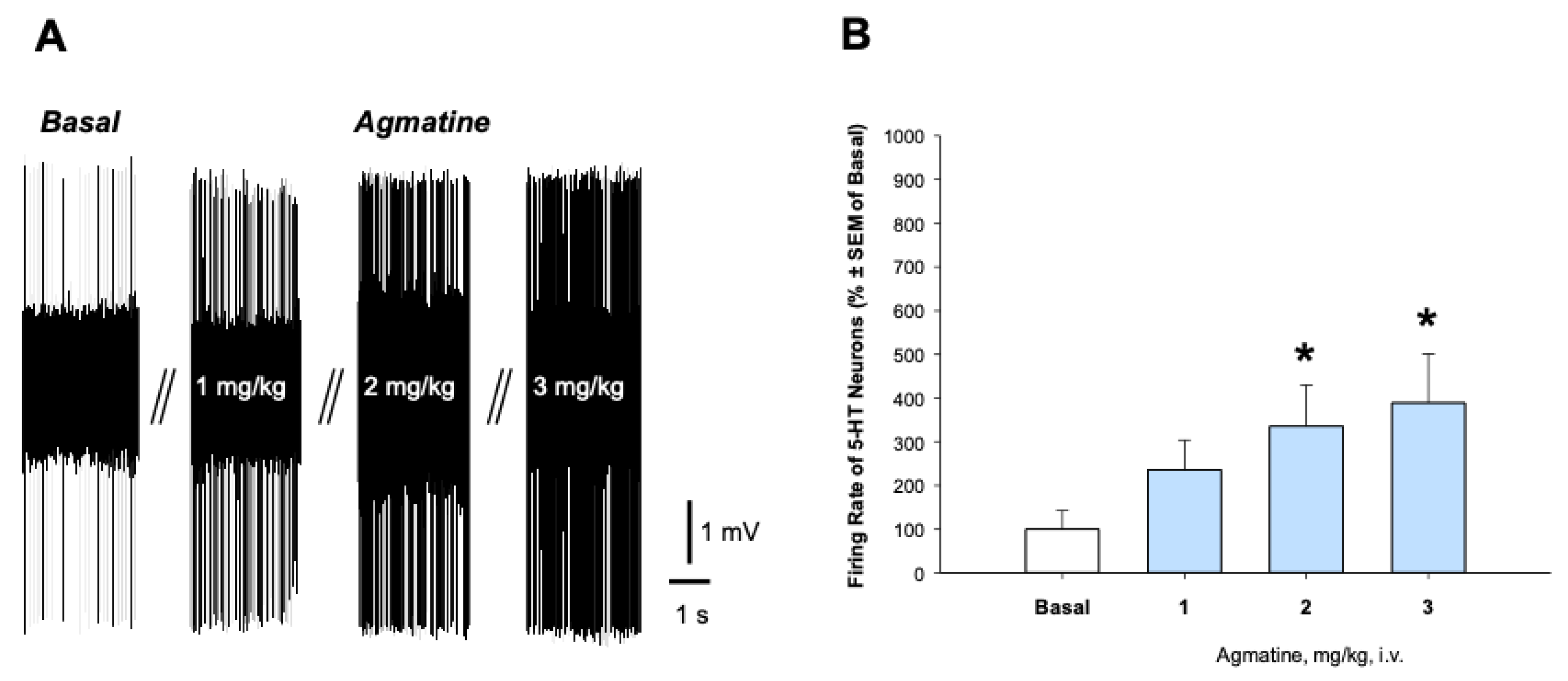
Figure 2.
Effect of chronic treatment with agmatine on the firing activity of 5-HT neurons. A: Representative recording from the selected DRN 5-HT neurons from the vehicle (up)- and agmatine (down)-treated rats. B: Summary effect from 75 neurons from 6 vehicle- and 100 neurons from 6 agmatine-treated rats. **p<0.01, two-tailed Student’s t-test. Data are presented as mean ± SEM.
Figure 2.
Effect of chronic treatment with agmatine on the firing activity of 5-HT neurons. A: Representative recording from the selected DRN 5-HT neurons from the vehicle (up)- and agmatine (down)-treated rats. B: Summary effect from 75 neurons from 6 vehicle- and 100 neurons from 6 agmatine-treated rats. **p<0.01, two-tailed Student’s t-test. Data are presented as mean ± SEM.
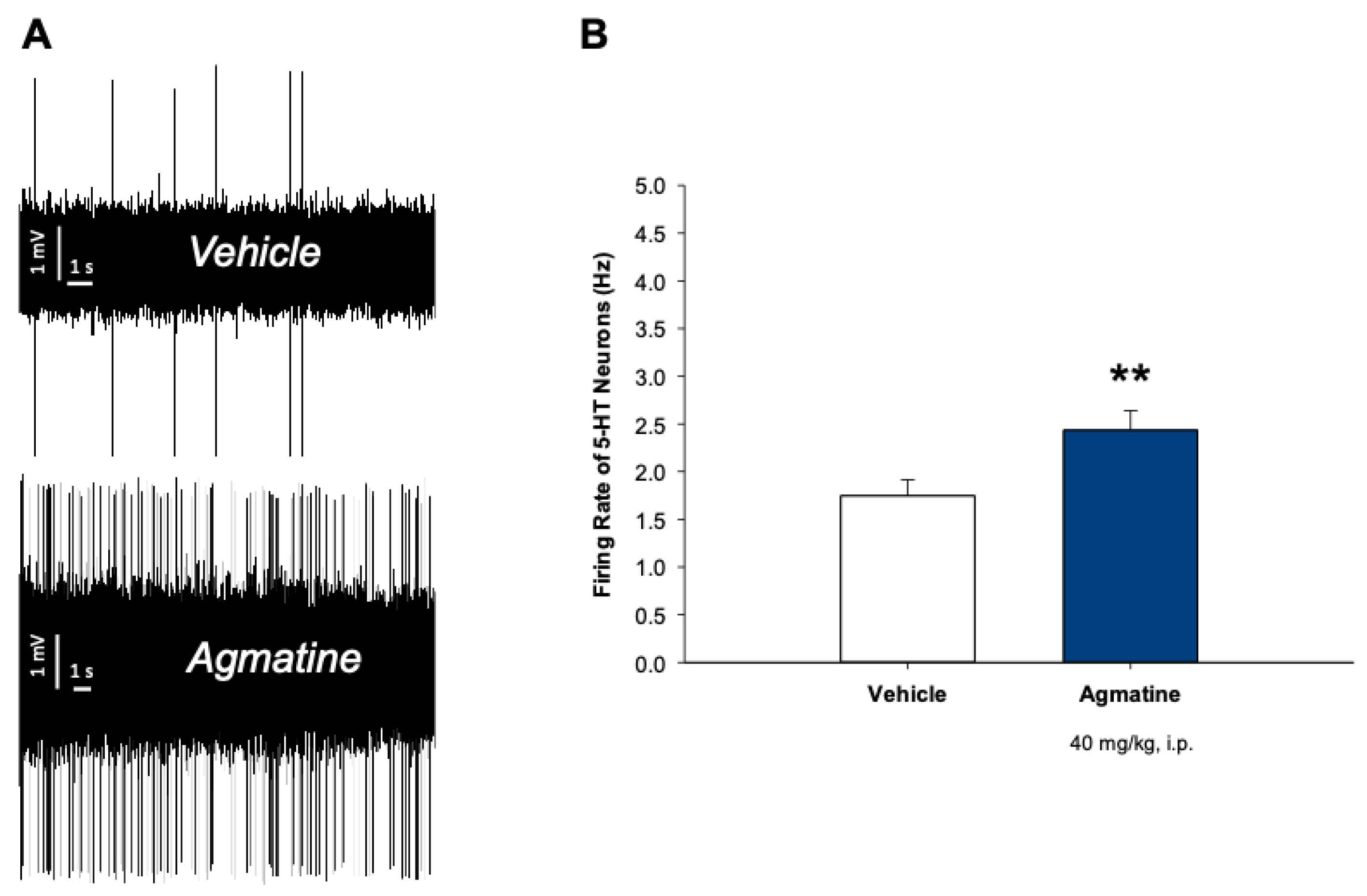
Figure 3.
Relative mRNA expression of nNOS and SERT in the DRN of vehicle- and agmatine-treated rats. Data are presented as mean ± SEM. (n=5 rats per group, two-tailed Student t-test; Mann-Whitney test, p>0.05).
Figure 3.
Relative mRNA expression of nNOS and SERT in the DRN of vehicle- and agmatine-treated rats. Data are presented as mean ± SEM. (n=5 rats per group, two-tailed Student t-test; Mann-Whitney test, p>0.05).
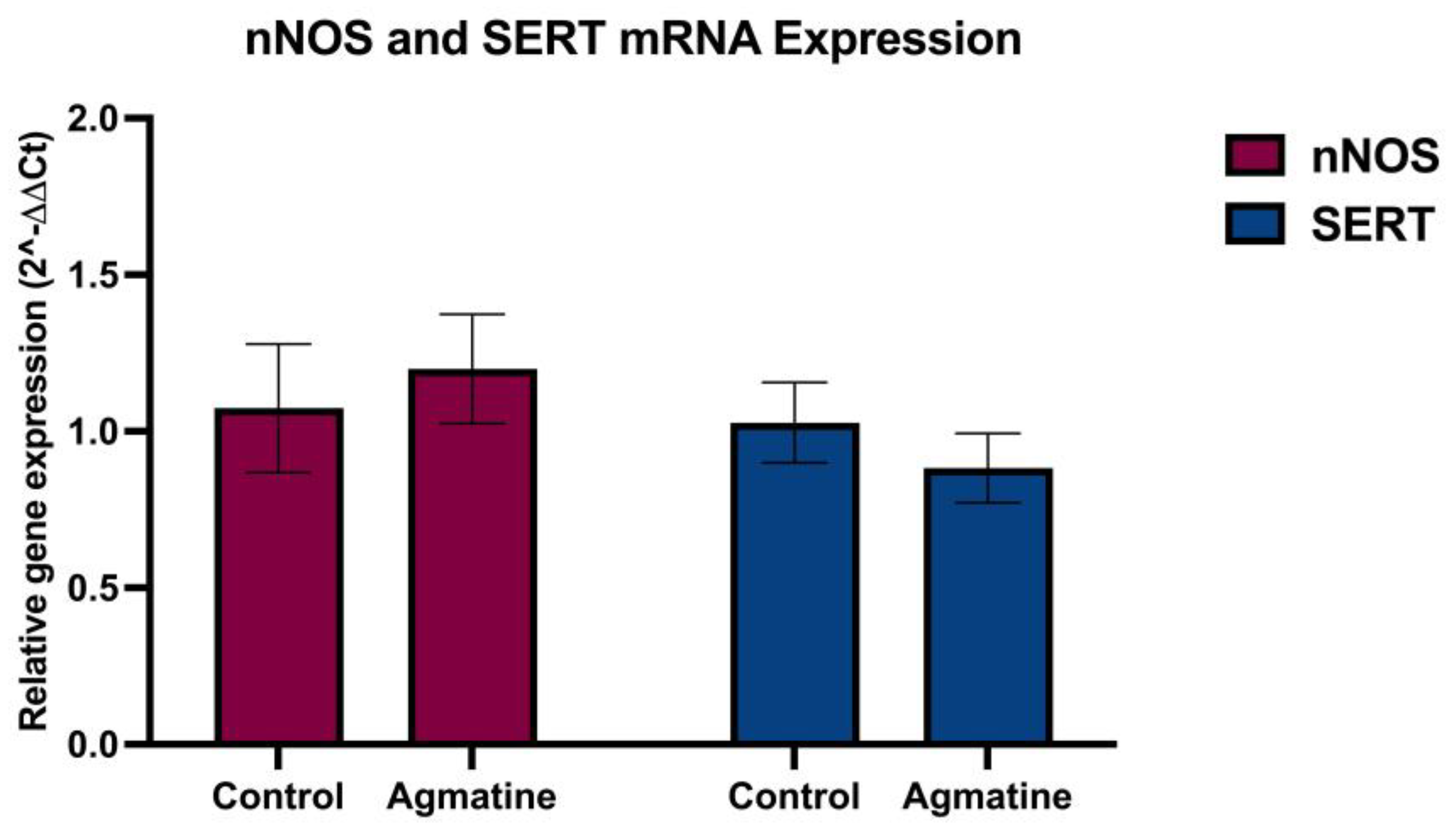
Figure 4.
Relative mRNA expression of 5-HT1A and 5-HT1B receptors in the DRN of vehicle- and agmatine-treated rats. Significant upregulation of 5-HT1B receptor mRNA expression was observed in agmatine-treated rats. Data are presented as mean ± SEM. (n=5 rats per group, two-tailed Student t-test; Mann-Whitney test, **p<0.01).
Figure 4.
Relative mRNA expression of 5-HT1A and 5-HT1B receptors in the DRN of vehicle- and agmatine-treated rats. Significant upregulation of 5-HT1B receptor mRNA expression was observed in agmatine-treated rats. Data are presented as mean ± SEM. (n=5 rats per group, two-tailed Student t-test; Mann-Whitney test, **p<0.01).
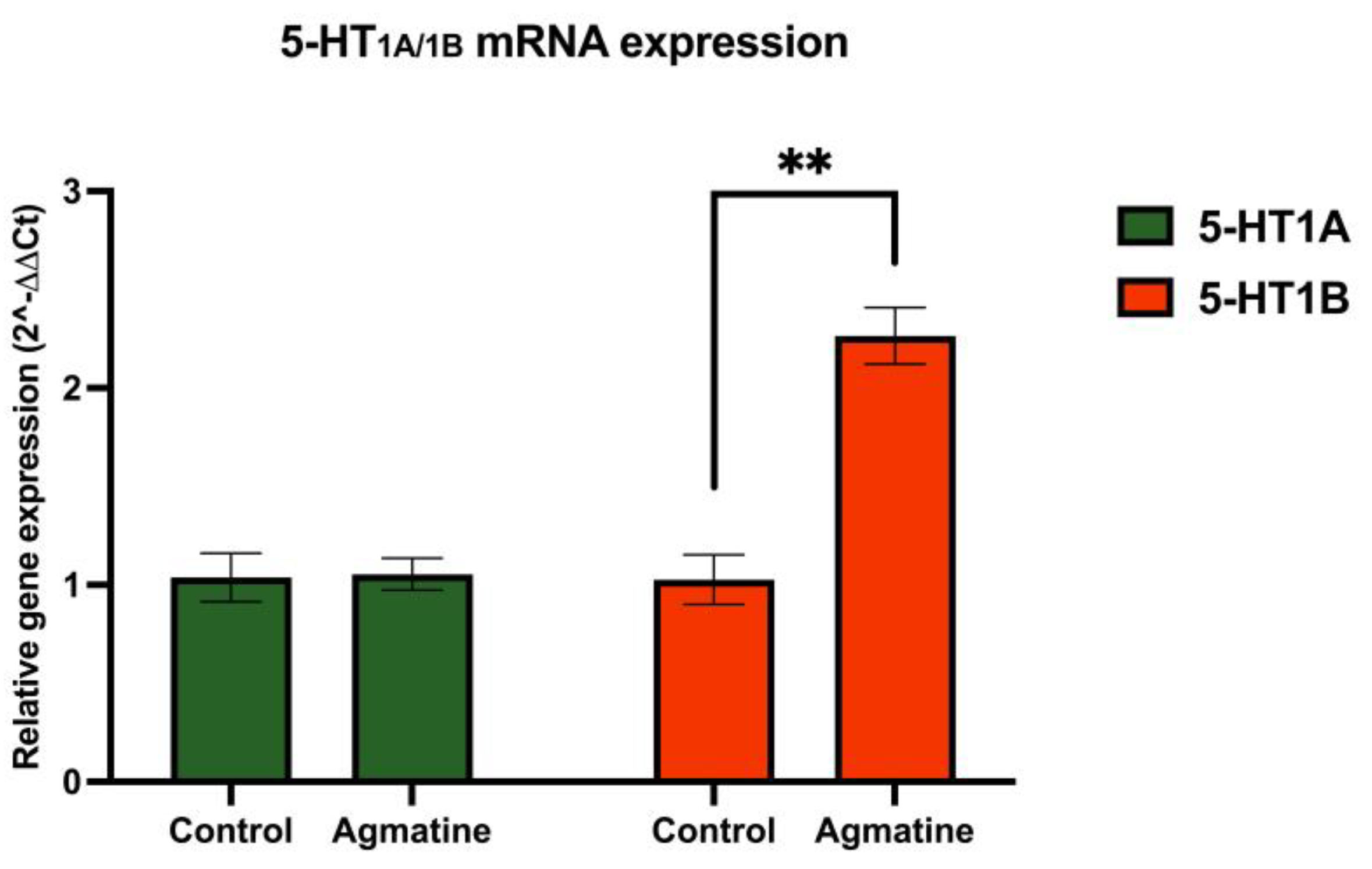
Figure 5.
Relative mRNA expression of 5-HT2A, 5-HT2B and 5-HT2C in the DRN. Significant upregulation of 5-HT2A receptor mRNA expression was observed in agmatine-treated rats. Data are presented as mean ± SEM. (n=5 rats per group, two-tailed Student t-test; Mann-Whitney test, *p<0.05).
Figure 5.
Relative mRNA expression of 5-HT2A, 5-HT2B and 5-HT2C in the DRN. Significant upregulation of 5-HT2A receptor mRNA expression was observed in agmatine-treated rats. Data are presented as mean ± SEM. (n=5 rats per group, two-tailed Student t-test; Mann-Whitney test, *p<0.05).
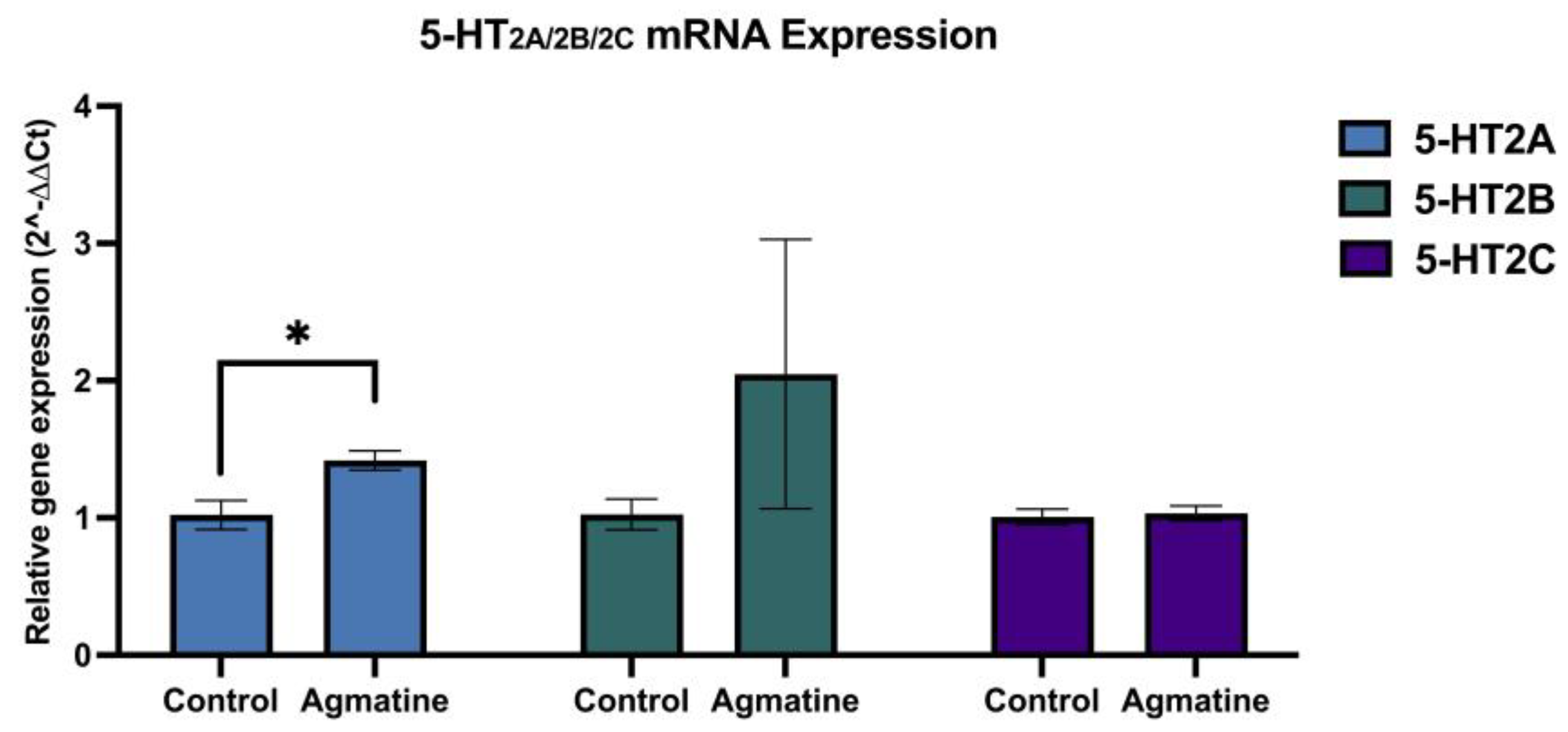

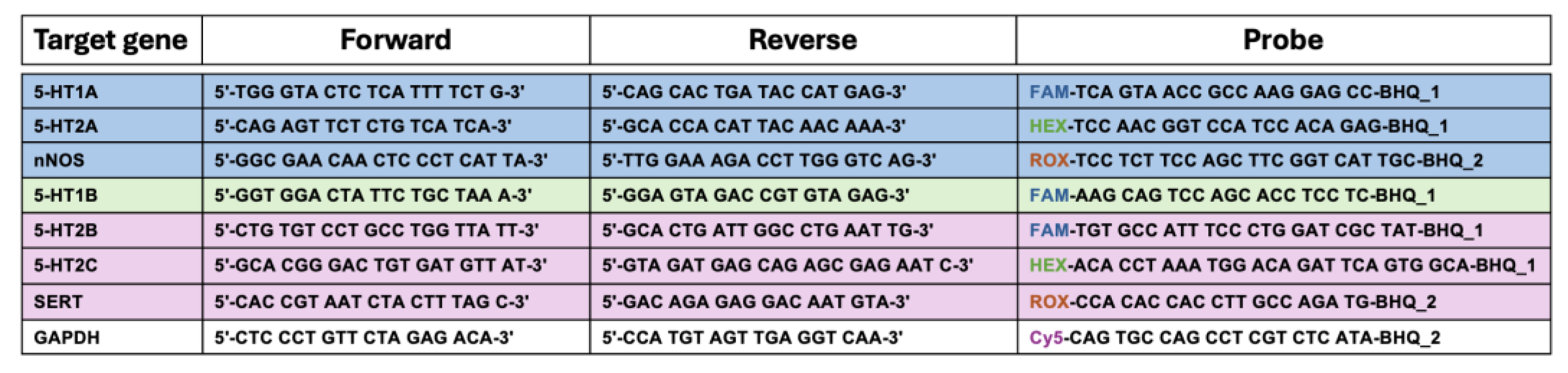
Table 1.
Forward and reverse primers and probe sequences used for RT-qPCR analysis of target genes.
 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



