1. Introduction
In many environments, the presence and movement of animals and humans can be revealed by their tracks and trails. Trails and tracks are the detectable signs of passage left by individuals or groups as they traverse the landscape. They can be transient or semi-permanent, depending on the substrate and frequency of use. Generally, tracks mark the passage of a single individual while trails denote repeated use. Their morphology depends on the substrate and frequency of use. Softer substrates and higher frequencies of use are expected to generate more distinct trails and tracks. Harder substrates and less-frequent passage would create less distinct features. Trails and tracks may be constructed purposefully to facilitate efficient movement (e.g., [
1]) or left behind by coincidental passage (e.g., [
2]).
Many ecological and wildlife-conservation applications require information on organism presence and movement, including habitat-selection studies (e.g., [3-4]), population-density assessments (e.g., [5-6]), and animal-movement models (e.g., [7-8]). Most contemporary researchers track wildlife through global navigation satellite systems (GNSS; e.g., [
9]), camera traps (e.g., [
10]), or genetics (e.g., [
11]). Each of these efforts could benefit from complementary information on trails and tracks.
Previous authors have pointed out the influence of trails on wildlife-detection probabilities in camera-trap studies. For example, Harmsen et al. [
12] reported that Neotropical mammals varied substantially in their use of existing trails. In the authors’ Belizean study area, Pumas (
Puma concolor) followed trails more closely than jaguars (
Panthera onca) in the same habitat type. As a result, density models of such species that fail to account for detection bias in cameras set on wildlife trails can produce misleading results. With reliable trail maps, researchers could not only parameterize their wildlife-density models more effectively, but also design camera-trap studies that take the existing trail network into account. This could help make efficient use of limited resources [
13].
Maps of human trails and tracks can also enhance our understanding and mitigation of anthropogenic disturbances on ecosystems. For example, pervious studies have demonstrated the negative effects of mechanical trails and tracks on soil characteristics [14-15], the distribution of invertebrates [
16], runoff patterns [
17], erosion [
18], and vegetation growth [
14,
19,
20]. Many of these effects are especially pronounced in boreal peatlands, where the disturbance of sensitive organic substrates can have far-reaching implications on soil physical and chemical properties [
21], plant community composition [
22], methane emissions [
23], and patterns of vegetation recovery [
24].
1.1. Mapping Trails and Tracks
Previous researchers have mapped trails and tracks directly through GNSS surveys (e.g., [25-26]) and volunteered geographic information (e.g., [27-28]), or indirectly through remote sensing. We concern ourselves here with the indirect remote-sensing approaches. On this subject, the literature is limited. The great majority of peer-reviewed research in this application domain has focused on mapping all-weather roads. Abdollahi et al. [
29] provides a recent review of this subject. Studies of this sort typically target wide, flat roads with distinct surface materials, making their methods less applicable to our task of detecting narrow, natural trails that blend with the surrounding environment. Therefore, while some techniques might be adapted, trail and track mapping applications differ from road detection.
There is a limited body of peer-reviewed research on the detection of trails and tracks in natural areas with remote sensing. Previous authors have analyzed the effects of concentrated livestock or wildlife activity on vegetation using spectral indices (e.g., [30-32]), but these works do not attempt to map trails and tracks directly. Welch et al. [
33] mapped 2,950 km of off-highway vehicle trails in Everglades National Park, Florida, using 1:40,000 color-infrared aerial photos. Kaiser et al. [
34] mapped smuggler-trail networks in the U.S.-Mexico border region of California using 60-cm four-band multispectral imagery. However, both studies relied extensively on heads-up digitizing. The Kaiser et al. work also evaluated image-enhancement and automated feature-extraction routines, but the authors’ best results relied on manual interpretation, feature delineation, and manual editing. Persistent noise, occlusions, and the variable nature of detectable mapping signals have largely prevented the use of automated workflows for mapping trails and tracks.
Fully automated strategies for detecting and mapping trails and tracks using modern datasets and processing workflows are just now beginning to emerge, powered largely by recent advances in artificial intelligence. Yamamoto et al. [
35] used convolutional neural networks (CNNs) to map dugong (
Dugong dugong) feeding trails from 0.47-cm resolution optical drone imagery in the intertidal seagrass beds of Trang Province, Thailand with F1 scores of up to 0.895. Bhatnagar et al. [
36] used a deep-learning image-segmentation algorithm on drone-based orthomosaics to map wheel-rut trails caused by mechanical harvesting machines in forest-harvest blocks in Norway with an average F1 score of 0.77.
There is a shortage of peer-reviewed research on the automated use of remote sensing to map trails and tracks in terrestrial environments. What literature currently exists relies on manual editing [
34] or deals only with mechanical ruts [
36]. The potential of CNNs and other forms of artificial intelligence to map trails and tracks over large terrestrial areas using high-resolution airborne datasets is largely untapped.
We are particularly interested in the capacity of high-density LiDAR (light detection and ranging) data to detect trails and tracks. While optical imagery also provides a promising information source, these data have limited canopy penetration abilities and are subject to frequent shadows in vegetated terrain. Other authors have recently begun using LiDAR to map drainage ditches (e.g., [37-40]). Whether these types of morphometric imprints could also be used to map trails and tracks more broadly has yet to be assessed.
1.2. Research Objectives
Our goal is to create tools and processing workflows for mapping trails and tracks automatically over large terrestrial areas using remote sensing. To help reach this goal, we established three research objectives, which are the subject of this paper:
To demonstrate the capacity of high-density LiDAR and CNNs to map trails and tracks automatically in a natural environment;
To compare the accuracy of trail/track maps developed with LiDAR from a drone platform (185 points/m2) and a piloted aircraft platform (30 points/m2); and
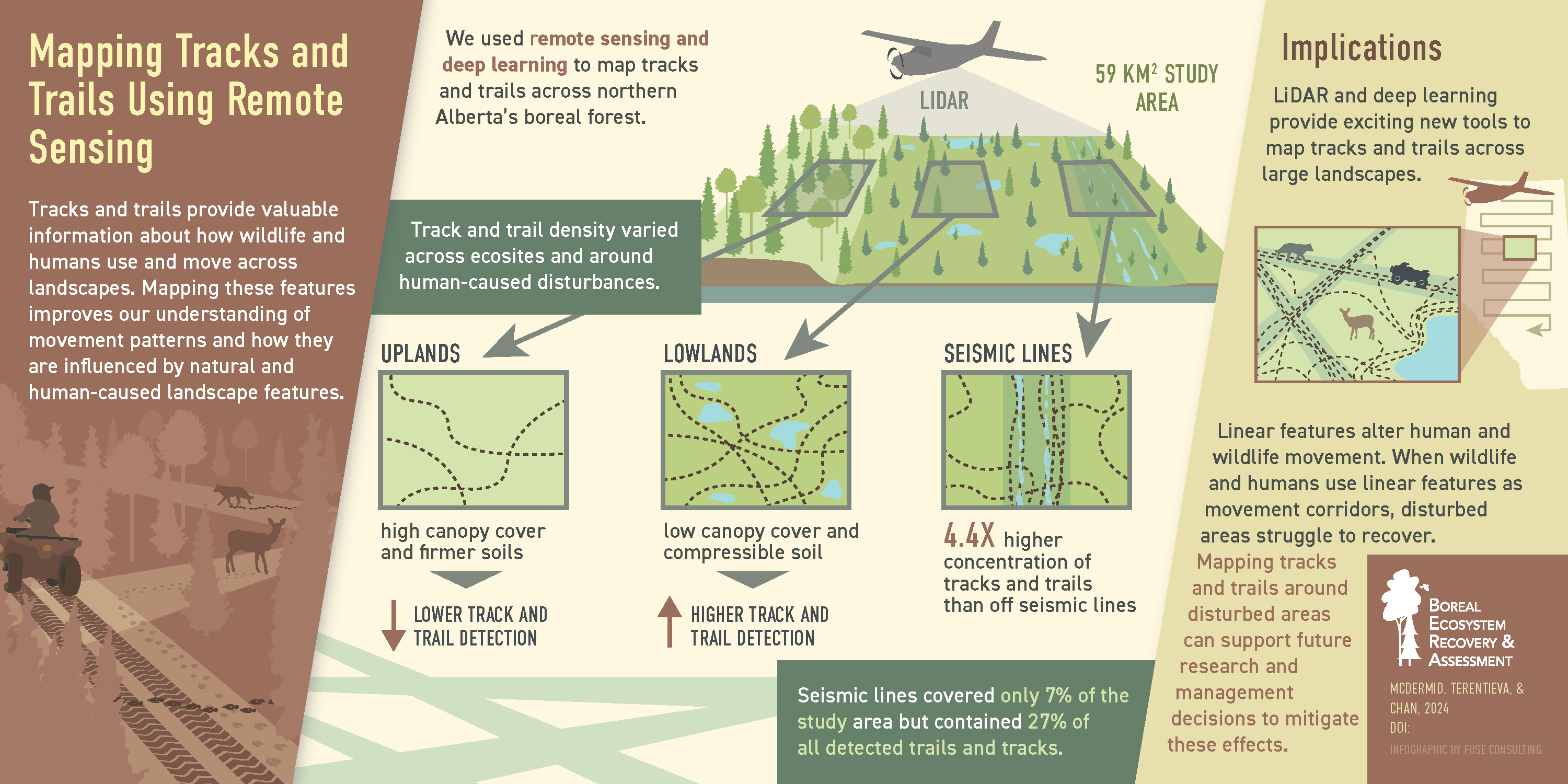
To measure the abundance and distribution of tracks and trails across different land-cover classes, and their co-location with anthropogenic disturbances across our study area in the Canadian boreal forest.
To our knowledge, this is the first study on the use of LiDAR and CNNs for mapping terrestrial trails and tracks to appear in the peer-reviewed literature. Our research demonstrates that not only can trails and tracks be detected accurately under the correct conditions, but that high-quality maps can be obtained over large areas using data from piloted-aircraft. While our work is a case study, our findings reveal the potential for these workflows across a variety of exciting applications.
2. Materials and Methods
2.1. Study Area
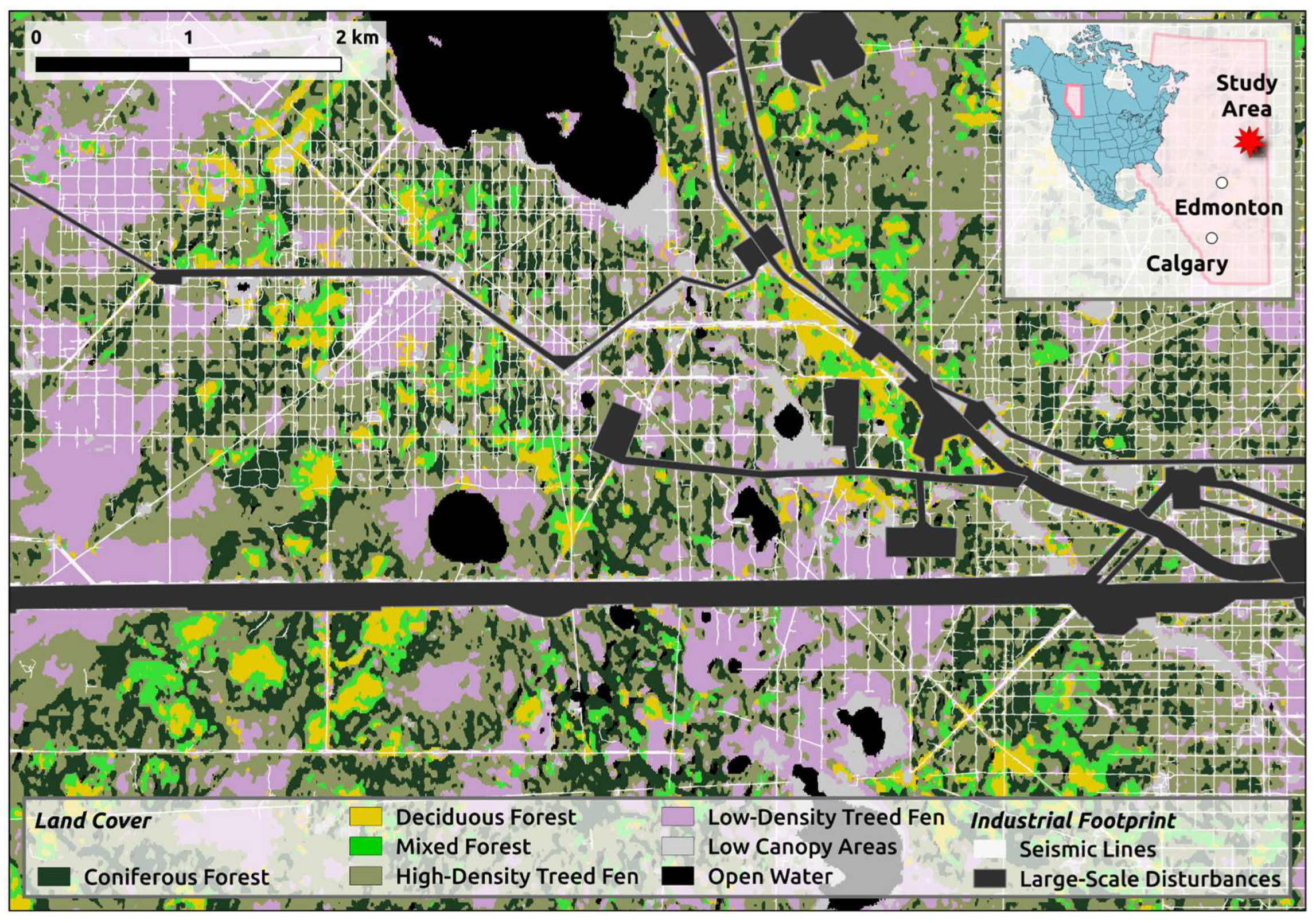
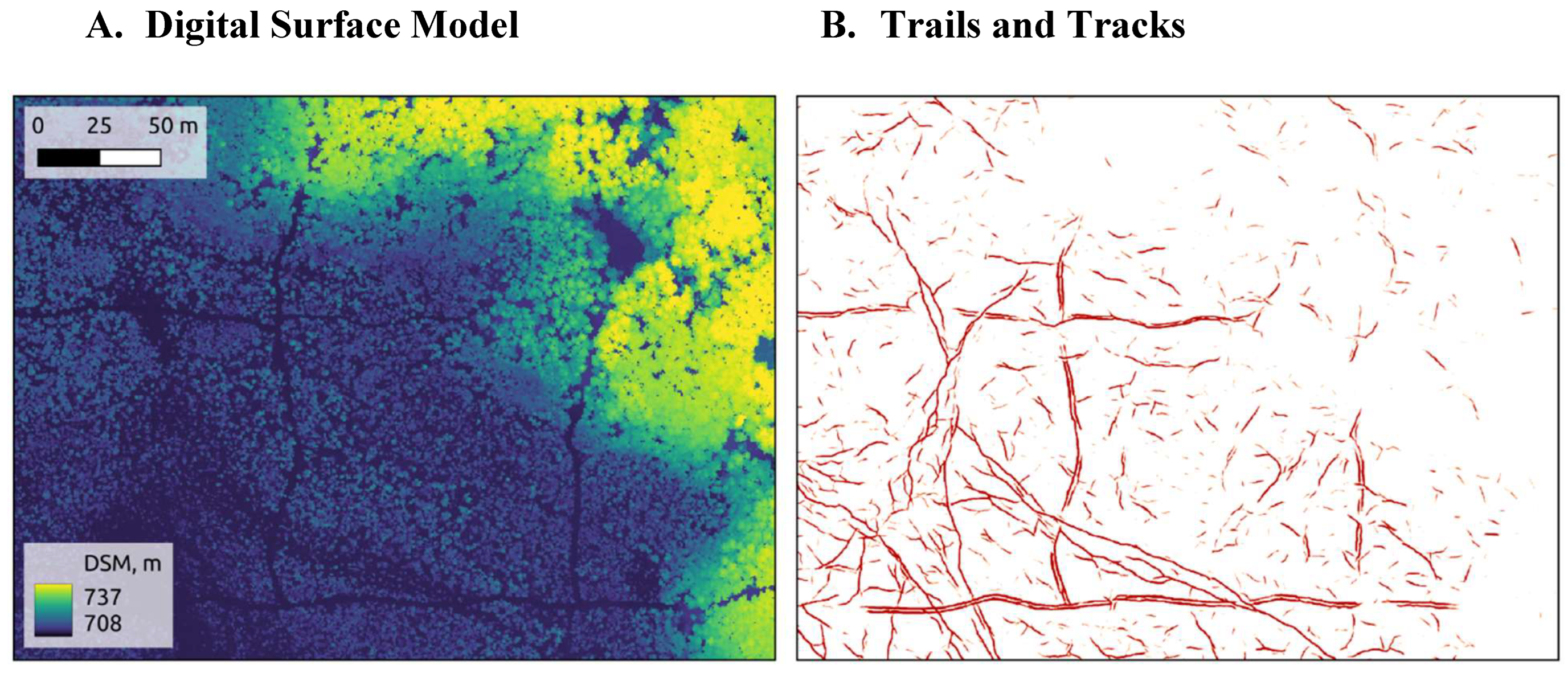
Our study area is in the boreal forest of northern-eastern Alberta, Canada (
Figure 1) at a site approximately 150 km south of Fort McMurray. The study area is in the central mixedwood natural subregion, located 500-600 meters above sea level. The central mixedwood subregion has a continental subarctic climate characterized by cold winters and short, warm summers. Precipitation is moderate, with an average annual rainfall of around 400 to 600 mm [
41].
Our 59-km
2 study area contains a diverse mosaic of land-cover types, from sparsely treed peatlands to thickly treed uplands (
Figure 1). Peatlands – wetlands with accumulated organic materials [
42] – cover 58% of the study area. Another 27% of the area is composed of uplands and transitional areas. This blend is typical of the central mixedwood parts of the boreal forest. Conifer trees dominate the peatland and transitional portions of the study area. Coniferous, deciduous, and mixedwood stands can be found on the upland portions. The remaining 15% of the area is covered by open water, roads, and large-scale industrial disturbances. We excluded these areas from most analyses.
There are three types of trails and tracks common to our study area: wildlife trails, off-highway vehicle (OHV) tracks, and seismic lines (
Figure 2). Wildlife trails at our site are created primarily by the movements of ungulates like woodland caribou (
Rangifer tarandus caribou) and white-tailed deer (
Odocoileus virginianus). OHV tracks are wheel ruts created by off-road vehicles. Seismic lines are linear corridors left behind by mulchers and other heavy machinery used to cut clearings for subsurface petroleum exploration [
43]. There are significant morphological differences between the three types. Seismic lines (
Figure 2c) are 4- to 6-meter-wide mechanical cuts that have been installed in a systematic grid, with a spacing of about 100 meters. Wildlife trails (
Figure 2a) are typically up to a meter wide and can form dense, intertwining networks. OHV tracks (
Figure 2b) are like wildlife trails, but normally appear as paired depressions created by OHV wheels. The models we developed for this study are aimed at wildlife trails and OHV trails and tracks. We did not target seismic lines in this work.
While there are substantial morphological differences between wildlife trails, OHV trails and tracks, and seismic lines in our study area, they are not always physically separated from one another. In fact, OHVs and wildlife often adopt seismic lines into their transportation networks. Quantifying the amount of wildlife- and OHV-adopted seismic lines that exist in our study area was one of our research objectives.
Data Acquisition and Processing
Aerial LiDAR was provided by Alberta-Pacific Forest Industries, Inc. through partnership in the Boreal Ecosystem Recovery and Assessment (BERA) project (
www.bera-project.org). The LiDAR was collected in the summer of 2022 with a Riegl VQ-1560ii system. The sensor was flown on a piloted aircraft at approximately 1800 m above ground level with a ± 60° maximum scan angle, 825 kHz pulse repetition frequency, and 50% swath side lap. The final point cloud had a density of 30 points/m
2. The vendor, Airborne Imaging, calibrated the dataset using precise point positioning (PPP) techniques to model and remove GNSS system errors. The horizontal and vertical accuracy were reported as 0.35 m and 0.20 m, respectively. Point-cloud classification was also performed by the vendor. We created 50-cm digital terrain models (DTMs) from classified ground points using the
las2dem tool in LAStools.
Drone data were collected for a subsection of our study area in the summer of 2022 using a Zenmuse L1 LiDAR sensor aboard a DJI Matrice 300 RTK. The sensor was flown approximately 100 m above ground level with triple-return mode and repetitive scanning enabled at a sampling rate of 160 kHz. The L1 sensor uses a near-infrared laser (905 nm), and the final point cloud had an average density of 185 points/m2. The drone had RTK (real-time kinematic) positioning enabled and was connected to a DJI D-RTK 2 GNSS base station. We used PPP to obtain the final position of the base station, then calculated the difference between the final and initial base station positions. This difference was then applied to shift the point cloud into a corrected position.
To evaluate the drone LiDAR’s vertical accuracy, we conducted an RTK-GNSS survey using six independent checkpoint targets. These checkpoints were 1 x 1-meter plastic targets that were laid out flat across the study area prior to drone flights. The targets coincided with 377 LiDAR points. The mean absolute error between the check point targets’ GNSS-surveyed elevation and the drone’s LiDAR-surveyed elevation was 3 cm.
We performed a variety of standard post-processing steps (e.g., noise removal, ground classification) on the drone-based point cloud using LAStools. We then generated a 10-cm DTM from the classified ground points using a k-nearest neighbor inverse-distance weighted algorithm (knnidw; k = 10, p = 2) in the R package lidR [44-45].
In the summer of 2023, we collected sub-centimeter resolution optical dataset over a subsection of our study area to provide reference data for independent testing. This subsection was a 0.25 km
2 portion of a low-density treed fen labeled
Testing Area in
Figure 3. These data were acquired using a DJI Zenmuse P1 sensor (full-frame 45-megapixel camera with a 35 mm lens) on the same drone platform that was used to collect the LiDAR. We flew 35 m above ground level with RTK positioning enabled. The side and front overlap was set to 70% and 80%, respectively. This imagery was processed into an ultra-high resolution (0.5 cm) true-color orthomosaic using PIX4Dmapper. Like the drone LiDAR processing, the orthomosaic was shifted based on the difference between the initial and final base station positions, following PPP processing. A GNSS survey of 20 independent check points was used to assess mean absolute error (0.11 m) and root mean square error (0.13 m) of the orthomosaic position.
We assume accurate coregistration between the three remote-sensing datasets used in this study: drone LiDAR, piloted-aircraft LiDAR, and drone orthomosaic. All three were captured following rigorous survey practices and produced very high spatial accuracy statistics. We further assume that there are no substantial differences in the pattern of trails and tracks within our remote study area between the summer of 2022, when the LiDAR data were flown, and the summer of 2023, when the optical data were collected. The LiDAR data were used to build the trail/track models, while the optical data were used for accuracy assessment. We contend that any differences, should they exist, would not serve to bias one LiDAR dataset over the other and would therefore have no material effect on our experiments.
Mapping Trails and Tracks
2.3.1. Training Data Preparation
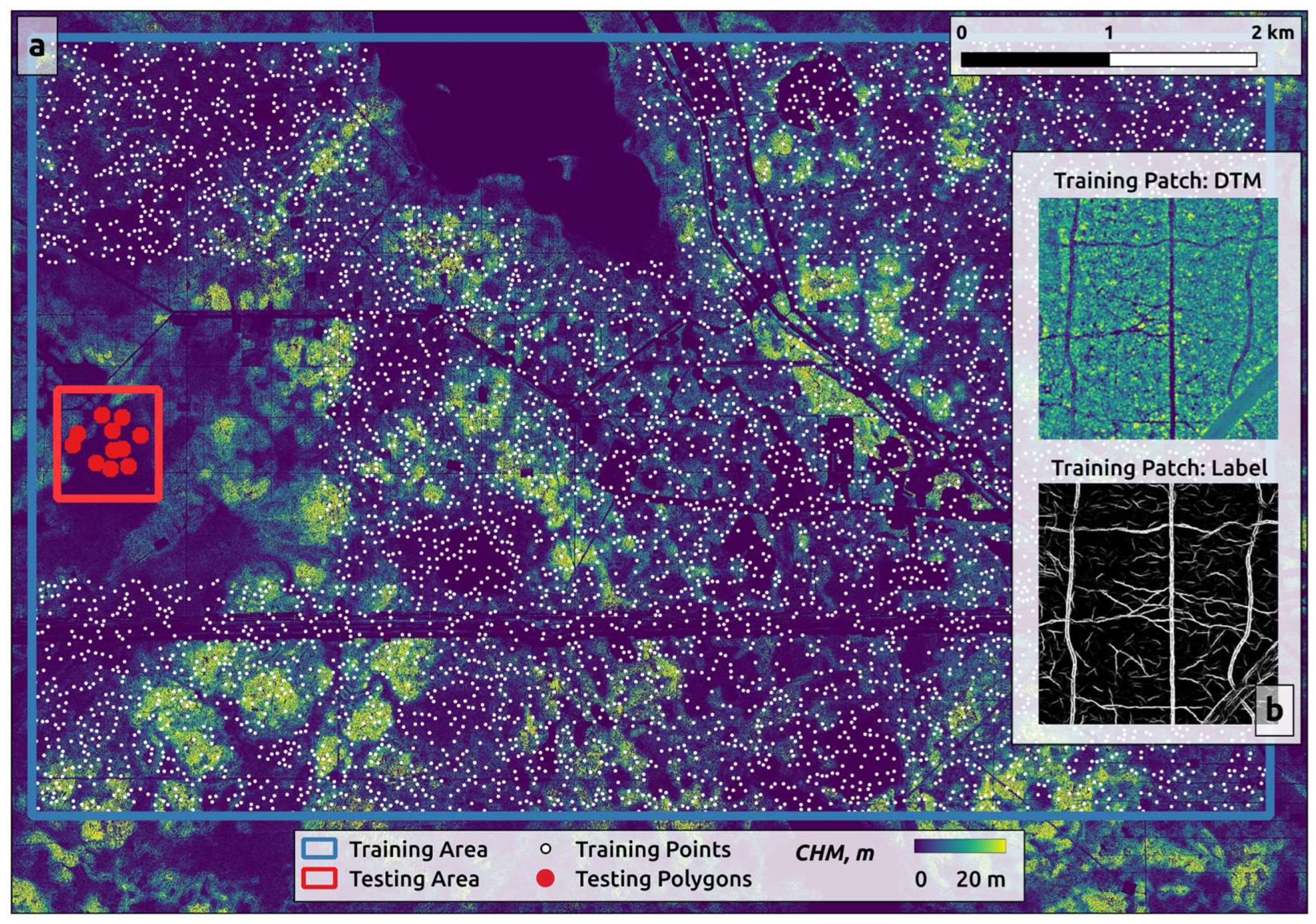
Our approach to training-data preparation and labeling involved a multi-stage strategy. First, we used visual interpretation of aerial LiDAR data to manually label a preliminary training dataset. This data was used to train an initial U-Net model. We then used the output of this initial model as input for training a second, more refined set of U-Net models: one for the 10-cm DTM and a second for the 50-cm DTM.
The main training set consisted of 6,395 training patches cropped around random points distributed across the training portion of our study area, which covered 45 km
2 (
Figure 3a). Each patch was 512 x 512 pixels in size and contained both DTM data and pixel-wise binary labels (
Figure 3b). To improve the segmentation quality at this second stage, we removed objects smaller than 30 pixels from the binary masks. Finally, we thinned the segmented features (trails and tracks) to a skeletal form, then buffering them back to 1-pixel width for 50-cm DTM, and 3-pixel width for 10-cm DTM. This process produced trails and tracks with standardized widths in the binary masks.
We used several augmentation techniques to increase the variability of our training dataset. First, training images were randomly rotated in increments of 90 degrees. Additionally, images were subject to random flipping. Moreover, the brightness and contrast of the images were adjusted randomly, with brightness changes up to 0.1 and contrast variations between 0.9 and 1.1.
2.3.2. U-Net
We employed a U-Net model [
46] using the Keras unet library. U-Net is a convolutional neural network renowned for its efficiency in image-segmentation tasks. The architecture uses concatenation and skip connections, which allows the network to use information from both deep, coarse layers and shallow, fine layers to improve the accuracy of segmentation. Our custom U-Net model includes an attention-gate function, incorporating an attention mechanism inspired by the Attention U-Net [
47]. This mechanism automatically learns to focus on relevant regions of the input image, suppressing irrelevant areas and highlighting salient features. This approach can improve segmentation accuracy by enhancing model sensitivity to important structures while maintaining computational efficiency. The implementation of the Attention U-Net was adapted from the open-source repository available at
https://github.com/karolzak/keras-unet.
Our U-Net architecture was configured to accept one-band input images 512 x 512 pixels in size. The initial number of filters was set to 32. A dropout rate of 0.3 was implemented to prevent overfitting. Dropout randomly ignores a subset of neurons during training to increase the model's generalization ability. Since the network was designed for binary segmentations, it used the sigmoid-activation function in the final layer for the classification tasks.
We used the Adam optimizer [
48]: a popular choice for deep learning applications due to its efficiency in handling sparse gradients on noisy problems. The Adam optimizer adjusts the weights of the CNN each step during training and is designed to improve convergence. The model was compiled with a binary cross-entropy loss function, suitable for binary classification tasks. To prevent overfitting and assist in escaping training plateaus, we implemented early stopping, ceasing training after one epoch without improvement in training loss. We also applied learning-rate reduction, halving the rate if no improvement in validation loss occurred over three epochs. The model was trained with a batch size of 2 due to computational resource constraints.
The CNN outputs a probability mask. To compare drone- and aerial platforms, these masks were thresholded at different values to generate a precision-recall curve and calculate accuracy metrics. To assess a trail distribution across the entire area, we chose a conservative threshold of 40%. Then, to calculate trail lengths, we converted the resulting binary mask into a vectorized product using the Zhang-Suen thinning algorithm [
49].
2.3.3. Accuracy Assessment
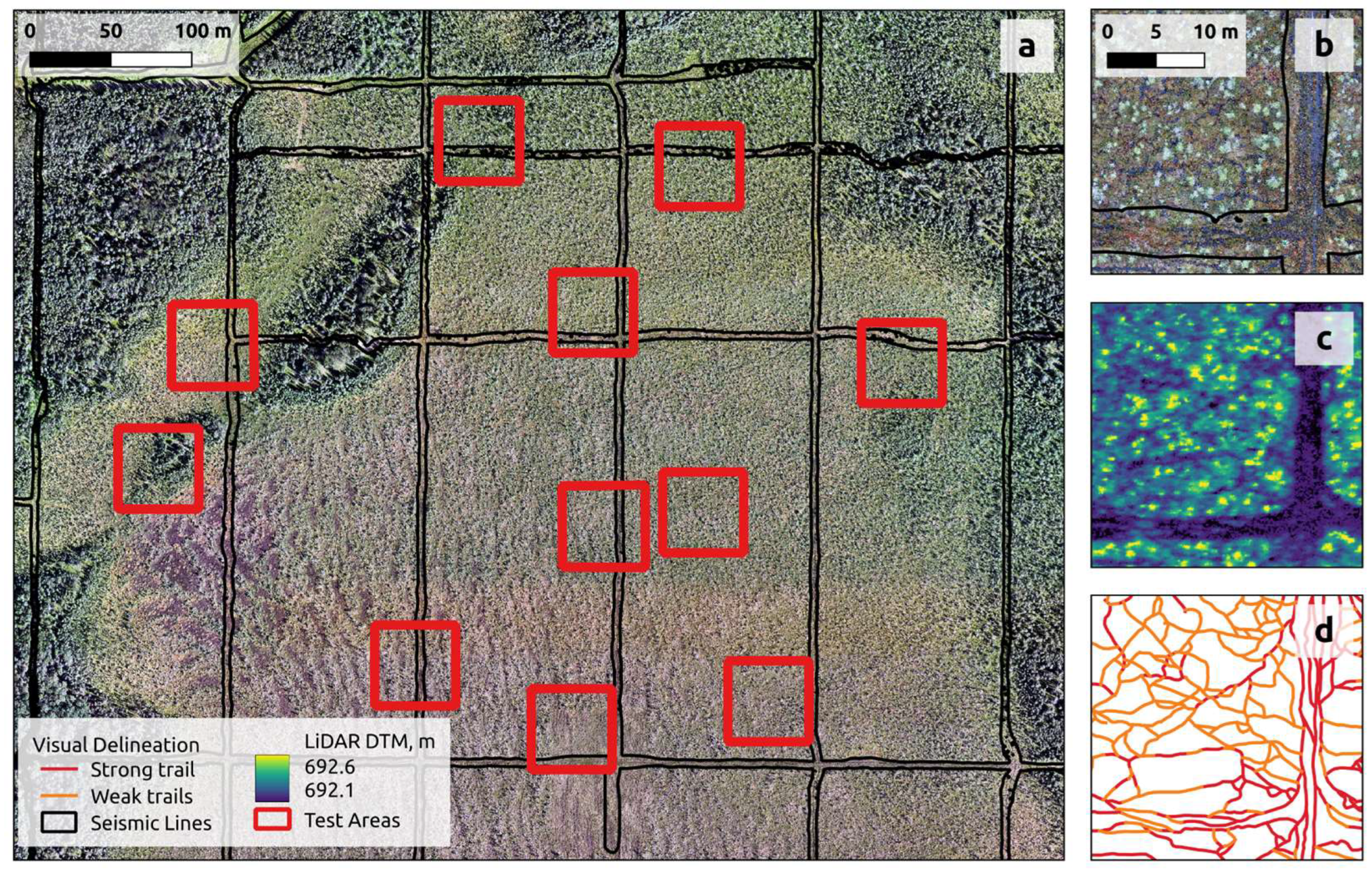
To demonstrate the capacity to map trails and tracks (Objective 1), we developed two U-Net models: one for the piloted-aircraft data and a second for the drone data. To compare the performance of these models on drone- and aerial-based platforms (Objective 2), we assessed the accuracy of the two map outputs. To accomplish this accuracy assessment, we created a census of all the trails and tracks located within 11 50 x 50-meter test squares distributed randomly across the testing portion of our study area (
Figure 4). Once again, the testing area is physically separate from the training area (
Figure 3). All test squares were located at least 500m away from any training patch.
The censuses of trails and tracks within the 11 test squares were developed using visual interpretation of the ultra-high resolution (0.5 cm) drone orthomosaic. Test squares were mapped by two different photo interpreters: both with subject-matter expertise and familiarity with the study area. The photo interpreters used a systematic photo-interpretation key (Figure S1) to map all the trails and tracks within the 11 test squares. The two photo-interpreted maps were combined by the lead author to create our final accuracy assessment reference.
We assessed the accuracy of our four U-Net trail predictions by comparing them to our photo-interpreted trail census. To create accuracy statistics, we generated 6,142 random points within our 11 test squares and distributed them across two strata: trails (2,842 points), and no trails (3,300 points). All non-trail points were located at least 1 meter away from existing trails.
A predicted trail point was considered a true positive if it occupied more than 25% of a 2x2 pixel area around a reference trail point. Average and standard deviation of precision and recall were calculated across the 11 test squares. We measured accuracy using confusion matrices, F1 statistics, and precision-recall curves.
Land Cover and Seismic Line Maps
To estimate the abundance and distribution of trails (Objective 3), we required supplementary maps of land cover and seismic line areas within our study area. We mapped seismic lines with the Forest Line Mapper [
50]: a semi-automated software tool for mapping linear features in a forest environment using LiDAR-derived canopy height models. The output of the Forest Line Mapper is a polygon feature-class layer depicting the precise extent of the ~4-meter-wide seismic lines that dissect our study area. This output can be seen in figures throughout the manuscript.
We opted to create our own landcover map (
Figure 1) to assess the distribution of trails across land-cover types. The map was developed in Google Earth Engine using a combination of satellite imagery and machine-learning classification techniques. The primary data sources were Sentinel-2 (S2) and Sentinel-1 (S1) satellite imagery. The S2 imagery was processed by selecting specific spectral bands (B3, B4, B8, B11) from images captured between 2015 and 2022. We generated seasonal composites (spring and summer) using median aggregation to capture diverse temporal aspects of the landscape. S1 radar data, utilizing the 'VV' polarization, was integrated into our analysis, combining data from both ascending and descending passes. The processed S2 and S1 data were then combined into a single 9-band image, leveraging both the spectral and radar features to enhance classification accuracy.
Labels for the classifier were based on distinguishing upland from lowland areas, as well as varying canopy properties, including low and high-density treed fens, deciduous stands, coniferous stands, and their mixtures in upland areas. We selected classes that cover substantial areas within the study region. A random forest classifier was trained using this labeled dataset. The classifier was configured with 200 trees, a minimum leaf population of 2, and a bag fraction of 0.5. The code is available in supplemental materials.
3. Results
3.1. Mapping Trails and Tracks
A five-meter resolution density map of all the trails and tracks detected across our 59-km
2 study area is shown in
Figure 5. A line-feature class database of the detected trails and tracks is provided in the supplementary materials. The map presented here was generated from the 50-cm DTM derived from the 30-points/m
2 piloted-aircraft data. The map contains 2,829 km of undifferentiated trails and tracks across all six land-cover classes. The accuracy of the map, its comparison to the alternative product generated from the 10-cm drone DTM, and an assessment of the distribution of trails and tracks across land cover types and anthropogenic disturbance features follows.
3.2. Accuracy of Piloted Aircraft- and Drone-Based Models
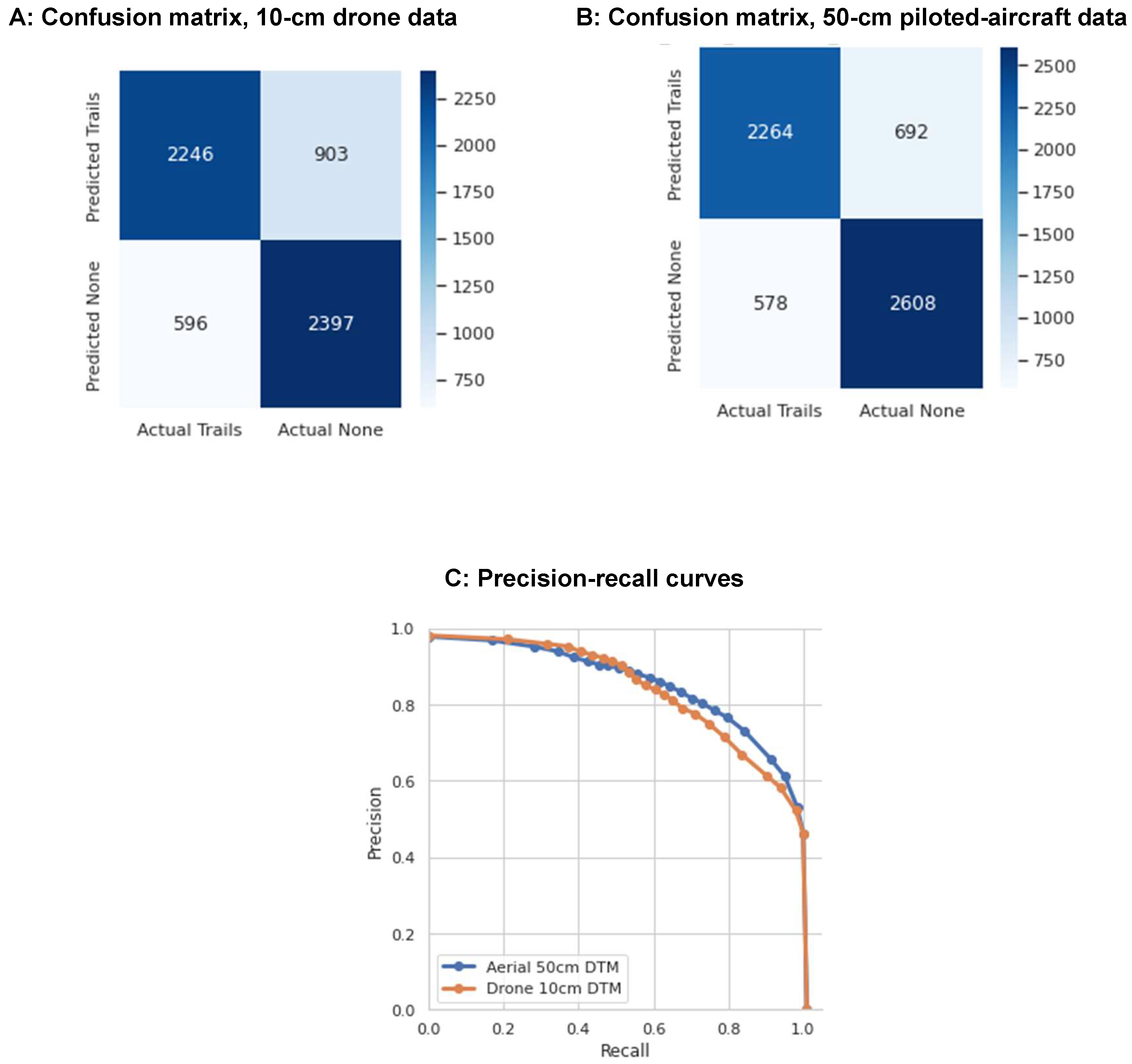
Both sets of CNNs demonstrated good performance in mapping trails and tracks over the testing portion of our study area (
Table 1). Surprisingly, there was no statistical difference in the overall accuracies of the piloted aircraft- (F1 = 77 ± 9%) and drone-based (F1 = 74 ± 6%) model products. The 50-cm piloted-aircraft data had slightly higher average precision (0.69) rates compared to the drone data (0.64). Both the confusion matrices and precision-recall curves were similar (
Figure 6), though the drone data produced a slightly higher rate of false positives. The precision-recall curves show that both models can detect virtually all the trails and tracks in the test squares, at the expense of a high rate of false positives. Alternatively, a lower-probability threshold would map about half of the existing trails and tracks in the test squares with virtually no false positives.
3.3. Distribution of Trails and Tracks Across Land-Cover Classes
Table 2 summarizes the distribution of the 2,829 km of trails and tracks mapped across the five land-cover classes present within our study area. This output was generated using a probability threshold of 40. Detected trail segments shorter than 1 m in length were filtered from the output.
Overall, we detected more trails and tracks in peatlands (2,320 km) than in upland (509 km) land-cover types. Low-density treed fens cover 18% of our study area but contain 33% (978 km) of all detected trails and tracks. High-density treed fens cover 41% of our study area and contain 47% (1,342 km) of all detected features. The three upland land-cover classes – coniferous forests (17% of our study area), deciduous forests (5%), and mixed forests (5%) – contain 14% (396 km), 2% (62 km), and 2% (51 km) of all detected trails and tracks, respectively.
The 98 km/km
2 density of trails and tracks mapped in low-density treed fens is 75% higher than that of the second-highest density land-cover type – high-density treed fens (56 km/km
2) – and more than double that of any other land-cover type (
Table 2). In uplands, the density of trails and tracks ranged from 17 km/km
2 (mixed forest) to 41 km/km
2 (coniferous forest).
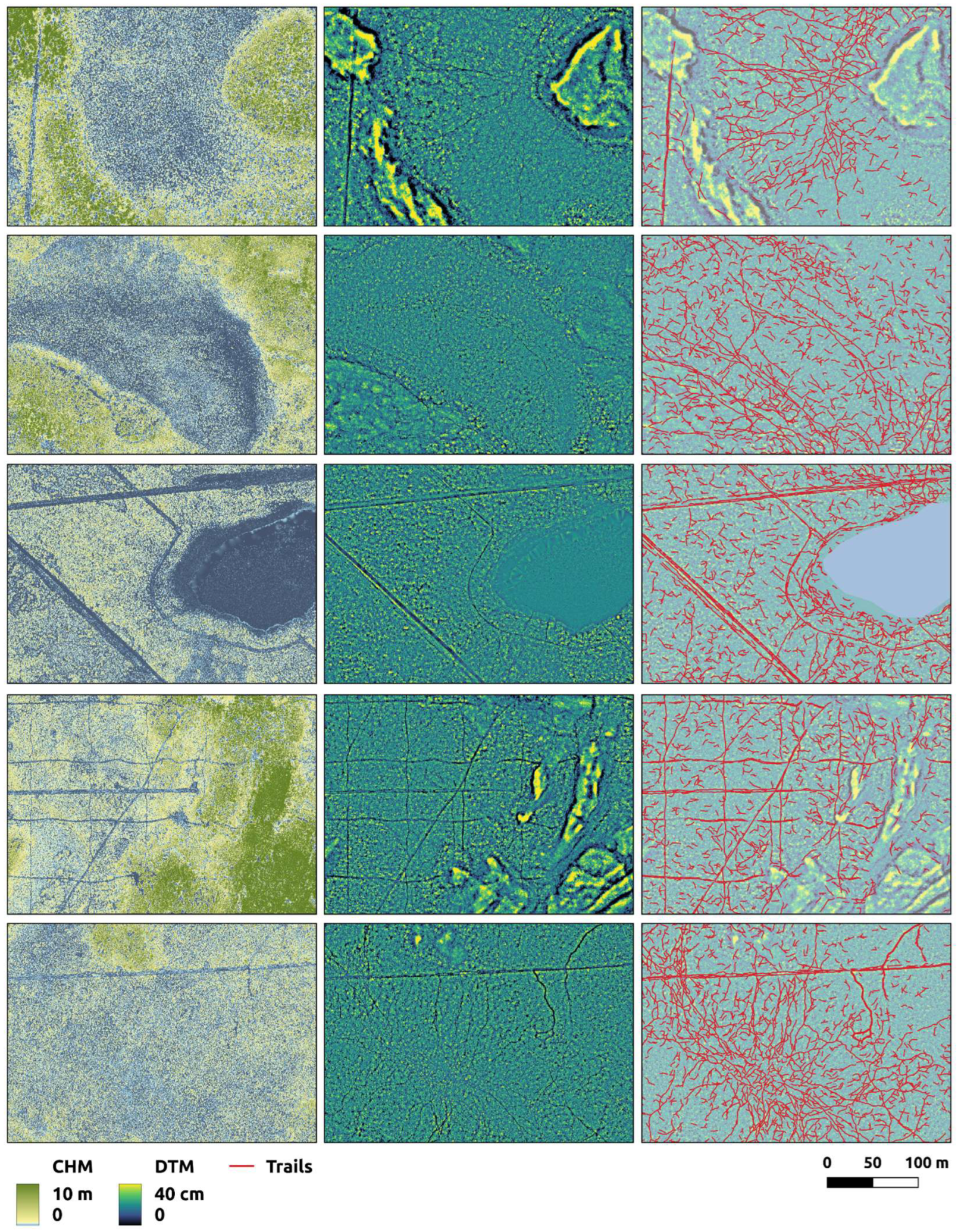
Our capacity to map trails and tracks using the present workflow is clearly influenced by the amount of overstory vegetation present. Trails and tracks mapped in the thickly forested upland portions of our study area are sparser and more fragmented than those detected in the more open peatland areas (
Figure 7).
3.4. Seismic Line Influence on Trails and Tracks
The density of trails and tracks detected on seismic lines (182 km/km2) is 4.4-times higher than that detected off seismic lines (41 km/km2;
Table 3). While seismic-line disturbances cover just 7% of our study area, they contain 27% of all detected trails and tracks.
4. Discussion
4.1. Boreal Trails and Tracks Can Be Mapped with LiDAR and Convolutional Neural Networks
Our work shows that trails and tracks – the detectable signs of passage of wildlife or OHVs – can be successfully mapped across a variety of land-cover types in the boreal forest using LiDAR and CNNs. Our piloted-aircraft model delivered an F1 score (the harmonic mean of precision and recall) of 77 ± 9% in the spatially distinct testing portion of our 59 km2 study area. To our knowledge, this is the first demonstration of fully automated terrestrial trail and track mapping to be published in the peer-reviewed remote-sensing literature.
It is important to note that the testing portion of our study area used to generate our accuracy statistics is a low-density treed fen. This area was chosen due to the straightforward detection of trails both with LiDAR and visual interpretation, making it ideal for testing this proof of concept. We would not expect to achieve the same levels of accuracy in more densely treed land-cover types.
Our CNN models predict the probability of a trail/track feature for every pixel in the study area. This model can be threshold at any user-specified probability level. The precision-recall curves (
Figure 6) show that it is possible to detect virtually all the trails and tracks present in the test squares by selecting a low probability threshold. However, such a selection would also produce a high rate of false positives. Peatlands in the boreal forest are characterized by patterns of hummocks and hollows [
23] that can be mistaken for trails or tracks under some conditions. Alternatively, end-users can achieve nearly perfect rates of precision in peatlands by selecting a high probability threshold with our models. Such a selection would capture the most obvious trails and tracks but may produce higher errors of omission for less-distinct features.
Our workflow succeeds for two reasons. First, LiDAR's ability to create high-quality DTMs captures the linear depressions created by trails and tracks in the substrate, especially in areas where the tree cover is not too dense. Second, CNNs can reliably recognize these patterns under diverse conditions.
Our experience shows that trails and tracks are not readily visible in LiDAR-based products for human interpreters (
Figure 8a). The typical depth of trails is around 20 cm, which is not well captured in standard CHM stretches when trees are present. Similarly, in DTMs, even small microforms can obscure 20-cm deep trails for human experts, making it challenging to identify these features unless one knows exactly what to look for. Well-constructed digital terrain models from either drone or piloted-aircraft platforms facilitate capturing the linear patterns (
Figure 8b).
We observed a similar, albeit smaller, effect in deep learning: when normalization stretches data across a larger scale (e.g., when tall trees are present in CHMs or hills in DTMs), the range of trail values narrows, affecting convergence and accuracy. Even in this case, though, CNNs are capable of handling noise and variability, capturing these features (
Figure 8c). In the future, we plan a follow-up study to compare ways of constructing DTMs.
Our accuracies are identical to those reported by Bhatnagar et al. [
36], who mapped the tracks of mechanical forest-harvest machines in Norwegian cut blocks with CNNs. Our results are lower than the 89.5% reported by Yamamoto et al. [
35], who mapped the feeding tracks of dugongs across intertidal seagrass beds in Thailand with CNNs. Performance of our models is substantially better than the 56% reported by Kaiser et al. [
34], who used semi-automated techniques to map human trails in the transboundary region of United States and Mexico.
4.2. Canopy Density and Substrate Materials Are Key Factors
Peatland land-cover types – mapped here as high- and low-density treed fens – make up 58% of our study area but account for 82% of the trails and tracks we detected. We expect that this disproportionate concentration is a function of two factors. First, there are likely more trails and tracks captured in soft-substrate peatlands compared to hard-substrate uplands. Peatlands are characterized by thick organic soils and high water tables [
42], whose soils are easily compressed [
51] compared to mineral soils. As a result, we expect peatlands to record the tracks and trails of animals and OHVs more easily than mineral-soiled uplands.
We also anticipate overstory vegetation cover to play a role in the detectability of whatever trails and tracks are present. While LiDAR has well-known canopy penetration abilities, previous authors have documented the influence of canopy density on the technology’s effectiveness to capture the terrain surface (e.g., [52-53]). The size of canopy openings varies widely within our study area [
54], and canopy cover can approach 100% in some locations. Trails and tracks mapped under thick upland tree canopies were commonly sparser and more fragmented than those mapped under thinner peatland tree canopies (
Figure 7). Future research would do well to investigate the role that overstory vegetation density plays in our ability to map trails and tracks.
4.3. Drone- and Piloted-Aircraft Map Accuracies Are Statistically Identical
There was no significant difference in the accuracy of models developed with the 50-cm piloted-aircraft data (77 ± 9%) and the 10-cm drone data (74 ± 6%). In fact, the accuracy statistics from the piloted-aircraft map are nominally better and demonstrate more balance among error types (77% precision vs 78% recall) than those from the drone map, which showed a slight tendency to over-represent trails and tracks (70% precision vs 80% recall).
These results are somewhat surprising. We anticipated that the higher-density drone LiDAR (185 points/m2) would provide a significantly better foundation for trail and track mapping than the lower-density piloted-aircraft data (30 points/m2). However, it appears that the benefits of the higher-powered Riegl VQ-1560ii system on a piloted aircraft are enough to compensate for the superior point density delivered by the Zenmuse L1 on a drone platform.
We interpret the equivalent results from both drone and piloted-aircraft data as promising, as it demonstrates the potential for application in both small-area research and large-area operational deployments.
4.3. Patterns of Trails and Tracks in Our Study Area
The density of trails and tracks in the peatland portions of our study area was 68 km/km
2 (
Table 3). We found that the width of detected trails and tracks in peatlands varied widely: from 20 cm to 80 cm. Assuming an average width of 30 cm, we estimate that 23 ± 4% of the low-density treed fen that comprised the testing portion of our study area has been disturbed by trails and tracks.
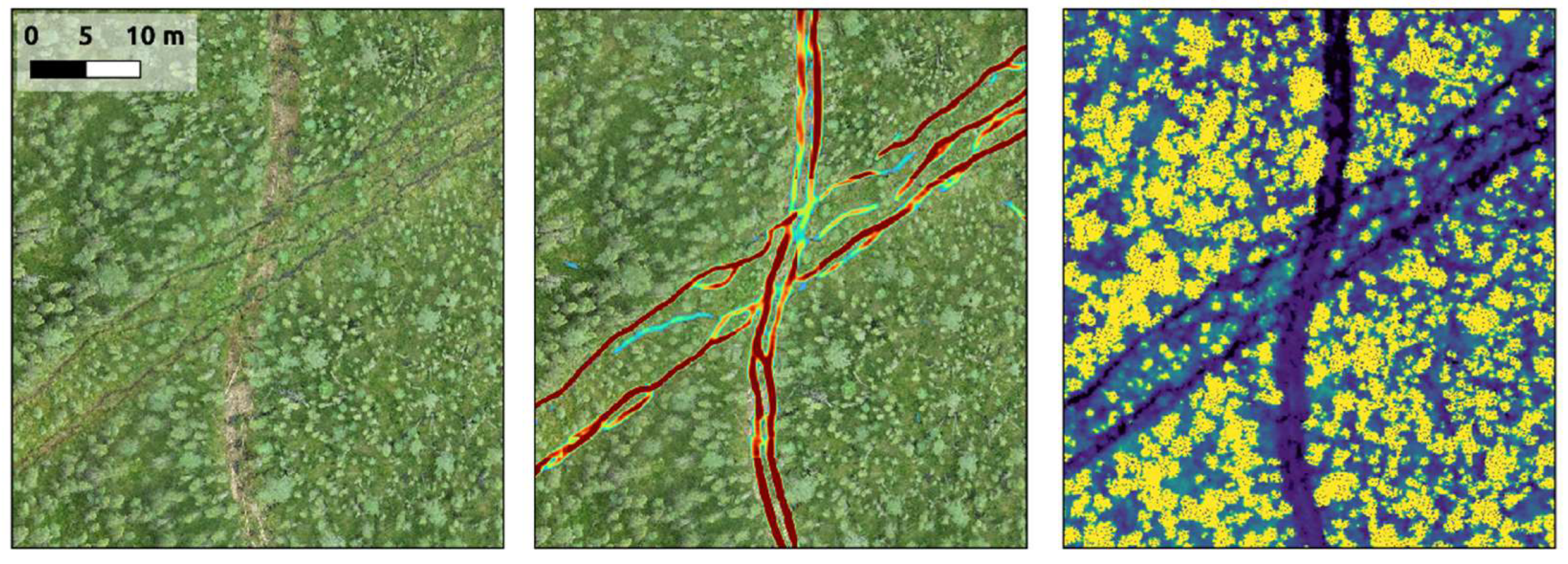
We clearly observed the tendency for wildlife and OHVs to adopt seismic lines and other linear disturbances into their transportation networks.
Figure 9 shows an example of this. The distinct tracks of a new seismic line running north-south are easily detectable in the soft organic substrate of a high-density treed fen. The wider tracks of an older seismic line running diagonally in the same figure are no longer visible, but the corridor has been adopted by wildlife trails. Overall, the density of trails and tracks detected on seismic lines within our study area is 4.4-times higher than those detected off seismic lines (
Table 3).
Previous researchers have demonstrated the effect of seismic lines on habitat-selection patterns (e.g., [55-56]) and animal-movement rates (e.g., [57-58]). Our work complements these studies and illustrates the funneling effect that linear anthropogenic disturbances have on wildlife and OHVs. Seismic lines are marked initially by distinct, parallel tracks caused by the machinery that constructed them. Over time, they become adopted as transportation corridors for large wildlife in the local area. In this process, animals adapt and reshape these disturbances. While the original machinery ruts are generally straight and parallel, the animal trails evolve into deeper and more intertwined routes. Even in cases where vegetation along the middle portion of a seismic line is recovering (e.g., the older diagonal line in
Figure 9) the deeply entrenched wildlife trails present along the edges of the feature may persist.
4.4. Ecosystem Effects of Trails and Tracks
The soft organic substrates that comprise peatlands are easily compressed by the passage of machinery [
21]. Such disturbances reduce forest regeneration [
14] [
20], increase methane emissions [
59], and amplify micro-erosion [
60]. Even after a single use, mechanical ruts may persist for years [
61] and generate wide-ranging ecological effects.
Wildlife movement can also generate substantial ecosystem effects. The trampling action of ungulates on peatlands has been studied in both arctic (e.g., Tuomi et al., 2021) and boreal (e.g., [62-63]) environments. High reindeer (
Rangifer tarandus) numbers have been associated with hummock fragmentation, waterlogged lawns, and altered vegetation communities in arctic peatlands [
64]. The effect of ungulate movement on vegetation regeneration, structure, and functioning can extend to upland land-cover types as well [
65].
Wildlife trails and tracks are particularly abundant in the peatlands portions of our study area, and we have observed this same pattern elsewhere. It could be that wildlife movement plays a key role in the formation and distribution of microtopography. Wildlife trampling can compress soft organic soils in such land-cover types, lowering the peatland surface and increasing water retention. This, in turn, alters the growth of hummock-forming
Sphagnum mosses [
66]. With long-term exposure, the cumulative effect of wildlife trampling may shape the ongoing evolution of peatland landforms. Our maps may capture the legacy of thousands of years of wildlife movement patterns within our study area and document the substantial alterations to these patterns by recent industrial disturbances.
4.5. Assumptions and Limitations
We acknowledge that trails and tracks are an imperfect representation of wildlife and OHV movement patterns. Our models detect linear depressions in the substrate associated primarily with ungulate movements and mechanical ruts. Many species leave no such detectable traces, and even target individuals (ungulates and OHVs) can move undetected over frozen or snow-covered surfaces. We further acknowledge that the detectability of such patterns is a function of substrate composition and overstory vegetation cover. We invite future research to investigate the specific role of these factors on our workflows.
We did not use field surveys to generate the reference dataset used to assess the accuracy of our models. It would have been extraordinarily challenging to create a census of all the trails and tracks in our test squares– the centerpiece of our accuracy assessment strategy – through field observations. Our experience showed that even a single 50 x 50-meter test polygon can contain up to 1,000 m of trails and tracks. Not only would documenting this density of trails and tracks on the ground be exceptionally time-consuming, but our experience is also that the aerial vantage provided by drones ultimately leads to more accurate data. For example, it is easier to distinguish microtopographic features from interlacing patterns of trails when viewed from above.
We acknowledge that our reference data set very likely contains errors of both omission and commission that influence our reported accuracy statistics. However, we contend that these errors would not serve to bias our experiments. Future authors may elect to use an alternative strategy for accuracy assessment.
Finally, we acknowledge that our work can only be regarded as a case study. Our 59-km2 study area represents a limited portion of the boreal forest, and our 0.25 km2 testing area is even more confined. The model was trained and validated across this broader region but was not explicitly tested outside of our peatland test area. While we expect the model to detect linear features similarly across various land cover types, the accuracy may vary due to differences in visibility under canopy and distinct ground characteristics. Those with soft substrates and less vegetation cover might produce better results. Areas with hard substrates and more vegetation cover are likely to produce less-accurate results.
4.6. Future Research Needs
We did not differentiate trails from tracks in our models since frequency of use does not necessarily determine the structural characteristics of detectable features. A single passage from an OHV might leave clear ruts in soft organic substrates, while regular passage over rocky or frozen substrates might be undetectable. Determining the frequency of use of detected features is an important topic for future research activities.
We also require research aimed at distinguishing wildlife trails and tracks from OHV trails and tracks. The challenge seems trivial initially but is quite difficult in practice. Humans and wildlife regularly use the same transportation corridors, and while parallel ruts can betray the presence of OHVs, they do not necessarily preclude the use of the same feature by wildlife. In fact, we regularly encountered instances in our study area where wildlife and OHV trails co-occurred.
Figure 10 illustrates an example of this. An OHV trail from the east merges onto a north-south oriented seismic line. The north-south seismic line has a well-defined wildlife trail, and all three features converge. While the track was originally an OHV trail, it remains an open question whether it has been reused or abandoned. In many areas where recreational ATV use is prohibited and industrial use is limited, these tracks may now only be used by wildlife. Distinguishing wildlife trails and tracks from OHV trails and tracks is important for practical purposes, as different deactivation strategies may be required depending on current usage. For instance, deactivating lines used primarily by wildlife might focus on the use of physical barriers like tree felling. Deactivating lines used by recreational traffic or Indigenous communities might require public outreach or community engagement.
Part of the challenge of mapping and attributing trails and tracks in natural environments is the underdeveloped definitions and lexica surrounding these features. We have already commented on the fact that the terms “trail” and “track” do not necessarily communicate utility or frequency of use, and that a single feature can serve multiple purposes. Also missing from these terms is any indication of age or utility of the feature in question, which can be similarly complicated. A single organism can leave footprints that can be preserved in rock for millions of years or create transient imprints in snow that disappear within hours. Alternatively, a single feature’s utility can transform dramatically over time. Legacy foot trails from ancient Rome can survive for centuries and provide the foundation for many modern road networks. Researchers and managers interested in inferring utility from trails and tracks in changing landscapes should be wary of these dynamics. Legacy trails created by declining wildlife populations can bias our assessments of contemporary conditions.
In this study, we elected to focus on mapping undifferentiated trails and tracks and have assigned no attributes to these features. However, future researchers may wish to develop lexica and attributes related to origin, utility, permanence, and frequency of use. To assess utility and frequency of use, ancillary data on habitat usage (camera traps, GNSS collars, DNA sampling) might be helpful. Extirpated wildlife populations (e.g., the Dawson's caribou on Canada’s west coast) may provide opportunities for mapping legacy trails and studying the rate of ground recovery. Finally, mapping trails both in summer and winter could provide a means of distinguishing legacy from contemporary trails. Such distinctions may be important, depending on the application.
Additional targets for next steps in research involve testing the current workflow in different environments and developing alterations as necessary. We have already stated the influence of overstory canopy cover on our models’ capacity to map trails and tracks. Our approach works well in relatively open environments. New approaches will have to be developed to deal with thickly forested landscapes. It is possible that disconnected trail and track fragments can be made whole by least-cost-path algorithms or other spatial approaches.
We are particularly interested in the transferability of our approaches to arid landscapes, where slower pedogenic processes [
67] might form the ideal environment for preserving trails and tracks. It would also be interesting to apply these workflows to built-up landscapes, and to test their applicability to tourism and outdoor-recreation applications (e.g., [
68]).
Finally, we could benefit from research that assesses optical and multisource datasets, perhaps combined with alternative analytical approaches. While we were drawn to LiDAR on account of its canopy penetration benefits, optical sensors provide a rich source of attribute information and well-developed processing workflows that could play an important role in this application domain.
5. Conclusions
We have demonstrated that trails and tracks can be mapped with good accuracy across diverse land-cover types in the boreal forest of northern Alberta using LiDAR and CNNs. In our case study, there was no significant difference between models developed for 50-cm DTMs from a piloted-aircraft platform (F1 score 77% ± 9%) and those developed for 10-cm DTMs from a drone platform (F1 score 74% ± 6%). This suggests that our workflows are both flexible and scalable. To our knowledge, this is the first-such demonstration of automated trail and track mapping to appear in the peer-reviewed literature.
Overall, our maps delineated 2,829 km of trails and tracks across our 59-km2 study area. Features were particularly abundant in peatlands, where a combination of soft organic substrate and relatively open canopy cover create good mapping conditions for our workflows. In those land-cover types, trail and track densities ranged from 56 km/km2 in high-density treed fens to 98 km/km2 in low-density treed fens. Upland land-cover types, where harder mineral-soil substrates underlie denser canopy cover, trails and tracks are generally more challenging to map. Even here, though, the detected trail and track density ranged from 17 km/km2 in mixed forests to 40 km/km2 in conifer forests.
Seismic line areas covered 7% of our study region but contained 27% of the detected trails and tracks. The density of trails and tracks on seismic line areas was 4.4-times higher than in the surrounding natural areas. This demonstrates the funneling effect of seismic lines on wildlife and OHV transportation corridors. This type of behaviour alters movement patterns of humans and wildlife across industrialized boreal landscapes and impedes the recovery of disturbed areas.
Our models and data sets are open-access and freely available (see Data Availability Statement, below). We particularly invite evaluations from experts working in other terrestrial ecosystems and application domains. The potential utility of accurate, spatially explicit trail and track maps generated by artificial intelligence is tremendous.
Author Contributions
Conceptualization, G.M. and I.T.; methodology, G.M., I.T.; software, I.T.; validation, I.T. and X.Y.C; formal analysis, I.T.; investigation, G.M., I.T., and X.Y.C.; resources, G.M.; data curation, G.M., I.T., and X.Y.C.; writing—original draft preparation, G.M., I.T., and X.Y.C.; writing—review and editing, G.M., I.T., and X.Y.C.; visualization, I.T.; supervision, G.M.; project administration, G.M.; funding acquisition, G.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research is part of the Boreal Ecosystem Recovery and Assessment (BERA) project (
www.bera-project.org), and was supported by a Natural Sciences and Engineering Research Council of Canada Alliance Grant (ALLRP 548285-19) in partnership with Alberta Environment and Protected Areas, Alberta-Pacific Forest Industries Inc., Canadian Natural Resources Ltd., Cenovus Energy, ConocoPhillips Canada Resources Corp., Imperial Oil Resources Ltd., Canadian Forest Service's Northern Forestry Centre, and the Alberta Biodiversity Monitoring Institute.
Data Availability Statement
Acknowledgments
The piloted-aircraft LiDAR was provided by Alberta-Pacific Forest Industries. Jim Herbers from the Alberta Biodiversity Monitoring Institute facilitated its use for this project. Jiaao Gou and Maverick Fong provided technical support and helped to process our baseline geospatial layers. Canadian Natural Resources Ltd provided access to the study site. Julia Linke coordinated most of these things, as well as many of the other moving parts that comprise the BERA program. Claudia Maurer provided administrative support. Thank you everyone!
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Abbreviations
The following abbreviations are used in this manuscript:
| LiDAR |
Light detection and ranging |
| GNSS |
Global navigation satellite system |
| CNN |
Convolutional neural netork |
| OHV |
Off-highway vehicle |
| BERA |
Boreal Ecosystem Recovery and Assessment |
| PPP |
Precise point positioning |
| RTK |
Real time kinematic |
| DTM |
Digital terrain model |
| S1 |
Sentinel 1 |
| S2 |
Sentinel 2 |
| CHM |
Canopy height model |
References
- Perna, A.; Latty, T. Animal transportation networks. Journal of the Royal Society Interface 2014, 11. [Google Scholar] [CrossRef] [PubMed]
- Myslajek, R.W.; Olkowska, E.; Wronka-Tomulewicz, M.; Nowak, S. Mammal use of wildlife crossing structures along a new motorway in an area recently recolonized by wolves. European Journal of Wildlife Research 2020, 66. [Google Scholar] [CrossRef]
- Newmark, W.D.; Rickart, E.A. High-use movement pathways and habitat selection by ungulates. Mammalian Biology 2012, 77, 293–298. [Google Scholar] [CrossRef]
- Squies, J.R.; Olson, L.E.; Roberts, E.K.; Ivan, J.S.; Hebblewhite, M. Winter recreation and Canada lynx: reducing conflict through niche partitioning. Ecosphere 2019, 10. [Google Scholar] [CrossRef]
- Hebblewhite, M.; Miguelle, D.G.; Murzin, A.A.; Aramilev, V.V.; Pikunov, D.G. Predicting potential habitat and population size for reintroduction of the Far Eastern leopards in the Russian Far East. Biological Conservation 2011, 144, 2403–2413. [Google Scholar] [CrossRef]
- Kondratenkov, I.A.; Oparin, M.L.; Oparina, O.S. Estimation of the Ecological Density of Some Species of Game according to Winter Route Censuses. Biology Bulletin 2023, 50, 2710–2718. [Google Scholar] [CrossRef]
- Whittington, J.; St Clair, C.C.; Mercer, G. Path tortuosity and the permeability of roads and trails to wolf movement. Ecology and Society 2004, 9. [Google Scholar] [CrossRef]
- Slovikosky, S.A.; Merrick, M.J.; Morandini, M.; Koprowski, J.L. Movement response of small mammals to burn severity reveals importance of microhabitat features. Journal of Mammalogy 2024, 105, 157–167. [Google Scholar] [CrossRef]
- Kays, R.; Crofoot, M.C.; Jetz, W.; Wikelski, M. Terrestrial animal tracking as an eye on life and planet. Science 2015, 348. [Google Scholar] [CrossRef]
- Muhly, T.B.; Semeniuk, C.; Massolo, A.; Hickman, L.; Musiani, M. Human Activity Helps Prey Win the Predator-Prey Space Race. Plos One 2011, 6. [Google Scholar] [CrossRef]
- Palm, E.C.; Landguth, E.L.; Holden, Z.A.; Day, C.C.; Lamb, C.T.; Frame, P.F.; Morehouse, A.T.; Mowat, G.; Proctor, M.F.; Sawaya, M.A.; et al. Corridor-based approach with spatial cross-validation reveals scale-dependent effects of geographic distance, human footprint and canopy cover on grizzly bear genetic connectivity. Molecular Ecology 2023, 32, 5211–5227. [Google Scholar] [CrossRef] [PubMed]
- Harmsen, B.J.; Foster, R.J.; Silver, S.; Ostro, L.; Doncaster, C.P. Differential Use of Trails by Forest Mammals and the Implications for Camera-Trap Studies: A Case Study from Belize. Biotropica 2010, 42, 126–133. [Google Scholar] [CrossRef]
- Rowcliffe, J.M.; Field, J.; Turvey, S.T.; Carbone, C. Estimating animal density using camera traps without the need for individual recognition. Journal of Applied Ecology 2008, 45, 1228–1236. [Google Scholar] [CrossRef]
- Cambi, M.; Certini, G.; Neri, F.; Marchi, E. The impact of heavy traffic on forest soils: A review. Forest Ecology and Management 2015, 338, 124–138. [Google Scholar] [CrossRef]
- Kormanek, M.; Dvorák, J.; Tylek, P.; Jankovsky, M.; Nuhlícek, O.; Mateusiak, L. Impact of MHT9002HV Tracked Harvester on Forest Soil after Logging in Steeply Sloping Terrain. Forests 2023, 14. [Google Scholar] [CrossRef]
- Battigelli, J.P.; Spence, J.R.; Langor, D.W.; Berch, S.M. Short-term impact of forest soil compaction and organic matter removal on soil mesofauna density and oribatid mite diversity. Canadian Journal of Forest Research 2004, 34, 1136–1149. [Google Scholar] [CrossRef]
- Croke, J.; Hairsine, P.; Fogarty, P. Soil recovery from track construction and harvesting changes in surface infiltration, erosion and delivery rates with time. Forest Ecology and Management 2001, 143, 3–12. [Google Scholar] [CrossRef]
- Robroek, B.J.M.; Smart, R.P.; Holden, J. Sensitivity of blanket peat vegetation and hydrochemistry to local disturbances. Science of the Total Environment 2010, 408, 5028–5034. [Google Scholar] [CrossRef]
- Ares, A.; Terry, T.A.; Miller, R.E.; Anderson, H.W.; Flaming, B.L. Ground-based forest harvesting effects on soil physical properties and Douglas-Fir growth. Soil Science Society of America Journal 2005, 69, 1822–1832. [Google Scholar] [CrossRef]
- Marra, E.; Cambi, M.; Fernandez-Lacruz, R.; Giannetti, F.; Marchi, E.; Nordfjell, T. Photogrammetric estimation of wheel rut dimensions and soil compaction after increasing numbers of forwarder passes. Scandinavian Journal of Forest Research 2018, 33, 613–620. [Google Scholar] [CrossRef]
- Davidson, S.J.; Goud, E.M.; Franklin, C.; Nielsen, S.E.; Strack, M. Seismic Line Disturbance Alters Soil Physical and Chemical Properties Across Boreal Forest and Peatland Soils. Frontiers in Earth Science 2020, 8. [Google Scholar] [CrossRef]
- Dabros, A.; Higgins, K.L.; Pinzon, J. Seismic line edge effects on plants, lichens and their environmental conditions in boreal peatlands of Northwest Alberta (Canada). Restoration Ecology 2022, 30. [Google Scholar] [CrossRef]
- Lovitt, J.; Rahman, M.M.; McDermid, G.J. Assessing the Value of UAV Photogrammetry for Characterizing Terrain in Complex Peatlands. Remote Sensing 2017, 9. [Google Scholar] [CrossRef]
- Filicetti, A.T.; Nielsen, S.E. Effects of wildfire and soil compaction on recovery of narrow linear disturbances in upland mesic boreal forests. Forest Ecology and Management 2022, 510. [Google Scholar] [CrossRef]
- Wimpey, J.F.; Marion, J.L. The influence of use, environmental and managerial factors on the width of recreational trails. Journal of Environmental Management 2010, 91, 2028–2037. [Google Scholar] [CrossRef]
- Costantino, C.; Mantini, N.; Benedetti, A.C.; Bartolomei, C.; Predari, G. Digital and Territorial Trails System for Developing Sustainable Tourism and Enhancing Cultural Heritage in Rural Areas: The Case of San Giovanni Lipioni, Italy. Sustainability 2022, 14. [Google Scholar] [CrossRef]
- Norman, P.; Pickering, C.M. Using volunteered geographic information to assess park visitation: Comparing three on-line platforms. Applied Geography 2017, 89, 163–172. [Google Scholar] [CrossRef]
- Loosen, A.; Capdevila, T.V.; Pigeon, K.; Wright, P.; Jacob, A.L. Understanding the role of traditional and user-created recreation data in the cumulative footprint of recreation. Journal of Outdoor Recreation and Tourism-Research Planning and Management 2023, 44. [Google Scholar] [CrossRef]
- Abdollahi, A.; Pradhan, B.; Shukla, N.; Chakraborty, S.; Alamri, A. Deep Learning Approaches Applied to Remote Sensing Datasets for Road Extraction: A State-Of-The-Art Review. Remote Sensing 2020, 12. [Google Scholar] [CrossRef]
- Washington-Allen, R.A.; Van Niel, T.G.; Ramsey, D.; West, N.E. Remote Sensing-Based Piosphere Analysis. GIScience & Remote Sensing 2004, 41, 136–154. [Google Scholar] [CrossRef]
- Dara, A.; Baumann, M.; Freitag, M.; Hölzel, N.; Hostert, P.; Kamp, J.; Müller, D.; Prishchepov, A.V.; Kuemmerle, T. Annual Landsat time series reveal post-Soviet changes in grazing pressure. Remote Sensing of Environment 2020, 239. [Google Scholar] [CrossRef]
- Chang, C.C.; Wang, J.; Zhao, Y.B.; Cai, T.Y.; Yang, J.L.; Zhang, G.L.; Wu, X.C.; Otgonbayar, M.; Xiao, X.M.; Xin, X.P.; et al. A 10-m annual grazing intensity dataset in 2015-2021 for the largest temperate meadow steppe in China. Scientific Data 2024, 11. [Google Scholar] [CrossRef] [PubMed]
- Welch, R.; Madden, M.; Doren, R.F. Mapping the Everglades. Photogrammetric Engineering and Remote Sensing 1999, 65, 163–170. [Google Scholar]
- Kaiser, J.V.; Stow, D.A.; Cao, L. Evaluation of remote sensing techniques for mapping transborder trails. Photogrammetric Engineering and Remote Sensing 2004, 70, 1441–1447. [Google Scholar] [CrossRef]
- Yamato, C.; Ichikawa, K.; Arai, N.; Tanaka, K.; Nishiyama, T.; Kittiwattanawong, K. Deep neural networks based automated extraction of dugong feeding trails from UAV images in the intertidal seagrass beds. Plos One 2021, 16. [Google Scholar] [CrossRef]
- Bhatnagar, S.; Puliti, S.; Talbot, B.; Heppelmann, J.B.; Breidenbach, J.; Astrup, R. Mapping wheel-ruts from timber harvesting operations using deep learning techniques in drone imagery. Forestry 2022, 95, 698–710. [Google Scholar] [CrossRef]
- Marra, E.; Wictorsson, R.; Bohlin, J.; Marchi, E.; Nordfjell, T. Remote measuring of the depth of wheel ruts in forest terrain using a drone. International Journal of Forest Engineering 2021, 32, 224–234. [Google Scholar] [CrossRef]
- Flyckt, J.; Andersson, F.; Lavesson, N.; Nilsson, L.; Gren, A. Detecting ditches using supervised learning on high-resolution digital elevation models. Expert Systems with Applications 2022, 201. [Google Scholar] [CrossRef]
- Lidberg, W.; Paul, S.S.; Westphal, F.; Richter, K.F.; Lavesson, N.; Melniks, R.; Ivanovs, J.; Ciesielski, M.; Leinonen, A.; Agren, A.M. Mapping Drainage Ditches in Forested Landscapes Using Deep Learning and Aerial Laser Scanning. Journal of Irrigation and Drainage Engineering 2023, 149. [Google Scholar] [CrossRef]
- Du, L.; McCarty, G.W.; Li, X.; Zhang, X.; Rabenhorst, M.C.; Lang, M.W.; Zou, Z.H.; Zhang, X.S.; Hinson, A.L. Drainage ditch network extraction from lidar data using deep convolutional neural networks in a low relief landscape. Journal of Hydrology 2024, 628. [Google Scholar] [CrossRef]
- Natural Regions Committee, Natural Regions and Subregions of Alberta. Compiled by D.J. Downing and W.W. Pettapiece 2006, Pub. No. T/852 .
- Vitt, D.H. An overview of factors that influence the development of Canadian peatlands. Memoirs of the Entomological Society of Canada 1994, 7–20. [Google Scholar] [CrossRef]
- Dabros, A.; Pyper, M.; Castilla, G. Seismic lines in the boreal and arctic ecosystems of North America: environmental impacts, challenges, and opportunities. Environmental Reviews 2018. [Google Scholar] [CrossRef]
- Roussel, J.R.; Auty, D.; Coops, N.C.; Tompalski, P.; Goodbody, T.R.H.; Meador, A.S.; Bourdon, J.F.; de Boissieu, F.; Achim, A. lidR: An R package for analysis of Airborne Laser Scanning (ALS) data. Remote Sensing of Environment 2020, 251. [Google Scholar] [CrossRef]
- Roussel, J.R.; Auty, D. Airborne LiDAR Data Manipulation and Visualization for Forestry Applications. R package version 4.1.2 2024, https://cran.r-project.org/package=lidR.
- Ronneberger, O.; Fischer, P.; Brox, T. U-Net: Convolutional Networks for Biomedical Image Segmentation. arXiv 2015, arXiv:1505.04597. [Google Scholar]
- Oktay, O.; Schlemper, J.; Le Folgoc, L.; Lee, M.; Heinrich, M.; Misawa, K.; Mori, K.; McDonagh, S.; Hammerla, N.Y.; Kainz, B.; Glocker, B.; Rueckert, D. Attention U-Net: Learning Where to Look for the Pancreas. arXiv 2018, arXiv:1804.03999. [Google Scholar]
- Kingma, D.P.; Ba, J. Adam: A Method for Stochastic Optimization. arXiv 2014, arXiv:1412.6980. [Google Scholar]
- Zhang, T.Y.; Suen, C.Y. A Fast Parallel Algorithm for Thinning Digital Patterns. Commun. ACM 1984, 27, 236–239. [Google Scholar] [CrossRef]
- Queiroz, G.L.; McDermid, G.J.; Rahman, M.M.; Linke, J. The Forest Line Mapper: A Semi-Automated Tool for Mapping Linear Disturbances in Forests. Remote Sens. 2020, 12, 4176. [Google Scholar] [CrossRef]
- Price, J.S.; Cagampan, J.; Kellner, E. Assessment of peat compressibility: is there an easy way? Hydrological Processes 2005, 19, 3469–3475. [Google Scholar] [CrossRef]
- Crow, P.; Benham, S.; Devereux, B.J.; Amable, G.S. Woodland vegetation and its implications for archaeological survey using LiDAR. Forestry 2007, 80, 241–252. [Google Scholar] [CrossRef]
- Razak, K.A.; Straatsma, M.W.; van Westen, C.J.; Malet, J.P.; de Jong, S.M. Airborne laser scanning of forested landslides characterization: Terrain model quality and visualization. Geomorphology 2011, 126, 186–200. [Google Scholar] [CrossRef]
- Dietmaier, A.; McDermid, G.J.; Rahman, M.M.; Linke, J.; Ludwig, R. Comparison of LiDAR and Digital Aerial Photogrammetry for Characterizing Canopy Openings in the Boreal Forest of Northern Alberta. Remote Sensing 2019, 11. [Google Scholar] [CrossRef]
- Finnegan, L.; Hebblewhite, M.; Pigeon, K.E. Whose line is it anyway? Moose (<i>Alces alces</i>) response to linear features. Ecosphere 2023, 14. [Google Scholar] [CrossRef]
- Finnegan, L.; Pigeon, K.E.; Cranston, J.; Hebblewhite, M.; Musiani, M.; Neufeld, L.; Schmiegelow, F.; Duval, J.; Stenhouse, G.B. Natural regeneration on seismic lines influences movement behaviour of wolves and grizzly bears. Plos One 2018, 13. [Google Scholar] [CrossRef]
- Dickie, M.; Serrouya, R.; McNay, R.S.; Boutin, S. Faster and farther: wolf movement on linear features and implications for hunting behaviour. Journal of Applied Ecology 2017, 54, 253–263. [Google Scholar] [CrossRef]
- Dickie, M.; Sherman, G.G.; Sutherland, G.D.; McNay, R.S.; Cody, M. Evaluating the impact of caribou habitat restoration on predator and prey movement. Conservation Biology 2023, 37. [Google Scholar] [CrossRef]
- Lovitt, J.; McDermid, G.J.; Kariyeva, J.; Mahoney, C.; Hall, R.J.; Franklin, S.E.; Saxena, A.; Kalacska, M. UAV Remote Sensing Can Reveal the Effects of Low-Impact Seismic Lines on Surface Morphology, Hydrology, and Methane (CH₄) Release in a Boreal Peatland. Remote Sens. 2018, 10, 1549. [Google Scholar]
- Evans, M.; Warburton, J. Erosion in peatlands: Recent research progress and future directions. Earth-Sci. Rev. 2010, 103, 30–49. [Google Scholar]
- Williams-Mounsey, J.; Crowle, A.; Grayson, R.; Lindsay, R.; Holden, J. Surface structure on abandoned upland blanket peatland tracks. Journal of Environmental Management 2023, 325. [Google Scholar] [CrossRef]
- Pellerin, S.; Huot, J.; Côté, S.D. Long term effects of deer browsing and trampling on the vegetation of peatlands. Biological Conservation 2006, 128, 316–326. [Google Scholar] [CrossRef]
- Persson, I.L.; Danell, K.; Bergström, R. Disturbance by large herbivores in boreal forests with special reference to moose. Annales Zoologici Fennici 2000, 37, 251–263. [Google Scholar]
- Holmgren, M.; Groten, F.; Carracedo, M.R.; Vink, S.; Limpens, J. Rewilding Risks for Peatland Permafrost. Ecosystems 2023. [Google Scholar] [CrossRef]
- Ramirez, J.I.; Jansen, P.A.; Poorter, L. Effects of wild ungulates on the regeneration, structure and functioning of temperate forests: A semi-quantitative review. Forest Ecology and Management 2018, 424, 406–419. [Google Scholar] [CrossRef]
- Bengtsson, F.; Rydin, H.; Baltzer, J.L.; Bragazza, L.; Bu, Z.J.; Caporn, S.J.M.; Dorrepaal, E.; Flatberg, K.I.; Galanina, O.; Galka, M.; et al. Environmental drivers of<i>Sphagnum</i>growth in peatlands across the Holarctic region. Journal of Ecology 2021, 109, 417–431. [Google Scholar] [CrossRef]
- Nir, N.; Davidovich, U.; Ullman, M.; Schütt, B.; Stahlschmidt, M.C. The environmental footprint of Holocene societies: a multi-temporal study of trails in the Judean Desert, Israel. Frontiers in Earth Science 2023, 11. [Google Scholar] [CrossRef]
- Godtman Kling, K.; Fredman, P.; Wall-Reinius, S. Trails for tourism and outdoor recreation: A systematic literature review. Tourism: An International Interdisciplinary Journal 2017, 65, 488–508. [Google Scholar]
Figure 1.
Our 59-km2 study site is in northeastern Alberta, Canada. It contains a diverse mosaic of land-cover types, from sparsely treed lowlands to thickly treed uplands.
Figure 1.
Our 59-km2 study site is in northeastern Alberta, Canada. It contains a diverse mosaic of land-cover types, from sparsely treed lowlands to thickly treed uplands.
Figure 2.
Wildlife trails (a), off-highway vehicle tracks (b), and seismic lines (c) are the three types of trails and tracks common to our study area. Ground-level (top row) and aerial-view (bottom row) examples of all three features are shown.
Figure 2.
Wildlife trails (a), off-highway vehicle tracks (b), and seismic lines (c) are the three types of trails and tracks common to our study area. Ground-level (top row) and aerial-view (bottom row) examples of all three features are shown.
Figure 3.
Working within the training portion of the study area (large blue square), we extracted training patches cropped around 6,395 random points (white dots in a). These 512 x 512 pixel training patches (b) were labeled using a preliminary U-Net model. These patches were then used to train a second, more refined set of U-Net models: one for the 10cm drone data and a second for the 50cm piloted-aircraft data. We tested the accuracy of these models in a physically independent test area (small red square) containing 11 randomly distributed testing polygons (red dots).
Figure 3.
Working within the training portion of the study area (large blue square), we extracted training patches cropped around 6,395 random points (white dots in a). These 512 x 512 pixel training patches (b) were labeled using a preliminary U-Net model. These patches were then used to train a second, more refined set of U-Net models: one for the 10cm drone data and a second for the 50cm piloted-aircraft data. We tested the accuracy of these models in a physically independent test area (small red square) containing 11 randomly distributed testing polygons (red dots).
Figure 4.
A quantitative accuracy assessment was performed across 11 non-overlapping test polygons (red squares in a) distributed randomly across the testing portion of the study area. Insets show the RGB orthomosaic (b), canopy height model (c), and mapped reference trails and tracks (d) in one of the test polygons.
Figure 4.
A quantitative accuracy assessment was performed across 11 non-overlapping test polygons (red squares in a) distributed randomly across the testing portion of the study area. Insets show the RGB orthomosaic (b), canopy height model (c), and mapped reference trails and tracks (d) in one of the test polygons.
Figure 5.
Trail/track density map of the study area (main map) and trail/track map (insets) for a portion of our study area transitioning from forested upland (left) to treed fen (right). Trails and tracks mapped in forested uplands are sparser and more fragmented than those in the peatland portions of our study area.
Figure 5.
Trail/track density map of the study area (main map) and trail/track map (insets) for a portion of our study area transitioning from forested upland (left) to treed fen (right). Trails and tracks mapped in forested uplands are sparser and more fragmented than those in the peatland portions of our study area.
Figure 6.
Confusion matrices and precision-recall curves for trails and tracks mapped with different sources of input data. Panel A shows the confusion matrix for in the model developed from the 10-cm drone digital terrain model (DTM). Panel B shows the confusion matrix for the model developed from the 50-cm piloted-aircraft DTM. Panel C shows the precision-recall curves for both models. Statistics were generated from 9,118 reference points from the testing portion of our study area.
Figure 6.
Confusion matrices and precision-recall curves for trails and tracks mapped with different sources of input data. Panel A shows the confusion matrix for in the model developed from the 10-cm drone digital terrain model (DTM). Panel B shows the confusion matrix for the model developed from the 50-cm piloted-aircraft DTM. Panel C shows the precision-recall curves for both models. Statistics were generated from 9,118 reference points from the testing portion of our study area.
Figure 7.
Digital surface model (A) and trails and tracks (B) for a portion of our study area transitioning from open peatlands (lower left) to thickly forested uplands (upper right). Trails and tracks mapped in are sparser and more fragmented in the thickly forested upland portions of our study area.
Figure 7.
Digital surface model (A) and trails and tracks (B) for a portion of our study area transitioning from open peatlands (lower left) to thickly forested uplands (upper right). Trails and tracks mapped in are sparser and more fragmented in the thickly forested upland portions of our study area.
Figure 8.
Trails and tracks within our study area are not generally visible in LiDAR canopy height models (Column A), which accentuate canopy altering landscape factors like anthropogenic disturbances and land-cover type. However, well-constructed digital terrain models (Column B) reveal linear patterns associated with the movement of wildlife and off-highway vehicles that can be captured as trails and tracks (Column C) by convolutional neural networks.
Figure 8.
Trails and tracks within our study area are not generally visible in LiDAR canopy height models (Column A), which accentuate canopy altering landscape factors like anthropogenic disturbances and land-cover type. However, well-constructed digital terrain models (Column B) reveal linear patterns associated with the movement of wildlife and off-highway vehicles that can be captured as trails and tracks (Column C) by convolutional neural networks.
Figure 9.
A comparison of new and wildlife-adopted older seismic-line disturbances. A new seismic line, cut in 2022, runs north-south through this section of high-density treed fen. The narrow track ruts of the mulcher that cut it are difficult to see in a 2-cm drone orthomosaic (a) but present in the LiDAR data (c) and easily detected by our models (b). The wider-spaced tracks from the older seismic line running diagonally are no longer visible, but the corridor has been adopted by wildlife into deeper and more intertwined routes, which are once again detectable by our models.
Figure 9.
A comparison of new and wildlife-adopted older seismic-line disturbances. A new seismic line, cut in 2022, runs north-south through this section of high-density treed fen. The narrow track ruts of the mulcher that cut it are difficult to see in a 2-cm drone orthomosaic (a) but present in the LiDAR data (c) and easily detected by our models (b). The wider-spaced tracks from the older seismic line running diagonally are no longer visible, but the corridor has been adopted by wildlife into deeper and more intertwined routes, which are once again detectable by our models.
Figure 10.
The co-occurrence of off-highway vehicle (OHV) and wildlife trails and tracks complicates the challenge of distinguishing among feature types. In this scene, an OHV trail coming from the east merges onto a north-south oriented seismic line. The seismic line has a well-defined wildlife trail on it, and all three features merge together. Is this a wildlife trail, an OHV trail, or both? The image set displayed includes an 0.5-cm orthomosaic (top), a 10-cm digital terrain model (middle); and LiDAR-derived trail/track map (bottom). The probability of trails and tracks in (c) increases from blue to red.
Figure 10.
The co-occurrence of off-highway vehicle (OHV) and wildlife trails and tracks complicates the challenge of distinguishing among feature types. In this scene, an OHV trail coming from the east merges onto a north-south oriented seismic line. The seismic line has a well-defined wildlife trail on it, and all three features merge together. Is this a wildlife trail, an OHV trail, or both? The image set displayed includes an 0.5-cm orthomosaic (top), a 10-cm digital terrain model (middle); and LiDAR-derived trail/track map (bottom). The probability of trails and tracks in (c) increases from blue to red.
Table 1.
Summary statistics of deep-learning models developed from 50-cm airborne digital terrain model (DTM) and 10-cm drone DTM. The metrics were obtained from 9,118 reference points from the test portion of our study area.
Table 1.
Summary statistics of deep-learning models developed from 50-cm airborne digital terrain model (DTM) and 10-cm drone DTM. The metrics were obtained from 9,118 reference points from the test portion of our study area.
| Data Source |
Precision
(%) |
Recall
(%) |
F1-score
(%) |
Average Precision |
| Aerial 50 cm DTM |
77 ± 9 |
78 ± 14 |
77 ± 9 |
0.69 |
| Drone 10 cm DTM |
70 ± 10 |
80 ± 8 |
74 ± 6 |
0.64 |
Table 2.
Numerical summaries of the length and density of trails and tracks across land-cover types within the study area.
Table 2.
Numerical summaries of the length and density of trails and tracks across land-cover types within the study area.
| |
|
Trails and Tracks |
| Land-cover Type |
Land-cover Area km2 (%) |
Length
km (%) |
Density (km/km2) |
| Coniferous forest |
10 (16.9) |
396 (14.0) |
40 |
| Deciduous forest |
3 (5.1) |
62 (2.2) |
21 |
| Mixed forest |
3 (5.1) |
51 (1.8) |
17 |
| High-density treed fen |
24 (40.7) |
1342 (47.4) |
56 |
| Low-density treed fen |
10 (16.9) |
978 (34.6) |
98 |
| Excluded areas (lakes, floodplains, roads, and dense industrial footprint) |
9 (15.2) |
N/A |
N/A |
| SUM |
59 (100) |
2829 (100) |
|
Table 3.
Numerical summaries of the length and density of trails and tracks on and off seismic lines within the study area.
Table 3.
Numerical summaries of the length and density of trails and tracks on and off seismic lines within the study area.
| |
Area,
km2 (%) |
Trails and Tracks Length
km (%) |
Trails and Tracks Density
km/km2 |
| On seismic lines |
4.2 (8) |
765 (27) |
182 |
| Off seismic lines |
45.7 (92) |
2,064 (73) |
41 |
| SUM |
49.9 (100) |
2,829 (100) |
|
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).