Submitted:
19 February 2025
Posted:
20 February 2025
You are already at the latest version
Abstract
(1) Background: Since there is no definite cure for Parkinsons Disease (PD) there is a growing interest for novel therapeutical agents with neuroprotective potential. Aim of this review is to update the knowledge of nuclear factor-erythroid 2-related factor 2(Nrf2) as neuroprotective agent ; (2) Methods: briefly describe the main methods or treatments applied; (3) Results: Activation of Nrf2, a transcription factor that regulates the expression of antioxidant and cytoprotective genes, has emerged as a promising therapeutic strategy for PD. Nrf2 is a master regulator of the cellular antioxidant response and is responsible for activating the expression of genes that encode antioxidant enzymes such as superoxide dismutase, catalase, and glutathione peroxidase. These enzymes help to neutralize ROS and protect cells from oxidative damage. Nrf2 also regulates the expression of genes involved in the detoxification of harmful substances and the repair of damaged DNA. In addition to its antioxidant and cytoprotective functions, Nrf2 has been shown to have anti-inflammatory properties that can help to alleviate the neuroinflammation associated with PD. Pharmacological agents such as sulforaphane, curcumin, and resveratrol have been shown to activate Nrf2 and upregulate the expression of antioxidant genes in preclinical PD models . These compounds work by disrupting the interaction between Nrf2 and its negative regulator, Keap1, leading to the translocation of Nrf2 into the nucleus where it can activate the expression of cytoprotective genes. Clinical trials are currently underway to evaluate the efficacy of these pharmacological agents in patients with PD. (4) Conclusions: While the neuroprotective role of Nrf2 in PD holds great promise for the development of novel therapies, there are several challenges and limitations that need to be addressed in order to harness the full potential of this pathway in the clinic. Overcoming these obstacles will require interdisciplinary collaborations, innovative research approaches, and a greater understanding of the complex pathophysiology of PD. By addressing these challenges, we can move closer to developing effective neuroprotective therapies that can slow or stop the progression of PD and improve the quality of life for patients with this devastating disease.
Keywords:
nuclear factor-erythroid 2-related factor 2 (Nrf2)
; Parkinson’s disease (PD)
; reactive oxygen species (ROS)
; neuroprotection
1. Introduction
Parkinson’s disease (PD) is a progressive neurodegenerative disorder characterized by the aggregation of misfolded a-synuclein and progressive loss of dopaminergic neurons (DA) in the substantia nigra (SN) pars compacta of the brain [1]. This loss leads to the motor symptoms of tremors, rigidity, and bradykinesia, as well as with non-motor symptoms such as cognitive impairments, depression, and sleep disturbances [1]. Since no definite cure exists, only symptomatic pharmaceutical options of levodopa, COMT inhibitors, anticholinergics agents, dopaminergic agonists, and inhibitors of MAO-B, are available with inconsistent results [2]. The etiology of PD is not fully understood, but both environmental and genetic (PINK1, leucine-rich repeat kinase 2 (LRRK2), Parkin, DJ-1, and α-synuclein) factors are involved in the progression of PD. However, oxidative stress and inflammation have been implicated in the pathogenesis of the disease. Oxidative stress is caused by an imbalance between the production of reactive oxygen species (ROS) and the ability of the body to detoxify these harmful molecules. Inflammation is an immune response that can exacerbate oxidative stress and further damage neurons in the brain. As the population ages, the incidence of PD in individuals over 50 years old is projected to rise approximately twofold, reaching an estimated 8.7 to 9.3 million cases by the year 2030 with enormous socioeconomic burden [3].
Activation of nuclear factor-erythroid 2-related factor 2 (Nrf2), a transcription factor that regulates the expression of antioxidant and cytoprotective genes, has been demonstrated to decrease alpha-synuclein aggregation and toxicity [4]. Nrf2 is a master regulator of the cellular antioxidant response and is responsible for activating the expression of genes that encode antioxidant enzymes such as superoxide dismutase, catalase, and glutathione peroxidase [5,6]. These enzymes help to neutralize ROS and protect cells from oxidative damage. Nrf2 also regulates the expression of genes involved in the detoxification of harmful substances and the repair of damaged DNA [5]. In addition to its antioxidant and cytoprotective functions, Nrf2 has been shown to have anti-inflammatory properties that can help to alleviate the neuroinflammation associated with PD [7]. Therefore, knowledge of the regulation and function of the Nrf2-KEAP1 pathway offers possibilities for alternative therapies approaches.
Several previous reviews have separately focused on the different Nrf2 pathways as promising therapeutic targets for PD [4,8,9,10]. However the aim of this narrative review is to unify the various Nrf2 pathways and delineate their neuroprotective role in a potential novel PD therapy. The connection between oxidative stress, neuroinflammation and the pathophysiology of PD are briefly reviewed in this article. After that, i gave a brief synopsis of the Keap1/Nrf2/ARE pathway and explained how Nrf2/HO-1 activation contributes to PD. Finally, i broaden our discussion to include the neuroprotective benefits of bioactive substances’ pharmacological manipulation of Nrf2 and the possible use of Nrf2 activators in the treatment of PD, as well as the challenges that limits the use of them from the clinical practice.
2. Materials and Methods
2.1. Data Collection
Articles were reviewed, focusing on Nrf2 neuroprotective pathways for treatment of PD. PUBMED, SCOPUS and EMBASE were searched until 1/2/2025. In more detail, the following Boolean search string was used: ((Parkinson) OR ((nuclear factor-erythroid 2-related factor 2) OR (Nrf2/ARE pathway) OR (Nrf2/HO-1 signaling pathway) OR (p62-Keap1-Nrf2-aldo-keto reductase 1C) OR (reactive nitrogen species), OR (Keap1–Nrf2-ARE), OR (reactive oxygen species), OR (methyl-4-phenyl-1,2,3,6-tetrahydropyridine),OR (Universal Asynchronous Receiver–Transmitter) OR (resveratrol), OR (curcumin), OR (naphthazarin), OR (genistein) , OR ( carnosic acid), OR ( berberin), OR (Sulforaphane), AND (neuroprotection).
No additional filters were applied in the literature search.
2.2. Inclusion and Exclusion Criteria
Including criteria were in vivo/ in vitro original research papers, reviews, and meta-analyses that have been published in peer-reviewed publications in English language.
The exclusion criteria are the following:
1. Books, book chapters, peer-reviewed papers, and publications issued as “letters,” “comments,” “perspectives,” “case reports,” or “surveys.”
2. Studies of non-English literature.
3. Research on illnesses other than PD.
4. Research that collected data without the activation of Nrf2 neuroprotective pathways for PD.
5. Research demonstrating the outcomes of a contest, challenge, or initiative.
The snowballing procedure was carried out to screen the references of each selected article for potential extra papers to cover the current key evidence following by a full-text evaluation of pertinent research. For the narrative synthesis, my initial categorization was based on the Nrf2 neuroprotective pathways for PD, followed by a discussion of limitations and potential future directions.
3. Results
3.1. Key Features of Nrf2
In 1994, the 605 amino acid protein known as nuclear factor-erythroid 2-related factor 2 (Nrf2) was cloned, which belongs to the Cap ’n’ Collar (CNC) family and has a basic leucine zipper (bZIP) [11]. Six structural domains (Neh1-6) that are largely conserved make up Nrf2 . A crucial link in the dimerization of Nrf2 with DNA is the Neh1 domain, which also dimerizes the broad-complex, tram track, bric-abrac (BTB), and CNC homolog (Bach) proteins. There are many lysine residues near the N-terminus of the Neh2 structural domain, which has a low affinity DLG motif and a high affinity ETGE motif [12]. Two motifs on a Neh2 will bind two Kelch-like ECH-associated protein 1 (Keap1), and when they bind to form a stable complex, Nrf2 is ubiquitinated [13]). An initial ETGE-mediated association forming an “open” KEAP1-NRF2 complex, and a subsequent DLG-mediated intra-complex association forming a “closed” KEAP1-NRF2 complex [12]. The transcriptional activity of Nrf2 is maintained by the C-terminal Neh3, whereas the two acidic residue-rich Neh4 and Neh5 interact with transcriptional coactivators and the CREB-binding protein (CBP) [14]. Neh6 is linked to the phosphorylation of glycogen synthase kinase-3 (GSK-3), has two motifs, DSGIS and DSAPGS, and has a lot of serine residues [15]. Nevertheless, new research has reported on the inhibitory Neh7 structural domain of Nrf2. By blocking the RXRα receptor, Neh7 influences Nrf2 activity, which in turn permits Nrf2 to prevent transcriptional coactivators from attaching to the Neh4 and Neh5 structural domains [16] (Figure 1).
3.2. The Neuroprotective Role of Nrf2 in the Iron Death PD Related Mechanism
A new cell death process called iron death influences gene expression and is a major factor in PD [17]. Free iron release and cytoplasmic iron efflux are necessary for the production of ferritin heavy chains, which guards against cell damage brought on by Fentonian iron death [18]. Nrf2 expression may be regulated by mitochondrial RNA (mtRNA), with distinct isoforms influencing Nrf2 activation or aggregation. A metabolite that produces Fe2+ and acts as a protective factor, heme oxygenase (HO) can control intracellular iron levels through its Nrf2/HO-1 pathway. The Nrf2 target genes HO-1 and NQO1 are impacted by the activation of the sequestosome 1 (SQSTM1) /p62-Keap1-Nrf2-aldo-keto reductase 1C (AKR1C) pathway, which controls iron death [19]. In DA neurons, microglia and astrocytes help maintain iron homeostasis and lessen iron excess, showing their neuroprotective properties. Astrocytic regulation of the Nrf2-ARE pathway is a viable target for therapies aiming at lowering or preventing neuronal death in PD, and Nrf2 expression limited to astrocytes is adequate to protect against MPTP [20].The Figure 2 summarized the main characteristics of Nrf2 neuroprotective role for PD
3.3. The Neuroprotective Role of Nrf2 in Oxidative Stress Related Mechanisms of PD Pathogenesis
One crucial component of organisms is the equilibrium between the negative and positive effects of free radicals. Mechanisms known as “redox regulation” are responsible for achieving this equilibrium. In PD, Nrf2 is an essential cellular defense system that shields cells from oxidative stress by upregulating the expression of about 200 cytoprotective genes. By promoting the production of genes for antioxidant enzymes, the Nrf2-Keap1 signaling pathway serves as a key defensive mechanism against oxidative stress [21]. Since Keap1 is a negative regulator of Nrf2, reactive chemicals such as reactive nitrogen species (RNS), electrophile reagents, ROS, and xenobiotics can oxidize it, increasing the nucleus and making Nrf2 seem unbound, which in turn controls the redox status of the cell. Additionally, Nrf2 controls mitochondrial biogenesis, redox homeostasis, and the upregulation of antioxidant/electrophilic response elements (ARE/ERE), which promotes the production of protective genes [22,23].
3.4. The Neuroprotective Role of Nrf2 in Neuroinflammation Related Mechanisms of PD Pathogenesis
Glial cells are essential for neuroprotection since they express a lot of Nrf2. Dopamine can stimulate them in order to keep dopamine neurons alive. Inhibiting inflammatory responses triggered by p65, IL-1β, TNF-α, caspase-1, and the NLRP3 inflammasome, Nrf2 is a crucial regulator of inflammation [23]. Nrf2 primarily controls the following proteins: HO-1, NQO-1, GSH, GPx, and a number of enzymes. Yim et al. [24] showed the translocation of Nrf2 from the cytosol to the nucleus after 6 h of ICM treatment, followed by the upregulation of heme-oxygenase 1 (HO-1) and NAD(P)H quinone oxidoreductase-1 (NQO1) after an additional 6 h, indicating that inflachromene (ICM) safeguards PC12 cells via the activation of the Nrf2 pathways [24]. Furthermore, ICM exhibited remarkable efficacy in inhibiting the loss of dopaminergic neurons and the activation of astrocytes and microglia. In the same line, the chemokine fractalkine (CX3CL1)/CX3CL1 receptor (CX3CR1) axis prevents microglia overactivation in PD by upregulating Nrf2 expression [25]. By preventing NF-κB signaling from being activated, Nrf2 has a positive neuroprotective effect on PD. In terms of PD genetics, DJ-1 stabilizes Nrf2, boosts its transcriptional activity, and stops Keap1 from inactivating it [26]. Nrf2 shields dopamine neurons from the neurotoxicity caused by oxidative stress by activating the transcriptional upregulation of PINK1(Figure 2).
3.5. The Neuroprotective Role of Nrf2 in Dysfunctional Mitochondria Related Mechanisms of PD Pathogenesis
Mitochondria-targeted gene therapy is a promising strategy to improve PD symptomatology [27]. Nrf2 activates genes related to mitochondrial functions and increases the expression of mitochondrial transcription factor A (TFAM). It controls the generation of ROS in mitochondria and NADPH oxidase [28]. Nrf2 affects mitochondrial membrane potential and ATP synthesis, but can also reduce ROS production in the presence of oxidative stress or cytokines. Dysfunctional mitochondria lead to dopamine neuron loss and PD symptoms [27]. Redox-critical cofactors like NADH and NADHP, whose synthesis is intimately related to Nrf2, are accessible through the pentose phosphate pathway of the cytoplasm and the tricarboxylic acid cycle of mitochondria [29]. Nrf2 regulation could improve PD symptoms by regulating the pentose phosphate pathway (Figure 2).
3.6. Activation of Nrf2 as Therapeutical Modulator of PD
There is growing evidence that in vitro or in vivo models of neurotoxicity caused by 6-OHDA (Table 2), MPP+ (Table 3), MPTP (Table 4), paraquat (Table 5), and rotenone showed far higher resistance to activators of the Nrf2/ARE pathway. Pharmacologic drugs that activate Nrf2 have been demonstrated to provide neuroprotection; on the other hand, a lack in Nrf2 increased the vulnerability of neurons to the neurotoxic. The published research is increasingly showing that Nrf2 activation, either prior to or concurrent with neurotoxin exposure, can provide protection against neurotoxicity caused by neurotoxins in PD. One promising option for treating PD is to target Nrf2 activation.
Heme is broken down into carbon monoxide, free iron, and biliverdin by the powerful antioxidant enzyme heme oxygenase-1 (HO-1), which is controlled by Nrf2. Higher levels of HO-1 have been correlated with a decreased right hippocampal volume in the plasma of PD patients [30]. Depending on the particular model used, the length and severity of HO-1 induction, and the chemistry of the local redox microenvironment, increased HO-1 activity may either lessen or worsen brain damage [31]. As a potential novel cure for Parkinson’s disease, HO-1 plays a role in neuroprotection against OS-dependent harm. By showing that elevation of HO-1 expression drives an adaptive cytoprotective response to OS in cultured human fibroblasts, Tyrrell and colleagues initially demonstrated the cytoprotective function of HO-1. The part HO-1 plays in PD is very intriguing.
3.6.1. Natural Compounds Targeting the Nrf2/HO-1 Signaling Pathway as a Neuroprotective Agent
Numerous bioactive substances have been identified as Nrf2 activators that have beneficial effects in as potential therapeutical agents of PD, including resveratrol, curcumin, naphthazarin, genistein, carnosic acid, and berberin (Table 1).
Among the known hormetic phytochemicals, naphthoquinones, such naphthazarin, a naphthoqui-none derivative, plays a neuroprotective role in the MPTP and MPP1-induced Parkinson’s disease-like pathogenesis models [32]. Rather of directly protecting neurons, naphthazarin-mediated neuroprotection is obtained via the regulation of astrocyte activity.
Similarly, others hormeticphytochemicals have been reported to exert neuro protective effects against PD. In vitro and in vivo, it has been demonstrated that curcumin, resveratrol, and green tea catechins protect against the development of Parkinson’s disease-like conditions caused by MPTP, MPP1, and 6-hydroxydopamine [33,34].In the rotenone mouse model of PD, decreased levels of p62 and LC3-II autophagic markers validated the impaired autophagy process, which in turn increased the aggregation of misfolded protein α-synuclein [35]. On the other hand, curcumin administration increased LC3-II expression and inhibited the apoptotic cascade, which improved autophagy-mediated clearance of misfolded α-synuclein proteins. The enhanced anti-oxidative activity is justified by the fact that curcumin treatment normalized the Nrf2-Keap1 pathway and increased Nrf2 expression showing a neuroprotective potential for novel PD therapy [35].
Sulforaphane, (SFN), an isothiocyanate compound found naturally in cruciferous vegetables like broccoli, may stimulate Nrf2 with antioxidant and anti-inflammatory responses. The results demonstrate that SFN directly activates BDNF transcription through Nrf2 binding with the Bdnf exon I promoter, resulting in an increased expression of BDNF protein, and ultimately shield nigral dopaminergic neurons from MPTP-induced cell death [36]. According to a prior research, SFN buffered wild-type mice against MPTP-induced neurotoxicity, but not Nrf2 KO mice, implying that Nrf2 plays a part in SFN’s neuroprotective properties [37]. Alongside the neuroprotective benefits, there was a reduction in pro-inflammatory cytokine production, microgliosis, and astrocytes.
Nitric oxide (NO) and lipid peroxidation (LPO) levels induced by RTN were decreased by BR treatment, which also improved antioxidative capacity, increased the activities of mitochondrial enzymes such as succinate dehydrogenase (SDH), ATPase, and the electron transport chain (ETC), and decreased striatal neuroinflammation and apoptosis markers (such as nitrite, TBARS, TNF-α, IL-1β, IL-6, and caspase-3) [38]. In particular, BBR’s protective effects were considerably reduced when trigonelline (TGN), an inhibitor of the Nrf2 pathway, was administered concurrently. This suggests that the Nrf2 pathway mediates BBR’s neuroprotective benefits. These findings highlight BBR’s capacity to improve PD-like motor deficits, indicating that it may be able to postpone or control PD symptoms.
4. Discussion
The Nrf2/ARE pathway is essential for cellular adaptation because it regulates coordinated cytoprotective proteins, such as HO-1, to counteract OS in PD [39]. This makes it a potentially ideal target for therapy against PD. Recent preclinical research employing a variety of PD-related neurotoxin-induced in vitro and in vivo models provide clear and convincing proof that the pharmacological manipulation of the Nrf2/ARE/HO-1 pathway contributes to neuroprotection in PD [4]. However, several limitations and challenges do not permit its use in clinical practice.
4.1. Challenges and Suggestions for Future Research
One of the major challenges in developing personalized medicine of Nrf2-based neuroprotective therapies for PD. PD is characterized by a wide range of symptoms and disease progression rates, and there is increasing evidence that different subtypes of PD may have distinct underlying pathophysiological mechanisms. This heterogeneity makes it difficult to develop a “one-size-fits-all” therapy for PD [40], and may explain why some clinical trials targeting the Nrf2 pathway have shown inconsistent results. Identifying patient-specific biomarkers, genetic factors, and environmental influences that affect Nrf2 activity and neuroprotection is critical for developing personalized treatment strategies that can maximize the efficacy of Nrf2 activators in PD.
There is an ongoing debate over whether animal models of PD are accurate representations of the disease’s course in humans. In this regard, the end stage of Parkinson’s disease, where non-dopaminergic neurodegeneration also appears, is not replicated by the mouse model. Furthermore, Lewy body development has never been reported in MPTP-treated animals, suggesting a significant distinction between the pathophysiology of idiopathic PD and MPTP-induced parkinsonism. Furthermore, the initial short-term toxic impact of MPTP in mice is limited to temporary behavioral abnormalities. Within 24 to 48 hours, they recover from hypersalivation, convulsions, piloerection, and hypokinesia. Lastly, we must keep in mind that whereas PD in people develops over decades, all changes driven by MPTP injection in mice manifest in a matter of days or weeks [41].
Another challenge in targeting the Nrf2 pathway for neuroprotection in PD is the limited ability of Nrf2 activators to penetrate the blood-brain barrier (BBB) [42]. The BBB is a highly selective barrier that regulates the entry of molecules into the brain, and many potential Nrf2 activators are large, hydrophobic compounds that have poor bioavailability and limited ability to cross the BBB. This limits their effectiveness as neuroprotective agents in PD, as they may not reach therapeutic levels in the brain when administered systemically [43]. For example, some Nrf2 activators can also activate other pathways, such as the nuclear factor kappa B (NF-κB) pathway, which is a pro-inflammatory pathway that may exacerbate neuroinflammation in PD [44]. Moreover, since Nrf2 activation has so far been shown to be protective against neurological disorders, the creation of Keap1-Nrf2 PPI inhibitors may one day be used to treat PD [45]. Its high polarity, however, makes it difficult to overcome. As a result, less polar group substitution is required, and predrugs and targeted system delivery designs would be further enhanced. Developing selective Nrf2 activators that specifically target the Nrf2 pathways without activating off-target pathways is a major challenge in the field.
The optimal timing for intervention with Nrf2 activators in PD is still unclear. Studies have shown that Nrf2 activation can be neuroprotective in animal models of PD when administered before the onset of symptoms, but the efficacy of Nrf2 activators in preventing or reversing established neurodegeneration in PD is less well studied [46]. The progressive nature of PD makes early intervention critical for maximizing neuroprotection, but identifying biomarkers that can reliably predict the onset of PD or the progression of the disease remains a question. Longitudinal studies are needed to investigate the effects of Nrf2 activation at different stages of PD and to determine the optimal treatment regimen. In addition to targeting the Nrf2 pathway, it is important to consider other factors that contribute to the pathogenesis of PD, such as protein aggregation, mitochondrial dysfunction, and neuroinflammation [47]. Combination therapies that target multiple pathways implicated in PD may be more effective in slowing disease progression and improving outcomes for PD patients.
Developing reliable biomarkers for monitoring Nrf2 activity and neuroprotection in PD is another major issue in the field. Currently, there are no validated biomarkers that can accurately assess the activation of the Nrf2 pathway in the brain, and the effects of Nrf2 activators on neuroprotection are often assessed using indirect measures such as changes in oxidative stress markers or expression of Nrf2 target genes [43]. Developing imaging techniques or biochemical assays that can directly measure Nrf2 activity in the brain would greatly facilitate the evaluation of Nrf2 activators as neuroprotective agents in PD in clinical trials.
A key difficulty in developing Nrf2-based therapies for PD is distinguishing between disease modification and symptomatic treatment. While Nrf2 activation may provide neuroprotection by reducing oxidative stress and inflammation, it is not clear whether Nrf2 activators can prevent the underlying neurodegeneration that drives the progression of PD [48]. Furthermore, the symptomatic relief provided by Nrf2 activators may mask the progression of the disease, making it difficult to assess the long-term effects of these therapies on disease progression [45,47]. Developing biomarkers that can differentiate between disease modification and symptomatic treatment is essential for evaluating the efficacy of Nrf2 activators in PD.
PD patients are already on multiple medications for the management of motor and non-motor symptoms, and there is a risk of drug-drug interactions with Nrf2 activators that can affect the safety and efficacy of these therapies. For example, some Nrf2 activators are metabolized by the cytochrome P450 enzymes, which are also responsible for metabolizing many commonly prescribed medications [49]. Drug-drug interactions that affect the pharmacokinetics of Nrf2 activators may alter their therapeutic levels in the brain, leading to unpredictable outcomes and potential side effects [45]. Understanding the potential drug-drug interactions of Nrf2 activators in PD patients is essential for their safe and effective use in the clinic.
Particulaly when it comes to screening natural product treatments based on their cytototy, pharmacological effects, and chemical properties, the discipline of microfluidics is an emerging player in the development of innovative techniques for drug discovery and screening. Microfluidic platforms have been developed to investigate specific molecular mechanisms associated with PD, such as ROS production and mitochondrial dysfunction [50]. These platforms have shown PD-relevant phenotypes, such as the diffusion of ROS from activated microglia to healthy neuroglioma cells. They have also been used to study the transport of mitochondria along dopaminergic axons and the mitochondrial dysfunctions associated with a genetic form of PD with dynamin-related GTPase optic atrophy type 1 (OPA1) mutations [51]. However, these newly developed microfluidic platforms and in vitro models of PD have not yet been used to study the impact of the Nrf2/HO-1 activator [4].The precise molecular mechanism of the Nrf2/HO-1 activator that regulates Nrf2 activation and HO-1 induction in thse newly developed in vitro models of PD is expected to be clarified by more research in order to develop novel drugs that target Nrf2/ARE/HO-1.
5. Conclusions
The Keap1–Nrf2-ARE signaling pathway holds great potential as a target for oxidative stress-related disease prevention and treatment. Nrf2 is an attractive therapy option for mitochondrial dysfunction in PD since it may have many important roles in mitochondrial function. One prospective treatment for PD is the activation of Nrf2 by naturally occurring bioactive substances. However the absence of reliable methods and tool compounds, incomplete comprehension of downstream regulators of neuroprotection or toxicity, and information shortages about the function of this system in idiopathic PD for clinical trial design and patient selection, however, make this endeavor difficult. The future of Nrf2 signaling is exciting and transformative because of its neuroprotective potential to improve patient outcomes and modify disease treatment.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study.
Conflicts of Interest
The author declares no conflicts of interest.
References
- Marino, B.L.B.; de Souza, L.R.; Sousa, K.P.A.; Ferreira, J.V.; Padilha, E.C.; da Silva, C.; Taft, C.A.; Hage-Melim, L.I.S. Parkinson’s Disease: A Review from Pathophysiology to Treatment. Mini reviews in medicinal chemistry 2020, 20, 754–767. [Google Scholar] [CrossRef] [PubMed]
- Mylius, V.; Möller, J.C.; Bohlhalter, S.; Ciampi de Andrade, D.; Perez Lloret, S. Diagnosis and Management of Pain in Parkinson’s Disease: A New Approach. Drugs & aging 2021, 38, 559–577. [Google Scholar] [CrossRef]
- Rossi, A.; Berger, K.; Chen, H.; Leslie, D.; Mailman, R.B.; Huang, X. Projection of the prevalence of Parkinson’s disease in the coming decades: Revisited. Movement disorders : official journal of the Movement Disorder Society 2018, 33, 156–159. [Google Scholar] [CrossRef]
- Wang, Y.; Gao, L.; Chen, J.; Li, Q.; Huo, L.; Wang, Y.; Wang, H.; Du, J. Pharmacological Modulation of Nrf2/HO-1 Signaling Pathway as a Therapeutic Target of Parkinson’s Disease. Frontiers in pharmacology 2021, 12, 757161. [Google Scholar] [CrossRef]
- Ngo, V.; Duennwald, M.L. Nrf2 and Oxidative Stress: A General Overview of Mechanisms and Implications in Human Disease. Antioxidants (Basel, Switzerland) 2022, 11. [Google Scholar] [CrossRef]
- Panieri, E.; Telkoparan-Akillilar, P.; Suzen, S.; Saso, L. The NRF2/KEAP1 Axis in the Regulation of Tumor Metabolism: Mechanisms and Therapeutic Perspectives. Biomolecules 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Saha, S.; Buttari, B.; Profumo, E.; Tucci, P.; Saso, L. A Perspective on Nrf2 Signaling Pathway for Neuroinflammation: A Potential Therapeutic Target in Alzheimer’s and Parkinson’s Diseases. Frontiers in cellular neuroscience 2021, 15, 787258. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Zhong, Y.; Gao, B.; Zheng, B.; Liu, Y. Nrf2-mediated therapeutic effects of dietary flavones in different diseases. Frontiers in pharmacology 2023, 14, 1240433. [Google Scholar] [CrossRef]
- Niu, Y.; Zhang, J.; Dong, M. Nrf2 as a potential target for Parkinson’s disease therapy. Journal of molecular medicine (Berlin, Germany) 2021, 99, 917–931. [Google Scholar] [CrossRef]
- Chakkittukandiyil, A.; Sajini, D.V.; Karuppaiah, A.; Selvaraj, D. The principal molecular mechanisms behind the activation of Keap1/Nrf2/ARE pathway leading to neuroprotective action in Parkinson’s disease. Neurochemistry international 2022, 156, 105325. [Google Scholar] [CrossRef]
- Moi, P.; Chan, K.; Asunis, I.; Cao, A.; Kan, Y.W. Isolation of NF-E2-related factor 2 (Nrf2), a NF-E2-like basic leucine zipper transcriptional activator that binds to the tandem NF-E2/AP1 repeat of the beta-globin locus control region. Proceedings of the National Academy of Sciences of the United States of America 1994, 91, 9926–9930. [Google Scholar] [CrossRef]
- Tong, K.I.; Katoh, Y.; Kusunoki, H.; Itoh, K.; Tanaka, T.; Yamamoto, M. Keap1 recruits Neh2 through binding to ETGE and DLG motifs: characterization of the two-site molecular recognition model. Molecular and cellular biology 2006, 26, 2887–2900. [Google Scholar] [CrossRef] [PubMed]
- Lo, S.C.; Li, X.; Henzl, M.T.; Beamer, L.J.; Hannink, M. Structure of the Keap1:Nrf2 interface provides mechanistic insight into Nrf2 signaling. The EMBO journal 2006, 25, 3605–3617. [Google Scholar] [CrossRef] [PubMed]
- Tonelli, C.; Chio, I.I.C.; Tuveson, D.A. Transcriptional Regulation by Nrf2. Antioxidants & redox signaling 2018, 29, 1727–1745. [Google Scholar] [CrossRef]
- Chowdhry, S.; Zhang, Y.; McMahon, M.; Sutherland, C.; Cuadrado, A.; Hayes, J.D. Nrf2 is controlled by two distinct β-TrCP recognition motifs in its Neh6 domain, one of which can be modulated by GSK-3 activity. Oncogene 2013, 32, 3765–3781. [Google Scholar] [CrossRef]
- Telkoparan-Akillilar, P.; Suzen, S.; Saso, L. Pharmacological Applications of Nrf2 Inhibitors as Potential Antineoplastic Drugs. International journal of molecular sciences 2019, 20. [Google Scholar] [CrossRef] [PubMed]
- Levy, O.A.; Malagelada, C.; Greene, L.A. Cell death pathways in Parkinson’s disease: proximal triggers, distal effectors, and final steps. Apoptosis : an international journal on programmed cell death 2009, 14, 478–500. [Google Scholar] [CrossRef]
- Chen, X.; Yu, C.; Kang, R.; Tang, D. Iron Metabolism in Ferroptosis. Frontiers in cell and developmental biology 2020, 8, 590226. [Google Scholar] [CrossRef]
- Chorley, B.N.; Campbell, M.R.; Wang, X.; Karaca, M.; Sambandan, D.; Bangura, F.; Xue, P.; Pi, J.; Kleeberger, S.R.; Bell, D.A. Identification of novel NRF2-regulated genes by ChIP-Seq: influence on retinoid X receptor alpha. Nucleic acids research 2012, 40, 7416–7429. [Google Scholar] [CrossRef]
- Chen, P.C.; Vargas, M.R.; Pani, A.K.; Smeyne, R.J.; Johnson, D.A.; Kan, Y.W.; Johnson, J.A. Nrf2-mediated neuroprotection in the MPTP mouse model of Parkinson’s disease: Critical role for the astrocyte. Proceedings of the National Academy of Sciences of the United States of America 2009, 106, 2933–2938. [Google Scholar] [CrossRef]
- Itoh, K.; Wakabayashi, N.; Katoh, Y.; Ishii, T.; Igarashi, K.; Engel, J.D.; Yamamoto, M. Keap1 represses nuclear activation of antioxidant responsive elements by Nrf2 through binding to the amino-terminal Neh2 domain. Genes & development 1999, 13, 76–86. [Google Scholar] [CrossRef]
- Dinkova-Kostova, A.T.; Abramov, A.Y. The emerging role of Nrf2 in mitochondrial function. Free radical biology & medicine 2015, 88, 179–188. [Google Scholar] [CrossRef]
- Tastan, B.; Arioz, B.I.; Genc, S. Targeting NLRP3 Inflammasome With Nrf2 Inducers in Central Nervous System Disorders. Frontiers in immunology 2022, 13, 865772. [Google Scholar] [CrossRef] [PubMed]
- Yim, J.; Hwang, Y.S.; Lee, J.J.; Kim, J.H.; Baek, J.Y.; Jeong, J.; Choi, Y.I.; Jin, B.K.; Park, S.B. Inflachromene ameliorates Parkinson’s disease by targeting Nrf2-binding Keap1. Chemical science 2024, 15, 3588–3595. [Google Scholar] [CrossRef]
- Pawelec, P.; Ziemka-Nalecz, M.; Sypecka, J.; Zalewska, T. The Impact of the CX3CL1/CX3CR1 Axis in Neurological Disorders. Cells 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Gan, L.; Johnson, D.A.; Johnson, J.A. Keap1-Nrf2 activation in the presence and absence of DJ-1. The European journal of neuroscience 2010, 31, 967–977. [Google Scholar] [CrossRef]
- Henrich, M.T.; Oertel, W.H.; Surmeier, D.J.; Geibl, F.F. Mitochondrial dysfunction in Parkinson’s disease—a key disease hallmark with therapeutic potential. Molecular neurodegeneration 2023, 18, 83. [Google Scholar] [CrossRef]
- Kasai, S.; Shimizu, S.; Tatara, Y.; Mimura, J.; Itoh, K. Regulation of Nrf2 by Mitochondrial Reactive Oxygen Species in Physiology and Pathology. Biomolecules 2020, 10. [Google Scholar] [CrossRef]
- Xiao, W.; Wang, R.S.; Handy, D.E.; Loscalzo, J. NAD(H) and NADP(H) Redox Couples and Cellular Energy Metabolism. Antioxidants & redox signaling 2018, 28, 251–272. [Google Scholar] [CrossRef]
- Sun, W.; Zheng, J.; Ma, J.; Wang, Z.; Shi, X.; Li, M.; Huang, S.; Hu, S.; Zhao, Z.; Li, D. Increased Plasma Heme Oxygenase-1 Levels in Patients With Early-Stage Parkinson’s Disease. Frontiers in aging neuroscience 2021, 13, 621508. [Google Scholar] [CrossRef]
- Schipper, H.M. Heme oxygenase expression in human central nervous system disorders. Free radical biology & medicine 2004, 37, 1995–2011. [Google Scholar] [CrossRef]
- Choi, S.Y.; Son, T.G.; Park, H.R.; Jang, Y.J.; Oh, S.B.; Jin, B.; Lee, J. Naphthazarin has a protective effect on the 1-methyl-4-phenyl-1,2,3,4-tetrahydropyridine-induced Parkinson’s disease model. Journal of neuroscience research 2012, 90, 1842–1849. [Google Scholar] [CrossRef]
- Lu, K.T.; Ko, M.C.; Chen, B.Y.; Huang, J.C.; Hsieh, C.W.; Lee, M.C.; Chiou, R.Y.; Wung, B.S.; Peng, C.H.; Yang, Y.L. Neuroprotective effects of resveratrol on MPTP-induced neuron loss mediated by free radical scavenging. Journal of agricultural and food chemistry 2008, 56, 6910–6913. [Google Scholar] [CrossRef]
- Chen, J.; Tang, X.Q.; Zhi, J.L.; Cui, Y.; Yu, H.M.; Tang, E.H.; Sun, S.N.; Feng, J.Q.; Chen, P.X. Curcumin protects PC12 cells against 1-methyl-4-phenylpyridinium ion-induced apoptosis by bcl-2-mitochondria-ROS-iNOS pathway. Apoptosis : an international journal on programmed cell death 2006, 11, 943–953. [Google Scholar] [CrossRef] [PubMed]
- Rathore, A.S.; Singh, S.S.; Birla, H.; Zahra, W.; Keshri, P.K.; Dilnashin, H.; Singh, R.; Singh, S.; Singh, S.P. Curcumin Modulates p62-Keap1-Nrf2-Mediated Autophagy in Rotenone-Induced Parkinson’s Disease Mouse Models. ACS chemical neuroscience 2023. [Google Scholar] [CrossRef] [PubMed]
- Cao, Q.; Zou, Q.; Zhao, X.; Zhang, Y.; Qu, Y.; Wang, N.; Murayama, S.; Qi, Q.; Hashimoto, K.; Lin, S.; et al. Regulation of BDNF transcription by Nrf2 and MeCP2 ameliorates MPTP-induced neurotoxicity. Cell death discovery 2022, 8, 267. [Google Scholar] [CrossRef]
- Jazwa, A.; Rojo, A.I.; Innamorato, N.G.; Hesse, M.; Fernández-Ruiz, J.; Cuadrado, A. Pharmacological targeting of the transcription factor Nrf2 at the basal ganglia provides disease modifying therapy for experimental parkinsonism. Antioxidants & redox signaling 2011, 14, 2347–2360. [Google Scholar] [CrossRef]
- Tseng, H.C.; Wang, M.H.; Fang, C.H.; Lin, Y.W.; Soung, H.S. Neuroprotective Potentials of Berberine in Rotenone-Induced Parkinson’s Disease-like Motor Symptoms in Rats. Brain sciences 2024, 14. [Google Scholar] [CrossRef]
- Huang, Y.; Li, W.; Su, Z.Y.; Kong, A.N. The complexity of the Nrf2 pathway: beyond the antioxidant response. The Journal of nutritional biochemistry 2015, 26, 1401–1413. [Google Scholar] [CrossRef]
- Sturchio, A.; Rocha, E.M.; Kauffman, M.A.; Marsili, L.; Mahajan, A.; Saraf, A.A.; Vizcarra, J.A.; Guo, Z.; Espay, A.J. Recalibrating the Why and Whom of Animal Models in Parkinson Disease: A Clinician’s Perspective. Brain sciences 2024, 14. [Google Scholar] [CrossRef]
- Schmidt, N.; Ferger, B. Neurochemical findings in the MPTP model of Parkinson’s disease. Journal of neural transmission (Vienna, Austria : 1996) 2001, 108, 1263–1282. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.T.; Dodson, M. The untapped potential of targeting NRF2 in neurodegenerative disease. Frontiers in aging 2023, 4, 1270838. [Google Scholar] [CrossRef]
- Yagishita, Y.; Gatbonton-Schwager, T.N.; McCallum, M.L.; Kensler, T.W. Current Landscape of NRF2 Biomarkers in Clinical Trials. Antioxidants (Basel, Switzerland) 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Guo, L.; Yang, Y.; Wang, Y.; Xia, S.; Gong, H.; Zhang, B.K.; Yan, M. Dissecting the Crosstalk Between Nrf2 and NF-κB Response Pathways in Drug-Induced Toxicity. Frontiers in cell and developmental biology 2021, 9, 809952. [Google Scholar] [CrossRef] [PubMed]
- Robledinos-Antón, N.; Fernández-Ginés, R.; Manda, G.; Cuadrado, A. Activators and Inhibitors of NRF2: A Review of Their Potential for Clinical Development. Oxidative medicine and cellular longevity 2019, 2019, 9372182. [Google Scholar] [CrossRef]
- Wang, X.; Kang, N.; Liu, Y.; Xu, G. Formononetin Exerts Neuroprotection in Parkinson’s Disease via the Activation of the Nrf2 Signaling Pathway. Molecules (Basel, Switzerland) 2024, 29. [Google Scholar] [CrossRef]
- Mayer, C.; Riera-Ponsati, L.; Kauppinen, S.; Klitgaard, H.; Erler, J.T.; Hansen, S.N. Targeting the NRF2 pathway for disease modification in neurodegenerative diseases: mechanisms and therapeutic implications. Frontiers in pharmacology 2024, 15, 1437939. [Google Scholar] [CrossRef]
- Dinkova-Kostova, A.T.; Copple, I.M. Advances and challenges in therapeutic targeting of NRF2. Trends in pharmacological sciences 2023, 44, 137–149. [Google Scholar] [CrossRef]
- Ashino, T.; Yamamoto, M.; Numazawa, S. Nrf2 Antioxidative System is Involved in Cytochrome P450 Gene Expression and Activity: A Delay in Pentobarbital Metabolism in Nrf2-Deficient Mice. Drug metabolism and disposition: the biological fate of chemicals 2020, 48, 673–680. [Google Scholar] [CrossRef]
- Kane, K.I.W.; Moreno, E.L.; Hachi, S.; Walter, M.; Jarazo, J.; Oliveira, M.A.P.; Hankemeier, T.; Vulto, P.; Schwamborn, J.C.; Thoma, M.; et al. Automated microfluidic cell culture of stem cell derived dopaminergic neurons. Scientific reports 2019, 9, 1796. [Google Scholar] [CrossRef]
- Bolognin, S.; Fossépré, M.; Qing, X.; Jarazo, J.; Ščančar, J.; Moreno, E.L.; Nickels, S.L.; Wasner, K.; Ouzren, N.; Walter, J.; et al. 3D Cultures of Parkinson’s Disease-Specific Dopaminergic Neurons for High Content Phenotyping and Drug Testing. Advanced science (Weinheim, Baden-Wurttemberg, Germany) 2019, 6, 1800927. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Summary of Key features of Nrf2.
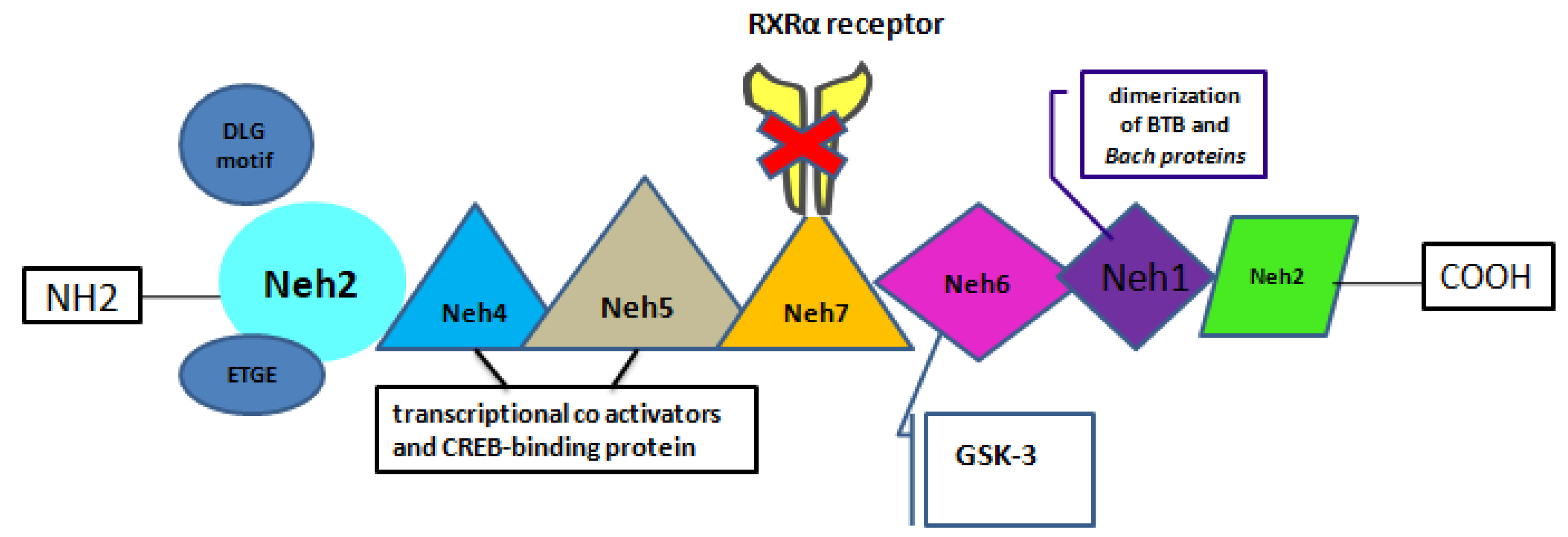
Figure 2.
Overview of the key points of Nrf2 neuroprotective role for PD.
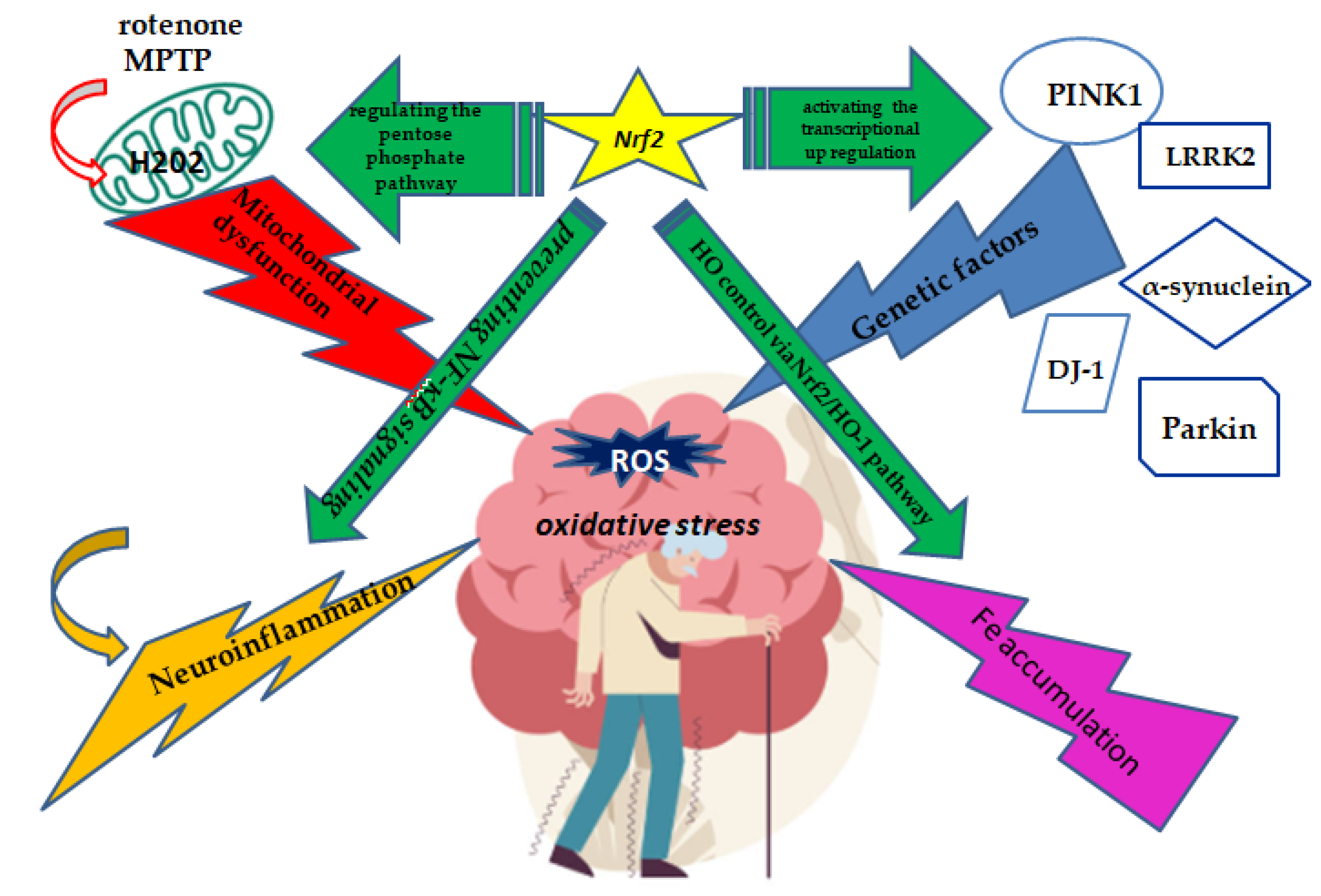

Table 1.
Overview of selected studies demonstrating the Nrf2 neuroprotective effect.
| Authors, year | Agent | Model | AdministrationRoute/ Dosage | Assessment tools/ measures | Neuroprotective Effect |
|---|---|---|---|---|---|
| Chen et al. 2006 | curcumin | PC12 cells | MTT solution (final concentration, 0.5 mg/ml) for 4 h | FCM iNOS and Bcl-2 |
decrease in cell viability in PC12 cells and overexpression of Bcl-2 |
| Jazwa et al. 2011 | SFN | Nrf2 + / + and Nrf2—/ - mice | Intraperitoneal | protein levels of MAO-B DAT |
Attenuates the MPTP-induced dopaminergic neuron damage and astrogliosis |
| Cao et al.2022 | SFN |
MPP+ treated SH-SY5Y cells
MPTP-treated mouse model |
i.p SFN (10 mg/kg) and MPTP (30 mg/kg) | rotarod test | Ameliorate dopaminergic neurotoxicity via activation of BDNF and suppression of MeCP2 |
| Rathore et al. 2023 | curcumin | subcutaneous injection | Nrf2, Keap1, p62, LC3, Bcl2, Bax, and caspase 3 | autophagy-mediated clearance of misfolded α-syn proteins by increasing the LC3-II expression and blocked apoptotic cascade | |
| Tseng et al. 2024 | BBR | RTN model rat |
subcutaneous RTN at 0.5 mg/kg for 21 days orally BBR at 30 or 100 mg/kg doses |
open-field, bar catalepsy, beam-crossing, rotarod, and grip strength TNF-α, IL-1β, and IL-6 levels |
modulation of the Nrf2-mediated pathway via the activation of PI3K/Akt, p38, and HO-1 expressions |
BBR: Berberine; BDNF: brain-derived neurotrophic factor ; ICV: Intracerebroventricular, FCM MMP: mitochondrial membrane potential; MPTP: methyl-4-phenyl-1,2,3,6-tetrahydropyridine; MPP+: ;Nrf2: nuclear factor erythroid-2-related factor 2; SFN: Sulforaphane.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



