
Submitted:
17 February 2025
Posted:
18 February 2025
You are already at the latest version
Abstract
The genus Trichogramma Westwood (Hymenoptera: Trichogrammatidae) includes insect egg parasitoids widely used worldwide as biological control agent of pests. The success of these parasitoids in pest management depends, among other factors, on their adaptation to the climatic conditions of the release area, particularly temperature. This study aimed to identify the Trichogramma species naturally occurring in Hungary. Parasitism was observed by trapping Trichogramma spp. individuals using bait cards containing Sitotroga ceraella eggs (provided by the AMW company) in apple orchard between July 8, 2024, and September 30, 2024. We evaluated the relationship between several variables – the rate of cards with observed parasitism, the parasitism rate, the number of host eggs and the hatching rate – and the time of release, the age of the host eggs and meteorological factors. We found that the parasitism rate significantly depends on the time of release. Pearson's correlation coefficients indicated that temperature parameters (average minimum, maximum and the mean temperatures during the exposure period of the cards) are significantly and negatively correlated with both parasitism rate and number of parasitised eggs. Air pressure showed a significant positive correlation with the number of parasitised eggs. The naturally occurring egg parasitoid species was identified as T. evanescens. These results suggest that these parasitoids are well adapted to the local climate and can be considered as candidates for pest management programs in Hungary and in other countries with a temperate continental climate.
Keywords:
Trichogramma
; native
; insect parasitoids
; host bait cards
; biological control
1. Introduction
Trichogramma wasps (Hymenoptera: Trichogrammatidae) are egg parasitoid insects that primarily parasitise the eggs of Lepidoptera, including moths. However, certain species of Trichogramma also parasitise eggs of insects from other orders (Coleoptera, Diptera, Heteroptera, Hymenoptera, Neuroptera) [1].
Generally, most Trichogramma spp. individuals are between 0.4 and 0.6 mm long, with females being slightly larger than males [2]. Trichogramma wasps have haplodiploid sex determination: unfertilised eggs develop into males, while fertilised eggs develop into females [3,4,5]. The female wasp locates host eggs with chemicals and visual clues. The chemical clues, called kairomones, are on the moth scales that the female moth leaves near the egg during the oviposition [1,5]Female parasitoids obtain sugars and certain proteins by utilising floral and extrafloral nectar, pollen, and honeydew produced by insects [6]. Parasitoid female wasps can also obtain nutrients from the host by feeding on the egg fluids that ooze from the wounds inflicted by the ovipositor [7,8,9,10]. Host feeding is important for the fecundity of the parasitoids [11,12,13]. The larvae undergo three developmental stages and pupates inside the host eggs. Parasitised host eggs undergo a change in coloration, becoming black, which makes them easily recognizable [1,11]. Trichogramma spp. overwinter via quiescence, diapause or a combination of these two strategies as immature stages within their host eggs [12].
Trichogramma wasps naturally occur in a wide range of terrestrial and aquatic habitats. Some of the most harmful caterpillar pests of field crops, forests, and fruit and nut trees are subject to attack by Trichogramma spp. However, in the majority of crop production systems, the number of caterpillar eggs destroyed by native Trichogramma populations is insufficient to prevent the pest from reaching damaging levels [1]. The use Trichogramma wasps in biological control began at the turn of the 20th century. In 1930 Flanders [13] developed a method of rearing Trichogramma wasps on eggs of the Angoumois grain moth Sitotroga cerealella [14]. Natural enemies offer several advantages over chemical pesticides: (1) Toxic residues on plants and soil are eliminated. (2) Application is simple. (3) Resistance by pests is prevented. (4) They are effective if used correctly. (5) They are harmless to natural enemies of pests. (6) The production of these items is possible in any country, including those with restricted access to traditional techniques [11]. The success of Trichogramma wasps in crop protection is favoured by its very short generation times, simple mass rearing systems, sustained economic efficiency and technologies adapted for commercial use [11,15]. Initially Trichogramma spp. are being mass-reared to control pests on corn, sugarcane, rice, cotton, soybean, sugar-beet, vegetables and pine [11] Today, the Trichogramma wasps became an integral part of pest management strategies to control agricultural and forest pests throughout the world, including the Americas [16,17], Europe [18,19], Africa [20,21,22] and Asia [23,24]
The process of parasitism by insect parasitoids comprises a series of interconnected steps, the outcome of which is influenced by a multitude of intrinsic and external factors [25,26]. The parasitism characteristics of Trichogramma species (parasitism, hatching and female offspring rates) are influenced by a number of abiotic (temperature, light and photoperiod) [3,27,28] and biotic factors (host species, age of individuals, age and number of female wasps, available food source) [3,29,30].
There is a growing interest in using indigenous Trichogramma species for pest control, because they are thought to be better suited to the local climatic conditions [31] and there are concerns about releasing exotic species [32].
Apple (Malus domestica Borkh) is considered to be the world’s oldest fruit crop. Apple is attacked by a large number of insect pests, including moths, such as the Codling Moth, Cydia pomonella and leaf miners [33]. In attempts to control C. pomonella, five species of Trichogramma wasps have been used: T. platneri Nagarkatti [34,35] T. minutum Riley and T. pretiosum Riley [36,37,38], T. dendrolimi Matsumura and T. cacoeciae (Marchal) [39,40].
The results show that Trichogramma wasps can be used effectively against a number of pests that are prevalent also in Hungary, such as Tomato Leaf Miner (Tuta absoluta) [41]; Tomato Fruit Borer, (Helicoverpa armigra) [42]; oriental fruit moth (Grapholita molesta) [43] and European corn borer (Ostrinia nubilalis) [19]. However, information about the efficacy of Trichogramma species in Hungarian apple orchards is limited. T. evanescens Westwood and T. cacoeciae Marchal species have been reported to infest C. pomonella and leafrollers [44,45,46].
The area under apple cultivation in Hungary is currently around 33,000 hectares [47]. Plant protection products based on Trichogramma species that are currently on the market are approved for use in maize and vegetable crops, and there is no Trichogramma product available for apple crops in Hungary [48]. Our goal is to survey the naturally occurring Trichogramma species in a Hungarian apple orchard. Our results could provide with the scientific basis for the development of a Trichogramma-based crop protection product in Hungary and other countries with a temperate continental climate.
2. Materials and Methods
2.1. Orchards/Location
The experiment was conducted in an apple (Malus domestica) orchard, located in the middle of the country, at Bács-Kiskun county, near to Kecskemét (GPS: 46°54'39.3"N 19°47'04.6"E). The age of the trees was 21 years, the row and stem distance were 4.5 x 2 m. The following varieties were present in the experimental area, with the varieties for which the bait cards were placed being marked in bold: Red Rose, Ten Rose, Ginson Golden, Mutsu, Regal Prince, Idared, Jonagold, Gloster, Close, Akane, Vista Bella. In addition to the mowing of plant vegetation between the rows, no other agrotechnical or plant protection methods were carried out in the experimental area and no previous applications of Trichogramma releases have been conducted. The experiment was set up between July 8, 2024, and September 30, 2024. The meteorological data were recorded at the National Meteorological Service meteorological station (GPS: 46°54'43.9"N 19°45'34.9"E).
2.2. Bait Cards
Folded cardboard bait cards containing Sitotroga cereaella eggs were used to collect the Trichogramma wasps. The bait cards and the eggs were provided by the company Appel-Melchior-Wührer (AMW, https://www.amwnuetzlinge.de/en/about-amw/about-us/). The age of the Sitotroga cereaella eggs used in the experiment ranged from one to four weeks. Prior to field use, the bait cards were stored at 6 °C to prevent S.cerealella eggs from hatching. The eggs were placed on the cards immediately prior to reliese (within 1 hour). To protect S. cereaella eggs from predators, the cards were covered with a net, that still allowed the Trichogramma wasps to enter.
2.3. Experimental Method/Field Collection
A total of 50 cards were placed at 10 different points on a weekly basis. The GPS coordinates of each placement point were recorded. The cards were replaced weekly. Following the collection of the bait cards, they were stored at room temperature (at 22–26 °C) for a further 2–3 days. Then, on the second or third day after collection, the eggs were examined under a stereomicroscope to check for parasitism. The parasitised eggs were closed in a 2 cm diameter, 4.5 cm high glass vial until the adults hatched. The hatched adults together with the parasitised but unhatched eggs were placed into 99% alcohol and stored in a freezer (at -20 °C) until molecular identification. The number of discovered cards (i.e. the number of cards containing at least one parasitised egg), the total number of eggs on the cards, the number of parasitised eggs, and the number of hatched individuals were recorded.
2.4. Molecular Identification
2.4.1. Polymerase Chain Reaction (PCR) of ITS2 Region
Thermo Scientific Phire Tissue Direct PCR Master Mix was used to amplify the ITS2 region using primers described by Stouthamer et al. [49]. Dilution Protocol was chosen and 5 Individuals (preserved in 99.5% Ethanol, stored at -20 °C) were placed into 20 μL of Dilution Buffer, add 0.5 μL of DNARelease Additive and mixed by vortexing the tube briefly, and spin down the solution. Incubated the reaction for 2−5 minutes at room temperature and then placed the tube into the pre-heated (98 °C) block for 2 minutes. PCR reaction was performed in 20 µl volumes containing H2O (20 µl), 2X Phire Tissue Direct PCR Master Mix (10 µl), Primer A (1 µl), Primer B (1 µl), Sample (1 µl). The PCR cycling programme was 5 min at 98 °C, followed by 40 cycles of 5 s at 98 °C, 5 s at 62.2 °C and 20 s at 72 °C, with 1 min at 72 °C after the last cycle. PCR products were visualised after electrophoresis on 1.5% agarose gel stained with Midorii green Advanced, to confirm amplification.
2.4.2. Restriction Analysis
For the PCR products restriction enzymes were used to find specific differences between the samples. Trichogramma wasps were identified using a proprietary method developed by AMW.
2.5. Data Analysis
Discovery rate (%) was calculated as the rate of cards detected (i.e. the number of cards containing at least one parasitised egg divided by the total number of cards released). Parasitism rate was calculated as the ratio of parasitised eggs to the total number of eggs released. Hatching rate (%) was calculated as the number of hatched eggs divided by the total number of parasitised eggs. Discovery rates (%), parasitism rates, hatching rates (%), age of host eggs (days) and the numbers of host eggs (pcs) were discretised according to Table 1.
From the daily data of mean, maximum and mean temperatures (°C), mean relative humidity (%), mean wind speed (m/sec) and mean air pressure (hPa), we calculated their averages in relation to the time intervals of the cards released.
We used a Chi-square test to find out whether parasitism rate is independent from the age of the eggs, the time of release, the number of the host eggs. Having significant result, we used adjusted standardised residuals (AdjRes) to detect under- and over-represented cases, with the criteria of the values being above 1.96 or below -1.96, respectively.
We used IBM SPSS Statistical software (v 29, Armonk, New York, 2023).
3. Results
First, we tested whether the placement points have a spatial block effect on parasitism rate or the hatching rate. Chi-square test indicated that both the parasitism and the hatching rates were independent from the placement of the cards (χ²(9)=6,221; p=0.718; χ²(27)=15.591; p=0.960, respectively).
3.1. The Dependence of Discovery Rate and Parasitism Rate on the Time of Release
Chi-square test indicated that the discovery rate was significantly dependent on the time of release (Fisher’s test value=10.49; p<0.05). According to the adjusted residuals, medium discovery rates (between 6–15%) were significantly more frequent between 21 August and 17 September 2024 (AdjRes >2) while high discovery rates (above 15%) were significantly more frequent between 24 September and 1 October 2024 (AdjRes >3).
The Chi-squared test showed that the detection rate was significantly dependent on the time of release (χ²(4)=18.549; p<0.001). According to the adjusted residuals, the low parasitism rates (between 0.001–0.1%) were significantly more frequent between 8 July 2024 and 13 August 2024 (AdjRes >2), while the high parasitism rates (between 0.101% and 0.4%) were significantly less frequent in this period (AdjRes <-2). In autumn, between 18 September and 1 October 2024, the high parasitism rates (between 0.101 and 0.4%) were significantly more frequent (AdjRes >2), while the zero-parasitism rate (0%) was significantly less frequent (AdjRes <-2).
3.2. Relationship Between Parasitism Rate and the Number and Age of the Host Eggs
Chi-square test indicated that the parasitism rate is independent from both the number and the age of the host eggs (χ²(8)=4.617; p=0.798; χ²(4)=5.511; p=0.24, respectively).
3.3. Relationship Between Hatching Rate and Time of Release and Age of the Host Eggs
Cases where parasitism was 0 were excluded. Chi-square test indicated that the hatching rate is not independent from the time of release (χ²(6)=26,816; p<0.001). According to the adjusted residuals, the low hatching rate (0%) was significantly more frequent between 8 July 2024 and 13 August 2024 (AdjRes >2) while later, between 17 September and 1 October 2024, high hatching rate (95–99%) was significantly more frequent (AdjRes >2).
Chi-square test indicated that the hatching rate is independent from the age of the host eggs (χ²(6)=4.926; p=0.553).
3.4. Relationship Between Parasitism, the Number of Parasitised Eggs and Meteorological Factors
The results of the correlation analysis between meteorological factors (Figure 1) and the parasitism rates with the number of parasitised eggs (Figure 2) are presented in Table 2.
Pearson's correlation coefficients indicated that the three temperature parameters (minimum, maximum and average temperatures during the time of the release of the cards) are significantly negatively correlated with both parasitism rate and the number of parasitised eggs. Meanwhile, air pressure has a significant positive correlation with the number of parasitised eggs. For relative humidity and wind speed, no significant correlation was found with either parasitism rate or the number of parasitised eggs.
4. Discussion
The natural occurrence of the Trichogramma species has been demonstrated in many locations worldwide, such as T. bourarachae Pintureau and Babault, T. cordubensis Vargas and Cabello, T. pretiosum and T. cacociae from Egypt [50], T. bourarache, T. cordubensis, T. evanescens, T. pintoi Voegele, and T. turkestanica from Portugal [51]. In the studies of Souza et al. [52] in total, 2,242 specimens of Trichogramma species were obtained, belonging to the species T. pretiosum Riley, T. manicobai Brun, Moraes & Soares, T. marandobai Brun, Moraes & Soares, and T. galloi Zucchi in Brazil.
In our studies, natural parasitism of Trichogramma species was observed by trapping with bait cards containing host eggs. The egg parasitoid species was identified as T. evanescens.
T. evanescens has been shown to occur naturally in many places around the world, including northern Moldova [53] , Portugal [51] and Russia [53]. In Europe Bírová [54,55] studied the natural parasitism of European Corn Borer (Ostrinia nubilalis) eggs by Trichogramma wasps in Slovakia as early as the late 1950s and early 1960s.
Bírová observed 65.4% parasitised European Corn Borer eggs in 1956, 49.8% in 1957 and 15.7% in 1958 and found very low parasitism in 1980–1985 [56]. The results of Cagán et al. [57] show that the percentage of natural parasitism of O. nubilalis eggs in south-western Slovakia is generally low, but can reach 20% or more depending on the year. Depending on the habitat, T. cacoeciae, T. embryophagum, T. cephalciae, T. minutum species have been identified in Czechoslovakia [58]. Trichogramma evanescens was observed by Ram [53] also in the eggs of O. nubilalis in Slovakia. The occurrence of four native species, T. cacoeciae, T. daumalae Dugast and Voegelé, T. evanescens and T. principium Sugonjaev and Sorokina were mentioned in a French vineyard by Barnay et al. [59]. In Germany [60], the survey of naturally occurring egg parasitoids using "trap cards" showed that only Trichogramma evanescens was present.
There are few literatures available on naturally occurring Trichogramma species in Hungary. Dudich [61] studied naturally occurring Trichogramma species on European Corn Borer (ECB) in and found no parasitism of Trichogramma spp. on ECB eggs. Nagy [62] confirmed his result and found no more than 1% parasitism in the region 30 km south-west of Budapest in 1976–1982.
In our studies, the activity of Trichogramma by trapping bait cards containing Sitotroga cereaella eggs was observed from July until September. Trichogramma wasps showed the greatest activity in the second part of September. When Barnay et al. [59] studied the spontaneous occurrence of Trichogramma wasps in a French vineyard, the results showed that April–May, July and September were the periods when Trichogramma individuals were the most active. Sengonca and Leisse [63,64] found similar results in Germany, where T. semblidis Auriv. was observed more frequently from mid-May to mid-June and, after a decline, egg parasitism increased again in September and October. In Germany, however, Kot [65] found that the Trichogramma population in an orchard was very high in July, August and the first half of September compared with the spring population. Similarly, in 1962, Bírová [55] observed maximum occurrence of the Trichogramma parasitism in Slovakia during the second and the fourth pentad of July.
As suggested by Kot [65], the degree of infestation of cards may not accurately reflect the number of Trichogramma wasps. If natural host eggs are abundant, Trichogramma wasps may not be eager to parasitise factitious hosts. Cagán et al. [57] found differences between parasitism rates in different years. In 1993, when no eggs were parasitised, there was very hot and dry weather in May, which probably affected the development of the parasitoid. These data suggest that extremely dry weather can have a negative impact on egg parasitoid populations.
In our studies, Trichogramma wasps showed the greatest activity in the second half of September. The results can be explained by the correlation between parasitism and meteorological data. We found that Pearson's correlation coefficients indicate that the three temperature parameters (minimum, maximum and the average temperature during the time of release) are significantly negatively correlated with both parasitism rate and number of parasitised eggs. Air pressure has a significant positive correlation with the number of parasitised eggs. For relative humidity and wind speed, no significant correlation was found with either parasitism rate or number of parasitised eggs. Between 08 July and 17 September, the parasitism rate was low (0–0.1%). During this period, the daily maximum temperature exceeded 34 °C and in some weeks even 37.5 °C. The temperature decreased between 10 and 17 of September, when the maximum temperatures dropped by about 10 °C. In the following two weeks, totally 596 and 333 parasitised eggs were found. Our results are consistent with those of Zouba et al. [66], who investigated the effect of the temperature sensitivity of four Trichogramma species. All Trichogramma species were able to develop and survive from 25 °C to 35 °C , but not at 40 °C. Results showed that temperature affects the lifespan and fertility of the female progeny, with both decreasing as temperature rises from 25 °C to 35 °C. In their laboratory tests, Schöller and Hassan [67] established that the parasitism of the T. evanescens decreased with increasing temperature, but T. evanescens was still able to parasitise hosts at 35 °C. These results support the results of our study, where we found 24 parasitised eggs per week even when the daily average maximum temperature reached almost 38 °C. In contrast to these results, Mohammad et al. [68] found that no development could be observed for T. evanescens at 35 °C . They reported that the optimum temperature for the development of T. evanescens ranged between 22 and 27 °C. Based on the results of Metwally et al. [69], we can estimate the optimum temperature for rearing T. evanescens as 30 °C.
In our results, air pressure has a significant positive correlation with the number of parasitised eggs. Fournier et al. [70] evaluated the effect of barometric pressure changes on flight initiation of female T. evanescens. The Trichogramma wasps did not respond to stable or slow changes in barometric pressure, but rapid barometric changes significantly reduced the flight initiation.
Information on naturally occurring species is needed for planning biological control projects. Natural populations of Trichogramma species could be observed in apple orchard in Hungary. However, their natural pest control potential will not be sufficient in itself to protect the crop, as their natural parasitism rate is low during the hot summer months when the average daily temperature is close to 29 °C. Nevertheless, the fact, that T. evanescens occur naturally in Hungary suggests that these parasitoids are well adapted to the local climate. Consequently, T. evanescens may be considered as potential candidates for inclusion in integrated pest management programs.
Author Contributions
Conceptualization, F.T., L.J. and M.L.; methodology, F.T., L.J., T.K-M. and M.L.; software, L.J. and M.L.; validation, L.J. and M.L.; formal analysis, L.J., T.K-M. and M.L.; investigation, L.J., T.K-M. and A.T.; resources, M.L., T.K-M. and F.T.; data curation, L.J. and M.L.; writing—original draft preparation, L.J. and M.L.; writing—review and editing, L.J. , F.T., T.K-M. and M.L.; visualization, L.J. and M.L.; supervision, F.T. and M.L.; All authors have read and agreed to the published version of the manuscript.”.
Funding
Please add: “This research received no external funding” or “This research was funded by NAME OF FUNDER, grant number XXX” and “The APC was funded by XXX”. Check carefully that the details given are accurate and use the standard spelling of funding agency names at https://search.crossref.org/funding. Any errors may affect your future funding.
Acknowledgments
This work was partly supported by the Research Excellence Programme of the Hungarian University of Agriculture and Life Sciences.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- A Knutson, A. The Trichogramma Manual. Bull. Agric. Ext. Serv. No 6071 1998.
- Pinto J.D.; Systematics of the North American Species of Trichogramma Westwood (Hymenoptera : Trichogrammatidae). Mem Entomol Soc Wash 1999, 22, 1–287.
- Yan, Z.; Yue, J.-J.; Zhang, Y.-Y. Biotic and Abiotic Factors That Affect Parasitism in Trichogramma Pintoi (Hymenoptera: Trichogrammatidae) as a Biocontrol Agent against Heortia Vitessoides (Lepidoptera: Pyralidae). Environ. Entomol. 2023, 52 (3), 301–308.
- Heimpel, G. E.; Boer, J. G. de. Sex Determination in the Hymenoptera. Annu. Rev. Entomol. 2008, 53 (Volume 53, 2008), 209–230. [CrossRef]
- Lewis, W. J.; Nordlund, D. A.; Gueldner, R. C.; Teal, P. E. A.; Tumlinson, J. H. Kairomones and Their Use for Management of Entomophagous Insects. J. Chem. Ecol. 1982, 8 (10), 1323–1331. [CrossRef]
- Jervis, M. A.; Kidd, N. A. C.; Walton, M. A Review of Methods for Determining Dietary Range in Adult Parasitoids. Entomophaga 1992, 37 (4), 565–574. [CrossRef]
- Godfray, H. C. J. Parasitoids: Behavioral and Evolutionary Ecology; Princeton University Press, 1994.
- Giron, D.; Pincebourde, S.; Casas, J. Lifetime Gains of Host-Feeding in a Synovigenic Parasitic Wasp. Physiol. Entomol. 2004, 29 (5), 436–442. [CrossRef]
- Ellers, J.; Sevenster, J. G.; Driessen, G. Egg Load Evolution in Parasitoids. Am. Nat. 2000, 156 (6), 650–665. [CrossRef]
- [10] Farahani, H. K.; Ashouri, A.; Zibaee, A.; Abroon, P.; Alford, L. The Effect of Host Nutritional Quality on Multiple Components of Trichogramma Brassicae Fitness. Bull. Entomol. Res. 2016, 106 (5), 633–641. [CrossRef]
- Hassan, S. A. The Mass Rearing and Utilization of Trichogramma to Control Lepidopterous Pests: Achievements and Outlook. Pestic. Sci. 1993, 37 (4), 387–391. [CrossRef]
- Boivin, G. Overwintering Strategies of Egg Parasitoids. Biol. Control Egg Parasit. 1994, 219–244.
- Flanders, S. E. Mass Production of Egg Parasites Oí the Genus Trichogramma. 1930.Accessed: Dec. 03, 2024. [Online]. Available: https://www.cabidigitallibrary.org/doi/full/10.5555/19310500283.
- Mills, N.; ‘Egg Parasitoids in Biological Control and Integrated Pest Management’, in Egg Parasitoids in Agroecosystems with Emphasis on Trichogramma, F. L. Consoli, J. R. P. Parra, and R. A. Zucchi, Eds., Dordrecht: Springer Netherlands, 2010, pp. 389–411. [CrossRef]
- Smith, S. M. Biological Control with Trichogramma: Advances, Successes, and Potential of Their Use. Annu. Rev. Entomol. 1996, 41 (Volume 41, 1996), 375–406. [CrossRef]
- Martel, V.; Johns, R. C.; Jochems-Tanguay, L.; Jean, F.; Maltais, A.; Trudeau, S.; St-Onge, M.; Cormier, D.; Smith, S. M.; Boisclair, J. The Use of UAS to Release the Egg Parasitoid Trichogramma Spp. (Hymenoptera: Trichogrammatidae) Against an Agricultural and a Forest Pest in Canada. J. Econ. Entomol. 2021, 114 (5), 1867–1881. [CrossRef]
- C Basso, C.; Chiaravalle, W.; Maignet, P.; Basso, C.; Chiaravalle, W.; Maignet, P. Effectiveness of Trichogramma Pretiosum in Controlling Lepidopterous Pests of Soybean Crops. Agrociencia Urug. 2020, 24 (SPE2). [CrossRef]
- Gavara, J.; Cabello, T.; Gámez, M.; Bastin, S.; Hernández-Suárez, E.; Piedra-Buena, A. Evaluation and Selection of New Trichogramma Spp. as Biological Control Agents of the Guatemalan Potato Moth (Tecia Solanivora) in Europe. Insects 2023, 14, 679, 2023. https://www.academia.edu/download/104845665/pdf.pdf (accessed 2024-12-03).
- Etilé, E.; Cabrera, P.; Boisclair, J.; Cormier, D.; Todorova, S.; Lucas, É. Field Evaluation of Trichogramma Ostriniae (Hymenoptera: Trichogrammatidae) and T. Brassicae as Biocontrol Agents of the European Corn Borer, Ostrinia Nubilalis (Lepidoptera: Crambidae), in Fresh Market Sweet Corn. Phytoprotection 2024, 104 (1), 35–46. [CrossRef]
- El, -Arnaouty S. A.; Galal, H. H.; Afifi, A. I.; Beyssat, V.; Pizzol, J.; Desneux, N.; Biondi, A.; Kortam, M. N.; Heikal, I. H. Assessment of Two Trichogramma Species for the Control of Tuta Absoluta in North African Tomato Greenhouses. Afr. Entomol. 2014, 22 (4), 801–809. [CrossRef]
- Sithanantham, S.; Abera, T. H.; Baumgärtner, J.; Hassan, S. A.; Löhr, B.; Monje, J. C.; Overholt, W. A.; Paul, A. V. N.; Wan, F. H.; Zebitz, C. P. W. EGG Parasitoids for Augmentative Biological Control of Lepidopteran Vegetable Pests in Africa: Research Status and Needs. Int. J. Trop. Insect Sci. 2001, 21 (3), 189–205. [CrossRef]
- Laminou, S. A.; Ba, M. N.; Karimoune, L.; Doumma, A.; Muniappan, R. Parasitism of Locally Recruited Egg Parasitoids of the Fall Armyworm in Africa. Insects 2020, 11 (7), 430. [CrossRef]
- Zang, L.-S.; Wang, S.; Zhang, F.; Desneux, N. Biological Control with Trichogramma in China: History, Present Status, and Perspectives. Annu. Rev. Entomol. 2021, 66 (Volume 66, 2021), 463–484. [CrossRef]
- Navik, O.; Yele, Y.; Kedar, S. C.; Sushil, S. N. Biological Control of Fall Armyworm Spodoptera Frugiperda (JE Smith) Using Egg Parasitoids, Trichogramma Species (Hymenoptera: Trichogrammatidae): A Review. Egypt. J. Biol. Pest Control 2023, 33 (1), 118. [CrossRef]
- Vinson, S. B.; Greenberg, S. M.; Rao, A.; Volosciuk, L. F. Biological Control of Pests Using Trichogramma: Current Status and Perspectives; Xi bei nong lin ke ji da xue chu ban she, 2015.
- Schmidt, J. M. Host Recognition and Acceptance by Trichogramma. Biol. Control Egg Parasit. 1994, 165–200.
- Foerster, M. R.; Foerster, L. A. Effects of Temperature on the Immature Development and Emergence of Five Species of Trichogramma. BioControl 2009, 54 (3), 445–450. [CrossRef]
- Kalyebi, A.; Overholt, W. A.; Schulthess, F.; Mueke, J. M.; Sithanantham, S. The Effect of Temperature and Humidity on the Bionomics of Six African Egg Parasitoids (Hymenoptera: Trichogrammatidae). Bull. Entomol. Res. 2006, 96 (3), 305–314. [CrossRef]
- Atashi, N.; Shishehbor, P.; Seraj, A. A.; Rasekh, A.; Hemmati, S. A.; Riddick, E. W. Effects of Helicoverpa Armigera Egg Age on Development, Reproduction, and Life Table Parameters of Trichogramma Euproctidis. Insects 2021, 12 (7), 569. [CrossRef]
- Tayat, E.; Özder, N. Preference Study of Trichogramma Pintoi (Voegele) (Hymenoptera:Trichogrammatidae) on Host Eggs of Different Ages and Species. Mustafa Kemal Üniversitesi Tarım Bilim. Derg. 2023, 28 (2), 355–362. [CrossRef]
- HASSAN, S. A. Strategies to Select Trichogramma Species for Use in Biological Control. Biol. Control Egg Parasit. 1994., Accessed: Dec. 05, 2024. [Online]. Available: https://cir.nii.ac.jp/crid/1571980075268517376.
- van Lenteren, J. C.; Babendreier, D.; Bigler, F.; Burgio, G.; Hokkanen, H. M. T.; Kuske, S.; Loomans, A. J. M.; Menzler-Hokkanen, I.; van Rijn, P. C. J.; Thomas, M. B.; Tommasini, M. G.; Zeng, Q.-Q. Environmental Risk Assessment of Exotic Natural Enemies Used in Inundative Biological Control. BioControl 2003, 48 (1), 3–38. [CrossRef]
- Sherwani, A.; Mukhtar, M.; Wani, A. A. Insect Pests of Apple and Their Management. Insect Pest Manag. Fruit Crops 2016, 295–306.
- Sigsgaard, L.; Herz, A.; Korsgaard, M.; Wührer, B. Mass Release of Trichogramma Evanescens and T. Cacoeciae Can Reduce Damage by the Apple Codling Moth Cydia Pomonella in Organic Orchards under Pheromone Disruption. Insects 2017, 8 (2), 41.
- Mansfield, S.; Mills, N. J. Host Egg Characteristics, Physiological Host Range, and Parasitism Following Inundative Releases of Trichogramma Platneri (Hymenoptera: Trichogrammatidae) in Walnut Orchards. Environ. Entomol. 2002, 31 (4), 723–731.
- Samara, R. Y.; Carlos Monje, J.; Zebitz, C. P. W. Comparison of Different European Strains of Trichogramma Aurosum (Hymenoptera: Trichogrammatidae) Using Fertility Life Tables. Biocontrol Sci. Technol. 2008, 18 (1), 75–86. [CrossRef]
- Yu, D. S. K.; Hagley, E. A. C.; Laing, J. E. Biology of Trichogramma Minutum Riley Collected from Apples in Southern Ontario. Environ. Entomol. 1984, 13 (5), 1324–1329. [CrossRef]
- Dolphin, R. E.; Cleveland, M. L.; Mouzin, T. E.; Morrison, R. K. Releases of Trichogramma Minutum1 and T. Cacoeciae1 in an Apple Orchard and the Effects on Populations of Codling Moths 2. Environ. Entomol. 1972, 1 (4), 481–484. [CrossRef]
- Hassan, S. A.; Kohler, E.; Rost, W. M. Mass Production and Utilization ofTrichogramma: 10. Control of the Codling mothCydia Pomonella and the Summer Fruit Tortrix mothAdoxophyes Orana [Lep.: Tortricidae]. Entomophaga 1988, 33 (4), 413–420. [CrossRef]
- Cherif, A.; Grissa-Lebdi, K. Field Releases of Trichogramma Cacoeciae (Marchal, 1927) (Hymenoptera: Trichogrammatidae) against the Codling Moth, Cydia Pomonella (Linnaeus, 1758) (Lepidoptera: Tortricidae). J. OASIS Agric. Sustain. Dev. 2024, 6 (03), 35–40. [CrossRef]
- Schäfer, L.; Herz, A. Suitability of European Trichogramma Species as Biocontrol Agents against the Tomato Leaf Miner Tuta Absoluta. Insects 2020, 11 (6), 357. [CrossRef]
- Abbas, S. S.; Shahzad, M. F.; Iqbal, J.; Ullah, A.; Batool, A.; Nadeem, M.; Begum, H. A.; Rehman, H.-; Muhammad, K. Trichogramma Chilonis as Parasitoid: An Eco-Friendly Approach Against Tomato Fruit Borer, Helicoverpa Armigera. J. Agric. Sci. 2020, 12 (2), 167. [CrossRef]
- Zhang, J.; Tang, R.; Fang, H.; Liu, X.; Michaud, J. P.; Zhou, Z.; Zhang, Q.; Li, Z. Laboratory and Field Studies Supporting Augmentation Biological Control of Oriental Fruit Moth, Grapholita Molesta (Lepidoptera: Tortricidae), Using Trichogramma Dendrolimi (Hymenoptera: Trichogrammatidae). Pest Manag. Sci. 2021, 77 (6), 2795–2803. [CrossRef]
- Reichart, G. Adatok a Magyarországi Gyümölcsösök Sodrómolyainak Ismeretéhez. Data Referring Knowl. Fruit Tree Leaf Roll. Hung. Orchards Ujabb Véd. Eljárások Kert. KártevHok És Bet. Ellen Bp. 1953, 21.
- Nagy, B. The Possible Role of Entomophagous Insects in the Genetic Control of the Codling Moth, with Special Reference toTrichogramma. Entomophaga 1973, 18 (2), 185–191. [CrossRef]
- [46] Bognár, S. Adatok Az Almamoly Magyarországi Természetes Ellenségeirôl És Szerepükrôl. (Data Concerning the Natural Enemies of the Apple Moth in Hungary and Their Role.). Kert. És Szőlészeti Főisk. Évkönyve 1962, 26, 31–43.
- Bokor, M.; Megújulóban van az almatermesztés Magyarországon. Agrofórum Online. https://agroforum.hu/agrarhirek/zoldseg-gyumolcs/megujuloban-van-az-almatermesztes-magyarorszagon/ (accessed 2024-12-08).
- Hedvig M. Mit tudnak a Trichogramma petefürkészek?. Biokontroll Hungária Nonprofit Kft. https://www.biokontroll.hu/mit-tudnak-a-trichogramma-petefuerkeszek/ (accessed 2024-12-08).
- Stouthamer, R.; Hu, J.; Van Kan, F. J. P. M.; Platner, G. R.; Pinto, J. D. [No Title Found]. BioControl 1999, 43 (4), 421–440. [CrossRef]
- Hegazi, E. M.; Herz, A.; Hassan, S.; Agamy, E.; Khafagi, W.; Shweil, S.; Zaitun, A.; Mostafa, S.; Hafez, M.; El-Shazly, A.; El-Said, S.; Abo-Abdala, L.; Khamis, N.; El-Kemny, S. Naturally Occurring Trichogramma Species in Olive Farms in Egypt. Insect Sci. 2005, 12 (3), 185–192. [CrossRef]
- Gonçalves, C. I.; Huigens, M. E.; Verbaarschot, P.; Duarte, S.; Mexia, A.; Tavares, J. Natural Occurrence of Wolbachia-Infected and Uninfected Trichogramma Species in Tomato Fields in Portugal. Biol. Control 2006, 37 (3), 375–381. [CrossRef]
- Souza, A. R. de; Giustolin, T. A.; Querino, R. B.; Alvarenga, C. D. Natural Parasitism of Lepidopteran Eggs by Trichogramma Species (Hymenoptera: Trichogrammatidae) in Agricultural Crops in Minas Gerais, Brazil. Fla. Entomol. 2016, 99 (2), 221–225. [CrossRef]
- Ram, P. Studies on Strains of Trichogramma Evanescens Westwood from Different Regions of Eurasia. Biocontrol Sci. Technol. 1995, 5 (3), 329–338. [CrossRef]
- Bírová H. Trichogramma Evanescens Westw. Als Beschrankungsfaktor Des Massenhaften Auftretens Des Maisziinslers (Pyrausta Nubilalis Hbn.).; 1956; Vol. The ontogeny of Insects, Acta symposii de evolutione insectorum Praha, 357-359.
- Bírová H. European Corn Borer-Pyrausta (i.e, Ostrinia) Nubilalis (Hbn.) (Lep. Pyralidae) in Czechoslovakia. 1962, a. Pol. Pismo Entomol. Bull., (Seria B,), 25–26, 25-29.
- írová H. VLskyt Vijacky Kukuricnej (Ostrinia Nubilalis Hbn.) v 19561985 v Oblasti Intenyivneho Pestovania Kukurice Na Slovensku. (Occurrence of the European Corn Borer (Ostrinia Nubilalis Hbn.) in 19561985 in a Region of Intensive Maize Production in Slovakia). XI. Czechoslovak Plant Protection Conference, Proceedings, 1988.
- Cagán, L.; Tancik, J.; Hassan, S. Natural Parasitism of the European Corn Borer Eggs Ostrinia Nubilalis Hbn. (Lep., Pyralidae) by Trichogramma in Slovakia—Need for Field Releases of the Natural Enemy. J. Appl. Entomol. 1998, 122 (1–5), 315–318. [CrossRef]
- Hochmut, R.; Martinek, V. Beitrag Zur Kenntnis Der Mitteleuropäischen Arten Und Rassen Der Gattung Trichogramma Westw. ( Hymenoptera, Trichogrammidae ). Z. Für Angew. Entomol. 1963, 52 (1–4), 255–274. [CrossRef]
- [59] Barnay, O.; Hommay, G.; Gertz, C.; Kienlen, J. C.; Schubert, G.; Marro, J. P.; Pizzol, J.; Chavigny, P. Survey of Natural Populations of Trichogramma (Hym., Trichogrammatidae) in the Vineyards of Alsace (France). J. Appl. Entomol. 2001, 125 (8), 469–477. [CrossRef]
- Castaneda-Samayoa, O.; Holst, H.; Ohnesorge, B. Evaluation of Some Trichogramma Species with Respect to Biological Control of Eupoecilia Ambiguella Hb. and Lobesia Botrana Schiff. (Lep., Tortricidae) / Evaluierung Einiger Trichogramma-Arten Hinsichtlich Ihrer Verwendbarkeit Zur Biologischen Bekämpfung von Eupoecilia Ambiguella Hb. Und Lobesia Botrana Schiff. (Lep., Tortricidae). Z. Für Pflanzenkrankh. Pflanzenschutz J. Plant Dis. Prot. 1993, 100 (6), 599–610.
- Dudich, E. Insect Parasites of the Corn Borer (Pyrausta Nubilalis Hb.) in Hungary. Int. Corn Borer Investig. Sci. Rep. 1928, 1, 184–190.
- B Nagy, B. On Some Aspects of the Investigations of the Maize Ecosystem in Hungary, with Special Respect to the European Corn Borer. Vedecke Pr. Vysk. Ustavu Kukurice V Trnave Czechoslov. 1985, No. 14.
- Sengonca, C.; Leisse, N. Vorkommen Und Bedeutung von Trichogramma Semblidis Auriv. (Hymenoptera, Trichogrammatidae) Als Eiparasit Beider Traubenwicklerarten Im Ahrtal. J. Appl. Entomol. 1987, 103 (1–5), 527–531. [CrossRef]
- Şengonca, Ç.; Leisse, N. Enhancement of the Egg Parasite Trichogramma Semblidis (Auriv.) (Hym., Trichogrammatidae) for Control of Both Grape Vine Moth Species in the Ahr Valley1,2. J. Appl. Entomol. 1989, 107 (1–5), 41–45. [CrossRef]
- Kot, J. Experiments in the Biology and Ecology of Species of the Genus Trichogramma Westw. and Their Use in Plant Protection. 1964.
- Zouba, A.; Zougari, S.; Mamay, M.; Kadri, N.; Ben Hmida, F.; Lebdi-Grissa, K. The Effect of Different Oviposition and Preadult Development Temperatures on the Biological Characteristics of Four Trichogramma Spp. Parasitoids (Hymenoptera: Trichogrammatidae) Species. Phytoparasitica 2024, 52 (1), 19. [CrossRef]
- Schöller, M.; Hassan, S. A. Comparative Biology and Life Tables of Trichogramma Evanescens and T. Cacoeciae with Ephestia Elutella as Host at Four Constant Temperatures. Entomol. Exp. Appl. 2001, 98 (1), 35–40. [CrossRef]
- Mohammad, J. K.; Al-Jassany, R. F.; Ali, A.-S. A. Influence of Temperature on Some Biological Characteristics of Trichogramma Evanescens (Westwood)(Hymenoptera: Trichogrammatidae) on the Egg of Lesser Date Moth Batrachedra Amydraula Meyrick. 2015.
- Metwally, M. M.; El-Kordy, M. W.; Mohamed, H. A.; El-Sebai, A.; Atta, A. A. Effect of temperature, photoperiod, biological and chemical factors of three host species on the performance of the eggs parasitoid Trichogramma Evanescens WEST. J. Plant Prot. Pathol. 2013, 4 (9), 781–793. [CrossRef]
- Fournier, F.; Pelletier, D.; Vigneault, C.; Goyette, B.; Boivin, G. Effect of Barometric Pressure on Flight Initiation by Trichogramma Pretiosum and Trichogramma Evanescens (Hymenoptera: Trichogrammatidae). Environ. Entomol. 2005, 34 (6), 1534–1540. [CrossRef]
Figure 1.
Meteorological data (mean daily temperature (°C); minimum daily temperature (°C); maximum daily temperature (°C); relative humidity (%) (A) and mean barometric pressure (hPa) (B) during the experimental period (8 July – 30 September 2024); Source: https://www.met.hu/rolunk/tevekenysegek/adattar/.
Figure 1.
Meteorological data (mean daily temperature (°C); minimum daily temperature (°C); maximum daily temperature (°C); relative humidity (%) (A) and mean barometric pressure (hPa) (B) during the experimental period (8 July – 30 September 2024); Source: https://www.met.hu/rolunk/tevekenysegek/adattar/.
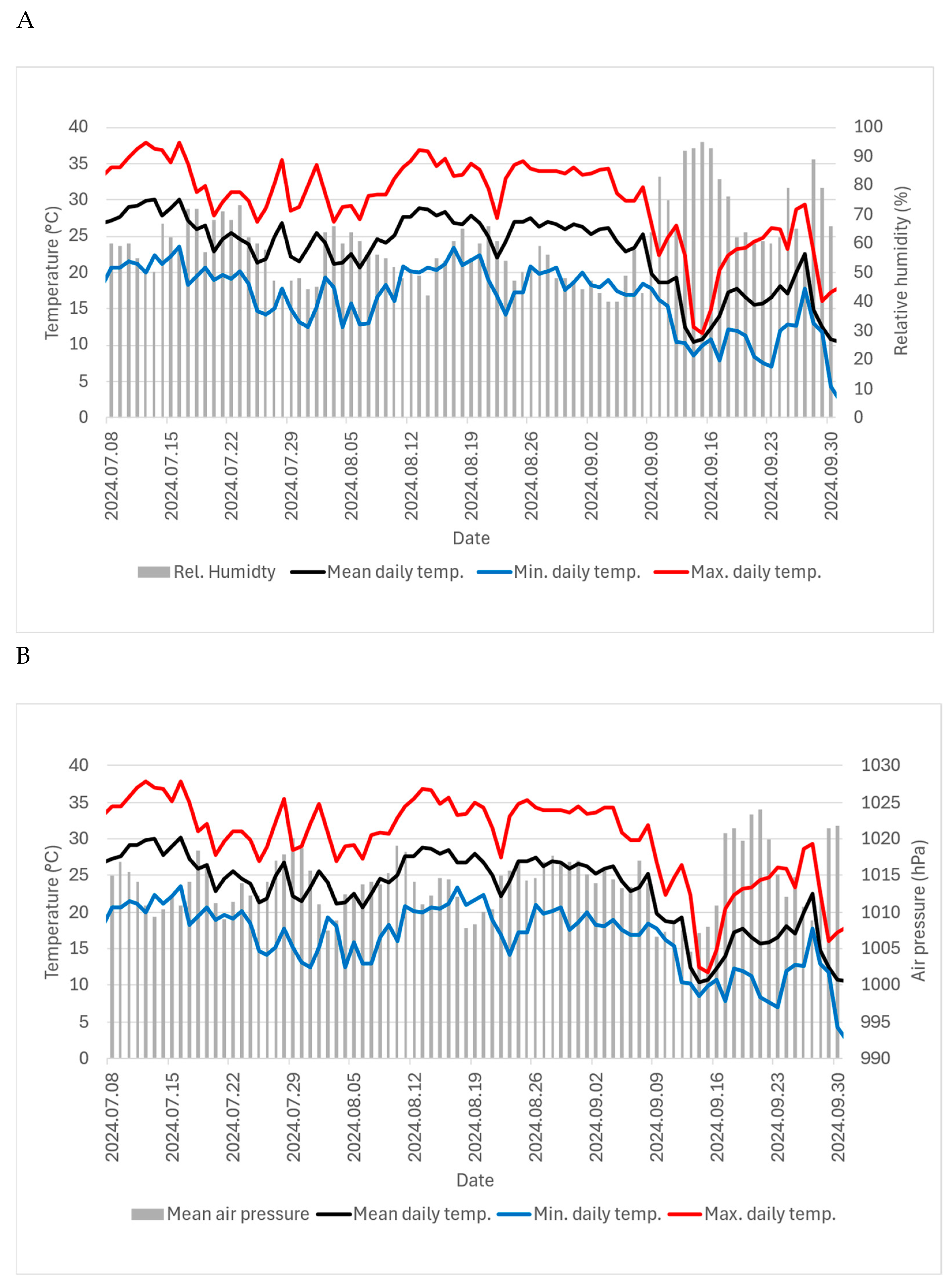
Figure 2.
Parasitism rate (%) and number of parasitised eggs plotted at collection time (15 July – 1 October 2024).
Figure 2.
Parasitism rate (%) and number of parasitised eggs plotted at collection time (15 July – 1 October 2024).
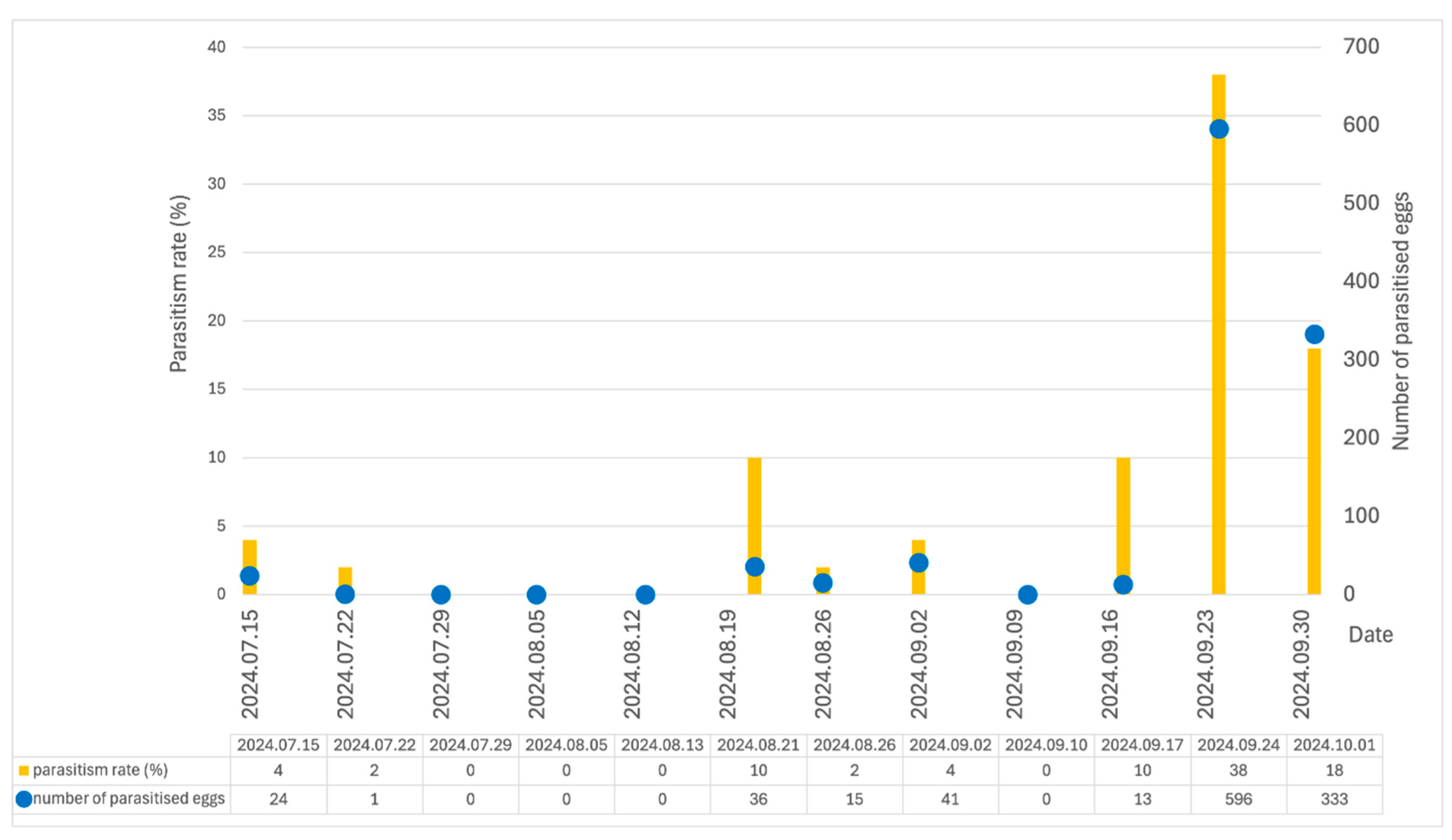

Table 1.
Categorisation of data for statistical analysis.
| Group Category Values | ||||||
|---|---|---|---|---|---|---|
| Group Category | 0 | 1 | 2 | 3 | 4 | 5 |
| Discovery rate* (%) | 0 | 1–5 | 6–15 | >16 | ||
| Parasitism rate† | 0 | 0.001–0.1 | 0.101–0.4 | - | - | |
| Hatching rate (%)‡ | 0 | 0–95 | 96–99 | 100 | - | |
| Date of the placement | - | 08.07.2024–13.08.2024 | 14.08.2024–17.09.2024 | 18.09.2024–01.10.2024 | - | - |
| Age of host eggs (days) | - | 1–7 | 8–14 | >14 | - | - |
| Number of host eggs (pcs) | - | 1–100 | 101–200 | 201–300 | 301–400 | >400 |
* the number of cards containing at least one egg parasitised by Trichogramma wasps divided by the total number of cards released (%). † the ratio of parasitised eggs to the total number of eggs released. ‡ the number of hatched eggs divided by the total number of parasitised eggs.
Table 2.
Correlation between parasitism rate (%), the number of parasitised eggs and the average meteorological parameters during the time of release: Pearson's correlation coefficient (R), test value of t-test of the significance of R with degree of freedom (t; df) and significance value (p). The only positive significant correlation is highlighted in bold.
Table 2.
Correlation between parasitism rate (%), the number of parasitised eggs and the average meteorological parameters during the time of release: Pearson's correlation coefficient (R), test value of t-test of the significance of R with degree of freedom (t; df) and significance value (p). The only positive significant correlation is highlighted in bold.
| Average mean temperature* (°C) |
Average minimum temperature* (°C) | Average maximum temperature* (°C) |
Average relative humidity* (%) |
Average wind speed* (m/sec) | Average air pressure* (hPa) | ||
|---|---|---|---|---|---|---|---|
| Parasitism rate (%) † |
R | -0.64 | -0.60 | -0.75 | 0.46 | -0.19 | 0.49 |
| t (df=10) | 2.64 | 2.38 | 3.59 | 1.62 | 0.60 | 1.78 | |
| p | 0.025 | 0.039 | 0.005 | 0.136 | 0.559 | 0.106 | |
| Number of parasitised eggs | R | -0.61 | -0.65 | -0.70 | 0.33 | -0.16 | 0.67 |
| t (df=10) | 2.46 | 2.74 | 3.06 | 1.09 | 0.50 | 2.83 | |
| p | 0.033 | 0.021 | 0.012 | 0.301 | 0.625 | 0.018 | |
† the ratio of parasitised eggs to the total number of eggs released; * average values are calculated from the daily data of mean, maximum and mean temperatures (°C), mean relative humidity (%), mean wind speed (m/sec) and mean air pressure (hPa), in relation to the time intervals of the cards released.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



