Submitted:
17 February 2025
Posted:
18 February 2025
You are already at the latest version
Abstract
Background: Staphylococcus aureus is a highly lethal Gram-positive bacterium that is responsible for over one million deaths annually. As a member of the ESKAPE pathogens, its methicillin-resistant strains (MRSA) are prevalent worldwide and exhibit significant antimicrobial resistance (AMR). Bacterial efflux pumps play a pivotal role in the development of AMR by facilitating the expulsion of a range of antimicrobial agents; Methods: The S. aureus strain SA-1199B, which overexpresses norA and carries a GrlA mutation, was utilized to comprehensively profile the mechanism of the compounds PQQ16P and PQK4F. To assess the toxicity and genotoxicity of these compounds, RAW macrophages, HEK 293T, and HepG2 cell lines were utilized. Female BALB/c mice were utilized to assess the in vivo synergism of EPIs with CPX; Results: NorA efflux pump inhibitors (EPIs), PQQ16P and PQK4F, enhanced the efficacy of the antibacterial ciprofloxacin (CPX) against resistant S. aureus strains. The mechanism of EPIs involved the inhibition of NorA efflux pump, without compromising bacterial membrane permeability, ATP levels, or mammalian calcium channels. Moreover, the EPIs significantly augmented the bactericidal and post-antibiotic effects of CPX, elevating its mutation prevention concentration without manifesting substantial toxicity to human cells. Furthermore, the EPIs reduced S. aureus invasiveness in macrophages, indicating a role for NorA in bacterial virulence. Notably, the in vivo synergism of these EPIs with CPX was observed in a mouse infection model; Conclusions: This study provides substantial evidence for the potential of employing EPIs in a combination with CPX to counteract AMR, both in vitro and in vivo.
Keywords:
efflux pump inhibitors (EPIs)
; Staphylococcus aureus
; antimicrobial resistance (AMR)
; antimicrobial resistance breakers (ARBs)
; ciprofloxacin
1. Introduction
Staphylococcus aureus is a Gram-positive bacterial pathogen that is a leading cause of both hospital and community-acquired infections [1]. Due to its capacity for adaptation to diverse environmental conditions, S. aureus is capable of causing a broad spectrum of infections, ranging from minor dermatological conditions to severe diseases such as pneumonia, endocarditis, and sepsis [2,3,4]. The pathogenicity of S. aureus is largely attributed to a diverse array of virulence factors, including toxins and enzymes that facilitate tissue invasion and immune evasion [2]. S. aureus is one of the most lethal bacterial pathogens, responsible for approximately 1.1 million deaths globally in 2019 [5]. In addition, it plays a significant role in the development of antimicrobial resistance (AMR) due to its capacity to evolve resistance to a range of antibiotics [6,7,8]. It is, therefore, unsurprising that it is part of the ESKAPE pathogens [9,10]. One of the most notable examples is methicillin-resistant S. aureus (MRSA), which exhibits resistance to methicillin and other beta-lactam antibiotics [11]. The treatment of MRSA infections presents a significant challenge, with higher morbidity and mortality rates associated with these infections [11]. The extensive utilization of antibiotics in both healthcare and agricultural contexts has facilitated the emergence of antibiotic-resistant strains of S. aureus [12,13,14]. This bacterium can obtain resistance genes through horizontal gene transfer and mutations, rendering it a formidable pathogen in both hospital and community settings [15]. The prevalence of MRSA and other resistant strains of S. aureus highlights the urgent necessity for the development of novel antibiotics and alternative therapeutic strategies to combat these infections [16]. Efflux pumps are of pivotal importance with regard to the antibiotic resistance of S. aureus [17,18]. These pumps are integral membrane proteins that actively expel a variety of antimicrobial agents out of the bacterial cell, thereby reducing the intracellular concentration of these drugs and diminishing their efficacy. In S. aureus, several efflux pumps have been identified, including NorA, NorB, NorC, MepA, and QacA/B [19,20,21]. These pumps are capable of transporting a diverse range of antibacterials, including macrolides, quinolones, and tetracyclines, as well as biocides such as quaternary ammonium compounds [19,20,21]. As a result, they play a pivotal role in enabling the bacterium to withstand the presence of these antimicrobial agents. Efflux pumps are frequently encoded on plasmids or within the bacterial chromosome, thereby facilitating the rapid acquisition and dissemination of resistance traits [22]. This renders them a pivotal element in the emergence of multidrug-resistant (MDR) and extensively drug-resistant (XDR) variants of S. aureus. In this regard, a challenge strategy is based on the approach of targeting and disrupting the mechanisms responsible for AMR. The intriguing concept of freezing resistance could potentially salvage ineffective antibiotics, thereby replenishing our arsenal of antimicrobial agents. The molecules that have been identified as capable of counteracting AMR have been referred to by a number of different names, including adjuvant molecules, helper compounds, and antimicrobial resistance breakers (ARBs) [23,24,25,26,27]. All are distinguished by a lack of antimicrobial activity and the capacity to interact synergistically with known antimicrobials, thereby restoring their efficacy against resistant strains. Since microorganisms primarily evolve resistance only to compounds that exert bactericidal or bacteriostatic effects [28,29], the absence of antimicrobial activity in ARBs appears to be a potential advantage for their future use. In light of the pivotal role of bacterial efflux pumps in the development of AMR, the development of efflux pump inhibitors (EPIs) represents a promising strategy [30].
The transmembrane protein NorA, belonging to the Major Facilitator Superfamily (MFS) and utilizing a proton motive force (PMF) as an energy source, is responsible for the extrusion of various chemicals (including the fluoroquinolone ciprofloxacin (CPX) and the dye ethidium bromide (EtBr)) and is the most extensively studied efflux pump in S. aureus [31,32,33]. Over time, numerous NorA EPIs have been identified through the application of three distinct methodologies: (i) screening of natural or synthetic compound libraries, (ii) drug repurposing, and (iii) the design and synthesis of novel compounds based on phenotypic screenings [34]. Conversely, only the recent release of two three-dimensional structures of NorA [33,35] and the lack of biophysical/biochemical assays using the isolated protein have significantly impeded the structure-based drug design and the identification of potent NorA EPIs, which have yet to reach clinical trials. Nevertheless, some chemical classes of NorA EPIs have been identified, including indoles [36,37,38,39], quinolines [40,41], quinazolines [42], boronic acids [43,44], chalcones [45], and piperine derivatives [46,47], as well as also various natural compounds. These include boeravinone B, capsaicin, α-terpinene, eugenol, isoeugenol, sesquiterpenes derived from Pilgerodendron uviferum and some flavonoids that have demonstrated promising results [48,49,50,51,52,53]. Furthermore, plant extracts or essential oils, including the ethanolic extract from Bauhinia forficata leaves, Chenopodium ambrosioides L. essential oil, and Nigella sativa essential oil, have demonstrated promising outcomes [54,55,56].
Our research group has been engaged in this field for years and has contributed to the development of various NorA EPIs through the application of different methodologies [57]. As a result of these endeavours, we recently reported the optimization of two classes of NorA EPIs derivatives: quinazolines and quinolines, from which the compounds PQK4F [42] and PQQ16P [58] emerged (Figure 1). Both demonstrated potent NorA inhibitory activity and a high capacity to synergize with CPX against resistant strains of S. aureus. In view of these encouraging outcomes, the present study was undertaken to conduct comprehensive investigations into the mechanisms of action of these two derivatives and their potential for combination with CPX, both in in vitro studies and in animal models.
2. Results
2.1. Impact of Compounds PQK4F and PQQ16P on CPX's Minimum Inhibitory Concentration (MIC)
Firstly, checkerboard assays of PQK4F [42] and PQQ16P [58] in combination with CPX were carried out in order to confirm previous data against the resistant S. aureus strain SA-1199B (overexpressing norA and bearing a GrlA mutation on the fluoroquinolone target) [59] (Table S1). Additionally, determination of MIC values of PQK4F and PQQ16P was also evaluated to confirm the absence of any antibacterial effect at low concentrations when tested individually (Table S1). Indeed, as anticipated in the introduction, EPIs are not expected to have antibacterial activity on their own. As expected, both compounds showed a promising synergism with CPX, reducing its MIC by 4-fold when tested at 3.13 µg/mL. To further confirm the synergism, Fractional Inhibitory Concentration Index (FICI) values were calculated and were ≤ 0.5 for both compounds, indicating a synergistic interaction with CPX (Table S1). In parallel, through checkerboard assays against S. aureus K-1758 (norA deletion strain), minimal or no modulation of the MIC values of CPX was observed for both compounds, as confirmed by FICI values >0.5 to <4, suggesting an indifferent interaction (Table S2). Therefore, the observed synergy with CPX against S. aureus SA-1199B, which overexpresses the norA gene, suggests that the compounds PQK4F and PQQ16P may function as inhibitors of the NorA efflux pump, and this is further corroborated by the absence of synergism against the S. aureus K-1758 strain that does not express norA gene.
2.2. Effect of the Combination of CPX with Compounds PQK4F and PQQ16P on the Time-Kill Kinetics on S. aureus SA-1199B
The time-kill kinetic study establishes the rate at which a compound kills a microorganism as a function of survival recorded at various exposure time points. Initially, CPX was used in this assay at inhibitory (8 µg/mL) and sub-inhibitory (2 µg/mL) concentrations alone. CPX at 8 µg/mL showed a reduction in SA-1199B growth of 2.25 log10 after 12 h of treatment, followed by a resurgence in growth. Conversely, CPX at 2 µg/mL showed a growth curve comparable to that of the untreated control bacterial cells. As expected, compounds PQK4F and PQQ16P tested individually at 12.48 µg/mL and 6.24 µg/mL, respectively, also demonstrated a comparable trend with no discernible impact on S. aureus growth. It is noteworthy that when sub-inhibitory concentrations of CPX (2 µg/mL and 1 µg/mL) were combined with PQK4F (12.48 µg/mL) or PQQ16P (6.24 µg/mL), a reduction of 3.69 log10 and 0.8 log10 and of 3.83 log10 and 2.54 log10, respectively, was observed after 12 h of treatment (Figure 2). Taken together, the data clearly demonstrate the significant impact of a combination of these two EPIs with CPX on the growth rate of the resistant S. aureus SA-1199B over time, thereby underscoring the potential benefits of a combination therapy.
2.3. Membrane Permeabilization and Depolarization Activity of Compounds PQK4F and PQQ16P on S. aureus SA-1199B
To exclude the possibility that the observed synergistic activity between CPX and PQK4F/PQQ16P was due to an unspecific effect generated by an increase in the S. aureus membrane permeability, the impact of PQK4F and PQQ16P on membrane permeabilization was assessed using propidium iodide (PI) (Figure 3.A). Polymyxin, used as a positive control, exhibited a significant increase in the fluorescence. In contrast, no such increase was observed in the case of PQK4F and PQQ16P at concentrations of 6.24 and 12.48 µg/mL, respectively. This indicates that PI was unable to enter the viable cell and bind to the target. Taken together, the results demonstrated that the compounds PQK4F and PQQ16P do not permeabilize the membrane.
An additional phenomenon that could result in unspecific synergism and produce extended human cell toxicity is that of membrane depolarization. To assess whether compounds PQK4F and PQQ16P possessed this unwanted property, the membrane potential-sensitive dye DiSC3(5) was used to quantify changes in the electrical potential gradient in intact bacteria (Figure 3.B). Valinomycin used as a positive control, caused massive fluorescence leakage, demonstrating its membrane-depolarizing property [60]. Considering the two tested compounds, the effect was more pronounced for the compound PQQ16P, which at a concentration of 12.48 µg/mL led to a significant depolarization of the S. aureus membrane. However, at a concentration of 6.25 µg/mL, the membrane depolarization was observed to be relatively mild, and it nearly disappeared at a concentration of 3.13 µg/mL. These findings align with those of our previous research [58]. In contrast, the compound PQK4F, for which no previous evaluations of its impact on bacterial membrane depolarization have been conducted, demonstrated a markedly less pronounced effect than PQQ16P, even at the highest concentration (12.48 µg/mL). Given that NorA is a PMF-dependent pump, it is plausible that the compounds at the highest tested concentration (12.48 µg/mL) may disturb the PMF, thereby partially “freezing” the activity of the pump and contributing to the synergism with CPX through this “dirty” mechanism. Nevertheless, at lower concentrations (3.13 µg/mL and 6.25 µg/mL, particularly for PQK4F), the impact on membrane depolarization is insignificant, and the synergism of PQK4F and PQQ16P with CPX appears to be exclusively due to the direct inhibition of NorA pump, rather than the disruption of PMF (Figure 3.B).
2.4. Effect of Compounds PQK4F and PQQ16P on the Electron Transport Chain by ATP Bioluminescence Detection Assay
Membrane dysfunction disturbs the respiratory chain functions, resulting in reduced ATP levels. The S. aureus SA-1199B strain was incubated with PQK4F and PQQ16P at 6.24, 12.48, and 24.96 µg/mL for 5 h, and the effect of compounds on the intracellular ATP level was evaluated (Figure 3.C). The ATP levels remained constant compared to the drug-free control, indicating no change in transmembrane potential, ruling out the possibility of ATP depletion as the cause of efflux inhibition. In contrast, a significant reduction in the intracellular ATP was observed in the case of CCCP (positive control), which is a potent mitochondrial uncoupling agent that increases the proton permeability across the mitochondrial inner membranes [30].
2.5. Effect of Compounds PQK4F and PQQ16P on Mammalian Ca2+ Channels
We were cognizant that many bacterial efflux inhibitors limit mammalian Ca2+ channel activity in clinical settings, such as verapamil inhibits host Ca2+ channels, eliciting human neurotoxicity [61,62]. The mammalian Ca2+ channel blocking was assessed using the Fluo-4 Direct calcium channel assay kit on HEK 293T cells (Figure 4). Notably, the Ca2+ channel activity was not affected by PQK4F and PQQ16P at concentrations of 12.48 and 6.24 µg/mL. In contrast, the positive control, verapamil, prevented the accumulation of Ca2+ ions in the cytoplasm. Taken together, the results showed that compounds did not disturb the Ca2+ channel activity, overcoming the problem that has hampered the development of some EPIs [63,64] (Figure 4).
2.6. Toxicity Studies of Compounds PQK4F and PQQ16P Towards Eukaryotic Cells
In order to gain preliminary insight into the toxicity of compounds PQK4F and PQQ16P, a viability assessment of mammalian cells was conducted on three different cell lines: RAW macrophage cells, human embryonic kidney cells (HEK 293T), and hepatoblastoma cell line (HepG2). Cell viability was determined by MTT assay at varying concentrations of PQK4F and PQQ16P ranging from 1.56 to 50 µg/mL (Figure 5). In general, both compounds demonstrated a certain degree of toxicity at concentrations of 50 and 25 µg/mL, while at lower concentrations, they exhibited significantly reduced toxicity. It is noteworthy that a comparison of the profiles of PQK4F and PQQ16P revealed that the former exhibited a superior safety profile and was significantly safer than the quinoline derivative PQQ16P. However, when the focus is narrowed to the concentrations utilized to achieve a notable synergistic effect with CPX, it can be observed that both compounds do not exhibit a considerable degree of toxicity against the three cell lines. This suggests that the synergistic effect can be attained at concentrations that are not inherently toxic.
2.7. Hemolytic Activity Studies of the Compounds PQK4F and PQQ16P
Additionally, the hemolytic activity of both compounds was evaluated by measuring the release of hemoglobin from rabbit erythrocytes as a function of concentration. It is noteworthy that the compounds PQK4F and PQQ16P exhibited minimal hemolytic activity, with values of 2.2% and 0.8%, respectively, at a concentration of 100 µg/mL (Figure 6).
2.8. Genotoxicity Assessment
The assessment of genotoxicity in preclinical candidates is of paramount importance for the early identification of compounds that may induce genetic mutations, which can ultimately lead to the development of cancer or heritable genetic defects. Indeed, the early detection of genotoxicity in the drug development process can result in significant savings in terms of time and resources. Such an approach prevents the advancement of potentially harmful compounds into later stages of development, where the costs and time investments are significantly higher. Accordingly, comet assays were conducted to assess the potential genotoxicity of PQK4F and PQQ16P using three distinct cell lines. It is noteworthy that the two compounds demonstrated no genotoxic effect (Figure 7, Supplementary Table S3). The ratio of means for the cells exposed to the compounds ranged between 0.78 and 1.37, while the ratio of means for the positive control (2 µM 4NQO) fluctuated between 3.23 and 5.45.
2.9. Enhanced Post-Antibiotic Life of CPX in the Presence of Compounds PQK4F and PQQ16P
The post-antibiotic effect (PAE) is defined as the suppression of bacterial growth for a specified duration following a brief exposure to an antimicrobial agent. A longer PAE allows for the administration of antibiotics at less frequent intervals, which can enhance patient compliance and mitigate the risk of missed doses. Moreover, extending the PAE can guarantee that bacteria are subjected to the antibiotic's effects for a more extended period, which may result in a more efficacious eradication of the infection. When an exponential-phase culture of S. aureus SA-1199B was subjected to 1 × MIC of CPX alone, a PAE of 1.3/1.4 h was observed. In contrast, a notable increase in the PAE was observed when CPX (2 µg/mL) was combined with PQK4F and PQQ16P at three distinct concentrations (12.48, 6.24, and 3.12 µg/mL). In particular, the increase in PAE in combination with PQK4F was of 3.7, 3.7, and 3.2 h, respectively, while in combination with PQQ16P, it was of 5.2, 4.8, and 4.1 h. The findings suggest that both EPIs contribute to an extension in the period of bacterial growth suppression (Figure 8).
2.10. Mutation Frequency Analysis of Compounds PQK4F and PQQ16P to Check the Development of Its Resistant Mutants
The capacity of a compound to impede or attenuate the emergence of resistant mutants is quantified by its mutation prevention concentration (MPC). It is imperative to maintain antibiotic concentrations above the MPC to prevent the development of resistance through single-step mutations. Indeed, the mutant selection window represents the range of antibiotic concentrations where resistant mutants are likely to emerge. A reduction in the MPC will result in a narrowing of the window, thereby reducing the probability of the emergence of resistant mutants. CPX alone displayed an MPC at 4 × MIC. In comparison, PQK4F at 12.48 and 6.24 µg/mL reduced the MPC of CPX by 8- and 4-folds, respectively, in the S. aureus SA-1199B strain (Table 1). Similar results were obtained when testing PQQ16P at 6.24 and 3.12 µg/mL in combination with CPX (Table 1).
2.11. Effectiveness of PQK4F and PQQ16P in Mitigating Macrophage Invasion by norA over-Expressing S. aureus
It is of great importance to mitigate the invasion of macrophages by S. aureus for a number of reasons, including the prevention of chronic infections, reduction of immune evasion, limitation of tissue damage, enhancement of antibiotic efficacy, and the prevention of dissemination [65]. Accordingly, the intracellular invasion of macrophage THP-1 cells (3 × 105 cells/well) by different S. aureus strains, including SA-1199B (norA overexpressing strain), SA-1199 (wild-type strain), and K-1758 (norA knockout strain), was evaluated in the presence or absence of PQK4F and PQQ16P (12.48 and 6.24 µg/mL) (Figure 9). The presence of PQK4F (12.48 and 6.24 µg/mL) led to a reduction in the invasion of S. aureus SA-1199B to 0.63 and 0.49 log10, respectively, compared to reductions of 0.30 and 0.05 log10, respectively, in the wild-type S. aureus SA-1199. Similarly, the presence of PQQ16P (12.48 and 6.24 µg/mL) resulted in decreased invasion of S. aureus SA-1199B to 0.75 and 0.61
log10, respectively, compared to reductions of 0.30 and 0.07 log10, respectively, in the wild-type SA-1199. The penetration of S. aureus K-1758 remained unaffected by PQK4F and PQQ16P treatments at concentrations of 12.48 and 6.24 µg/mL. Taken together, the data not only underscore the potential efficacy of the compounds PQK4F and PQQ16P in mitigating macrophage invasion by S. aureus but also indirectly reinforce the importance of efflux pumps in the invasion process. Indeed, it can be observed that the strain SA-1199B, which overexpresses norA, exhibits a markedly enhanced invasion capacity relative to both the wild-type strain and the strain that does not express norA. In particular, the difference in macrophage invasion capacity between the latter and the former is readily apparent. Moreover, these findings suggest that EPIs may serve as potential anti-virulence agents, given that when administered alone, they are capable of significantly reducing the macrophage invasion capacity of S. aureus.
2.12. Mouse Thigh Infection Model In Vivo
The impressive efficacy of the combination prompted us to assess the impact of compounds PQK4F and PQQ16P in combination with CPX on a mouse soft-tissue infection model. Using a neutropenic mouse thigh infection model, we investigated the in vivo effectiveness of these compounds (Figure 10). Mice were infected intramuscularly with 107 CFU of S. aureus SA-1199B. After 4 h, the first control group was sacrificed to confirm successful infection establishment, and their right thigh tissue was homogenized and plated. The CFU count in this control group was approximately 7.7 log10. Treatment with CPX (10 mg/Kg) or the EPIs PQK4F and PQQ16P (5 and 10 mg/Kg) alone, administered 4 h post-infection, did not significantly reduce bacterial counts after 20 h of treatment. However, combining PQK4F at doses of 5 mg/Kg and 10 mg/Kg with CPX at 10 mg/Kg, administered 4 h post-infection, showed notable efficacy, resulting in a reduction of approximately 0.86 and 1.5 log10, respectively, compared to the untreated control after 24 h. Similarly, combining PQQ16P at doses of 5 mg/Kg and 10 mg/Kg with CPX at 10 mg/Kg, administered 4 h post-infection, demonstrated significant efficacy, resulting in a reduction of approximately 1.86 and 2.34 log10, respectively, compared to the untreated control after 24 h. CPX (50 mg/Kg) showed a reduction of 1.9 log10 that is approximately equal to that of CPX (10 mg/Kg) + PQK4F (10 mg/Kg). Notably, the combination of CPX with PQQ16P shows higher CFU reduction than CPX alone at 50 mg/Kg.
3. Discussion
S. aureus exhibits remarkable adaptability to environmental changes and rapidly acquires resistance to antibiotics. This study focuses on a key mechanism that contributes to heightened resistance against various antimicrobial agents, namely the expulsion of antibiotics through efflux pumps. Consequently, compounds capable of inhibiting these pumps may prove effective in countering or mitigating resistance. Therefore, the objective of our investigation was to conduct a thorough investigation of the impact of PQQ16P and PQK4F (Figure 1), two previously reported S. aureus NorA EPIs [42,58], on fluoroquinolone resistance or tolerance in S. aureus. The selection of these two compounds was based on their proven efficacy as NorA inhibitors, resulting from a series of medicinal chemistry optimization cycles. Indeed, both compounds have been demonstrated to exhibit synergistic activity with CPX at low concentrations against the S. aureus SA-1199B strain. Moreover, their activity is comparable to that of some of the most potent analogues reported in the literature as NorA EPIs [34,39,57]. Thus, in this study, we initially confirmed the ability of PQK4F and PQQ16P to synergize with CPX against SA-1199B (Table S1). We then proceeded to ascertain that the observed effect was primarily due to the actual inhibition of the NorA efflux pump. The initial indication of this phenomenon was observed in the absence of a synergistic effect between the two compounds and CPX when tested against S. aureus K1758, which lacks the norA gene (Table S2). Indeed, it is to be expected that compounds which act as NorA inhibitors will be unable to synergize with CPX against a strain which does not possess the NorA pump. Conversely, compounds that exhibit synergistic activity with CPX, even in strains lacking NorA, could potentially function as inhibitors of other efflux pumps that are otherwise basally expressed in this strain or act through non-specific mechanisms, such as alteration of the bacterial membrane. Therefore, to eliminate the potential influence of PQK4F and PQQ16P on the bacterial membrane, we also assessed their impact on membrane depolarization, membrane permeability, ATP depletion (Figure 3.A-C), and their potential to counteract the activation of eukaryotic Ca2+ channels (Figure 4). Indeed, also the inhibition of the eukaryotic Ca2+ channels can contribute to identification of false EPIs, as demonstrated by compounds like verapamil [61,62]. It is noteworthy that both compounds did not exhibit the aforementioned undesirable activities, with the exception of membrane depolarization at the highest concentrations tested. In contrast, at lower concentrations that are useful for obtaining the synergistic effect with CPX, membrane depolarization is either modest or absent. This indicates that both compounds can act as direct NorA inhibitors devoid of a "dirty" effect. Once confirmed that the synergistic effect of PQK4F and PQQ16P was based on the inhibition of NorA, additional studies have been conducted to investigate the potential biological effect of combining NorA inhibitors with CPX against the resistant S. aureus strain. Time-kill curves demonstrated that the combination of CPX at concentrations of 2 µg/mL (1/4 MIC) and 1 µg/mL (1/8 MIC) with PQK4F at 12.48 µg/mL and PQQ16P at 6.24 µg/mL resulted in a notable reduction of S. aureus growth after 12 h of treatment, in comparison to the treatment involving CPX alone (Figure 2). This finding highlights the potential of this adjunctive therapy to enhance the bactericidal effects of CPX, even at sub-MIC concentrations, against the target pathogens. Furthermore, both PQK4F and PQQ16P facilitated the enhancement of PAE, impeding bacterial cell recovery. At a concentration of 12.48 µg/mL, PQK4F and PQQ16P augmented the PAE of CPX at 2 µg/mL (1/4 MIC) significantly, extending it by 3.7 h and 5.2 h, respectively, in comparison to CPX at its MIC (8 µg/mL), which exhibited a PAE of 1.3 h. In addition to enhancing the efficacy of antibiotic treatment in suppressing the growth of S. aureus, both compounds demonstrated a notable reduction (8-fold) in the MPC of CPX when administered in combination, as evidenced by mutation frequency analysis (Table 1). These findings highlight the potential of a concurrent therapy in minimizing the development of antibiotic resistance. Indeed, the enhancement of efflux pump expression in bacteria represents a crucial initial step in the development of target-based resistance [17,66]. Considering these findings, the combination of EPIs and antibacterials may represent a promising avenue for future therapeutic development. This approach has the potential to prevent the emergence of resistance, even when utilizing lower concentrations of antibacterials. Furthermore, the inhibition of the NorA efflux pump may prove an effective method of reducing the capacity of S. aureus to invade macrophages (Figure 9). Although S. aureus is typically not considered an intracellular pathogen, there is a possibility for it to survive and propagate disease by briefly occupying an intracellular niche [65,67]. It is worth noting that our observations indicate that overexpression of the NorA efflux pump contributes to the intracellular invasion of S. aureus SA-1199B in THP-1 macrophage cell lines. This significantly enhances the invasion ability of SA-1199B compared to the wild-type strain SA-1199 and the norA-deleted strain SA-K1758. In addition, the NorA EPI compounds PQK4F and PQQ16P were observed to prevent invasion by SA-1199B, thereby further confirming their ability to inhibit NorA and corroborating the evidence that NorA plays a role in aiding S. aureus in invading macrophages.
To ascertain the potential utility of these two compounds in an animal model, the toxicity profile was also investigated using three distinct mammalian cell lines (RAW, HEK, and HepG2 cell lines – Figure 5). Both compounds demonstrated only modest toxicity at the highest concentrations tested, which are considerably higher than those required to achieve the potent synergistic effect with CPX. Furthermore, when evaluated via comet assays at concentrations up to 12.5 µg/mL on the same three cell lines, neither compound exhibited any notable genotoxicity (Figure 7). Moreover, both compounds demonstrated minimal haemolytic activity, with a rate of less than 3% (Figure 6).
Subsequently, encouraged by these promising results, we decided to assess the in vivo effectiveness of the combination of PQK4F and PQQ16P with CPX. It is crucial to emphasize the significance of these studies in animal models, as they facilitate the assessment of the actual potential of the NorA EPI-CPX combination. Indeed, while numerous examples of NorA EPIs exist in the literature, regrettably, there is a dearth of data pertaining to their potential applicability in animal models, which would facilitate more comprehensive preclinical studies. In this study, our findings revealed a significant reduction in CFU of PQK4F or PQQ16P in combination with CPX, both compared to infected animals treated with CPX alone and the control group of untreated animals. Indeed, combination of NorA EPIs with 10 mg/Kg of CPX showed an activity comparable or slightly better (in the case of PQQ16P) than CPX alone at 50 mg/Kg, strongly highlighting the advantage to combine NorA EPIs with CPX in animal models.
4. Materials and Methods
4.1. Bacterial Strains, Cell Lines, and Growth Medium
The S. aureus SA-1199B (overexpressing the NorA MDR efflux pump and bearing an A116E GrlA substitution), SA-1199 (norA wild-type), and SA-K-1758 (NCTC 8325-4 norA deletion mutant) were used. Bacterial media included Cation adjusted BBLTM Mueller Hinton II Broth (CA-MHB), and Mueller–Hinton agar (MHA).
4.2. MIC Determination and Synergy Assay
Using broth microdilution by CLSI standards, the minimum inhibitory concentration (MIC) was ascertained [68,69]. The experiment used 96 round-bottom well plates with a total reaction volume of 200 µL. The bacteria were diluted until each well contained 5 × 105 CFU/mL and their OD600nm was set to 0.25–0.3. The plates were then incubated at 37°C for 18 h.
The checkerboard experiment was used to determine the synergistic effect of the compounds PQK4F and PQQ16P with CPX (Sigma Aldrich) [68,69]. The following equation was used to calculate the fractional inhibitory concentration index (FICI), which was used to determine the synergistic effect of the compounds PQK4F and PQQ16P:
FICI= MIC (antibiotic in the presence of EPI)/MIC (antibiotic alone) + MIC (EPI in the presence of antibiotic)/ MIC (EPI alone)
The FICI values of ≤0.5, >0.5-4, and >4 were considered to be synergistic, indifferent, and antagonistic, respectively [70].
4.3. Membrane Potentiation Assay
Mid-exponential S. aureus SA-1199B was cultivated at 37 ˚C. The cells were centrifuged, washed three times for 10 minutes at 4,000 × g, and then resuspended in 50 mM HEPES buffer with 0.1% glucose. At last, the OD600nm was adjusted to 0.1. The cells were then treated with DiSC3(5) (Sigma Aldrich) dye at a concentration of 1 µM and 300 mM KCl in HEPES buffer (to balance the outer and cytoplasmic K+ concentrations) before being incubated for 10 minutes at room temperature in the dark[71,72,73,74]. Valinomycin (Sigma Aldrich) (MIC = 16 µg/mL) was used as the positive control and the compounds PQK4F and PQQ16P were added at 0.78, 1.56, 3.12, 6.24 and 12.48 µg/mL concentrations. After that, the bacterial OD600nm was normalized for both the control and all treatment groups. Then the fluorescence leakage was measured after 30 min at a λexcitation/λemission of 622/670 nm.
4.4. Membrane Permeability Assay
The InvitrogenTM Propidium Iodide (PI) kit (Thermo Fisher Scientific) methodology was modified slightly for the membrane permeabilization assay to quantify the fluorescence of propidium iodide (PI) permeating the membrane. S. aureus SA-1199B was cultured and allowed for growth in CA-MHB until it reached the mid-exponential stage with an OD600 nm of 0.3. The compounds PQK4F and PQQ16P at 6.24 µg/mL and 12.48 µg/mL concentrations were added to the 500 µL bacterial suspension for 30 min. Following treatment, the culture was washed three times and resuspended in PBS. The OD600nm was normalized for treated and untreated cells. The suspensions were given a 30 µM PI treatment and incubated for 15–20 minutes at room temperature. The suspensions were then suspended in 500 µL of PBS after being centrifuged at 13,000 × g for 5 minutes to remove any unbound PI. After 30 minutes, the fluorescence of 100 µL of suspensions was measured in a 96-well plate at 490 and 635 nm for excitation and emission, respectively [39].
4.5. Determination of Intracellular ATP Levels
Bacteria's intracellular ATP levels were measured by the manufacturer's instructions using an ATP determination kit (Invitrogen, Life Technologies, U.S.). 500 µL of the S. aureus SA-1199B bacterial culture was treated with CCCP (Sigma Aldrich) at 6.24, 12.48, 24.96 µg/mL in MHB and the compounds PQK4F and PQQ16P separately at a sub-inhibitory concentration for 4 h at 37 °C and 200 rpm. After treatment, the OD600nm of the treated and untreated culture was normalized. 100 µL of freshly prepared solution of BacTiter-Glo reagent were added in each well and then add equal volume of cells and incubated for 5 min. All steps are performed at room temperature (22–25°C). Total ATP was measured using relative luminescence [39,75]. An 0.25–1 second integration time per well should be a guideline.
4.6. Mammalian Ca2+ Channel Blocking Assay
The Fluo-4 Direct Ca2+ channel test kit (Life Technologies, Carlsbad, CA) was utilized to investigate the effect of the compounds PQK4F and PQQ16P on human calcium channels by the manufacturer's recommendations [62]. Fluorescence was evaluated when Probenecid (5 mM) and Fluo-4 dye were added to HEK-293T cells (5 × 104) for 1 hour. The compounds PQK4F and PQQ16P at 6.24 µg/mL and 12.48 µg/mL concentrations, and verapamil at 50 μg/mL were then applied to the cells. After 2 minutes, the Ca2+ channel stimulator carbachol (50 μg/mL) was applied, and the fluorescence was measured at 494/516 nm using a microplate reader.
4.7. Genotoxicity Test
4.7.1. Chemical and Reagents
Minimum Essential Medium Eagle, Dulbecco′s Modified Eagle′s Medium, foetal bovine serum (FBS), trypsin-EDTA, L-glutamine, antibiotics (penicillin and streptomycin), non-essential amino acid, β-mercaptoethanol and HEPES were bought from Euroclone SpA (Milan, Italy). Hydrochloric acid (HCl), dimethyl sulfoxide (DMSO), ethanol, ethylenediaminetetraacetic acid disodium (Na2EDTA) and tetrasodium (Na4EDTA) salt, sodium chloride (NaCl), and sodium hydroxide (NaOH) were obtained from Carlo Erba Reagenti Srl (Milan, Italy). Dulbecco’s phosphate-buffered saline, pH 7.4 (PBS), ethidium bromide, low- and normal-melting point agarose (LMPA and NMPA, respectively), 4-nitroquinoline N-oxide (4NQO), tris(hydroxymethyl)aminomethane (Tris base), Triton X-100, and GlutaMax were obtained from Merck Life Science (Merck KGaA, Darmstadt, Germany). Conventional microscope slides and coverslips were purchased from Knittel-Glaser GmbH (Braunschweig, Germany). Distilled water was used throughout the experiments.
4.7.2. Alkaline Single-Cell Microgel Electrophoresis (Comet) Assay
According to the Organization for Economic Cooperation and Development (OECD) guidelines [76,77], to avoid any conditions which would lead to false-positive results, only non-cytotoxic concentrations (i.e. 3.125, 6.25, 12.5 µg/mL) of PQK4F or PQQ16P were considered in the comet assay. This latter was performed following previous procedures [77,78], with minor modifications.
Firstly, each cell line (HEK 293T from passage 2 to 4, RAW 264.7 from passage 2 to 4 and Hep G2 from passage 7 to 10) was seeded at approximately 2.65 × 105 cells/well in 12-well plates (Corning Inc.). The day after, the growth medium was removed and cells were exposed to the selected non-cytotoxic scalar concentration (see above). Proper negative (untreated cells) and positive (2 µM NQO) controls were carried out. In order to prevent any additional DNA damage, each step was carried out under a yellow light throughout the comet test, which was set up in triplicate.
After 4 hours of treatment, cells were detached and centrifuged (1,000 × g, 8 min, 8 °C). Subsequently, they were resuspended in 0.7% LMPA (in Ca2+/Mg2+-free DPBS (w/v)) and layered onto microscope slides precoated with 1% NMPA in Ca2+/Mg2+-free DPBS (w/v). Coverslips were placed onto the samples and removed after allowing the agarose to solidify for 10 min at 4 °C, followed by another top layer of 75 µL of 0.7% LMPA. Slides were then dipped overnight at 4 °C in a lysing solution, composed of 2.5 M NaCl, 100 mM Na2EDTA, and 10 mM Tris-HCl, pH 10, containing 1% Triton X-100.
Slides were maintained for 20 min in an alkaline buffer solution (pH > 13) of 10 mM Na4EDTA and 300 mM NaOH to allow the expression of alkali-labile damage and the unwinding of DNA. Electrophoresis was performed with a horizontal box (HE99; Hoefer Scientific, Holliston, MA, USA) at an electric field strength of 1 V/cm, with current at 300 mA (Power Supply E411; Consort, Turnhout, Belgium) in an ice bath for 20 min. Microgels were later neutralised with 0.4 M Tris-HCl buffer, pH 7.5.
Slides were finally fixed in ethanol for 10 min and stored at room temperature.
To rate DNA damage, the slides were stained with ethidium bromide (50 µL, 20 µg/mL) and examined using an Olympus BΧ41 (Japan) fluorescence microscope equipped with a high-sensitivity charge-coupled device (CCD) camera connected to a computerised image analysis system (“Comet Assay III”, Perspective Instruments Ltd., Suffolk, UK). One hundred randomly selected cells (50 cells/replicate slides) were analysed, with tail intensity (%) being chosen as the damage parameter. The median of the scored comets for each slide was used to calculate the group means [79].
Results were expressed as the ratio of means (treated cells % tail intensity/untreated cells % tail intensity), for each concentration. Results were expressed as the ratio of means with the 95% confidence interval of the triplicate test. Statistical significance was rated through one-way ANOVA and Dunnet’s post hoc test considering a significant level of p < 0.05.
4.8. Post-Antibiotic Assay
The S. aureus SA-1199B strain was cultivated to the mid-log phase in CA-MHB. After that, the culture was set up to an OD600nm of 0.25. The bacterial culture was further diluted a thousand times before being treated with CPX at concentrations of 8 and 2 µg/mL, and the compounds PQK4F and PQQ16P at 3.12, 6.24, 12.48 µg/mL concentrations or a combination of CPX at 2 µg/mL with PQK4F or PQQ16P at 3.12, 6.24, 12.48 µg/mL concentrations, at 37 °C for two h. Following that, the bacterial suspension OD600nm for each treatment group and the control was normalized. Then, to remove the drug's carry-over effect, the bacterial cultures were centrifuged for 10 min. at 1200 × g. The control was the untreated bacterial suspension. In a 96-well flat-bottom plate, a 300 µL bacterial suspension was dispensed and subjected to incubation at 37°C for 18 h. The Post-Antibiotic Effect (PAE) was determined using the formula PAE = T50 - C50, where T50 and C50 represent the durations required for the treated and untreated bacterial cultures, respectively, to achieve 50% of the optical density (OD) observed in the untreated culture [80].
4.9. Mutation Prevention Concentration (MPC)
Bacterial suspensions of S. aureus SA-1199B (109 CFU) were applied to MHA plates at different CPX concentrations (0.25 ×, 0.5 ×, 1 ×, 2 ×, 4 ×, 8 × MIC) alone and in combination with PQK4F at 6.24 and 12.48 µg/mL and PQQ16P at 3.12 and 6.24 µg/mL concentrations, and the plates were then incubated at 37 °C. Colonies were counted after 48 h, and the mutation frequency was calculated by dividing the number of survivors by the total CFU plated. The concentration at which no colonies formed was referred to as the MPC [81].
4.10. Time-Kill Kinetics Study
S. aureus SA-1199B cultures were cultivated to mid-exponential phase, and a suspension with an optical density at 600 nm (OD600nm) of approximately 0.3 was prepared. The initial inoculum of S. aureus SA-1199B was exposed to CPX at concentrations of 8 µg/mL and 2 µg/mL and PQK4F and PQQ16P at its sub-inhibitory concentrations of 12.48 µg/mL and 6.24 µg/mL respectively. Also a combination of CPX with PQK4F and PQQ16P separately at 12.48 µg/mL and 6.24 µg/mL respectively. Bacterial counts were determined at various intervals (0, 2, 4, 6, 8, 12, and 24 h) and quantified as colony-forming units per milliliter (CFU/mL) [75].
4.11. Hemolysis Assay
For the hemolysis assay[75], fresh rabbit blood erythrocytes were employed. Blood was drawn into a heparin tube, then centrifuged and washed with PBS three times. The RBCs were resuspended in PBS to achieve a final concentration of 4% (vol/vol). In 200 µL of freshly prepared 4% RBC suspension, PQK4F and PQQ16P were added at final concentrations ranging from 0.78 to 100 µg/mL. Additionally, 0.1% Triton-X (positive control) was included. The suspension was then incubated for 1 hour at 37°C before being centrifuged for 5 minutes at 4°C at 1000 × g. Following centrifugation, 100 µL of the supernatant was transferred to a 96-well flat-bottom plate, and the absorbance was measured at 570 nm using a BioTek spectrophotometer. % Hemolysis = [(Absorbance of treatment – Absorbance of blank)/(Absorbance of control – Absorbance of blank)] × 100
4.12. Mammalian Cytotoxicity
The cytotoxicity of compounds PQK4F and PQQ16P was evaluated on Human Embryonic Kidney 293T cells (HEK), RAW cells, and HepG2 cells [39,75]. Each well was seeded with approximately 10,000 cells separately in individual plates and then incubated in a CO2 incubator with 5% CO2 at 37°C for 24 h. Following incubation, varying concentrations of compounds PQK4F and PQQ16P (1.56, 3.125, 6.25, 12.5, 25, 50, 100 µg/mL) were added to the respective plates and further incubated at 37°C in a CO2 incubator. After further 24 h of incubation, MTT solution (500 µg/mL) was added to the wells after removing the medium. Formazan crystals formed were then solubilized with a 100 µL stopping solution comprising 40% (vol/vol) dimethylformamide (DMF) in 2% (vol/vol) glacial acetic acid and 16% (wt/vol) sodium dodecyl sulfate (SDS) at pH 4.7. The absorbance of the resulting solution was measured at 570 nm using a BioTek spectrophotometer
The given equation was used to calculate the percent viability:
% Cell viability = (At/ Ac) × 100;
At = (absorbance of test compound – absorbance of blank);
Ac = (absorbance of control – absorbance of blank)
4.13. Macrophage Invasion Assay
The THP-1 macrophage cell line was utilized to investigate the intracellular invasion of S. aureus in the presence of compounds PQK4F and PQQ16P [39,75]. S. aureus strains SA-1199B, SA-1199, and K-1758 (106 CFU/well) were used to infect macrophages (105 cells/well), with or without PQK4F and PQQ16P (12.48 and 6.24 µg/mL), and incubated for 2 h. Subsequently, extracellular bacteria were removed by washing the cells three times with PBS, followed by treatment with gentamicin (50 µg/mL) for 30 minutes. The cells were then briefly treated with 0.1% saponin, and the viability of intracellular bacteria was determined by plating on MHA.
4.14. In Vivo Thigh Infection Model
Eight groups of mice, each consisting of six female BALB/c mice, were employed for the study. To induce neutropenia, female mice were treated with two intraperitoneal doses of 150 mg/Kg and 100 mg/Kg of cyclophosphamide, administered 4 and 1 days prior to infection, respectively. Following this, mice were intramuscularly injected with 50 µL of S. aureus SA-1199B (107 CFU/mL). Four h post-infection, various treatments were administered subcutaneously. Two groups received 5 mg/Kg and 10 mg/Kg of either PQK4F or PQQ16P individually. Another set of two groups received a combination of 5 mg/Kg and 10 mg/Kg of PQK4F or PQQ16P with 10 mg/Kg of CPX (Sigma Aldrich). Subsequently, two groups were administered with 10 mg/Kg and 50 mg/Kg of CPX, respectively. The final two groups served as untreated controls. One control group was euthanized before treatment by cervical dislocation after 4 h of infection. The right thighs were then excised, homogenized, and CFUs were determined. The remaining groups were euthanized by cervical dislocation 20 h after receiving the administered doses. CFUs from all groups were enumerated from the right thigh [39,82].
4.15. Statistical Analysis
Data are presented as mean ± standard deviation. Two-tailed t-tests were conducted to detect group variances. Statistical significance was defined as P < 0.05 (*), highly significant as P < 0.01 (**), P < 0.001 (***), and P < 0.0001 (****).
5. Conclusions
In this study, a comprehensive analysis has been conducted to elucidate the mechanism of action of two NorA EPIs that have been previously identified. Subsequently, an investigation was conducted to assess the potential benefits of combining both EPIs with CPX, with the aim of identifying the potential advantages of this combination. In conclusion, our findings demonstrate that the NorA EPI-CPX combination is advantageous in several aspects. These include increasing the efficacy and potency of CPX against resistant strains of S. aureus, as well as preventing the ability of S. aureus to generate mutations and become resistant. Moreover, NorA EPIs can also exert an anti-virulence action, limiting the invasion of macrophages by S. aureus. It is also noteworthy that the combination of NorA EPIs with CPX has been demonstrated to be effective in the treatment of S. aureus infection in an animal model. It is crucial to acknowledge that this animal model study employed the resistant strain SA-1199B, which exhibits both overexpression of the NorA efflux pump and a single mutation on the fluoroquinolone target. Nevertheless, it is essential to underscore the notion of combining a NorA EPI with CPX also to prevent the emergence of resistance. It is therefore our recommendation that the concept of exclusively utilizing EPIs to treat infections with pre-existing resistance be reconsidered. Conversely, it would be prudent to endeavor to demonstrate that the EPI-antibacterial combination has the potential to serve as a significant future strategy for the prevention of the emergence of resistant strains. The efficacy of this approach has been validated in this study through in vitro MPC experiments. Further studies will be needed to elucidate the potential advantages of this combination as a prophylactic strategy against resistance development in animal models.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: MIC and checkerboard synergy assay of the compounds PQK4F and PQQ16P with CPX on S. aureus SA-1199B (norA overexpressed strain); Table S2: MIC and checkerboard synergy assay of the compounds PQK4F and PQQ16P with CPX on S. aureus K-1758 (norA deletion strain); Table S3: Genotoxicity assessment of PQK4F and PQQ16P at different concentrations.
Author Contributions
Conceptualization, T.F., H.N. and S.S.; formal analysis, N.C., N.S., T.R., and M.A.; investigation, N.C., N.S., T.R., and M.A.; data curation, N.C. and T.F.; writing—original draft preparation, N.C. and T.F.; writing—review and editing, G.C., T.F., H.N. and S.S.; supervision, H.N. and S.S.; project administration, H.N. and S.S.; funding acquisition, H.N. and S.S. All authors have read and agreed to the published version of the manuscript.
Funding
This work has been funded by the European Union – NextGenerationEU under the Italian Ministry of University and Research (MUR) National Innovation Ecosystem grant ECS00000041 - VITALITY - CUP J97G22000170005.
Acknowledgments
All authors acknowledge CSIR-Institute of Microbial Technology, Chandigarh India for providing the infrastructure and laboratory facilities to execute the biological experiments. All authors are also thankful to Council of Scientific & Industrial Research (CSIR), New Delhi for execution of international collaboration between CSIR-Institute of Microbial Technology, Chandigarh INDIA and Department of Pharmaceutical Sciences, University of Perugia, Italy.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
AMR, antimicrobial resistance; ARBs, antimicrobial resistance breakers; CPX, ciprofloxacin; EPI, efflux pump inhibitor; EtBr, ethidium bromide; FICI, fractional inhibitory concentration index; HEK 293T, human embryonic kidney cells; HepG2, hepatoblastoma cell line; MDR, multidrug-resistance; MFS, Major Facilitator Superfamily; MIC, minimum inhibitory concentration; MPC, mutation prevention concentration; PAE, post-antibiotic effect; MRSA, methicillin-resistant S. aureus; PMF, proton motive force.
References
- Gherardi, G. Staphylococcus Aureus Infection: Pathogenesis and Antimicrobial Resistance. Int J Mol Sci 2023, 24, 8182. [CrossRef]
- Cheung, G.Y.C.; Bae, J.S.; Otto, M. Pathogenicity and Virulence of Staphylococcus Aureus. Virulence 2021, 12, 547. [CrossRef]
- Thomer, L.; Schneewind, O.; Missiakas, D. Pathogenesis of Staphylococcus Aureus Bloodstream Infections. Annual Review of Pathology: Mechanisms of Disease 2016, 11, 343–364. [CrossRef]
- Kwiecinski, J.M.; Horswill, A.R. Staphylococcus Aureus Bloodstream Infections: Pathogenesis and Regulatory Mechanisms. Curr Opin Microbiol 2020, 53, 51–60. [CrossRef]
- Ikuta, K.S.; Swetschinski, L.R.; Aguilar, G.R.; Sharara, F.; Mestrovic, T.; Gray, A.P.; Weaver, N.D.; Wool, E.E.; Han, C.; Hayoon, A.G.; et al. Global Mortality Associated with 33 Bacterial Pathogens in 2019: A Systematic Analysis for the Global Burden of Disease Study 2019. The Lancet 2022, 400, 2221–2248. [CrossRef]
- Aguilar, G.R.; Swetschinski, L.R.; Weaver, N.D.; Ikuta, K.S.; Mestrovic, T.; Gray, A.P.; Chung, E.; Wool, E.E.; Han, C.; Hayoon, A.G.; et al. The Burden of Antimicrobial Resistance in the Americas in 2019: A Cross-Country Systematic Analysis. The Lancet Regional Health - Americas 2023, 25, 100561. [CrossRef]
- Murray, C.J.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Robles Aguilar, G.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global Burden of Bacterial Antimicrobial Resistance in 2019: A Systematic Analysis. The Lancet 2022, 399, 629–655. [CrossRef]
- Mestrovic, T.; Robles Aguilar, G.; Swetschinski, L.R.; Ikuta, K.S.; Gray, A.P.; Davis Weaver, N.; Han, C.; Wool, E.E.; Gershberg Hayoon, A.; Hay, S.I.; et al. The Burden of Bacterial Antimicrobial Resistance in the WHO European Region in 2019: A Cross-Country Systematic Analysis. Lancet Public Health 2022, 7, 897–913. [CrossRef]
- Boucher, H.W.; Talbot, G.H.; Bradley, J.S.; Edwards, J.E.; Gilbert, D.; Rice, L.B.; Scheld, M.; Spellberg, B.; Bartlett, J. Bad Bugs, No Drugs: No ESKAPE! An Update from the Infectious Diseases Society of America. Clinical Infectious Diseases 2009, 48, 1–12. [CrossRef]
- Miller, W.R.; Arias, C.A. ESKAPE Pathogens: Antimicrobial Resistance, Epidemiology, Clinical Impact and Therapeutics. Nat Rev Microbiol 2024, 1–19. [CrossRef]
- Turner, N.A.; Sharma-Kuinkel, B.K.; Maskarinec, S.A.; Eichenberger, E.M.; Shah, P.P.; Carugati, M.; Holland, T.L.; Fowler, V.G. Methicillin-Resistant Staphylococcus Aureus: An Overview of Basic and Clinical Research. Nat Rev Microbiol 2019, 17, 203–218. [CrossRef]
- Abebe, A.A.; Birhanu, A.G. Methicillin Resistant Staphylococcus Aureus: Molecular Mechanisms Underlying Drug Resistance Development and Novel Strategies to Combat. Infect Drug Resist 2023, 16, 7641. [CrossRef]
- Mann, A.; Nehra, K.; Rana, J.S.; Dahiya, T. Antibiotic Resistance in Agriculture: Perspectives on Upcoming Strategies to Overcome Upsurge in Resistance. Curr Res Microb Sci 2021, 2, 100030. [CrossRef]
- Singh, B.; Bhat, A.; Ravi, K. Antibiotics Misuse and Antimicrobial Resistance Development in Agriculture: A Global Challenge. Environment and Health 2024. [CrossRef]
- Haaber, J.; Penadés, J.R.; Ingmer, H. Transfer of Antibiotic Resistance in Staphylococcus Aureus. Trends Microbiol 2017, 25, 893–905. [CrossRef]
- Douglas, E.J.A.; Wulandari, S.W.; Lovell, S.D.; Laabei, M. Novel Antimicrobial Strategies to Treat Multi-drug Resistant Staphylococcus Aureus Infections. Microb Biotechnol 2023, 16, 1456. [CrossRef]
- Papkou, A.; Hedge, J.; Kapel, N.; Young, B.; MacLean, R.C. Efflux Pump Activity Potentiates the Evolution of Antibiotic Resistance across S. Aureus Isolates. Nat Commun 2020, 11, 1–15. [CrossRef]
- Rajabi, S.; Shivaee, A.; Khosravi, M.A.; Eshaghi, M.; Shahbazi, S.; Hosseini, F. Evaluation of Multidrug Efflux Pump Expression in Clinical Isolates of Staphylococcus Aureus. Gene Rep 2020, 18, 100537. [CrossRef]
- The Major Facilitator Superfamily, M.; Kolar, M.; Auxiliadora Dea-Ayuela, M.; Stephen, J.; Salam, F.; Lekshmi, M.; Kumar, S.H.; Varela, M.F. The Major Facilitator Superfamily and Antimicrobial Resistance Efflux Pumps of the ESKAPEE Pathogen Staphylococcus Aureus. Antibiotics 2023, 12, 343. [CrossRef]
- Costa, S.S.; Viveiros, M.; Amaral, L.; Couto, I. Multidrug Efflux Pumps in Staphylococcus Aureus: An Update. Open Microbiol J 2013, 7, 59–71. [CrossRef]
- Hassanzadeh, S.; Ganjloo, S.; Pourmand, M.R.; Mashhadi, R.; Ghazvini, K. Epidemiology of Efflux Pumps Genes Mediating Resistance among Staphylococcus Aureus; a Systematic Review. Microb Pathog 2020, 139, 103850. [CrossRef]
- Schindler, B.D.; Kaatz, G.W. Multidrug Efflux Pumps of Gram-Positive Bacteria. Drug Resistance Updates 2016, 27, 1–13. [CrossRef]
- Laws, M.; Shaaban, A.; Rahman, K.M. Antibiotic Resistance Breakers: Current Approaches and Future Directions. FEMS Microbiol Rev 2019, 43, 490–516. [CrossRef]
- Dhanda, G.; Acharya, Y.; Haldar, J. Antibiotic Adjuvants: A Versatile Approach to Combat Antibiotic Resistance. ACS Omega 2023, 8, 10757–10783. [CrossRef]
- Kumar, V.; Yasmeen, N.; Pandey, A.; Ahmad Chaudhary, A.; Alawam, A.S.; Ahmad Rudayni, H.; Islam, A.; Lakhawat, S.S.; Sharma, P.K.; Shahid, M. Antibiotic Adjuvants: Synergistic Tool to Combat Multi-Drug Resistant Pathogens. Front Cell Infect Microbiol 2023, 13, 1293633. [CrossRef]
- Wright, G.D. Antibiotic Adjuvants: Rescuing Antibiotics from Resistance. Trends Microbiol 2016, 24, 862–871. [CrossRef]
- Martins, M.; Dastidar, S.G.; Fanning, S.; Kristiansen, J.E.; Molnar, J.; Pagès, J.-M.; Schelz, Z.; Spengler, G.; Viveiros, M.; Amaral, L. Potential Role of Non-Antibiotics (Helper Compounds) in the Treatment of Multidrug-Resistant Gram-Negative Infections: Mechanisms for Their Direct and Indirect Activities. Int J Antimicrob Agents 2008, 31, 198–208. [CrossRef]
- Christaki, E.; Marcou, M.; Tofarides, A. Antimicrobial Resistance in Bacteria: Mechanisms, Evolution, and Persistence. J Mol Evol 2019, 88, 26–40. [CrossRef]
- Zhang, F.; Cheng, W. The Mechanism of Bacterial Resistance and Potential Bacteriostatic Strategies. Antibiotics 2022, 11, 1215. [CrossRef]
- Sharma, A.; Gupta, V.K.; Pathania, R. Efflux Pump Inhibitors for Bacterial Pathogens: From Bench to Bedside. Indian J Med Res 2019, 149, 129. [CrossRef]
- Yoshida, H.; Bogaki, M.; Nakamura, S.; Ubukata, K.; Konno, M. Nucleotide Sequence and Characterization of the Staphylococcus Aureus NorA Gene, Which Confers Resistance to Quinolones. J Bacteriol 1990, 172, 6942–6949.
- Costa, S.S.; Sobkowiak, B.; Parreira, R.; Edgeworth, J.D.; Viveiros, M.; Clark, T.G.; Couto, I. Genetic Diversity of NorA, Coding for a Main Efflux Pump of Staphylococcus Aureus. Front Genet 2019, 10, 421435. [CrossRef]
- Li, J.; Li, Y.; Koide, A.; Kuang, H.; Torres, V.J.; Koide, S.; Wang, D.N.; Traaseth, N.J. Proton-Coupled Transport Mechanism of the Efflux Pump NorA. Nat Commun 2024, 15, 1–12. [CrossRef]
- Lamut, A.; Peterlin Mašič, L.; Kikelj, D.; Tomašič, T. Efflux Pump Inhibitors of Clinically Relevant Multidrug Resistant Bacteria. Med Res Rev 2019, 39, 2460–2504. [CrossRef]
- Brawley, D.N.; Sauer, D.B.; Li, J.; Zheng, X.; Koide, A.; Jedhe, G.S.; Suwatthee, T.; Song, J.; Liu, Z.; Arora, P.S.; et al. Structural Basis for Inhibition of the Drug Efflux Pump NorA from Staphylococcus Aureus. Nature Chemical Biology 2022 2022, 1–7. [CrossRef]
- Lepri, S.; Buonerba, F.; Goracci, L.; Velilla, I.; Ruzziconi, R.; Schindler, B.D.; Seo, S.M.; Kaatz, G.W.; Cruciani, G. Indole Based Weapons to Fight Antibiotic Resistance: A Structure-Activity Relationship Study. J Med Chem 2016, 59, 867–891. [CrossRef]
- Buonerba, F.; Lepri, S.; Goracci, L.; Schindler, B.D.; Seo, S.M.; Kaatz, G.W.; Cruciani, G. Improved Potency of Indole-Based NorA Efflux Pump Inhibitors: From Serendipity toward Rational Design and Development. J Med Chem 2017, 60, 517–523. [CrossRef]
- Hequet, A.; Burchak, O.N.; Jeanty, M.; Guinchard, X.; Le Pihive, E.; Maigre, L.; Bouhours, P.; Schneider, D.; Maurin, M.; Paris, J.M.; et al. 1-(1H-Indol-3-Yl)Ethanamine Derivatives as Potent Staphylococcus Aureus NorA Efflux Pump Inhibitors. ChemMedChem 2014, 9, 1534–1545. [CrossRef]
- Chandal, N.; Tambat, R.; Kalia, R.; Kumar, G.; Mahey, N.; Jachak, S.; Nandanwar, H. Efflux Pump Inhibitory Potential of Indole Derivatives as an Arsenal against NorA over-Expressing Staphylococcus Aureus. Microbiol Spectr 2023, 11, e0487622. [CrossRef]
- Felicetti, T.; Mangiaterra, G.; Cannalire, R.; Cedraro, N.; Pietrella, D.; Astolfi, A.; Massari, S.; Tabarrini, O.; Manfroni, G.; Barreca, M.L.; et al. C-2 Phenyl Replacements to Obtain Potent Quinoline-Based Staphylococcus Aureus NorA Inhibitors. J Enzyme Inhib Med Chem 2020, 35, 584–597. [CrossRef]
- Felicetti, T.; Cedraro, N.; Astolfi, A.; Cernicchi, G.; Mangiaterra, G.; Vaiasicca, S.; Massari, S.; Manfroni, G.; Barreca, M.L.; Tabarrini, O.; et al. New C-6 Functionalized Quinoline NorA Inhibitors Strongly Synergize with Ciprofloxacin against Planktonic and Biofilm Growing Resistant Staphylococcus Aureus Strains. Eur J Med Chem 2022, 241, 114656. [CrossRef]
- Cedraro, N.; Cannalire, R.; Astolfi, A.; Mangiaterra, G.; Felicetti, T.; Vaiasicca, S.; Cernicchi, G.; Massari, S.; Manfroni, G.; Tabarrini, O.; et al. From Quinoline to Quinazoline-Based S. Aureus NorA Efflux Pump Inhibitors by Coupling a Focused Scaffold Hopping Approach and a Pharmacophore Search. ChemMedChem 2021, 16, 1–17. [CrossRef]
- Fontaine, F.; Hequet, A.; Voisin-Chiret, A.S.; Bouillon, A.; Lesnard, A.; Cresteil, T.; Jolivalt, C.; Rault, S. First Identification of Boronic Species as Novel Potential Inhibitors of the Staphylococcus Aureus NorA Efflux Pump. J Med Chem 2014, 57, 2536–2548. [CrossRef]
- Fontaine, F.; Héquet, A.; Voisin-Chiret, A.S.; Bouillon, A.; Lesnard, A.; Cresteil, T.; Jolivalt, C.; Rault, S. Boronic Species as Promising Inhibitors of the Staphylococcus Aureus NorA Efflux Pump: Study of 6-Substituted Pyridine-3-Boronic Acid Derivatives. Eur J Med Chem 2015, 95, 185–198. [CrossRef]
- Holler, J.G.; Slotved, H.C.; Molgaard, P.; Olsen, C.E.; Christensen, S.B. Chalcone Inhibitors of the NorA Efflux Pump in Staphylococcus Aureus Whole Cells and Enriched Everted Membrane Vesicles. Bioorg Med Chem 2012, 20, 4514–4521. [CrossRef]
- Kumar, A.; Khan, I.A.; Koul, S.; Koul, J.L.; Taneja, S.C.; Ali, I.; Ali, F.; Sharma, S.; Mirza, Z.M.; Kumar, M.; et al. Novel Structural Analogues of Piperine as Inhibitors of the NorA Efflux Pump of Staphylococcus Aureus. Journal of Antimicrobial Chemotherapy 2008, 61, 1270–1276. [CrossRef]
- Sangwan, P.L.; Koul, J.L.; Koul, S.; Reddy, M. V; Thota, N.; Khan, I.A.; Kumar, A.; Kalia, N.P.; Qazi, G.N. Piperine Analogs as Potent Staphylococcus Aureus NorA Efflux Pump Inhibitors. Bioorg Med Chem 2008, 16, 9847–9857. [CrossRef]
- dos Santos, J.F.S.; Tintino, S.R.; da Silva, A.R.P.; Cristina, C.R.; Scherf, J.R.; de S. Silveira, Z.; de Freitas, T.S.; de Lacerda Neto, L.J.; Barros, L.M.; Irwin, I.R.; et al. Enhancement of the Antibiotic Activity by Quercetin against Staphylococcus Aureus Efflux Pumps. J Bioenerg Biomembr 2021, 53, 157–167. [CrossRef]
- Muniz, D.F.; dos Santos Barbosa, C.R.; de Menezes, I.R.A.; de Sousa, E.O.; Pereira, R.L.S.; Júnior, J.T.C.; Pereira, P.S.; de Matos, Y.M.L.S.; da Costa, R.H.S.; de Morais Oliveira-Tintino, C.D.; et al. In Vitro and in Silico Inhibitory Effects of Synthetic and Natural Eugenol Derivatives against the NorA Efflux Pump in Staphylococcus Aureus. Food Chem 2021, 337, 127776. [CrossRef]
- Espinoza, J.; Urzúa, A.; Sanhueza, L.; Walter, M.; Fincheira, P.; Muñoz, P.; Mendoza, L.; Wilkens, M. Essential Oil, Extracts, and Sesquiterpenes Obtained From the Heartwood of Pilgerodendron Uviferum Act as Potential Inhibitors of the Staphylococcus Aureus NorA Multidrug Efflux Pump. Front Microbiol 2019, 10, 337. [CrossRef]
- Kalia, N.P.; Mahajan, P.; Mehra, R.; Nargotra, A.; Sharma, J.P.; Koul, S.; Khan, I.A. Capsaicin, a Novel Inhibitor of the NorA Efflux Pump, Reduces the Intracellular Invasion of Staphylococcus Aureus. Journal of Antimicrobial Chemotherapy 2012, 67, 2401–2408. [CrossRef]
- Diniz-Silva, H.T.; Magnani, M.; de Siqueira, S.; de Souza, E.L.; de Siqueira-Júnior, J.P. Fruit Flavonoids as Modulators of Norfloxacin Resistance in Staphylococcus Aureus That Overexpresses <i>NorA<i/>. LWT - Food Science and Technology 2017, 85, 324–326. [CrossRef]
- Singh, S.; Kalia, N.P.; Joshi, P.; Kumar, A.; Sharma, P.R.; Kumar, A.; Bharate, S.B.; Khan, I.A. Boeravinone B, A Novel Dual Inhibitor of NorA Bacterial Efflux Pump of Staphylococcus Aureus and Human P-Glycoprotein, Reduces the Biofilm Formation and Intracellular Invasion of Bacteria. Front Microbiol 2017, 8, 1868. [CrossRef]
- de Morais Oliveira-Tintino, C.D.; Tintino, S.R.; Limaverde, P.W.; Figueredo, F.G.; Campina, F.F.; da Cunha, F.A.B.; da Costa, R.H.S.; Pereira, P.S.; Lima, L.F.; de Matos, Y.M.L.S.; et al. Inhibition of the Essential Oil from Chenopodium Ambrosioides L. and α-Terpinene on the NorA Efflux-Pump of Staphylococcus Aureus. Food Chem 2018, 262, 72–77. [CrossRef]
- Mouwakeh, A.; Kincses, A.; Nové, M.; Mosolygó, T.; Mohácsi-Farkas, C.; Kiskó, G.; Spengler, G. Nigella Sativa Essential Oil and Its Bioactive Compounds as Resistance Modifiers against Staphylococcus Aureus. Phytother Res 2019, 33, 1010–1018. [CrossRef]
- de Sousa, J.N.; de Oliveira, A.B.M.; Ferreira, A.K.; Silva, E.; de Sousa, L.M.S.; França Rocha, M.C.; De, J.P.; Júnior, S.; William Kaatz, G.; da Silva Almeida, J.R.G.; et al. Modulation of the Resistance to Norfloxacin in Staphylococcus Aureus by Bauhinia Forficata Link. Nat Prod Res 2021, 35, 681–685. [CrossRef]
- Cernicchi, G.; Felicetti, T.; Sabatini, S. Microbial Efflux Pump Inhibitors: A Journey around Quinoline and Indole Derivatives. Molecules 2021, 26, 6996. [CrossRef]
- Felicetti, T.; Cannalire, R.; Pietrella, D.; Latacz, G.; Lubelska, A.; Manfroni, G.; Barreca, M.L.; Massari, S.; Tabarrini, O.; Kieć-Kononowicz, K.; et al. 2-Phenylquinoline S. Aureus NorA Efflux Pump Inhibitors: Evaluation of the Importance of Methoxy Group Introduction. J Med Chem 2018, 61, 7827–7848. [CrossRef]
- Kaatz, G.W.; Seo, S.M. Mechanisms of Fluoroquinolone Resistance in Genetically Related Strains of Staphylococcus Aureus. Antimicrob Agents Chemother 1997, 41, 2733–2737. [CrossRef]
- Uribe, E.G. ATP Synthesis Driven by a K+-Valinomycin-Induced Charge Imbalance across Chloroplast Grana Membranes. FEBS Lett 1973, 36, 143–147. [CrossRef]
- Koh, J.Y.; Cotman, C.W. Programmed Cell Death: Its Possible Contribution to Neurotoxicity Mediated by Calcium Channel Antagonists. Brain Res 1992, 587, 233–240. [CrossRef]
- Blanchard, C.; Barnett, P.; Perlmutter, J.; Dunman, P.M. Identification of Acinetobacter Baumannii Serum-Associated Antibiotic Efflux Pump Inhibitors. Antimicrob Agents Chemother 2014, 58, 6360. [CrossRef]
- Gandhi, S.; Fleet, J.L.; Bailey, D.G.; McArthur, E.; Wald, R.; Rehman, F.; Garg, A.X. Calcium-Channel Blocker–Clarithromycin Drug Interactions and Acute Kidney Injury. JAMA 2013, 310, 2544–2553. [CrossRef]
- AlMatar, M.; Albarri, O.; Makky, E.A.; Köksal, F. Efflux Pump Inhibitors: New Updates. Pharmacological Reports 2020, 73, 1–16. [CrossRef]
- Pidwill, G.R.; Gibson, J.F.; Cole, J.; Renshaw, S.A.; Foster, S.J. The Role of Macrophages in Staphylococcus Aureus Infection. Front Immunol 2020, 11, 620339. [CrossRef]
- Kumawat, M.; Nabi, B.; Daswani, M.; Viquar, I.; Pal, N.; Sharma, P.; Tiwari, S.; Sarma, D.K.; Shubham, S.; Kumar, M. Role of Bacterial Efflux Pump Proteins in Antibiotic Resistance across Microbial Species. Microb Pathog 2023, 181, 106182. [CrossRef]
- Wertheim, H.F.L.; Vos, M.C.; Ott, A.; Van Belkum, A.; Voss, A.; Kluytmans, J.A.J.W.; Van Keulen, P.H.J.; Vandenbroucke-Grauls, C.M.J.E.; Meester, M.H.M.; Verbrugh, H.A. Risk and Outcome of Nosocomial Staphylococcus Aureus Bacteraemia in Nasal Carriers versus Non-Carriers. Lancet 2004, 364, 703–705. [CrossRef]
- Abbey, T.C.; Deak, E. What’s New from the CLSI Subcommittee on Antimicrobial Susceptibility Testing M100, 29th Edition. Clin Microbiol Newsl 2019, 41, 203–209. [CrossRef]
- Hemaiswarya, S.; Kruthiventi, A.K.; Doble, M. Synergism between Natural Products and Antibiotics against Infectious Diseases. Phytomedicine 2008, 15, 639–652. [CrossRef]
- Zimmermann, S.; Klinger-Strobel, M.; Bohnert, J.A.; Wendler, S.; Rödel, J.; Pletz, M.W.; Löffler, B.; Tuchscherr, L. Clinically Approved Drugs Inhibit the Staphylococcus Aureus Multidrug NorA Efflux Pump and Reduce Biofilm Formation. Front Microbiol 2019, 10, 492781. [CrossRef]
- Wu, M.; Maier, E.; Benz, R.; Hancock, R.E.W. Mechanism of Interaction of Different Classes of Cationic Antimicrobial Peptides with Planar Bilayers and with the Cytoplasmic Membrane of Escherichia Coli. Biochemistry 1999, 38, 7235–7242. [CrossRef]
- Morin, N.; Lanneluc, I.; Connil, N.; Cottenceau, M.; Pons, A.M.; Sablé, S. Mechanism of Bactericidal Activity of Microcin L in Escherichia Coli and Salmonella Enterica. Antimicrob Agents Chemother 2011, 55, 997–1007. [CrossRef]
- Cheng, M.; Huang, J.X.; Ramu, S.; Butler, M.S.; Cooper, M.A. Ramoplanin at Bactericidal Concentrations Induces Bacterial Membrane Depolarization in Staphylococcus Aureus. Antimicrob Agents Chemother 2014, 58, 6819. [CrossRef]
- French, S.; Farha, M.; Ellis, M.J.; Sameer, Z.; Côté, J.P.; Cotroneo, N.; Lister, T.; Rubio, A.; Brown, E.D. Potentiation of Antibiotics against Gram-Negative Bacteria by Polymyxin B Analogue SPR741 from Unique Perturbation of the Outer Membrane. ACS Infect Dis 2020, 6, 1405–1412. [CrossRef]
- Tambat, R.; Jangra, M.; Mahey, N.; Chandal, N.; Kaur, M.; Chaudhary, S.; Verma, D.K.; Thakur, K.G.; Raje, M.; Jachak, S.; et al. Microbe-Derived Indole Metabolite Demonstrates Potent Multidrug Efflux Pump Inhibition in Staphylococcus Aureus. Front Microbiol 2019, 10, 2153. [CrossRef]
- OECD Overview of the Set of OECD Genetic Toxicology Test Guidelines and Updates Performed in 2014-2015 Series On. 2016, 60. [CrossRef]
- di Vito, R.; Levorato, S.; Fatigoni, C.; Acito, M.; Sancineto, L.; Traina, G.; Villarini, M.; Santi, C.; Moretti, M. In Vitro Toxicological Assessment of PhSeZnCl in Human Liver Cells. Toxicol Res 2023, 39, 105. [CrossRef]
- Rondini, T.; Branciari, R.; Franceschini, E.; Acito, M.; Fatigoni, C.; Roila, R.; Ranucci, D.; Villarini, M.; Galarini, R.; Moretti, M. Olive Mill Wastewater Extract: In Vitro Genotoxicity/Antigenotoxicity Assessment on HepaRG Cells. Int J Environ Res Public Health 2024, 21, 1050. [CrossRef]
- Lovell, D.P.; Omori, T. Statistical Issues in the Use of the Comet Assay. Mutagenesis 2008, 23, 171–182. [CrossRef]
- Rázquin-Olazarán, I.; Shahrour, H.; Martínez-De-Tejada, G. A Synthetic Peptide Sensitizes Multi-Drug Resistant Pseudomonas Aeruginosa to Antibiotics for More than Two Hours and Permeabilizes Its Envelope for Twenty Hours. J Biomed Sci 2020, 27, 1–19. [CrossRef]
- Drugeon, H.B.; Juvin, M.E.; Bryskier, A. Relative Potential for Selection of Fluoroquinolone-Resistant Streptococcus Pneumoniae Strains by Levofloxacin: Comparison with Ciprofloxacin, Sparfloxacin and Ofloxacin. J Antimicrob Chemother 1999, 43, 55–59. [CrossRef]
- Ling, L.L.; Schneider, T.; Peoples, A.J.; Spoering, A.L.; Engels, I.; Conlon, B.P.; Mueller, A.; Schäberle, T.F.; Hughes, D.E.; Epstein, S.; et al. A New Antibiotic Kills Pathogens without Detectable Resistance. Nature 2015, 517, 455–459. [CrossRef]
Figure 1.
Chemical structures of NorA EPIs PQK4F and PQQ16P.
Figure 2.
Time-kill kinetic assay, the combination of CPX (2 and 1 µg/mL) and PQK4F (A) and PQQ16P (B) at 12.48 and 6.24 µg/mL, respectively on S. aureus SA-1199B strain. The experiment is the representation of three individual repeats ± SD.
Figure 2.
Time-kill kinetic assay, the combination of CPX (2 and 1 µg/mL) and PQK4F (A) and PQQ16P (B) at 12.48 and 6.24 µg/mL, respectively on S. aureus SA-1199B strain. The experiment is the representation of three individual repeats ± SD.
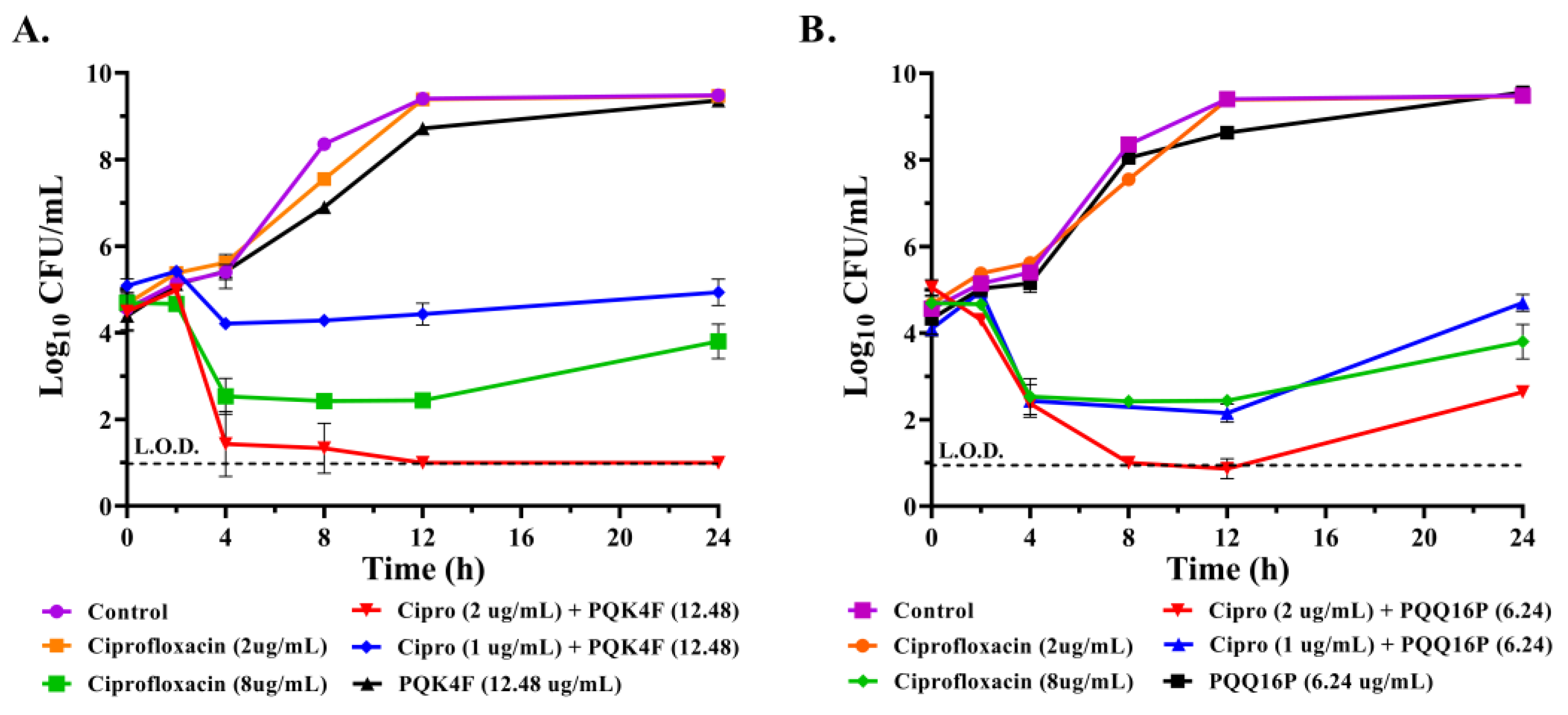
Figure 3.
A. Membrane permeability was measured using nucleic acid binding dye propidium iodide, polymyxin B was used as the positive control, and drug-free control was used as the negative control, B. Membrane depolarization was detected using fluorescence of DiSC3(5) showing high fluorescence of positive control valinomycin, drug-free control was used as the negative control, C. ATP bioluminescence detection assay, CCCP the positive control quenches ATP compared to control. All the results are representation of three individual readings ± SD on SA-1199B.
Figure 3.
A. Membrane permeability was measured using nucleic acid binding dye propidium iodide, polymyxin B was used as the positive control, and drug-free control was used as the negative control, B. Membrane depolarization was detected using fluorescence of DiSC3(5) showing high fluorescence of positive control valinomycin, drug-free control was used as the negative control, C. ATP bioluminescence detection assay, CCCP the positive control quenches ATP compared to control. All the results are representation of three individual readings ± SD on SA-1199B.
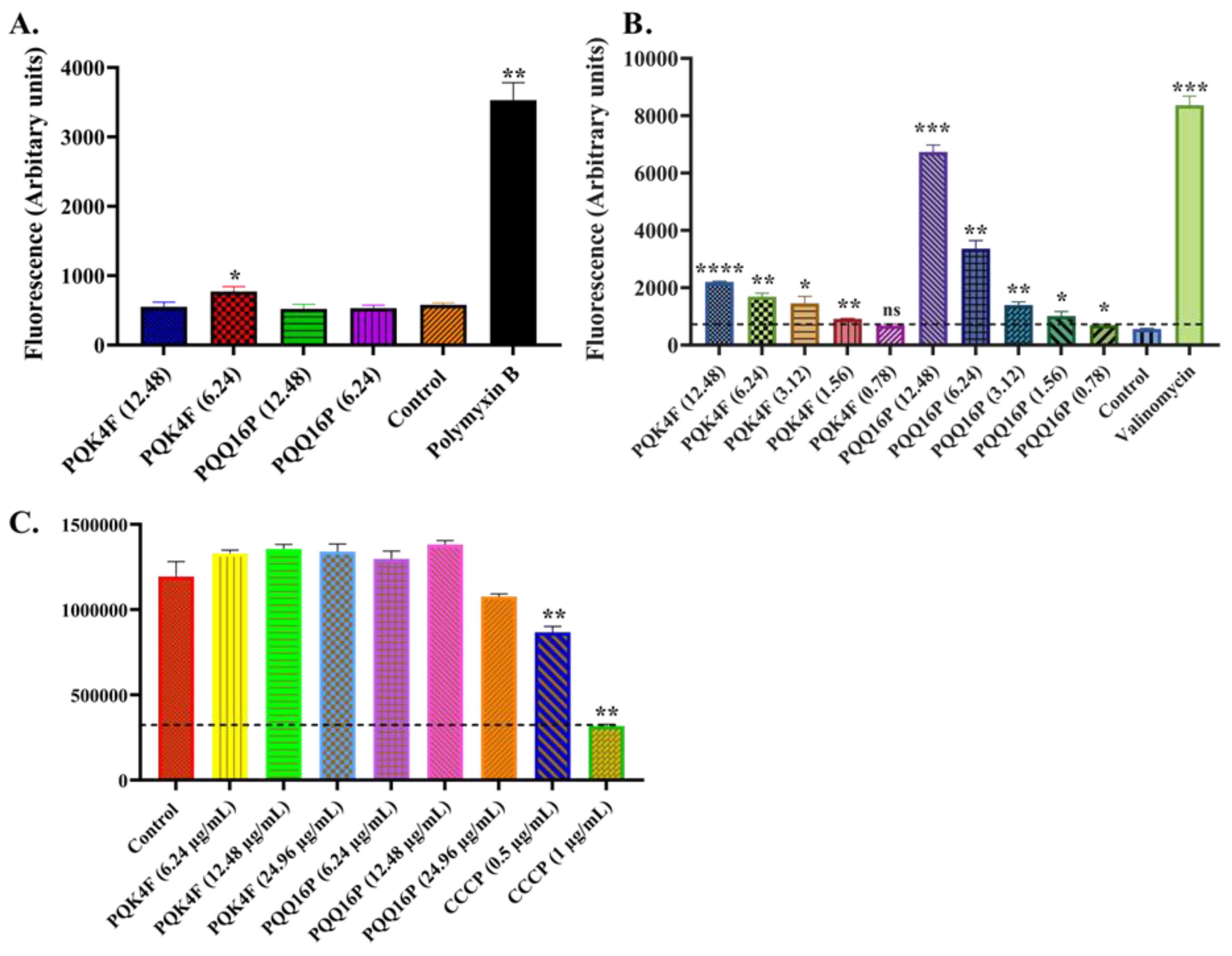
Figure 4.
Activity of PQK4F and PQQ16P on mammalian Ca2+ channels of HEK cells. The results are the representation of three individual readings ± SD.
Figure 4.
Activity of PQK4F and PQQ16P on mammalian Ca2+ channels of HEK cells. The results are the representation of three individual readings ± SD.
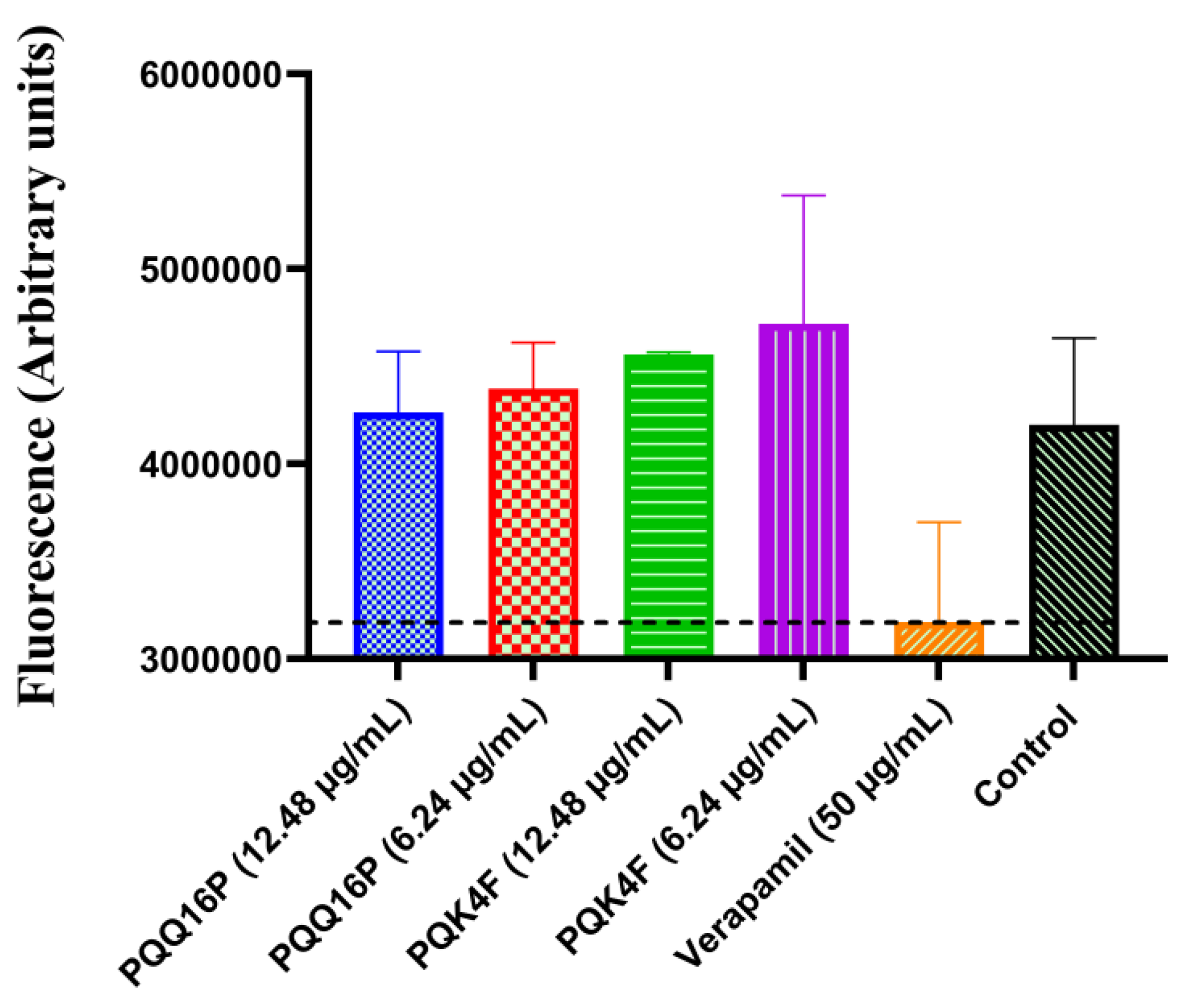
Figure 5.
Cytotoxicity assay on human embryonic kidney cells (HEK 293T), RAW, and HepG2 cells A. of PQK4F and B. PQQ16P at concentrations ranging from 50 to 1.56 µg/mL concentration. The results are the representation of three individual readings ± SD.
Figure 5.
Cytotoxicity assay on human embryonic kidney cells (HEK 293T), RAW, and HepG2 cells A. of PQK4F and B. PQQ16P at concentrations ranging from 50 to 1.56 µg/mL concentration. The results are the representation of three individual readings ± SD.
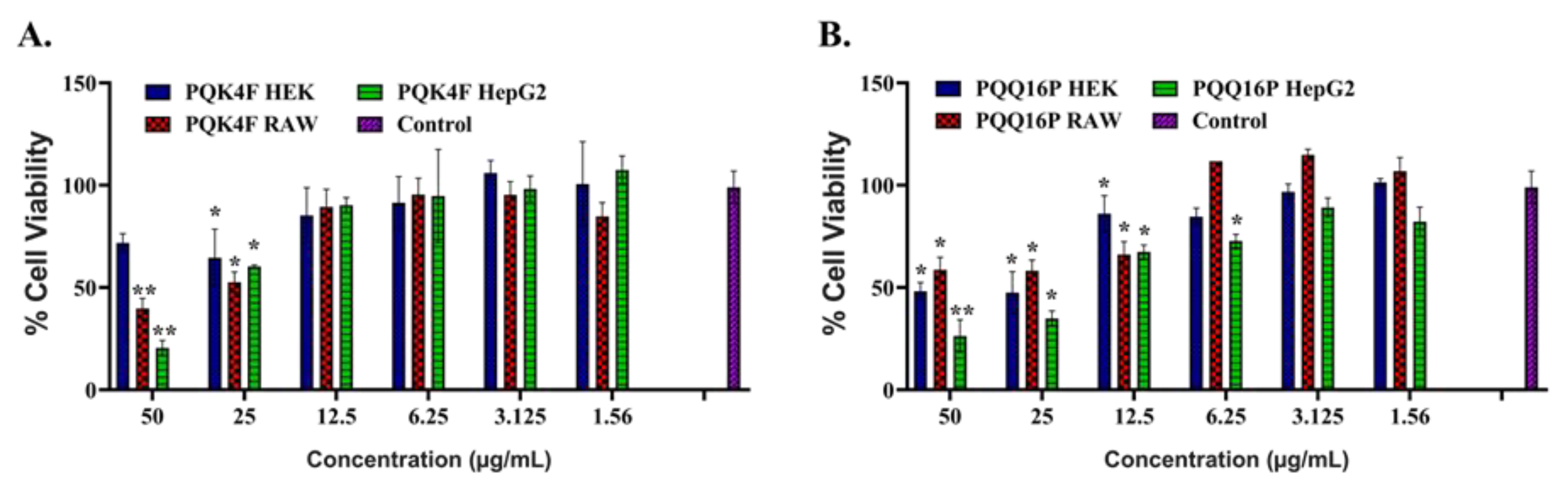
Figure 6.
Hemolysis assay on 4% RBC at 100 to 0.78 µg/mL concentration. The results are the representation of three individual readings ± SD.
Figure 6.
Hemolysis assay on 4% RBC at 100 to 0.78 µg/mL concentration. The results are the representation of three individual readings ± SD.
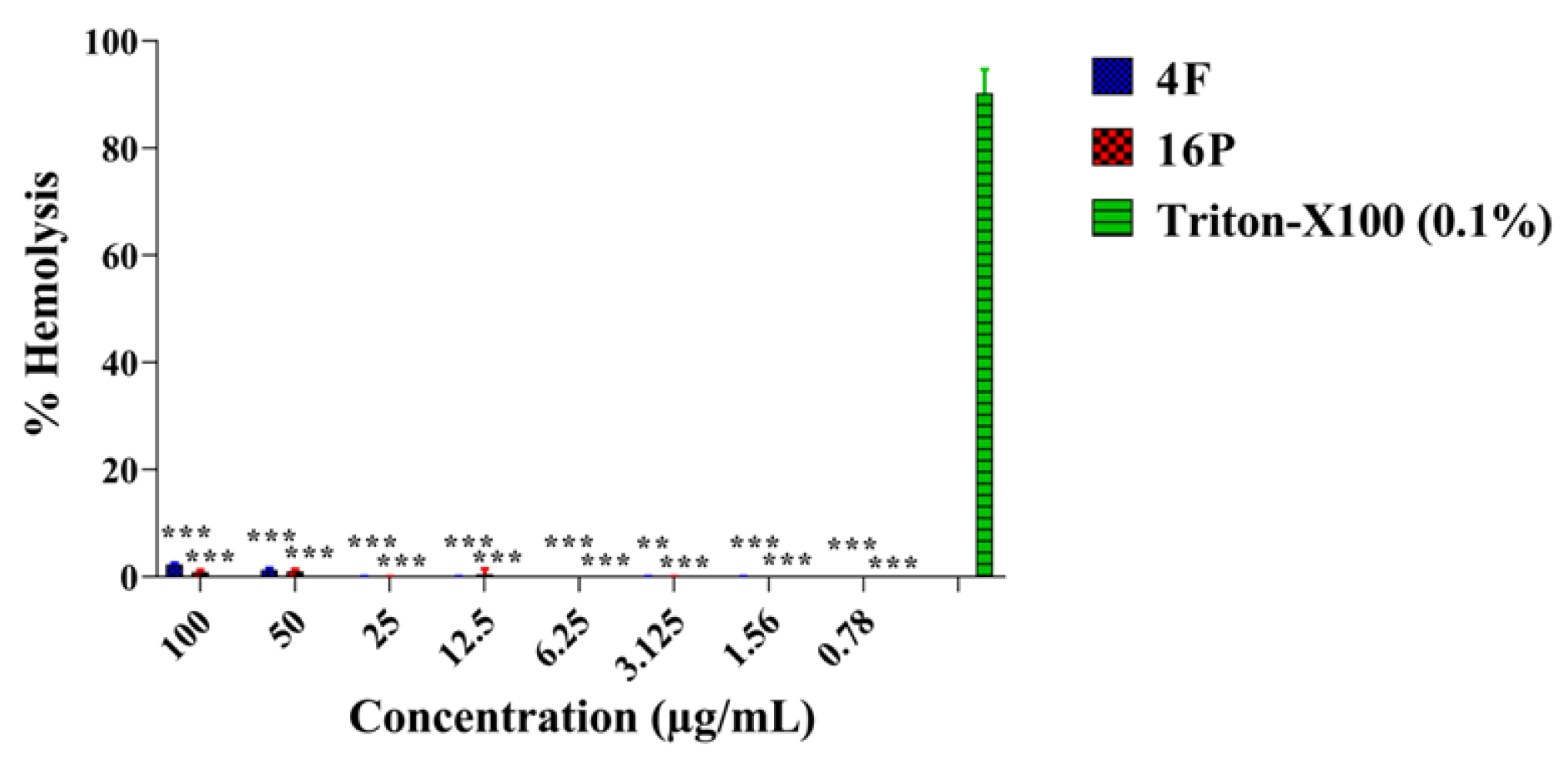
Figure 7.
Genotoxic effects of PQK4F and PQQ16P antibiotics in HEK 293T, RAW 264.7 and Hep G2 cells after 4 hours of exposure. Each result is expressed as the ratio of means with 95% confidence interval of three independent experiments. Negative Control (N.C., untreated cells); Positive control (2 μM 4NQO).
Figure 7.
Genotoxic effects of PQK4F and PQQ16P antibiotics in HEK 293T, RAW 264.7 and Hep G2 cells after 4 hours of exposure. Each result is expressed as the ratio of means with 95% confidence interval of three independent experiments. Negative Control (N.C., untreated cells); Positive control (2 μM 4NQO).
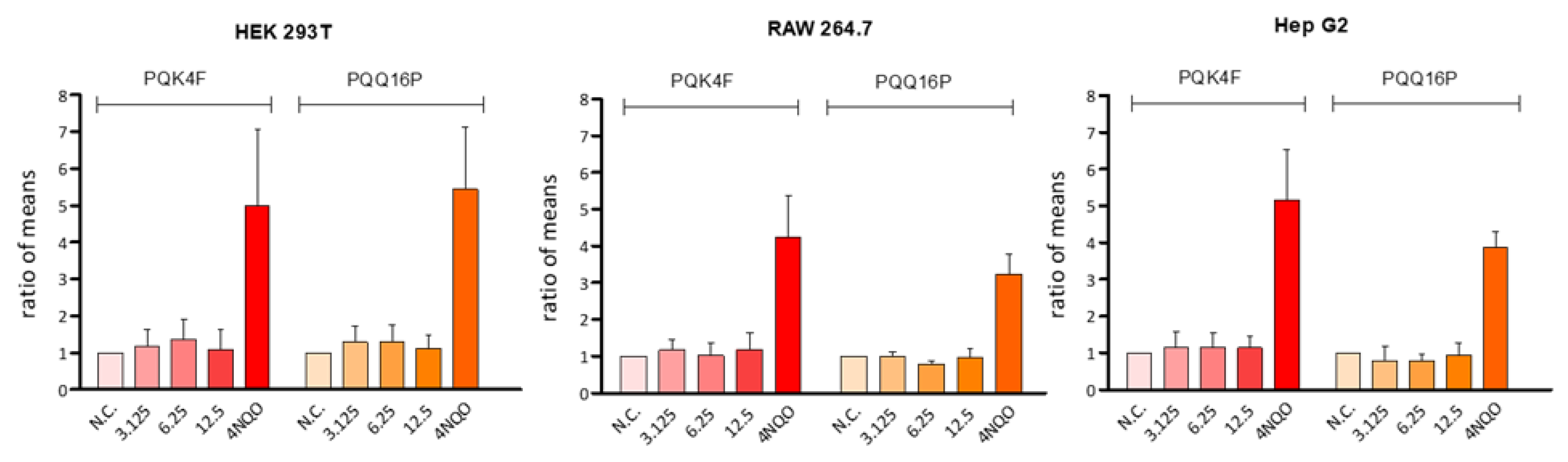
Figure 8.
Post-antibiotic effect of CPX at 2 and 8 µg/mL alone and in combination with PQK4F and PQQ16P at 12.48, 6.24, and 3.12 µg/mL alone and in combination with CPX at 2 µg/mL by turbidity method on S. aureus SA-1199B strain in h; the results are the representation of three individual readings ± SD.
Figure 8.
Post-antibiotic effect of CPX at 2 and 8 µg/mL alone and in combination with PQK4F and PQQ16P at 12.48, 6.24, and 3.12 µg/mL alone and in combination with CPX at 2 µg/mL by turbidity method on S. aureus SA-1199B strain in h; the results are the representation of three individual readings ± SD.
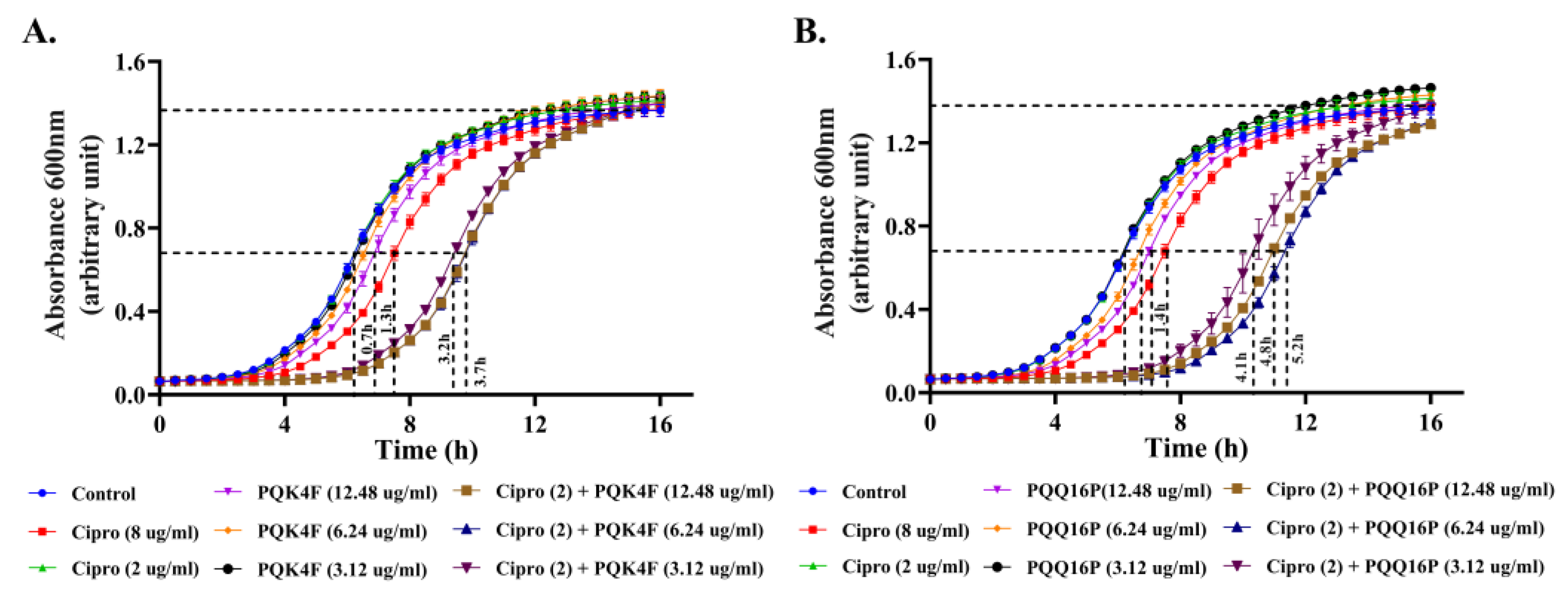
Figure 9.
Macrophage invasion assays were conducted on S. aureus strains SA-1199B (norA over-expressing), SA-1199 (norA wild type), and K-1758 (norA knockout) in the presence of PQK4F and PQQ16P at 12.48 and 6.24 µg/mL concentrations, the results are the representation of three individual readings ± SD.
Figure 9.
Macrophage invasion assays were conducted on S. aureus strains SA-1199B (norA over-expressing), SA-1199 (norA wild type), and K-1758 (norA knockout) in the presence of PQK4F and PQQ16P at 12.48 and 6.24 µg/mL concentrations, the results are the representation of three individual readings ± SD.
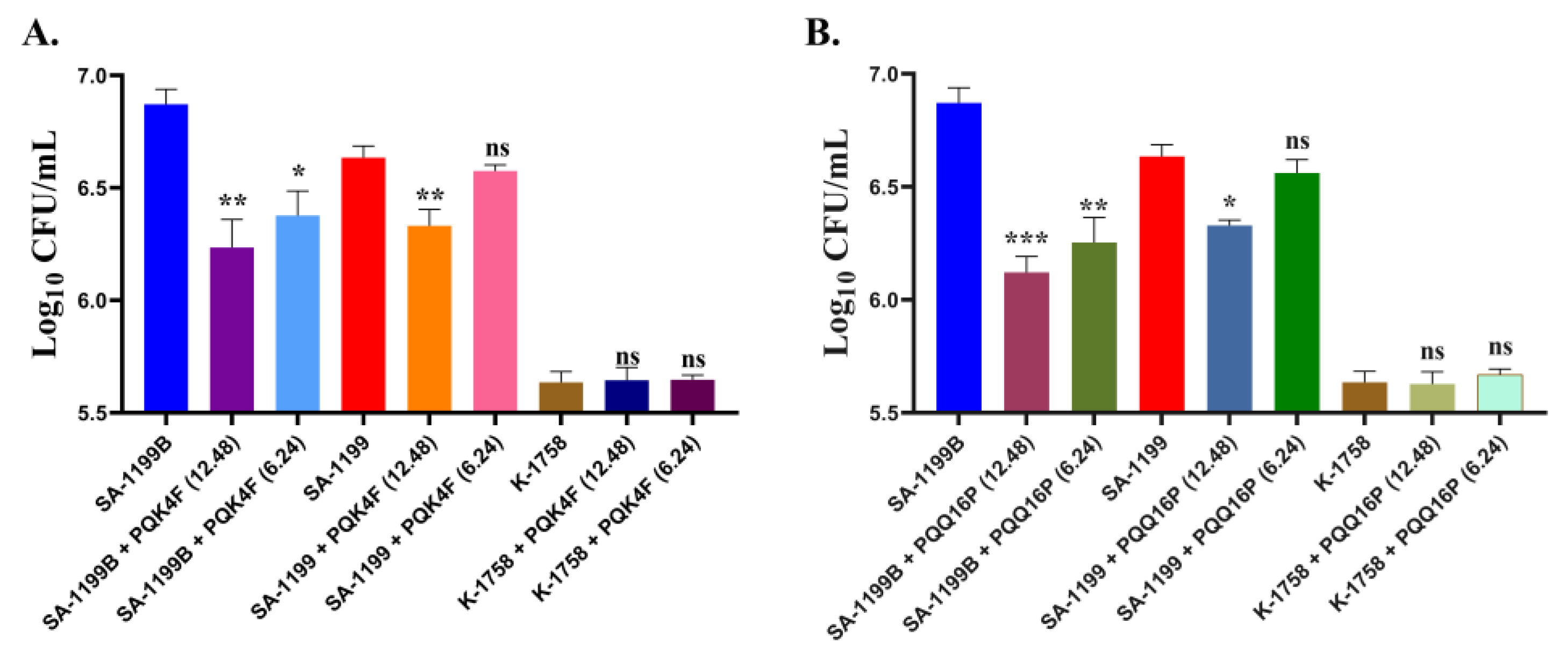
Figure 10.
Neutropenic mouse thigh infection model: Single subcutaneous dose treatment of 5 and 10 mg/Kg PQK4F and PQQ16P, alone and in combination with 10 mg/Kg CPX, administered 4 h post-infection. Six mice per group were used. CFU determined 24 h post-infection for drug-treated mice; controls assessed at 4 and 24 h post-infection for CFU calculation.
Figure 10.
Neutropenic mouse thigh infection model: Single subcutaneous dose treatment of 5 and 10 mg/Kg PQK4F and PQQ16P, alone and in combination with 10 mg/Kg CPX, administered 4 h post-infection. Six mice per group were used. CFU determined 24 h post-infection for drug-treated mice; controls assessed at 4 and 24 h post-infection for CFU calculation.
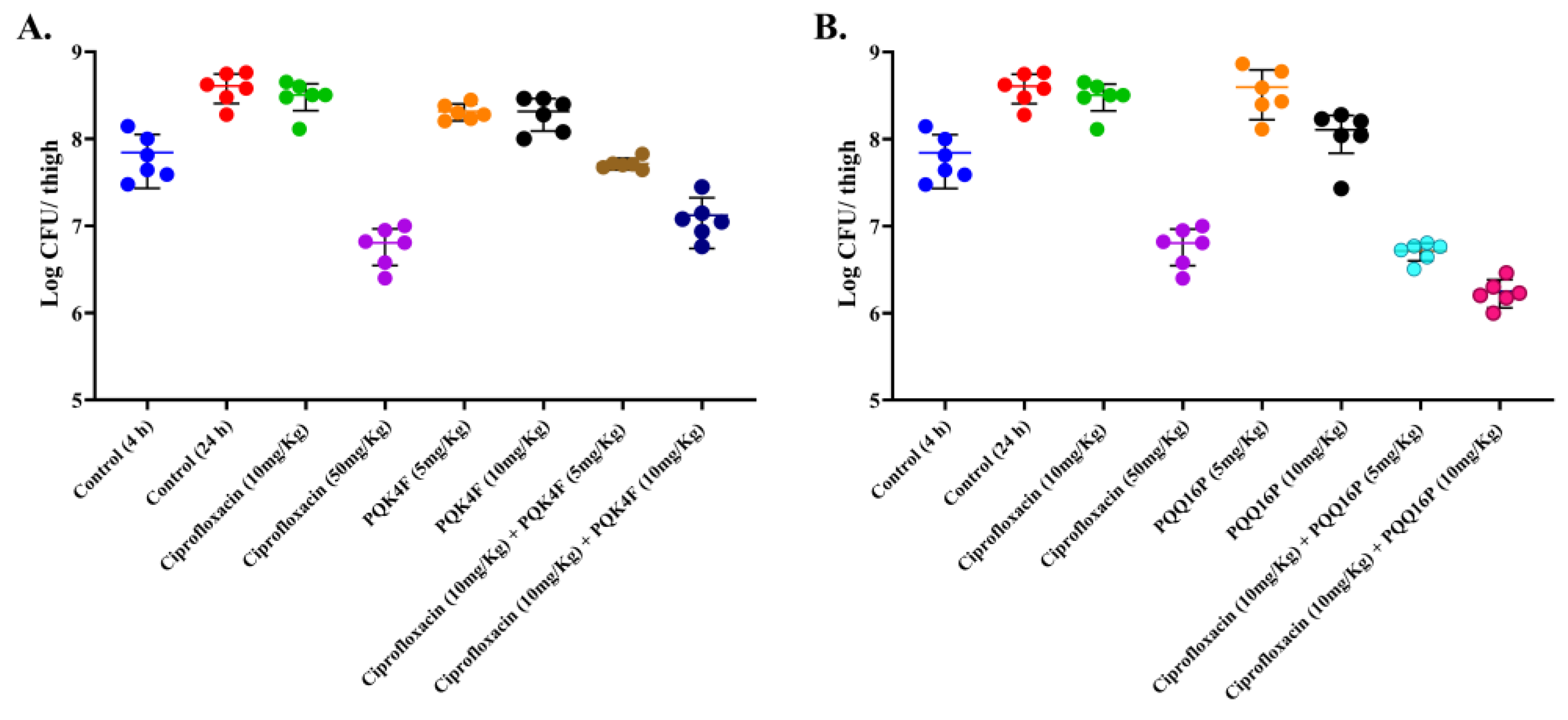

Table 1.
Mutation Prevention concentration of CPX alone at various concentrations (0.25 to 8 × MIC) and in combination with PQQ16P at 6.24 and 12.48 µg/mL and PQK4F at 3.12 and 6.24 µg/mL on S. aureus SA-1199B.
Table 1.
Mutation Prevention concentration of CPX alone at various concentrations (0.25 to 8 × MIC) and in combination with PQQ16P at 6.24 and 12.48 µg/mL and PQK4F at 3.12 and 6.24 µg/mL on S. aureus SA-1199B.
| CPX Conc. | 0.25×MIC | 0.5×MIC | 1×MIC | 2×MIC | 4×MIC | 8×MIC |
|---|---|---|---|---|---|---|
| CFU count | ||||||
| CPX | UC | UC | UC | 2.75 × 10-9 | 2.3 × 10-8 | < 10-9 |
| + PQK4F (6.24 µg/mL) | UC | 1 × 10-9 | < 10-9 | < 10-9 | < 10-9 | < 10-9 |
| + PQK4F (12.48 µg/mL) | 5.26 × 10-9 | 3 × 10-7 | < 10-9 | < 10-9 | < 10-9 | < 10-9 |
| + PQQ16P (3.12 µg/mL) | UC | 1.67 × 10-9 | < 10-9 | < 10-9 | < 10-9 | < 10-9 |
| + PQQ16P (6.24 µg/mL) | UC | 1 × 10-7 | < 10-9 | < 10-9 | < 10-9 | < 10-9 |
1 UC = Uncountable, CPX (MIC) = 8 µg/mL.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



