Submitted:
16 February 2025
Posted:
17 February 2025
You are already at the latest version
Abstract
Background: Adult B-cell acute lymphoblastic leukaemia (aB-ALL) is characterised by abnormal differentiation and proliferation of lymphoid progenitors. Despite a significant improvement in relapse-free and overall survival for children with B-ALL, aB-ALL has a particularly poor prognosis with a 5-year survival rate of 20%. First remission is achieved for most patients but relapse is common with a high associated mortality. New treatments such as immunotherapy offer an opportunity to extend remission and prevent relapse. Methods: aB-ALL antigens were identified using different sources - immunoscreening, protoarrays, two microarrays and one cancer-testis antigen (CTA) database, and a review of the genomic analyses of aB-ALL. 385 aB-ALL-associated gene products were examined for their association with patient survival. Results: We identified 87 transcripts with differential expression between aB-ALL and healthy donors, and 42 that were associated with survival. Enrichr analysis showed that the Transforming Growth Factor-β (TGFβ), Wnt and hippo pathways were highly represented (p<0.02). We found that SOX4 was upregulated in all types of B-ALL, while ROCK1 was upregulated in all types of B-ALL (p<0.001 except t(8;14)) and along with SMAD3 and TEAD4 were associated with survival (p=0.0008, 0.05 and 0.001 respectively). Expression of each aB-ALL antigen was verified by qPCR but only TEAD4 showed significant transcript upregulation in aB-ALL compared to healthy donor CD19+ cells (p=0.01). Conclusions: We have identified a number of antigens and their pathways that play key roles in aB-ALL and may act as useful targets for future immunotherapy strategies.
Keywords:
adult B-cell acute lymphocytic leukaemia
; antigen discovery
; cancer vaccines
; SMAD3
; TEAD4
; immunotherapy
1. Introduction
Adult B-cell acute lymphoblastic leukaemia (aB-ALL) occurs due to an impairment in B-cell differentiation leading to an accumulation of blast cells in the bone marrow. ALL has a bimodal distribution in terms of age, with peak incidences in children aged 2-5 years, and in adults (those aged 18 and over) when they over 40 years of age. Adults over 60 account for 20% of ALL cases and 50% of ALL-related deaths [1]. Although early induction treatment can result in first remission for 80-90% of B-ALL patients, long-term survival is hampered by resistance, relapse and extramedullary infiltration [2]. For this reason, the monitoring of minimal residual disease has been used to determine prognosis, best practise treatment options and risk of relapse for patients [3].
First line treatment for aB-ALL involves pre-phase and induction therapy followed by consolidation and maintenance. Although 80-90% aB-ALL patients achieve first remission more than half relapse. In recent years a number of immunotherapy strategies including antibody and chimeric antigen receptor (CAR) T-cell therapies have been used to break immunological tolerance and changed treatment outcomes for patients with relapsed/refractory (R/R) ALL following chemotherapy and haematopoietic stem cell transplant. Blinatumomab, a bispecific T cell engager that directs CTL to CD19 expressing cells, inotuzumab ozogamicin, an anti-CD22 antibody conjugated to calichaemicin, and tisangeneucel, a CAR T cell therapy have all achieved improved outcomes for patients with R/R ALL when compared to standard therapies [4,5,6]. Despite its efficacy, CAR-T cell therapy is associated with adverse effects including B-cell aplasia, increased infection susceptibility due to the impairment of antibody production, and more serious events such as cytokine release syndrome leading to neurotoxicity and multiple organ dysfunction [7]. In addition, the time-consuming process of CAR-T cell engineering, antigenic variability due to ALL being a heterogeneous disease and trying to develop off-the-shelf therapies represent a real challenge [8].
Recently the paradigm has shifted to immune-checkpoint inhibitors, although these can lead to excessive immune stimulation [9]. To address this issue, "armoured" chimeric antigen receptors (CARs) that secrete PD-1-blocking cytokines or co-express dominant-negative transforming growth factor-B receptor type II to create a proinflammatory environment [10] were utilised. However, long-term use of immunotherapy can lead to the selection of leukaemic clones that are resistant to treatment. This evolutionary process hampers the effectiveness of immunotherapy by selecting subclones that are unresponsive to current therapy, resulting in escape variants and relapse. This phenomenon has been observed in patients with the KMT2A rearrangement who were treated with CAR-T cells and CD19 targetting-immunotherapy for B-ALL, which eventually became ineffective due to the presence of an expanding population of CD19 negative clones [11].
In addition to the immunotherapy targets supra vide, the identification of antigens associated with survival in leukaemia, especially for subtypes associated with poor prognosis and phenotypic plasticity, remains an attractive approach for B-ALL treatment. Sero-profiling of B-ALL compared to age- and sex-matched healthy donor samples revealed three differentially recognised tumour antigens, bone marrow tyrosine kinase (BMX), DCTPP1, and VGLL4 [12]. BMX is found on chromosome Xp22.2, and expressed in bone marrow cells. It belongs to the Tec kinase family which is a non-receptor tyrosine kinase involved in many signal transduction processes such as PI3K and TNF pathways. In addition to its expression in bone marrow cells, BMX is upregulated in prostate cancer and non-malignant conditions such as wound healing where it causes the proliferation of skin keratinocytes [13]. Targeting of BMX has been achieved using the epidermal growth factor receptor inhibitor, BMX-IN-1, and CTN06, which are small molecule inhibitors of both BMX and BTK, suppressing tumour growth and migration via the induction of autophagy and apoptosis. Ibrutinib and zanubrutinib are BTK inhibitors, the latter with improved specificity for BTK, and both of which have been used in clinical trials as treatments for mature B-malignancies such as R/R chronic lymphoblastic leukaemia and small lymphocytic lymphoma [14]. Ibrutinib was found to cause a higher than expected level of adverse events and premature discontinuation in clinical trials [14] while zanubrutinib has been found to have a more acceptable safety profile with improved response rates and progression-free survival [15,16].
Boullosa et al. [17] found that baculoviral IAP repeat containing 5 (BIRC5)/Survivin was expressed in aB-ALL but not healthy donors (p= 0.015). This anti-apoptotic protein is found at low levels in terminally differentiated healthy tissues but is upregulated in many cancers such as lung and breast cancer due to the high expression of oncogenes (JAK/STAT, Akt/PI3K and TCF-β-catenin pathways) and loss of tumour suppressor genes (P53 pathway) promoting tumour proliferation and survival [18]. BIRC5 overexpression is also associated with chemotherapy resistance and tumour aggression while BIRC5 knockdown results in the induction of apoptosis of leukaemia cells and increased chemosensitivity in-vitro [19]. BIRC5 and BMX are newly identified TAAs in B-ALL [12,17] and offer an opportunity for existing therapies to be repurposed to aB-ALL.
Due to the existence of immune escape variants, the heterogeneity between and within aB-ALL tumour cells, and the limitations with existing immunotherapy treatments (generating a lasting anti-tumour immune response), it is essential to broaden the number, specificity and sensitivity of TA targets available for future clinical use [20]. The search for leukaemia associated and ideally cancer-restricted antigens, that are effective targets to stimulate the immune destruction of aB-ALL cells, may help in the development of new treatments that boost anti-leukaemia immune responses, while retaining the specificity needed to minimise off-target effects. To maximise the identification of new targets for treatment, and enable the repurposing of already approved drugs for B-ALL patients, aB-ALL TAAs were identified from six sources - immunoscreening, protoarrays, two microarrays and one cancer testis-antigen (CTA) database, and a review of the genomic analyses of aB-ALL, and their relationship with patient survival was determined.
2. Materials and Methods
2.1. Patient Samples
This study received local ethical approval (REC Reference: 07/H0606/88; LREC 228/02/T) and samples were collected (Table 1 and Table 2) following informed consent. On the day of sample receipt, serum was collected from clotted PB samples following centrifugation at 1200xg for 10 min. Bone marrow (BM) and peripheral blood (PB) samples collected in K2-EDTA tubes, were incubated with 5 volumes of red cell lysis buffer (155 mM NH4Cl, 10 mM KHCO3, and 0.1 mM EDTA) for 30 min at room temperature. Leukocytes were isolated by centrifugation for 10 min at 800xg. Healthy donors were self-selecting individuals who responded to a local request for samples. They had no signs of illness and were not undergoing any investigations by a health care professional. CD19+ B-cells were used as a control as pre-normal B cells could not be sourced. CD19+ is expressed on most B-ALL cells, normal pre-B and B-cells up until maturation to plasma cells and were purchased from cambridge bioscience (Cambridge, UK) after isolation via negative immunomagnetic separation from leukophoresate. The donor was a female aged 18-66 and cells were shipped on dry ice. Isolated leukocytes were resuspended at 5 x 106/ml in PBS and 5ul spotted onto two sites on each microscope slide. Once dried for 4-16h, slides were saran wrapped and stored at -20oC for future immunocytochemistry (ICC) analysis. Remaining leukocytes, were pelleted and along with sera samples were immediately stored at −80°C for later use. K562 (ATCC) and A549 (Sigma-Aldrich Co. Ltd) were cultured in RPMI or DMEM, respectively, 10% foetal calf serum, 1% L-Glutamine and 1% penicillin streptomycin (all Sigma-Aldrich Co Ltd) in a 5% CO2 in a humidified chamber. K562 cells were used as a positive control for the expression of all antigens of interest in this study except YAP1, while A549 was used as a positive control for YAP1 expression.
2.2. Antigen Identification
Genes and antigens were identified from six sources as follows:-
- (i) Serological analysis of recombinant cDNA expression libraries (SEREX)
The testis cDNA expression library was made using the ZAP Express® cDNA synthesis and ZAP Express® cDNA Gigapack® III Gold cloning kit (Stratagene Europe) as described previously [21]. SEREX was performed following an optimised protocol [22] based on the original method [23]. The reactivity of serum was confirmed through agglutination tests and >106 plaques in the testes cDNA library were immunoscreened using pre-cleared serum from five representative aALL patients (ALL004;005;010;017;018; Table 1). cDNA in plaques that were reactive in two independent rounds of immunoscreening were amplified using T3 and T7 primers and Ready Mix Taq PCR Reaction Mix with MgCl2, (Sigma Aldrich, UK). PCR products were gel purified (QIAQuick gel extraction kit, Qiagen Ltd., Lancashire, UK), Sanger sequenced (Department of Biochemistry, University of Cambridge) and analysed using Applied Biosystems’ Sequencing Analysis Software. Sequences were compared to expressed sequence tag (EST) and protein databases, including the National Centre for Biotechnology Information (NCBI) BLAST and these aALL antigens were added to the pool for survival and pathways analysis.
- (ii) & (iii) Differentially expressed genes (DEGs) identified through the analysis of two microarray databases
DEGs in aB-ALL were identified using (i) the GSE38403 [25] and (ii) the Microarray Innovations in leukaemia (MILE; GSE13204) microarray datasets via BloodSpot.eu. 649 aB-ALL samples and 74 HVs were utilised to compare gene transcription levels in ALL patient subgroups. Patient samples were collected from eighteen diagnostic gold standard categories . The read count was downloaded from GEO and gene expression was calculated by the Fragments per Kilobase per Million Mapped Fragments (FPKM) method. Data was normalised using the limma method and DEGs were identified (fold change ≥1.5 for upregulated genes and ≤1.5 for downregulated one) by the DESeq2 method which is part of the 'Bioconductor' package. The adjust p-value (qp) and p-value ≤0.05were calculated using the Fisher’s test [24].
The MILE study includes a limited number of samples from childhood B-ALL (cB-ALL) for C3 (c-ALL/pre-B-ALL with t(9;22)) and C8 (c-ALL/pre-B-ALL without t(9;22) which is known as Philadelphia (Ph) and Philadelphia-like ALL (Ph-like ALL) cytogenetics) subclasses [27] as well as a pre-ponderance of aB-ALL.
- (iv) A review of genomic studies
We also analysed all of the genes identified by Iacobucci and Mullighan’s [28] review of genomic studies. Genes with a corrected Spearman’s correlation false detection rate (FDR) corrected to p>0.05 were filtered out and the remaining genes were arranged in accordance with decreasing Spearman’s value, thus creating a ranked correlation file. These aB-ALL antigens were added to the pool of genes for survival and pathways analysis.
- (v) Proto-array analysis
18 antigens had previously been identified using protoarrays, by virtue of their preferential recognition by antibodies in patients but not healthy donor sera [12] and these were added to the pool of antigens for further analysis.
- (vi) the CTA database
2.3. Pathway Analysis and Survival
The molecular pathways utilised by the antigens were determined using Enrichr [33,34] and the core genes in these selected pathways were examined for their protein-protein interaction networks using the Search Tool for the Retrieval of Interacting Genes (STRING) analytical program [35]. We determined which antigens, when transcribed at above and below median levels were associated with aB-ALL patient survival using the MILE dataset accessed via bloodspot.eu [32]. This identified gene expression data from 649 B-ALL patients. We then priority-ranked the list of cancer antigens based on predefined and preweighted objective criteria following the analytical hierarchy process developed by a panel of 80 experts and described by Cheever et al [36].
2.4. qPCR
RNA extraction was performed on 11 PB and 5 BM samples from 15 aALL patients and 6 PB samples from healthy donors, including one sample of CD19+ purified cells, using the RNeasy Mini Kit® (Qiagen) and following the manufacturer’s guidelines. The genomic DNA elimination mix was prepared in a RNAse and DNAse free PCR tube according to the manufacturer’s instructions (MBI Fermentas). Quantitative PCR (qPCR) was performed using the RT² SYBR® Green Master-mix, RT² qPCR Primer Assay and cDNA synthesis reaction (all Qiagen) on a 96-well qPCR plate (Applied Biosystems, USA). TATA-box binding protein (TBP1) and protein kinase cGMP-dependent 1 (PRKG1) were used as reference genes following MIQE guidelines [37]. Primers from Qiagen were used to detected each transcript: SRY-Box Transcription Factor 4 (SOX4; PPH01950A), Rho associated coiled-coil containing protein kinase 1 (ROCK1; PPH01966C), Yes-associated protein (YAP1; PPH13459A), TEA Domain Transcription Factor 4 (TEAD4; PPH10558A-200), SMAD family member 3 (SMAD3; PPH01921C), and T cell receptor 4 (TCF4; PPH02770A) and BIRC5 (PH00271E) and at least one human cancer cell line that was known to express the gene of interest, based on previously published studies [38,39] was used as a positive control. Every sample was analysed for expression of each gene in triplicate. To control for gDNA contamination within the qPCR reagents, a no cDNA control was included on every qPCR plate whereby cDNA was replaced by RNase-free H2O. Otherwise the reaction volumes were 10 µL 2X QuantiNova SYBR green PCR Mix, 2 µL QN ROX reference dye, 2 µL QuantiTect primer assay and 5 µL RNase-free H2O, making a total volume of 19 µL added to each well in the 96-well plate. In 1 µL final volume, ≤100 ng of template cDNA was added to achieve a final volume of 20 µl. The plate was then loaded into the thermocycler (StepOne Plus Real-Time PCR system, Applied Biosystems).
The cycle was 2 minutes at 95°C, then 40 cycles of 5 s at 95°C followed by 10 s at 60°C. This was immediately followed by a melt curve stage of 15 s at 95°C, 1 min at 60°C and 15 s at 95°C, to verify the specificity of the amplification. Data was collected using StepOne software v2.0 (Applied Biosystems) and the results normalised using the TBP1 and PRKG1 housekeeping genes (ΔCT = CT genes of interest – CT housekeeping genes). The 2-ΔΔCT method [40] log transforms the ΔΔCT (where ΔΔCT = ΔCT patient sample – ΔCT healthy donor) to determine the relative expression of the seven genes of interest (BIRC5, ROCK1, SMAD3, SOX4, TCF4, TEAD4 and YAP1).
2.5. Immunocytochemistry
Immunocytochemistry was performed as described previously [41] using monoclonal rabbit anti-human antibodies against TEAD4 (1D10) and SMAD3 (2C12) (ThermoFisher Scientific, Leicestershire, UK)(Table S1). Mouse anti-human monoclonal Actin antibody (ACTN05 (C4), Fisher Scientific) was used as a positive control for immunolabelling, BIRC5 as a comparator [17], while isotype controls (mouse or rabbit anti-human IgG antibodies) were used to determine whether there was any non-specific antibody binding. Secondary antibodies were provided as part of the EnVision+ System/HRP, Rabbit(DAB+) kit (DAKO, USA). Lillies-Mayer Haematoxylin was used as a counterstain. Staining intensities were scored according to a five-tiered scale described originally by [42] as follows: - 0: no staining; 1: background; 2: weak staining; 3: moderate staining; 4: strong staining. The percentage of positively stained cells was based on the cell count of stained cells per microscopic view and represented on a five-tiered scale (0: 0 %; 1: 1–10%; 2: 11–50%; 3: 51–80%; 4:>80%). The final immunoreactivity score was obtained by multiplication of the percentage of positive stained cells scored over ≥5 microscopic views by the value for staining intensity within the same [43].
3. Results
3.1. Immunoscreening Identified 72 aB-ALL Associated Antigens
Primary immunoscreening of the testes cDNA library with pre-cleared aB-ALL patient sera identified 310 sero-positive plaques. In total 134 phage colonies were confirmed as expressing immunogenic polypeptides through a second independent round of immunoscreening. PCR amplification of cDNA inserts identified 69 independent known genes (Table S2) and three novel sequences (UOH-ALL-104, UOH-ALL-105, UOH-ALL-106). The function and distribution of the identified genes in cancer and healthy tissues were determined (Table S3) through a comprehensive search of the literature. The expression of SEREX identified genes were examined using data from the MILE study and compared to bone marrow from HVs from the same (Table S3).
3.2. Association Between Leukaemia Associated Antigen (LAA) Expression in aB-ALL Cells and Patient Survival
Of the 385 genes identified from the six sources ((i) SEREX; (ii) GSE38403; (iii) GSE13204; (iv) Iacobucci and Mullighan’s review of genomic studies [28]; (v) proto-array analysis and (vi); CTA database; Figure 1), 87 transcripts were found to have differential expression (Figure 2) and 42 of these had expression levels (above and below media) that were associated with survival in aB-ALL patients, determined using the MILE study data (Table S4).
3.3. TGFβ, Wnt and Hippo Pathways Were Highly Represented by the DEGs in aB-ALL Samples
Transforming Growth Factor-β (TGFβ), Wnt and hippo pathways were highly represented by the genes (p≤0.001, ≤0.01 and ≤0.01 respectively), when analysed by Enrichr (Figure 3A). The core genes from these pathways (TGFβ, Wnt and hippo pathways) were then examined using the MILE dataset for their association with patient survival (n=11) (Table S4) and literature searches were performed to identify genes known to be involved in B-ALL. SOX4 and YAP1 were added to the antigen ranking as SOX4 was considered to be non-conical activator for three pathways [44] whereas YAP1 has restricted expression in healthy tissues (excluding stem/progenitor populations) [45].
STRING showed that PMP2, PMP4, CDKN2B, RBX, CUL1 were involved in TGFβ enrichment (Figure S1) while Wnt2B, CDKN2B, RBX, PMP2, PTGS2 were involved in cancer pathways. SEREX identified genes that were involved in ubiquitination including UBC, UBE2C, UBE2D2 and CUL1 and genes involved in regulating the intrinsic apoptosis process (BCLAF2, PARL, SERINC3, TRAP1, TPT1, MCL1 and PTPN1).
Enrichr showed that the SEREX-identified antigens were mostly involved in molecular functions involving binding and catalytic activity, and the Wiki pathway (Table 3). In addition, envelope proteins (TCF7L2, Wnt2B) essential to maintaining the structure and function of the nucleus, mainly correlated with Emery-Dreifuss muscular dystrophy (EDMD). From MSigDB Hallmark 2020, the mitotic spindle pathway had a high p-value of 0.001 and this pathway plays a crucial role in cell division and chromosomal separation. Using PANTHER.db, the most enriched pathway was the Cadherin signalling pathway (14.3%) and PTPN1, TCF7L2, FER and Wnt2B were also identified by SEREX.
3.4. Most of the aB-ALL Antigens Were Differentially Expressed in Solid Tumours
Of the antigens that were ranked by the Cheever system (Figure 2A), ubiquitin C (HMG20/UBC) and MOB kinase activator 1B (MOB1B) were rarely detected in the healthy tissues (Figure S2A), while SOX4 was found at low to moderate levels in most solid tumours. ROCK1 and YAP1 had very similar patterns of expression in solid tumours except lymphomas where ROCK1 was upregulated and YAP1 was not detected. SMAD3 was upregulated in renal cancers. TGFβ1 was not found in most solid tumours or found at low levels, while TCF4, HMG20 and septin 9 (SEPT 9) weren’t found in any of the solid tumours investigated (Figure S2B).
3.5. SOX4 and TGFβ1 Were the Top-Ranking aB-ALL Vaccine Targets
aB-ALL antigens that impacted survival were ranked using the criteria described by Cheever et al [36] to prioritise cancer antigens on their capacity to act as vaccine targets in clinical studies (Figure 3B). SOX4 (0.89) and TGFβ1 (0.81) had the top scores while ROCK1, YAP1, TEAD4, SMAD3 and TCF4 had the next highest cumulative scores of 0.41, 0.36, 0.34, 0.33, and 0.32 respectively. TGFβ1 was downregulated in aB-ALL compared to healthy donor samples (p<0.0001) reflecting its activity as a tumour suppressor [46] and was excluded from further analysis.
3.6. TCF4, SOX4 and SMAD3 Transcription Was Increased in Almost All aB-ALL Subtypes
We analysed the expression levels of the six most highly ranked aB-ALL antigens (SOX4, ROCK1, TCF4, TEAD4, SMAD3 and YAP1), identified using the Cheever system, and used BIRC5 as a comparator, in 16 aALL PB and BM samples compared to four HV PB and one CD19+ sample by qPCR. The K562 cell line was used as a positive control for BIRC5, TCF4, SMAD3 and ROCK1 expression while A549 cells were used as a positive control for YAP1. Transcription of all aALL-antigens were elevated, while BIRC5, SMAD3 and TEAD4 had the highest mean fold change in aALL patient samples compared to HV samples (Figure 4A).
In the MILE study, ROCK1 was found to be upregulated in all types of aB-ALL (p<0.05), except t(8;14)/t(1;19)/Pre-B-ALL t(9;22) which were not significant (NS). TEAD4 was upregulated in aB-ALL with t(8;14), t(1;19), c-/Pre-B-ALL t(9;22) - p<0.001. There was no consistent difference in YAP1 expression between aB-ALL and HV samples, with variation occurring depending on the probe set used. TCF4 was upregulated in all subtypes of aB-ALL- p<0.001, SOX4 was upregulated in all subtypes of B-ALL - p<0.001 except t(8;14) and SMAD3 was upregulated in all subtypes of aB-ALL- p<0.05.
3.7. TEAD4 and SMAD3 Protein Expression Was Elevated in aB-ALL Samples
ICC was performed on six B-ALL samples compared to HV CD19+ B-cells to examine SMAD3 and TEAD4 expression. TEAD4 and SMAD3 levels were significantly higher in aB-ALL samples compared to HV CD19+ B-cells (p<0.05; Figure 4B). TEAD4 protein was mostly found in the nucleus whereas SMAD3 was found in both the nucleus and cytoplasm. TEAD4 protein levels were significantly different between PB and BM. We found no statistical significance in immunoreactivity scores between TEAD4, SMAD3 and historical BIRC5 data, when we compared the levels of TEAD4 and SMAD3 in this study and BIRC5 levels in the same three patient samples analysed in our previous study [17].
4. Discussion
A number of immunotherapy strategies have been used to improve treatment outcomes for patients with relapsed/refractory (R/R) ALL following chemotherapy and haematopoietic stem cell transplant. However, long-term use of immunotherapy can lead to the selection of leukaemic clones that are resistant to treatment. This evolutionary process hampers the effectiveness of immunotherapy by selecting subclones that are unresponsive to current therapy, resulting in escape variants and relapse. To overcome their existence, the heterogeneity between and within aB-ALL tumour cells, and the limitations with existing immunotherapy treatments, it is essential to broaden the number, specificity and sensitivity of tumour antigen targets available for future clinical use. To this end we immunoscreened a testes cDNA library with sera from five representative aALL sera. We identified 134 sero-positive plaques that encoded 72 independent sequences and three (UOH-ALL-104, UOH-ALL-105 and UOH-ALL-106) were novel providing a source of previously uncharacterised gene products for future analysis.
The majority of SEREX-identified antigens (67/72) had already been shown to be involved in tumour pathogenesis, four were CTAs (TUBA3C/UOH-ALL-3, C10orf82/UOH-ALL-20, CT152/UOH-ALL-83 and PRM1/UOH-ALL-101), three were highly enriched in the testis and involved in spermatogenesis (TEX43/UOH-ALL-43, CCDC89/UOH-ALL-49 and LOC338963/UOH-ALL-85) and three were non-coding RNAs that had testis-restricted expression (LINC00661/UOH-ALL-58, LOC338963/UOH-ALL-85 and LINC00251/ UOH-ALL-98).
In addition we collated CTAs and LAAs from our previous studies [12] and that of others [25,26,28,30,31], determined the pathways most often utilised by the antigens and the core genes in these selected pathways, and then examined the association between levels of expression as determined by the MILE study in aB-ALL samples with survival. We priority ranked the antigens using the system described by Cheever et al [36] that assesses antigens for their utility as vaccine targets, however, this mathematical model has not been updated since 2009, has the disadvantage of giving a high weighting to therapeutic function and may not be precise in reflecting the fairness and reliability of the data [36,47]. However, in the absence of a better criteria, we used the model to identify the five aB-ALL antigens that scored highest for vaccine target potential and the correlation of their expression with patient survival. Levels of SOX4 and YAP1 expression levels were not associated with survival, while TGFβ1, ROCK1 and TEAD4 were. We found that SOX4 and TEAD4 were upregulated in aB-ALL patients samples while lower SOX4 expression showed a trend towards better outcomes and survival (p=0.07). SOX4 belongs to the sex-determining region Y-related HMG box family. Its abnormal expression is associated with cancer, reflecting its role in regulating cell stemness, proliferation and differentiation in healthy cells. SOX4 is known to regulate several cancer-promoting signalling pathways including Wnt, TGFβ and PI3K [48] and the TGFβ, Wnt and hippo pathways were all highly represented by the aB-ALL antigens. SEREX also identified ROCK1/UOH-ALL-65 which has been shown previously to regulate normal haematopoiesis through the negative regulation of erythropoietic stress and inflammation [49]. ROCK1 is an established oncogene in AML, where it’s overexpression is associated with poor survival (p < 0.01) [50]. Of note, using data from the MILE study we identified TEAD4, SMAD3, SOX4 and TCF4 as being upregulated in cB-ALL compared to HVs, while ROCK1 and YAP1 were not. ROCK1 oncogenes may be an attractive target for future treatments as a small molecule inhibitor, GSK269962A, has already been investigated in an AML mouse model [51]. GSK269962A was shown to inhibit tumour growth, apoptosis and clonogenicity, significantly prolonging survival with some mice remaining disease-free for more than 140 days. Knocking out ROCK1 was shown to at least double the survival time in a KIT(D814V)-induced myeloid leukaemia model when compared to controls [52]. The anti-proliferative activity of GSK269962A occurs via the inhibition of cell cycle kinase (CDK6) which induces cell cycle arrest. It also causes apoptosis via increased expression and phosphorylation of p53 as well as decreased expression of antiapoptotic genes including survivin, Bcl-xL and it has been shown to cause the cleavage of PARP in AML cells [51].
TGFβ signalling in haemopoietic and lymphoid cells is complex and context dependent [53]. The TGFβ1 pathway is involved in a variety of cellular functions, including inhibiting cell proliferation, and is often inactivated in lymphoma and acute myeloid leukaemia [54]. Although the MILE study showed that TGFβ1 was downregulated in aB-ALL, in contrast to paediatric B-ALL [55], TGFβ1 acts as a potent immune suppressant that inhibits NK effector function and TGFβ blocking monoclonal antibodies have been shown to reverse ALL-mediated NK cell dysfunction [55]. In contrast, adding exogenous TGFβ1 to healthy donor NK cells induced an inhibitory phenotype similar to ALL-NK cells that has been shown to act through the SMAD signalling pathway [55].
The Wnt/β-catenin pathway regulates cell proliferation, survival and differentiation in hematopoietic stem cells (HSCs), while deregulation of this pathway leads to haematological malignancies including B-ALL [56]. SEREX identified four Wnt pathway components (WLS, Wnt2b, TCF7L2, and DKK3) that were recognised by antibodies in aB-ALL patient sera. Targeting Wnt ligands and receptors can reduce leukaemic stemness [57] while knock-down of FZD6/WNT10B reduced the proliferation of leukaemic stem cells [58]. However, TGFβ and NFAT pathways sustain Wnt activation increasing the leukaemic burden [58]. KIBRA, is a promising tumour suppressor in drosophila and mammalian cells [59] and an upstream regulator of the hippo pathway. It was found to be frequently silenced due to hypermethylation in cB-ALL [60], suggesting one mode by which the Hippo pathway could be targeted.
We also identified eukaryotic initiation factor 2α (eIF2α) through the immunoscreening of a testis cDNA library using ALL sera. The activation of protein kinase RNA-like endoplasmic reticulum kinase (PERK) leads to the phosphorylation of eIF2α. This phosphorylation event promotes the translation of activating transcription factor 4 (ATF4) while reducing overall protein synthesis [61]. ATF4, as a transcription factor, has the ability to increase the expression of CCAAT/enhancer-binding protein homologous protein (CHOP). ATF4 and CHOP together can also upregulate the production of growth arrest and DNA damage-inducible protein-34 (GADD34), which acts to dephosphorylate phospho-eIF2α [61]. Therefore, prolonged endoplasmic reticulum (ER) stress, without the restoration of protein synthesis homeostasis, can ultimately result in apoptotic cell death which may be worthy of further investigation in the context of treating aB-ALL.
Our investigation of genes that can act as possible targets for immunotherapy has identified antigens that are involved in patient survival (ROCK1, TEAD4, SMAD3, and TCF4), in maintaining tumour stem cell features (SOX4, YAP1 and TCF4) as well as the tumour microenvironment (ROCK1, YAP1 and TEAD4). Several of the antigens we identified have been found to play a role in childhood B-ALL (SMAD3 [62]; TCF4 [63]) and demonstrate the broad potential of treatments targeting these proteins/pathways.
The identification of immunogenic epitopes within antigens and their capacity to stimulate the immune response are the usual next step in the development of immunotherapy strategies for patient treatment [64,65]. However, there is a need to break immune tolerance which is exacerbated by the immunosuppressive microenvironment created by tumour cells, through the secretion of IL-10 and TGFβ [66]. The direct targeting of proteins such as TEAD4 and SMAD3 may be especially challenging because they are involved in the maintenance of homeostatic functions and may cause significant off-target effects. Additional investigations using small inhibitors/protein-protein interaction targeting are necessary to determine the clinical utility of direct targeting and the specificity of such treatment for diseased cells [67]. Although BIRC5 is a novel target, phase I trial using antisense oligonucleotide targeting BIRC5 has been examined in children who had relapsed with B-ALL (cB-ALL). However, the trial was discontinued due to high toxicity and off-target effects [19]. One reason for this limited success may be that many immunodominant TAAs are self-proteins. As such, they are subject to both central and peripheral tolerances that dampen immune responses to avoid autoimmunity. This situation demonstrates the need to break self-tolerance in a controlled manner, replicating stem cell transplant strategies that have been used and evolved to effectively treat at least a proportion of patients with haematological malignancies since the 1930s.
5. Conclusions
Current treatments for aB-ALL have involved integrating chemotherapy and targeted therapy. B-ALL is a complex disease with various factors influencing the overall prognosis, including cytogenetic abnormalities, MDR, and response to therapy. Simply targeting the tumour antigens on and in malignant cells is not enough to eradicate tumour growth, as the tumour microenvironment within the BM plays a crucial role in treatment outcomes. Sensitivity to immunotherapy in B-ALL is not solely determined by intrinsic biological factors but also by the diverse interactions between leukaemia cells and the bone marrow microenvironment [68]. Leukaemia cells exploit this microenvironment to sustain their proliferation and survival, taking advantage of the tightly controlled signalling pathways and transcriptional factors that regulate normal lymphopoiesis. Breaking immune tolerance towards leukaemic cells will require a multifaceted approach that minimizes the opportunities for escape mutants to survive the treatment process.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org., Table S1 Antibodies used for ICC; Table S2 Characterisation of cDNA sequences confirmed by two rounds of immunoscreening; Table S3 Expression of SEREX-identified genes and their function in health and cancer tissues; Table S4 Gene expression and association with above and below median expression using MILE data; Figure S1 Protein-protein interaction of LAAs using STRING. Most of the LAAs were involved in binding followed by transcription regulator activity. PMP2, PMP4, CDKN2B, RBX, CUL1 involved in TGFβ enrichment. Wnt2B, CDKN2B, RBX, PMP2, PTGS2 were involved in cancer pathways. The lack of interactions among other genes in this signature is likely explained by a variety of other biological pathways utilised within B-ALL. SEREX identified genes that were involved in ubiquitination included UBC, UBE2C, UBE2D2, CUL1, RBX1 and SKP2. Genes involved in regulating the intrinsic apoptosis process were BCLAF2, PARL, SERINC3, TRAP1, TPT1, MCL1 and PTPN1; Figure S2. In silico analysis of the antigens revealed the levels of expression of each shortlisted antigen in healthy and cancerous tissues. Heatmap showing the expression levels of each protein in (A) healthy cells and (B) different tumour types, with the y-axes represent the proportion of patients expressing each tumour antigen. The purple, red and orange colours indicate very high to low levels of expression, pale yellow represents no expression detected, and white represents no data available in the corresponding histology in Human Protein Atlas.
Author Contributions
Conceptualization, E.M. and B.G.; methodology, E.M. and B.G.; validation, E.M., S.G. and L.C.; formal analysis, E.M., S.G. and L.C.; investigation, E.M. S.G. and L.C.; resources, V.B.B. and K.H.O.; data curation, E.M., D.M.F., K.I.M. and K.H.O.; writing—original draft preparation, E.M., S.G., L.C. and B.G.; writing—review and editing, E.M., and B.G.; visualization, E.M. and B.G.; supervision, B.G.; project administration, B.G.; funding acquisition, B.G. All authors have read and agreed to the published version of the manuscript.
Funding
E.M. and L.C. were funded by University of Hull cluster PhD studentships and V.B. by Breakthrough Cancer Research (previously known as Cork Cancer Research Centre). We would like to thank the patients for their kind donation of samples.
Institutional Review Board Statement
Informed consent and the collection of aB-ALL samples was performed according to the guidelines of the Declaration of Helsinki, and approved by a Local Ethics Committee in the UK as well as the Southampton University Hospitals Trust and University of Hull ethics committees.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Acknowledgments
We would like to thank the patients for their kind donation of samples.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Abbreviations
The following abbreviations are used in this manuscript:
| aB-ALL | Adult B-cell acute lymphoblastic leukaemia |
| BIRC5 | baculoviral IAP repeat containing 5 |
| BMX | bone marrow tyrosine kinase |
| CAR | chimeric antigen receptor |
| cB-ALL | children with B-ALL |
| CTA | cancer testis antigen |
| DEGs | Differentially expressed genes |
| HV | Healthy volunteer |
| LAA | leukaemia associated antigens |
| NS | Not significant |
| PB | Peripheral blood |
| PRKG1 | protein kinase cGMP-dependent 1 |
| R/R | relapsed/refractory |
| ROCK1 | Rho associated coiled-coil containing protein kinase 1 |
| SMAD3 | SMAD family member 3 |
| SOX4 | SRY-Box Transcription Factor 4 |
| TAAs | Tumour associated antigens |
| TBP1 | TATA-box binding protein |
| TCF4 | T cell Factor 4 |
| TEAD4 | TEA Domain Transcription Factor 4 |
| TGFβ | Transforming Growth Factor-β |
| YAP1 | Yes-associated protein1 |
References
- Samra B, Jabbour E, Ravandi F, Kantarjian H, Short NJ. Evolving therapy of adult acute lymphoblastic leukemia: state-of-the-art treatment and future directions. J Hematol Oncol. 2020;13(1):70. [CrossRef]
- DeAngelo DJ, Jabbour E, Advani A. Recent advances in managing acute lymphoblastic leukemia. American Society of Clinical Oncology Educational Book. 2020;40:330-42. [CrossRef]
- Gokbuget N, Dombret H, Giebel S, Bruggemann M, Doubek M, Foa R, et al. Minimal residual disease level predicts outcome in adults with Ph-negative B-precursor acute lymphoblastic leukemia. Hematology. 2019;24(1):337-48. [CrossRef]
- Kantarjian HM, Vandendries E, Advani AS. Inotuzumab Ozogamicin for Acute Lymphoblastic Leukemia. N Engl J Med. 2016;375(21):2100-1. [CrossRef]
- Brown PA, Ji L, Xu X, Devidas M, Hogan LE, Borowitz MJ, et al. Effect of Postreinduction Therapy Consolidation With Blinatumomab vs Chemotherapy on Disease-Free Survival in Children, Adolescents, and Young Adults With First Relapse of B-Cell Acute Lymphoblastic Leukemia: A Randomized Clinical Trial. JAMA. 2021;325(9):833-42. [CrossRef]
- Stackelback AV, Jaschke K, Jousseaume E, Templin C, Jeratsch U, Kosmides D, et al. Tisagenlecleucel vs. historical standard of care in children and young adult patients with relapsed/refractory B-cell precursor acute lymphoblastic leukemia. Leukemia. 2023;37(12):2346-55. [CrossRef]
- Paul S, Kantarjian H, Jabbour EJ, editors. Adult acute lymphoblastic leukemia. Mayo Clinic Proceedings; 2016: Elsevier. [CrossRef]
- Martino M, Alati C, Canale FA, Musuraca G, Martinelli G, Cerchione C. A Review of Clinical Outcomes of CAR T-Cell Therapies for B-Acute Lymphoblastic Leukemia. International Journal of Molecular Sciences. 2021;22(4):2150. [CrossRef]
- Webster J, Luskin MR, Prince GT, DeZern AE, DeAngelo DJ, Levis MJ, et al. Blinatumomab in combination with immune checkpoint inhibitors of PD-1 and CTLA-4 in adult patients with relapsed/refractory (R/R) CD19 positive B-cell acute lymphoblastic leukemia (ALL): preliminary results of a phase I study. Blood. 2018;132:557. [CrossRef]
- Namuduri M, Brentjens RJ. Enhancing CAR T cell efficacy: the next step toward a clinical revolution? Expert review of hematology. 2020;13(5):533-43. [CrossRef]
- Gardner R, Wu D, Cherian S, Fang M, Hanafi L-A, Finney O, et al. Acquisition of a CD19-negative myeloid phenotype allows immune escape of MLL-rearranged B-ALL from CD19 CAR-T-cell therapy. Blood, The Journal of the American Society of Hematology. 2016;127(20):2406-10. [CrossRef]
- Jordaens S, Cooksey L, Freire Boullosa L, Van Tendeloo V, Smits E, Mills KI, et al. New targets for therapy: antigen identification in adults with B-cell acute lymphoblastic leukaemia. Cancer Immunol Immunother. 2020;69(5):867-77. [CrossRef]
- Paavonen K, Ekman N, Wirzenius M, Rajantie I, Poutanen M, Alitalo K. Bmx tyrosine kinase transgene induces skin hyperplasia, inflammatory angiogenesis, and accelerated wound healing. Mol Biol Cell. 2004;15(9):4226-33. [CrossRef]
- Coutre SE, Byrd JC, Hillmen P, Barrientos JC, Barr PM, Devereux S, et al. Long-term safety of single-agent ibrutinib in patients with chronic lymphocytic leukemia in 3 pivotal studies. Blood Adv. 2019;3(12):1799-807. [CrossRef]
- Hillmen P, Eichhorst B, Brown JR, Lamanna N, O'Brien SM, Tam CS, et al. Zanubrutinib Versus Ibrutinib in Relapsed/Refractory Chronic Lymphocytic Leukemia and Small Lymphocytic Lymphoma: Interim Analysis of a Randomized Phase III Trial. J Clin Oncol. 2023;41(5):1035-45. [CrossRef]
- Brown JR, Eichhorst B, Hillmen P, Jurczak W, Kazmierczak M, Lamanna N, et al. Zanubrutinib or Ibrutinib in Relapsed or Refractory Chronic Lymphocytic Leukemia. N Engl J Med. 2023;388(4):319-32. [CrossRef]
- Boullosa LF, Savaliya P, Bonney S, Orchard L, Wickenden H, Lee C, et al. Identification of survivin as a promising target for the immunotherapy of adult B-cell acute lymphoblastic leukemia. Oncotarget. 2018;9(3):3853. [CrossRef]
- Li Y, He W, Gao X, Lu X, Xie F, Um S-W, et al. Cullin7 induces docetaxel resistance by regulating the protein level of the antiapoptotic protein Survivin in lung adenocarcinoma cells. Journal of Thoracic Disease. 2023;15(9). [CrossRef]
- Li F, Aljahdali I, Ling X. Cancer therapeutics using survivin BIRC5 as a target: what can we do after over two decades of study? Journal of Experimental & Clinical Cancer Research. 2019;38(1):368. [CrossRef]
- Valipour B, Abedelahi A, Naderali E, Velaei K, Movassaghpour A, Talebi M, et al. Cord blood stem cell derived CD16+ NK cells eradicated acute lymphoblastic leukemia cells using with anti-CD47 antibody. Life Sciences. 2020;242:117223. [CrossRef]
- Boncheva VB, Linnebacher M, Kdimati S, Draper H, Orchard L, Mills KI, et al. Identification of the Antigens Recognised by Colorectal Cancer Patients Using Sera from Patients Who Exhibit a Crohn's-like Lymphoid Reaction. Biomolecules. 2022;12(8). [CrossRef]
- Liggins AP, Guinn BA, Banham AH. Identification of lymphoma-associated antigens using SEREX. Methods Mol Med. 2005;115:109-28. [CrossRef]
- Sahin U, Tureci O, Schmitt H, Cochlovius B, Johannes T, Schmits R, et al. Human neoplasms elicit multiple specific immune responses in the autologous host. Proc Natl Acad Sci U S A. 1995;92(25):11810-3. [CrossRef]
- Akhmedov M, Martinelli A, Geiger R, Kwee I. Omics Playground: a comprehensive self-service platform for visualization, analytics and exploration of Big Omics Data. NAR Genom Bioinform. 2020;2(1):lqz019.
- Geng H, Brennan S, Milne TA, Chen W-Y, Li Y, Hurtz C, et al. Integrative epigenomic analysis identifies biomarkers and therapeutic targets in adult B-acute lymphoblastic leukemia. Cancer discovery. 2012;2(11):1004-23. [CrossRef]
- Kohlmann A, Kipps TJ, Rassenti LZ, Downing JR, Shurtleff SA, Mills KI, et al. An international standardization programme towards the application of gene expression profiling in routine leukaemia diagnostics: the Microarray Innovations in LEukemia study prephase. Br J Haematol. 2008;142(5):802-7. [CrossRef]
- Haferlach T, Kohlmann A, Wieczorek L, Basso G, Kronnie GT, Béné M-C, et al. Clinical utility of microarray-based gene expression profiling in the diagnosis and subclassification of leukemia: report from the International Microarray Innovations in Leukemia Study Group. Journal of clinical oncology. 2010;28(15):2529-37. [CrossRef]
- Iacobucci I, Mullighan CG. Genetic basis of acute lymphoblastic leukemia. Journal of Clinical Oncology. 2017;35(9):975. [CrossRef]
- Naik A, Lattab B, Qasem H, Decock J. Cancer testis antigens: Emerging therapeutic targets leveraging genomic instability in cancer. Mol Ther Oncol. 2024;32(1):200768. [CrossRef]
- Almeida LG, Sakabe NJ, deOliveira AR, Silva MC, Mundstein AS, Cohen T, et al. CTdatabase: a knowledge-base of high-throughput and curated data on cancer-testis antigens. Nucleic acids research. 2009;37(Database issue):816-9. [CrossRef]
- Mohamed E. Identification of tumour antigens that may facilitate effective cancer detection and treatment: University of Hull; 2023.
- Gislason MH, Demircan GS, Prachar M, Furtwangler B, Schwaller J, Schoof EM, et al. BloodSpot 3.0: a database of gene and protein expression data in normal and malignant haematopoiesis. Nucleic Acids Res. 2024;52(D1):D1138-D42. [CrossRef]
- Kuleshov MV, Jones MR, Rouillard AD, Fernandez NF, Duan Q, Wang Z, et al. Enrichr: a comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 2016;44(W1):W90-7. [CrossRef]
- Chen EY, Tan CM, Kou Y, Duan Q, Wang Z, Meirelles GV, et al. Enrichr: interactive and collaborative HTML5 gene list enrichment analysis tool. BMC Bioinformatics. 2013;14:128. [CrossRef]
- Szklarczyk D, Gable AL, Lyon D, Junge A, Wyder S, Huerta-Cepas J, et al. STRING v11: protein–protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic acids research. 2019;47(D1):D607-D13. [CrossRef]
- Cheever MA, Allison JP, Ferris AS, Finn OJ, Hastings BM, Hecht TT, et al. The prioritization of cancer antigens: a national cancer institute pilot project for the acceleration of translational research. Clinical cancer research : an official journal of the American Association for Cancer Research. 2009;15(17):5323-37. [CrossRef]
- Lossos IS, Czerwinski DK, Wechser MA, Levy R. Optimization of quantitative real-time RT-PCR parameters for the study of lymphoid malignancies. Leukemia. 2003;17(4):789-95. [CrossRef]
- Gomarasca M, Maroni P, Banfi G, Lombardi G. microRNAs in the antitumor immune response and in bone metastasis of breast cancer: from biological mechanisms to therapeutics. International Journal of Molecular Sciences. 2020;21(8):2805. [CrossRef]
- Zhang Y, Chen H-x, Zhou S-y, Wang S-x, Zheng K, Xu D-d, et al. Sp1 and c-Myc modulate drug resistance of leukemia stem cells by regulating survivin expression through the ERK-MSK MAPK signaling pathway. Molecular cancer. 2015;14:1-18. [CrossRef]
- Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods. 2001;25(4):402-8. [CrossRef]
- Khan G, Brooks S, Mills K, Guinn B. Infrequent expression of the cancer–testis antigen, PASD1, in ovarian cancer. Biomark Cancer. 2015;7:31-8. [CrossRef]
- Biesterfeld S, Veuskens U, Schmitz F, Amo-Takyi B, Böcking A. Interobserver reproducibility of immunocytochemical estrogen-and progesterone receptor status assessment in breast cancer. Anticancer research. 1996;16(5A):2497-500.
- Deng Z, Hasegawa M, Aoki K, Matayoshi S, Kiyuna A, Yamashita Y, et al. A comprehensive evaluation of human papillomavirus positive status and p16INK4a overexpression as a prognostic biomarker in head and neck squamous cell carcinoma. International journal of oncology. 2014;45(1):67-76. [CrossRef]
- Mehta GA, Angus SP, Khella CA, Tong K, Khanna P, Dixon SA, et al. SOX4 and SMARCA4 cooperatively regulate PI3k signaling through transcriptional activation of TGFBR2. NPJ breast cancer. 2021;7(1):1-15. [CrossRef]
- Steinhardt AA, Gayyed MF, Klein AP, Dong J, Maitra A, Pan D, et al. Expression of Yes-associated protein in common solid tumors. Human pathology. 2008;39(11):1582-9. [CrossRef]
- Mostufi-Zadeh-Haghighi G, Veratti P, Zodel K, Greve G, Waterhouse M, Zeiser R, et al. Functional Characterization of Transforming Growth Factor-β Signaling in Dasatinib Resistance and Pre-BCR+ Acute Lymphoblastic Leukemia. Cancers. 2023;15(17):4328. [CrossRef]
- Silva WA, Gnjatic S, Ritter E, Chua R, Cohen T, Hsu M, et al. PLAC1, a trophoblast-specific cell surface protein, is expressed in a range of human tumors and elicits spontaneous antibody responses. Cancer Immunity Archive. 2007;7(1).
- Luo X, Ji X, Xie M, Zhang T, Wang Y, Sun M, et al. Advance of SOX transcription factors in hepatocellular carcinoma: From role, tumor immune relevance to targeted therapy. Cancers. 2022;14(5):1165. [CrossRef]
- Mali RS, Kapur S, Kapur R. Role of Rho kinases in abnormal and normal hematopoiesis. Current opinion in hematology. 2014;21(4):271. [CrossRef]
- Liu Y, Gao X, Tian X. High expression of long intergenic non-coding RNA LINC00662 contributes to malignant growth of acute myeloid leukemia cells by upregulating ROCK1 via sponging microRNA-340-5p. European Journal of Pharmacology. 2019;859:172535. [CrossRef]
- Pan T, Wang S, Feng H, Xu J, Zhang M, Yao Y, et al. Preclinical evaluation of the ROCK1 inhibitor, GSK269962A, in acute myeloid leukemia. Front Pharmacol. 2022;13:1064470. [CrossRef]
- Mali RS, Ramdas B, Ma P, Shi J, Munugalavadla V, Sims E, et al. Rho kinase regulates the survival and transformation of cells bearing oncogenic forms of KIT, FLT3, and BCR-ABL. Cancer Cell. 2011;20(3):357-69. [CrossRef]
- Tamayo E, Alvarez P, Merino R. TGFβ superfamily members as regulators of B cell development and function—implications for autoimmunity. International journal of molecular sciences. 2018;19(12):3928. [CrossRef]
- Vicioso Y, Gram H, Beck R, Asthana A, Zhang K, Wong DP, et al. Combination Therapy for Treating Advanced Drug-Resistant Acute Lymphoblastic Leukemia. Cancer Immunology Research. 2019;7(7):1106-19. [CrossRef]
- Rouce RH, Shaim H, Sekine T, Weber G, Ballard B, Ku S, et al. The TGF-β/SMAD pathway is an important mechanism for NK cell immune evasion in childhood B-acute lymphoblastic leukemia. Leukemia. 2016;30(4):800-11. [CrossRef]
- Khoury H, Pinto R, Meng Y, He R, Wu S, Minden MD. Noggin Overexpression Enhances Leukemic Progenitors Self-Renewal in AML by Abrogating the BMP Pathway Activation. Blood. 2006;108(11):2214-. [CrossRef]
- Chiarini F, Paganelli F, Martelli AM, Evangelisti C. The Role Played by Wnt/β-Catenin Signaling Pathway in Acute Lymphoblastic Leukemia. International Journal of Molecular Sciences. 2020;21(3):1098. [CrossRef]
- Cassaro A, Grillo G, Notaro M, Gliozzo J, Esposito I, Reda G, et al. FZD6 triggers Wnt–signalling driven by WNT10BIVS1 expression and highlights new targets in T-cell acute lymphoblastic leukemia. Hematological Oncology. 2021;39(3):364-79. [CrossRef]
- Yu J, Zheng Y, Dong J, Klusza S, Deng W-M, Pan D. Kibra functions as a tumor suppressor protein that regulates Hippo signaling in conjunction with Merlin and Expanded. Developmental cell. 2010;18(2):288-99. [CrossRef]
- Hill VK, Dunwell TL, Catchpoole D, Krex D, Brini AT, Griffiths M, et al. Frequent epigenetic inactivation of KIBRA, an upstream member of the Salvador/Warts/Hippo (SWH) tumor suppressor network, is associated with specific genetic event in B-cell acute lymphocytic leukemia. Epigenetics. 2011;6(3):326-32. [CrossRef]
- Fun XH, Thibault G. Lipid bilayer stress and proteotoxic stress-induced unfolded protein response deploy divergent transcriptional and non-transcriptional programmes. Biochimica et Biophysica Acta (BBA) - Molecular and Cell Biology of Lipids. 2020;1865(1):158449. [CrossRef]
- Liu SX, Xiao HR, Wang GB, Chen XW, Li CG, Mai HR, et al. Preliminary investigation on the abnormal mechanism of CD4(+)FOXP3(+)CD25(high) regulatory T cells in pediatric B-cell acute lymphoblastic leukemia. Exp Ther Med. 2018;16(2):1433-41. [CrossRef]
- Takahashi H, Kajiwara R, Kato M, Hasegawa D, Tomizawa D, Noguchi Y, et al. Treatment outcome of children with acute lymphoblastic leukemia: the Tokyo Children's Cancer Study Group (TCCSG) Study L04-16. Int J Hematol. 2018;108(1):98-108. [CrossRef]
- Caron E, Aebersold R, Banaei-Esfahani A, Chong C, Bassani-Sternberg M. A case for a human immuno-peptidome project consortium. Immunity. 2017;47(2):203-8. [CrossRef]
- Backert L, Kohlbacher O. Immunoinformatics and epitope prediction in the age of genomic medicine. Genome medicine. 2015;7:1-12. [CrossRef]
- Thomas DA, Massagué J. TGF-β directly targets cytotoxic T cell functions during tumor evasion of immune surveillance. Cancer cell. 2005;8(5):369-80. [CrossRef]
- Chen M, Wang J, Yao SF, Zhao Y, Liu L, Li LW, et al. Effect of YAP Inhibition on Human Leukemia HL-60 Cells. Int J Med Sci. 2017;14(9):902-10. [CrossRef]
- Brown G. Lessons to cancer from studies of leukemia and hematopoiesis. Frontiers in cell and developmental biology. 2022;10:993915. [CrossRef]
Figure 1.
Workflow for the identification and analysis of aB-ALL antigens from six sources. Antigens were analysed for their differential expression, expression in gene pathways and association with survival. Antigens were prioritised using the model devised by Cheever et al [55]. From the final antigens analysed for RNA transcription and translation, we found SMAD3 and TEAD4 to be elevated in aB-ALL samples compared to healthy donor samples.
Figure 1.
Workflow for the identification and analysis of aB-ALL antigens from six sources. Antigens were analysed for their differential expression, expression in gene pathways and association with survival. Antigens were prioritised using the model devised by Cheever et al [55]. From the final antigens analysed for RNA transcription and translation, we found SMAD3 and TEAD4 to be elevated in aB-ALL samples compared to healthy donor samples.
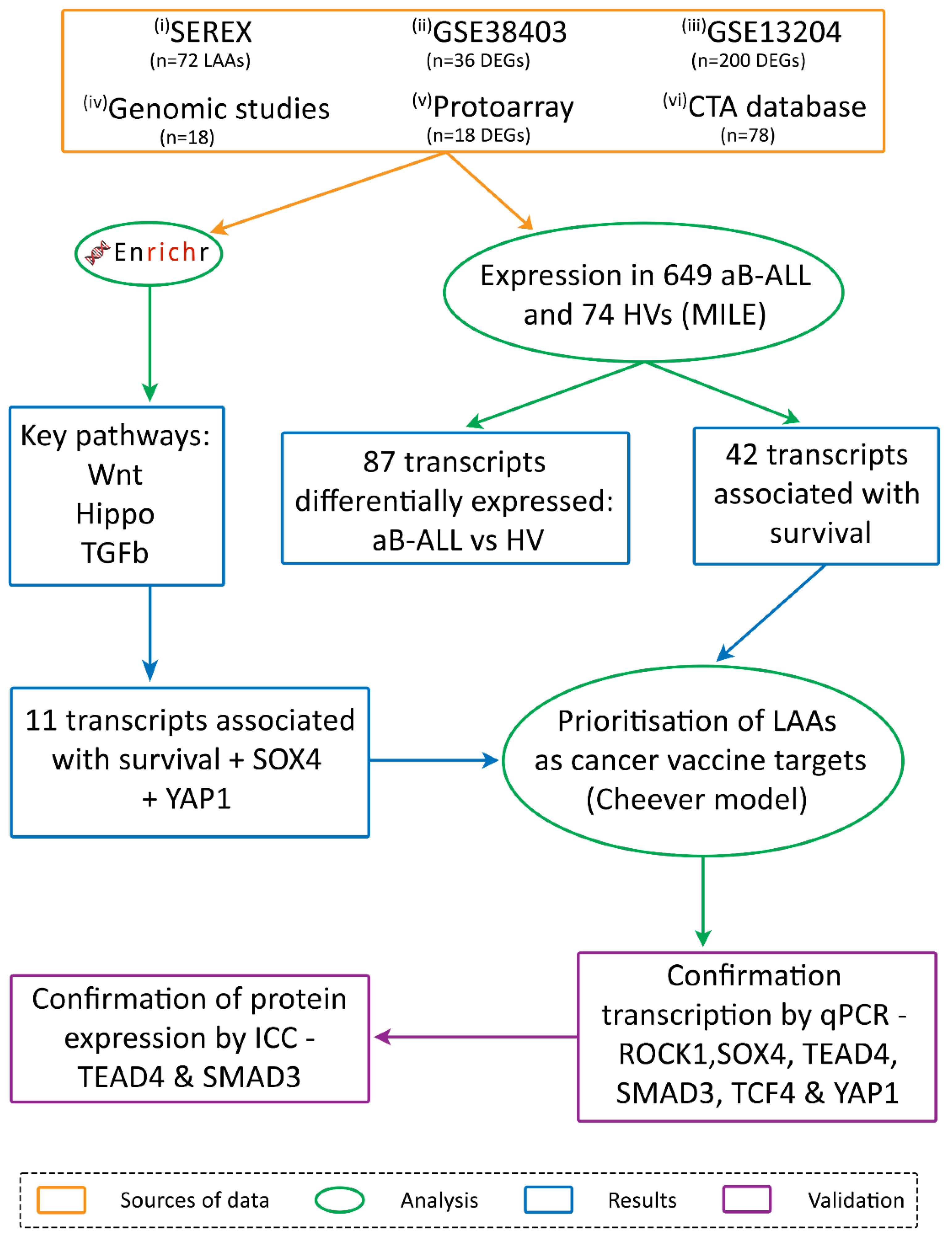
Figure 2.
Relative expression of aB-ALL associated genes from each source. Differentially expressed genes which were upregulated (red) or downregulated (green) are shown as a fold change (FC) and by a key, ranging in FC from +1 to +2.5 and -1 to -2, respectively. The * indicates genes that were associated with survival (p<0.05). Only BMX was common between two studies (protoarray and MILE study).
Figure 2.
Relative expression of aB-ALL associated genes from each source. Differentially expressed genes which were upregulated (red) or downregulated (green) are shown as a fold change (FC) and by a key, ranging in FC from +1 to +2.5 and -1 to -2, respectively. The * indicates genes that were associated with survival (p<0.05). Only BMX was common between two studies (protoarray and MILE study).
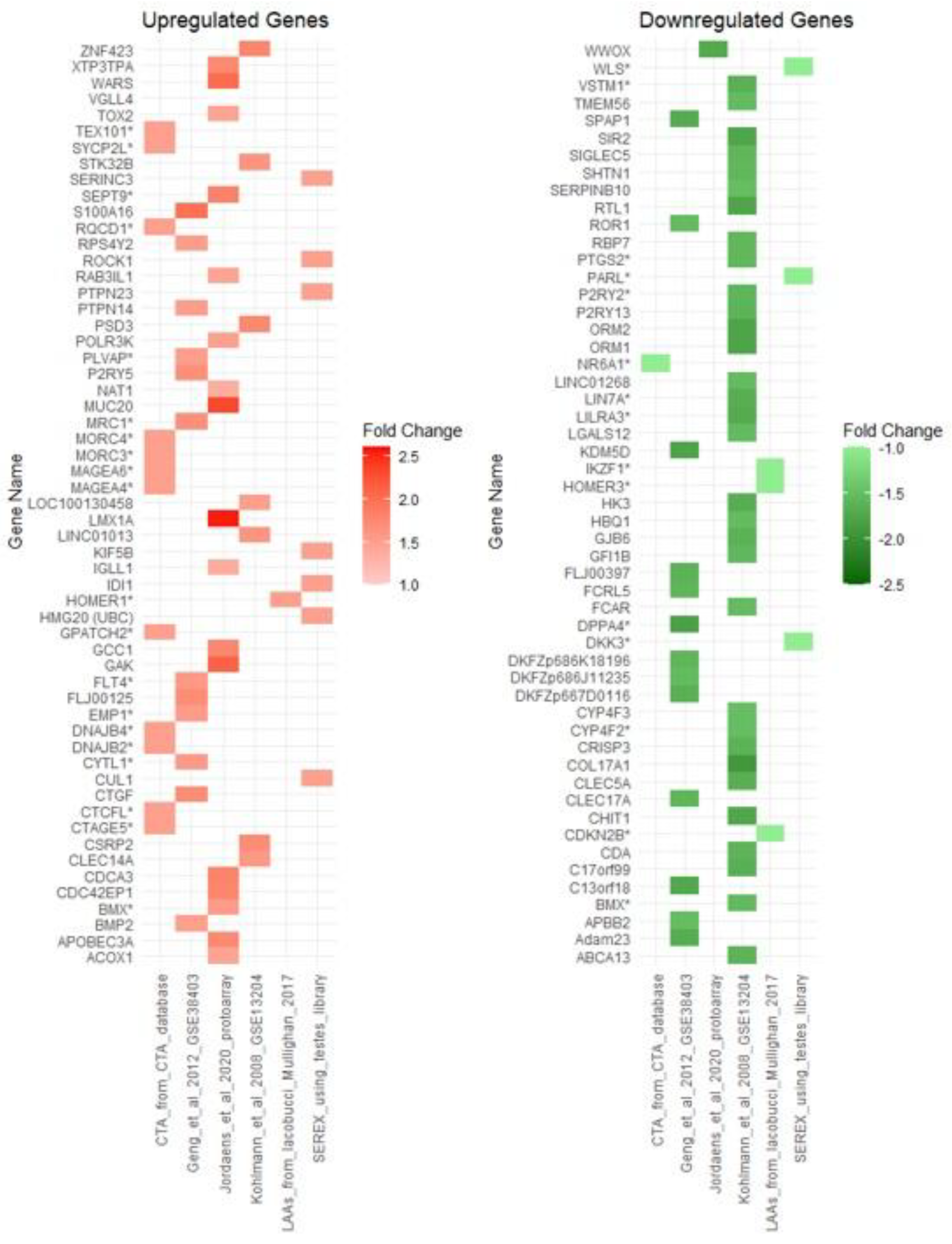
Figure 3.
Role of gene products in key pathways and prioritising of gene products to act as vaccine targets. (A) Three key pathways were found to be enriched within the list of genes identified. These were Hippo, TGFβ and Wnt pathways each involved in promoting leukaemogenesis via the upregulation of anti-apoptotic genes and pro-proliferative genes that causes blasts to continue growing and resist to apoptosis; (B) Antigens that were identified using SEREX, pathway enrichment (PE), literature search (LS), CTA database and microarrays (LAA). Key shows evaluation criteria and the maximum attainable score in parenthesis; Genes indicated in red font were ranked highly within the Cheever system and were selected for further analysis.
Figure 3.
Role of gene products in key pathways and prioritising of gene products to act as vaccine targets. (A) Three key pathways were found to be enriched within the list of genes identified. These were Hippo, TGFβ and Wnt pathways each involved in promoting leukaemogenesis via the upregulation of anti-apoptotic genes and pro-proliferative genes that causes blasts to continue growing and resist to apoptosis; (B) Antigens that were identified using SEREX, pathway enrichment (PE), literature search (LS), CTA database and microarrays (LAA). Key shows evaluation criteria and the maximum attainable score in parenthesis; Genes indicated in red font were ranked highly within the Cheever system and were selected for further analysis.
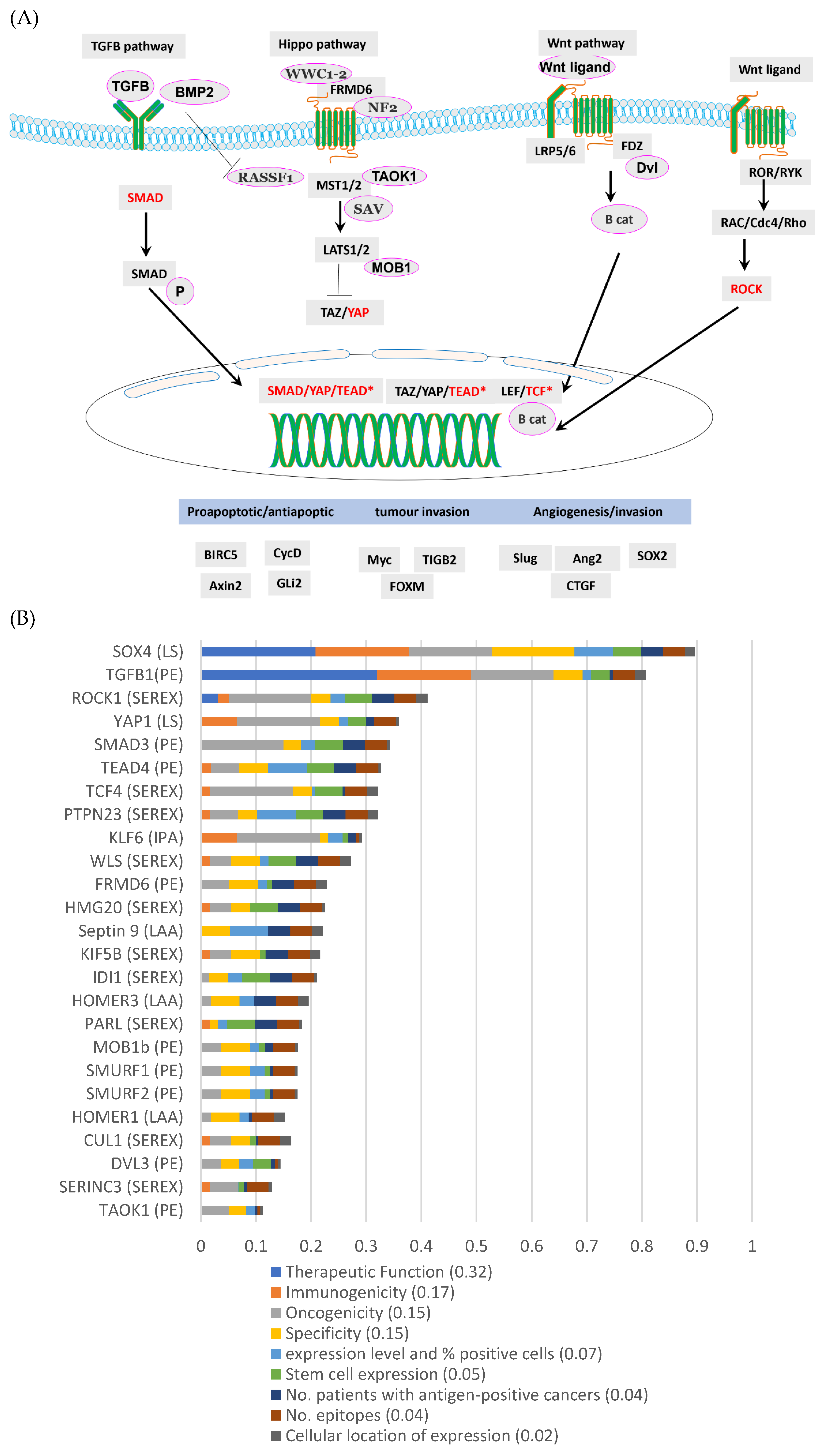
Figure 4.
Expression of the key genes in aALL patient samples. (A) 2-ΔΔCT was used to determine relative gene expression of (i) BIRC5; (ii) ROCK1; (iii) SMAD3; (iv) SOX4; (v) TCF4; (vi) TEAD4 and (vii) YAP1 when compared to the average of two reference genes – TBP and PRKG1. Expression was analysed in 11 PB and five BM samples from ALL patients and five PB samples from healthy donors. (B) Immunolabelling of SMAD3 and TEAD4 in HV CD19+ B-cells and patient samples at X 400 magnification were represented by an immunoreactivity score (0-400; shown in black text on the bottom right corner of each image). Isotype caused little immunolabelling while actin provided a positive control for immunolabelling (stained dark brown). (C)(i) SMAD3 and (ii) TEAD4 immunolabelling were detected in all patient samples with immunoreactivity scores (y-axis) of 200-350, indicating their varied but elevated levels in aB-ALL samples when compared to CD19+ cells from the PB of a HV.
Figure 4.
Expression of the key genes in aALL patient samples. (A) 2-ΔΔCT was used to determine relative gene expression of (i) BIRC5; (ii) ROCK1; (iii) SMAD3; (iv) SOX4; (v) TCF4; (vi) TEAD4 and (vii) YAP1 when compared to the average of two reference genes – TBP and PRKG1. Expression was analysed in 11 PB and five BM samples from ALL patients and five PB samples from healthy donors. (B) Immunolabelling of SMAD3 and TEAD4 in HV CD19+ B-cells and patient samples at X 400 magnification were represented by an immunoreactivity score (0-400; shown in black text on the bottom right corner of each image). Isotype caused little immunolabelling while actin provided a positive control for immunolabelling (stained dark brown). (C)(i) SMAD3 and (ii) TEAD4 immunolabelling were detected in all patient samples with immunoreactivity scores (y-axis) of 200-350, indicating their varied but elevated levels in aB-ALL samples when compared to CD19+ cells from the PB of a HV.
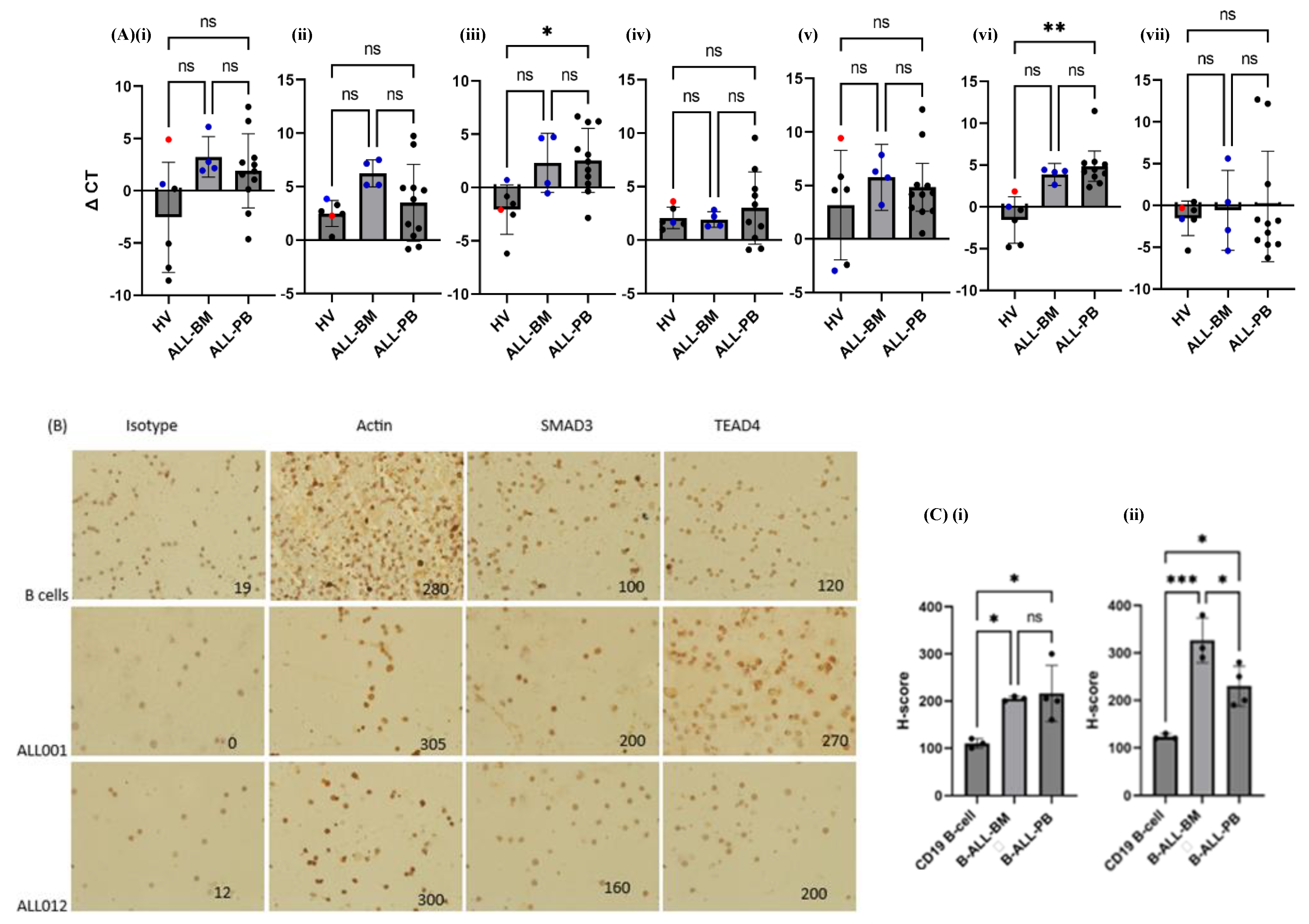

Table 1.
Patient information.
| ID | Disease stage | Cytogenetics | WCC (109/L) |
BM blast % | Relapse | Survival post-sample (mo) | Age≠ | Sex | Sample type |
| ALL001 | Diagnosis Ph+ ALL | Ph+ ALL: t(9;22) | 4.9 | 40 | NK | NK | 39 | M | PB |
| ALL002 | Diagnosis T-ALL | 46,XY,t(1;7)(p36;p15) | 232 | 88 | No | Alive (post allo) | 19 | M | PB |
| ALL003 | Diagnosis B-ALL | t(1;19) | 28.6 | 91 | No | Alive | 26 | F | PB |
| ALL004* | Diagnosis T-ALL | Complex karyotype | 591 | NK | No | Died 19mo (post allo) | 19 | M | PB |
| ALL005* | ¶Post-allo T-ALL | No result | NK | NK | No | Died 3.5mo (post allo) | 46 | M | PB |
| ALL007 | Diagnosis Pre B-ALL | Loss of one copy of ETV6 (12p13) and gain of one copy of ABL1 (9q34) by FISH | 8.2 | 92 | No | Alive | 24 | M | PB |
| ALL008 | Diagnosis Pre-B-ALL | 46XY, 5,del(5)(q15q33),dic(9;16)(p11;q11),del(13)(q12q14) | 10.4 | 72 | No | Alive | 19 | M | PB |
| ALL009 | Diagnosis Pre-B-ALL | 46,XY,t(1;7)(q25;q3?5),add(3)(p1?3) | 48.1 | 89.6 | Yes | Died 94mo (post allo, CART) | 19 | M | BM |
| ALL010* | Diagnosis Pre-B-ALL | Complex including t(4;11) | 229.1 | 85 | Yes | Died 3mo | 64 | M | PB |
| ALL011 | Diagnosis Pre-B-ALL | No result – external referral | NK | NK | NK | NK | 19 | F | PB |
| ALL012 | Diagnosis Pre-B-ALL | t(11;14)(q24;q32) | 4.3 | 83 | No | Alive (post allo) | 33 | M | BM |
| ALL014 | Diagnosis Pre-B-ALL | 47,XY,+2,add(2)(p1)[3]/46,XY[47].nucish (CRLF2)x2[100] | 4.9 | 53 | Yes | Alive (post allo & CART) | 56 | M | BM |
| ALL015 | Diagnosis Pre-B-ALL | Gain of one copy of CRLF2 (Xp22.3/Yp11.3) and loss of one copy of CSFR1 (5q32) and EBF1 (5q33.3) detected by FISH | 68.0 | 94 | No | Alive | 20 | F | PB |
| ALL016 | Diagnosis Pre-B-ALL | Hyperdiploid; 56-57 XX +X,+4,+6,+9,+10,+14,+17,+18,+21,+marker | 1.9 | 96 | No | Alive | 27 | F | BM |
| ALL017† | Diagnosis, Pre-B-ALL | No cytogenetics available, no FISH | 3.0 | 96 | No | Alive | 19 | F | PB |
| ALL018† | Diagnosis, T-ALL | Normal cytogenetics, SET/CAN fusion detected by FISH | 58.8 | 74 | No | Alive | 22 | M | PB |
| ALL020 | Diagnosis Pre-B-ALL | 46,XY, t(1;7)(q25;q3?5), add(3)(p1?3) TCF3 ex16-PBX1 ex3 fusion transcript detected |
31.9 | 92 | Yes | Died 34.5mo (Post allo, relapse, salvage chemo then CART & relapse) | 56 | F | PB/BM |
≠ - age at time of sampling; ¶- status on 30 Oct 2024; Allo: allogeneic stem cell transplant; NK: not known; * - samples used in immunoscreening, qPCR and ICC; †: serum samples used for immunoscreening only.
Table 2.
Healthy volunteer demographics.
| HV control | Age≠ | Sex | Sample type |
| HV008 | 40 | F | PB |
| HV010 | 22 | M | PB |
| HV012 | 46 | F | PB |
| HV021 | 34 | M | PB |
| HV043 | NK | M | PB |
| CD19+ cells | 18-66 | F | PB |
BM: bone marrow; HV: healthy volunteer; NK; not known; PB: peripheral blood.
Table 3.
Pathways identified as involving members of the SEREX identified genes.
| Pathway | p-value | q-value | SEREX-identified genes involved in this pathway | Database used |
| Envelope proteins and their potential roles in EDMD physiopathology | 0.0007 | 0.0892 | TCF7L2, WNT2B | Wiki Pathway |
| Hematopoietic stem cell gene regulation | 0.001 | 0.064 | DNMT1, MCL1 | |
| TGFβ signalling pathway | 0.015 | 0.229 | TRAP1, ROCK1, CUL1 | |
| Mitotic spindle | 0.001 | 0.038 | ROCK1, KIF5B, FLNB, KIF1B, MYH10 | MSigDB Hallmark 2020 |
| Notch signalling | 0.007 | 0.117 | TCF7L2, CUL1 | |
| Apoptosis | 0.026 | 0.286 | ROCK1, TIMP1, MCL1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



