Submitted:
10 February 2025
Posted:
13 February 2025
You are already at the latest version
Abstract
Climate change has a negative impact on crop productivity, yield and product quality. Abiotic stresses such as increased drought, elevated temperatures, flooding, salinity and extreme weather events may severely affect plant development, reproduction, yield and fruit quality. Plants have evolved mechanisms to face stressful conditions and adapt to increased environmental pressures. Intricate molecular processes involving genetic and epigenetic factors and interactions with the sphere of the microbiota have been implicated in the response of plants to abiotic stresses. Deciphering the mechanisms whereby plants perceive and respond to stress and acquire resilience is crucial for developing strategies to counteract current and future climate challenges. The advances in new technologies and methodologies such as next-generation sequencing, multi-omics, GWAS, New Plant Breeding Techniques (NPBTs), have led to the elucidation of various aspects of the complex gene networks and metabolic pathways underlying plant processes as well as plant-microbiota crosstalk involved in the response to abiotic stresses. Although most of this knowledge has arisen from research on model and annual herbaceous plants, recent efforts have begun illuminating the molecular basis of abiotic stress tolerance in woody perennial species. In this review, we will provide a comprehensive overview and discuss the recent findings concerning genetic, epigenetic and microbiome aspects shaping abiotic stress responses, in the context of enhancing tolerance to environmental stressors in important Mediterranean woody fruit crops.
Keywords:
abiotic stress tolerance
; resilience
; drought
; environmental
; climate change
; woody fruit crop
; grapevine
; olive
; date palm
; pomegranate
; microbiome
; microbiota
; epigenetic
1. Introduction
The ongoing climate change poses major challenges to plant development leading to a severe reduction in crop productivity. Abiotic stresses such as drought, elevated temperatures, increased salinity, and extreme weather phenomena, exert major pressures on crop sustainability and threaten global food security [1,2].
In the upcoming decades, Europe, especially the Mediterranean region, will be significantly impacted by climate change, according to the estimates of the Intergovernmental Panel on Climate Change-IPCC Sixth Assessment Report [3]. Depending on greenhouse gas emissions, rainfall is predicted to decrease by 4–22%, whereas the intensity and frequency of extreme weather events such as drought spells and heat waves as well as erratic precipitation patterns and storm surges in certain areas, are anticipated to increase. These alarming forecasts call for concerted actions by agriculturists and researchers to mitigate the severe effects of unfavorable environmental conditions on crop production and agricultural sustainability.
Developing ways to safeguard crop and food systems relies greatly on illuminating the cellular processes and molecular mechanisms underlying abiotic stress perception, stress response, and acquisition of tolerance by crop species [2,4]. The response of crops to adverse environmental conditions involves the intricate relations of gene networks and gene expression programmes induced by various abiotic stressors. Furthermore, epigenetic factors, such as DNA methylation, histone modifications, and the action of noncoding RNAs, play crucial roles in regulating the molecular mechanisms associated with the abiotic stress response and establishment of tolerance [5,6,7] (Figure 1). Moreover, multiple and ever-increasing studies have highlighted the vital role of the microbiome (soil, rhizosphere, phyllosphere, carposphere) on plant development and environmental stress resilience [8,9,10].
The implementation of molecular tools such as genomics, epigenomics, transcriptomics, proteomics, metabolomics (-omics), genome-wide association studies (GWAS) and genome-editing approaches (e.g. CRISPR/Cas) have greatly facilitated the study of abiotic stress responses and tolerance in plants [11,12,13,14]. Moreover, the tools above have been implemented in a variety of genetic resources including indigenous material, wild species, and commercial varieties with diverse characteristics to elucidate stress-related gene networks, metabolic pathways, and cellular processes associated with specific resilient phenotypes. Ultimately, this knowledge could lead to superior genotypes with improved abiotic stress tolerance, through genomic selection and modern breeding approaches [5,15,16,17].
Another aspect of high importance in the quest for plant resilience is the action of the microbial communities associated with the plant entity. Plant-associated microbiota could be either pathogenic or beneficial. Beneficial microbiota comprise several classes of microorganisms, including epiphytes, endophytes and arbuscular mycorrhiza. It is well-recognized that these microorganisms have the capacity to exert beneficial effects on plants by promoting growth, increasing water and nutrient uptake, and enhancing resilience to environmental stresses [8,18,19]. Numerous studies have shown that both the host plant and beneficial microorganisms are able to confer abiotic stress tolerance through a wide range of physiological and molecular mechanisms. These include a) osmotic adjustment through the accumulation of osmoprotectant molecules (e.g. proline, betaine, trehalose), b) modulation of critical phytohormones such as auxin and ethylene which reinforces plant growth under unfavorable conditions, c) scavenging of damaging reactive oxidative species (ROS) through the induction of oxidative enzymes and d) regulating the expression of stress-related genes [11,19,20,21].
Considering the re-emerging notion of the plant as an ‘holobiont’, which refers to the assemblage of the host plant along with its associated microbiota, our view of the plant’s responses has expanded to include the microbiome of the epiphytic and endophytic microbiota (e.g. bacteria, fungi, viruses) associated with the plant’s organs and adjacent soil [22,23,24]. A wealth of investigations has explored the genetic and environmental drivers of microbiota diversity, structure of microbial communities, and host-microbiota associations, both belowground and aboveground (soil, rhizosphere, phyllosphere, carposphere) as well as the interactions within these complex systems [9,10,18,19,20,25,26,27,28,29] (Figure 2).
The majority of the studies described above have focused on annual herbaceous plants including important crops such as legumes, tomato, and cereals, whereas in the last several years research efforts have begun to unravel the molecular dynamics of abiotic stress response in woody perennial species. In the current review, we present a comprehensive survey of the progress achieved regarding the genetic, epigenetic, and microbial facets of abiotic stress responsiveness and tolerance in key woody fruit crops of the Mediterranean basin. Furthermore, we discuss the significance of these multidimensional aspects in successfully designing innovative strategies for mitigating the negative impact of global climate change on woody fruit crop sustainability.
The ongoing climate change increases the intensity and frequency of abiotic stressors such as drought, temperature, and fluctuating patterns of rainfalls, storms and cold spells, especially in the Mediterannean area, a ‘hot spot’ of climate risk. This has negative impacts on the sustainable production of essential crops of the region and on key woody fruit crops such as grapevine, olive, date palm and pomegranate. Stress induces complex processes involving the host genome and epigenome and the associated microbiome activating stress-resonsive mechanisms that may lead to crop tolerance to abiotic stress and adaptation to environmental changes.
2. Grapevine
2.1. Climate Challenges
Grapevine (Vitis vinifera L.) is a woody perennial plant belonging to the genus Vitis which harbors 60–80 species (Vitis spp.). Grapevine domestication dates back to 11000-8000 B.C. and it has been estimated that the species Vitis vinifera encompasses 6000-10000 cultivars with very high inter- and intra-varietal genetic diversity [30,31,32]. Nevertheless, only a portion of this wide genetic pool is exploited for commercial use [15,32,33].
Cultivated grapevine constitutes a crop of high economic value, intimately linked to the history and cultural heritage of viticultural regions around the world [32]. Global vineyard surface area expands to approximately 7.2 million hectares (mha) with annual grape production of ~77.8 million tons of which 47.4%, 44.5%, and 8% correspond to wine, table, and dried grape, respectively [34]. It has been well acknowledged that mild drought stress is beneficial for winegrape varieties as it promotes higher sugar content and lower acidity as well as the synthesis of metabolites (polyphenols, tannins, volatile organic compounds-VOCs) determining colour, aroma, and flavor, all leading to improved wine quality [35]. However, in recent years, grapevine cultivation has witnessed severe environmental pressures, characterized by prolonged drought periods, escalating temperatures and enhanced rainfalls, that pose major threats both for wine and table grape productivity and the sustainability of vitiviniculture [32,36]. Moreover, increased salinization of agricultural lands in grape-growing regions reduces grapevine production and raises major concerns for the future of viticulture.
In 2023, global wine production declined by 10% as compared to 2022, whereas a further decline is anticipated for 2024 [34]. Extreme weather phenomena including early frosts, protracted drought as well as heavy rainfall leading to the exacerbation of fungal diseases, dramatically impacted vineyard harvests worldwide. To address adverse environmental changes and dire climate forecasts, viticulturists, producers, the wine industry and researchers have adopted various strategies such as a) improving vineyard management and implementing effective cultivation practices adapted to current challenges [36,37,38] b) capitalizing on grapevine’s great genetic diversity and explore the wide allele pool of Vitis spp. to develop superior genotypes with enhanced stress resilience[15,17,33]; c) Exploit the growing knowledge on the grapevine microbiome, especially the capacity of rhizosphere microbiota to confer stress tolerance, an approach gaining significant attention lately [19,39,40]. An important aspect across all strategies is the effective use of rootstocks to optimize scion performance. The ancient practice of grafting scions onto suitable rootstock significantly improves plant qualities as it imparts the plant with advantageous adaptive traits. Historically, European grapevine has been grafted onto interspecific North American hybrid rootstocks to counteract the catastrophic infestations of phylloxera (Daktulosphaira vitifoliae) [41,42]. Additionally, cultivated grapevine varieties are grafted onto compatible rootstocks to ensure productivity, fruit quality and resistance to disease [33,42,43]. Certain rootstocks have the potential to provide abiotic stress tolerance, whereas successful graft performance relies on the compatibility of rootstock and scion genotypes [33,42,43,44,45]. Consequently, proper management of rootstocks represents a sound strategy to augment the resilience of grapevines to unfavorable environmental conditions. Nevertheless, the genetic basis of grapevine rootstocks is quite narrow, and improved existing or novel varieties have become necessary. Equally, appropriate and improved scion varieties are sought to satisfy enhanced resilience requirements. Thus, another effective approach to mitigate the negative effects of abiotic stresses is implementing technology to develop abiotic stress-tolerant grapevine cultivars. In this regard, delineating genetic and epigenetic mechanisms underlying abiotic stress responsiveness and rootstock-scion crosstalk will be crucial for crop improvement of both grafting partners [45]. Similarly, comprehending microbiome structure/function and harnessing the potential of the grapevine microbial communities to confer tolerance to abiotic stresses that damage grapevine productivity, could lead to alternative protecting approaches [46]. The latter may be based on the development of effective bioinoculants and ultimately serve in synergy with the methods mentioned above to counteract climate risks, enhance fitness and adaptability, and safeguard sustainable viticulture [33,47].
2.2. Genetic and Epigenetic Attributes on Abiotic Stress
Elucidating the molecular mechanisms of tolerance to abiotic stresses and implementing this knowledge to crop improvement, relies intensely on studying genetic and epigenetic components shaping grapevine performance (Table 1).
The high genetic variability and rich allelic reservoir characterizing Vitis spp. constitute valuable resources for understanding stress-response mechanisms and designing improvement strategies toward enhanced tolerance to abiotic stresses [15,33]. A plethora of genetic studies have identified SSR and SNP markers related to specific grapevine varieties and important agronomic characteristics, thereby providing molecular means that could substantially aid the selection of desired genotypes in breeding programmes [48,49,50,51,52,53,54,55,56].
Moreover, the advancement in DNA sequencing technologies has led to the release of improved versions of grapevine reference genome assemblies [57] as well as a non-reference pangenome of wild grapevine accessions [58]. These genomic resources serve as a valuable tool to understand the molecular basis of morphological, physiological, and biochemical characteristics related to yield, fruit quality and stress resilience. Importantly, the recent construction of a grapevine pangenome reference map, encompassing resequencing data from 466 grapevine cultivars and including a remarkable range of short variants as well as structural variants (SV), is anticipated to decipher complex polygenic trait associations and contribute to genomic breeding [59,60].
Recently, Genome-Wide Association Studies (GWAS) have been conducted to dissect the genetic basis of abiotic stress-tolerance-related traits in grapevine [15,33]. GWAS was used to explore stomatal conductance during water deficit stress utilizing 100 grapevine distinct genotypes which included interspecific hybrids and various rootstock varieties. Genomic regions encompassing 24 significant SNP marker-trait associations with relevance to drought tolerance were detected and thirteen candidate genes possibly responsive to water deficit were identified [49]. Notably, one of these candidate genes encoded a raffinose synthase, indicating a role for this enzyme in the early response to drought stress and agreeing with the known protective function of raffinose oligosaccharides against abiotic stressors [61]. Moreover, whole genome re-sequencing of 77 rootstock genotypes (including North American and Asian Vitis species and their hybrids) and GWAS analysis revealed six sets of 631, 13, 9, 2, 810, and 44 SNPs, with substantial relevance to resistance/tolerance to phylloxera, root-knot nematodes, salt, drought, cold, and waterlogging, respectively [62].
Transcriptomic profiling based on genome-wide RNA sequencing has highlighted important aspects of gene expression programmes and molecular mechanisms associated with abiotic stress responsiveness. A study of drought-tolerant and drought-susceptible grapevine rootstocks revealed differential expression of genes encoding transcription factors, VvAGL15, VvBD41, and VvMYB86 and a range of proteins implicated in antioxidant pathways suggesting their putative roles in the regulation of the molecular processes underlying drought response and tolerance [63]. Investigation of the effects of combined drought and heat stress at the physiological and molecular level revealed the co-expression of gene networks encompassing signal transduction cascades, phenyl propanoid metabolism, sugar metabolizing enzymes, heat-shock protein and transcription factor regulation, and histone modification factors. Notably, most of the differentially expressed genes were unique to the individual or combined stress types, and only a few were shared among the two, pointing to the operation of distinct mechanisms [64]. Moreover, transcriptomic analysis of grapevine genotypes with variable responses to heat stress, showed differential regulation of genes encoding transcription factor families of the type HSP, WRKYs, MYBs and NACs, or regulators involved in auxin and ABA signaling and starch and sucrose metabolic networks, highlighting the importance of these key pathways in the response to high-temperature stress [65].
Concerning epigenetic regulation and transcriptional reprogramming during grapevine development and stress response, several investigations have indicated altered cytosine methylation patterns, differential histone modifications and the action of diverse classes of non-coding RNAs (miRNAs and siRNAs), in the response to drought, temperature, salt and UV-B radiation stresses [38,45,66,67]. More specifically, Pagliarani et al. reported that water deficit conditions induced significant downregulation of the well-conserved microRNA 159, miR159, in a drought-tolerant hybrid (M4) but not in a drought-sensitive variety (Cabernet Sauvignon) in both greenhouse and field conditions [68]. In addition, transcript abundance of conserved microRNA156, miR156, was reduced upon drought stress in both genotypes. Conversely, a microRNAome study demonstrated up-regulation of miRNAs miR159c, miR156b and miR156f under drought conditions, in the susceptible genotype (Cabernet Sauvignon) but not in the tolerant genotype (110 Richter rootstock) [69]. Recent investigations on the response to drought of indigenous grapevine varieties from traditional viticultural regions identified substantial variability in drought tolerance among distinct genotypes, accompanied by differential miRNA regulation [70]. miRNA159 displayed a marked induction in the drought-tolerant variety ‘Dichali’, both in self-rooted plants and those grafted on 110R rootstock, but not in the drought-sensitive variety ‘Debina’. On the other hand, miR156, showed a significant induction only in severely stressed self-rooted plants of the drought-tolerant ‘Dichali’ [70]. These findings suggest that miRNA159 and miR156 expression patterns are associated with grapevine drought responsiveness and tolerance in a genotype- and grafting-dependent manner. Furthermore, miRNA up- or down regulation was linked to anticorrelated expression patterns of putative target genes, MYB1 and TRP, encoding key transcription factors involved in development and stress response processes[68,70]. These results suggested the involvement of ‘miRNA-Transcription factor’ regulatory modules characterizing the drought response and are in line with previous studies in grapevine and other plant species [71,72,73]. Furthermore, other investigations have shown differential expression of large sets of miRNAs under salinity or cold stress and indicated miRNA subgroups as regulators of downstream key genes associated with stress responsiveness [74,75]. Studies on DNA methylation and histone remodeling have begun to emerge. Treatment of Cabernet Sauvignon seedlings with 5-azacytidine and subsequent decrease in global cytosine methylation led to an effective reduction of aluminum toxicity presumably by up-regulating stress-responsive genes [63,76]. Additionally, histone modification profiling of V. amurensis leaves demonstrated massive genome-wide H3K27 trimethylation modifications upon chilling stress, associated with various pathways including stress-responsive gene networks [77].
Collectively, the findings described above underscore the important role of genetic and epigenetic regulation in abiotic stress responses and unveil the multiple factors that may shape stress tolerance (genotype, grafting, developmental stage, type of stress). A deeper comprehension of the molecular mechanisms underlying key responses to abiotic stressors, engaging both scion and rootstock, will expand our knowledge of grapevine stress tolerance. Ultimately, research outcomes will be translated into practical application by developing abiotic stress-tolerant varieties in breeding programmes. Similarly, it will enable further advancements in crop improvement biotechnology related to genome editing approaches (e.g. CRISPR-Cas) [78] paving the way to climate-resilient viticulture.

Table 1.
Abiotic stress responses associated with genomics and epigenomics.
| Species | Stress type | Molecular tools | Molecular response/ tolerance-associated genes | References |
|---|---|---|---|---|
| Grapevine | Drought | Genome-wide identification studies (GWAS) | Candidate genes and SNPs associated with stomatal conductance and drought responsiveness e.g raffinose synthase | [49,62] |
| Transcriptomics-RNA Seq/ Quantitative PCR |
co-expression of gene networks related to signal transduction cascades, phenyl propanoid metabolism, sugar metabolizing enzymes, heat-shock protein transcription factor regulation, and histone modification factor TF families-VvAGL15, VvLBD41, and VvMYB86 Up- and down regulation of responsive miRNAs-VvmiR159, VvmiR156 Induction of miRNAs VvmiR159, VvmiR156 and anticorrelated expression TF genes, MYB1 and TPR Drought-induced VvmiR169d and VvmiR156b upregulation and VvmiR398a downregulation Activation of the module: miR156b-VvSBP8/13 |
[64] [63] [68] [70] [69] [72] |
||
| Heat | Transcriptomics-RNA seq/ Quantitative PCR | Transcription factor families -WRKYs, MYBs and NACs, Auxin and ABA signaling, Starch and sucrose metabolism Induction of heat stress-responsive miRNAs-VvmiR167 |
[65] [55] |
|
| Aluminum (Al) toxicity | Whole Genome Bisulfite Sequencing (WGBS) | DNA methylation reduction / enhanced tolerance to Al | [76] | |
| Cold | Chromatin immunoprecipitation (ChIP) Transcriptomics-RNA seq |
H3K27 trimethylation alterations / gene target downregulation Novel cold stress-responsive microRNAs |
[77] [75] |
|
|
Olive tree |
Drought | Transcriptomics/RNA-seq | Transmembrane transport and metal ion binding processes, abscisic acid, gibberellin, brassinosteroids, and ethylene-activated signaling |
[79] |
| Salt | Transcriptomics/RNA-seq | TF families, JERF and bZIP Up regulation of OeNHX7, OeP5CS, OeRD19A and OePetD |
[80] [81] |
|
| Date Palm | Combined heat and drought |
Proteomics | Increased abundance of Heat Shock Proteins (HSP), redox homeostasis proteins and proteins involved in isoprene production | [82] |
|
Salt |
Multi-omics |
Converging gene expression and protein abundance associated with osmotic adjustment, reactive oxygen species scavenging in leaves, and remodeling of the ribosome-associated proteome in salt-exposed root cells. Induction of Salt Overly Sensitive (SOS) genes, PdSOS2;1, PdSOS2;2, PdSOS4, PdSOS5, and PdCIPK11 |
[83] [84] |
|
|
Whole Genome Bisulfite Sequencing (WGBS) |
Differential DNA methylation and gene expression alterations in roots |
[85] |
||
| Pomegranate | Salt | Transcriptomics/RNA-seq | Spatiotemporal regulation of SWEET genes DEGs associated with ABA- and Ca2+-related and MAPK signal transduction pathways (ABA-receptors, Ca2+-sensors, MAPK cascades, TFs) and downstream functional genes coding for HSPs, LEAs, AQPs and PODs. Induction of proline, total soluble sugar, and SOD/POD activities and differential gene expression |
[86] [87] [88] |
|
Cold |
Transcriptomics/RNA-seq |
Upregulation of CBFs genes PgCBF3, PgCBF7 Differentially expressed genes related to TFs, photosynthesis, osmotic regulation system, and hormone signal transduction, sucrose metabolism Induction of beta-amylase, PgBAM4, and increase in soluble sugar content |
[89] [90] [91] |
2.3. Microbiota Attributes Related to Abiotic Stress
Concerning the microbiome aspect as it pertains to abiotic stress, several studies have shown that grapevine microbiota are capable of conferring resilience to abiotic stress factors such as drought, elevated temperature and high salinity by activating plant growth mechanisms, increasing photosynthetic capacity, synthesizing phytohormones (e.g. IAA, ABA), accumulating osmoprotectants (e.g. proline, trehalose) and ROS scavenging antioxidant molecules, inducing stress-responsive genes, and triggering other cell protecting mechanisms [39,40] (Table 2).
Earlier laboratory and greenhouse studies identified plant growth promoting (PGP) bacterial strains capable of colonizing the grapevine rhizoplane and supporting plant growth under water deficit conditions by increasing shoot/leaf biomass and shoot length, inducing photosynthetic activity and accumulating hormones as well as producing defense-related products such as terpenes [92,93]. Importantly, bacterial subsets originating from grapevine roots and rhizosphere were found to exert beneficial effects on field-grown grapevines, in various biogeographical settings [94].
In addition, mycorrhizal and endophytic fungi have also been shown to alleviate drought, temperature and salt stress through a variety of mechanisms such as modulating ABA and auxin metabolism and inducing antioxidant activity [39,95]. Recently, arbuscular mycorrhizal fungi (AMF) inoculation of a Vitis vinifera L. cv. Ecolly was demonstrated to confer drought tolerance by increasing the accumulation of osmolytes, triggering antioxidant processes and regulating the expression of key stress-responsive genes, VvNCED, VvP5CS, VvSIP, VvPIP1;2, and VvTIP2;1 [96]. Moreover, field experiments conducted in periods of rising temperatures and reduced water availability showed that grapevine rootstocks inoculated with AMF displayed enhanced growth and survival compared to non-AMF-associated plants [97]. In another study, growth-promoting rhizobacteria consortia from the marine plant significantly improved the resilience of grapevine to heat stress, highlighting the potential of microbial consortia to promote fitness and protect grapevine from frequent heatwave exposure [98,99]
Notably, global biogeography surveys have linked microbiomes with entire viticultural regions. Large-scale studies of vineyard rhizosphere microbiomes across continents revealed remarkable associations among microbiomes, terroir, and wine characteristics [100,101]. Microbiome ‘’signatures’’ could be ascribed to distinct vineyards within and between continents and countries, highlighting the microbiome’s potential as a fingerprinting tool to trace the geographical origin of grapevine germplasm and ensure the authenticity of end products. Considering that terroir-associated microbiomes are shaped by rootstock/scion genotype, soil and climate factors [46,102,103] it is conceivable that microbiota consortia from hot and dry areas may confer tolerance to grapevines from more temperate zones currently undergoing temperature and drought pressures [19,46]
Finally, concerning the mechanistic complexities of plant-microbe interactions, a major challenge will be to decode the intricate molecular interplay between the genetic, epigenetic, and microbial components driving the response to abiotic stress. In this respect, little progress has been made, and involves a few annual herbaceous plants like Arabidopsis, chickpea and herbs [28]. For example, it was shown that Pseudomonas putida (a plant growth-promoting rhizobacteria-PGPR), known to improve performance under abiotic stress, mediates the regulation of specific chickpea (Cicer arietinum L.) miRNAs and their targets, in response to drought and salt stress [104]. Interestingly, the Pseudomonas putida-induced microRNA, car-miR166, was found to play a stress-mitigating role in PGPR-inoculated Arabidopsis transgenic lines under drought conditions [99].
Furthermore, Pokeweed (Phytolacca americana L.), a North American native perennial herb with great potential for bioremediation of heavy metal-contaminated soils, displayed altered DNA methylation in roots upon inoculation with PGPR. Importantly, the DNA methylation modification was associated with enhanced plant growth and persisted even after removal of the inoculum [105].
Recently, investigations have emerged aiming to explore the molecular dynamics of the ‘grapevine–microbiota–stress’ interactions. Campos et al. studied V. vinifera cv. Touriga Nacional grafted onto 1103 Paulsen rootstock, inoculated with AMF and subjected to high-temperature stress. AMF inoculation of grapevine roots enhanced physiological indices under temperature stress. Notably, it also up- or down-regulated specific miRNAs that target genes encoding stress-related transcription factors or proteins involved in antioxidant pathways, suggesting that mycorrhiza-mediated miRNA regulatory networks act in response to heat stress [106]. Furthermore, the variation in miRNA expression was observed in leaf tissue, pointing to molecular signaling control between rootstock and scion by mycorrhization and stress factors. Interestingly, in another study, a comparative transcriptome analysis of ten grapevine rootstocks revealed the induction of common as well as unique sets of genes after mycorrhiza inoculation, indicating genotype-specific gene expression upon mycorrhization [107] that may have stress-response relevance. These findings set the foundation for further investigations of the complex molecular interplay between the grapevine plant and associated microbiota under abiotic stress conditions.
A deeper understanding of the response to abiotic stress, the role of genetic and epigenetic factors involved, and the grapevine-microbe interplay, will illuminate the molecular processes engaged in abiotic stress responsiveness and tolerance, and lead to innovative approaches for climate-smart viticulture.
Table 2.
Abiotic stress responses associated with microbiomes.
| Species | Stress type | Microbe type | Microbial effect – molecular response | References |
|---|---|---|---|---|
| Grapevine | Drought |
Rhizosphere associated bacteria | Protection against Reactive Oxygen Species (ROS) – accumulation of terpenes | [93] |
| Drought | Root associated microbiome | Water stress-protection |
[92] |
|
| Drought | Arbuscular mycorrhizal fungi (AMF) | Drought tolerance by increasing the accumulation of osmolytes, triggering antioxidant processes and regulating the expression of key stress-responsive genes | [96] |
|
| Heat |
Marine Plant Growth-Promoting Rhizobacteria Consortia | Heat stress tolerance |
[98] | |
| Heat | AMF |
Enhancement of physiological indices Modulation of miRNAs and stress-related transcription factors and proteins related to antioxidant pathways |
[106] |
|
| Olive tree | Drought |
Pseudomonas reactans Ph3R3 | Enhancement of plant performance by reducing water loss, improving N levels, net CO2 assimilation rate, and antioxidant capacity. | [108] |
| Drought | PGPB consortia sampled from soil and rhizosphere of Tunisian olive orchards | Conferred tolerance to both drought-susceptible and drought-tolerant cultivars |
[109] |
|
| Drought | AMF (Rhizophagus irregularis) |
Reinforced tolerance to water deficit by enhancing olive plant growth, improving water status, accumulation of osmolytes and antioxidants and phytohormone regulation | [110] |
|
| Drought | AMF (Rhizophagus irregularis) | Enhanced water deficit tolerance by increasing net carbon fixation, water use efficiency and antioxidant defenses | [111] |
|
| Salt | PGPB Bacillus G7 |
Improved physiological and metabolic parameters, increased photosynthetic capacity, net carbon fixation, water use efficiency, and accumulation of osmolytes and antioxidant | [106] 131 |
|
| Salt | AMF mixtrure of Glomus deserticola and Gigaspora margarita | Alleviation of the stress imposed by irrigation with salt-enriched wastewater. | [112] | |
| Date Palm | Drought |
Selected date plam root bacterial endophytes | Increased the biomass of date palms exposed to recurrent drought stress cycles in greenhouse experiment |
[113] |
| Salt |
Piriformospora indica endophyte |
Mitigated the detrimental effects of salt stress through ion homeostasis and nutrient uptake, antioxidant activity, and upregulation of stress-responsive genes. | [114,115] |
|
| Salt |
Enterobacter cloacae SQU-2 (SQU-2)’ | Improved the growth of Arabidopsis thaliana Columbia (Col-0) seedlings under both normal and salt stress conditions through production of microbial volatile compounds mVOCs. | [116] | |
| Pomegranate | Drought | AMF strains Rhizophagus intraradices (GA5 and GC2) | Early inoculation with AMF, especially for the GC2 strain, offers protection against drought. Enhanced antioxidant defenses, specifically the ROS-scavenging enzymes superoxide dismutase (SOD), catalase (CAT), and ascorbate peroxidase (APX), in shoots. | [117] |
3. Olive Tree
3.1. Climate Challenges
Olive belongs to the botanical family Oleaceae, harboring about 30 genera and 600 species. The Olea L. genus comprises 33 species with Olea europaea L. being the only cultivated species with more than 2000 cultivars [118]. The olive tree is thought to have been domesticated since 7000 B.C in the Mediterranean region [119]. Cultivated olive (Olea europaea subsp. europaea var. europaea) is a woody fruit crop of major economic and social importance. Total olive cultivation area amounts to approximately 10.4 million hectares (mha), worldwide. In the Mediterranean region alone, olive cultivation occupies 9 mha accounting for 80% of the global table and 98% of global olive oil production [120].
Well-adapted to the semiarid climate of the Mediterranean, olive has been historically planted in low-density systems in rainfed environments, nevertheless, the increasing demand for olive products has led to intensive planting under irrigation systems [121]. In recent years, olive productivity has been threatened by the ever-growing frequency and intensity of abiotic stressors experienced in the Mediterranean basin [122]. In this respect, olive yields were severely affected by drought and extreme weather events in the south of EU, throughout 2022 and the spring and summer of 2023. As a result, the production of olive oil dropped to its lowest level since 1994–1995, and market prices escalated (https://worldpopulationreview.com). Model climate forecasts indicate that intensified warming and drought are anticipated in many parts of the planet in the coming decades. According to these projections, heat and drought conditions will deteriorate significantly in areas already facing climate risks. In this context, olive trees will encounter severe climate pressures due to increased temperatures and altered precipitation patterns anticipated in the Mediterranean region [81,123]. In addition, environmental changes are expected to exacerbate susceptibility to disease and have severe effects on olive production [123]. Recently, olive-grove-management systems have focused on incorporating sustainable agroecological practices including effective soil management, composting, and reutilization of waste products, to cope with the negative consequences of climate pressures [124]. In addition, exploring olive genetic variability, genetic/epigenetic mechanisms and olive-microbiota interactions related to abiotic stress responsiveness are alternative approaches of great promise and discussed below.
3.2. The Genetic and Epigenetic Component
Multiple studies have attempted to investigate the genetic/molecular basis of abiotic stress responses employing molecular markers and a suite of multi-omic tools across a wide range of olive genotypes [125]. SSR and SNP markers have been used extensively to discriminate among different varieties and link distinct genotypes to crucial agronomic traits [126]. Recently the most complete genomic variation map based on 89 olive tree genotypes was reported [127] providing a valuable tool for illuminating the genetic diversity among varieties as well as the molecular basis of fruit quality and stress response traits.
Transcriptomic studies using a salt-tolerant variety (Kalamon) and a salt-sensitive variety (Chondrolia Chalkidikis) subjected to salt stress demonstrated differential gene expression between the two genotypes and suggested transcription factors of the JERF and bZIP families as putative modulators of salt-responsive gene regulatory networks [80]. Similarly, comparative morpho-physiological and transcriptomic studies under high-salt conditions revealed marked upregulation of the genes OeNHX7, OeP5CS, OeRD19A and OePetD, in a salt-tolerant variety (Royal de Cazorla) as compared to medium-tolerance and susceptible varieties, indicating their key role in salt-tolerance mechanisms[128].
A meta-analysis of twenty-six RNA-Seq samples from Olea europaea and other fruit tree species identified sets of genes commonly regulated in drought stress conditions most likely involving transmembrane transport and metal ion binding or differentially regulated sets related to abscisic acid, gibberellin, brassinosteroids, and ethylene-activated signaling [79]. Noteworthy, an Olive Atlas including 70-RNA-seq experiments spanning developmental processes and the response to a range of abiotic and biotic stresses, has been reported recently [129], constituting a valuable tool for analyzing gene networks and metabolic pathways implicated in stress response and tolerance.
Little has been reported on epigenetic factors associated with abiotic stress mechanisms. Differential DNA methylation patterns were observed under high salinity conditions among varieties with variable salt tolerance. Further analysis of the differentially methylated regions identified a set of underlying genes encoding OePIP1.1 (an aquaporin), OePetD (a cytochrome b6), OePI4Kg4 (a phosphatidylinositol 4-kinase) and OeXylA (a xylose isomerase), suggesting their implication and regulation by epigenetic mechanisms in the response to salt stress [81].
Overall, unraveling the molecular mechanism of abiotic stress responsiveness at the genetic and epigenetic level is crucial for comprehending the regulatory networks and cellular signaling pathways linked to abiotic stress tolerance in olive and will have significant ramifications for climate-resilient oliviculture.
3.3. The Microbiota Component
A series of studies have explored the effects of microbiota such as Plant Growth-Promoting Bacteria (PGPB) and Plant Growth-Promoting Fungi (PGPF) on olive tolerance to a variety of abiotic stressors in greenhouse and field conditions [130] (Table 2).
PGPB used in these studies were sourced from soil or rhizosphere of olive or other species as well as from PGPB collections. For example, Dias et al. explored the effects of the bacterium Pseudomonas reactans Ph3R3 in potted Olea europaea L. plants cv. Arbequina subjected to water deficiency conditions. It was demonstrated that Ph3R3 treatment enhanced plant performance by reducing water loss, improving N levels, net CO2 assimilation rate, and antioxidant capacity [108]. Similarly, inoculation of cv. Arbequina with PGPB Bacillus G7 protected plants from high salt stress. Bacillus G7 treatment improved physiological and metabolic parameters by increasing photosynthetic capacity, net carbon fixation, water use efficiency, and accumulation of osmolytes and antioxidant molecules [131]. Transcriptional reprogramming was also evidenced involving induction of ABA pathway- and ion homeostasis-related genes. In another study, PGPB consortia sampled from soil and rhizosphere of Tunisian olive orchards were used to inoculate drought-susceptible and drought-tolerant potted olive plantlets that were subsequently exposed to water deficit stress. The PGPB consortia were able to confer drought tolerance to olive plants from both cultivars [109]. Following a different approach, Sallami et al. determined microbial profiles from olive rhizosphere samplings across arid and semi-arid regions of Morocco and assessed plant growth bacterial performance under various conditions [132]. A large proportion of isolates displayed marked tolerance to high salinity conditions highlighting their potential as candidates for bioinoculant formulations.
Concerning the effect of beneficial fungi, several studies have shown the positive impact of AMF on olive trees under abiotic stress exposure. Inoculation of potted olive plants with AMF (Rhizophagus irregularis) reinforced tolerance to water deficit. This was attributed to enhanced olive plant growth, improved water status, increased accumulation of osmolytes and antioxidants and modulation of phytohormones [110,111].
Furthermore, certain AMF mixtures confer tolerance to high salt stress conditions. Notably, inoculating young potted olive plants cv. Chetoui (salt-sensitive) with a mixture of Glomus deserticola and Gigaspora margarita alleviated the stress imposed by irrigation with salt-enriched wastewater. This has important implications for certain Mediterranean agricultural regions where the reuse of treated wastewater, often rich in salts, constitutes common practice [112].
Thus far, information regarding the molecular mechanisms and potential implication of epigenetic factors in the olive tree-associated microbiome interactions is obscure. On the other hand, noteworthy research has been conducted characterizing olive microbiomes and determining genotype and environmental drivers that may shape microbial community composition and diversity [133,134,135,136]. Further investigations and a deeper understanding of the molecular pathways governing the interplay between olive trees and microbiota during abiotic stress will help dissect stress tolerance from the perspective of the holobiont and contribute to alternative strategies for protecting oliviculture against climate challenges.
4. Other Woody Fruit Crops
4.1. Date Palm
Date palm (Phoenix dactylifera L.) is a socioeconomically significant crop throughout the Middle East and North Africa contributing to food security in semiarid and arid regions, where it remains the main woody plant. The date palm can grow in a wide range of climates, from Australia to Asia, Africa, and the Americas. Since it is acclimated to a wide range of temperatures (12.7 to 27.5°C on average), it can tolerate both frost and very high temperatures of -5 and +50 °C [137] together with prolonged water shortage [138]. Although P. dactylifera can withstand drought and salt stress, intense water scarcity and ever-rising soil salinity have jeopardized the crop's productivity. Specifically, the date palm suffers from high soil salinity despite being a relatively salt-tolerant plant. Consequently, P. dactylifera is a useful model for analyzing the physiological and molecular mechanisms that enable plants to withstand harsh weather conditions[139]. Studies over the last several years on date palm have begun to elucidate the physiological mechanisms of abiotic stress tolerance and the genes and biochemical pathways that control the response to these stresses.
4.1.1. Genetic/Epigenetic Factors in Abiotic Stress
Recent reviews have reported on the genetic and genomic advancements made in date palm for facilitating targeted crop improvement strategies, such as abiotic tolerance in harsh environments [140,141,142]. Several of these studies have focused on transcriptomic analyses to unravel the genes and mechanisms that contribute to the development of salinity tolerance [141,143]. Using transcriptome and metabolomic profiling, Safronov et al. (2017)[144] investigated the adaptation methods of date palms under mild heat, drought, and combined heat and drought. In all three situations (drought, heat, and combined heat and drought), transcriptome data revealed transcriptional activation of genes linked to reactive oxygen species, indicating enhanced activity of enzymatic antioxidant systems in the cytosol, chloroplast, and peroxisome. Of note, there was a considerable enrichment for circadian and diurnal rhythm motifs in the genes that were differentially expressed under heat and combined heat and drought conditions, indicating new stress avoidance methods. For instance, since certain genes encoding heat shock factors (HSP) harbor circadian control motifs in promoter regions, it is conceivable that the synthesis of HSP is synchronized with the time of the day as required by the circadian clock. Another transcriptome study identified genes implicated in detoxifying cadmium (Cd) toxicity [145]. Numerous defense and detoxification-related genes, including those encoding heavy metal (HM)-chelators and HM-transporters, were successfully predicted in response to Cd stress. This provided a strong basis for the study of the molecular regulation mechanisms of heavy metal accumulation and tolerance in date palm leaves and roots. The primary and secondary metabolic profiles, along with memory effects on water relations, appear to prime date palm foliar characteristics for recurrent summer drought occurrences, according to a study of the metabolic profile of date palm seedlings subjected to a drought-recovery regime in both summer and winter simulated climates [146]. Date palms that have a well-coordinated metabolic network—which include the anti-oxidative system, the accumulation of appropriate solutes, osmotic adjustment, and cell-membrane stability—seem to be less vulnerable to drought. These drought-compensating strategies may be required more frequently during the summer. Xu et al. demonstrated the implication of salt overly sensitive (SOS) pathway genes, including PdSOS2;1, PdSOS2;2, PdSOS4, PdSOS5, and PdCIPK11, in the salt response [84]. Gene expression was in line with changes in physiological parameters such as amino acid profile and Na+/K+ homeostasis, which in turn inhibited plant growth. The "spliceosome" pathway was interestingly enriched in the upregulation category, suggesting that alternative splicing (AS) may play a role in the date palm's response to salt stress.
Climate chamber experiments and proteomics analysis provided a complete view of how the date palm leaf proteome may adapt to Saudi Arabia's natural environment [82]. It appears that the date palm has evolved a complex multi-mechanism based on increasing abundances of Heat Shock Proteins involved in abiotic stress defense as well as redox homeostasis proteins and proteins involved in isoprene production to counteract the stress imposed by summer temperature conditions and the soil aridity of the Arabian Peninsula.
Mueller et al. [83] used integrative multi-omics studies, followed by focused metabolomics, hormone, and ion investigations, after exposing date palm to a salt stress dose equal to seawater for up to four weeks. A strikingly high degree of convergence between gene expression and protein abundance was discovered when proteomic data were integrated with transcriptome data. This clarifies the mechanisms of acclimatization that are used, which rely on reprogramming protein production. Date palm successfully combines several salt-tolerance mechanisms present in both halophytes and glycophytes for growth in highly saline environments: "acclimation" through osmotic adjustment, reactive oxygen species scavenging in leaves, and remodeling of the ribosome-associated proteome in salt-exposed root cells, and "avoidance" through effective sodium and chloride exclusion at the roots.
Further attempts to identify and functionally characterize salt and drought tolerance-related genes in date palm have been made by several research groups [116,143,147]. Due to the lengthy regeneration time and restricted research implementation, genetic engineering techniques have not yet been effectively applied in date palm breeding, despite their immense value [148].
The role of epigenetic mechanisms in date palm gene regulation under abiotic stresses has not yet been thoroughly studied, despite its significance. Comparative miRNA profiling under salt stress showed that differential expression of miRNAs in leaf and root tissue whereas most miRNA sequences were upregulated in both tissue types by salinity treatment [149]. Al-Harrasi et al. investigated the DNA methylation status and transcriptome profile in date palm under salt stress. Whole genome bisulfite sequence (WGBS) revealed differential DNA methylation in roots of date palm plants upon salinity treatment [85]. Gene expression was impacted by 5mC methylation alterations evidenced in different gene regions, however, DNA methylation was not the only factor affecting global transcript abundance. These findings emphasize the significance of DNA methylation in stress-induced epigenetic remodeling and may be useful for crop breeding efforts.
4.1.2. Microbiota Aspects and Abiotic Stress
Microbial communities in oasis habitats enable plants to withstand harsh environmental conditions, which is where Phoenix dactylifera flourishes [150]. Even though endophytic bacteria promote plant growth in the face of abiotic stress [92], little is known about P. dactylifera endophytic bacteria and how they contribute to the development of abiotic tolerance, including drought and salt tolerance.
Date palm roots choose a variety of endophytic communities that can support plant growth in drought-prone environments, according to a study by Cherif et al. on the ecology of date palm root endophytes from oasis desert farms in southern Tunisia [113]. In another study, the morphological and growth characteristics of date palm seedlings were negatively impacted by prolonged exposure to salt stress. Since the rhizosphere is the primary source of endophytes, changes in rhizosphere populations are likely to directly affect the makeup of the endophytic community. Yaish et al. 2016 identified endophytic bacterial and fungal communities in P. dactylifera grown under salt stress and showed that the composition of those microbial communities changed significantly in response to changes in salinity [151].
Bacterial strains isolated from the rhizosphere of date palms cultivated in an orchard with high soil salinity were examined for their ability to promote plant growth through the generation of microbial volatile compounds (mVOC) [116]. The strain ‘Enterobacter cloacae SQU-2 (SQU-2)’ was found to produce mVOCs, which improved the growth of Arabidopsis thaliana Columbia (Col-0) seedlings under both normal and salt stress conditions. Inspection of the rhizobacterium's genome using next-generation sequencing techniques demonstrated the existence of mVOC gene clusters suggesting the function of mVOC synthesis pathways in growth enhancement of date palm plants confronted with high salinity stress. These findings encourage additional research on the mechanisms underlying the plant-microbe interaction that promotes development under unfavorable conditions and its potential use in agriculture. Sabeem et al. 2022, reported that root colonization of date palm seedlings with the beneficial endophyte Piriformospora indica significantly reduced the detrimental effects of salt stress through enhanced growth through ion homeostasis and nutrient uptake, antioxidant activity, and modulation of stress-responsive genes [114]. Ensuing studies of root transcriptomes demonstrated that P. indica colonization resulted in up-regulation of multiple genes involved in metabolic and signaling pathways relevant to salt tolerance [115]. Date palm root colonization by P. indica is a remarkable example of a beneficial microbial symbiosis. Presently, the underlying molecular mechanisms by which it establishes itself and exerts its beneficial effects are poorly understood and it is anticipated to be the focus of rigorous future research.
4.2. Pomegranate
Pomegranate (Punica granatum L.), an ancient perennial species indigenous to Central Asia, is commercially cultivated across over 30 countries, including India, Iran, Spain, China, and the United States. With a history of cultivation exceeding 2000 years, pomegranate is an important subtropical fruit crop that can be used for its fruits, leaves and other parts. It has been widely cultivated in arid and semi-arid regions that have been faced with salinity[152] and many other hazardous environmental conditions. Like other fruit trees, pomegranate also suffers from drought and salinity affecting its transplantation survival rate, fruit yield and quality [152,153].
On the other hand, Pomegranate trees thrive in warm climates and have a low tolerance for cold temperatures, leading to their predominant cultivation in tropical and subtropical regions. Cold stress limits plant growth, development and yields, specifically cold snaps during winter and late spring are prone to inducing freezing damage in pomegranate trees, which can significantly impair fruit yield and quality, consequently diminishing market availability [154].
4.2.1. Genetic Aspects and Transcriptional Regulation
Gene expression studies have revealed numerous gene sets involved in a variety of metabolic, antioxidant, and signaling pathways during pomegranate exposure to abiotic stress factors.
In-plant kingdom, the Sugar Will Eventually Be Exported Transporters (SWEET) gene family is widely dispersed and is involved in various biochemical and physical processes [155]. Earlier studies have demonstrated the role of SWEET genes under many abiotic stresses (such as salt, hot or cold stress) by regulating the redistribution of soluble sugar, resulting in the maintenance of osmotic balance and growth [156,157]. In a study by Kumawat et al. 2022, 15 SWEET genes were identified in the pomegranate genome, and their chromosome distribution, exon/intron structure, phylogeny, conserved motif, and expression features were characterized [86]. RNA-seq data analysis and qPCR evaluation demonstrated tissue-specific SWEET transcript accumulation and differential expression under high salinity stress. These findings will help to understand the spatio-temporal regulation of sugar transport in pomegranate under abiotic stress conditions.
Several studies have investigated the genetic basis of salt stress responses to unveil the molecular and physiological strategies pomegranate plants employ to adapt and survive in high-salt environments [87]. Transcriptomic analysis of pomegranate roots and leaves under salinity stress, demonstrated massive induction of differential gene expression, including 1080 up-regulated and 1175 down-regulated genes [87]. Most DEGs showed tissue- and temporal-dependent transcript accumulation. Among root DEGs, genes associated with cell wall organization and transmembrane transport were suppressed, and most metabolism-related genes were over-represented. On the other hand, in leaves, ion transport and oxidation-reduction processes were restricted. Many DEGs were associated with ABA- and Ca2+-related and MAPK signal transduction pathways (ABA-receptors, Ca2+-sensors, MAPK cascades, TFs) and downstream functional genes coding for HSPs, LEAs, AQPs and PODs.
Similarly, Tang et al. reported significant changes in physiological parameters and gene expression, during a time course experiment at different salt concentrations, in pomegranate seedlings. Induction of proline, total soluble sugar, and superoxide dismutase and peroxidase (SOD/POD) activities were accompanied by differential gene expression, 72 hours after salinity stress [88]. 6571 DEGs were identified across different treatments including 374 transcription factors. Six special modules and 180 hub genes were obtained by WGCNA (the weighted gene co-expression network analysis) which is used to find modules and networks of highly related genes. Functional annotation highlighted primary and secondary metabolism and signaling cascades as significant pathways.
Furthermore, several studies have explored the effect of abscisic acid (ABA) a common phytohormone known to play key roles in regulating plant growth, development, and environmental adaptation [158,159]. Morpho-physiological analysis and transcriptome profiling in pomegranate under drought stress demonstrated that exogenous ABA application substantially enhanced drought resistance by reinforcing metabolic pathways, such as brassinosteroid synthesis, peroxisome biogenesis, photosynthesis and hemicelluloses synthesis [153].
Although grown in warm environments pomegranate may face cold spells in winter and early spring with catastrophic consequences. Recently, the molecular basis of pomegranate response and tolerance to cold stress has been explored. Genes encoding C-repeat binding factors (CBFs) are known to have an important function in plant cold resistance. Two CBFs genes (PgCBF3 and PgCBF7) were identified from a cold-tolerant pomegranate cultivar and their overexpression was found to enhance cold tolerance in transgenic Arabidopsis plants [89]. Moreover, PgICE1 induced promoter activity via MYC-recognition site-binding leading to increased PgCBF3 and PgCBF7 transcript abundance suggesting putative regulation of downstream cold-responsive (COR) genes by PgCBF3 and PgCBF7. These findings provided insights into the signaling cascades that may be implicated in cold response and highlighted potential gene targets for breeding aiming at cold-tolerant genotypes. Furthermore, Guan et al. (2023) reported on the physiological changes and transcriptome profiles of Tunisian soft-seed pomegranate exposed to cold (6 °C) and freezing (0°C) stress. Differentially expressed genes under cold stress were related to TFs, photosynthesis, osmotic regulation system, and hormone signal transduction. Potential cold response/resistance genes involved in plant hormone signal transduction, photosynthetic systems, carbon fixation in the C4 pathway, and sucrose and galactose metabolism were also identified laying a foundation for the selection of major candidate genes for molecular breeding [90].
Other studies have investigated the involvement of Beta-amylases (BAMs), a protein family known to have an important role in breaking down starch into soluble sugars contributing significantly to sugar accumulation during cold stress. These soluble sugars are then translocated from the chloroplasts to the cytoplasm, where they participate in energy metabolism, strengthening the plant’s defense against cold-induced damage [91,160]. Liu et al. examined eight PgBAM genes from the pomegranate genome dataset. Transcriptome analysis of the cold-sensitive cultivar ‘Tunisia’ subjected to cold stress for 0 and 12 hours highlighted the starch and sucrose metabolism pathway as a critical component in cold stress response. In particular, PgBAM4 displayed marked induction under cold conditions. Notably, variations in soluble sugar content and PgBAM4 expression correlated with differences in cold stress resistance between cv. ‘Tunisia’ (cold-sensitive) and cv. ‘Sanbai’ (cold-tolerant) seedlings. One-hybrid assays indicated that PgCBF7, a key transcription factor functioning in freezing tolerance, binds to the promoter region of PgBAM4. These findings offer a comprehensive understanding of the PgBAM gene family function and reveal new insights into the regulatory mechanisms behind cold stress tolerance in pomegranate [156].
4.2.2. Microbiome and Abiotic Stress
Few studies have been reported thus far concerning pomegranate microbiomes and plant-microbe interactions under abiotic stress conditions. Bompadre et al. investigated the impact of inoculating two strains of arbuscular mycorrhizal fungi (AMF), Rhizophagus intraradices (N.C. Schenck & G.S. Smith) C. Walker & A. Schüßler (GA5 and GC2), on pomegranate plants grown under two different irrigation regimes [117]. The plants' response to oxidative stress varied depending on the type of tissue and the severity of stress. Their results indicated that mycorrhizal plants enhanced their antioxidant defenses—specifically the ROS-scavenging enzymes superoxide dismutase (SOD), catalase (CAT), and ascorbate peroxidase (APX)—in shoots across both irrigation treatments, while the root response was inconsistent. AMF inoculation helped regulate malondialdehyde (MDA) levels, possibly by swiftly boosting antioxidant defenses and preventing lipid peroxidation. The study concluded that early inoculation with AMF, especially for the GC2 strain, offers protection to pomegranate plants against abiotic stress during propagation. Characterizing microbiota communities' diversity and structure in different conditions has begun to be reported. For instance, Ravinath et al. (2022) [161] utilized a 16S rRNA amplicon-based metagenomics to identify the dominant and abundant bacterial species in the rhizosphere of the pomegranate Bhagwa variety at various soil depths. Considering that beneficial microorganisms in the rhizosphere play a functional role in plant protection and resilience to abiotic factors, future research efforts are expected to focus on the pomegranate microbiome and pomegranate-microbe interactions.
5. Conclusions and Future Perspectives
Climate change and the exponential growth of the human population raise major concerns for crop production, sustainable agriculture and food security. Abiotic stress pressures such as increased drought, elevated temperatures, extreme salinity, and unpredictable rainfall patterns, negatively affect plant development, yield, and quality.
Plants have evolved complex mechanisms to counteract stress and adapt to changing environments.
A deep comprehension of the sophisticated cellular processes that render plants tolerant to stress is vital for adopting appropriate strategies to mitigate the impacts of adverse environmental conditions. Molecular mechanisms operating at the genetic and epigenetic level, engaging multiple gene networks and metabolic pathways as well as the intricate interactions between the plant and its microbial communities, have been shown to play crucial roles in abiotic stress responsiveness and tolerance. Most of the studies involved in understanding the molecular basis of abiotic stress tolerance have been performed on annual herbaceous species including important crops such tomato, legumes, and cereals. In recent years, molecular mechanisms underlying abiotic stress responses have begun to be characterized in key woody fruit crops of high economic value, cultivated in the Mediterranean region, such as grapevine, olive, date palm and pomegranate.
Lately, the Mediterranean basin has experienced enhanced climatic pressures including rising drought, high temperatures and salinity whereas climate forecasts predict increased water scarcity and warming conditions for the decades ahead. These current and future climate risks jeopardize the viability of key woody perennial fruit crop cultivations. In this context, further research is needed to address environmental threats and develop ways to impart resilience while sustaining good yields and fruit quality. Insights of abiotic stress responses at the genetic and epigenetic level and the host plant-microbiota interaction will greatly contribute to a deeper understanding of stress tolerance molecular processes. Subsequently, this will facilitate the development of biomarkers to discriminate among cultivars with variable susceptibility and link genotypes to stress-relevant traits. The knowledge acquired will allow for selecting suitable genotypes in breeding programs or for precise genome editing during NPBT applications, ultimately leading to stress-tolerant varieties. In recent years, genomic technologies and the employment of molecular tools have led to advances in unlocking plant genetic diversity, developing improved tolerant varieties, and harnessing the potential of the microbiome to confer stress resilience in several woody perennial fruit crops. Further research should focus on a more detailed investigation of these domains, including more woody fruit species. In addition, future efforts should be directed at epigenomic remodeling and plant-microbiome interactions while conceptualizing the plant as a holobiont entity. In this context, exciting new ideas have been proposed regarding the holobiont as a breeding target and the microbiome as a trait [162].
Expanding our knowledge of the genetic, epigenetic and microbial factors governing abiotic stress responsiveness and tolerance will enable optimal use of NPBTs. Genome editing methods like CRISPR/Cas offer exciting new opportunities for genetic/epigenetic chromatin modification, targeting genomic regions with abiotic stress relevance [163]. CRISPR/Cas can expedite crop improvement as it circumvents the long and cumbersome cycles of the breeding process, especially in woody perennial species. Moreover, rapid advancements in the CRISPR/Cas field concerning transgene-free systems make this biotechnology particularly attractive. Nevertheless, low regeneration capacity of genome-edited plants and regulatory framework limitations (regarding cultivation, market release, and consumer use approval) hinder the effective implementation of genome-editing technology [164]. In this context, more progress is needed towards efficient transformation and regeneration systems across different genotypes as well as easing regulations and harmonizing the legal framework among countries.
Finally, characterizing woody crop microbiomes under different conditions, determining genotype and environmental drivers of microbial community structure/function, and exploring the potential of beneficial microbiota to confer tolerance is of major importance. Elucidating the role of host plant-microbiota interaction in stress protection will allow embracing new perspectives in crop improvement such as the emerging concept of microbiota-assisted breeding. In addition, it could result in effective bio-inoculations with suitable consortia cocktails, enabling crops to alleviate the damaging effects of stress and withstand environmental pressures in the field. Overall, the research endeavors and outcomes mentioned above will lead to effective synergistic strategies to tackle climate pressures, enhance fitness and adaptability, and safeguard sustainable woody crop cultivation under unfavorable environmental conditions.
Further insights into the intertwined genetic, epigenetic, and microbial mechanisms underlying abiotic stress tolerance will unravel the complex ‘crop-microbiome-stress’ network and ultimately enhance the resilience of key woody perennial fruit crops in an era of water scarcity and global warming.
Author Contributions
Conceptualization, A.K.; writing—original draft preparation, A.K, E.T., V.P.; writing—review and editing, A.K., E.T., V.P., S.L., M.G., E.A., P.B.; supervision, A.K.; All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Jägermeyr, J.; Müller, C.; Ruane, A.C.; Elliott, J.; Balkovic, J.; Castillo, O.; Faye, B.; Foster, I.; Folberth, C.; Franke, J.A.; et al. Climate Impacts on Global Agriculture Emerge Earlier in New Generation of Climate and Crop Models. Nat Food 2021, 2, 873–885. [Google Scholar] [CrossRef] [PubMed]
- Eckardt, N.A.; Ainsworth, E.A.; Bahuguna, R.N.; Broadley, M.R.; Busch, W.; Carpita, N.C.; Castrillo, G.; Chory, J.; DeHaan, L.R.; Duarte, C.M.; et al. Climate Change Challenges, Plant Science Solutions. Plant Cell 2023, 35, 24–66. [Google Scholar] [CrossRef]
- IPCC. Intergovernmental Panel on Climate Change. Sixth Assessment Report. Impacts, Adaptation and Vulnerability.
- Verslues, P.E.; Bailey-Serres, J.; Brodersen, C.; Buckley, T.N.; Conti, L.; Christmann, A.; Dinneny, J.R.; Grill, E.; Hayes, S.; Heckman, R.W.; et al. Burning Questions for a Warming and Changing World: 15 Unknowns in Plant Abiotic Stress. Plant Cell 2023, 35, 67–108. [Google Scholar] [CrossRef] [PubMed]
- Varotto, S.; Tani, E.; Abraham, E.; Krugman, T.; Kapazoglou, A.; Melzer, R.; Radanović, A.; Miladinović, D. Epigenetics: Possible Applications in Climate-Smart Crop Breeding. J Exp Bot 2020, 71, 5223–5236. [Google Scholar] [CrossRef]
- Ventouris, Y.E.; Tani, E.; Avramidou, E.V.; Abraham, E.M.; Chorianopoulou, S.N.; Vlachostergios, D.N.; Papadopoulos, G.; Kapazoglou, A. Recurrent Water Deficit and Epigenetic Memory in Medicago Sativa L. Varieties. Applied Sciences 2020, 10, 3110. [Google Scholar] [CrossRef]
- Drosou, V.; Kapazoglou, A.; Letsiou, S.; Tsaftaris, A.S.; Argiriou, A. Drought Induces Variation in the DNA Methylation Status of the Barley HvDME Promoter. J Plant Res 2021, 134, 1351–1362. [Google Scholar] [CrossRef]
- de Vries, F.T.; Griffiths, R.I.; Knight, C.G.; Nicolitch, O.; Williams, A. Harnessing Rhizosphere Microbiomes for Drought-Resilient Crop Production. Science (1979) 2020, 368, 270–274. [Google Scholar] [CrossRef]
- Trivedi, P.; Batista, B.D.; Bazany, K.E.; Singh, B.K. Plant–Microbiome Interactions under a Changing World: Responses, Consequences and Perspectives. New Phytologist 2022, 234, 1951–1959. [Google Scholar] [CrossRef]
- Sharma, B.; Singh, B.N.; Dwivedi, P.; Rajawat, M.V.S. Interference of Climate Change on Plant-Microbe Interaction: Present and Future Prospects. Frontiers in Agronomy 2022, 3. [Google Scholar] [CrossRef]
- Diwan, D.; Rashid, M.d.M.; Vaishnav, A. Current Understanding of Plant-Microbe Interaction through the Lenses of Multi-Omics Approaches and Their Benefits in Sustainable Agriculture. Microbiol Res 2022, 265, 127180. [Google Scholar] [CrossRef]
- Joshi, S.; Patil, S.; Shaikh, A.; Jamla, M.; Kumar, V. Modern Omics Toolbox for Producing Combined and Multifactorial Abiotic Stress Tolerant Plants. Plant Stress 2024, 11, 100301. [Google Scholar] [CrossRef]
- Ravikiran, K.T.; Thribhuvan, R.; Anilkumar, C.; Kallugudi, J.; Prakash, N.R.; Adavi B, S.; Sunitha, N.C.; Abhijith, K.P. Harnessing the Power of Genomics to Develop Climate-Smart Crop Varieties: A Comprehensive Review. J Environ Manage 2025, 373, 123461. [Google Scholar] [CrossRef] [PubMed]
- Ali, N.; Singh, S.; Garg, R. Unlocking Crops’ Genetic Potential: Advances in Genome and Epigenome Editing of Regulatory Regions. Curr Opin Plant Biol 2025, 83, 102669. [Google Scholar] [CrossRef]
- Lazaridi, E.; Kapazoglou, A.; Gerakari, M.; Kleftogianni, K.; Passa, K.; Sarri, E.; Papasotiropoulos, V.; Tani, E.; Bebeli, P.J. Crop Landraces and Indigenous Varieties: A Valuable Source of Genes for Plant Breeding. Plants 2024, 13, 758. [Google Scholar] [CrossRef] [PubMed]
- Agius, D.R.; Kapazoglou, A.; Avramidou, E.; Baranek, M.; Carneros, E.; Caro, E.; Castiglione, S.; Cicatelli, A.; Radanovic, A.; Ebejer, J.-P.; et al. Exploring the Crop Epigenome: A Comparison of DNA Methylation Profiling Techniques. Front Plant Sci 2023, 14. [Google Scholar] [CrossRef]
- Kapazoglou, A.; Gerakari, M.; Lazaridi, E.; Kleftogianni, K.; Sarri, E.; Tani, E.; Bebeli, P.J. Crop Wild Relatives: A Valuable Source of Tolerance to Various Abiotic Stresses. Plants 2023, 12, 328. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Dias, M.C.; Freitas, H. Drought and Salinity Stress Responses and Microbe-Induced Tolerance in Plants. Front Plant Sci 2020, 11. [Google Scholar] [CrossRef]
- Mikiciuk, G.; Miller, T.; Kisiel, A.; Cembrowska-Lech, D.; Mikiciuk, M.; Łobodzińska, A.; Bokszczanin, K. Harnessing Beneficial Microbes for Drought Tolerance: A Review of Ecological and Agricultural Innovations. Agriculture 2024, 14, 2228. [Google Scholar] [CrossRef]
- Sharma, A.; Choudhary, P.; Chakdar, H.; Shukla, P. Molecular Insights and Omics-Based Understanding of Plant–Microbe Interactions under Drought Stress. World J Microbiol Biotechnol 2024, 40, 42. [Google Scholar] [CrossRef]
- Salvi, P.; Mahawar, H.; Agarrwal, R.; Kajal; Gautam, V.; Deshmukh, R. Advancement in the Molecular Perspective of Plant-Endophytic Interaction to Mitigate Drought Stress in Plants. Front Microbiol 2022, 13. [Google Scholar] [CrossRef]
- Vandenkoornhuyse, P.; Quaiser, A.; Duhamel, M.; Le Van, A.; Dufresne, A. The Importance of the Microbiome of the Plant Holobiont. New Phytologist 2015, 206, 1196–1206. [Google Scholar] [CrossRef] [PubMed]
- Li, J.-H.; Muhammad Aslam, M.; Gao, Y.-Y.; Dai, L.; Hao, G.-F.; Wei, Z.; Chen, M.-X.; Dini-Andreote, F. Microbiome-Mediated Signal Transduction within the Plant Holobiont. Trends Microbiol 2023, 31, 616–628. [Google Scholar] [CrossRef] [PubMed]
- Ravanbakhsh, M.; Kowalchuk, G.A.; Jousset, A. Targeted Plant Hologenome Editing for Plant Trait Enhancement. New Phytologist 2021, 229, 1067–1077. [Google Scholar] [CrossRef]
- Trivedi, P.; Leach, J.E.; Tringe, S.G.; Sa, T.; Singh, B.K. Plant–Microbiome Interactions: From Community Assembly to Plant Health. Nat Rev Microbiol 2020, 18, 607–621. [Google Scholar] [CrossRef]
- Ling, N.; Wang, T.; Kuzyakov, Y. Rhizosphere Bacteriome Structure and Functions. Nat Commun 2022, 13, 836. [Google Scholar] [CrossRef] [PubMed]
- Jain, A.; Sarsaiya, S.; Singh, R.; Gong, Q.; Wu, Q.; Shi, J. Omics Approaches in Understanding the Benefits of Plant-Microbe Interactions. Front Microbiol 2024, 15. [Google Scholar] [CrossRef]
- Doddavarapu, B.; Lata, C.; Shah, J.M. Epigenetic Regulation Influenced by Soil Microbiota and Nutrients: Paving Road to Epigenome Editing in Plants. Biochimica et Biophysica Acta (BBA) - General Subjects 2024, 1868, 130580. [Google Scholar] [CrossRef]
- Xu, L.; Dong, Z.; Chiniquy, D.; Pierroz, G.; Deng, S.; Gao, C.; Diamond, S.; Simmons, T.; Wipf, H.M.-L.; Caddell, D.; et al. Genome-Resolved Metagenomics Reveals Role of Iron Metabolism in Drought-Induced Rhizosphere Microbiome Dynamics. Nat Commun 2021, 12, 3209. [Google Scholar] [CrossRef]
- Zombardo, A.; Meneghetti, S.; Morreale, G.; Calò, A.; Costacurta, A.; Storchi, P. Study of Inter- and Intra-Varietal Genetic Variability in Grapevine Cultivars. Plants 2022, 11, 397. [Google Scholar] [CrossRef]
- Dong, Y.; Duan, S.; Xia, Q.; Liang, Z.; Dong, X.; Margaryan, K.; Musayev, M.; Goryslavets, S.; Zdunić, G.; Bert, P.-F.; et al. Dual Domestications and Origin of Traits in Grapevine Evolution. Science (1979) 2023, 379, 892–901. [Google Scholar] [CrossRef]
- Wolkovich, E.M.; García de Cortázar-Atauri, I.; Morales-Castilla, I.; Nicholas, K.A.; Lacombe, T. From Pinot to Xinomavro in the World’s Future Wine-Growing Regions. Nat Clim Chang 2018, 8, 29–37. [Google Scholar] [CrossRef]
- Ollat, N.; Marguerit, E.; de Miguel, M.; Coupel-Ledru, A.; Cookson, S.J.; van Leeuwen, C.; Vivin, P.; Gallusci, P.; Segura, V.; Duchêne, E. Moving towards Grapevine Genotypes Better Adapted to Abiotic Constraints. Vitis - Journal of Grapevine Research 2023, 62, 67–76. [Google Scholar] [CrossRef]
- OIV 2024, I.O. of V. and Wine. Available online: Http://Www.Oiv.Int/En/the-International-Organisation-of-Vine-and-Wine (Accessed on 10 January 2024).
- Bernardo, S.; Dinis, L.-T.; Machado, N.; Moutinho-Pereira, J. Grapevine Abiotic Stress Assessment and Search for Sustainable Adaptation Strategies in Mediterranean-like Climates. A Review. Agron Sustain Dev 2018, 38, 66. [Google Scholar] [CrossRef]
- Venios, X.; Korkas, E.; Nisiotou, A.; Banilas, G. Grapevine Responses to Heat Stress and Global Warming. Plants 2020, 9, 1754. [Google Scholar] [CrossRef] [PubMed]
- Mavromatis, T.; Koufos, G.C.; Koundouras, S.; Jones, G.V. Adaptive Capacity of Winegrape Varieties Cultivated in Greece to Climate Change: Current Trends and Future Projections. OENO One 2020, 54, 1201–1219. [Google Scholar] [CrossRef]
- Venios, X.; Gkizi, D.; Nisiotou, A.; Korkas, E.; Tjamos, S.; Zamioudis, C.; Banilas, G. Emerging Roles of Epigenetics in Grapevine and Winegrowing. Plants 2024, 13, 515. [Google Scholar] [CrossRef]
- Pacifico, D.; Squartini, A.; Crucitti, D.; Barizza, E.; Lo Schiavo, F.; Muresu, R.; Carimi, F.; Zottini, M. The Role of the Endophytic Microbiome in the Grapevine Response to Environmental Triggers. Front Plant Sci 2019, 10. [Google Scholar] [CrossRef]
- Francioli, D.; Strack, T.; Dries, L.; Voss-Fels, K.P.; Geilfus, C. Roots of Resilience: Optimizing Microbe-rootstock Interactions to Enhance Vineyard Productivity. PLANTS, PEOPLE, PLANET, 2024. [Google Scholar] [CrossRef]
- Mudge, K.; Janick, J.; Scofield, S.; Goldschmidt, E.E. A History of Grafting. In Horticultural Reviews; Wiley, 2009; pp. 437–493. [Google Scholar]
- Warschefsky, E.J.; Klein, L.L.; Frank, M.H.; Chitwood, D.H.; Londo, J.P.; von Wettberg, E.J.B.; Miller, A.J. Rootstocks: Diversity, Domestication, and Impacts on Shoot Phenotypes. Trends Plant Sci 2016, 21, 418–437. [Google Scholar] [CrossRef]
- Migicovsky, Z.; Cousins, P.; Jordan, L.M.; Myles, S.; Striegler, R.K.; Verdegaal, P.; Chitwood, D.H. Grapevine Rootstocks Affect Growth-related Scion Phenotypes. Plant Direct 2021, 5. [Google Scholar] [CrossRef]
- Chen, Y.; Fei, Y.; Howell, K.; Chen, D.; Clingeleffer, P.; Zhang, P. Rootstocks for Grapevines Now and into the Future: Selection of Rootstocks Based on Drought Tolerance, Soil Nutrient Availability, and Soil PH. Aust J Grape Wine Res 2024, 2024, 1–23. [Google Scholar] [CrossRef]
- Kapazoglou, A.; Tani, E.; Avramidou, E.V.; Abraham, E.M.; Gerakari, M.; Megariti, S.; Doupis, G.; Doulis, A.G. Epigenetic Changes and Transcriptional Reprogramming Upon Woody Plant Grafting for Crop Sustainability in a Changing Environment. Front Plant Sci 2021, 11. [Google Scholar] [CrossRef]
- Darriaut, R.; Lailheugue, V.; Masneuf-Pomarède, I.; Marguerit, E.; Martins, G.; Compant, S.; Ballestra, P.; Upton, S.; Ollat, N.; Lauvergeat, V. Grapevine Rootstock and Soil Microbiome Interactions: Keys for a Resilient Viticulture. Hortic Res 2022, 9. [Google Scholar] [CrossRef] [PubMed]
- Bettenfeld, P.; Cadena i Canals, J.; Jacquens, L.; Fernandez, O.; Fontaine, F.; van Schaik, E.; Courty, P.-E.; Trouvelot, S. The Microbiota of the Grapevine Holobiont: A Key Component of Plant Health. J Adv Res 2022, 40, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Avramidou, E.; Masaoutis, I.; Pitsoli, T.; Kapazoglou, A.; Pikraki, M.; Trantas, E.; Nikolantonakis, M.; Doulis, A. Analysis of Wine-Producing Vitis Vinifera L. Biotypes, Autochthonous to Crete (Greece), Employing Ampelographic and Microsatellite Markers. Life 2023, 13, 220. [Google Scholar] [CrossRef]
- Trenti, M.; Lorenzi, S.; Bianchedi, P.L.; Grossi, D.; Failla, O.; Grando, M.S.; Emanuelli, F. Candidate Genes and SNPs Associated with Stomatal Conductance under Drought Stress in Vitis. BMC Plant Biol 2021, 21, 7. [Google Scholar] [CrossRef]
- Tsivelikas, A.L.; Avramidou, E.V.; Ralli, P.E.; Ganopoulos, I.V.; Moysiadis, T.; Kapazoglou, A.; Aravanopoulos, F.A.; Doulis, A.G. Genetic Diversity of Greek Grapevine ( Vitis Vinifera L.) Cultivars Using Ampelographic and Microsatellite Markers. Plant Genet Resour 2022, 20, 124–136. [Google Scholar] [CrossRef]
- De Lorenzis, G.; Mercati, F.; Bergamini, C.; Cardone, M.F.; Lupini, A.; Mauceri, A.; Caputo, A.R.; Abbate, L.; Barbagallo, M.G.; Antonacci, D.; et al. SNP Genotyping Elucidates the Genetic Diversity of Magna Graecia Grapevine Germplasm and Its Historical Origin and Dissemination. BMC Plant Biol 2019, 19, 7. [Google Scholar] [CrossRef]
- Marinov, L.; Magris, G.; Di Gaspero, G.; Morgante, M.; Maletić, E.; Bubola, M.; Pejić, I.; Zdunić, G. Single Nucleotide Polymorphism (SNP) Analysis Reveals Ancestry and Genetic Diversity of Cultivated and Wild Grapevines in Croatia. BMC Plant Biol 2024, 24, 975. [Google Scholar] [CrossRef]
- Villano, C.; Procino, S.; Blaiotta, G.; Carputo, D.; D’Agostino, N.; Di Serio, E.; Fanelli, V.; La Notte, P.; Miazzi, M.M.; Montemurro, C.; et al. Genetic Diversity and Signature of Divergence in the Genome of Grapevine Clones of Southern Italy Varieties. Front Plant Sci 2023, 14. [Google Scholar] [CrossRef]
- Villano, C.; Aiese Cigliano, R.; Esposito, S.; D’Amelia, V.; Iovene, M.; Carputo, D.; Aversano, R. DNA-Based Technologies for Grapevine Biodiversity Exploitation: State of the Art and Future Perspectives. Agronomy 2022, 12, 491. [Google Scholar] [CrossRef]
- Zhang, L.; Fan, D.; Li, H.; Chen, Q.; Zhang, Z.; Liu, M.; Liu, J.; Song, Y.; He, J.; Xu, W.; et al. Characterization and Identification of Grapevine Heat Stress-Responsive MicroRNAs Revealed the Positive Regulated Function of Vvi-MiR167 in Thermostability. Plant Science 2023, 329, 111623. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Song, Y.; Li, J.; Liu, J.; Zhang, Z.; Xu, Y.; Fan, D.; Liu, M.; Ren, Y.; Xi, X.; et al. Development, Identification and Validation of a Novel SSR Molecular Marker for Heat Resistance of Grapes Based on MiRNA. Horticulturae 2023, 9, 931. [Google Scholar] [CrossRef]
- Shi, X.; Cao, S.; Wang, X.; Huang, S.; Wang, Y.; Liu, Z.; Liu, W.; Leng, X.; Peng, Y.; Wang, N.; et al. The Complete Reference Genome for Grapevine ( Vitis Vinifera L.) Genetics and Breeding. Hortic Res 2023, 10. [Google Scholar] [CrossRef]
- Cochetel, N.; Minio, A.; Guarracino, A.; Garcia, J.F.; Figueroa-Balderas, R.; Massonnet, M.; Kasuga, T.; Londo, J.P.; Garrison, E.; Gaut, B.S.; et al. A Super-Pangenome of the North American Wild Grape Species. Genome Biol 2023, 24, 290. [Google Scholar] [CrossRef]
- Liu, Z.; Wang, N.; Su, Y.; Long, Q.; Peng, Y.; Shangguan, L.; Zhang, F.; Cao, S.; Wang, X.; Ge, M.; et al. Grapevine Pangenome Facilitates Trait Genetics and Genomic Breeding. Nat Genet 2024, 56, 2804–2814. [Google Scholar] [CrossRef]
- Magon, G.; De Rosa, V.; Martina, M.; Falchi, R.; Acquadro, A.; Barcaccia, G.; Portis, E.; Vannozzi, A.; De Paoli, E. Boosting Grapevine Breeding for Climate-Smart Viticulture: From Genetic Resources to Predictive Genomics. Front Plant Sci 2023, 14. [Google Scholar] [CrossRef]
- Yan, S.; Liu, Q.; Li, W.; Yan, J.; Fernie, A.R. Raffinose Family Oligosaccharides: Crucial Regulators of Plant Development and Stress Responses. CRC Crit Rev Plant Sci 2022, 41, 286–303. [Google Scholar] [CrossRef]
- Wang, P.; Zhao, F.; Zheng, T.; Liu, Z.; Ji, X.; Zhang, Z.; Pervaiz, T.; Shangguan, L.; Fang, J. Whole-Genome Re-Sequencing, Diversity Analysis, and Stress-Resistance Analysis of 77 Grape Rootstock Genotypes. Front Plant Sci 2023, 14. [Google Scholar] [CrossRef]
- Zhang, X.; Zheng, Q.; Hao, Y.; Zhang, Y.; Gu, W.; Deng, Z.; Zhou, P.; Fang, Y.; Chen, K.; Zhang, K. Physiology and Transcriptome Profiling Reveal the Drought Tolerance of Five Grape Varieties under High Temperatures. J Integr Agric 2024. [Google Scholar] [CrossRef]
- Tan, J.W.; Shinde, H.; Tesfamicael, K.; Hu, Y.; Fruzangohar, M.; Tricker, P.; Baumann, U.; Edwards, E.J.; Rodríguez López, C.M. Global Transcriptome and Gene Co-Expression Network Analyses Reveal Regulatory and Non-Additive Effects of Drought and Heat Stress in Grapevine. Front Plant Sci 2023, 14. [Google Scholar] [CrossRef] [PubMed]
- Dou, F.; Phillip, F.O.; Liu, G.; Zhu, J.; Zhang, L.; Wang, Y.; Liu, H. Transcriptomic and Physiological Analyses Reveal Different Grape Varieties Response to High Temperature Stress. Front Plant Sci 2024, 15. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Zeng, Z.; Liu, Z.; Xia, R. Small RNAs, Emerging Regulators Critical for the Development of Horticultural Traits. Hortic Res 2018, 5, 63. [Google Scholar] [CrossRef]
- Ferrandino, A.; Pagliarani, C.; Pérez-Álvarez, E.P. Secondary Metabolites in Grapevine: Crosstalk of Transcriptional, Metabolic and Hormonal Signals Controlling Stress Defence Responses in Berries and Vegetative Organs. Front Plant Sci 2023, 14. [Google Scholar] [CrossRef]
- Pagliarani, C.; Vitali, M.; Ferrero, M.; Vitulo, N.; Incarbone, M.; Lovisolo, C.; Valle, G.; Schubert, A. The Accumulation of MiRNAs Differentially Modulated by Drought Stress Is Affected by Grafting in Grapevine. Plant Physiol 2017, 173, 2180–2195. [Google Scholar] [CrossRef]
- Guo, S.; Xu, T.; Ju, Y.; Lei, Y.; Zhang, F.; Fang, Y.; Zhang, Z.; Jin, L.; Meng, J. MicroRNAs Behave Differently to Drought Stress in Drought-Tolerant and Drought-Sensitive Grape Genotypes. Environ Exp Bot 2023, 207, 105223. [Google Scholar] [CrossRef]
- Maniatis, G.; Tani, E.; Katsileros, A.; Avramidou, E.V.; Pitsoli, T.; Sarri, E.; Gerakari, M.; Goufa, M.; Panagoulakou, M.; Xipolitaki, K.; et al. Genetic and Epigenetic Responses of Autochthonous Grapevine Cultivars from the ‘Epirus’ Region of Greece upon Consecutive Drought Stress. Plants 2023, 13, 27. [Google Scholar] [CrossRef]
- Zang, E.; Jiang, L.; Cui, H.; Li, X.; Yan, Y.; Liu, Q.; Chen, Z.; Li, M. Only Plant-Based Food Additives: An Overview on Application, Safety, and Key Challenges in the Food Industry. Food Reviews International 2023, 39, 5132–5163. [Google Scholar] [CrossRef]
- Guo, S.; Zhang, M.; Feng, M.; Liu, G.; Torregrosa, L.; Tao, X.; Ren, R.; Fang, Y.; Zhang, Z.; Meng, J.; et al. MiR156b-Targeted VvSBP8/13 Functions Downstream of the Abscisic Acid Signal to Regulate Anthocyanins Biosynthesis in Grapevine Fruit under Drought. Hortic Res 2024, 11. [Google Scholar] [CrossRef]
- Feng, C.; Zhang, X.; Du, B.; Xiao, Y.; Wang, Y.; Sun, Y.; Zhou, X.; Wang, C.; Liu, Y.; Li, T.-H. MicroRNA156ab Regulates Apple Plant Growth and Drought Tolerance by Targeting Transcription Factor MsSPL13. Plant Physiol 2023, 192, 1836–1857. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Du, Y.; Xiang, J.; Zheng, T.; Cheng, J.; Wu, J. Integrated MRNA and MiRNA Transcriptome Analysis of Grape in Responses to Salt Stress. Front Plant Sci 2023, 14. [Google Scholar] [CrossRef]
- Rooy, S.S.B.; Ghabooli, M.; Salekdeh, G.H.; Fard, E.M.; Karimi, R.; Fakhrfeshani, M.; Gholami, M. Identification of Novel Cold Stress Responsive MicroRNAs and Their Putative Targets in ‘Sultana’ Grapevine (Vitis Vinifera) Using RNA Deep Sequencing. Acta Physiol Plant 2023, 45, 2. [Google Scholar] [CrossRef]
- Zhang, Y.; Qiao, Z.; Li, X.; Ren, Z.; Xu, S.; Liu, Z.; Wang, K. 5-Azacytidine Enhances the Aluminum Tolerance of Grapes by Reducing the DNA Methylation Level. J Soil Sci Plant Nutr 2024, 24, 6922–6937. [Google Scholar] [CrossRef]
- Zhu, Z.; Li, Q.; Gichuki, D.K.; Hou, Y.; Liu, Y.; Zhou, H.; Xu, C.; Fang, L.; Gong, L.; Zheng, B.; et al. Genome-Wide Profiling of Histone H3 Lysine 27 Trimethylation and Its Modification in Response to Chilling Stress in Grapevine Leaves. Hortic Plant J 2023, 9, 496–508. [Google Scholar] [CrossRef]
- Ren, C.; Mohamed, M.S.M.; Aini, N.; Kuang, Y.; Liang, Z. CRISPR/Cas in Grapevine Genome Editing: The Best Is Yet to Come. Horticulturae 2024, 10, 965. [Google Scholar] [CrossRef]
- Benny, J.; Marchese, A.; Giovino, A.; Marra, F.P.; Perrone, A.; Caruso, T.; Martinelli, F. Gaining Insight into Exclusive and Common Transcriptomic Features Linked to Drought and Salinity Responses across Fruit Tree Crops. Plants 2020, 9, 1059. [Google Scholar] [CrossRef]
- Bazakos, C.; Manioudaki, M.E.; Therios, I.; Voyiatzis, D.; Kafetzopoulos, D.; Awada, T.; Kalaitzis, P. Comparative Transcriptome Analysis of Two Olive Cultivars in Response to NaCl-Stress. PLoS One 2012, 7, e42931. [Google Scholar] [CrossRef]
- Mousavi, S.; Regni, L.; Bocchini, M.; Mariotti, R.; Cultrera, N.G.M.; Mancuso, S.; Googlani, J.; Chakerolhosseini, M.R.; Guerrero, C.; Albertini, E.; et al. Physiological, Epigenetic and Genetic Regulation in Some Olive Cultivars under Salt Stress. Sci Rep 2019, 9, 1093. [Google Scholar] [CrossRef]
- Ghirardo, A.; Nosenko, T.; Kreuzwieser, J.; Winkler, J.B.; Kruse, J.; Albert, A.; Merl-Pham, J.; Lux, T.; Ache, P.; Zimmer, I.; et al. Protein Expression Plasticity Contributes to Heat and Drought Tolerance of Date Palm. Oecologia 2021, 197, 903–919. [Google Scholar] [CrossRef]
- Mueller, H.M.; Franzisky, B.L.; Messerer, M.; Du, B.; Lux, T.; White, P.J.; Carpentier, S.C.; Winkler, J.B.; Schnitzler, J.; El-Serehy, H.A.; et al. Integrative Multi-omics Analyses of Date Palm ( Phoenix Dactylifera ) Roots and Leaves Reveal How the Halophyte Land Plant Copes with Sea Water. Plant Genome 2024, 17. [Google Scholar] [CrossRef]
- Xu, Z.; Zhang, N.; Fu, H.; Wang, F.; Wen, M.; Chang, H.; Wu, J.; Abdelaala, W.B.; Luo, Q.; Li, Y.; et al. Salt Stress Modulates the Landscape of Transcriptome and Alternative Splicing in Date Palm (Phoenix Dactylifera L.). Front Plant Sci 2022, 12. [Google Scholar] [CrossRef]
- Al-Harrasi, I.; Al-Yahyai, R.; Yaish, M.W. Differential DNA Methylation and Transcription Profiles in Date Palm Roots Exposed to Salinity. PLoS One 2018, 13, e0191492. [Google Scholar] [CrossRef]
- Kumawat, S.; Sharma, Y.; Vats, S.; Sudhakaran, S.; Sharma, S.; Mandlik, R.; Raturi, G.; Kumar, V.; Rana, N.; Kumar, A.; et al. Understanding the Role of SWEET Genes in Fruit Development and Abiotic Stress in Pomegranate (Punica Granatum L.). Mol Biol Rep 2022, 49, 1329–1339. [Google Scholar] [CrossRef]
- Liu, C.; Zhao, Y.; Zhao, X.; Wang, J.; Gu, M.; Yuan, Z. Transcriptomic Profiling of Pomegranate Provides Insights into Salt Tolerance. Agronomy 2019, 10, 44. [Google Scholar] [CrossRef]
- Tang, H.; Wang, C.; Mei, J.; Feng, L.; Wu, Q.; Yin, Y. Transcriptome Analysis Revealed the Response Mechanism of Pomegranate to Salt Stress. Agronomy 2024, 14, 2261. [Google Scholar] [CrossRef]
- Wang, L.; Wang, S.; Tong, R.; Wang, S.; Yao, J.; Jiao, J.; Wan, R.; Wang, M.; Shi, J.; Zheng, X. Overexpression of PgCBF3 and PgCBF7 Transcription Factors from Pomegranate Enhances Freezing Tolerance in Arabidopsis under the Promoter Activity Positively Regulated by PgICE1. Int J Mol Sci 2022, 23, 9439. [Google Scholar] [CrossRef]
- Guan, S.; Chai, Y.; Hao, Q.; Ma, Y.; Wan, W.; Liu, H.; Diao, M. Transcriptomic and Physiological Analysis Reveals Crucial Biological Pathways Associated with Low-Temperature Stress in Tunisian Soft-Seed Pomegranate ( Punica Granatum L.). J Plant Interact 2023, 18. [Google Scholar] [CrossRef]
- Liu, L.; Xu, S.; Zhang, L.; Zheng, J. A Genome-Wide Analysis of the BAM Gene Family and Identification of the Cold-Responsive Genes in Pomegranate (Punica Granatum L.). Plants 2024, 13, 1321. [Google Scholar] [CrossRef]
- Rolli, E.; Marasco, R.; Vigani, G.; Ettoumi, B.; Mapelli, F.; Deangelis, M.L.; Gandolfi, C.; Casati, E.; Previtali, F.; Gerbino, R.; et al. Improved Plant Resistance to Drought Is Promoted by the Root-associated Microbiome as a Water Stress-dependent Trait. Environ Microbiol 2015, 17, 316–331. [Google Scholar] [CrossRef]
- Salomon, M.V.; Purpora, R.; Bottini, R.; Piccoli, P. Rhizosphere Associated Bacteria Trigger Accumulation of Terpenes in Leaves of Vitis Vinifera L. Cv. Malbec That Protect Cells against Reactive Oxygen Species. Plant Physiology and Biochemistry 2016, 106, 295–304. [Google Scholar] [CrossRef]
- Rolli, E.; Marasco, R.; Saderi, S.; Corretto, E.; Mapelli, F.; Cherif, A.; Borin, S.; Valenti, L.; Sorlini, C.; Daffonchio, D. Root-Associated Bacteria Promote Grapevine Growth: From the Laboratory to the Field. Plant Soil 2017, 410, 369–382. [Google Scholar] [CrossRef]
- Torres, N.; Goicoechea, N.; Zamarreño, A.M.; Carmen Antolín, M. Mycorrhizal Symbiosis Affects ABA Metabolism during Berry Ripening in Vitis Vinifera L. Cv. Tempranillo Grown under Climate Change Scenarios. Plant Science 2018, 274, 383–393. [Google Scholar] [CrossRef]
- Ye, Q.; Wang, H.; Li, H. Arbuscular Mycorrhizal Fungi Enhance Drought Stress Tolerance by Regulating Osmotic Balance, the Antioxidant System, and the Expression of Drought-Responsive Genes in Vitis Vinifera L. Aust J Grape Wine Res 2023, 2023, 1–13. [Google Scholar] [CrossRef]
- Cardinale, M.; Minervini, F.; De Angelis, M.; Papadia, P.; Migoni, D.; Dimaglie, M.; Dinu, D.G.; Quarta, C.; Selleri, F.; Caccioppola, A.; et al. Vineyard Establishment under Exacerbated Summer Stress: Effects of Mycorrhization on Rootstock Agronomical Parameters, Leaf Element Composition and Root-Associated Bacterial Microbiota. Plant Soil 2022, 478, 613–634. [Google Scholar] [CrossRef]
- Carreiras, J.; Cruz-Silva, A.; Fonseca, B.; Carvalho, R.C.; Cunha, J.P.; Proença Pereira, J.; Paiva-Silva, C.; A. Santos, S.; Janeiro Sequeira, R.; Mateos-Naranjo, E.; et al. Improving Grapevine Heat Stress Resilience with Marine Plant Growth-Promoting Rhizobacteria Consortia. Microorganisms 2023, 11, 856. [Google Scholar] [CrossRef]
- Yadav, V.; Zhong, H.; Patel, M.K.; Zhang, S.; Zhou, X.; Zhang, C.; Zhang, J.; Su, J.; Zhang, F.; Wu, X. Integrated Omics-Based Exploration for Temperature Stress Resilience: An Approach to Smart Grape Breeding Strategies. Plant Stress 2024, 11, 100356. [Google Scholar] [CrossRef]
- Gobbi, A.; Acedo, A.; Imam, N.; Santini, R.G.; Ortiz-Álvarez, R.; Ellegaard-Jensen, L.; Belda, I.; Hansen, L.H. A Global Microbiome Survey of Vineyard Soils Highlights the Microbial Dimension of Viticultural Terroirs. Commun Biol 2022, 5, 241. [Google Scholar] [CrossRef]
- Anand, L.; Gentimis, T.; Downie, A.B.; Lopez, C.M.R. Grapevine Rootstock and Scion Genotypes’ Symbiosis with Soil Microbiome: A Machine Learning Revelation for Climate-Resilient Viticulture 2024.
- Griggs, R.G.; Steenwerth, K.L.; Mills, D.A.; Cantu, D.; Bokulich, N.A. Sources and Assembly of Microbial Communities in Vineyards as a Functional Component of Winegrowing. Front Microbiol 2021, 12. [Google Scholar] [CrossRef]
- Colautti, A.; Mian, G.; Tomasi, D.; Bell, L.; Marcuzzo, P. Exploring the Influence of Soil Salinity on Microbiota Dynamics in Vitis Vinifera Cv. “Glera”: Insights into the Rhizosphere, Carposphere, and Yield Outcomes. Diversity (Basel) 2024, 16, 247. [Google Scholar] [CrossRef]
- Jatan, R.; Chauhan, P.S.; Lata, C. Pseudomonas Putida Modulates the Expression of MiRNAs and Their Target Genes in Response to Drought and Salt Stresses in Chickpea (Cicer Arietinum L.). Genomics 2019, 111, 509–519. [Google Scholar] [CrossRef]
- Chen, C.; Wang, M.; Zhu, J.; Tang, Y.; Zhang, H.; Zhao, Q.; Jing, M.; Chen, Y.; Xu, X.; Jiang, J.; et al. Long-Term Effect of Epigenetic Modification in Plant–Microbe Interactions: Modification of DNA Methylation Induced by Plant Growth-Promoting Bacteria Mediates Promotion Process. Microbiome 2022, 10, 36. [Google Scholar] [CrossRef] [PubMed]
- Campos, C.; Coito, J.L.; Cardoso, H.; Marques da Silva, J.; Pereira, H.S.; Viegas, W.; Nogales, A. Dynamic Regulation of Grapevine’s MicroRNAs in Response to Mycorrhizal Symbiosis and High Temperature. Plants 2023, 12, 982. [Google Scholar] [CrossRef]
- Sportes, A.; Hériché, M.; Mounier, A.; Durney, C.; van Tuinen, D.; Trouvelot, S.; Wipf, D.; Courty, P.E. Comparative RNA Sequencing-Based Transcriptome Profiling of Ten Grapevine Rootstocks: Shared and Specific Sets of Genes Respond to Mycorrhizal Symbiosis. Mycorrhiza 2023, 33, 369–385. [Google Scholar] [CrossRef]
- Dias, M.C.; Silva, S.; Galhano, C.; Lorenzo, P. Olive Tree Belowground Microbiota: Plant Growth-Promoting Bacteria and Fungi. Plants 2024, 13, 1848. [Google Scholar] [CrossRef]
- Azri, R.; Lamine, M.; Bensalem-Fnayou, A.; Hamdi, Z.; Mliki, A.; Ruiz-Lozano, J.M.; Aroca, R. Genotype-Dependent Response of Root Microbiota and Leaf Metabolism in Olive Seedlings Subjected to Drought Stress. Plants 2024, 13, 857. [Google Scholar] [CrossRef]
- Aganchich, B.; Wahbi, S.; Yaakoubi, A.; El-Aououad, H.; Bota, J. Effect of Arbuscular Mycorrhizal Fungi Inoculation on Growth and Physiology Performance of Olive Trees under Regulated Deficit Irrigation and Partial Rootzone Drying. South African Journal of Botany 2022, 148, 1–10. [Google Scholar] [CrossRef]
- Tekaya, M.; Dabbaghi, O.; Guesmi, A.; Attia, F.; Chehab, H.; Khezami, L.; Algathami, F.K.; Ben Hamadi, N.; Hammami, M.; Prinsen, E.; et al. Arbuscular Mycorrhizas Modulate Carbohydrate, Phenolic Compounds and Hormonal Metabolism to Enhance Water Deficit Tolerance of Olive Trees (Olea Europaea). Agric Water Manag 2022, 274, 107947. [Google Scholar] [CrossRef]
- Ben Hassena, A.; Zouari, M.; Trabelsi, L.; Decou, R.; Ben Amar, F.; Chaari, A.; Soua, N.; Labrousse, P.; Khabou, W.; Zouari, N. Potential Effects of Arbuscular Mycorrhizal Fungi in Mitigating the Salinity of Treated Wastewater in Young Olive Plants (Olea Europaea L. Cv. Chetoui). Agric Water Manag 2021, 245, 106635. [Google Scholar] [CrossRef]
- Cherif, H.; Marasco, R.; Rolli, E.; Ferjani, R.; Fusi, M.; Soussi, A.; Mapelli, F.; Blilou, I.; Borin, S.; Boudabous, A.; et al. Oasis Desert Farming Selects Environment-specific Date Palm Root Endophytic Communities and Cultivable Bacteria That Promote Resistance to Drought. Environ Microbiol Rep 2015, 7, 668–678. [Google Scholar] [CrossRef]
- Sabeem, M.; Abdul Aziz, M.; Mullath, S.K.; Brini, F.; Rouached, H.; Masmoudi, K. Enhancing Growth and Salinity Stress Tolerance of Date Palm Using Piriformospora Indica. Front Plant Sci 2022, 13. [Google Scholar] [CrossRef]
- Ahmad, M.; Abdul Aziz, M.; Sabeem, M.; Kutty, M.S.; Sivasankaran, S.K.; Brini, F.; Xiao, T.T.; Blilou, I.; Masmoudi, K. Date Palm Transcriptome Analysis Provides New Insights on Changes in Response to High Salt Stress of Colonized Roots with the Endophytic Fungus Piriformospora Indica. Front Plant Sci 2024, 15. [Google Scholar] [CrossRef]
- Jana, G.A.; Yaish, M.W. Isolation and Functional Characterization of a MVOC Producing Plant-Growth-Promoting Bacterium Isolated from the Date Palm Rhizosphere. Rhizosphere 2020, 16, 100267. [Google Scholar] [CrossRef]
- Bompadre, M.J.; Silvani, V.A.; Bidondo, L.F.; Ríos de Molina, M.D.C.; Colombo, R.P.; Pardo, A.G.; Godeas, A.M. Arbuscular Mycorrhizal Fungi Alleviate Oxidative Stress in Pomegranate Plants Growing under Different Irrigation Conditions. Botany 2014, 92, 187–193. [Google Scholar] [CrossRef]
- Diez, C.M.; Trujillo, I.; Martinez-Urdiroz, N.; Barranco, D.; Rallo, L.; Marfil, P.; Gaut, B.S. Olive Domestication and Diversification in the Mediterranean Basin. New Phytologist 2015, 206, 436–447. [Google Scholar] [CrossRef]
- Langgut, D.; Garfinkel, Y. 7000-Year-Old Evidence of Fruit Tree Cultivation in the Jordan Valley, Israel. Sci Rep 2022, 12, 7463. [Google Scholar] [CrossRef]
- FAOSTATS. https://www.fao.org/faostat/en/#data/QCL;IOC, https://www.internationaloliveoil.org/) FAOSTATS,Https://Www.Fao.Org/Faostat/En/#data/QCL;IOC, Https://Www.Internationaloliveoil.Org/).
- El Yamani, M.; Cordovilla, M. del P. Tolerance Mechanisms of Olive Tree (Olea Europaea) under Saline Conditions. Plants 2024, 13, 2094. [Google Scholar] [CrossRef]
- Fraga, H.; Moriondo, M.; Leolini, L.; Santos, J.A. Mediterranean Olive Orchards under Climate Change: A Review of Future Impacts and Adaptation Strategies. Agronomy 2020, 11, 56. [Google Scholar] [CrossRef]
- Cardoni, M.; Mercado-Blanco, J. Confronting Stresses Affecting Olive Cultivation from the Holobiont Perspective. Front Plant Sci 2023, 14. [Google Scholar] [CrossRef]
- Hrameche, O.; Tul, S.; Manolikaki, I.; Digalaki, N.; Kaltsa, I.; Psarras, G.; Koubouris, G. Optimizing Agroecological Measures for Climate-Resilient Olive Farming in the Mediterranean. Plants 2024, 13, 900. [Google Scholar] [CrossRef]
- Nteve, G.-M.; Kostas, S.; Polidoros, A.N.; Madesis, P.; Nianiou-Obeidat, I. Adaptation Mechanisms of Olive Tree under Drought Stress: The Potential of Modern Omics Approaches. Agriculture 2024, 14, 579. [Google Scholar] [CrossRef]
- De Ollas, C.; Morillón, R.; Fotopoulos, V.; Puértolas, J.; Ollitrault, P.; Gómez-Cadenas, A.; Arbona, V. Facing Climate Change: Biotechnology of Iconic Mediterranean Woody Crops. Front Plant Sci 2019, 10. [Google Scholar] [CrossRef]
- Bazakos, C.; Alexiou, K.G.; Ramos-Onsins, S.; Koubouris, G.; Tourvas, N.; Xanthopoulou, A.; Mellidou, I.; Moysiadis, T.; Vourlaki, I.; Metzidakis, I.; et al. Whole Genome Scanning of a Mediterranean Basin Hotspot Collection Provides New Insights into Olive Tree Biodiversity and Biology. The Plant Journal 2023, 116, 303–319. [Google Scholar] [CrossRef]
- Mousavi, S.; Mariotti, R.; Valeri, M.C.; Regni, L.; Lilli, E.; Albertini, E.; Proietti, P.; Businelli, D.; Baldoni, L. Characterization of Differentially Expressed Genes under Salt Stress in Olive. Int J Mol Sci 2021, 23, 154. [Google Scholar] [CrossRef]
- Bullones, A.; Castro, A.J.; Lima-Cabello, E.; Alché, J.d.D.; Luque, F.; Claros, M.G.; Fernandez-Pozo, N. OliveAtlas: A Gene Expression Atlas Tool for Olea Europaea. Plants 2023, 12, 1274. [Google Scholar] [CrossRef]
- Dias, M.C.; Araújo, M.; Ma, Y. A Bio-Based Strategy for Sustainable Olive Performance under Water Deficit Conditions. Plant Stress 2024, 11, 100342. [Google Scholar] [CrossRef]
- Galicia-Campos, E.; García-Villaraco, A.; Montero-Palmero, Ma.B.; Gutiérrez-Mañero, F.J.; Ramos-Solano, B. Bacillus G7 Improves Adaptation to Salt Stress in Olea Europaea L. Plantlets, Enhancing Water Use Efficiency and Preventing Oxidative Stress. Sci Rep 2023, 13, 22507. [Google Scholar] [CrossRef]
- Sallami, A.; Rachidi, F.; Lahsini, A.I.; El Khedri, H.; Douira, A.; El Modafar, C.; Medraoui, L.; Filali-Maltouf, A. Plant Growth Promoting (PGP) Performances and Diversity of Bacterial Species Isolated from Olive (Olea Europaea L.) Rhizosphere in Arid and Semi-Arid Regions of Morocco. J Pure Appl Microbiol 2023, 17, 2165–2178. [Google Scholar] [CrossRef]
- Malacrinò, A.; Mosca, S.; Li Destri Nicosia, M.G.; Agosteo, G.E.; Schena, L. Plant Genotype Shapes the Bacterial Microbiome of Fruits, Leaves, and Soil in Olive Plants. Plants 2022, 11, 613. [Google Scholar] [CrossRef]
- Vita, F.; Sabbatini, L.; Sillo, F.; Ghignone, S.; Vergine, M.; Guidi Nissim, W.; Fortunato, S.; Salzano, A.M.; Scaloni, A.; Luvisi, A.; et al. Salt Stress in Olive Tree Shapes Resident Endophytic Microbiota. Front Plant Sci 2022, 13. [Google Scholar] [CrossRef]
- Kakagianni, M.; Tsiknia, M.; Feka, M.; Vasileiadis, S.; Leontidou, K.; Kavroulakis, N.; Karamanoli, K.; Karpouzas, D.G.; Ehaliotis, C.; Papadopoulou, K.K. Above- and below-Ground Microbiome in the Annual Developmental Cycle of Two Olive Tree Varieties. FEMS Microbes 2023, 4. [Google Scholar] [CrossRef] [PubMed]
- Vergine, M.; Vita, F.; Casati, P.; Passera, A.; Ricciardi, L.; Pavan, S.; Aprile, A.; Sabella, E.; De Bellis, L.; Luvisi, A. Characterization of the Olive Endophytic Community in Genotypes Displaying a Contrasting Response to Xylella Fastidiosa. BMC Plant Biol 2024, 24, 337. [Google Scholar] [CrossRef]
- Chao, C.T.; Krueger, R.R. The Date Palm (Phoenix Dactylifera L.): Overview of Biology, Uses, and Cultivation. HortScience 2007, 42, 1077–1082. [Google Scholar] [CrossRef]
- Du, B.; Kruse, J.; Winkler, J.B.; Alfarray, S.; Schnitzler, J.-P.; Ache, P.; Hedrich, R.; Rennenberg, H. Climate and Development Modulate the Metabolome and Antioxidative System of Date Palm Leaves. J Exp Bot 2019, 70, 5959–5969. [Google Scholar] [CrossRef]
- Arab, L.; Kreuzwieser, J.; Kruse, J.; Zimmer, I.; Ache, P.; Alfarraj, S.; Al-Rasheid, K.A.S.; Schnitzler, J.-P.; Hedrich, R.; Rennenberg, H. Acclimation to Heat and Drought—Lessons to Learn from the Date Palm (Phoenix Dactylifera). Environ Exp Bot 2016, 125, 20–30. [Google Scholar] [CrossRef]
- Hazzouri, K.M.; Flowers, J.M.; Nelson, D.; Lemansour, A.; Masmoudi, K.; Amiri, K.M.A. Prospects for the Study and Improvement of Abiotic Stress Tolerance in Date Palms in the Post-Genomics Era. Front Plant Sci 2020, 11. [Google Scholar] [CrossRef]
- Rahman, H.; Vikram, P.; Hammami, Z.; Singh, R.K. Recent Advances in Date Palm Genomics: A Comprehensive Review. Front Genet 2022, 13. [Google Scholar] [CrossRef]
- Pandey, N.; Tripathi, P.; Pandey, N.; Nakum, H.; Vala, Y.S. Advancements in Date Palm Genomics and Biotechnology Genomic Resources to the Precision Agriculture: A Comprehensive Review 2024.
- Yaish, M.W.; Patankar, H.V.; Assaha, D.V.M.; Zheng, Y.; Al-Yahyai, R.; Sunkar, R. Genome-Wide Expression Profiling in Leaves and Roots of Date Palm (Phoenix Dactylifera L.) Exposed to Salinity. BMC Genomics 2017, 18, 246. [Google Scholar] [CrossRef]
- Safronov, O.; Kreuzwieser, J.; Haberer, G.; Alyousif, M.S.; Schulze, W.; Al-Harbi, N.; Arab, L.; Ache, P.; Stempfl, T.; Kruse, J.; et al. Detecting Early Signs of Heat and Drought Stress in Phoenix Dactylifera (Date Palm). PLoS One 2017, 12, e0177883. [Google Scholar] [CrossRef]
- Rekik, I.; Chaâbene, Z.; Kriaa, W.; Rorat, A.; Franck, V.; Hafedh, M.; Elleuch, A. Transcriptome Assembly and Abiotic Related Gene Expression Analysis of Date Palm Reveal Candidate Genes Involved in Response to Cadmium Stress. Comparative Biochemistry and Physiology Part C: Toxicology & Pharmacology 2019, 225, 108569. [Google Scholar] [CrossRef]
- Du, B.; Kruse, J.; Winkler, J.B.; Alfarraj, S.; Albasher, G.; Schnitzler, J.-P.; Ache, P.; Hedrich, R.; Rennenberg, H. Metabolic Responses of Date Palm ( Phoenix Dactylifera L.) Leaves to Drought Differ in Summer and Winter Climate. Tree Physiol 2021, 41, 1685–1700. [Google Scholar] [CrossRef] [PubMed]
- Hussain, M.I.; Danish, S.; Naqvi, S.A.; Jaskani, M.J.; Asghar, M.A.; Khan, I.A.; Munir, M.; Muscolo, A. Physiological, Biochemical, and Comparative Genome Analysis of Salt and Drought Stress Impact on Date Palm (Phoenix Dactylifera L.): Tolerance Mechanism and Management. Plant Growth Regul 2024, 104, 1261–1283. [Google Scholar] [CrossRef]
- Kordrostami, M.; Mafakheri, M.; Al-Khayri, J.M. Date Palm (Phoenix Dactylifera L.) Genetic Improvement via Biotechnological Approaches. Tree Genet Genomes 2022, 18, 26. [Google Scholar] [CrossRef]
- Yaish, M.W.; Sunkar, R.; Zheng, Y.; Ji, B.; Al-Yahyai, R.; Farooq, S.A. A Genome-Wide Identification of the MiRNAome in Response to Salinity Stress in Date Palm (Phoenix Dactylifera L.). Front Plant Sci 2015, 6. [Google Scholar] [CrossRef]
- Kumar, K.; Amaresan, N.; Bhagat, S.; Madhuri, K.; Srivastava, R.C. Isolation and Characterization of Rhizobacteria Associated with Coastal Agricultural Ecosystem of Rhizosphere Soils of Cultivated Vegetable Crops. World J Microbiol Biotechnol 2011, 27, 1625–1632. [Google Scholar] [CrossRef]
- Yaish, M.W.; Al-Harrasi, I.; Alansari, A.S.; Al-Yahyai, R.; Glick, B.R. The Use of High Throughput DNA Sequence Analysis to Assess the Endophytic Microbiome of Date Palm Roots Grown under Different Levels of Salt Stress. Int Microbiol 2016, 19, 143–155. [Google Scholar] [CrossRef]
- Khayyat, M.; Vazifeshenas, M.; Akbari, M. Pomegranate and Salt Stress Responses—Assimilation Activities and Chlorophyll Fluorescence Performances: A Review. Applied Fruit Science 2024, 66, 2455–2468. [Google Scholar] [CrossRef]
- Qian, J.; Zhang, X.; Yan, Y.; Wang, N.; Ge, W.; Zhou, Q.; Yang, Y. Unravelling the Molecular Mechanisms of Abscisic Acid-Mediated Drought-Stress Alleviation in Pomegranate (Punica Granatum L.). Plant Physiology and Biochemistry 2020, 157, 211–218. [Google Scholar] [CrossRef]
- Adiba, A.; Haddioui, A.; Hamdani, A.; Kettabi, Z.E.; Outghouliast, H.; Charafi, J. Impact of Contrasting Climate Conditions on Pomegranate Development and Productivity: Implications for Breeding and Cultivar Selection in Colder Environments. Vegetos 2024. [Google Scholar] [CrossRef]
- Eom, J.-S.; Chen, L.-Q.; Sosso, D.; Julius, B.T.; Lin, I.; Qu, X.-Q.; Braun, D.M.; Frommer, W.B. SWEETs, Transporters for Intracellular and Intercellular Sugar Translocation. Curr Opin Plant Biol 2015, 25, 53–62. [Google Scholar] [CrossRef]
- ZHAO, L.; YAO, J.; CHEN, W.; LI, Y.; LÜ, Y.; GUO, Y.; WANG, J.; YUAN, L.; LIU, Z.; ZHANG, Y. A Genome-Wide Analysis of SWEET Gene Family in Cotton and Their Expressions under Different Stresses. Journal of Cotton Research 2018, 1, 7. [Google Scholar] [CrossRef]
- Feng, C.-Y.; Han, J.-X.; Han, X.-X.; Jiang, J. Genome-Wide Identification, Phylogeny, and Expression Analysis of the SWEET Gene Family in Tomato. Gene 2015, 573, 261–272. [Google Scholar] [CrossRef]
- Sah, S.K.; Reddy, K.R.; Li, J. Abscisic Acid and Abiotic Stress Tolerance in Crop Plants. Front Plant Sci 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Waterland, N.L.; Campbell, C.A.; Finer, J.J.; Jones, M.L. Abscisic Acid Application Enhances Drought Stress Tolerance in Bedding Plants. HortScience 2010, 45, 409–413. [Google Scholar] [CrossRef]
- Sun, S.; Hu, C.; Qi, X.; Chen, J.; Zhong, Y.; Muhammad, A.; Lin, M.; Fang, J. The AaCBF4-AaBAM3.1 Module Enhances Freezing Tolerance of Kiwifruit (Actinidia Arguta). Hortic Res 2021, 8, 97. [Google Scholar] [CrossRef] [PubMed]
- Ravinath, R.; Das, A.J.; Usha, T.; Ramesh, N.; Middha, S.K. Targeted Metagenome Sequencing Reveals the Abundance of Planctomycetes and Bacteroidetes in the Rhizosphere of Pomegranate. Arch Microbiol 2022, 204, 481. [Google Scholar] [CrossRef]
- Corbin, K.R.; Bolt, B.; Rodríguez López, C.M. Breeding for Beneficial Microbial Communities Using Epigenomics. Front Microbiol 2020, 11. [Google Scholar] [CrossRef]
- Hwarari, D.; Radani, Y.; Ke, Y.; Chen, J.; Yang, L. CRISPR/Cas Genome Editing in Plants: Mechanisms, Applications, and Overcoming Bottlenecks. Funct Integr Genomics 2024, 24, 50. [Google Scholar] [CrossRef]
- Pal, P.; Pal, S. CRISPR Genome Editing of Woody Trees: Current Status and Future Prospects. In CRISPRized Horticulture Crops; Elsevier, 2024; pp. 401–418. [Google Scholar]
Figure 1.
Abiotic stress due to climate change in key woody perennial fruit crops.

Figure 2.
A schematic view of the factors influencing response to stress, growth and plant-microbe interactions in woody fruit species. Soil properties, water and nutrient availability and the synergies of the genome the epigenome and the microbiome are crucial elements for proper plant growth and stress resilience. Beneficial microorganisms such as rhizosphere microbial communities are involved in many physiological and molecular processes and can promote plant growth, increase water and nutrient uptake, and enhance resilience to environmental stresses. These activities depend on genetic, epigenetic, microbial factors and soil physicochemical properties and entail intricate gene networks, expression programmes, protein and metabolite synthesis and epigenetic regulation such as DNA methylation, histone modifications, and the action of small noncoding RNAs.
Figure 2.
A schematic view of the factors influencing response to stress, growth and plant-microbe interactions in woody fruit species. Soil properties, water and nutrient availability and the synergies of the genome the epigenome and the microbiome are crucial elements for proper plant growth and stress resilience. Beneficial microorganisms such as rhizosphere microbial communities are involved in many physiological and molecular processes and can promote plant growth, increase water and nutrient uptake, and enhance resilience to environmental stresses. These activities depend on genetic, epigenetic, microbial factors and soil physicochemical properties and entail intricate gene networks, expression programmes, protein and metabolite synthesis and epigenetic regulation such as DNA methylation, histone modifications, and the action of small noncoding RNAs.
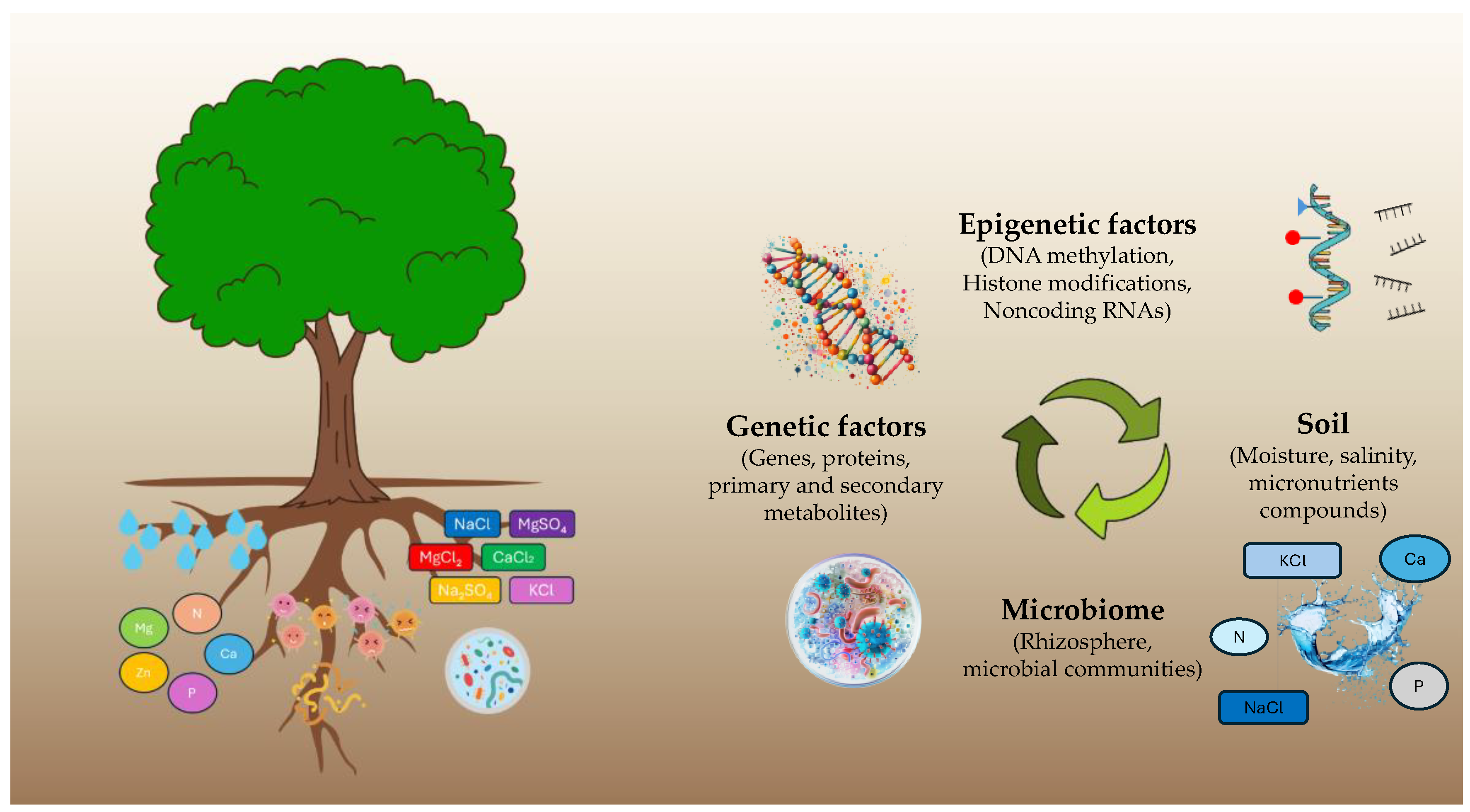
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



