Submitted:
11 February 2025
Posted:
12 February 2025
You are already at the latest version
Abstract
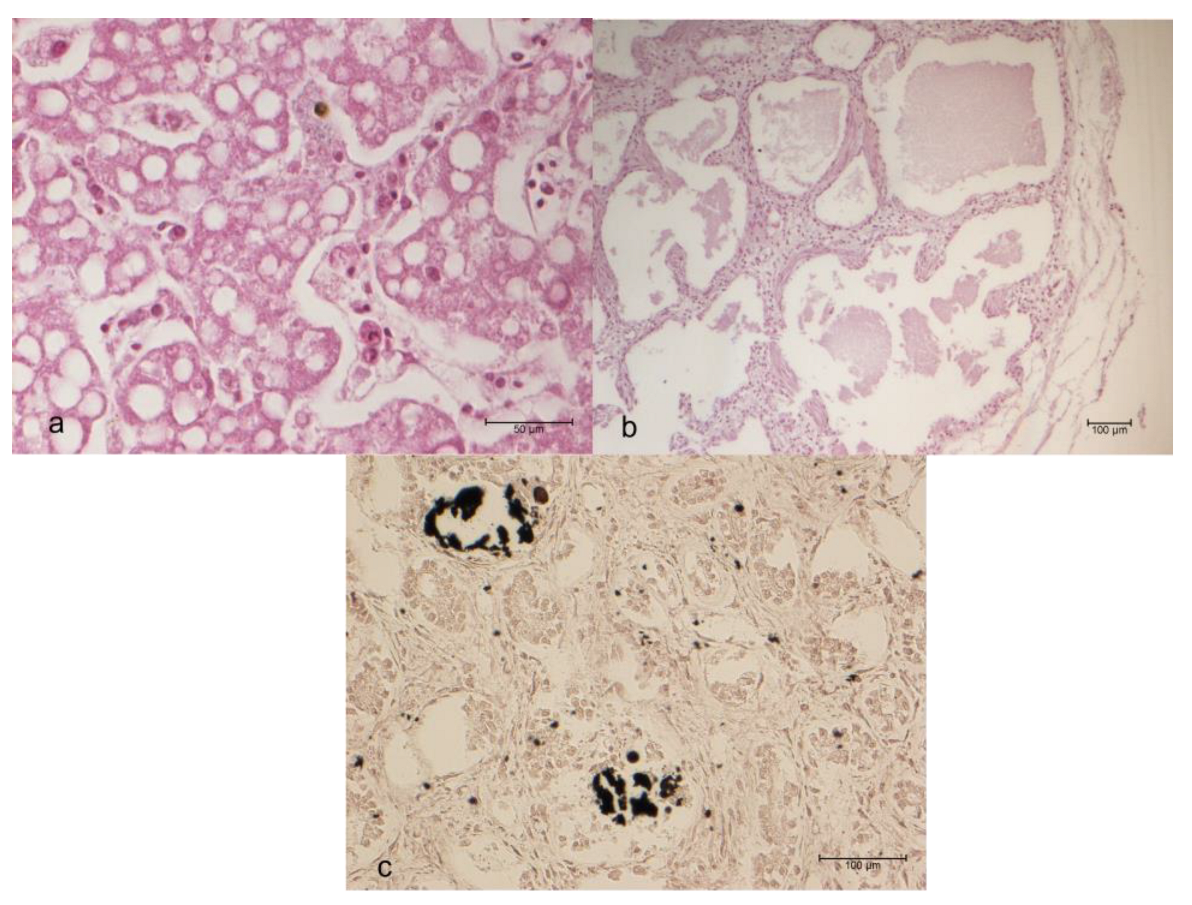
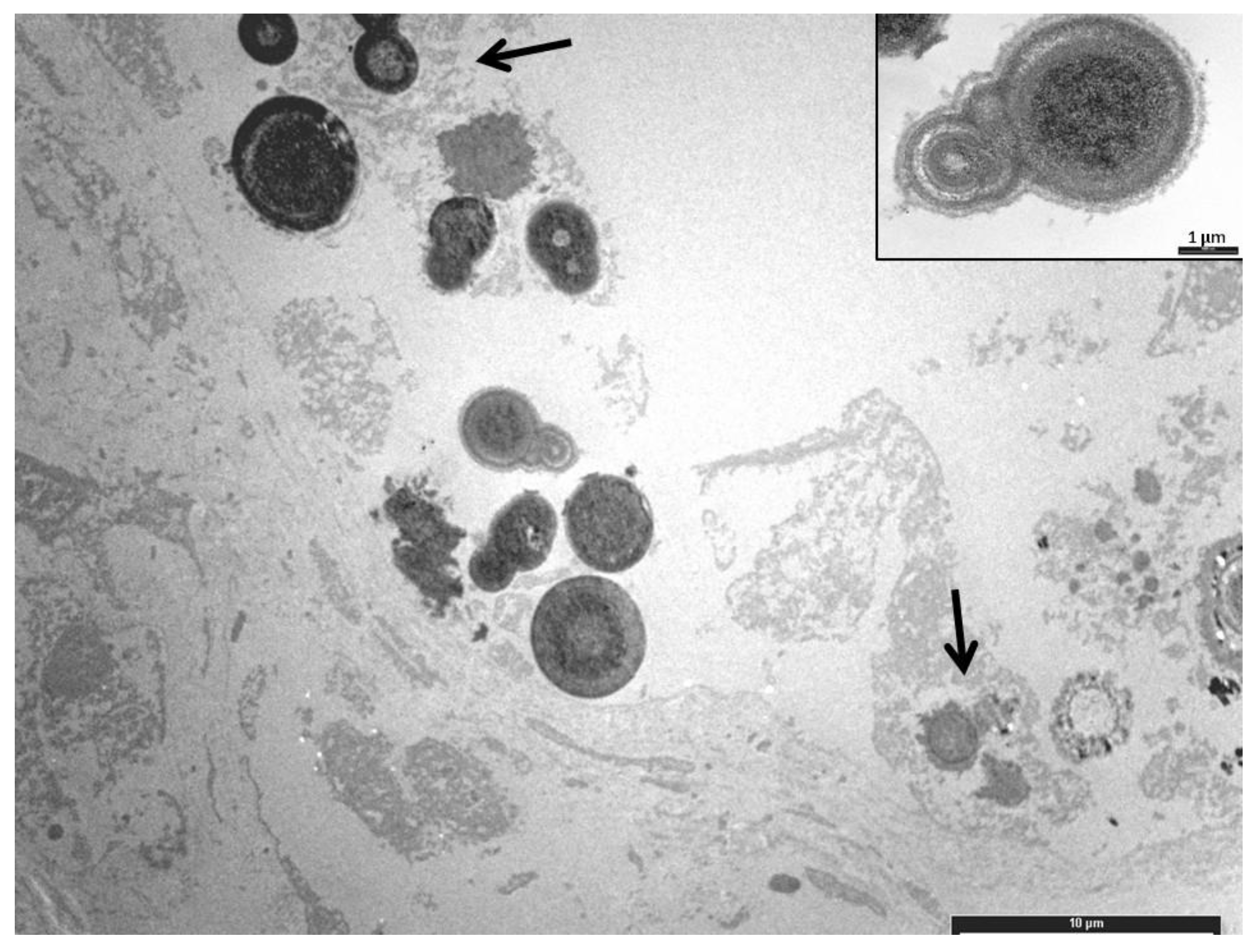
On the beach of Linosa Island (Italy), forty-three loggerhead sea turtle (Caretta caretta) unhatched eggs were recovered from nests, formalin-fixed and necropsied. The tissue samples were stained with Haematoxylin-Eosin (HE), Grocott, von Kossa, Periodic Ac-id-Schiff (PAS), and Movat pentachrome stains. Histologically, vacuolar degeneration (100.0%) and increased numbers of melanomacrophages (18.6%) in the liver, and edema (14.0%) in the lungs were observed. Twenty-five kidneys (58.1%) showed deposition of blue amorphous material with HE staining, which also appeared PAS-positive and black with von Kossa staining, allowing a diagnosis of calcium oxalate, confirmed by transmis-sion electron microscopy. The hepatic lesions may be indicative of toxicosis, infection, or a defense mechanism. A statistically significant association between the nest position and renal calculosis was observed. Renal calculosis was probably due to the exceptionally high summer temperatures, which were statistically higher compared to the temperatures recorded in the previous two years.
Keywords:
1. Introduction
2. Materials and Methods
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| IUCN | International Union for Conservation of Nature |
| SCL | Standard carapace length |
| SCW | standard carapace width |
| HL | head length |
| HW | head width |
| HE | Haematoxylin-Eosin stain |
| PAS | Periodic Acid Schiff |
| TEM | Transmission Electron Microscopy |
References
- Pritchard, P.; Mortimer, J. Taxonomy, External Morphology, and Species Identification. In Research and management techniques for the conservation of sea turtles.; SSC/IUCNN marine turtle specialist group.: Gland, 1999; pp. 21–38. [Google Scholar]
- Mingozzi, T.; Masciari, G.; Paolillo, G.; Pisani, B.; Russo, M.; Massolo, A. Discovery of a Regular Nesting Area of Loggerhead Turtle Caretta Caretta in Southern Italy: A New Perspective for National Conservation. 2006; 7. [Google Scholar] [CrossRef]
- Balletto, E.; Giacoma, C.; Paolillo, G.; Mari, F.; Dell’Anna, L. Piano d’azione per La Conservazione Della Tartaruga Marina Caretta Caretta Nelle Isole Pelagie. In; Edi. Tur. S.r.l.: Roma, 2003; pp. 1–62. [Google Scholar]
- Casale, P. Caretta Caretta (Mediterranean Subpopulation); IUCN Red List of Threatened Species Available online:. Available online: https://www.iucn.it/scheda.php?id=1108177324 (accessed on 7 December 2023).
- Casale, P. Sea Turtles in the Mediterranean : Distribution, Threats and Conservation Priorities; IUCN, 2010; ISBN 978-2-8317-1240-6.
- Casale, P.; Broderick, A.C.; Camiñas, J.A.; Cardona, L.; Carreras, C.; Demetropoulos, A.; Fuller, W.J.; Godley, B.J.; Hochscheid, S.; Kaska, Y.; et al. Mediterranean Sea Turtles: Current Knowledge and Priorities for Conservation and Research. Endangered Species Research 2018, 36, 229–267. [Google Scholar] [CrossRef]
- Depellegrin, D.; Menegon, S.; Farella, G.; Ghezzo, M.; Gissi, E.; Sarretta, A.; Venier, C.; Barbanti, A. Multi-Objective Spatial Tools to Inform Maritime Spatial Planning in the Adriatic Sea. Science of The Total Environment 2017, 609, 1627–1639. [Google Scholar] [CrossRef]
- Vlachogianni, T.; Fortibuoni, T.; Ronchi, F.; Zeri, C.; Mazziotti, C.; Tutman, P.; Varezić, D.B.; Palatinus, A.; Trdan, Š.; Peterlin, M.; et al. Marine Litter on the Beaches of the Adriatic and Ionian Seas: An Assessment of Their Abundance, Composition and Sources. Marine Pollution Bulletin 2018, 131, 745–756. [Google Scholar] [CrossRef] [PubMed]
- Santoro, M.; Hernández, G.; Caballero, M.; García, F. AEROBIC BACTERIAL FLORA OF NESTING GREEN TURTLES (CHELONIA MYDAS) FROM TORTUGUERO NATIONAL PARK, COSTA RICA. zamd 2006, 37, 549–552. [Google Scholar] [CrossRef] [PubMed]
- Durmuş, S.H.; Ilgaz, Ç.; Gü çlü, Ö.; Özdemir, A. The Effect of the Predicted Air Temperature Change on Incubation Temperature, Incubation Duration, Sex Ratio and Hatching Success of Loggerhead Turtles. Animal Biology 2011, 61, 369–383. [Google Scholar] [CrossRef]
- Caracappa, S.; Persichetti, M.F.; Piazza, A.; Caracappa, G.; Gentile, A.; Marineo, S.; Crucitti, D.; Arculeo, M. Incidental Catch of Loggerhead Sea Turtles (Caretta Caretta) along the Sicilian Coasts by Longline Fishery. PeerJ 2018, 6, e5392. [Google Scholar] [CrossRef]
- Gleason, F.H.; Allerstorfer, M.; Lilje, O. Newly Emerging Diseases of Marine Turtles, Especially Sea Turtle Egg Fusariosis (SEFT), Caused by Species in the Fusarium Solani Complex (FSSC). Mycology 2020, 11, 184–194. [Google Scholar] [CrossRef]
- Risoli, S.; Sarrocco, S.; Terracciano, G.; Papetti, L.; Baroncelli, R.; Nali, C. Isolation and Characterization of Fusarium Spp. From Unhatched Eggs of Caretta Caretta in Tuscany (Italy). Fungal Biology 2023, 127, 1321–1327. [Google Scholar] [CrossRef]
- Esposito, M.; Canzanella, S.; Iaccarino, D.; Pepe, A.; Di Nocera, F.; Bruno, T.; Marigliano, L.; Sansone, D.; Hochscheid, S.; Gallo, P.; et al. Trace Elements and Persistent Organic Pollutants in Unhatched Loggerhead Turtle Eggs from an Emerging Nesting Site along the Southwestern Coasts of Italy, Western Mediterranean Sea. Animals 2023, 13, 1075. [Google Scholar] [CrossRef]
- Chemello, G.; Trotta, E.; Notarstefano, V.; Papetti, L.; Di Renzo, L.; Matiddi, M.; Silvestri, C.; Carnevali, O.; Gioacchini, G. Microplastics Evidence in Yolk and Liver of Loggerhead Sea Turtles (Caretta Caretta), a Pilot Study. Environmental Pollution 2023, 337, 122589. [Google Scholar] [CrossRef]
- Duncan, E.M.; Broderick, A.C.; Fuller, W.J.; Galloway, T.S.; Godfrey, M.H.; Hamann, M.; Limpus, C.J.; Lindeque, P.K.; Mayes, A.G.; Omeyer, L.C.M.; et al. Microplastic Ingestion Ubiquitous in Marine Turtles. Global Change Biology 2019, 25, 744–752. [Google Scholar] [CrossRef] [PubMed]
- Pietroluongo, G.; Centelleghe, C.; Sciancalepore, G.; Ceolotto, L.; Danesi, P.; Pedrotti, D.; Mazzariol, S. Environmental and Pathological Factors Affecting the Hatching Success of the Two Northernmost Loggerhead Sea Turtle (Caretta Caretta) Nests. Sci Rep 2023, 13, 2938. [Google Scholar] [CrossRef]
- Jensen, M.P.; Allen, C.D.; Eguchi, T.; Bell, I.P.; LaCasella, E.L.; Hilton, W.A.; Hof, C.A.M.; Dutton, P.H. Environmental Warming and Feminization of One of the Largest Sea Turtle Populations in the World. Current Biology 2018, 28, 154–159e4. [Google Scholar] [CrossRef]
- Hall, J.M.; Sun, B. Heat Tolerance of Reptile Embryos: Current Knowledge, Methodological Considerations, and Future Directions. Journal of Experimental Zoology Part A: Ecological and Integrative Physiology 2021, 335, 45–58. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.D.; Mortimer, J.A.; Limpus, C.J. A Field Key to the Developmental Stages of Marine Turtles (Cheloniidae) with Notes on the Development of Dermochelys. ccab 2017, 16, 111–122. [Google Scholar] [CrossRef]
- Wyneken, J. The Anatomy of Sea Turtles; Technical Memorandum.; Department of Commerce, NOAA, U.S.: Miami (FL), 2001. [Google Scholar]
- Merendi, F.; Zaccaroni, A.; Zucchini, M.; Affronte, M.; Scaravelli, D.; Simoni, P. Rilievi Necroscopici, Esami Citologici, Istologici e Determinazione Dei Metalli Pesanti e Arsenico in Tartarughe Marine (Caretta Caretta) Spiaggiate in Emilia Romagna. In Proceedings of the Proceedings of the 1th annual congress of the Italian association of pathologists.
- Prearo, M.; Squadrone, S.; Tarasco, R.; Zizzo, N.; Appino, S.; Pavoletti, E.; Abete, M. Presenza Di Metalli Pesanti in Tartarughe Comuni (Caretta Caretta) Spiaggiate Lungo Le Coste Pugliesi. In Proceedings of the Proceedings of the 11th annual congress of Italian Society of Veterinary Diagnostic Laboratory.
- Storelli, M.M.; Marcotrigiano, G.O. Chlorobiphenyls, HCB, and Organochlorine Pesticides in Some Tissues of Caretta Caretta (Linnaeus) Specimens Beached along the Adriatic Sea, Italy. Bull Environ Contam Toxicol 2000, 64, 481–488. [Google Scholar] [CrossRef]
- Alam, S.K.; Brim, M.S. Organochlorine, PCB, PAH, and Metal Concentrations in Eggs of Loggerhead Sea Turtles (Caretta Caretta) from Northwest Florida, USA. J Environ Sci Health B 2000, 35, 705–724. [Google Scholar] [CrossRef]
- van de Merwe, J.P.; Hodge, M.; Whittier, J.M.; Ibrahim, K.; Lee, S.Y. Persistent Organic Pollutants in the Green Sea Turtle <em>Chelonia Mydas</Em>: Nesting Population Variation, Maternal Transfer, and Effects on Development. Marine Ecology Progress Series 2010, 403, 269–278. [Google Scholar]
- Stewart, K.R.; Keller, J.M.; Templeton, R.; Kucklick, J.R.; Johnson, C. Monitoring Persistent Organic Pollutants in Leatherback Turtles (Dermochelys Coriacea) Confirms Maternal Transfer. Marine Pollution Bulletin 2011, 62, 1396–1409. [Google Scholar] [CrossRef]
- Cañas, J.E.; Anderson, T.A. Organochlorine Contaminants in Eggs: The Influence of Contaminated Nest Material. Chemosphere 2002, 47, 585. [Google Scholar] [CrossRef]
- Johnson, J.C.; Schwiesow, T.; Ekwall, A.K.; Christiansen, J.L. Reptilian Melanomacrophages Function under Conditions of Hypothermia: Observations on Phagocytic Behavior. Pigment Cell Research 1999, 12, 376–382. [Google Scholar] [CrossRef] [PubMed]
- Agius, C.; Roberts, R.J. Melano-Macrophage Centres and Their Role in Fish Pathology. Journal of Fish Diseases 2003, 26, 499–509. [Google Scholar] [CrossRef]
- Mader, D. Reptile Medicine and Surgery. In; Saunders Elsevier: Philadelphia, 2006. [Google Scholar]
- Frye, F. Biomedical and Surgical Aspects of Captive Reptile Husbandry.; Krieger publishing company.: Malabar, 1991. [Google Scholar]
- Miller, J. Embryology of Marine Turtles. In Biology of the reptilia.; Wiley-Interscience: Hoboken, 1985; pp. 269–328. [Google Scholar]
- Lutz, P.; Musick, J.; Wyneken, J. The Biology of Sea Turtles.; Crc press: London, 1997. [Google Scholar]
- Ackerman, R. The Nest Environment and the Embryonic Development of Sea Turtles. In The biology of sea turtles.; Crc press: London, 1997; pp. 83–106. [Google Scholar]
- Packard, G.; Packard, M. The Physiological Ecology of Reptilian Eggs and Embryos. In Biology of the reptilia.; Academic press.: Cambridge, 1988; pp. 523–605. [Google Scholar]
- Moran, K.; Bjorndal, K.; Bolten, A. Effects of the Thermal Environment on the Temporal Pattern of Emergence of Hatchling Loggerhead Turtles Caretta Caretta. Mar. Ecol. Prog. Ser. 1999, 189, 251–261. [Google Scholar] [CrossRef]
- Matsuzawa, Y.; Sato, K.; Sakamoto, W.; Bjorndal, K. Seasonal Fluctuations in Sand Temperature: Effects on the Incubation Period and Mortality of Loggerhead Sea Turtle (Caretta Caretta) Pre-Emergent Hatchlings in Minabe, Japan. Marine Biology 2002, 140, 639–646. [Google Scholar] [CrossRef]
- Davenport, J. Temperature and the Life-History Strategies of Sea Turtles. Journal of Thermal Biology 1997, 22, 479–488. [Google Scholar] [CrossRef]

| Depth | Nest 1 | Nest 2 | Nest 3 | Nest 4 | Nest 5 |
|---|---|---|---|---|---|
| 13-18 cm | - | - | 1 | - | - |
| 19-23 cm | 3 | - | - | - | 1 |
| 24-28 cm | 3 | - | 8 | 3 | - |
| 29-33 cm | 6 | 2 | 3 | 1 | - |
| 34-38 cm | - | 3 | 1 | 2 | - |
| 39-43 cm | - | 6 | - | - | - |
| TOTAL | 12 | 11 | 13 | 6 | 1 |
| JULY | AUGUST | ||
| 2 years before | 25.7°C | 2 years before | 26.1°C |
| 1 year before | 25.8°C | 1 year before | 25.8°C |
| Sampling year | 30.2°C | Sampling year | 30.7°C |
| 1 year after | 25.7°C | 1 year after | 27.2°C |
| 2 years after | 26.8°C | 2 years after | 27.1°C |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).



