Submitted:
04 February 2025
Posted:
05 February 2025
You are already at the latest version
Abstract
Continental rift zones on the early Earth provided essential conditions for the emergence of the first cells. These conditions included an abundant supply of raw materials, cyclic fluctuations in pressure and temperature over millions of years, and transitions of gases between supercritical and subcritical phases. While evidence supports vesicle formation and the chemical evolution of peptides, the mechanism by which information was stored remains unresolved. This study pro-poses a model illustrating how interactions among organic molecules may have enabled the encoding of amino acid sequences in RNA. The model highlights the interplay between three key molecular components: a proto-tRNA, the vesicle membrane, and short peptides. The vesicle membrane acted as a reservoir for hydrophobic amino acids and facilitated their attachment to proto-tRNA. As a single strand, proto-tRNA also served as proto-mRNA, enabling it to be read by charged tRNAs. Through the replication of this information and the assembly of RNA strands, the first catalytically active peptides could form, thereby enhancing the long-term sta-bility of vesicles. This model further outlines how these vesicles may have evolved into the ear-liest cells, with enzymes and larger RNA molecules giving rise to tRNA and ribosomal struc-tures. Shearing forces may have facilitated the first cellular divisions, representing a pre-LUCA stage.
Keywords:
origin of life
; hydrothermal biochemistry
; information storage
; continental crust model
; super-critical fluids
; pre-LUCA stage
1. Introduction
The origin of life remains one of the greatest unsolved mysteries, with the precise environment for the emergence of the first cell yet to be conclusively identified. Fundamental questions persist: where did the building blocks of life form in sufficient abundance, and how did they concentrate into reactive mixtures [1]? What energy sources drove the critical chemical reactions for life, and how did these processes sustain life despite the constraints of increasing entropy [2,3]? How was the high thermodynamic barrier for peptide condensation in aqueous solutions—commonly referred to as the "water paradox"—overcome [4,5]? If life began on Earth’s surface, what roles did solar wind and ultraviolet radiation play [6]? Furthermore, what impact did meteorite bombardment, erosion, and sedimentation have on ponds that might have existed over millions of years, fostering the conditions necessary for life [7]? What were the prevailing temperatures on early Earth's surface, and how did they influence cloud formation and precipitation patterns—key factors for sustaining dry-wet cycles? How did vesicles evolve as precursors to cells, and how did complex molecular sequences, which could not have arisen by chance, come into existence [8,9,10]? What accounts for the handedness of chiral molecules in living systems [11,12]? Lastly, how did information storage originate during the earliest stages of life [13]?
Until recently, no widely accepted model described a realistic, experimentally testable environment for the origin of life, despite the necessity of such a model to advance our understanding of the peptide/RNA system. Two primary models have dominated the literature: the hydrothermal vent ("black and white smokers") model [14] and the "warm little ponds" model, which incorporates both hydrothermal inputs and extra-terrestrial organic compounds [15,16,17]. However, both models face significant challenges. Black smokers exhibit extreme temperatures and high concentrations of metal compounds unsuitable for life, while modern white smokers are often associated with calcareous sediments from biogenic reefs, conditions unlikely to have existed on early Earth [7,18]. The "warm little ponds" model is similarly fraught with difficulties, such as exposure to ultraviolet radiation, solar wind particles, rapid sediment or salt accumulation, and the potential destruction of ponds by meteorite impacts, which could cause catastrophic flooding and erosion. Additionally, no reliable estimate exists for the volume of organic material delivered to such ponds by meteorites, as organic concentrations in meteorites are low, with weathering releasing only trace amounts over extended periods [19].
Recent studies on early continental crust environments and the origin of life propose a novel model that can be tested both in the laboratory and in nature [20,21,22]. This model centers on the evolution of the continental crust, which is thought to have formed soon after Earth’s surface cooled. By the time life is believed to have emerged [23], the continental crust may have already constituted over 20% of its current mass [24]. Tectonic stresses likely created fault zones in the early continental crust, facilitating the release of magma and gases to the surface. These fault zones may have remained active for tens to hundreds of millions of years. Volcanic regions would have exhibited a range of temperatures, with cooler zones influenced by artesian water flows, potentially giving rise to cold-water geysers driven by CO₂ and N₂ gases. Such conditions in cold-water geysers can be reproduced in high-pressure laboratory experiments [21,22]. Additionally, the chemical composition of fluids from these fault zones can be studied through fluid inclusions in hydrothermally formed fissure minerals [25,26]. This approach offers new insights into the conditions that may have fostered the origin of life.
2. Environment and Resources
Tectonic faults in the upper continental crust, filled with water and gases, presented a diverse range of pressure and temperature conditions (Figure 1) [20]. Processes such as Haber-Bosch and Fischer-Tropsch syntheses may have produced life’s building blocks from abiotic materials, utilizing various metallic catalysts naturally available in these environments. These conditions closely resemble the early atmospheric conditions studied by Peters et al. (2023), which involved similar gas partial pressures and iron compounds derived from volcanic ash and meteorites acting as catalysts [27]. Today, subsurface ecosystems influenced by geological degassing represent microbial hotspots within the deep biosphere [28]. Precursors to lipid molecules, such as aldehydes, have been identified in ancient hydrothermal quartz from Western Australia and, more recently, in Quaternary volcanic calcites in the Eifel region [25,26].
Hennet et al. demonstrated that amino acids could form under hydrothermal conditions (150°C, 10 atm, pH 7) with mineral catalysts and starting compounds like formaldehyde, ammonia, and cyanide [29]. Glycine was detected in the highest concentrations, with other amino acids (aspartic acid, serine, glutamic acid, alanine, and isoleucine) present at concentrations roughly two orders of magnitude lower. Marshall [30] extended this work by identifying 12 amino acids formed under hydrothermal conditions, including proline, leucine, lysine, valine, threonine, and phenylalanine. These results suggest that tectonic fault zones could have supplied at least 12 amino acids crucial for early peptide synthesis, although their concentrations likely varied. Furthermore, LaRowe and Regnier demonstrated the synthesis of five nucleobases (adenine, cytosine, guanine, thymine, and uracil) as well as ribose and deoxyribose under hydrothermal conditions typical of such environments [31].
Fluids in crustal faults often exhibit low pH (<3) due to high pressures where CO₂ dominates the gas phase [32]. Sulfur compounds can further decrease pH, while nitrogen tends to increase it. Geyser eruptions can cause transient pH increases of up to 0.5 due to rapid pressure changes. High pressure, temperature, and low pH promote the alteration of marginal rocks, releasing phosphates from apatite, a mineral abundant in igneous rocks. Cations from rock-forming minerals can act as catalysts for organic reactions or form new reactive surfaces, such as sulfide ore veins, carbonates, or arsenites on fault walls. Sulfide ores, often containing radioactive elements, may have contributed to RNA evolution and reaction networks [33].
Clay minerals lining fault surfaces could have served as templates for nucleotide linkage, a critical step in RNA formation [34,35]. Quartz, commonly found in fault fissures, is often accompanied by colloidal SiO₂-rich fluids that, like quartz surfaces, exhibit catalytic properties [36]. Additionally, during the early history of Earth, stronger tidal forces from a closer Moon influenced crustal activity [37]. While piezoelectric effects in crustal rocks typically cancel out during seismic events due to random mineral distribution, they can create regular voltage gradients within hydrothermally formed quartz in faults. These gradients could have generated weak cyclical electrical currents, and voltage spikes associated with seismic activity may have caused lightning-like discharges in gas-filled cavities or facilitated the splitting of water into H₂ and O₂ [38].
The stable conditions deep within Earth’s crust, shielded from solar wind, UV radiation, surface erosion, and flooding, could have persisted for millions of years. Organic molecules from surface environments may have been transported to these depths via artesian water flow and geyser backflow, potentially delivering meteoritic organic material. CO₂ and N₂ gases in geysers played a crucial role in this context. The upward flow of gas bubbles facilitated substance transport, continuously replenishing reactants and removing reaction products. This dynamic prevented equilibrium conditions and reduced tar formation, which could otherwise inhibit further reactions [39]. Hydrophobic and amphiphilic molecules may have been carried within supercritical gas bubbles, while amphiphilic molecules associated with hydrophilic compounds could have been transported upwards, in a process resembling flotation.
Pure CO₂, with a critical point of 30.98°C and 73.77 bar, enters a supercritical state (scCO₂) at depths of around 1000 m, depending on temperature and the density of gas bubbles in the overlying water column. Similarly, pure N₂, with a critical point of -146.9°C and 33.96 bar, achieves supercriticality at approximately 400 m in an open water column. Mixtures of scCO₂ and scN₂ exhibit intermediate critical points depending on their respective proportions, a principle that also applies to trace gases such as NH₃, CH₄, and SO₂ when present in sufficient concentrations.
Under these conditions, supercritical gases function as non-polar solvents, enabling the accumulation and interaction of organic molecules. Cavities where scCO₂/scN₂ fills the upper region and water occupies the lower region act as autoclave-like reaction chambers with distinct two-phase boundaries. In such environments, amino acids can condense into peptides, releasing water as a by-product. This process is enhanced by pressure drops during cyclic geyser eruptions, which trigger local phase transitions from scCO₂ to subcritical CO₂ (gCO₂). The accompanying entropy increase during these phase transitions facilitates peptide chain elongation without requiring catalysts [21]. Organic molecules transported from deeper crustal regions via supercritical gas bubbles would likely accumulate here due to reduced solubility in subcritical gases.
3. The First RNA: A Precursor to tRNA?
The formation of RNA under hydrothermal conditions remains unproven, but thermodynamic calculations suggest that RNA building blocks—adenine, guanine, cytosine, thymine, uracil, ribose, and deoxyribose—could synthesize from precursors such as CH₂O and HCN under the temperature, pressure, and fluid composition characteristic of hydrothermal systems [31]. Given that these RNA components could form within the upper continental crust, the basic prerequisites for an RNA-based information storage system may have existed. However, as a random product of chemical reactions, RNA would not initially have served as an information storage system akin to modern mRNA or DNA. Instead, early RNA may have had an autocatalytic function, acting as a ribozyme [40,41,42].
RNA takes on a unique role if viewed as a precursor to tRNA rather than as a direct informational molecule like mRNA. RNA is most stable in acidic conditions, such as those found in low-temperature hydrothermal waters within tectonic faults [43]. Under these conditions, RNA remains protonated at low pH levels. However, when scN₂ dominates the gas phase, protonation is avoided, allowing the formation of longer RNA strands. Even so, RNA strands tend to form duplexes with complementary sequences, hindering replication, as duplex structures prevent the incorporation of free nucleotides. Single-stranded RNA would eventually transition into duplexes, rendering them non-replicable and effectively forming a dead-end product [44].
Fault zones containing low-temperature hydrothermal water with excess N₂ (and possibly CO₂) offer optimal conditions for RNA replication. At depths below 400 m, temperatures on early Earth could have exceeded 50°C due to surface water infiltration from artesian aquifers. During geyser eruptions, the phase change from supercritical to gaseous N₂/CO₂ induces cooling due to gas expansion (Joule-Thomson effect). When water refills the fault and compresses the gas, temperatures rise, restoring supercritical conditions. These cycles, with peak temperatures surpassing 60°C, could separate RNA duplexes, enabling replication to proceed with available building blocks once cooling occurs. The lower melting point of RNA compared to DNA may explain DNA's absence during early cellular evolution. Early RNA strands likely differed significantly from their modern counterparts, potentially incorporating additional organic bases and sugars in both D- and L-enantiomeric forms.
Even if enantiomerically pure RNA formed, this alone would not guarantee the evolution of a robust information system. Longer strands often develop complementary base pairings between the first and last thirds, folding into loops. In modern tRNA, these loops form structures where three outward-facing bases act as anticodons that bind complementary nucleotides. A similar structure could have arisen during early chemical evolution, serving as a precursor to tRNA. In modern cells, tRNA translates specific amino acids into the genetic code. Proto-tRNAs may have played a comparable role. For such a function, the end opposite the anticodon would need to remain single-stranded, analogous to the acceptor arm of modern tRNA, which carries the conserved base sequence CCA.
4. The Formation of Vesicles
In laboratory experiments, the conditions of cold water geyser systems were recreated within a high-pressure cell, where water, CO₂, lipids, and amino acids were subjected to cyclic pressure fluctuations. This experimental setup allowed for the demonstration of vesicle formation and the chemical evolution of peptides [9,21]. The experiments focused on the transition zone from supercritical CO₂ (scCO₂) to gas at depths around 1,000 meters, occasionally incorporating varying proportions of nitrogen (N₂) at shallower depths. The process unfolds as follows: along fracture edges, numerous cavities form on fault surfaces, facilitating the accumulation of CO₂. These cavities act as reaction chambers, with water occupying the lower section and scCO₂ or gCO₂ residing in the upper section.
When scCO₂ is present, a decrease in pressure during a geyser eruption triggers a phase transition in the transition zone, producing gCO₂, in which dissolved water condenses into mist. Experimental evidence suggests that this condensation facilitates vesicle formation, as lipids from scCO₂, which cannot remain in the gas phase, accumulate to form a primary envelope on the surface of the mist droplets. As these droplets sink towards the water-gas interface, where lipids are also present, they undergo further coating, ultimately forming vesicles with bilayer membranes in the aqueous phase. During the pressure drop, amino acids link to form peptides, which can interact with the vesicle membranes. Mayer et al. [21] demonstrated that repeated pressure fluctuation cycles can drive chemical evolution, leading to the mutual stabilization of peptides and vesicles [9,22]. However, the vast diversity of potential peptide combinations poses a challenge to the formation of identical amino acid sequences, representing a key limitation of this process. Importantly, no mechanism exists to store sequence information, such as that provided by RNA or DNA.
A secondary vesicle formation process occurs in the gas phase alongside mist formation. During pressure drops in a geyser eruption, dissolved gas in the water is released, resulting in an eruption so vigorous that, similar to the opening of a champagne bottle, it induces turbulence and forms foam within the cavities. In this case, droplets containing lipids from the water-gas interface are propelled into the gas phase. The initial lipid envelope forms around these droplets, which then receive a secondary coating upon re-entering the interface. Notable distinctions exist between the two vesicle formation mechanisms. While mist droplets contain quasi-distilled water (with regard to dissolved inorganic substances) and forcibly absorb organic molecules from previously supercritical gas, "champagne droplets" contain salts from the fluid and organic molecules that are readily water-soluble. Additionally, the droplet sizes differ significantly, with mist droplets measuring a few microns in diameter and "champagne droplets" being approximately two orders of magnitude larger.
5. The Hypothetical Model for Storing Information
The model proposed here suggests that the development of an information storage system requires the presence of a proto-tRNA composed of only 12 nucleobases. This proto-tRNA would involve a single base pair, with a single strand featuring the nucleobases CCA at one end, and a loop at the other, where three bases rotate outward to form an anticodon (Figure 2).
During vesicle formation at depths exceeding 400 meters, hydrothermally generated hydrophobic amino acids can form short peptides that integrate into the vesicle membrane, while hydrophilic amino acids remain in the aqueous phase. Glycine, as a neutral amino acid, can reside in either the water or scCO₂/scN₂ environments, depending on the specific physicochemical conditions. Given the hydrophobic nature of adenine, the single strand of the proposed proto-tRNA could embed itself within the membrane, allowing interaction with the membrane-associated hydrophobic amino acids (Figure 3). This arrangement would facilitate linkage at the 2'-OH position of the terminal ribose, occurring cyclically within the geyser system.
A critical question is how a specific bond between the corresponding amino acid and the anticodon’s base triplet could be formed. The base triplet, which ultimately defines the code, consists of three out of four possible bases, each exhibiting distinct hydrophobic or hydrophilic properties. As each base and its position within the triplet vary in hydrophobicity, they produce differing entropic forces due to the hydrophobic effect, which influences the acceptor arm’s penetration depth into the membrane. Thus, greater hydrophobicity within the anticodon region would drive the proto-tRNA’s tip deeper into the membrane, where it encounters the most hydrophobic amino acids, likely located in the innermost zone of the membrane. It is hypothesized that these amino acids were the initial constituents of short peptides and detached from the membrane as they bonded to the proto-tRNA.
Fine-tuned positioning of the CCA arm within the membrane could arise from various base combinations, allowing distinct hydrophobic amino acids to attach at specific membrane depths. When more hydrophilic bases are present within the anticodon, the CCA arm does not penetrate as deeply into the membrane, facilitating the attachment of hydrophilic amino acids from the water side at the 3'-OH position. If the terminal ribose aligns with the lipid heads at the inner edge of the membrane, no amino acid binds, corresponding to a stop codon.
Figure 3.
Variations in the penetration depths of the CCA acceptor arm of a proto-tRNA into a vesicle membrane (depicted in green) relative to the hydrophobicity of the anticodon. The first amino acid in each peptide sequence attaches to the CCA arm while simultaneously detaching from the peptide chain. Hydrophobic amino acids are represented by yellow and red dots, while hydrophilic amino acids are depicted as blue and purple dots. The nucleobases A (adenine), G (guanine), C (cytosine), and U (uracil) are shown.
Figure 3.
Variations in the penetration depths of the CCA acceptor arm of a proto-tRNA into a vesicle membrane (depicted in green) relative to the hydrophobicity of the anticodon. The first amino acid in each peptide sequence attaches to the CCA arm while simultaneously detaching from the peptide chain. Hydrophobic amino acids are represented by yellow and red dots, while hydrophilic amino acids are depicted as blue and purple dots. The nucleobases A (adenine), G (guanine), C (cytosine), and U (uracil) are shown.
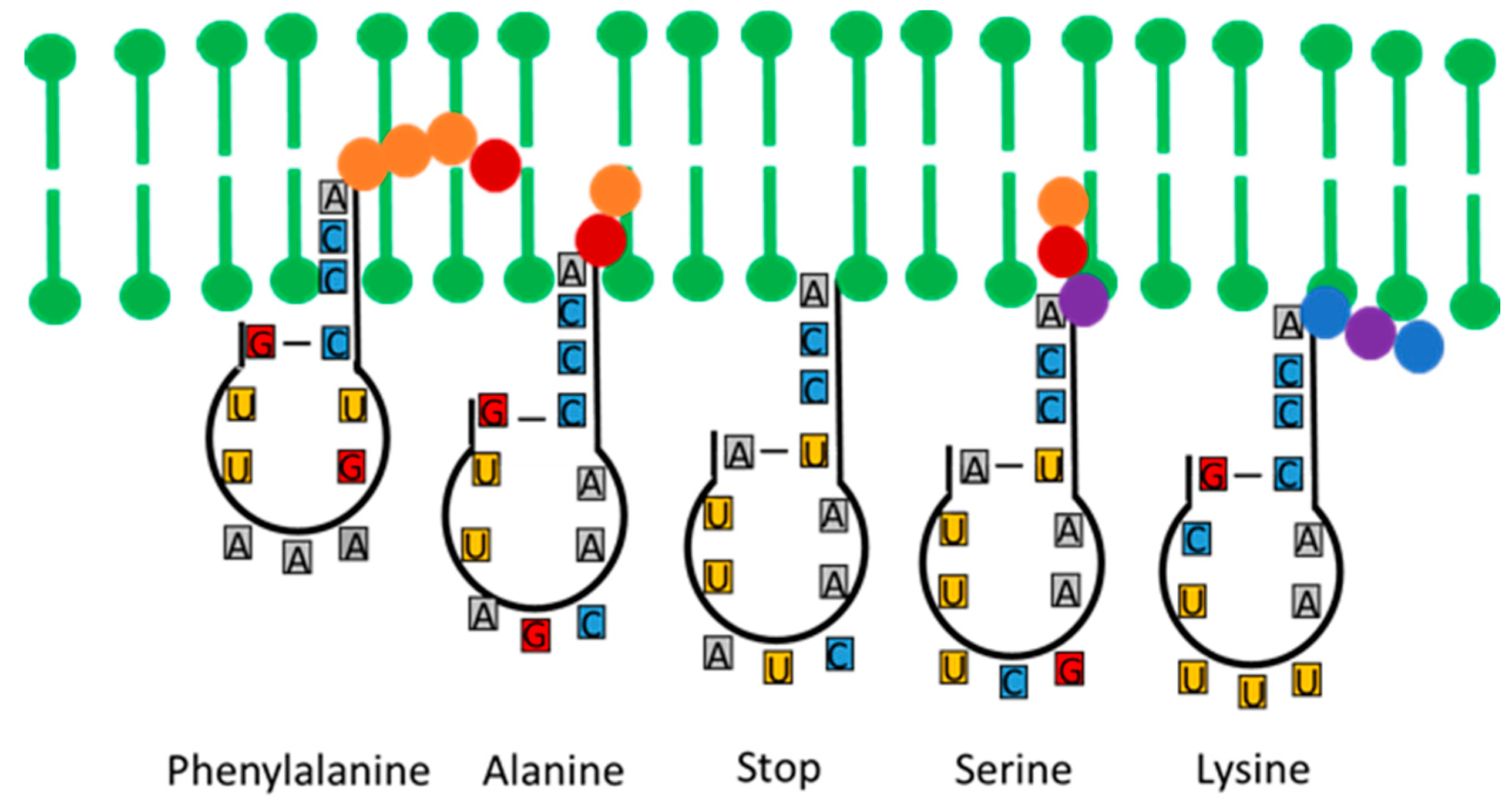
This model allows an important conclusion: If the hydrophobicity of the nucleobases played a role in the selection of amino acids, then there must be a systematic correlation between these two groups of molecules in the genetic code with regard to their physicochemical properties. Figure 4 illustrates this relationship, based on Jungck's [45] findings, particularly highlighting the correlation among amino acids that can form hydrothermally. This relationship underscores the potential evolutionary role of physicochemical properties in the development of the genetic code.
6. Next Evolutionary Step: Repurposing tRNA as mRNA
The only double-stranded region of the proto-tRNA readily dissociates under increased temperatures or turbulence typical of eruptive cycles. Single strands formed this way could then be replicated, acting as proto-mRNA templates. After linking with complementary, weakly specifically loaded proto-tRNAs and the subsequent linking of amino acids, the first short peptide chains (tetrapeptides) would be formed, the sequences of which would be preserved in the RNA. Initially, many proto-tRNA templates may not have contributed directly to peptide synthesis, as they encoded amino acids that were absent in the environment. However, specific proto-mRNA strands could sporadically enable peptide formation, suggesting that certain sequences, under appropriate conditions, could yield functional peptides, marking an early step in protein evolution.
Given that proto-tRNA must be homochiral (existing in both D- and L-forms), a preferential linkage emerges between L-amino acids and D-ribose in RNA (and vice versa) [46]. Consequently, the peptides formed were also homochiral, existing in equal amounts of both enantiomers. With a maximum of 12 amino acids potentially linking specifically to corresponding proto-tRNAs, at least an equal number of tetrapeptide templates would have existed, likely many more. The fusion of two same-handedness tetrapeptides could yield chains long enough to traverse membrane thickness, potentially forming pores crucial for ion and molecule exchange and acting as early energy generators. Since these peptide sequences were storable, evolutionary development shifted towards combining tetrapeptide chains or proto-mRNA templates, fostering the emergence of functional molecules with catalytic properties. If these molecules supported the synthesis of further functional molecules with the same chirality, they helped to displace the other chirality in the fight for resources. In this way, their own chirality finally prevailed. At the same time, they formed the basis for the formation of more complex peptides, so that the origin of life can be defined at this point in time.
Through millions of years, peptides with novel catalytic functions emerged, enhancing reactions that stabilized vesicle formation. At a certain evolutionary point, biochemical selection enabled cells to replicate essential molecules using external building blocks. When a protocell reached sufficient size, flow-induced shear forces triggered division, allowing daughter cells to independently thrive, provided that the mother cell had accumulated surplus resources. This process led to a population of ancestor cells with distinct (quantitative) compositions, all operating with a unified “language and grammar” in molecular replication and interaction.
7. Conclusions Drawn from Contemporary Biochemistry
In modern biochemistry, two distinct classes of aminoacyl-tRNA synthetases catalyze the specific attachment of amino acids to tRNAs [47,48]. Class I synthetases, which attach amino acids at the 2'-OH end of the terminal ribose, are generally associated with larger, less polar amino acids, while Class II synthetases target the 3'-OH end, attaching smaller, more polar amino acids. Biochemical and bioinformatic analyses, alongside protein engineering studies, suggest that these two synthetase classes may have evolved from complementary strands of a common ancestral gene [47]. This proposition aligns with the hypothesis that modern aminoacyl-tRNA synthetases trace their origins back to early molecular evolution events, specifically to a template-driven mechanism akin to the proto-tRNA formation described here. The peptides synthesized on proto-tRNA and its complementary strand may later have been encoded in DNA using bidirectional genetic coding in a sense-antisense manner.
8. Summary
The ancient continental crust provided an optimal setting for the emergence of early life, offering a stable, sheltered environment that persisted over millions of years. This habitat was rich in essential resources and offered diverse conditions conducive to organic molecule formation, such as variations in pressure, temperature, and pH, alongside natural catalysts. The crust’s structure allowed for continuous substance exchange as an open system. Supercritical gases dissolved nonpolar compounds, thereby facilitating organic synthesis and polymer formation. The cyclic phase transitions induced by cold-water geysers generated entropy fluctuations, promoting chemical reactions without the need for activation energy. Though short-lived, vesicles were readily formed, enabling the spontaneous linkage of molecular chains driven by entropic forces. Temperature variations enabled RNA replication, while vesicle membranes supported interactions with organic molecules, including proto-tRNA. Acting as molecular bridges, proto-tRNAs linked specific amino acids and conveyed information, simultaneously establishing the precursors to mRNA. In summary, the development of life was fundamentally propelled by the dynamic interaction of three critical components: vesicles, peptides, and RNA. These interactions set the stage for the evolution of life, where entropic forces and cyclic environmental conditions drove the emergence of complex molecular systems.
Funding
This research received no external funding
Acknowledgments
I am grateful to Christian Mayer (University of Duisburg-Essen) for the extensive discussions and helpful comments.
Conflicts of Interest
The author declares no conflicts of interest.
References
- Simoneit, B.R.T. Prebiotic organic synthesis under hydrothermal conditions: An overview. Adv. Space Res. 2004, 33, 88–94. [CrossRef]
- Schrödinger, E. What Is Life—The Physical Aspect of the Living Cell; Cambridge University Press: Cambridge, UK, 1944.
- Kleidon, A.; Lorenz, R.D. (Eds.). Non-Equilibrium Thermodynamics and the Production of Entropy: Life, Earth, and Beyond; Springer Science & Business Media: Dordrecht, The Netherlands, 2004.
- Marshall, M. How the first life on Earth survived its biggest threat—water. Nature 2020, 588, 210–213. [CrossRef]
- Russell, M.J. The “Water Problem” (sic), the Illusory Pond and Life’s Submarine Emergence—A Review. Life 2021, 11, 429. [CrossRef]
- Cnossen, I.; Sanz-Forcada, J.; Favata, F.; Witasse, O.; Zegers, T.; Arnold, N.F. Habitat of early life: Solar X-ray and UV radiation at Earth's surface 4–3.5 billion years ago. J. Geophys. Res. Planets 2007, 112, E02.
- Schreiber, U.; Mayer, C. The First Cell—The Mystery Surrounding the Beginning of Life; Springer: Cham, Switzerland, 2020.
- Deamer, D. The role of lipid membranes in life’s origin. Life 2017, 7, 5. [CrossRef]
- Mayer, C.; et al. Molecular Evolution in a Peptide-Vesicle System. Life 2018, 8, 16. [CrossRef]
- Dávila, M.J.; Mayer, C. Membrane structure obtained in an experimental evolution process. Life 2022, 12, 145. [CrossRef]
- Meierhenrich, U. Amino Acids and the Asymmetry of Life: Caught in the Act of Formation; Springer Science & Business Media: Dordrecht, The Netherlands, 2008.
- Hazen, R.M.; Filley, T.R.; Goodfriend, G.A. Selective adsorption of L- and D-amino acids on calcite: Implications for biochemical homochirality. Proc. Natl. Acad. Sci. USA 2001, 98, 5487–5490. [CrossRef]
- Chatterjee, S.; Yadav, S. The origin of prebiotic information system in the peptide/RNA world: A simulation model of the evolution of translation and the genetic code. Life 2019, 9, 25. [CrossRef]
- Martin, W.; Russell, M.J. On the origin of biochemistry at an alkaline hydrothermal vent. Philos. Trans. R. Soc. B Biol. Sci. 2007, 362, 1887–1926. [CrossRef]
- Deamer, D.; Damer, B.; Kompanichenko, V. Hydrothermal chemistry and the origin of cellular life. Astrobiology 2019, 19, 1523–1537. [CrossRef]
- Damer, B.; Deamer, D. The hot spring hypothesis for an origin of life. Astrobiology 2020, 20, 429–452. [CrossRef]
- Pearce, B.K.; Pudritz, R.E.; Semenov, D.A.; Henning, T.K. Origin of the RNA world: The fate of nucleobases in warm little ponds. Proc. Natl. Acad. Sci. USA 2017, 114, 11327–11332. [CrossRef]
- Kelley, D.S.; Früh-Green, G.L.; Karson, J.A.; Ludwig, K.A. The Lost City hydrothermal field revisited. Oceanography 2007, 20, 90–99. [CrossRef]
- Pierazzo, E.; Chyba, C.F. Amino acid survival in large cometary impacts. Meteorit. Planet. Sci. 1999, 34, 909–918. [CrossRef]
- Schreiber, U.; Locker-Grütjen, O.; Mayer, C. Hypothesis: Origin of life in deep-reaching tectonic faults. Orig. Life Evol. Biosph. 2012, 42, 47–54. [CrossRef]
- Mayer, C.; Schreiber, U.; Dávila, M.J. Periodic vesicle formation in tectonic fault zones—An ideal scenario for molecular evolution. Orig. Life Evol. Biosph. 2015, 45, 139–148. [CrossRef]
- Mayer, C.; Schreiber, U.; Dávila, M.J. Selection of prebiotic molecules in amphiphilic environments. Life 2017, 7, 3. [CrossRef]
- Dhuime, B.; Hawkesworth, C.J.; Cawood, P.A.; Storey, C.D. A change in the geodynamics of continental growth 3 billion years ago. Science 2012, 335, 1334–1336. [CrossRef]
- Rosas, J.C.; Korenaga, J. Rapid crustal growth and efficient crustal recycling in the early Earth: Implications for Hadean and Archean geodynamics. Earth Planet. Sci. Lett. 2018, 494, 42–49. [CrossRef]
- Schreiber, U.; et al. Organic compounds in fluid inclusions of Archean quartz—Analogues of prebiotic chemistry on early Earth. PLoS ONE 2017, 12, e0177570. [CrossRef]
- Großmann, Y.; Schreiber, U.; Mayer, C.; Schmitz, O.J. Aliphatic aldehydes in the Earth’s crust—Remains of prebiotic chemistry? Life 2022, 12, 925. [CrossRef]
- Peters, S.; Semenov, D.A.; Hochleitner, R.; Trapp, O. Synthesis of prebiotic organics from CO₂ by catalysis with meteoritic and volcanic particles. Sci. Rep. 2023, 13, 6843. [CrossRef]
- Bornemann, T.L.V.; et al. Genetic diversity in terrestrial subsurface ecosystems impacted by geological degassing. Nat. Commun. 2022, 13, 284. [CrossRef]
- Hennet, R.J.C.; Holm, N.G.; Engel, M.H. Abiotic synthesis of amino acids under hydrothermal conditions and the origin of life: A perpetual phenomenon. Naturwissenschaften 1992, 79, 361–365. [CrossRef]
- Marshall, W.L. Hydrothermal synthesis of amino acids. Geochim. Cosmochim. Acta 1994, 58, 2099–2106. [CrossRef]
- LaRowe, D.E.; Regnier, P. Thermodynamic potential for the abiotic synthesis of adenine, cytosine, guanine, thymine, uracil, ribose, and deoxyribose in hydrothermal systems. Orig. Life Evol. Biosph. 2008, 38, 383. [CrossRef]
- Peng, C.; Crawshaw, J.P.; Maitland, G.C.; Trusler, J.M.V.; Vega-Maza, D. The pH of CO₂-saturated water at temperatures between 308 K and 423 K at pressures up to 15 MPa. J. Supercrit. Fluids 2013, 82, 129–137. [CrossRef]
- Yi, R.; et al. A continuous reaction network that produces RNA precursors. Proc. Natl. Acad. Sci. USA 2020, 117, 13267–13274. [CrossRef]
- Ferris, J.P. Mineral catalysis and prebiotic synthesis: montmorillonite-catalyzed formation of RNA. Elements 2005, 1, 145–149. [CrossRef]
- Cleaves II, H.J.; Scott, A.M.; Hill, F.C.; Leszczynski, J.; Sahai, N.; Hazen, R. Mineral–organic interfacial processes: Potential roles in the origins of life. Chem. Soc. Rev. 2012, 41, 5502–5525. [CrossRef]
- Criado-Reyes, J.; Bizzarri, B.M.; García-Ruiz, J.M.; Saladino, R.; Di Mauro, E. The role of borosilicate glass in Miller–Urey experiment. Sci. Rep. 2021, 11, 1. [CrossRef]
- Ćuk, M.; Hamilton, D.P.; Lock, S.J.; Stewart, S.T. Tidal evolution of the Moon from a high-obliquity, high-angular-momentum Earth. Nature 2016, 539, 402–406. [CrossRef]
- Finkelstein, D.; Hill, U.S.; Powell, J.R. The piezoelectric theory of earthquake lightning. J. Geophys. Res. 1973, 78, 992–993. [CrossRef]
- Benner, S.A.; Kim, H.J.; Carrigan, M.A. Asphalt, water, and the prebiotic synthesis of ribose, ribonucleosides, and RNA. Acc. Chem. Res. 2012, 45, 2025–2034. [CrossRef]
- Fedor, M.J.; Williamson, J.R. The catalytic diversity of RNAs. Nat. Rev. Mol. Cell Biol. 2005, 6, 399–412. [CrossRef]
- Gesteland, R.F.; Cech, T.; Atkins, J.F. The RNA World, 3rd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2006.
- Lincoln, T.A.; Joyce, G.F. Self-sustained replication of an RNA enzyme. Science 2009, 323, 1229–1232. [CrossRef]
- Järvinen, P.; Oivanen, M.; Lönnberg, H. Interconversion and phosphorester hydrolysis of 2’,5’- and 3’,5’-dinucleoside monophosphates: Kinetics and mechanisms. J. Org. Chem. 1991, 56, 5396–5401. [CrossRef]
- Szostak, J.W. The eightfold path to non-enzymatic RNA replication. J. Syst. Chem. 2012, 3, 2. [CrossRef]
- Jungck, J.R. The genetic code as a periodic table. J. Mol. Evol. 1978, 11, 211–224. [CrossRef]
- Bailey, J.M. RNA-directed amino acid homochirality. FASEB J. 1998, 12, 503–507. [CrossRef]
- Carter, C.W. Coding of Class I and II aminoacyl-tRNA synthetases. Protein Rev. 2017, 103–148. [CrossRef]
- Eriani, G.; Delarue, M.; Poch, O.; Gangloff, J.; Moras, D. Partition of tRNA synthetases into two classes based on mutually exclusive sets of sequence motifs. Nature 1990, 347, 203–206.
Figure 1.
Schematic Block Profile of the Early Continental Crust: The average thickness of the modern continental crust is approximately 30 km; however, during early Earth, it was likely considerably thinner. Despite the overall higher global temperatures at the time, artesian water within the upper layers could have facilitated localized cooling, resulting in a temperature gradient within the crust.
Figure 1.
Schematic Block Profile of the Early Continental Crust: The average thickness of the modern continental crust is approximately 30 km; however, during early Earth, it was likely considerably thinner. Despite the overall higher global temperatures at the time, artesian water within the upper layers could have facilitated localized cooling, resulting in a temperature gradient within the crust.
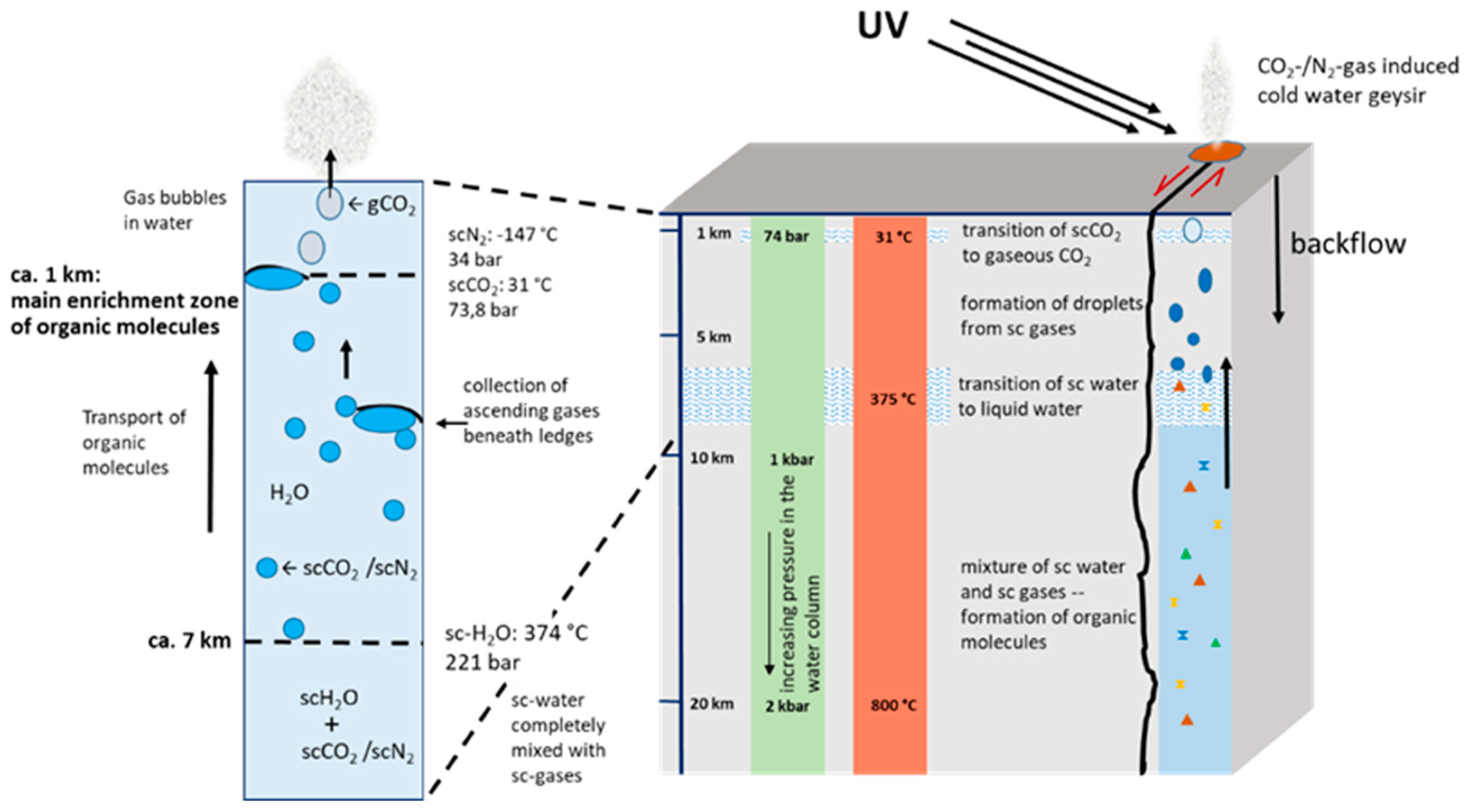
Figure 2.
Example of a proto-tRNA consisting of only 12 nucleobases: This form of proto-tRNA would have been selected from a diverse pool of RNAs, varying in length, conformation, and molecular variations, reflecting the diversity of possible sugar molecules in both chiral forms. The selection of a specific enantiomerically pure tRNA type likely occurred over an extended period, during which the processes outlined below would have played a crucial role. The sequence would include the bases A (adenine), G (guanine), C (cytosine), and U (uracil).
Figure 2.
Example of a proto-tRNA consisting of only 12 nucleobases: This form of proto-tRNA would have been selected from a diverse pool of RNAs, varying in length, conformation, and molecular variations, reflecting the diversity of possible sugar molecules in both chiral forms. The selection of a specific enantiomerically pure tRNA type likely occurred over an extended period, during which the processes outlined below would have played a crucial role. The sequence would include the bases A (adenine), G (guanine), C (cytosine), and U (uracil).

Figure 4.
Complementary amino acid groups reflecting the utility of proto-tRNA templates and their complementary strands, following Jungck [45]. Amino acids synthesized within hydrothermal systems are highlighted in color, with stop codons indicated in red. Hydrophilic amino acids (shown in shades of blue) are positioned in the most hydrophilic anticodon region.
Figure 4.
Complementary amino acid groups reflecting the utility of proto-tRNA templates and their complementary strands, following Jungck [45]. Amino acids synthesized within hydrothermal systems are highlighted in color, with stop codons indicated in red. Hydrophilic amino acids (shown in shades of blue) are positioned in the most hydrophilic anticodon region.
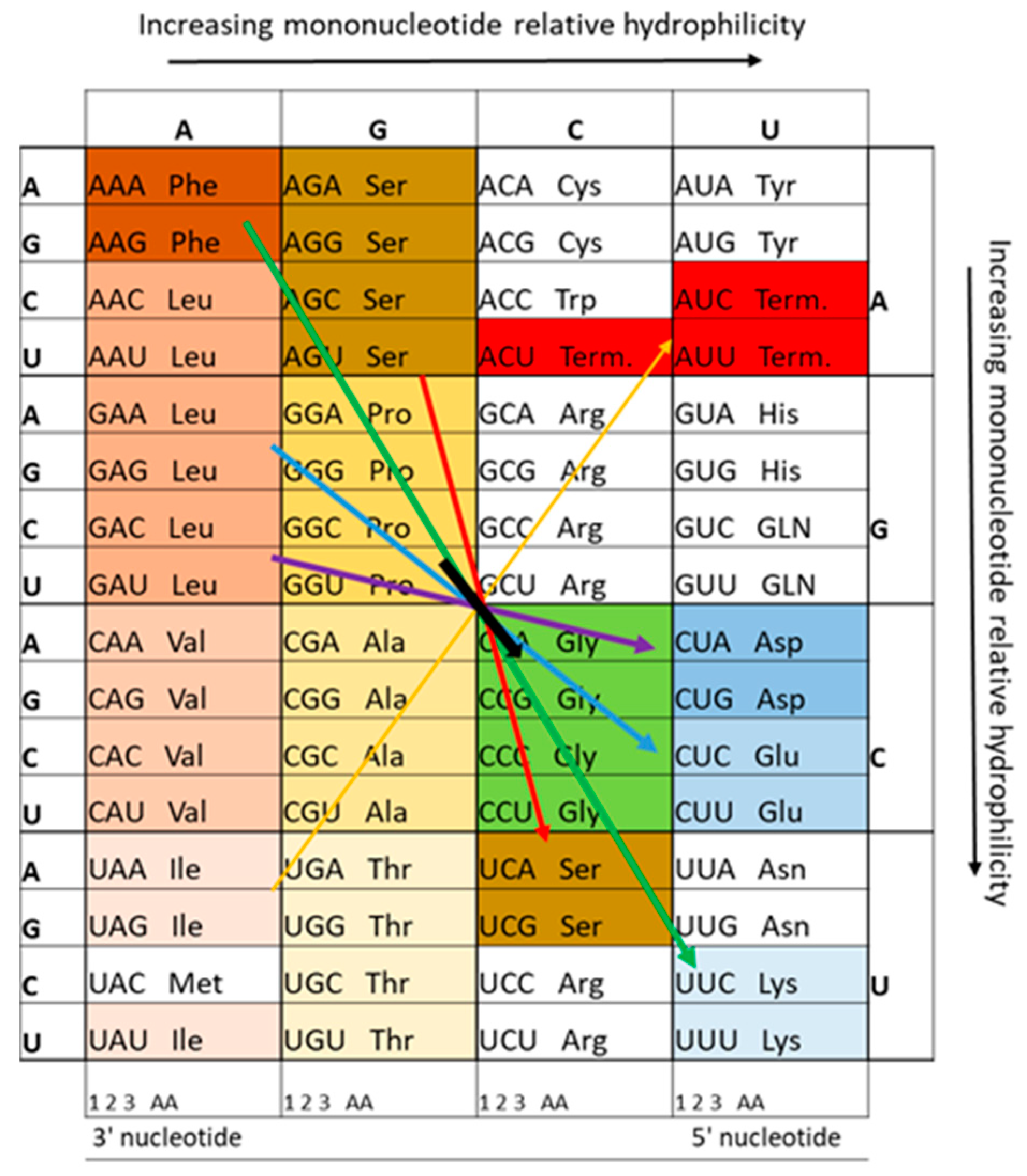
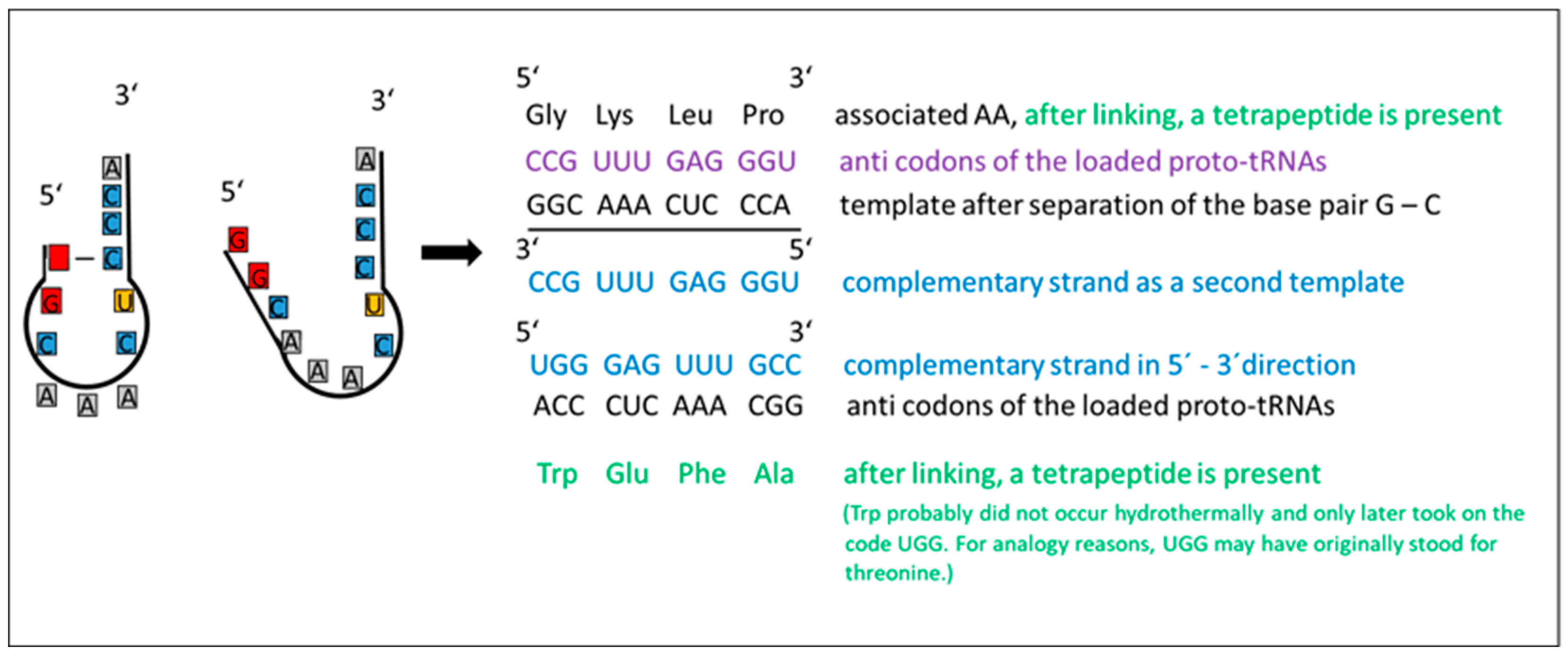
Figure 5.
Template utilization in proto-tRNA duplication - An example illustrating the utilization of separated proto-tRNA strands as a template for peptide assembly. Once the complementary strand is synthesized, a peptide chain comprising four amino acids (shown in green) is formed along the template strand.
Figure 5.
Template utilization in proto-tRNA duplication - An example illustrating the utilization of separated proto-tRNA strands as a template for peptide assembly. Once the complementary strand is synthesized, a peptide chain comprising four amino acids (shown in green) is formed along the template strand.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



