
Submitted:
04 February 2025
Posted:
05 February 2025
You are already at the latest version
Abstract
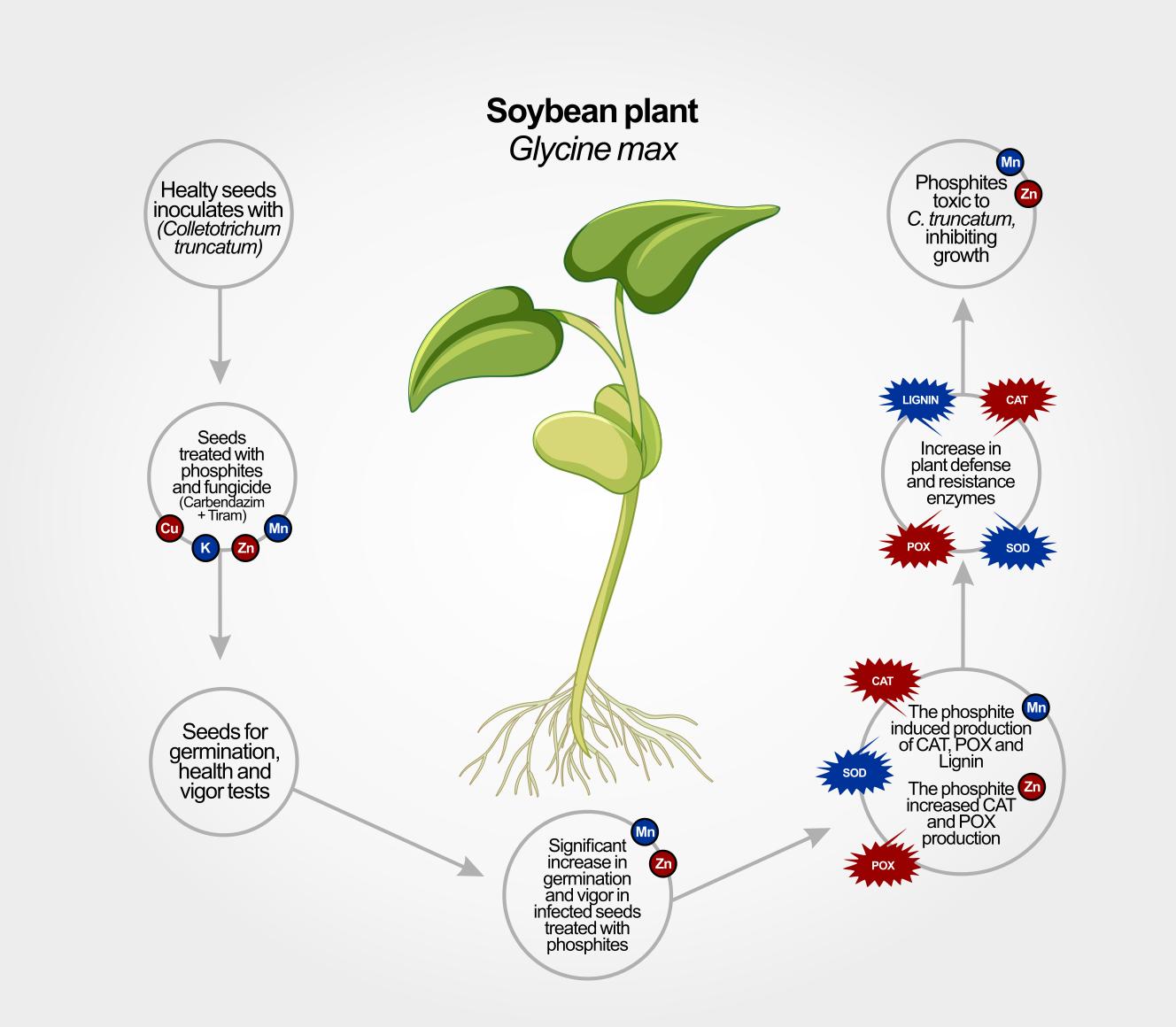
Soybean is one of the main agricultural commodities, and its productivity is limited by several diseases, such as anthracnose, which is caused by a complex of fungal species, with Colletotrichum truncatum being the most prevalent. Management is mainly carried out through chemical seed treatment. However, a reduction in the sensitivity of C. truncatum to fungicides was observed. Therefore, it is extremely important to search for products that are effective in controlling the disease. The objectives of this study were to evaluate the efficacy of commercial formulations of Cu, K, Mn and Zn phosphites in the treatment of soybean seeds infected by C. truncatum and their ability to induce defense mechanisms against the fungus. For this purpose, seeds inoculated with C. truncatum were treated with phosphites and a fungicide (carbendazim + thiram). The seeds were subjected to germination, health and vigor tests. Fungal toxicity and the ability of phosphites to induce defense through the activities of catalase, peroxidase and superoxide dismutase enzymes, as well as the levels of lignin and total soluble phenols, were also evaluated. Mn and Zn phosphites were toxic to C. truncatum and effective in controlling anthracnose in infected soybean seeds, showing no difference in relation to the fungicide. Mn phosphite induced the production of CAT, POX and lignin, while Zn phosphite increased the production of CAT and POX. These results demonstrate the effectiveness of Mn and Zn phosphites in controlling anthracnose in infected soybean seeds, their direct toxic action and their ability to induce defense mechanisms.
Keywords:
Glycine max L.
; Seed treatment
; Potassium phosphite
; Zinc phosphite
; Manganese phosphite
; Copper phosphite
1. Introduction
Soybean (Glycine max [L.] Merril) is one of the most important commodities in the world [1]. Rich in proteins (40%) and moderate in oil (20%), it is used in both human and animal feed [2,3] and is of vital importance for ensuring global food security [4,5]. However, the crop can be affected by various pathogens from seed germination to grain filling. Anthracnose is one of the most significant fungal diseases affecting soybean [6] and is prevalent in nearly all soybean-growing areas worldwide, causing yield losses that can reach up to 100% [7,8]. Approximately 90 kg/ha of grain yield is lost for each 1% increase in disease incidence in commercial soybean fields [9]. Although new species have recently been reported to be associated with anthracnose, Colletotrichum truncatum (Schw.) Andrus & WD Moore is the predominant species in soybean fields [6,10]. C. truncatum is one of the most common and harmful seed-borne pathogens affecting soybean, particularly when the crop is grown under warm and humid conditions [11]. The fungus can attack all parts of the plant at all developmental stages [12,13]. In infected seeds, it causes a significant reduction in seed quality, leading to a decrease in plant stand [14]. Typical anthracnose symptoms include pre- and post-emergence damping-off; dark, sunken, and irregular lesions on cotyledons, stems, petioles, and pods, where acervuli are observed [10]; immature pod opening, resulting in seed germination [15]; and necrotic lesions on the abaxial leaf veins, which can lead to premature defoliation [7,10]. Recurring reports of severe epidemics and significant productivity losses have increased each year [10]. Management of this disease is carried out through seed chemical treatment or genetic resistance [16,17]. However, highly resistant cultivars are not known in Brazil. Therefore, chemical control remains the most commonly used method for this purpose. However, studies have demonstrated a reduction in C. truncatum sensitivity to fungicides recommended for the crop [9,18,19]. C. truncatum exhibits high genetic variability [10], which allows the fungus to adapt to various environmental conditions, hosts, and control measures implemented in its management. This high variability complicates genetic improvement efforts in the search for soybean cultivars with durable resistance and results in a reduction in the fungus's sensitivity to fungicides used for its control. Moreover, due to the harmful effects of pesticides on the environment and human health, as well as a reduction in Bradirrizhobium viability and consequent damage to nodulation [20], new alternative seed treatment methods for controlling anthracnose in soybeans are necessary. Among the alternative products available on the market, phosphites stand out. These products are obtained through the reaction of phosphorous acid with a base (K+, Ca2+, among others). They can act on nutrition, direct toxicity to the pathogen, and the induction of resistance in the host [21]. Another important feature of phosphites is their low cost compared to chemical fungicides [22]. Thus far, no studies have been found regarding soybean seed treatment with phosphites for the control of anthracnose, and few studies have tested phosphites in seed treatment. Given the increasing importance of anthracnose worldwide and reports of C. truncatum resistance to fungicides recommended for the crop, this study aimed to evaluate the effectiveness of four commercial phosphite formulations (Phi): copper phosphite (CuPhi), manganese phosphite (MnPhi), potassium phosphite (KPhi), and zinc phosphite (ZnPhi) in treating soybean seeds infected by C. truncatum and characterize their mechanism of action in controlling anthracnose via seed treatment.
2. Materials and Methods
2.1. Fungal Strains
The isolate of C. truncatum LAPS-473, obtained from the Seed Analysis and Pathology Laboratory Collection at the Federal University of Lavras (UFLA), was used. Fragments of the fungus were placed at the center of a Petri dish containing PDA (Potato-Dextrose-Agar at 4%). The Petri dishes were then kept in a growth chamber at 27±1°C with a 12-hour photoperiod at 40 μmol s−1 m−2 for seven days. A monosporic culture was obtained from the colony, following the method described by Magalhães et al. [23], and was used throughout the experiment.
2.2. Source and Doses of Phosphites and Fungicide
Four commercial phosphite formulations (Phi) obtained from Agrichem Brazil were tested: Reforce Mn® (MnPhi; 10% Mn + 51.5% P2O5), Reforce Zn® (ZnPhi; 10% Zn + 34% P2O5), Yantra® (KPhi; 29% K2O + 33.6% P2O5), and Reforce Cu® (CuPhi; 4% Cu + 20.3% P2O5) at doses of 500, 500, 500, and 250 mL.100 kg⁻¹ of seeds, respectively. The commercial fungicide Protreat® (150 g of carbendazim.L⁻¹ + 350 g of thiram.L⁻¹) from Novozymes BioAq Produtos para Agricultura Ltda, recommended for seed treatment to control anthracnose, was used as a positive control treatment at a dose of 200 mL.100 kg⁻¹ of seeds.
2.3. Inoculation of Soybean Seeds with C. truncatum
Soybean seeds of the TMG 1176 RR cultivar, highly susceptible to the fungus C. truncatum, were obtained from the Mato Grosso Foundation. Initially, the seeds were subjected to germination and health tests to determine the quality of the seed lot, following the Rules for Seed Analysis [24]. Subsequently, the seeds were inoculated using the water restriction method [25]. For this purpose, the PDA culture medium was prepared at an osmotic potential of -1.0 MPa by adding mannitol (water restrictor), in the quantity indicated by the SPPM software [26]. Ten mL portions of the sterilized PDA medium were transferred to 90 mm Petri dishes and allowed to solidify. Three 5-mm-diameter disks containing C. truncatum mycelium were placed at the center of each Petri dish and incubated in a growth chamber at 27±1°C with a photoperiod of 12 h at 40 μmol s−1 m−2 until the fungus grew uniformly across the plate. Subsequently, a total of 100 seeds were placed in each Petri dish, remaining in contact with the fungus for 28 hours. After this period, the seeds were removed and dried in a laminar flow cabinet for 24 hours.
2.4. Seed Treatments
Soybean seeds inoculated with C. truncatum were subjected to the treatments described above. The seeds were placed in 1 L plastic bags with the treatments and manually agitated for 2 minutes. Then, the bags were kept open for drying and for the treatments to adhere to the seeds. The control treatment consisted of inoculated seeds that were not treated, and non-inoculated seeds treated with water.
2.5. Effects of Phosphites on Seed Health and Germination
The standard germination test (roll paper method) and blotter test described in the manual issued by MAPA [24], were employed to determine the incidence of C. truncatum and the percentage of germinated seedlings that emerged from inoculated treated seeds, inoculated non-treated seeds, and non-inoculated non-treated seeds, as well as their health quality. A total of 400 seeds per treatment were used, divided into eight replicates, each containing 50 seeds. For the standard germination test, the seeds were incubated in a growth chamber at 25±1°C with a 12-hour photoperiod at 40 μmol s−1 m−2 for 7 days. For the blotter test, seeds were subsequently maintained at 20°C under a 12:12 h photoperiod for eight days and then examined individually under a stereomicroscope to determine the level of C. truncatum infection. The experiment was conducted in a completely randomized design.
2.6. Emergence and Development of Soybean Seedlings Under Greenhouse Conditions
The seeds were sown in trays containing sand, previously autoclaved for 1 h at 121ºC and 1 atm, and were irrigated daily using a controlled micro-spray system. The emergence speed index (ESI) was determined according to the methodology described by Maguire [27]. Evaluations were conducted daily by counting the number of seedlings that emerged until the stabilization of the plant population. On the 7 and 14th day after sowing, initial (IS) and final (FS) stand establishment were assessed. On the 28th day after sowing, each plantlet was cut at the soil surface and its height (H) and root length (R) were recorded. To determine the dry weight biomass of the aerial part (BA) and roots (BR), plantlets were transferred to paper bags and maintained in a forced air oven at 60ºC until constant weight was attained. The whole experiment was conducted according to a randomized block design with 8 treatments replicated two times with 8 plots of 50 seeds per replicate totaling 400 seeds per treatment.
2.7. Fungitoxic Activity of Phosphites
The fungicide carbendazim + thiram, MnPhi, and ZnPhi were tested for their efficacy in controlling anthracnose in infected soybean seeds. The fungicidal capacity of the phosphites (Phi) against C. truncatum was evaluated according to the methodology of Araújo et al. [2010]. Discs of 5 mm in diameter, containing C. truncatum mycelium, were placed at the center of Petri dishes and incubated in a growth chamber at 27±1°C with a 12-hour photoperiod at 40 μmol s−1 m−2 for eight days. MnPhi and ZnPhi were tested at doses of 1.25, 2.5, 5, and 10 mL.L⁻¹, and the fungicide was applied at the commercial dose (2 mL.L⁻¹) to the molten PDA culture medium, which was then poured into 9 cm diameter Petri dishes. After the medium solidified, 5 mm diameter mycelial discs of C. truncatum were placed in the center of each Petri dish. PDA medium without any treatment was used as the control. The plates were incubated in a growth chamber at 25±1°C with a 12-hour photoperiod at 40 μmol s−1 m−2. Two orthogonal measurements of colony diameter were taken at a fixed time each day from the 2nd to the 9th day. The mycelial growth index (MGI) was calculated using the expression MGI = Σ(D – Da) / N, where D is the actual mean colony diameter, Da is the previous mean diameter, and N is the number of growth days [29]. Based on the MGI, the lethal dose that inhibited 50% of mycelial growth (LD50) of C. truncatum was calculated according to the methodology proposed by Torrez-Calzada et al. [30]. The experiment was conducted in a completely randomized design with 4 replications, each consisting of one Petri dish.
2.8. Activation of Biochemical Defense Mechanisms
Soybean seeds inoculated and treated with MnPhi and ZnPhi were subjected to the germination roll paper test, as described earlier. A total of 200 seeds per treatment were used, divided into eight replications, with 25 seeds per replication. The number of seeds used in this assay was lower than in the previous one to avoid root entangling. The seeds were incubated in a growth chamber at 25±1°C with a 12-hour photoperiod at 40 μmol s−1 m−2. Plant material was collected at 24, 48, 72, 96, and 192 hours after incubation (hai). The plant material was wrapped in aluminum foil and stored in a freezer at -80°C until the beginning of the analyses. Three grams of plant material from each treatment were macerated in liquid nitrogen. An aliquot of 0.5 g and another of 30 mg were collected from each treatment for enzymatic analyses and lignin content determination, respectively. The 0.5 g aliquot was added to 2 mL microtubes, followed by the addition of 1.5 mL of 100 mM phosphate buffer, pH 7.8, and 1.5 mL of 0.1 mM EDTA and 10 mM ascorbic acid. The microtubes were centrifuged at 12000 g, at 4°C, for 15 minutes, and the supernatant was collected for enzymatic analyses. The protein concentration in the suspension was determined according to the Bradford [31] method, using bovine serum albumin as the standard, and adjusted for microquantities. For this, 10 µL of the sample, 20 µL of 100 mM potassium phosphate buffer, pH 7.8, and 170 µL of Bradford reagent were added to 96-well microplates. The absorbance was measured using a spectrophotometer (Biotek®) at 595 nm. A standard curve was prepared with bovine serum albumin (BSA) between 0 and 100 µg, and the results were expressed as the average of three replicates.
Peroxidase (POX) activity was evaluated using the guaiacol oxidation method, in which the enzyme mixture consisted of 30 μL of sample, 130 μL of 19.2 mM phosphate potassium buffer (pH 7), 20 μL of 50 mM guaiacol, and 20 μL of 125 mM hydrogen peroxide. The absorbance of the reaction mixture at 480 nm was measured after 10 minutes of incubation at 30°C [32], and peroxidase activity was expressed as μmol of tetraguaiacol produced per minute per mg of protein. The molar extinction coefficient of 1.23 mM⁻¹ cm⁻¹ was used to calculate POX activity [33].
Catalase (CAT) activity was assessed by the decrease in absorbance at 240 nm for 3 minutes at 25°C. The enzyme mixture consisted of 10 μL of the enzymatic extract, 60 μL of sterilized distilled water, 100 μL of 200 mM potassium phosphate buffer, pH 7, and 20 μL of 250 mM hydrogen peroxide. The molar extinction coefficient of 18 M⁻¹ cm⁻¹ was used to calculate the activity of this enzyme [34].
Superoxide dismutase (SOD) activity was determined by the enzyme's ability to inhibit the photoreduction of nitro blue tetrazolium (NBT), as described by Giannopolitis and Ries [35]. In each well of a 96-well microplate, 10 μL of the enzymatic extract, 30 μL of sterilized distilled water, 2 μL of 0.2 mM riboflavin, 2 μL of 10 μM EDTA, 40 μL of 70 mM methionine, 100 μL of 100 mM potassium phosphate buffer, pH 7.8, and 15 μL of 1 mM NBT were added. The plates were incubated for 7 minutes in a closed chamber with a 30W fluorescent lamp. Absorbance was measured at 560 nm, and the enzyme unit was defined as its ability to inhibit 50% of NBT photoreduction. Based on the activities of the CAT, POX, and SOD enzymes, the area under the activity progress curve (AUAPC) was calculated for CAT (AUAPCCat), POX (AUAPCPox), and SOD (AUAPCSod), following the methodology proposed by Shanner & Finney [36].
Soluble lignin was determined using the method proposed by Doster and Bostock [37]. A 30 mg sample of freeze-dried plant material was added to 1.5 mL of a solution of thioglycolic acid and hydrochloric acid (HCl) 2M (1:10 ratio). The samples were then incubated in a water bath for 4 hours. Absorbance was measured at 280 nm in a spectrophotometer, and the value was expressed in μg.mg⁻¹ dry weight. The evaluation of total soluble phenolic compounds was conducted according to the methodology of Spanos and Wrolstad [38]. Absorbance was measured at 725 nm, and the amount was expressed in μg.mg⁻¹ dry weight. The experiment was conducted in a completely randomized design.
2.9. Statistical Analysis
The experiments were conducted in duplicates, and the results were presented as the average of both experiments. Firstly, the assumptions of the analysis of variance were checked. To assess normality and homoscedasticity, the Shapiro-Wilk and Bartlett tests were used, respectively. The data did not follow the assumptions of the analysis of variance and needed to be transformed. In cases of significance, the data were transformed using √(x+1). To accept or reject H0 (null hypothesis), the data were analyzed using the F test, and in case of significance, regression analysis or mean comparison tests were performed to differentiate the treatments. Scott-Knott, Tukey, and Dunnet tests (α < 0.05) were applied to determine the significance of the differences between the mean values. These analyses were carried out using the R program from the R Development Core Team [39]. The standard error was also calculated.
3. Results
3.1. Effects of Phosphites on Seed Health and Germination
The test for the physiological and sanitary quality of the seeds showed that the seed lot had good sanitary quality, with less than 4% incidence of C. truncatum and a germination rate of 85.5% (Table 1). All the phosphites increased the germination of the inoculated seeds when compared to the inoculated and untreated seeds. Inoculated seeds treated with MnPhi and ZnPhi exhibited germination percentages similar to those of the non-inoculated seeds and seeds treated with KPhi and CuPhi also increased the germination of the inoculated seeds but were less effective than MnPhi, ZnPhi, and the fungicide Protreat®. The phosphites also reduced the incidence of C. truncatum in the seeds. As with germination, MnPhi and ZnPhi did not differ from the fungicide Protreat® and were the treatments that most reduced the incidence of C. truncatum in the inoculated seeds, followed by KPhi and CuPhi. The inoculated and untreated seeds had an incidence of C. truncatum above 96%.
3.2. Emergence and Development of Soybean Seedlings Under Greenhouse Conditions
Colletotrichum truncatum significantly reduced the emergence speed index (ESI), initial stand at seven days (IS), and final stand (FS) of the inoculated and untreated seeds (Table 2). Consequently, there was lower plant height (H), root length (R), dry biomass of the aerial part (BA), and dry biomass of the roots (BR) of plants derived from these seeds. However, inoculated seeds treated with Phi and the fungicide showed superior results in all of these parameters compared to the inoculated, untreated seeds. Among the treatments, the highest ESI was observed when the inoculated seeds were treated with ZnPhi, MnPhi, and fungicide, followed by KPhi and CuPhi. Seven days after sowing, the treatments that showed the highest IS were ZnPhi and fungicide, followed by MnPhi and KPhi. Among the Phi, CuPhi had the lowest IS. Fourteen days after sowing, MnPhi, ZnPhi, and the fungicide achieved the highest FS, followed by KPhi. As with EI, CuPhi was the phosphite that showed the lowest FS, but it was still superior to the inoculated, untreated seeds. Regarding plant height, it was observed that MnPhi, KPhi, ZnPhi, and the fungicide induced the greatest plant growth, surpassing the plants derived from non-inoculated, untreated seeds, which did not differ from the seeds treated with CuPhi. The roots of plants derived from inoculated seeds treated with MnPhi, ZnPhi, and the fungicide exhibited greater growth, not differing from the roots of plants derived from non-inoculated seeds, followed by the treatments based on KPhi and CuPhi. Regarding BA, ZnPhi, KPhi, MnPhi, and the fungicide showed the best results, not differing from the non-inoculated seeds, followed by CuPhi. As for BR, the treatments that achieved the best results were MnPhi, ZnPhi, and fungicide, followed by CuPhi and KPhi, which were lower than those of the non-inoculated seeds.
3.3. Fungitoxic Activity of Phosphites
ZnPhi and MnPhi reduced the mycelial growth of C. truncatum with increasing doses (Figure 1), demonstrating that both have fungitoxic activity against the fungus. The DL50 and MIC of ZnPhi and MnPhi against C. truncatum were very similar, corresponding to a DL50 of 2.35 mL.L-1 and 2.48 mL.L-1, and MICs of 8.44 mL.L-1 and 8.64 mL.L-1, respectively (Table 3). At a dose of 8.6 mL.L-1, the phosphites completely inhibited the mycelial growth of the fungus. At a dose of 10 mL.L-1 (2x the dose used in seed treatment), it would correspond to the dose of the fungicide carbendazim + thiram used for soybean seed treatment to control C. truncatum (Figure 2).
3.4. Activation of Biochemical Defense Mechanisms
An increase in the activity of the CAT and POX enzymes in soybean seeds infected with C. truncatum and treated with MnPhi and ZnPhi was observed (Figure 3). The increase in CAT activity was observed over time, with two activation peaks at 72 and 196 hours after incubation when treated with ZnPhi, and at 96 hours after incubation when treated with MnPhi (Figure 3A). No difference in the AUAPCCat at was observed in seeds treated with both Phi (Figure 3D). For the POX and SOD enzymes, activation peaks occurred at 192 hours after incubation for seeds treated with both Phi (Figure 3B,C). However, seeds treated with ZnPhi presented higher AUAPCPox compared to seeds treated with MnPhi (Figure 3D). For AUAPCSod, no significant difference was found between MnPhi and ZnPhi, nor in relation to the control treatment (Figure 3D).
No increase in the total soluble phenolic content was observed in the seeds treated with both phosphites (Figure 4). However, a significant increase in the lignin content was observed in the seeds treated with MnPhi.
4. Discussion
New products for soybean seed treatment to control C. truncatum are critically needed due to the increasing occurrence of anthracnose in productive fields in recent years and the resistance of the fungus to active ingredients of fungicides widely used in the main soybean growing areas worldwide. Additionally, there is a global trend toward seeking products that cause less environmental and public health impact and, in the case of soybeans, products that do not negatively interfere with nodulation. The irrational use of synthetic pesticides in soybean plantations has caused serious environmental problems, such as reduced biodiversity in agroecosystems and reduced nodulation by nitrogen-fixing bacteria, such as those of the Bradyrhizobium genera [20,40]. However, in recent decades, considerable evidence has been provided to establish the value of phosphites (Phi) in suppressing a range of plant diseases [41]. Despite Yáñez-Juárez et al. [40] reporting that phosphites are generally less effective in reducing damage caused by plant pathogens and could not completely replace fungicides, we found that ZnPhi and MnPhi were as effective as the commercial fungicide in controlling anthracnose in soybeans. Similarly, CuPhi, ZnPhi, and K/MnPhi phosphites were also more effective than the fungicide in controlling powdery mildew in eucalyptus seedlings [42]. Soybean seeds treated with MnPhi were more efficient than the commercial fungicide based on fludioxonil + metalaxyl in controlling damping-off caused by Pythium aphanidermatum, P. irregulare, and P. ultimum, with 97% of seeds germinating, while only 73% and 10% of seeds treated with the fungicide and untreated seeds germinated, respectively [41]. Phi has been mainly used in the management of foliar diseases in different crops [42,43,44], including soybeans [45]. Some studies have also shown their effectiveness in seed treatment, such as in cucumber seeds for controlling Pythium spp. [46], in hemp seeds against damping-off [47], in potato seeds for controlling Fusarium solani, Rhizoctonia solani, and Phytophthora infestans [48], and in soybean seeds for controlling Macrophomina phaseolina [49] and Pythium spp. [41]. Several commercial products containing Phi are available at the market. Most formulations vary in composition, properties, application rates, and the type of cation bound to the Phi molecule [41]. Phi has the advantages of low cost and excellent absorption by plants, being easily transported within plants, and displaying the unique property of moving through the plant not only via xylem but also via phloem [50,51]. Due to their efficient and rapid translocation in plant tissue, they can be applied to the canopy, stems, roots, or fruits [40], accumulate in regions of rapid growth, such as roots and shoots [52] and because they are not metabolized by the plants [50,53] they can remain there for months or even years, preventing pathogen attacks [54,55]. These characteristics bring important advantages when considering soybean seed treatment for controlling anthracnose, which occurs both in seeds and in the aerial part of the plant. In addition to possessing protective and curative properties [56], Phi has shown low phytotoxicity [41,46], stimulate arbuscular mycorrhizal colonization [57,58], and can be mixed with other defense inducers, biological control agents, or fungicide molecules without causing inactivation or reduction in the activity of biocontrol agents [49,59,60]. The application of Bacillus subtilis in combination with Phi showed an additive effect, significantly reducing charcoal root rot (Macrophomina phaseolina) in soybean plants compared to treatments with B. subtilis or Phi separately, as well as the application of MnPhi in association with Pseudomonas fluorescens [49]. A reduction in the number of Meloidogyne javanica eggs in soybean roots treated with MnPhi was also observed [61]. These results demonstrate that MnPhi can be used in soybean seed treatment for controlling various plant pathogens.
MnPhi and ZnPhi caused direct toxicity to C. truncatum, inhibiting its growth in culture media, promoting control of anthracnose in soybean seeds, stimulating seedling growth, and inducing catalase (CAT) and peroxidase (POX) enzyme activity, as well as promoting increases in lignin content in plant tissues, in the case of MnPhi. Different studies have also demonstrated the direct action of phosphites that limits the growth, development, and reproduction of plant pathogens. This is due to a combination of factors, such as the formation of pores and lysis of the fungal cell wall and membrane, leading to the efflux of cellular content [42,62] and transcriptional changes in several genes that code for proteins involved in the biosynthesis of cell wall components, amino acid synthesis, protein metabolism, and oxidative stress [21,63]. The decrease in the pH of the culture medium is also a factor that has led to the reduction of microorganism growth [28,64]. The level of growth reduction is determined by the tested organism, the amount of phosphite added, the pH produced in the culture medium, and the type of ion bound to the phosphite [28,64]. It is likely that the level of resistance of pathogens to phosphites is minimal because phosphites act on various targets within the pathogens [54,65]. However, there have been reports of resistance in isolates, such as Bremia lactucae in lettuce [66] and P. cinnamomi in avocados [67]. Therefore, the use of phosphites should be implemented within an integrated management program.
These changes and the rupture of fungal structures can lead to the release of elicitor molecules, triggering plant defense mechanisms [63,68]. The ability to induce resistance in plants when treating them with phosphites has also been documented. Biochemical and structural defense mechanisms, such as the production of pathogenesis related proteins, phytoalexins, reactive oxygen species, deposition of callose, lignin, and suberin that restrict pathogen penetration and survival in the plant, have been observed when the phosphite ion enters plant tissue cells [40,69,70,71]. Daniel and Guest [72] demonstrated that when Arabidopsis thaliana seedlings were treated with KPhi and inoculated with zoospores of Phytophthora palmivora, the infected cells exhibited increased cytoplasmic activity, development of cytoplasmic aggregates, hydrogen peroxide release, localized cell death, and an increase in phenolic compounds. We observed that seedlings originated from seeds treated with MnPhi had increases in CAT and POX activities, as well as increases in lignin content. ZnPhi also promoted significant increases in CAT and POX activities. The activation of CAT and POX enzymes is likely due to Mn and Zn acting as cofactors for these enzymes. According to Zhao et al. [73], increased Mn supply led to higher activity of SOD and CAT enzymes in Phytolacca americana plants, while increased Zn supply raised POX activity. Higher doses of Mn also promoted significant increases in CAT and POX activities in barley [74]. MnPhi induced higher activities of the enzymes ascorbate peroxidase (APX), CAT, SOD, and polyphenol oxidase (PPO) in coffee seedlings inoculated with Hemileia vastatrix [44]. Regarding lignin, Rengel et al. [75] found that increased Mn concentration in soil induced higher lignin content in wheat (Triticum aestivum) plants. These authors correlated the increased lignin content with plant resistance to the fungus Geumannomyces graminis. According to Dordas [76], Mn controls lignin and suberin biosynthesis by activating numerous enzymes in the shikimate and phenylpropanoid pathways, where peroxidase can act. In this study, both ZnPhi and MnPhi significantly increased POX activity. Peroxidase catalyzes the oxidation and eventual polymerization of hydroxycinnamic alcohol in the presence of hydrogen peroxide, resulting in lignin [77]. Therefore, the increase in lignin is related to higher POX activity. According to Lobato et al. [48], resistance-inducing products may involve a metabolic cost resulting from the production of defense compounds or structures. However, these authors observed that CaPhi and KPhi induced defense mechanisms in seed tubers without being detrimental to plant growt [48], which was also observed in our study. We observed that the defense response in soybean seedlings was rapidly activated. This may be due to the rapid absorption of the phosphites by the roots. Griffith, Coffey, and Grant [78] suggested that plant cells may absorb phosphites more quickly than phosphorus.
We observed that the phosphites, except CuPhi, induced greater growth of the aerial part and roots of soybean, whose seeds were previously treated. However, there is much controversy, as studies question the effects of phosphite on growth promotion [79,80], while others have shown their capacity to stimulate aerial part and root growth, as observed in wheat, canola, sugarbeet, ryegrass and Arabidopsis thaliana [44,45,58,81]. Phosphite cannot be converted into phosphate, so it does not improve plant growth through a nutritional mechanism [80]. However, further studies are needed to elucidate these mechanisms.
Although KPhi and CuPhi showed intermediate results in treating soybean seeds infected with C. truncatum, some studies have demonstrated their effectiveness in controlling certain diseases and inducing resistance in plants, such as KPhi against Phytophthora infestans in potatoes [70,82,83], Pythium spp. in soybeans [41], CuPhi controlling powdery mildew in eucalyptus [42,62] and Pythium spp. in cucumbers [46].
This study presents sufficient data and evidence to show that ZnPhi and MnPhi can be used as an effective alternative in soybean seed treatments for controlling C. truncatum. The use of both Phi in soybean seed treatments has shown numerous benefits, such as efficient control of anthracnose in already infected seeds; the ability to control various soybean diseases, as observed in other studies; the potential to protect the aerial parts against anthracnose due to their rapid absorption and translocation; the ability to stimulate soybean growth and induce resistance mechanisms; low cost; the amount of Phi required to treat seeds is much lower than what is needed for foliar application; and the use of these products constitutes a viable and sustainable method for effective soybean seed protection against anthracnose. As far as we are aware, this study is the first report of using Phi as a soybean seed treatment for controlling C. truncatum. Further studies are needed to evaluate the duration of protection provided by Phi, both in the aerial and root parts, and the spectrum of action against different diseases affecting soybean crops. This will allow for a better understanding of the performance of these products in disease management.
5. Conclusions
MnPhi and ZnPhi at doses of 5 mL.kg⁻¹ promoted control of anthracnose in soybean seeds infected with C. truncatum, similar to the fungicide recommended for soybean seed treatment. Both Phi showed fungi toxic activity against C. truncatum and were able to induce the production of catalase and peroxidase enzymes, which are involved in the plant defense mechanisms against pathogens. Manganese phosphite also induced lignin accumulation in tissues infected by C. truncatum. KPhi, MnPhi, and ZnPhi induced greater soybean growth.
Author Contributions
MBSJ conceptual idea, methodology design, data collection, data analysis and interpretation, writing and editing. MLVR conceptual idea, methodology design. EAP conceptual idea, data analysis and interpretation, writing and editing. ARMR conceptual idea, data analysis and interpretation, writing and editing. GCDS conceptual idea, data analysis and interpretation, writing and editing. JDVdata analysis and interpretation, writing and editing. ACS conceptual idea, data analysis and interpretation, writing and editing. JMO data analysis and interpretation, writing and editing, translation.
Funding
This research was supported by the “Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), “Fundação de Amparo à Pesquisa do Estado de Minas Gerais (FAPEMIG)”, and the “Instituto Nacional de Ciência e Tecnologia do Café (INCT)”.
Data Availability Statement
The datasets generated and/or analyzed during the current study are available in the Zenodo repository (Batista da Silva Júnior et al., 2025) at https://doi.org/10.5281/zenodo.14787619.
Acknowledgments
We thank the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), the Instituto Nacional de Ciência e Tecnologia do café (INCT-Coffee), and the Fundação de Amparo à Pesquisa do Estado de Minas Gerais (FAPEMIG) for contributing to this study.
Conflicts of Interest
The authors declare that they have no conflict of interest.
References
- Chen, H., Li, H., Liu, Z., Zhang, C., Zhang, S., Atkinson, PM, 2023. A novel Greenness and Water Content Composite Index (GWCCI) for soybean mapping from single remotely sensed multispectral images. Remote Sensing of Environment, 295. [CrossRef]
- Belewu, MA., Belewu, KY, 2007. Comparative physico-chemical evaluation of tiger-nut, soybean and coconut milk sources. International Journal of Agriculture and Biology. 9, 785-7.
- Pagano, MC., Miransari, M., 2016. The importance of soybean production worldwide. In: Miransari, M. (Ed.) Abiotic and Biotic Stresses in Soybean Production. Cambridge, MA, USA: Academic Press, 1-26. [CrossRef]
- Inglada, J., Arias, M., Tardy, B., Hagolle, O., Valero, S., Morin, D., Dedieu, G., Sepulcre, G., Bontemps, S., Defourny, P., Koetz, B., 2015. Assessment of an operational system for crop type map production using high temporal and spatial resolution satellite optical imagery. Remote Sens. 7, 12356-12379. [CrossRef]
- Matton, N., Canto, G., Waldner, F., Valero, S., Morin, D., Inglada, J., Arias, M., Bontemps, S., Koetz, B., Defourny, P., 2015. An automated method for annual cropland mapping along the season for various globally-distributed agrosystems using high spatial and temporal resolution time series. Remote Sens. 7, 13208-13232. [CrossRef]
- Rogério, F., de Castro, RRL., Massola Júnior, NS., Boufleur, TR., dos Santos, RF., 2024. Multiple resistance of Colletotrichum truncatum from soybean to QoI and MBC fungicides in Brazil. Journal of Phytopathology, 172, e13341. [CrossRef]
- Hartman, GL., Sinclair, JB., Rupe, JC., 1999. Compendium of Soybean Diseases, 4th ed.; APS Press: St. Paul, MN, USA. [CrossRef]
- Billores, SDO., Joshi, P., Bhatia, VS., Ramesh, A., 2011. Soybean Based Intercropping Systems in India- A Review. Soybean Research, 9, 1-30.
- Dias, MD., Pinheiro, VF., Cafe-Filho, AC., 2016. Impact of anthracnose on the yield of soybean subjected to chemical control in the north region of Brazil. Summa Phytopathologica. 42, 1, 18-23. [CrossRef]
- Dias, MD., Dias-Neto, JJ., Santos, MDM., Formento, AN., Bizerra, LVAS., Fonseca, MEN., Boiteux, LS., Café-Filho, AC., 2019. Current Status of Soybean Anthracnose Associated with Colletotrichum truncatum in Brazil and Argentina. Plants (Basel). 8, 11, 459. [CrossRef]
- Silva, AC., Souza, PE., Machado, JC., Silva, BM., Pinto, JEBP., 2012. Effectiveness of essential oils in the treatment of Colletotrichum truncatum infected soybean seeds. Topical Plant Pathology. 37, 305-313. [CrossRef]
- Galli, JÁ., Panizzi, RC., Vieira, RD., 2007. Resistência de variedades de soja à morte de plântulas causada por Colletotrichum truncatum. Arquivos do Instituto Biológico, 74, 163-165. [CrossRef]
- Sharma, SK., Gupta, GK., Ramteke, R., 2011. Colletotrichum truncatum [(Schw.) Andrus and W.D. Moore], the causal agent of anthracnose of soybean [Glycine max (L.) Merrill.] –a review. Soybean Research, 9, 31-52. https://www.apsnet.org/publications/phytopathology/backissues/Documents/1974Articles/Phyto64n01_154.pdf.
- Júnior, MBd., Resende, MLV., Pozza, EA., 2021. Effect of temperature on Colletotrichum truncatum growth, and evaluation of its inoculum potential in soybean seed germination. Eur J Plant Pathol, 160, 999–1004. [CrossRef]
- Boufleur, TR., Ciampi-Guillardi, M., Tikami, Í., Rogério, F., Thon, MR., Sukno, SA., Massola Júnior, NS., Baroncelli, R., 2021. Soybean anthracnose caused by Colletotrichum species: Current status and future prospects. Mol Plant Pathol. 22, 4, 393-409. [CrossRef]
- Pereira, CE., Oliveira, JÁ., Rosa, MCM., Oliveira, GE., Neto, JC., 2009. Tratamento fungicida de sementes de soja inoculadas com Colletotrichum truncatum. Ciência Rural, 39, 2390-2395. [CrossRef]
- Yang, HC., Hartman, GL., 2015. Methods and evaluation of soybean genotypes for resistance to Colletotrichum truncatum. Plant Disease, 99, 143-148. [CrossRef]
- Poti, T., Mahawan, K., Cheewangkoon, R., Arunothayanan, H., Akimitsu, K., Nalumpang, S., 2020. Detection and molecular characterization of carbendazim-resistant Colletotrichum truncatum isolates causing anthracnose of soybean in Thailand. Journal of Phytopathology, 168, 267–278.
- Mello, FE., Mathioni, SM., Matos, VORL., 2024. Sensitivity of Colletotrichum plurivorum and C. truncatum isolated from soybean in Brazil to SDHIs and DMIs fungicides. Trop. plant pathol. 49, 83–92. [CrossRef]
- Begum, MM., Sariah, M., Puteh, AB., Zainal Aabidin, MA., Rahman, MA., Siddiqui, Y., 2010. Field performance of bio-primed seeds to supress Colletotrichum truncatum causing dampping-off and seedling stand of soybean. Biological Control. 53, 18-23. [CrossRef]
- Dalio, RJD., Ribeiro Júnior, PM., Resende, MLV., Silva, AC., Blumer, S., Pereira, VF., Osswald, W., Pascholati, SF., 2012. O triplo modo de ação dos fosfitos em plantas. In: Wilmar C. Luz. (Org.). Revisão Anual de Patologia de Plantas, 20, 206-242.
- Buffara, CRS., Angelotti, F., Tessmann, DJ., Souza, CD., Vida, JB., 2013. Atividade de fosfito de potássio na pré e pós-infecção de Phakospsora euvitis em folhas de videira. Semina: Ciências Agrárias, 34, 6, 3333-3340. [CrossRef]
- Magalhães, LPP., de Lima Pereira Sales, N., Barroso, PD., da Silva, RAF., Pinho, DB., Zanuncio, JC., Silva, AC., 2024. Pseudoplagiostoma humilis sp. nov., a New Fungal Species Causing Shoot Blight and Dieback in Anacardium humile in Brazil. Curr Microbiol 81, 378. [CrossRef]
- Brasil. 2009. Ministério da Agricultura, Pecuária e Abastecimento. Regras para análise de sementes. Brasília: Secretaria de Defesa Agropecuária. Mapa/ACS. 399 p.
- Machado, JC., Oliveira, JÁ., Vieira, MGCG., Alves, MC., 2001. Inoculação artificial de sementes de soja por fungos, utilizando solução de manitol. Revista Brasileira de Sementes, Londrina, 23, 2, 95-101. [CrossRef]
- Michel, BE., Radcliffe, DA., 1995. Computer program relating solute potential to solution composition for five solutes. Agronomy Journal, 87, 1, 131-136. [CrossRef]
- Maguire, JD., 1962. Speed of germination-aid in selection and evaluation for seedling emergence and vigor. Crop Science, 2, 2, 176-177. [CrossRef]
- Araújo, L., Valdebenito-Sanhueza, RM., Stadnik, MJ., 2010. Avaliação de formulações de fosfito de potássio sobre Colletotrichum gloeosporioides in vitro e no controle pós-infeccional da mancha foliar de Glomerella em macieira. Tropical Plant Pathology. v. 35, n. 1, p. 54-59. [CrossRef]
- Araújo, L., Stadnik, MJ., Borsato, LC., Valdebenito-Sanhueza, RM., 2008. Fosfito de potássio e ulvana no controle da mancha foliar da gala em macieira. Tropical Plant Pathology. 33, 2, 148-152. [CrossRef]
- Torres-Calzada, C., Tapia-Tussel, R., Higuera-Ciapara, I., Nexticapan-Garcez, A., Perez- Brito, D., 2015. Sensitivity of Colletotrichum truncatum to four fungicides and characterization of thiabendazole-resistant isolates. Plant Disease. 99, 1590-1595. [CrossRef]
- Bradford, MM., 1976. A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical Biochemistry, Washington, 72, 1/2, 248-254. [CrossRef]
- Urbanek, H., Kuzniak-Gebarowska, E., Herka, H., 1991. Elicitation of defence responses in common bean leaves by Botrytis cinerea polygalacturonase. Acta Phisiologiae Plantarum. 13, 1, 43-50.
- Chance, B., Maehley, AC., 1955. Assay of catalases and peroxidases. Methods in Enzymology, 2, 764-775.
- Havir, EA., McHale, NA., 1987. Biochemical and developmental characterization of multiple forms of catalase in toacco leaves. Plant Physiology, 84, 2, 450-455. [CrossRef]
- Gianopolitis, CN., Ries, SK., 1977. Superoxide dismutases: I. Occourrence in higher plants. Plant Phisiology, 59, 2, 309-314. [CrossRef]
- Shaner, G., Finney, RF., 1977. The effect of nitrogen fertilization on the expression of slow-mildewing resistance in knox wheat. Phytopathology, 67, 1051-1056.
- Doster, MA., Bostock, RM., 1988. Quantification of lignin formation in almond bark in response to wounding and infection by Phytophthora species. Phytopathology, 78, 473-477. https://www.apsnet.org/publications/phytopathology/backissues/Documents/1988Articles/Phyto78n04_473.PDF.
- Spanos, GA., Wrolstad, RE., 1990. Influence of processing and storage on the phenolic composition of Thompson sedless grape juice. Journal of Agricultural & Food Chemistry, 38, 7, 1565-1571. https://pubs.acs.org/doi/10.1021/jf00097a030.
- R Development Core Team, 2009. R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing.
- Yáñez-Juárez, MG., López-Orona, CA., Ayala-Tafoya, F., Partida-Ruvalcaba, L., Velázquez-Alcaraz, TJ., Medina-López, R., 2018. Phosphites as alternative for the management of phytopathological problems. Revista mexicana de fitopatología. 36, 1, 79-94. [CrossRef]
- Carmona, MA., Sautua, FJ., Grijalba, PE., Cassina, M., Pérez-Hernández, O., 2017. Effect of potassium and manganese phosphites in the control of Pythium damping-off: a feasible alternative to fungicide seed treatments. Pest Management Science. 74, 366-374. 10.1002/ps.4714.
- da Silva, AC., Resende, MLV., de Souza, PE., Silva, NCN., Silva, MB., Vitorino, LRR., 2013. Coffee-leaf extract and phosphites on the curative control of powdery mildew in eucalyptus mini-stumps. For. Path., 43, 297-305. [CrossRef]
- Yogev, E., Sadowsky, A., Solel, Z., Oren, Y., Orbach, Y., 2006. The performance of potassium phosphite for controlling Alternaria brown spot of citrus fruit. J. Plant Dis. Prot. 113, 207-213. [CrossRef]
- Monteiro, ACA., Resende, MLV., Valente, TCT., Ribeiro Júnior, PM., Pereira, VF., Costa, JR., Silva, JAG., 2016. Manganese phosphite in coffee defence against Hemileia vastatrix, the coffee rust fungus: Biochemical and molecular analyses. Journal of Phytopathology. 164, 1043-1053. [CrossRef]
- Silva, OC., Santos, HAA., Dalla Pria, M., May-De Mio, LL., 2011. Potassium phosphite for control of downy mildew of soybean. Crop Protection. 30, 598-604. [CrossRef]
- Abbasi, PA., Lazarovits, G., 2006. Seed treatment with phosphonate (AG3) suppresses Pythium damping-off of cucumber seedlings. Plant Disease. 90, 459-464. [CrossRef]
- Mayton, H., Amirkhani, M., Loos, M., Johnson, B., Fike, J., Johnson, C., Myers, K., Starr, J., Bergstrom, GC., Taylor, A., 2022. Evaluation of Industrial Hemp Seed Treatments for Management of Damping-Off for Enhanced Stand Establishment. Agriculture. 12, 591. [CrossRef]
- Lobato, MC., Olivieri, FP., González Altamiranda, EA., Wolski, EA., Daleo, GR., Caldiz, DO., Andreu, AB., 2008. Phosphite compounds reduce disease severity in potato seed tubers and foliage. European Journal of Plant Pathology. 122, 349-358. [CrossRef]
- Simonetti, E., Pin Viso, N., Montecchia, M., Zilli, C., Balestrasse, K., Carmona, M., 2015. Evaluation of native bacteria and manganese phosphite for alternative control of charcoal root rot of soybean. Microbiological Research. 180, 40-48. [CrossRef]
- McDonald, AE., Grant, BR., Plaxton, WC., 2001. Phosphite (phosphorous acid): its relevance in the environment and agriculture and influence on plant phosphate starvation response. Journal Plant Nutrition, 24, 1505-1519. [CrossRef]
- Tkaczyk, M., Kubiak, KA., Sawicki, J., Nowakowska, JA., Oszako, T., 2016. The use of phosphates in forestry. Forest Research Papers. 77, 76-81. [CrossRef]
- Whiley, AW., Hargreaves, PA., Pegg, KG., Doogan, VJ., Ruddle, LJ., Saranah, JB., Langdon, PW., 1995. Changing sink strengths influence translocation of phosphonate in avocado (Persea Americana Mill.) trees. Aust. J. Agric. Res. 46, 1079-1090. [CrossRef]
- Förster, H., Adaskaveg, J., Kim, D., Stanghellini, M., 1998. Effect of Phosphite on Tomato and Pepper Plants and on Susceptibility of Pepper to Phytophthora Root and Crown Rot in Hydroponic Culture. Plant. Dis. 82, 1165–1170. [CrossRef]
- Guest, DI., Grant, BR., 1991. The complex action of phosphonates as antifungal agents. Biol. Rev. 66, 159-187.
- Malusa, E., Tosi, L., 2005. Phosphorous acid residues in apples after foliar fertilization: results of field trials. Food Addit. Contam. 22, 541-548. https://pubmed.ncbi.nlm.nih.gov/16019827/.
- Marks, GC., Smith, W., 1992. Metalaxyl and phosphonate as prophylactic and curative agents against stem infection of Leucadendron caused by Phytophthora cinnamomi. Aust. J. Exp. Agric. 32, 255-259. [CrossRef]
- Howard, K., Dell, B., Hardy, GE., 2000. Phosphite and mycorrhizal formation in seedlings of three Australian Myrtaceae. Aust J Bot. 48, 6, 725-729. [CrossRef]
- Tambascio, C., Covacevich, F., Lobato, MC., De Lasa, C., Caldiz, D., Dosio, G., Andreu, A., 2014. The application of K phosphites to seed tubers enhanced emergence, early growth and mycorrhizal colonization in potato (Solanum tuberosum). Am J Plant Sci. 5, 132-137. [CrossRef]
- Martinez, S., 2016. Effects of combined application of potassium phosphite and fungicide on stem and sheath disease control, yield, and quality of rice. Crop Prot. 89, 259-264. [CrossRef]
- Liljeroth, E., Lankinen, Å., Wiik, L., Burra, DD., Alexandersson, E., Andreasson, E., 2016. Potassium phosphite combined with reduced doses of fungicides provides efficient protection against potato late blight in large-scale field trials. Crop Prot. 86, 42-55. [CrossRef]
- Puerari, HH., Dias-Arieira, C., Tavares Silva, CA., Arieira, JO., Biela, F., Poletine, JP., 2013. Ecolife nd manganese phosphite in the control of Meloidogyne javanica and in the development of soybean cultivars susceptible and resistant to the nematode. Nematropica. 43, 105-112. https://www.cabidigitallibrary.org/doi/pdf/10.5555/20133372345.
- Silva, AC., Resende, MLV., Souza, PE., Possa, KF., Júnior, MBS., 2016. Plant extract, zinc phosphite and zinc sulphate in the control of powdery mildew in the eucalyptus. Revista Ciência Agronômica, 47, 1, 93-100. [CrossRef]
- King, M., Reeve, W., Van der Hoek, MB., Williams, N., McComb, J., O’Brien, PA., Hardy, GE., 2010. Defining the phos phite-regulated transcriptome of the plant pathogen Phyto phthora cinnamomi. Mol Genet Genomics 284:425-35. [CrossRef]
- Lobato, MC., Olivieri, FP., Daleo, GR., Andreu, AB., 2010. Antimicrobial activity of phosphites against different potato pathogens. Journal of Plant Diseases and Protection. 117, 102-109. [CrossRef]
- Dobrowolski, MP., Shearer, BL., Colquhoun, IJ., O’Brienand, PA., Hardy, GESTJ., 2008. Selection for decreased sensitivity to phosphite in Phytophthora cinnamomi with prolonged use of fungicide. Plant. Pathol. 58, 928-936. [CrossRef]
- Brown, S., Koike, ST., Ochoa, OE., Laemmlen, F., Michelmore, RW., 2004. Insensitivity to the fungicide fosetyl-aluminum in California isolates of the lettuce downy mildew pathogen Bremia lactucae. Plant Dis. 88, 502-508. [CrossRef]
- Duvenhage, JA., 1994. Monitoring the Resistance of Phytophthora cinnamomito Fosetyl-Al and H3PO3. Duivelskloof: Yearbook, South African Avocado Growers’ Association. 17, 35-37.
- Perez, V., Mamdouh, AM., Huet, JC., Pernollet, JC., Bompeix, G., 1995. Enhanced secretion of elicitins by Phytophthora fungi exposed to phosphonate. Cryptogamie. Mycologia. 16, 191-994. ⟨hal-02707440⟩.
- Pilbeam, RA., Howard, K., Shearer, BL., Hardy, GEJ., 2011. Phosphite stimulated histological responses of Eucalyptus marginata to infection by Phytophthora cinnamomi. Tree. 25, 1121-1131. [CrossRef]
- Machinandiarena, MF., Lobato, MC., Feldman, ML., Daleo, GR., Andreu, AB., 2012. Potassium phosphite primes defense responses in potato against Phytophthora infestans. Journal of Plant Physiology, 169, 14, 1417-1424. [CrossRef]
- Han, X., Xi, Y., Zhang, Z., Mohammadi, M., Joshi, J., Borza, T., Wang-Pruski, G., 2021. Effects of phosphite as a plant biostimulant on metabolism and stress response for better plant performance in Solanum tuberosum. Ecotox. Environ. Safety 210, 111873. [CrossRef]
- Daniel, R., Guest. D., 2006. Defence responses induced by potassium phosphonate in Phytophthora palmivora-cha llenged Arabidopsis thaliana. Physiologycal and Molecular Plant Pathology, 67, 194-201. [CrossRef]
- Zhao, H., Wu, L., Chai. T., Zhang. Y., Tan, J., Ma, S., 2012. The effects of copper, manganese and zinc on plant growth and elemental accumulation in the manganese- hyperacumulatior Phytolacca americana. Journal of Plant Physiology. 169, 1243-1252. [CrossRef]
- Demirevska-Kepova. K., Simova-Stoilova, L., Stoyanova, Z., Holzer, R., Feller, U., 2004. Biochemical changes in barley plants after excessive supply of copper and manganese. Enviromental and Experimental Botany, 52, 3, 253-266. [CrossRef]
- Rengel, Z., Graham, RD., Pedler, JF., 1993. Manganese nutrition and accumulation of phenolics and lignina as related to differential resistance of wheat genotypes to the take-all fungus. Plant and Soil, 151, 255-263. [CrossRef]
- Dordas, C., 2008. Role of nutrients in controlling plant diseases in sustainable agriculture: A review. Agronomy for Sustainable Development, 28, 1, 33-46. [CrossRef]
- Stangarlin, JR., Kuhn, OJ., Toledo, MV., Portz, RL., Schan-Estrada, KRF., Pascholati, SF., 2011. A defesa vegetal contra fitopatógenos. Scientia Agraria Paranaensis, 10, 18-46.
- Griffith, J., Coffey, M., Grant, B., 1993. Phosphonate inhibition as a function of phosphate concentration in isolates of Phytophthora palmivova. J. Gen. Microbiol. 139, 2109-2116. [CrossRef]
- Varadarajan, DK., Karthikeyan, AS., Matilda, PD., Raghothama, KG., 2002. Phosphite, an analog of phosphate, suppresses the coordinated expression of genes under phosphate starvation. Plant Physiol. 129, 3, 1232-1240. [CrossRef]
- Thao, HTB., Yamakawa, T., 2009. Phosphite (phosphorous acid): Fungicide, fertilizer or bio-stimulator? Soil Sci. Plant Nutr. 55, 2, 228-234. [CrossRef]
- Rossall, S., Qing, C., Paneri, M., Bennett, M., Swarup, R., 2016. A ‘growing’ role for phosphites in promoting plant growth and development. Acta Hortic. 1148. [CrossRef]
- Mohammadi, M., Zhang, Z., Xi, Y., Han, H., Lan, F., Zhang, B., Wang-Pruski, G., 2019. Effects of potassium phosphite on biochemical contents and enzymatic activities of Chinese potatoes inoculated by Phytophthora infestans. Appl. Ecol. Environ. Res, 17, 4499-4514. [CrossRef]
- Mohammadi, M., Han, X., Zhang, Z., Xi, Y., Boorboori, M., Wang-Pruski G., 2020. Phosphite.
- Application Alleviates Pythophthora infestans by Modulation of Photosynthetic and Physio-Biochemical Metabolites in Potato Leaves. Pathogens. 9, 170.
Figure 1.
Doses of manganese (A) and zinc (B) phosphites on the mycelial growth index (mm/day) of Colletotrichum truncatum.
Figure 1.
Doses of manganese (A) and zinc (B) phosphites on the mycelial growth index (mm/day) of Colletotrichum truncatum.
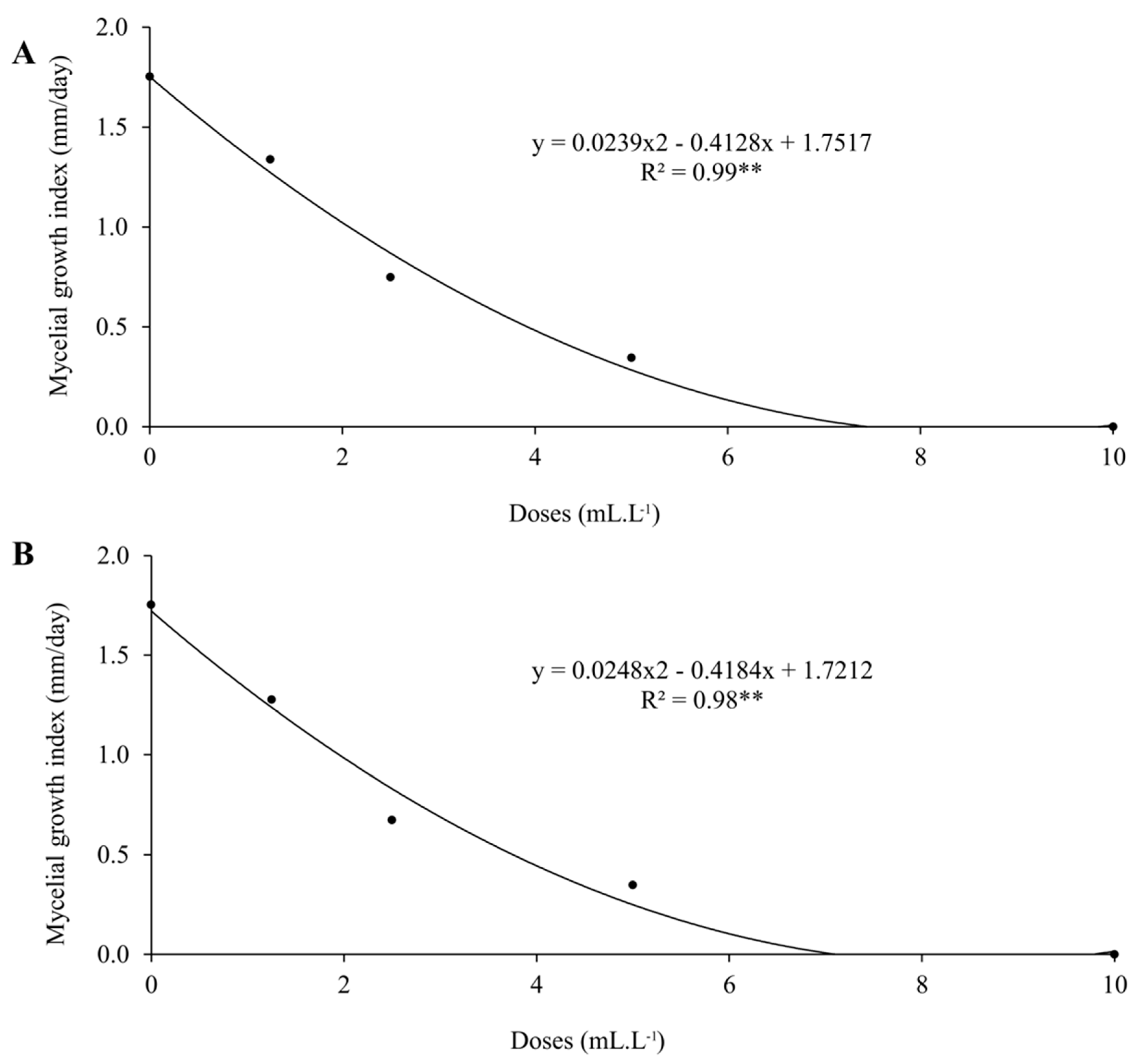
Figure 2.
Dunnett test comparing the tested doses of Mn phosphite (A) and Zn phosphite (B) with the fungicide carbendazim + thiram.
Figure 2.
Dunnett test comparing the tested doses of Mn phosphite (A) and Zn phosphite (B) with the fungicide carbendazim + thiram.
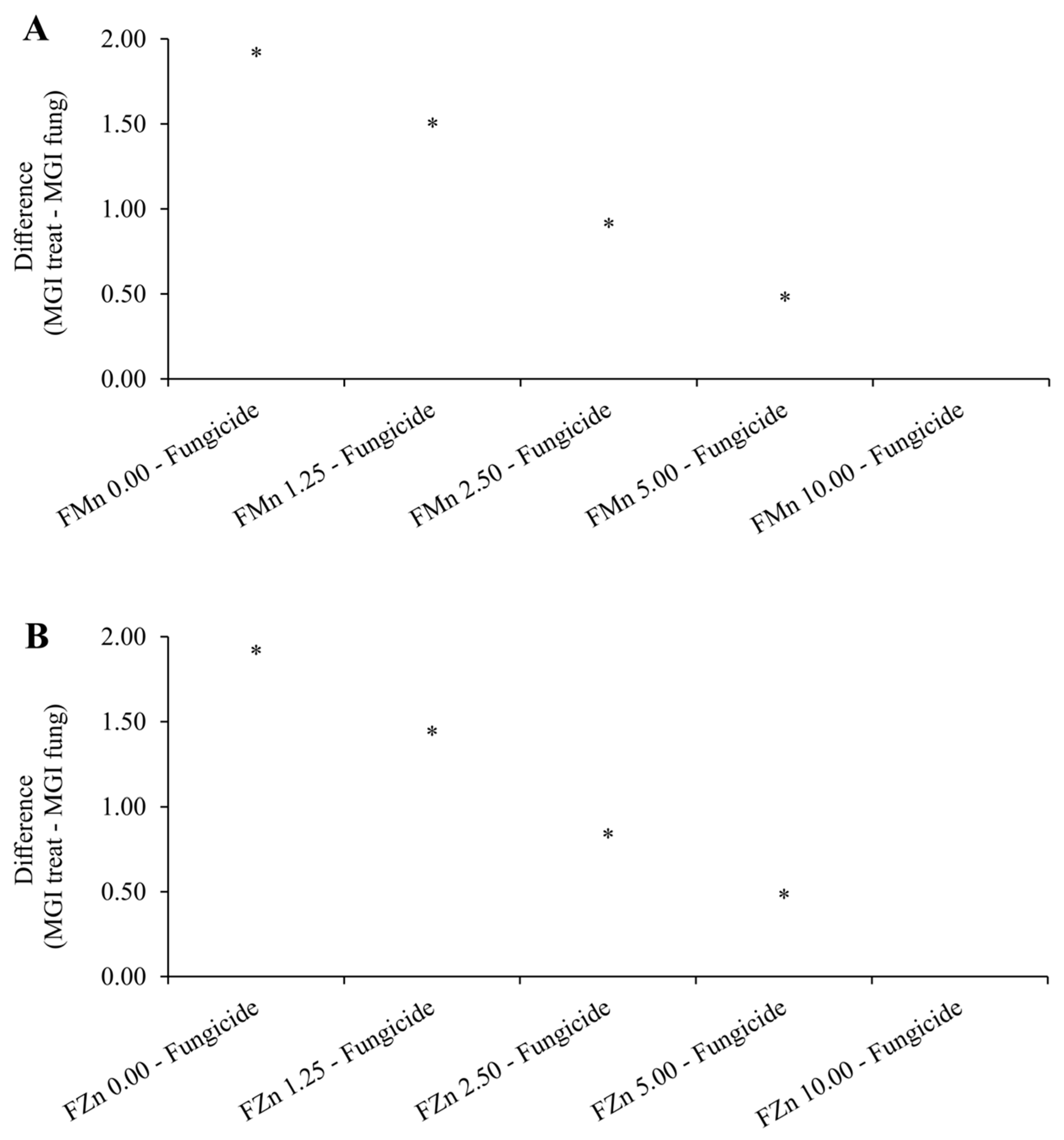
Figure 3.
Activation of defense-related enzymes in soybean seeds inoculated with Colletotrichum truncatum and treated with Mn and Zn phosphite after incubation in paper rolls. (A) Catalase (CAT) activity, (B) Peroxidase (POX) activity, and (C) Superoxide dismutase (SOD) activity over 192 hours, and (D) area under the progress curve for catalase (AUAPCCat), peroxidase (AUAPCPox), and superoxide dismutase (AUAPCSod). Columns with the same letter above did not differ according to the Tukey test, P ≤ 0.05.
Figure 3.
Activation of defense-related enzymes in soybean seeds inoculated with Colletotrichum truncatum and treated with Mn and Zn phosphite after incubation in paper rolls. (A) Catalase (CAT) activity, (B) Peroxidase (POX) activity, and (C) Superoxide dismutase (SOD) activity over 192 hours, and (D) area under the progress curve for catalase (AUAPCCat), peroxidase (AUAPCPox), and superoxide dismutase (AUAPCSod). Columns with the same letter above did not differ according to the Tukey test, P ≤ 0.05.
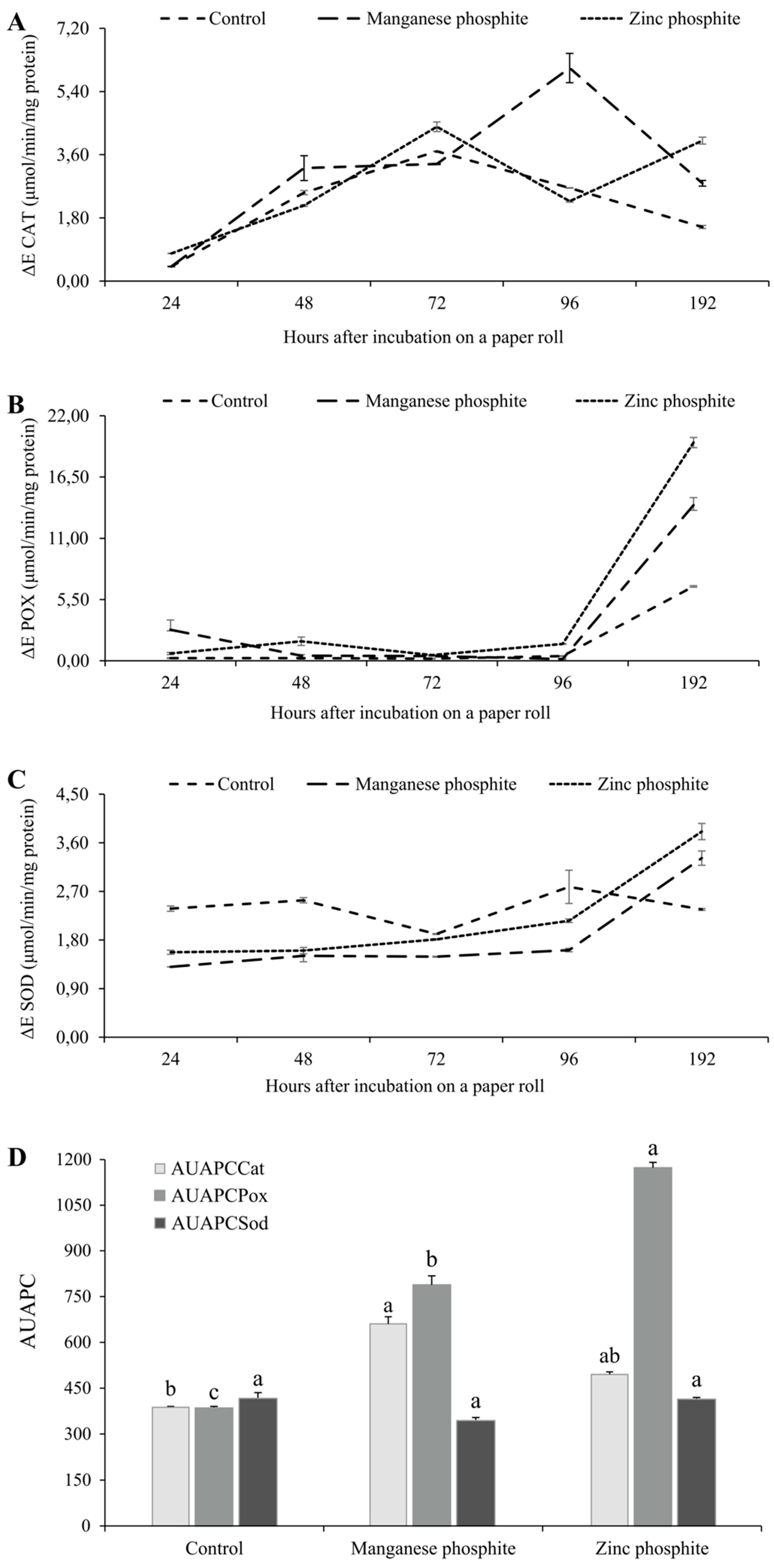
Figure 4.
Total soluble phenolic content and soluble lignin content in seeds infected with Colletotrichum truncatum and treated with Zn and Mn phosphite. Columns with the same letter above did not differ according to the Tukey test, P ≤ 0.05.
Figure 4.
Total soluble phenolic content and soluble lignin content in seeds infected with Colletotrichum truncatum and treated with Zn and Mn phosphite. Columns with the same letter above did not differ according to the Tukey test, P ≤ 0.05.
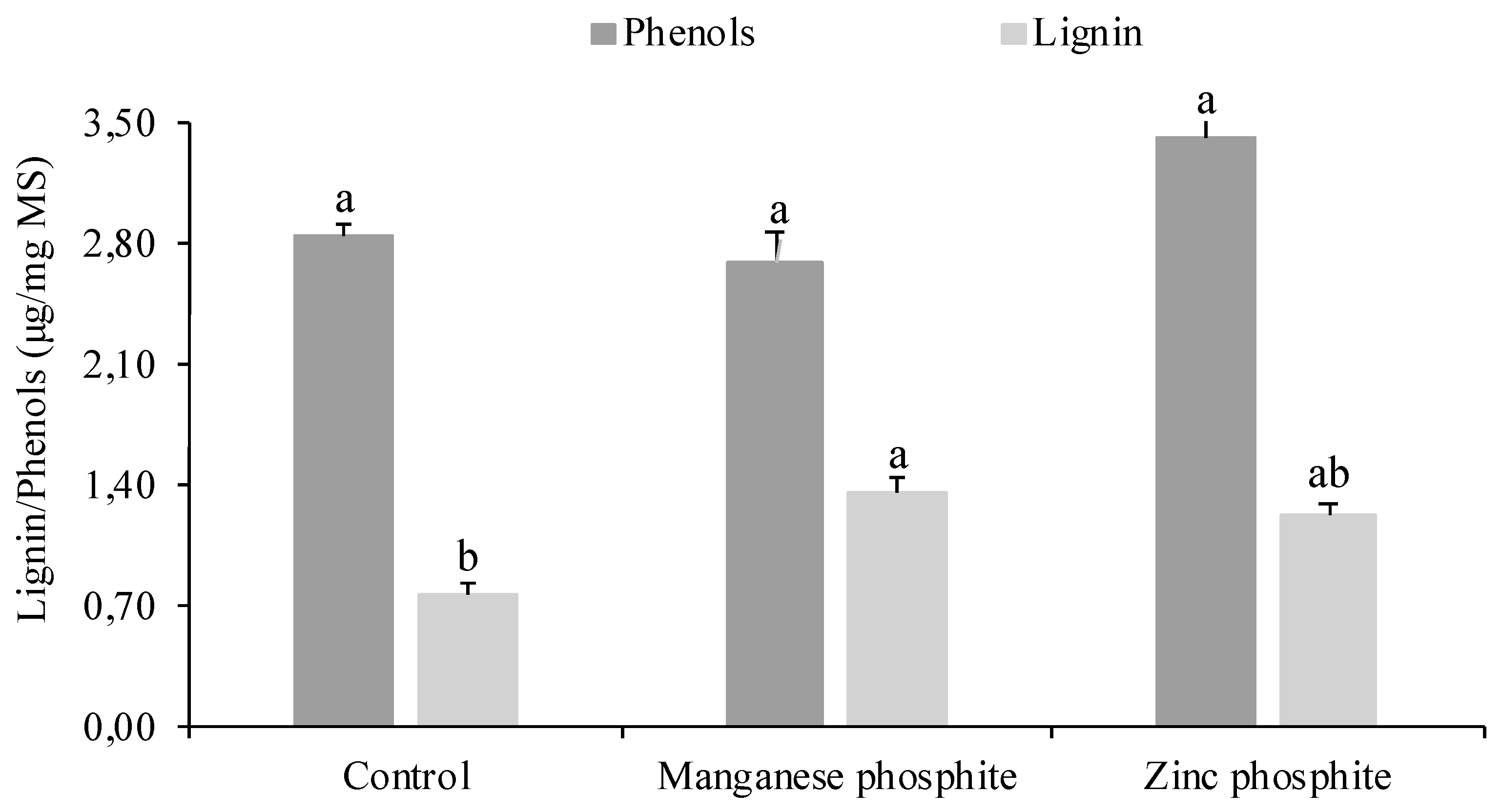

Table 1.
Percentage of germination and sanitary quality of soybean seeds non-inoculated and inoculated with Colletotrichum truncatum and treated with Cu, K, Zn, and Mn phosphites and with the fungicide Protreat®.
Table 1.
Percentage of germination and sanitary quality of soybean seeds non-inoculated and inoculated with Colletotrichum truncatum and treated with Cu, K, Zn, and Mn phosphites and with the fungicide Protreat®.
| Treatments | Germination (%) | Incidence (%) | Incidence Reduction (%) |
|---|---|---|---|
| Inoculated Control | 25 ± 1.0d* | 96 ± 3.7e* | 0 |
| Uninoculated control | 85 ± 3.4a | 4 ± 4.4a | 95 |
| Fungicide | 83 ± 10.8a | 27 ± 6.9b | 72 |
| Copper phosphite | 69 ± 3.4b | 70 ± 7.2d | 27 |
| Manganese phosphite | 81 ± 3.4a | 29 ± 5.7b | 69 |
| Potassium phosphite | 59 ± 10.8c | 49 ± 4.8c | 49 |
| Zinc phosphite | 85 ± 5.2a | 33 ± 4.1b | 65 |
*Mean values ± standard errors obtained from eight replicates. The means followed by same lower-case letter in the column do not differ by the Scott Knott test, P ≤ 0.05.
Table 2.
Emergence speed index (ESI), initial stand at seven days (IS), final stand at 14 days (FS), plant height (H), root length (R), dry biomass of the aerial part (BA) and roots (BR) at 28 days, of seedlings derived from soybean seeds inoculated or not with Colletotrichum truncatum, subjected to different treatments based on Cu, K, Zn, and Mn phosphites and the fungicide Protreat®.
Table 2.
Emergence speed index (ESI), initial stand at seven days (IS), final stand at 14 days (FS), plant height (H), root length (R), dry biomass of the aerial part (BA) and roots (BR) at 28 days, of seedlings derived from soybean seeds inoculated or not with Colletotrichum truncatum, subjected to different treatments based on Cu, K, Zn, and Mn phosphites and the fungicide Protreat®.
| Treatments | ESI* | IS* (%) | FS* (%) | H*(cm) | R*(cm) | BA*(g) | BR*(g) |
|---|---|---|---|---|---|---|---|
| Control inoculated | 21.5 ± 1.2d* | 32.2 ± 1.5e* | 34.0 ± 4.1e* | 24.3 ± 0.7c* | 20.4 ± 2.5c* | 2.9 ± 0.4c* | 1.6 ± 0.1d* |
| Uninoculated control | 65.7 ± 2.8a | 93.0 ± 2.9a | 95.0 ± 2.1a | 28.9 ± 1.7b | 32.8 ± 1.1a | 7.5 ± 0.7a | 5.1 ± 0.6a |
| Fungicide | 45.9 ± 3.2b | 71.0 ± 3.5b | 76.7 ± 5.6b | 30.3 ± 0.7a | 30.2 ± 3.0a | 6.8 ± 0.7a | 3.8 ± 0.7b |
| Copper phosphite | 33.3 ± 5.4c | 51.5 ± 7.0d | 55.0 ± 9.6d | 28.6 ± 1.0b | 25.0 ± 2.1b | 4.6 ± 0.6b | 2.6 ± 0.8c |
| Manganese phosphite | 44.1 ± 2.7b | 62.5 ± 7.5c | 80.2 ± 1.2b | 30.5 ± 1.6a | 30.2 ± 0.9a | 7.0 ± 0.3a | 4.2 ± 0.2b |
| Potassium phosphite | 39.0 ± 5.8c | 60.0 ± 8.2c | 64.0 ± 6.9c | 30.2 ± 2.5a | 24.4 ± 4.0b | 5.8 ± 0.97a | 2.4 ± 0.3c |
| Zinc phosphite | 47.7 ± 2.9b | 75.7 ± 5.3b | 80.0 ± 3.3b | 31.9 ± 1.6a | 30.5 ± 0.1a | 6.5 ± 0.2a | 3.7 ± 0.4b |
*Mean values ± standard errors obtained from eight replicates. Means followed by same lower-case letter in the column do not differ by the Scott Knott test, P ≤ 0.05.
Table 3.
Lethal dose (LD50) of Mn and Zn phosphites that inhibited 50% of the mycelial growth of Colletotrichum truncatum and the maximum concentration for mycelial growth inhibition (MCI).
Table 3.
Lethal dose (LD50) of Mn and Zn phosphites that inhibited 50% of the mycelial growth of Colletotrichum truncatum and the maximum concentration for mycelial growth inhibition (MCI).
| Treatments | LD50 (mL.L-1) | MCI (mL.L-1) |
|---|---|---|
| Manganese phosphite | 2.48 | 8.64 |
| Zinc phosphite | 2.35 | 8.44 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



