Submitted:
30 January 2025
Posted:
04 February 2025
You are already at the latest version
Abstract
Ecological and genetic studies of malaria mosquitoes of the Black Sea coast have not been recently conducted despite increasing human-caused environmental changes in the area. In this study, we investigated the species composition, geographical distribution, ecological preferences, and chromosomal polymorphism of malaria mosquitoes of the Crimean Peninsula and the Black Sea coast of the Caucasus. Species were diagnosed using a combination of morphological, cytogenetic, and molecular markers. The ecological conditions of the larval habitats, such as dissolved oxygen content in the water, acidity, salinity, and temperature, were measured. Seven species of malaria mosquitoes were identified in the pool of 2229 individual mosquitoes collected at 56 breeding sites including An. atroparvus, An. claviger, An. daciae (formely identified as An. messeae s. l.), An. hyrcanus, An. maculipennis s. s., An. plumbeus and An. melanoon. The previously recorded species of An. algeriensis, An. messeae s. s., An. sacharovi, An. superpictus were not found in this study. Anopheles maculipennis was dominant in typical anophylogenic water bodies. Anopheles plumbeus, which used to breed mainly in tree holes in coastal forests, has spread to urban settlements along the Black Sea coast and breeds in artificial containers. Chromosomal polymorphism was studied and found in An. atroparvus and An. daciae populations. Differences in the chromosomal composition of An. daciae populations in Crimea and on the Black Sea coast of the Caucasus were revealed. The Crimean population had a low level of polymorphism in autosomal inversions. The data obtained in this study can be used to inform a better control of potential malaria vectors in the Black Sea coastal region.
Keywords:
Anopheles
; The Crimean Peninsula
; Black Sea coast of Caucasus
; distribution
; breeding sites
; chromosomal polymorphism
1. Introduction
The environmental and ecological conditions of the Crimean Peninsula and the Black Sea coast of the Caucasus have changed dramatically over the last 20 years [1,2]. Human population growth, expansion of economic activities and increased anthropogenic pressure from resort development, have led to the destruction and further degradation of natural ecosystems and landscapes, and massive environmental pollution. In addition, global warming has an increasingly strong impact on ecosystems, thus constantly changing the epidemiologic situation in the world, including those at the Black Sea coast [3,4]. Although the implementation of antimalarial interventions led to the elimination of autochthonous malaria in Europe [5,6], the territories of Crimea and the Black Sea coast of the Caucasus are currently classified as high-risk areas for resurgence of malaria transmission [7]. Cases of imported malaria are reported annually in the Crimea and Krasnodar Krai [8,9,10]. Therefore, understanding the current state of epidemiologically important groups of insects, such as malaria mosquitoes of the genus Anopheles (Diptera, Culicidae), has become urgent.
Most of the data on species composition and geographical distribution of malaria mosquito in the southern European part of Russia were collected between 1920 and 1960 [11] and are largely out of date. Moreover, the taxonomic status of mosquitoes in the Maculipennis complex has changed. Originally, the sibling species of this complex were considered as subspecies of the broadly polytypic species An. maculipennis s. l. [12]. Hybridization experiments and numerous data on morphology, ecology, and cytogenetics supported the taxonomic status of the following species of the Maculipennis group: An. atroparvus Van Theil, 1927; An. maculipennis Meigen, 1818; An. messeae Falleroni, 1926; An. melanoon Hackett, 1934; An. sacharovi Favre, 1903; An. subalpinus Hackett & Lewis, 1937 [13,14,15,16,17,18]. Later, several sibling species of the Maculipennis group were diagnosed using cytogenetic analysis [13,19,20,21,22,23]. Two new species of Palearctic malaria mosquitoes have been distinguished by differences in banding patterns of polytene chromosomes: An. beklemishevi Stegniy & Kabanova, 1976 [24] and An. martinius Shingarev, 1926 [25]. The reproductive isolation of new species has been demonstrated by studying experimental interspecific hybrids [26] obtained by the method of forced mating [27].
Molecular genetic methods provided additional opportunities to discover new malaria mosquito species and became an important approach in determining their taxonomic status [28,29,30]. The most valuable data for Anopheles systematics have been obtained by studying the nucleotide composition of the second internal transcribed spacer (ITS2) of the ribosomal gene cluster that separates the 5,8S and 28S genes of ribosomal RNA [31]. Three new species have been identified in the Maculipennis complex based on ITS2 sequence: An. persiensis Linton, Sedaghat & Harbach, 2003 [32,33]; An. daciae Linton, Nicolescu & Harbach, 2004 [34]; An. artemievi Gordeev, Zvantsov, Goryacheva, Shaikevich & Yezhov, 2005 [35]. Anopheles subalpinus has been synonymized with An. melanoon because the ITS2 sequences were identical in both species [36]. In addition, the ITS2 region of ribosomal DNA served as a tool for reconstructing phylogenetic relationships in malaria mosquitoes [37,38,39,40,41,42]. Further advances in species recognition and phylogeny reconstruction in the Maculipennis group of the malaria mosquitoes have been achieved through whole-genome sequencing [43,44].
According to 2003 and 2008 studies, the list of malaria mosquitoes of the North Caucasus and the Southern Russian Plain included 10 species [45,46,47]. Nine of which belong to the subgenus Anopheles: An. algeriensis Theobald, 1903; An. atroparvus Van Theil, 1927; An. claviger Meigen, 1804; An. hyrcanus Pallas, 1771; An. maculipennis Meigen, 1818; An. melanoon (subalpinus) Hackett, 1934; An. messeae Falleroni, 1926; An. plumbeus Stephens, 1828; An. sacharovi Favre,1903. One species was a member of the subgenus Cellia – An. superpictus Grassi, 1899. However, the present geographical distribution of malaria mosquitoes on the Black Sea Coast of the Caucasus and in Crimea is poorly understood.
The aim of this work was to revisit and describe in detail the species composition, geographic distribution, breeding sites and chromosomal polymorphism of the malaria mosquitoes of the Crimean Peninsula and the Black Sea coast of the Caucasus focusing on the sibling species of the Maculipennis group. The dynamics of the epidemic situation under the conditions of global warming and urbanization in the study area was discussed.
2. Materials and Methods
2.1. Field Collection and Material Preservation
Malaria mosquito larvae were collected from 2009 to 2024 at 56 locations along the Black Sea coast of the Caucasus and the Crimean Peninsula. A total of 2229 individual mosquitoes were included in this study. The coordinates of the collection sites and the number of mosquitoes collected are shown in Table 1. Fourth instar Anopheles larvae were collected from the surface of water by dipping method [48] and then were placed in Clark's solution (glacial acetic acid and 95% ethanol in a 1:3 ratio). Each larva was removed from Clark's solution and was divided into two parts. The head and thorax were placed back into Clark's solution for inversion polymorphism analysis. The abdomen was fixed in 70% ethanol for molecular identification. All samples were placed in a 1.5 ml Eppendorf tube and stored at −20°C. The obtained samples were used for preliminary species identification based on morphological characters [49,50]. Morphologically identical species were identified by cytogenetic and molecular genetic markers.
Ecological characteristics of malaria mosquito breeding sites were determined in local habitats. Water temperature (T), potential of hydrogen (pH), and total salinity (ppt) were measured using a Hanna Combo HI 98129 (Hanna Instruments, Woonsocket, Rhode Island, USA) conductometer. Dissolved oxygen content in water was measured using an ExStik DO600 oximeter (Extech Instruments, Waltham, Massachusetts, USA). The density of larvae of 1-4 stages (number of individuals per m2) was estimated in natural habitats (ponds, lakes, river outfalls).
2.2. Karyotyping
Polytene chromosome preparations were obtained from the salivary glands of fourth instar larvae according to the standard technique [23,51]. Salivary glands were extracted from larval thorax with dissecting needles in Clark's solution. The glands were stained with lacto-aceto-orcein (2% orcein in 80% lactic acid and 100% acetic acid in a 1:1 ratio). The time for lacto-aceto-orsine staining was increased from 40 to 80-90 minutes. The stained glands were squoshed under a coverslip in 50% acetic acid for 15-20 min.. Mosquito karyotypes were analyzed using an Eclipse E200 light microscope (Nikon, BioVitrum, Moscow, Russia). Karyotypes were used to identify sibling species of malaria mosquitoes. Chromosomal inversions were determined by comparing banding patterns with cytogenetic maps of the studied species [52,53]. Differences in inversion frequencies between mosquito populations were assessed using the Chi-square (χ2) test [54]. A total of 2229 individual mosquitoes were karyotyped, of which 75 An. atroparvus, 158 An. claviger, 27 An. hyrcanus, 908 An. maculipennis, 750 An. daciae, 309 An. plumbeus and 2 An. melanoon larvae.
2.3. Genotyping by RLFP-RCR
Karyotyped mosquitoes were used for molecular identification of the sibling species An. daciae/An. messeae. All samples were analyzed individually. Total DNA was extracted from alcohol-fixed larval fragments (abdomen) using the phenol-chloroform method [55]. DNA concentration was determined spectrophotometrically using Implen NanoPhotometer NP80 (Implen, Munich, Germany). The concentration was adjusted to 30–60 ng/µl. Polymerase chain reaction (PCR) was performed in a final volume of 20 μl using EncycloPlus PCR kit (Eurogen, Moscow, Russia) according to the manufacturer's instructions.
Species An. daciae and An. messeae were distinguished by the ITS2 fragment of the rDNA. The ITS2 fragment was amplified with the forward primer its2_vdir: 5′-TGTGAACTGCAGGACACATG-3′, and reverse primer its2_nrev: 5′-ATGCTTAAATTTAGGGGGTA-3′, as described previously [53]. RsaI endonuclease (SibEnzyme, Novosibirsk, Russia) was used for PCR-restriction fragment length polymorphism (RFLP) analysis [53]. PCR product ITS2 in An. daciae has 3 restriction sites for RsaI, the length of restriction fragments is 10, 47, 71 and 307 bp. PCR product ITS2 in An. messeae has 4 restriction sites, the length of restriction fragments is 10, 47, 71, 72 and 235 bp. The obtained PCR products were stained with ethidium bromide and analyzed by electrophoresis in 1.5% agarose gel and TBE buffer.
2.4. Genotyping by Sequencing
The rDNA-ITS2 of mosquitoes from two Crimean and two Caucasian populations were sequenced. Each larval abdomen was homogenized separately in liquid nitrogen. Genomic DNA was extracted using a standard protocol from the Qiagen DNeasy Blood and Tissue Kit (Qiagen, Germantown, MD, USA). DNA elution was performed in 100 μl of water. ITS2 from An. maculipennis s. l. which do not differ significantly in electrophoretic mobility of PCR products (An. messeae, An. daciae, An. atroparvus, and An. maculipennis) was amplified with the forward universal primers: its2_ndir 5′-ATCACTCGGCTCGTGGATCG-3′, or its2_vdir 5′-TGTGAACTGCAGGACACATG-3′ and the reverse primer its2_nrev 5′-ATGCTTAAATTTAGGGGGTA-3′, or its2_rev 5′-ATGCTTAAATTTAGGGGGTAGTC-3′, with modifications [56,57]. The HotStarTaq Plus Master Mix Kit (Qiagen, Germantown, MD, USA) was used for PCR amplification. The PCR mix consisted of a total volume of 20 µl of ~40 ng DNA, 0.5 µM of each forward and reverse primer, and 10 µl of 2× HotStarTaq Plus reaction mix. PCR was performed on thermocycler Applied Biosystems GeneAmp PCR system 2700 (Applied Biosystems, Waltham, Massachusetts, USA) under the following conditions: initial denaturation at 95°C for 5 min, followed by 25–35 cycles of 95°C for 15 s, 58°C for 30 s and 72°C for 30 s and a final elongation step at 72°C for 5 min. The resulting reaction mix was placed in a stand-by mode at 4°C. The amplicons were visualized by gel electrophoresis in a 2% agarose gel. The DNA amplicons were purified using the Wizard™ PCR Clean Up Kit (Promega, Fitchburg, WI, USA). PCR products were sequenced by Sanger using the BigDye Terminator v3.1 Cycle Sequencing Kit (Thermo Fisher Scientific, USA) from suitable forward or reverse primers. The DNA products of the sequencing reactions have been purified by ethanol precipitation and analyzed at the Genomics Core Facility of the Siberian Branch of the Russian Academy of Sciences (http://sequest.niboch.nsc.ru). Nucleotide positions in ITS2 sequences of An. daciae and An. messeae were compared with the reference sequence AY648982 in An. messeae [43]. Only nucleotides AC in positions 412 and 432 were considered as species-specific for An. daciae, respectively [43].
Anopheles melanoon / An. maculipennis s. s. mosquitoes were identified by the BOLD fragment of the mitochondrial COI gene, using the following Folmer primers: 5‘-TTTCAACAAACCATAAGGATATTGG-3’ and 5‘-TATACTTCAGGATGACCAAAAAATCA-3’, which were adapted using the “Primer3” program [58]. Total DNA was extracted individually from alcohol-fixed larval fragments (abdomen) using the phenol-chloroform method [55]. PCR amplification was performed at an annealing temperature of 59°C. PCR was performed in a final volume of 20 μl using EncycloPlus PCR kit (Eurogen, Moscow, Russia). Elution of fragments from the gel was performed using Zymoclean™ Gel DNA Recovery Kit (Zymo Research, Los Angeles, California, USA). The obtained fragments were sequenced by Sanger sequencing. The nucleotide sequence of PCR fragments was determined from forward and reverse primers on a 3500 Genetic Analyzer using BigDye®Terminator v3.1 Cycle Sequencing Kit (Applied Biosystems, Waltham, Massachusetts, USA).
3. Results
3.1. Species Composition, Geographical Distribution and Ecological Preferences
Our study found 7 species of malaria mosquito in the Crimea and the Black Sea coast of the Caucasus: An. atroparvus Van Thiel, 1923; An. claviger Meigan, 1904; An. daciae Linton, Nicolescu & Harbach, 2004; An. hyrcanus Pallas, 1771; An. maculipennis Meigan, 1818; An. melanoon Hackett 1934; An. plumbeus Stephens, 1828 (Table 1). Four of them were sibling species of the Maculipennis group: An. atroparvus, An. daciae, An. maculipennis s. s., and An. melanoon. The geographical distribution of the species is shown in Figure 1. Six of them were present in both regions, one species An. melanoon was found only in the Imereti Valley on the Black Sea coast of the Caucasus (Table 1, location number 51 in the vicinity of Sochi). The previously reported in this area species An. algeriensis, An. messeae s. s., An. sacharovi, An. superpictus were not found [47].
Anopheles maculipennis s. s. mosquitoes dominated everywhere along the Black Sea coast and in the mountainous forest zone of the Crimean Peninsula, as well as in the foothills of the northern Caucasus. This species was found in 28 out of 56 locations (50%). Anopheles maculipennis s. s. larvae were found in most typical Anopheles breeding sites, and mosquitoes of this species alone were found in 14 locations (Table 1). Immature stages developed in ponds, water reservoirs, drainage, and irrigation canals, as well as in temporary pools of water. Mass larval development occurred in habitats with pH 6.48-8.85, but predominantly in the pH range of 7.0-8.4 (Table 2). Total dissolved solids (TDS) of water in larval habitats ranges ranged from 0.15 to 2.56 ppt, rarely exceeding 1.5. Larvae are sensitive to dissolved oxygen in the water. The oxygen content varied in the range of 4.4-10.2 mg/l in most breeding sites. The density of An. maculipennis s. s. larvae was low at oxygen levels of 0.5-2.3 mg/l. Larval development occurred over a wide range of daytime temperatures from 16.5 to 33°C, but the highest density of 160 larvae per square meter was observed at a water temperature of 20.7°C and an oxygen level of 4.5 mg/l (Table 2; location 3).
Anopheles daciae was the most abundant species in the plains located north of the Greater Caucasus Range. Anopheles daciae mosquitoes were recorded on the Black Sea coast of the Caucasus near Novorossiysk and Gelendzhik (Figure 1; locations 41-43). A single population of An. daciae in Crimea was found near Sevastopol (Figure 1; location 10). The larvae of An. daciae developed separately (Table 1; locations 40, 41) or together with An. maculipennis s. s. (Table 1; locations 10, 34-37, 42, 43). In some breeding sites shared with An. maculipennis s. s., An. daciae larvae dominated. At An. daciae breeding sites, water pH ranged from 7.0 to 8.4; TDS varied between 0.20 and 1.58 ppt; dissolved oxygen content varied between 0.5 and 8.1 mg/L; and water temperature ranged from 20.5 to 33.3°C (Table 2).
Anopheles atroparvus mosquitoes were found in steppe biotopes in the Crimea and the North Caucasus. Only this species larvae were able to develop in a highly saline water reservoirs with a TDS of 5.99 ppt (Table 2; location 33). Anopheles atroparvus larvae were found together with An. maculipennis s. s. and An. daciae mosquitoes in ponds and lakes with less saline water with TDS range of 0.59-1.46 ppt (Table 2; locations 10, 32). pH value at An. atroparvus breeding sites ranged from 7.14 to 9.1; daytime water temperature ranged from 18.0 to 28.0°C; and dissolved oxygen content ranged from 7.7 to 16.5 mg/L.
We found a single breeding site of An. melanoon mosquitoes in the Imereti Valley near Sochi (Figure 1; location 51). Two larvae were caught in a temporary water reservoir such as a stream overflow. Anopheles melanoon larvae were identified by the BOLD fragment of the mitochondrial COI gene. Anopheles melanoon larvae developed together with An. maculipennis s. s. in permanent water reservoirs with abundant aquatic vegetation in the floodplain of the Psou River. Many natural breeding sites in the Psou River estuary are now lost due to large-scale construction. Anopheles maculipennis s. s. and An. plumbeus mosquitoes numerically outnumber An. melanoon in the Imereti Valley and neighboring in Abkhazia (locations 47-49, 52-56).
Anopheles hyrcanus breeding sites were found in lakes along the eastern zone of the southern Crimean coast. Larvae developed in dense sedge and reed thickets, perennial grasses of the Cyperaceae and Poaceae families, where other malaria mosquito species were absent (Figure 1; locations 28-30). An. hyrcanus mosquitoes were found together with An. maculipennis s. s. and An. daciae larvae in two biotopes in the south of the Azov-Kuban Plain (Figure 1; locations 36-37). Water pH ranged from 7,00 to 8,24 An. hyrcanus breeding places; daytime temperature varies between 22,4 and 33,3°C; TDS changed from 0,81 to 3.02 ppt; the dissolved oxygen content in the water reached 10.0 mg/L (Table 2).
Because Anopheles claviger is a highly specialized malaria mosquito species, its larvae were found in springs, ditches with running water or in water reservoirs fed by groundwater. Water temperatures ranged from 16.5-23.7°C at most of the breeding sites (Table 2). Higher water temperatures of up to 27.5°C were only recorded in one biotope, with flowing water and a low density of An. claviger larvae (Table 2; location 50). Water pH varied within a narrow range of 7.14-7.72, but was 9.26 in one biotope (Table 2; location 50). TDS in all locations varied between 0.24 and 2.08 ppt. The amount of dissolved oxygen in water was 2.5 and 7.6 mg/L (Table 2; locations 45 and 50).
Anopheles plumbeus mosquitoes were found in the mountainous forested part of the southern Crimean coast. Larvae developed in shallow forest lakes, overflow of small rivers, in temporary micro-habitats such as tree holes, condensation puddles between piles of stones (Table 1; locations 7, 9, 11-13, 15, 16, 18, 19). Breeding sites of this species were found in tree holes on the Black Sea coast of the Caucasus (locations 48, 53) and in mountains at altitudes up to 1700 m (Table 1; locations 54, 55). Breeding sites of An. plumbeus mosquitoes were found in old car tires in Caucasian resorts (Table 1; locations 44, 53, 56). Water composition in larval habitats was highly variable (Table 2). Water acidity correlated with organic matter content, and pH varied from 5.20 to 8.32 (in open biotopes mainly in the range from 7.14 to 7.62). TDS varied between 0.03 and 2.78 ppt. Daytime water temperatures at breeding sites ranged from 14.9 to 32.3°C. Dissolved oxygen content was measured in a number of micro-watersheds and ranged from 2.5 to 7.5 mg/L. Larval densities did not exceed 9 per square meter at most breeding sites, but in habitats with high saprobic conditions larval densities reached 32-90 mosquitoes of all instars per square meter. Anopheles plumbeus larvae developed together with An. maculipennis s. s. in the two largest of the listed breading sites - in a forest lake and in an ornamental pond of the Nikitsky Botanical Gardens (Table 1; locations 7, 18).
3.2. Chromosomal Inversion Polymorphism
The mitotic chromosome set in malaria mosquitoes includes three pairs of chromosomes [59]. Salivary gland cells of the larvae contain polytene chromosomes. Homologues chromosomes are paired during polytenization. The karyotype of polytene chromosomes consists of 5 arms (short arm of sex chromosome XL and arms of autosomes 2R, 2L, 3R, 3L). All 5 arms are connected by a common chromocenter. The long arm of the XR sex chromosome and both arms of the Y chromosome consist of heterochromatin and are not polytenized. Male karyotypes have only one polytene X chromosome that is thinner than the X chromosome of females.
Chromosomal inversions were studied in populations of An. atroparvus and An. daciae. We identified chromosomal inversions in accordance with the previously published photo maps of polytene chromosomes in An. atroparvus and An. daciae/An. messeae [52,53].
Anopheles daciae differed from other species by a high degree of chromosomal polymorphism (Table 3).
Homo- and heterozygotes for three paracentric inversions were common in populations of this species: XL1 (2a-5b); 3R1 (23c/24a-26c/27a); 3L1 (34b/c-37a/b-38c/39a-39c/d), whereas the 3L1 inversion consists of two overlapping inversions. Only one heterozygote for new inversion 2R5 (11c-14a) was found in the Novorossiysk population (Figure 2; location 42). There were no inversions in the 2L arm.
We described, for the first time, the chromosomal polymorphism of marginal populations of An. daciae in the southern part of the species range. The only one population of An. daciae in the Crimean Peninsula (Figure 3; location 10) differed in chromosomal composition from three Caucasian populations (Figure 3; locations 36, 40, 42).
A high level of inversion polymorphism of sex chromosome XL is observed in all populations of An. daciae on the Black Sea coast (Figure 3), but the frequency of chromosomal variants with XL0 inversion was significantly higher in the Crimean population in males (χ2=4.47; number of degrees of freedom df=1; p<0.05) and females (χ2=6.16; df=2; p<0.05). XL0 inversion serves as a species marker of An. daciae. The alternative inversion XL1 is considered ancestral to the two cryptic species An. daciae and An. messeae [53]. According to molecular genetic analyses, the separation of An. daciae and An. messeae happened about 2 Ma during the glaciation in Eurasia [44]. It is assumed that An. daciae mosquitoes were isolated in a refugium in southern Europe. The XL0 inversion, which occurs with high frequency in populations of the steppe zone and broad-leaved forest zone, probably arose during this period [60].
Autosomal inversions 3R1 and 3L1 are present with low frequency in coastal populations of An. daciae (Figure 3). These inversions were probably derived from the common ancestor of An. daciae and An. messeae, since polymorphism on inversions 3R1 and 3L1 is present in populations of both species. Geographical gradients in longitude suggest the influence of climatic factors on the frequencies of these inversions [56]. The frequencies of homo- and heterozygotes with 3R1 and 3L1 inversions were significantly higher in Caucasian populations (χ2=4.48 and 7.40; df=1; p<0.05 and p<0.01, respectively). In general, the level of chromosomal polymorphism was significantly higher in the Caucasian populations than in the Crimean population.
The inversion polymorphism was observed in An. atroparvus populations. Heterozygotes for inversion 3L1 (34b/c-38b) were found in two populations from Crimea and the Stavropol Region (Figure 4).
Heterozygotes 3L1 occurred with a frequency of 33.3% in both populations (Table 1; locations 2, 19). No homozygotes for this inversion were detected. Chromosome 3L has an identical disc pattern in An. atroparvus and An. daciae. Interestingly, the breakpoints of the 3L1 inversion of An. atroparvus (34b/c and 38b) are close to two breakpoints of the overlapping 3L1 inversion of An. daciae (34b/c-38c/39a). Perhaps this localization of inversions determines similar adaptive effects, so individuals of two species with these inversions are found in the same regions.
Discussion
In this study, we examined the malaria mosquito species composition, geographical distribution, and ecological preferences in the Crimean Peninsula and the Black Sea coast of the Caucasus. Anopheles maculipennis s. s. was the most abundant species in the foothills and coastal areas. Anopheles daciae dominated in freshwaters in the Kuban-Priazov lowland. Anopheles atroparvus larvae developed in saline lakes, but they may also be subdominant in freshwater pools. Anopheles melanoon mosquitoes were found only in the humid subtropical zone of the Black Sea coast of the Caucasus. Anopheles claviger mosquitoes preferred cool springs and groundwater outflows. Anopheles hyrcanus larvae developed in reed thickets along the shores of shallow lakes. Anopheles plumbeus larvae inhabited tree holes and shallow temporary water pools. The diversity of coastal landscapes allowed the above species with different ecological preferences to co-occur in the same area. The number of malaria mosquitoes was limited by the lack of suitable water pools during dry and hot summers, especially in the steppe zone.
Anopheles maculipennis s. s. appeared to out-compete mosquitoes of other Maculipennis species in water pools with standing fresh water on the southern coast of the Crimea and in the foothills of the Caucasus. This species was found together with An. daciae and An. messeae s. s. over a wide area of the Russian Plain [44]. Anopheles maculipennis s. s. larvae also occupied temporary water pools dominated by other closely related malaria mosquito species. It has been shown that climate warming is contributing to the expansion of the species range of An. maculipennis s. s. in the north and east of European Russia [61,62]. Chromosomal polymorphism in An. maculipennis populations has been noticed but it was not evaluated in the current study. Populations of this species are considered as chromosomally monomorphic throughout the territory of the Russian Plain. Only one single heterozygous inversion on the left arm of chromosome 2L, at the 5c-20c region, was observed in the population of Falesti [52,63].
Before the description of An. daciae in 2004 [34], both An. daciae and An. messeae were diagnosed as An. messeae Fall., 1926 [11,45,46,47]. We were unable to find any An. messeae s. s. mosquitoes in the studied regions. This result was verified by sequencing ITS2 fragments of 166 larvae from three habitats (Table 1; locations 10, 40, 42) [56]. The ecological niches of An. daciae and An. maculipennis partially overlap in the Russian Plain; larvae of both species can develop in the same water reservoirs under similar temperate conditions. However, more suitable breeding sites for An. maculipennis were found in the subtropical zone, on the southern coast of the Crimea and on the southern part of the Black Sea coast of the Caucasus, where Anopheles daciae mosquitoes were not found. The southern range limit of An. daciae was in the Pshadsky district of Krasnodar Krai (Figure 1; location 43), where the transition from temperate to subtropical climate occurs. Winter temperatures are significantly higher in southern areas, in the Tuapse-Sochi coastal strip making over-wintering conditions as the main limiting factor for Palearctic species of malaria mosquitoes in the south [64]. It is likely that diapausing females of An. daciae cannot tolerate warm winters in the humid subtropical zone. In contrast, An. maculipennis s. s. females can blood feed repeatedly during the over-wintering period [7,64,65,66]. Feeding on blood during diapause helps species to survive in excessively warm winter shelters whereas fat reserves of diapausing females are rapidly depleted. So far, only An. maculipennis s. s. has been considered as the main potential vector of malaria throughout the southern Russian Plain, especially in the mountainous and foothill areas of the North Caucasus [7]. However, we believe that the epidemiologic role of An. daciae is currently underestimated and needs to be further evaluated.
Our study indicated that populations of An. daciae and An. maculipennis s. s. on the Crimean Peninsula were isolated from those of the Kuban-Priazov lowland and the Black Sea lowland. A natural barrier for these mosquitoes is the dry steppes with saline lakes in the northern Crimea and the Taman Peninsula. The isolation of populations likely occurred after the last glaciation in Europe because of the Black Sea natural disaster [67]. The Black Sea was a freshwater lake with the level of 120 m below the present-day level. The rupture of the Bosphorus, followed by the intrusion of saline water from the Mediterranean Sea, occurred around 9300 BC. Prior to this, a significant part of the present underwater continental shelf of the Black Sea was a terrestrial area with a common flora and fauna. Flooding led to the breakage of a continuous coastal strip and the formation of the Crimean Peninsula. The subsequent climatic aridification in the northen part of Crimea led to the isolation of the mosquito fauna of the southern Crimean coast. The use of inversions as genetic markers allows reconstructing the genetic history of An. daciae populations on the Black Sea coast. Apparently, long-term isolation caused differences in the chromosomal composition of An. daciae populations in Crimea and on the Black Sea coast of the Caucasus. The Crimean population had a low level of polymorphism in autosomal inversions: 0% homo- and heterozygotes for inversion 3R1 and only 1.5% heterozygotes for inversion 3L1 (Tab. 3; location 10). In Caucasian populations, the frequencies of homo- and heterozygotes for 3R1 and 3L1 inversions were much higher and vary between 7.4-10.0% and between 10.2-18.7%, respectively (Tab. 3; location 36, 40, 42). We determined that the Crimean and the Black Sea coast populations of this species were more homogeneous than in the center of the Russian Plain. The long-term isolation of the Crimean populations has led to even more pronounced differences in the chromosomal polymorphism of the An. daciae populations there. Local populations within Crimea differ mainly in the frequency of inversions in the sex chromosome XL. The autosomal inversions 3R1 and 3L1 occur with lower frequency than in the center of the Russian Plain [43]. The level of chromosomal polymorphism in An. daciae/An. messeae mosquitoes was shown to be associated with specific landscape-climatic zones [60]. We believe that the high level of inversion polymorphism is associated with optimal landscape-climatic zones for this species. The sub-taiga and forest-steppe zones are probably more favorable for the development of An. daciae mosquitoes than the steppes of the Kuban-Priazov lowland.
In contrast to An. daciae and An. maculipennis s. s., An. atroparvus mosquitoes develop en masse in the steppe salt lakes in the northern Crimea [68]. The steppes and semi-deserts of the northern Crimea belong to the same natural zones of the Black Sea lowlands. The steppes of the Kerch Peninsula in Crimea are separated from the steppes of the Taman Peninsula by the relatively narrow Kerch Strait. Thus, the range of An. atroparvus covered the coastal areas as a continuous strip. Similar chromosomal variability occurred in geographically distant populations of this species. For example, the chromosomal inversion 3L1 is present in the Crimea and on the Black Sea coast of the Caucasus in this mosquito species. The northern limit of the An. atroparvus range is the southern Russian Plain, approximately south of the 48th parallel [47] but is not well defined.
The malaria mosquito An. melanoon was one of the rarest species of the Maculipennis complex on the Black Sea coast of the Caucasus. This species lives in sympatry with An. maculipennis s. s. on the territory of Bulgaria, Moldova, Romania, Georgia and Turkey [34,69,70,71,72,73]. We assumed that the northern limit of the An. melanoon distribution was the subtropical zone of the Black Sea coast of the Caucasus and may have shifted southwards to the Imereti Valley under conditions of anthropogenic transformation of coastal landscapes. The species was chromosomally monomorphic [18]. The polytene chromosomes of An. melanoon and An. maculipennis s. s. in ovarian nurse cells have the same banding patterns except for pericentromeric regions [74] that attach to the nuclear periphery, which causes the differentiation of chromosomal morphology between these species [75,76,77].
The malaria mosquito An. hyrcanus inhabited steppe plains and was one of the most highly specialized species. The larvae of this exophilic mosquito developed mainly in the coastal zone of lakes and in wetlands covered with reeds, rushes, sedges, cattails, and other aquatic vegetation. In the Krasnodar Krai, rice fields created favorable breeding conditions for An. hyrcanus mosquitoes. Anopheles hyrcanus breeding sites have been found in the Tien Shan and Pamir-Alai valleys of Central Asia at altitudes of up to 1000 m [78]. Chromosomal polymorphism in An. hyrcanus populations has not been studied. A high level of chromosomal variability was found in populations of another species of the Oriental Hyrcanus group, An. kleini Rueda, 2005, which lives in the Far East [79,80].
Anopheles claviger mosquitoes were widespread in the foothills of the Caucasus and Crimea. We studied several An. claviger breeding sites on the southern coast of Crimea. The natural complexes of the southern coast of Crimea were formed under the conditions of a hot sub-Mediterranean climate [81]. The sub-Mediterranean climate zone consists of a western part (from Cape Aya to Alushta) and a more arid eastern part (from Alushta to Feodosia). Anopheles claviger mosquitoes inhabited both parts of the sub-Mediterranean climate zone, especially in mountain forests at the northern limit of this zone. Two population peaks of An. claviger mosquitoes have been recorded during the summer season: in mid-late June and late October [82]. The summer decline in abundance was specific for An. claviger throughout the southern Russian Plain. Anopheles claviger can be found at altitudes up to of 2200 m in the Tien Shan Mountains and numerically outnumbers other Anopheles species in the foothills throughout the breeding season [83]. A sharp decline in An. claviger mosquito numbers was shown to occur during the summer months in the lowlands of Central Asia, as reservoirs of cold clear water are required for larval development of this species. Chromosomal variability in the wide distribution of An. claviger has not been studied.
Anopheles plumbeus mosquitoes were usually found in the mountainous forests. The larval stages of this species developed in the rotten tree holes filled with rainwater with a high organic content, as well as in small water pools of natural origin [82]. Anopheles plumbeus females layed eggs in water pools just above the waterline and the larvae hatch during the first floods of the rainy season [84]. Anopheles plumbeus was originally considered a dendrolimnetic species, but the ecological preferences of this species have now changed [85,86]. In several European countries, this species has begun to breed in water pools of artificial origin, such as rainwater barrels, lagoons, septic tanks, car tires, cemetery vases, liquid manure collection tanks, and manure puddles [85,86,87,88,89]. According to our observations, An. plumbeus has started to breed in the territory of Caucasian resorts since 2018. The anthropogenic transformation of coastal landscapes has created favorable conditions for the development of this species in artificial water containers with hard walls polluted with organic matter. An increase in the number of An. plumbeus in Crimea and the Black Sea coast of the Caucasus began in 2018. The expansion of the An. plumbeus range from the vicinity of Sochi сity (Tab. 1; location 53) to Tuapse district (Tab. 1; location 44). occurred in several stages and was similar to the spread of the invasive Asian tiger mosquito Aedes albopictus Scuse, 1895 [90]. Unlike other Eurasian malaria mosquito species, An. plumbeus has been shown to be capable of transmitting malaria parasites Plasmodium falciparum and Plasmodium vivax [85,91,92]. This malaria mosquito was considered the probable cause of two autochthonous cases of Pl. falciparum malaria in Germany [66,93]. Moreover, An. plumbeus was shown to be susceptible to West Nile virus and, because of its double ornithophagic and anthropophagic behavior, was considered as one of the potential vectors of this virus from birds to humans [94,95]. Clearly, the epidemiological significance of this species as a secondary vector of vector-borne diseases in Crimea and the Black Sea coast of the Caucasus needs to be reconsidered. It remains unknown how changes in the ecological preferences of An. plumbeus are related to the genetic variability of natural populations. Chromosomal polymorphism in An. plumbeus populations has not been studied.
Although An. algeriensis, An. sacharovi, and An. superpictus were previously recorded in the Crimea and the Black Sea coast of the Caucasus [47], we have not found any breeding sites of these species. Previously, An. algeriensis was found in spring water in the foothills of the North Caucasus and in the Kuban-Priazov lowland [96]. Anopheles algeriensis larvae begin to emerge in spring waters in the Caucasus at a temperature of about 5°C [97]. The breeding sites of mosquitoes of this species have not been found less than 3-5 km from any settlement [98]. The most recent record of An. algeriensis mosquitoes was made in Kalmykia [99]. The northern limit of the range of An. algeriensis is south of the 48th parallel on the Russian Plain and is not well defined. Anopheles algeriensis has been shown to be a competent vector for Plasmodium parasites [100]. However, this species is rare, lives far from human settlements and may not play a role in the malaria transmission in the Caucasus. The karyotype composition and chromosomal polymorphism in An. algeriensis populations have not been studied.
The malaria mosquito An. sacharovi has not been recorded in Crimea and the Black Sea coast of the Caucasus, but it has been found the adjacent plains of Transcaucasia and Dagestan [101]. This species was the main vector of three-day malaria caused by Pl. vivax in the valleys of Transcaucasia (Georgia, Armenia, Azerbaijan) during the malaria outbreak in the late 20th and early 21st centuries [7]. The malaria mosquitoes An. sacharovi and An. superpictus are among the most epidemiologically important species in the Palearctic [7]. Global warming may contribute to the expansion of the range of these species to the southern Russian Plain [102,103,104]. For example, the northern limit of the range of An. sacharovi has shifted from Dagestan to the territory of Kalmykia, where this species was not previously recorded [101]. Environmental changes can affect not only the geographical distribution but also the abundance and ecological preferences of malaria mosquitoes. Finally, An. superpictus was considered a mountain stream species in Central Asia (Tajikistan). Currently, mosquitoes of this species are found in lowland waters with increased eutrophication, including rice fields. The abundance of An. superpictus increased significantly as a result of changes in ecological preferences [78].
Our findings demonstrate that malaria mosquitoes of the Crimean Peninsula and the Black Sea coast of the Caucasus consists of ecologically specialized species. This study helped to better understand the dynamics of the malaria mosquito species distribution and their ecological preferences under the effects of global warming and increasing human activities. Further monitoring of the composition of malaria vector species and their geographic distribution represents an important component of entomological surveillance, which stimulates the development of appropriate mosquito control strategies aimed at preventing the re-emergence and spread of malaria in areas where it was previously eliminated.
Author Contributions
Conceptualization, A.V.M., A.G.B. and M.I.G.; Methodology A.V.M., A.G.B., I.V.S., M.V.S., B.V.A. and M.I.G.; Supervision, A.V.M. and M.I.G.; Resources, A.V.M., A.G.B., V.N.R., I.V.S., M.V.S. and M.I.G.; Investigation, A.V.M., I.I.B., A.N.N., D.A.K., A.G.B., V.N.R., B.V.A., I.I.G., E.Y.L., V.I.P., I.V.S., M.V.S. and M.I.G.; Writing—Original Draft Preparation, A.V.M., A.G.B. and M.I.G.; Writing—Review and Editing, A.V.M., A.G.B., I.V.S., M.V.S. and M.I.G.; Funding Acquisition, A.V.M. All authors have read and agreed to the published version of the manuscript.
Funding
The study was funded by the Russian Science Foundation (RSF) grant No. 24-44-10003, https://rscf.ru/project/24-44-10003/ “Genetic and ecological analysis of populations of the malaria mosquito Anopheles plumbeus as an important potential vector of vector-borne diseases in the Russian Federation and the Republic of Belarus” (the project is implemented by a scientific team together with a foreign scientific team selected by the Belarusian Republican Foundation for Basic Research (BRFFR), grant No. B23RNFM-068).
Data Availability Statement
All the data are available in the text, figures and tables of this article. The Sanger sequence data from ITS2 for individual mosquitoes from Mazanka, Crimea (Table 1, location 5), are available in GeneBank [101] under accession numbers PQ510968–PQ511024; from Tylovoye, Crimea (Table 1, location 10B and 10D) – numbers PQ554994–PQ555014 and PQ550752–PQ550804; from Abinsk, Krasnodar Krai (Table 1, location 40) – numbers PQ526514–PQ526598. COI sequence data for individual mosquitoes from Verkhneveseloye Krasnodar Krai (Table 1, location 50) are available in GeneBank under access numbers ID PQ740514–PQ740518; from Mazanka, Crimea (Table 1, location 51) – number PQ740513.
Acknowledgements
We would like to express our gratitude to the staff of the Anti-Plague Station of the Republic of Crimea for their assistance in collecting mosquitoes.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
References
- Mironova, L.P. Socio-ecological problems of Eastern Crimea in the past and present: causes of emergence, ways of solution. History and Modernity 2017, 1, 79–106. [Google Scholar]
- Kudaktin, A.N. Ecological threats to the resorts of the south of Russia. Fundamental researches 2006, 10, 56–58. Available online: https://fundamental-research.ru/ru/article/view?id=5473 (accessed on 20 September 2024).
- Rossati, A.; Bargiacchi, O.; Kroumova, V.; Zaramella, M.; Caputo, A.; et al. Climate, environment and transmission of malaria. Infez Med 2016, 24, 93–104. [Google Scholar] [PubMed]
- Fischer, L.; Gultekin, N.; Kaelin, M. B.; Fehr, J.; Schlagenhauf, P. Rising temperature and its impact on receptivity to malaria transmission in Europe: A systematic review. Travel Med Infect Dis 2020, 36, 101815. [Google Scholar] [CrossRef] [PubMed]
- Regional strategy: from malaria control to elimination in the WHO European Region 2006–2015. World Health Organization. Regional Office for Europe: Copenhagen, Denmark. 2006, p. 40. Available online: https://iris.who.int/bitstream/handle/10665/107760/E88840.pdf?sequence=1&isAllowed=y (accessed on 20 September 2024).
- Ejov, M.; Sergiev, V.; Baranova, A.; Kurdova-Mintcheva, R; Emiroglu, N.; et al. Malaria in the WHO European Region: on the road to elimination 2000-2015: summary; World Health Organization. Regional Office for Europe: Copenhagen, Denmark, 2018; p. 40. Available online: https://iris.who.int/bitstream/handle/10665/342148/9789289053112-eng.pdf?sequence=1&isAllowed=y (accessed on 20 September 2024).
- Lysenko, A. J.; Kondrashin, A.V. Malariology. World Health Organization: Geneva, Switzerland, 1999. Available online: https://fundamental-research.ru/ru/article/view?id=5473 (accessed on 20 September 2024).
- Morenets, T.M.; Isaeva, E.B.; Gorodin, V.N.; Avdeeva, M.G.; Grechanaya, T.V. Clinical and epidemiologic aspects of malaria in Krasnodar Krai. Epidemiology and Infectious Diseases 2016, 21, 253–261. [Google Scholar] [CrossRef]
- Karimov, I.Z.; Los’-Yatsenko, N.G.; Midikari, A.S.; Gorovenko, M.V.; Arshinov, P.S. Clinical and epidemiological features of imported malaria in the Republic of Crimea for a twenty-year period (1994-2014). Kazan medical journal 2014, 95, 916–920. [Google Scholar] [CrossRef]
- Baranova, A.M.; Sergiev, V.P.; Guzeeva, T.M.; Tokmalaev, A.K. Clinical suspicion to imported malaria: transfusion cases and deaths in Russia. Infectious Diseases: News, Opinions, Training 2018, 7, 97–101. [Google Scholar] [CrossRef]
- Gornostaeva, R.M. A checklist of the mosquitoes (fam. Culicidae) in the European part of Russia. Parazitologiia 2000, 34, 428–434. [Google Scholar]
- Gutsevich, A.V.; Dubitskiy, A.M. New species of mosquitoes in the fauna of the USSR. Parazitologicheskiy sbornic 1981, 30, 97–165. [Google Scholar]
- Kitzmiller, J.B.; Frizzi, G.; Baker, R. Wright, J.W., Ed.; Evolution and speciation within the Maculipennis complex of the genus Anopheles. In Genetics of Insect Vectors of Disease; Elsevier Publishing Company: Amsterdam, The Netherlands; London, UK; New York, NY, USA, 1967; pp. 151–210. [Google Scholar]
- Coluzzi, M. Sibling species in Anopheles and their importance in malariology. Miscellaneous Publ. Entomol. Soc. Amer. 1970, 7, 63–77. [Google Scholar] [CrossRef]
- Knight, K.L.; Stone, A. A catalog of the mosquitoes of the world (Diptera, Culicidae). 2nd edition. Thomas Say Found. Entomol. Soc. Amer. 1977. [Google Scholar]
- White, G.B. The place of morphological studies in the investigation of Anopheles species complexes. Mosquito Systematics 1977, 9, 1–24. [Google Scholar]
- White, G.B. Systematic reappraisal of the Anopheles maculipennis complex. Mosquito Systematics 1978, 10, 13–44. [Google Scholar]
- Stegniy, V.N. Population Genetics and Evolution of Malaria Mosquitoes; Tomsk State University Publisher: Tomsk, Russia, 1991; pp. 1–137. ISBN 5-7511-0073-5. [Google Scholar]
- Frizzi, G. Salivary gland chromosomes of Anopheles. Nature (London) 1947, 160, 226–227. [Google Scholar] [CrossRef] [PubMed]
- Frizzi, G. Nuovi contributi e prospetti di ricerca nel gruppo Anopheles maculipennis in base allo studio del dimorfismo cromosomico. Symposia Genetica 1952, 3, 231–265. [Google Scholar]
- Frizzi, G. Etude cytogénétique d'Anopheles maculipennis en Italie. Bull World Health Organ. 1953, 9, 335–344. [Google Scholar]
- Kiknadze, I.I. Chromosomes of Diptera. Evolutionary and practical significance. Genetika 1967, 7, 145–165. [Google Scholar]
- Kabanova, V.M.; Kartashova, N.N.; Stegnii, V.N. Karyological study of natural populations of malarial mosquitoes in the Middle Ob river. I. Characteristics of the karyotype of Anopheles maculipennis messeae. Tsitologiia 1972, 14, 630–636. [Google Scholar]
- Stegnii, V.N.; Kabanova, V.M. Cytoecological study of natural populations of malaria mosquitoes on the USSR territory. 1. Isolation of a new species of Anopheles in Maculipennis complex by the cytodiagnostic method. Med Parazitol (Mosk) 1976, 45, 192–198. [Google Scholar]
- Stegniy, V.N. Detection of chromosomal races in the malaria mosquito Anopheles sacharovi. Tsitologiia 1976, 18, 1039–1041. [Google Scholar]
- Stegniy, V.N. Reproductive interrelations of malaria mosquitos of the complex Anopheles maculipennis (Diptera, Culicidae). Zool Zh. 1980, 59, 1469–1475. [Google Scholar]
- Baker, R.H.; French, W.L.; Kitzmiller, J.B. Induced copulation in Anopheles mosquitoes. Mosq News 1962, 22, 16–17. [Google Scholar]
- Collins, F.H.; Paskewitz, S.M. A review of the use of ribosomal DNA (rDNA) to differentiate among cryptic Anopheles species. Insect Mol. Biol. 1996, 5, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Harbach, R.E. The classification of genus Anopheles (Diptera: Culicidae): a working hypothesis of phylogenetic relationships. Bull Entomol Res. 2004, 94, 537–553. [Google Scholar] [CrossRef] [PubMed]
- Harbach, R.E. Review of the internal classification of the genus Anopheles (Diptera: Culicidae): the foundation for comparative systematics and phylogenetic research. Bulletin of Entomological Research 1994, 84, 331–342. [Google Scholar] [CrossRef]
- Fedoroff, N.V. On spacers. Cell 1979, 16, 697–710. [Google Scholar] [CrossRef]
- Sedaghat, M.M.; Linton, Y.-M.; Oshaghu, M.A.; Vatandoost, H.; Harbach, R.E. The Anopheles maculipennis complex in Iran: Molecular characterization and recognition of a new species. Bull. Entomol. Res. 2003, 93, 527–535. [Google Scholar] [CrossRef]
- Sedaghat, M.M.; Howard, T.; Harbach, R.E. Morphological study and description of Anopheles (Anopheles) persiensis, a member of the Maculipennis Group (Diptera: Culicidae: Anophelinae) in Iran. Journal of Entomological Society of Iran 2009, 28, 35–25. [Google Scholar]
- Nicolescu, G.; Linton, Y.-M.; Vladimirescu, A.; Howard, T.M.; Harbach, R. E. Mosquitoes of the Anopheles maculipennis group (Diptera: Culicidae) in Romania, with the discovery and formal recognition of a new species based on molecular and morphological evidence. Bull. Entomol. Res. 2004, 94, 525–535. [Google Scholar] [CrossRef]
- Gordeev, M.I.; Zvantsov, A.B.; Goriacheva, I.I.; Shaĭkevich, E.V.; Ezhov, M.N. Description of the new species Anopheles artemievi sp.n. (Diptera, Culicidae). Med Parazitol (Mosk) 2005, 2, 4–5. [Google Scholar]
- Linton, Y.-M.; Smith, L.; Harbach R., E. Observations on the taxonomic status of Anopheles subalpinus Hackett & Lewis and An. melanoon Hackett. Eur. Mosq. Bull. 2002, 13, 1–7. [Google Scholar]
- Collins, F.H.; Paskewitz, S.M. A review of the use of ribosomal DNA (rDNA) to differentiate among cryptic Anopheles species. Insect Mol. Biol. 1996, 5, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Marinucci, M.; Romi, R.; Mancini, P.; Di Luca, M.; Severini, C. Phylogenetic relationships of seven palearctic members of the maculipennis complex inferred from ITS2 sequence analysis. Insect Mol Biol. 1999, 8, 469–480. [Google Scholar] [CrossRef] [PubMed]
- Proft, J.; Maier, W.A.; Kampen, H. Identification of six sibling species of the Anopheles maculipennis complex (Diptera: Culicidae) by a polymerase chain reaction assay. Parasitol. Res. 1999, 85, 837–843. [Google Scholar] [CrossRef] [PubMed]
- Harbach, R.E. The classification of genus Anopheles (Diptera: Culicidae): a working hypothesis of phylogenetic relationships. Bull Entomol Res. 2004, 94, 537–553. [Google Scholar] [CrossRef]
- Harbach, R. E. Manguin, S., Ed.; The phylogeny and classification of Anopheles. In Anopheles mosquitoes - New insights into malaria vectors; Chapter 1; InTech: Rijeka, Croatia, 2013; pp. 3–55. [Google Scholar] [CrossRef]
- Hodge, J.M.; Yurchenko, A.A.; Karagodin, D.A.; Masri, R.A.; Smith, R.C.; Gordeev, M.I.; Sharakhova, M.V. The new Internal Transcribed Spacer 2 diagnostic tool clarifies the taxonomic position and geographic distribution of the North American malaria vector Anopheles punctipennis. Malar J. 2021, 20, 141. [Google Scholar] [CrossRef]
- Naumenko, A.N.; Karagodin, D.A.; Yurchenko, A.A.; Moskaev, A.V.; Martin, O.I.; Baricheva, E.M.; Sharakhov, I.V.; Gordeev, M.I.; Sharakhova, M.V. Chromosome and Genome Divergence between the Cryptic Eurasian Malaria Vector-Species Anopheles messeae and Anopheles daciae. Genes 2020, 11, 165. [Google Scholar] [CrossRef]
- Yurchenko, A.A.; Naumenko, A.N.; Artemov, G.N.; Karagodin, D.A.; Hodge, Ja.M.; Velichevskaya, A.I.; Kokhanenko, A.A.; Bondarenko, S.M.; Abai, M.R.; Kamali, M.; Gordeev, M.I.; Moskaev, A.V.; Caputo, B.; Aghayan, S.A.; Baricheva, E.M.; Stegniy, V.N.; Sharakhova, M.V.; Sharakhov, I.V. Phylogenomics revealed migration routes and adaptive radiation timing of Holarctic malaria mosquito species of the Maculipennis Group. BMC Biol. 2023, 21, 63. [Google Scholar] [CrossRef]
- Khalin, A.V.; Gornostaeva, R.M. On the taxonomic composition of mosquitoes (Diptera: Culicidae) of the world and Russian fauna (critical review). Parazitologiia 2008, 42, 360–381. [Google Scholar]
- Gornostaeva, R.M. Analysis of modern data on the fauna and ranges of malaria mosquitoes (Diptera: Culicidae: Anopheles) on the territory of Russia. Parazitologiia 2003, 37, 298–305. [Google Scholar]
- Gornostaeva, R.M.; Danilov, A.V. On ranges of the malaria mosquitoes (Diptera: Culicidae: Anopheles) of the Maculipennis complex on the territory of Russia. Parazitologiia 2002, 36, 33–47. [Google Scholar]
- Benedict, M.; Dotson, E. Methods in Anopheles research. 2015. Atlanta: Malaria Research and Reference Reagent Resource Center.
- Gutsevich, A.V.; Monchadskii, A.S.; Shtakelberg, A.A. Fauna of the USSR. Diptera. Mosquitoes; family Culicidae. Zoological Institute, USSR Academy of Science: Leningrad, Russia, 1971; p. 407. [Google Scholar] [CrossRef]
- Fedorova, M.V.; Sycheva, K.A. Blood-sucking mosquitoes (Diptera:Culicidae) of the Krasnodar Territory and the Crimean Peninsula: an identifier Akimkin, V.G., Eds.; FBIS Central Research Institute of Epidemiology: Moscow, Russia, 2024. [Google Scholar]
- Moskaev, A.V.; Gordeev, M.I.; Kuzmin, O.V. Chromosomal composition of populations of malaria mosquito Anopheles messeae in the centre and on the periphery of the species range. Bulletin of Moscow State Regional University. Natural Sciences 2015, 1, 29–36. [Google Scholar]
- Stegniy, V. N.; Kabanova, V. M. Chromosomal analysis of malaria mosquitoes Anopheles atroparvus and A. maculipennis (Diptera, Culicidae). Zool. zh. 1978, 57, 613–619. [Google Scholar]
- Artemov, G.N.; Fedorova, V.S.; Karagodin, D.A.; Brusentsov, I.I.; Baricheva, E.M.; Sharakhov, I.V.; Gordeev, M.I.; Sharakhova, M.V. New cytogenetic photomap and molecular diagnostics for the cryptic species of the malaria mosquitoes Anopheles messeae and Anopheles daciae from Eurasia. Insects 2021, 12, 835. [Google Scholar] [CrossRef] [PubMed]
- Corder, G.W.; Foreman, D.I. Nonparametric statistics: a step-by-step approach, 2nd ed; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2014; p. 288. [Google Scholar] [CrossRef]
- Sambrook, J.; Russell, D.W. Purification of nucleic acids by extraction with phenol:chloroform. CSH protocols 2006, 1. [Google Scholar] [CrossRef]
- Brusentsov, I.I.; Gordeev, M.I.; Yurchenko, A.A.; Karagodin, D.A.; Moskaev, A.V.; Hodge, J.M.; Burlak, V.A.; Artemov, G.N.; Sibataev, A.K.; Becker, N.; Sharakhov, I.V.; Baricheva, E.M.; Sharakhova, M.V. Patterns of genetic differentiation imply distinct phylogeographic history of the mosquito species Anopheles messeae and Anopheles daciae in Eurasia. Mol Ecol. 2023, 32, 5609–5625. [Google Scholar] [CrossRef]
- Rozen, S.; Skaletsky, H.J. Misener, S., Krawetz, S., Eds.; Primer 3 on the WWW for general users and biologist programmers. In In Methods in molecular biology: Bioinformatics methods and protocols; Humana Press: Totowa, NJ, USA, 2000; Volume 132, pp. 365–386. [Google Scholar] [CrossRef]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3 — new capabilities and interfaces. Nucleic Acids Res. 2012, 40, 115. [Google Scholar] [CrossRef]
- Rai, K.S. A Comparative Study of Mosquito Karyotypes. Ann. Ent. Soc. Amer. 1963, 56, 160–170. [Google Scholar] [CrossRef]
- Gordeev, M.I.; Temnikov, A.A.; Panov, V.I.; Klimov, K.S.; Lee, E.Yu.; Moskaev, A.V. Chromosomal variability in populations of malaria mosquitoes in different landscape zones of Eastern Europe and the Southern Urals. Bulletin of the Moscow State Regional University (Geographical Environment and Living Systems 2022, 4, 48–66. [Google Scholar] [CrossRef]
- Novikov, Y.M.; Vaulin, O.V. Expansion of Anopheles maculipennis s.s. (Diptera: Culicidae) to northeastern Europe and northwestern Asia: causes and consequences. Parasit Vectors. [CrossRef]
- Moskaev, A.V.; Bega, A.G.; Panov, V.I.; Perevozkin, V.P.; Gordeev, M.I. (2024). Chromosomal polymorphism of malaria mosquitoes of Karelia and expansion of northern boundaries of species ranges. Russian Journal of Genetics 2024, 60, 754–762. [Google Scholar] [CrossRef]
- Gordeev, M.I.; Moskaev, A.V.; Bezzhonova, O.V. Chromosomal polymorphism in the populations of malaria vector mosquito Anopheles messeae at the south of Russian plain. Russian Journal of Genetics 2012, 48, 962–965. [Google Scholar] [CrossRef]
- Beklemishev, V.N. Ecology of the malaria mosquito; Medgiz: Moscow, Russia, 1944; pp. 1–299. [Google Scholar]
- Jetten, T.H.; Takken, W. Anophelism without malaria in Europe: a review of the ecology and distribution of the genus Anopheles in Europe; Agric Univ Pap.: Wageningen, Netherlands, 1994; Volume 94, pp. 1–69. [Google Scholar]
- Bertola, M.; Mazzucato, M.; Pombi, M.; Montarsi, F. Updated occurrence and bionomics of potential malaria vectors in Europe: a systematic review (2000–2021). Parasit Vectors 2022, 15, 88. [Google Scholar] [CrossRef] [PubMed]
- Yanchilina, A.G.; Ryan, W.B.F.; McManus, J.F.; Dimitrov, P.; Dimitrov, D.; Slavova, K.; Filipova-Marinova, M. Compilation of geophysical, geochronological, and geochemical evidence indicates a rapid Mediterranean-derived submergence of the Black Sea's shelf and subsequent substantial salinification in the early Holocene. Marine Geology 2017, 383, 14–34. [Google Scholar] [CrossRef]
- Alekseev, E.V.; Razumeiko, V.N. Blood-sucking mosquitoes (Diptera, Culicidae) of anthropogenic landscapes of flat Crimea. Ecosystems, their optimisation and protection 2005, 16, 120–129. [Google Scholar]
- Hubenov, Z. Species composition and distribution of the dipterans (Insecta: Diptera) in Bulgaria. Advanced Books. National Museum of Natural History: Sofia, Bulgaria, 2021; Sep 9. [CrossRef]
- Șuleșco, T.; Sauer, F.G.; Lühken, R. Update on the distribution of Anopheles maculipennis s. l. members in the Republic of Moldova with the first record of An. daciae. 2024; Aug 17. [CrossRef]
- Bezzhonova, O.V.; Babuadze, G.A.; Gordeev, M.I.; Goriacheva, I.I.; Zvantsov, A.B.; Ezhov, M.N.; Imnadze, P.; Iosava, M.; Kurtsikashvili, G. Malaria mosquitoes of the Anopheles maculipennis (Diptera, Culicidae) complex in Georgia. Med Parazitol (Mosk) 2008, 3, 32–36. [Google Scholar]
- Akiner, M.M.; Cağlar, S.S. Identification of Anopheles maculipennis group species using polymerase chain reaction (PCR) in the regions of Birecik, Beyşehir and Cankiri. Turkiye Parazitol Derg. 2010, 34, 50–54. [Google Scholar]
- Simsek, F.M.; Ulger, C.; Akiner, M.M.; Tuncay, S.S.; Kiremit, F.; Bardakci, F. Molecular identification and distribution of Anopheles maculipennis complex in the Mediterranean region of Turkey. Biochemical Systematics and Ecology 2011, 39, 258–265. [Google Scholar] [CrossRef]
- Sharakhova, M.V.; Stegnii, V.N.; Braginets, O.P. Interspecies differences in the ovarian trophocyte precentromere heterochromatin structure and evolution of the malaria mosquito complex Anopheles maculipennis. Genetika 1997, 33, 1640–1648. [Google Scholar] [PubMed]
- Stegnii, V.N. , Structure reorganization of the interphase nuclei during ontogenesis and phylogenesis of malaria mosquitoes. Dokl. Akad. Nauk SSSR 1979, 249, 1231–1234. [Google Scholar]
- Stegnii, V.N. Systemic reorganization of the architectonics of polytene chromosomes in the onto- and phylogenesis of malaria mosquitoes. Genetika 1987, 23, 821–827. [Google Scholar]
- Stegnii, V.N.; Sharakhova, M.V. Systemic reorganization of the architechtonics of polytene chromosomes in onto- and phylogenesis of malaria mosquitoes. Structural features regional of chromosomal adhesion to the nuclear membrane. Genetika 1991, 27, 828–835. [Google Scholar] [PubMed]
- Kadamov, D.S.; Zvantseva, A.B.; Karimov, S.S.; Gordeev, M.I.; Goriacheva, I.I.; Ezhov, M.N.; Tadzhiboev, A. Malaria mosquitoes (Diptera, Culicidae, Anopheles) of North Tajikistan, their ecology, and role in the transmission of malaria pathogens. Med Parazitol (Mosk) 2012, 3, 30–34. [Google Scholar]
- Perevozkin, V.P. Chromosomal polymorphism of malarial mosquitoes (Diptera, Culicidae) of Primorsky Krai. Tomsk State Pedagogical University Bulletin 2009, 11, 181–185. [Google Scholar]
- Khrabrova, N.V.; Andreeva, Y.V.; Sibataev, A.K.; Alekseeva, S.S.; Esenbekova, P.A. Mosquitoes of Anopheles hyrcanus (Diptera, Culicidae) group: species diagnostic and phylogenetic relationships. Am J Trop Med Hyg. 2015, 93, 619–622. [Google Scholar] [CrossRef]
- Agarkova-Lyakh, I.V. Natural complexes of the coastal zone of the Southern coast of Crimea. Scientific Notes of the V. I. Vernadsky Crimean Federal University. Geography. Geology 2015, 1, 42–58. [Google Scholar]
- Razumeiko, V.N.; Ivashov, A.V.; Oberemok, V.V. Seasonal activity and density dynamics of blood-sucking mosquitoes (Diptera, Culicidae) in water bodies of the southern coast of Crimea. Scientific Notes of the V. I. Vernadsky Tauride National University 2010, 23, 114–128. [Google Scholar]
- Gordeev, M.I.; Zvantsov, A.B.; Goriacheva, I.I.; Shaĭkevich, E.V.; Ezhov, M.N.; Usenbaev, N.T.; Shapieva, Zh.Zh.; Zhakhongirov, Sh.M. Anopheles mosquitoes (Diptera, Culicidae) of the Tien Shan: morphological, cytogenetic, and molecular genetic analysis. Med Parazitol (Mosk) 2008, 3, 25–32. [Google Scholar]
- Service, M.W. 1968. Observations on feeding and oviposition in some British mosquitoes. Entomologia Experimentalis et Applicata 1968, 11, 277–285. [Google Scholar] [CrossRef]
- Schaffner, F.; Thiéry, I.; Kaufmann, C.; Zettor, A.; Lengeler, C.; Mathis, A.; Bourgouin, C. Anopheles plumbeus (Diptera: Culicidae) in Europe: a mere nuisance mosquito or potential malaria vector? Malar J. 2012, 11, 393. [Google Scholar] [CrossRef]
- Heym, E.C.; Kampen, H.; Fahle, M.; Hohenbrink, T.L.; Schäfer, M.; Scheuch, D.E.; Walther, D. Anopheles plumbeus (Diptera: Culicidae) in Germany: updated geographic distribution and public health impact of a nuisance and vector mosquito. Trop Med Int Health 2017, 22, 103–112. [Google Scholar] [CrossRef]
- Becker, N.; Petric, D.; Zgomba, M.; Boase, C.; Madon, M.; Dahl, C.; Kaiser, A. Mosquitoes and Their Control; Springer: Berlin, Heidelberg, Germany, 2010. [Google Scholar] [CrossRef]
- Bueno-Marí, R.; Jiménez-Peydró, R. Anopheles plumbeus Stephens, 1828: a neglected malaria vector in Europe. Malaria Reports 2011, 1, 2. [Google Scholar] [CrossRef]
- Dekoninck, W.; Hendrickx, F.; Vasn Bortel, W.; Versteirt, V.; Coosemans, M.; Damiens, D.; Hance, T.; De Clercq, E.M.; Hendrickx, G.; Schaffner, F.; Grootaert, P. Human-induced expanded distribution of Anopheles plumbeus, experimental vector of West Nile virus and a potential vector of human malaria in Belgium. J Med Entomol. 2011, 48, 924–928. [Google Scholar] [CrossRef] [PubMed]
- Bega, A.G.; Moskaev, A.V.; Gordeev, M.I. Ecology and distribution of the invasive mosquito species Aedes albopictus (Skuse, 1895) in the south of the European Part of Russia. Russian Journal of Biological Invasions 2021, 12, 148–56. [Google Scholar] [CrossRef]
- Marchant, P.; Eling, W.; van Gemert, G.J.; Leake, C.J.; Curtis, C.F. Could british mosquitoes transmit falciparum malaria? Parasitol Today 1998, 14, 344–345. [Google Scholar] [CrossRef]
- Bueno-Marí, R.; Jiménez-Peydró, R. Study of the malariogenic potential of Eastern Spain. Trop Biomed. 2012, 29, 39–50. [Google Scholar]
- Krüger, A.; Rech, A.; Su, X.Z.; Tannich, E. Two cases of autochthonous Plasmodium falciparum malaria in Germany with evidence for local transmission by indigenous Anopheles plumbeus. Trop Med Int Health 2001, 6, 983–985. [Google Scholar] [CrossRef]
- Medlock, J.M.; Snow, K.R.; Leach, S. Potential transmission of West Nile virus in the British Isles: an ecological review of candidate mosquito bridge vectors. Med Vet Entomol. 2005, 19, 2–21. [Google Scholar] [CrossRef]
- Medlock, J.M.; Snow, K.R.; Leach, S. Possible ecology and epidemiology of medically important mosquito-borne arboviruses in Great Britain. Epidemiol Infect. 2007, 135, 466–482. [Google Scholar] [CrossRef]
- Shcherbina V., P. Materials on the fauna of blood-sucking mosquitoes (Diptera, Culicidae) of the Lower Don and North Caucasus. Parasitol. sb. of ZIN AS USSR 1974, 26, 205–217. [Google Scholar]
- Enikolopov, S.K. Biology of Anopheles algeriensis Theo. Med Parazitol (Mosk) 1944, 13, 68–69. [Google Scholar]
- Enikolopov, S.K. On the ecology of Anopheles algeriensis Theo. 1903. Med Parazitol (Mosk) 1937, 6, 354–359. [Google Scholar]
- Savitskiĭ, B.P. Blood-sucking mosquitoes (Culicidae) attacking man in the region of the Eastern Manych (Kalmyk ASSR). Parazitologiia 1982, 16, 163–165. [Google Scholar] [PubMed]
- Tippelt, L.; Walther, D.; Scheuch, D.E.; Schäfer, M.; Kampen, H. Further reports of Anopheles algeriensis Theobald, 1903 (Diptera: Culicidae) in Germany, with evidence of local mass development. Parasitol. Res. 2018, 117, 2689–2696. [Google Scholar] [CrossRef] [PubMed]
- Perevozkin, V.P.; Bondarchuk, S.S.; Gordeev, M.I. The population-and-species-specific structure of malaria (Diptera, Culicidae) mosquitoes in the Caspian Lowland and Kuma-Manych Hollow. Med Parazitol (Mosk). 2012, 1, 12–17. [Google Scholar]
- Martens, W.J.M.; Kovats, R.S.; Nijhof, S.; deVries, P.; Livermore, M.J.T.; Mc Michael, A.J.; Bradley, D.; Cox, J. Climate change and future populations at risk of malaria. Global Environ Change 1999, 9, S89–S107. [Google Scholar] [CrossRef]
- Yasjukevich, V.V. Malaria in Russia and its immediate geographical environment: analysis of the situation in connection with the expected climate change. Problems of ecological monitoring and modelling of ecosystems 2002, 18, 142–157. [Google Scholar]
- Yasjukevich, V.V.; Titkina, S.N.; Popov, I.O.; Davidovich, E.A.; Yasjukevich, N.V. Climate-dependent diseases and arthropod vectors: possible impact of climate change observed in Russia. Problems of ecological monitoring and modelling of ecosystems 2013, 25, 314–359. [Google Scholar]
- NCBI. Available online: https://www.ncbi.nlm.nih.gov/ (accessed on 20 December 2024).
Figure 1.
Geographical distribution of malaria mosquitoes on the Crimean Peninsula and the Black Sea coast of the Caucasus. Numbers indicate locations as in Table 1.
Figure 1.
Geographical distribution of malaria mosquitoes on the Crimean Peninsula and the Black Sea coast of the Caucasus. Numbers indicate locations as in Table 1.
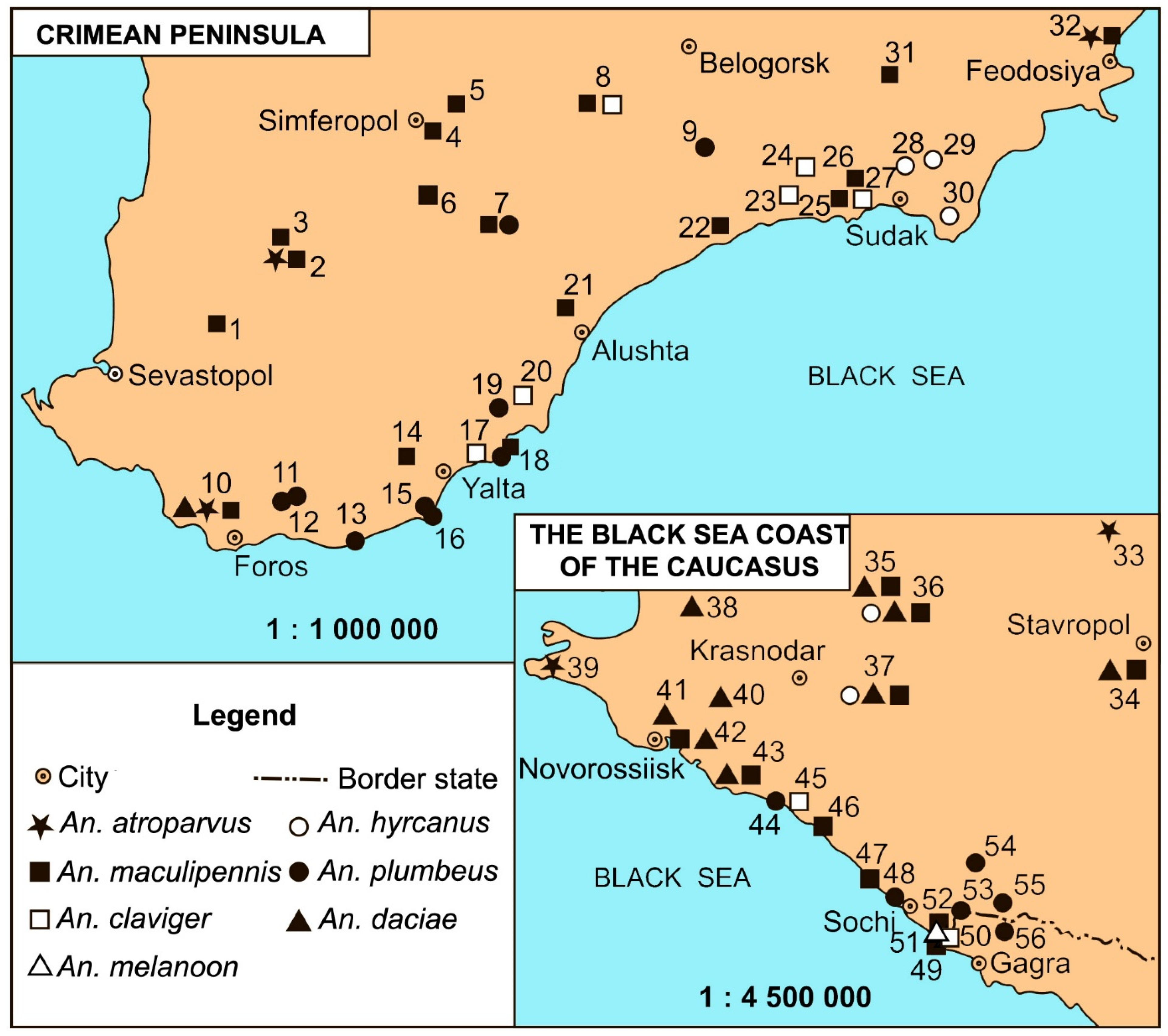
Figure 2.
A chromosomal complement of a squashed preparation of salivary gland cells in an An. daciae female, stained by lacto-aceto-orcein. Panel (a) shows the standard karyotype XL11, 2R00, 2L00, 3R00, and 3L00, where XL, 2R, 2L, 3R, and 3L represent chromosome arms and the numbers 11 and 00 are chromosomal variants (objective lens – Nikon Plan Fluor 60x/0,85). Chromosome arms XL, 2R, 2L, 3R, 3L are indicated. Panel (b) shows the inversion heterozygote 2R05 (11c-14a) in An. daciae (objective lens – Nikon Plan 100x/1,25). The arrows indicate the points of breaks and homologues exchanges in the inversion loop. CC – stands for the chromocenter. Scale bar equals 50 μm.
Figure 2.
A chromosomal complement of a squashed preparation of salivary gland cells in an An. daciae female, stained by lacto-aceto-orcein. Panel (a) shows the standard karyotype XL11, 2R00, 2L00, 3R00, and 3L00, where XL, 2R, 2L, 3R, and 3L represent chromosome arms and the numbers 11 and 00 are chromosomal variants (objective lens – Nikon Plan Fluor 60x/0,85). Chromosome arms XL, 2R, 2L, 3R, 3L are indicated. Panel (b) shows the inversion heterozygote 2R05 (11c-14a) in An. daciae (objective lens – Nikon Plan 100x/1,25). The arrows indicate the points of breaks and homologues exchanges in the inversion loop. CC – stands for the chromocenter. Scale bar equals 50 μm.
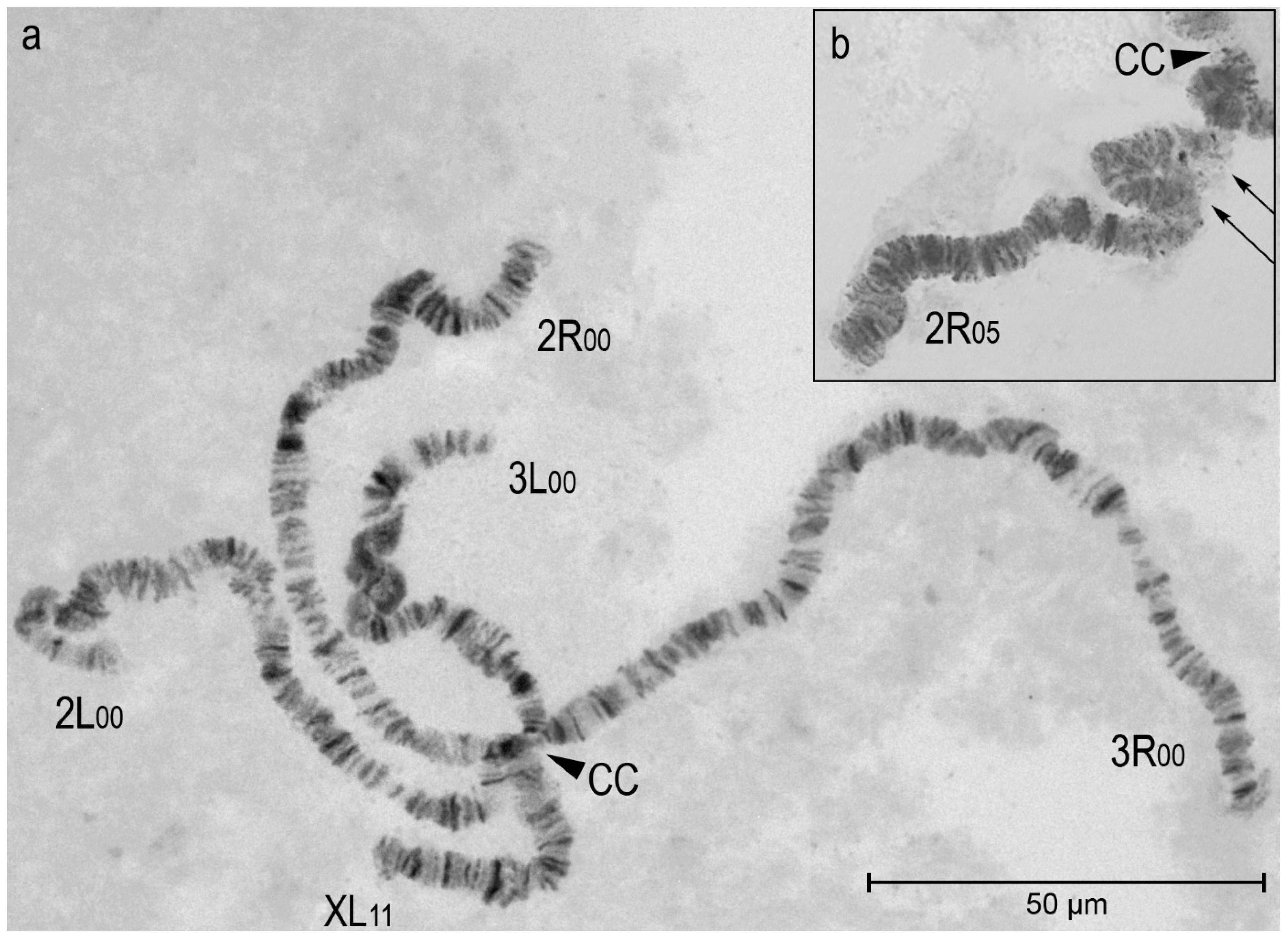
Figure 3.
Frequencies of inversion homo- and heterozygotes in populations of Anopheles daciae in Crimea (location 10) and on the Black Sea coast of the Caucasus (locations 36, 40, 42). The frequencies of inversions of sex chromosome XL in males and females are given separately. The frequencies of inversions of 3R and 3L autosomes are shown for individuals of both sexes.
Figure 3.
Frequencies of inversion homo- and heterozygotes in populations of Anopheles daciae in Crimea (location 10) and on the Black Sea coast of the Caucasus (locations 36, 40, 42). The frequencies of inversions of sex chromosome XL in males and females are given separately. The frequencies of inversions of 3R and 3L autosomes are shown for individuals of both sexes.
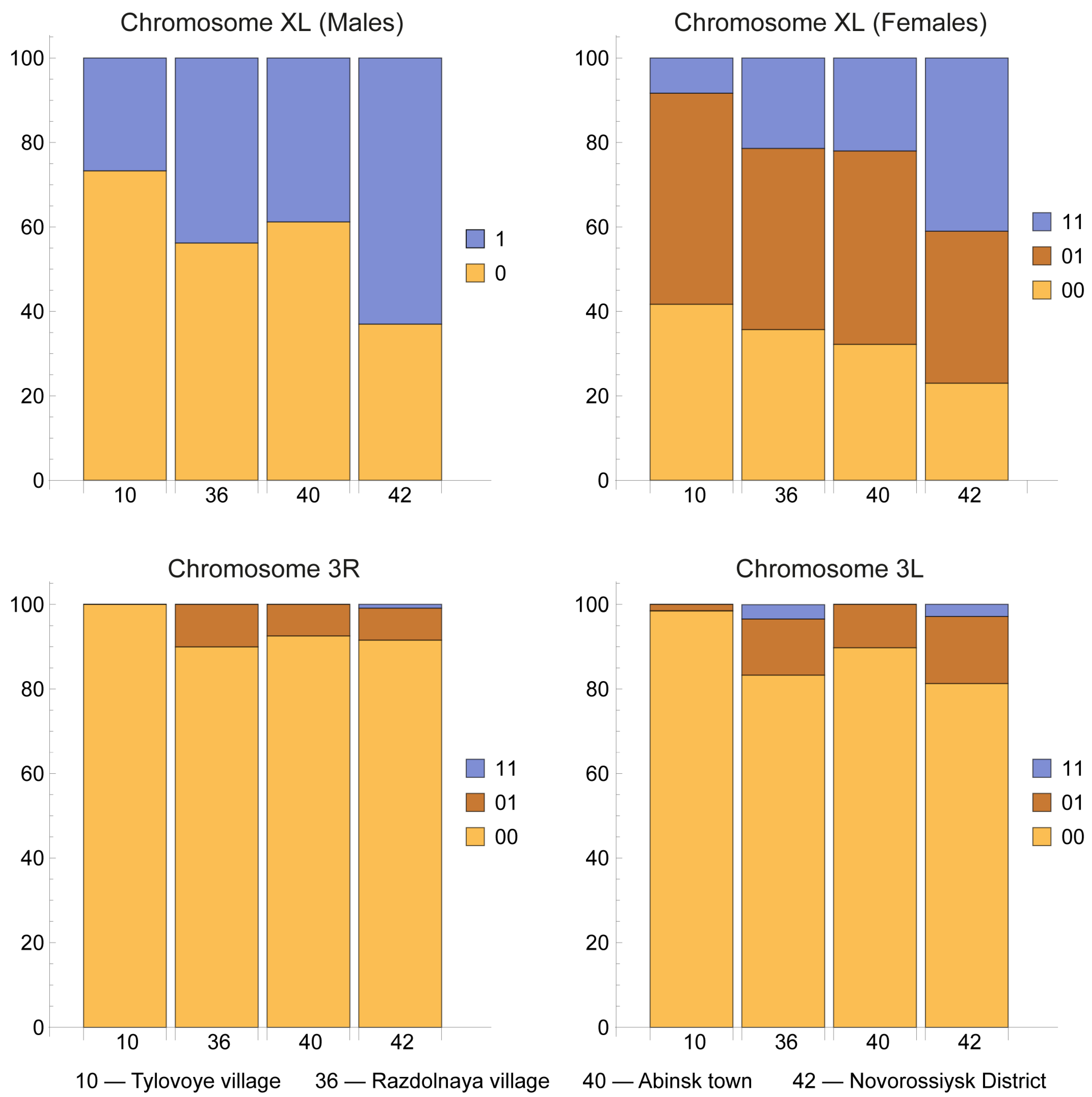
Figure 4.
A chromosomal complement of a squashed preparation of salivary gland cells in an Anopheles atroparvus female, with karyotype XL00, 2R00, 2L00, 3R00, and 3L01, stained by lacto-aceto-orcein, where XL, 2R, 2L, 3R, and 3L represent chromosome arms and the numbers 00 and 01 are chromosomal variants (objective lens – Nikon Plan 100x/1,25). The inversion heterozygous variant 3L01 (34b/c-38b) in An. atroparvus appears as a loop in the chromosome. CC – stands for the chromocenter. Chromosome arms XL, 2R, 2L, 3R, 3L are indicated. Scale bar equals 20 μm.
Figure 4.
A chromosomal complement of a squashed preparation of salivary gland cells in an Anopheles atroparvus female, with karyotype XL00, 2R00, 2L00, 3R00, and 3L01, stained by lacto-aceto-orcein, where XL, 2R, 2L, 3R, and 3L represent chromosome arms and the numbers 00 and 01 are chromosomal variants (objective lens – Nikon Plan 100x/1,25). The inversion heterozygous variant 3L01 (34b/c-38b) in An. atroparvus appears as a loop in the chromosome. CC – stands for the chromocenter. Chromosome arms XL, 2R, 2L, 3R, 3L are indicated. Scale bar equals 20 μm.

Table 1.
Species composition of malaria mosquitoes in breeding places of the Crimean Peninsula and the Black Sea coast of the Caucasus.
Table 1.
Species composition of malaria mosquitoes in breeding places of the Crimean Peninsula and the Black Sea coast of the Caucasus.
| No. | Location / breeding place | Latitude | Longitude | Date of sampling | Number (%) of mosquitoes | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Total | AT | CL | HY | MA | DA | PL | ML | |||||
| Crimean Peninsula | ||||||||||||
| 1 | Pirogovka village, Nakhimov district of Sevastopol /water storage | 44.685296 | 33.739026 | 11.09.2016 | 18 | - | - | - | 18 (100) | - | - | - |
| 2 | Bakhchisaray town / pond | 44.763889 | 33.853056 | 10.09.2016 | 12 | 3 (25,0) | - | - | 9 (75,0) | - | - | - |
| 3 | Bakhchisaray town / dried-up creek | 44.763889 | 33.853611 | 10.09.2016 | 10 | - | - | - | 10 (100) | - | - | - |
| 4 | Simferopol city, botanical garden / pond | 44.939167 | 34.133056 | 10.09.2016 | 15 | - | - | - | 15 (100) | - | - | - |
| 5*1 | Mazanka village, Simferopol district / pond | 45.014861 | 34.235861 | 12.07.2016 | 57 | - | - | - | 57 (100) | - | - | - |
| 6 | Konstantinovka village, Simferopol district / lake | 44.856389 | 34.123333 | 20.08.2016 | 3 | - | - | - | 3 (100) | - | - | - |
| 7 | Mramornoye village, Simferopol district / lake | 44.813889 | 34.237222 | 20.06.2016 | 9 | - | - | - | 7 (77,8) | - | 2 (22,2) | - |
| 8 | Mezhgorye village, Belogorsky district / river | 44.970556 | 34.416111 | 20.06.2016 | 9 | - | 5 (55,6) | - | 4 (44,4) | - | - | - |
| 9 | Krasnosyolovka village, Belogorsky district / river spill | 44.917778 | 34.633333 | 20.06.2016 | 2 | - | - | - | - | - | 2 (100) | - |
| 10A | Tylovoye village, Balaklava district of Sevastopol / pond | 44.441389 | 33.728056 | 13.09.2016 | 128 | 2 (1,6) | - | - | 60 (46,9) | 66 (51,5) | - | - |
| 10B*2 | 44.443570 | 33.739879 | 12.08.2017 | 21 | - | - | - | 12 (57,1) | 9 (42,9) | - | - | |
| 10C | 84 | 1 (1,2) | - | - | 83 (98,8) | - | - | - | ||||
| 10D*3 | 44.441740 | 33.727469 | 08.08.2019 | 53 | - | - | - | 23 (43,4) | 30 (56,6) | - | - | |
| 10E | 95 | - | - | - | 55 (57,9) | 40 (42,1) | - | - | ||||
| 11 | Rodnikovoye village, Balaklava district of Sevastopol / puddle | 44.453611 | 33.862222 | 20.07.2016 | 3 | - | - | - | - | - | 3 (100) | - |
| 12 | Rodnikovoye village, Balaklava district of Sevastopol / tree hollow | 44.457222 | 33.872778 | 20.07.2016 | 7 | - | - | - | - | - | 7 (100) | - |
| 13 | Simeiz Settlement, Yalta district / mountain puddle | 44.403611 | 33.991667 | 20.06.2016 | 5 | - | - | - | - | - | 5 (100) | - |
| 14 | Yalta district / forest puddle | 44.516389 | 34.143889 | 20.07.2016 | 37 | - | - | - | 37 (100) | - | - | - |
| 15 | Gaspra settlement, Yalta district / water in rock cracks | 44.433611 | 34.130000 | 20.06.2016 | 3 | - | - | - | - | - | 3 (100) | - |
| 16 | Gaspra settlement, Yalta district / tree hollow | 44.445278 | 34.118889 | 20.06.2016 | 2 | - | - | - | - | - | 2 (100) | - |
| 17 | Voskhod settlement, Yalta district / lake | 44.517417 | 34.219796 | 20.08.2016 | 8 | - | 8 (100) | - | - | - | - | - |
| 18 | Nikitsky Botanical Gardens, Yalta district / pond | 44.508831 | 34.233093 | 13.09.2016 | 7 | - | - | - | 1 (14,3) | - | 6 (85,7) | - |
| 19 | Krasnokamenka village, Yalta district / forest puddle | 44.577500 | 34.255556 | 20.06.2016 | 2 | - | - | - | - | - | 2 (100) | - |
| 20 | Zaprudnoye village, Alushta district / lake | 44.599444 | 34.305000 | 20.08.2016 | 14 | - | 14 (100) | - | - | - | - | - |
| 21 | Nizhnyaya Kutuzovka village, Alushta district / pond | 44.709842 | 34.377544 | 14.09.2016 | 31 | - | - | - | 31 (100) | - | - | - |
| 22 | Alushta district / pond | 44.814167 | 34.657778 | 14.09.2016 | 12 | - | - | - | 12 (100) | - | - | - |
| 23 | Gromovka village, Sudak district / pond | 44.857778 | 34.791389 | 20.06.2016 | 3 | - | 3 (100) | - | - | - | - | - |
| 24 | Voron village, Sudak district / spring | 44.892222 | 34.820278 | 20.08.2016 | 3 | - | 3 (100) | - | - | - | - | - |
| 25 | Veseloye village, Sudak district / water reserve | 44.849722 | 34.883611 | 15.09.2016 | 9 | - | - | - | 9 (100) | - | - | - |
| 26 | Sudak district / pond | 44.868611 | 34.900556 | 15.09.2016 | 3 | - | - | - | 3 (100) | - | - | - |
| 27 | Veseloye village, Sudak district / lake | 44.851944 | 34.883333 | 20.08.2016 | 2 | - | 2 (100) | - | - | - | - | - |
| 28 | Dachnoye village, Sudak district / river spill | 44.888889 | 34.990278 | 20.08.2016 | 1 | - | - | 1 (100) | - | - | - | - |
| 29 | Dachnoye village, Sudak district / lake | 44.897362 | 35.040173 | 20.08.2016 | 4 | - | - | 4 (100) | - | - | - | - |
| 30 | Mindalnoye village, Sudak district / lake | 44.831756 | 35.082243 | 20.07.2016 | 3 | - | - | 3 (100) | - | - | - | - |
| 31 | Grushevka village, Sudak district / lake | 45.010570 | 34.971796 | 15.09.2016 | 48 | - | - | - | 48 (100) | - | - | - |
| 32 | Feodosia city / pond | 45.063792 | 35.341071 | 16.09.2016 | 9 | 7 (77,8) | - | - | 2 (22,2) | - | - | - |
| Black Sea coast of the Caucasus | ||||||||||||
| 33 | Krasnogvardeyskoye village, Stavropol Krai / dried up river | 45.850476 | 41.482381 | 12.08.2015 | 27 | 27 (100) | - | - | - | - | - | - |
| 34 | Stavropol city / pond | 45.013332 | 41.974723 | 12.08.2015 | 30 | - | - | - | 26 (86,7) | 4 (13,3) | - | - |
| 35 | Malevanyi settlement, Krasnodar Krai / river | 45.531517 | 39.461591 | 21.08.2015 | 50 | - | - | - | 1 (2,0) | 49 (98,0) | - | - |
| 36 | Razdolnaya village, Korenovsky district, Krasnodar Krai / lake | 45.383469 | 39.537257 | 01.08.2024 | 100 | - | - | 18 (18,0) | 52 (52,0) | 30 (30,0) | - | - |
| 37 | Shengzhiy settlement, Republic of Adygeya / channel | 44.883810 | 39.075139 | 05.08.2009 | 54 | - | - | 1 (1,9) | 2 (3,7) | 51 (94,4) | - | - |
| 38 | Novonikolayevskaya village, Krasnodar Krai / pond | 45.581165 | 38.369233 | 04.08.2019 | 120 | - | - | - | - | 120 (100) | - | - |
| 39 | Tamanskoye Settlement, Temryuksky district, Krasnodar Krai / lake | 45.144803 | 36.700849 | 07.07.2016 | 35 | 35 (100) | - | - | - | - | - | - |
| 40*4 | Abinsk town, Krasnodar Krai / pond | 44.862380 | 38.183556 | 14.08.2018 | 108 | - | - | - | - | 108 (100) | - | - |
| 41 | Gaiduk village, Novorossiysk district, Krasnodar Krai / pond | 44.781486 | 37.679653 | 03.08.2018 | 100 | - | - | - | - | 100 (100) | - | - |
| 42 | Novorossiysk district, Krasnodar Krai / water storage | 44.780000 | 37.815833 | 09.07.2016 | 108 | - | - | - | 1 (0,9) | 107 (99,1) | - | - |
| 43 | Pshada village, Gelendzhik district, Krasnodar Krai / river | 44.452257 | 38.346501 | 19.08.2015 | 106 | - | - | - | 70 (66,0) | 36 (34,0) | - | - |
| 44 | Community Zarya, Tuapse district, Krasnodar Krai /car tire | 44.082778 | 39.131667 | 31.07.2021 | 36 | - | - | - | - | - | 36 (100) | - |
| 45 | Novomikhailovsky settlement, Tuapse district, Krasnodar Krai / drainage ditch | 44.247752 | 38.844524 | 18.08.2015 | 100 | - | 100 (100) | - | - | - | - | - |
| 46 | Agui-Shapsug village, Tuapse district, Krasnodar Krai / river | 44.174722 | 39.066944 | 11.07.2016 | 35 | - | - | - | 35 (100) | - | - | - |
| 47 | Zubova Shchel village, Sochi district, Krasnodar Krai / river | 43.837451 | 39.441109 | 13.07.2016 | 34 | - | - | - | 34 (100) | - | - | - |
| 48 | Sochi city, Krasnodar Krai / tree hollow | 43.675833 | 39.608889 | 30.07.2021 | 60 | - | - | - | - | - | 60 (100) | - |
| 49 | Adler town, Krasnodar Krai / swamp | 43.432222 | 39.947222 | 17.07.2016 | 32 | - | - | - | 32 (100) | - | - | - |
| 50A | Verkhneveseloye village, Sochi district, Krasnodar Krai / drainage ditch | 43.426067 | 39.973288 | 04.08.2023 | 14 | - | 14 (100) | - | - | - | - | - |
| 50B*5 | 6 | - | 6 (100) | - | - | - | - | - | ||||
| 50C | 43.426306 | 39.973515 | 04.08.2024 | 3 | - | 3 (100) | - | - | - | - | - | |
| 51*6 | Sochi city, Krasnodar Krai / stream | 43.410321 | 39.983947 | 07.08.2024 | 2 | - | - | - | - | - | - | 2 (100) |
| 52 | Sirius settlement, Krasnodar Krai / fire pond | 43.412778 | 39.937778 | 16.07.2016 | 156 | - | - | - | 156 (100) | - | - | - |
| 53 | Vesyoloye microdistrict, Sochi city, Krasnodar Krai / car tire | 43.409722 | 40.008330 | 11.08.2018 | 32 | - | - | - | - | - | 32 (100) | - |
| 54 | Krasnaya Polyana Resort, Sochi district, Krasnodar Krai / hollow tree | 43.711944 | 40.209167 | 25.07.2021 | 94 | - | - | - | - | - | 94 (100) | - |
| 55 | Rosa Khutor resort, Sochi district, Krasnodar Krai /hollow tree | 43.638978 | 40.307983 | 29.07.2021 | 39 | - | - | - | - | - | 39 (100) | - |
| 56 | Ritsinsky National Park, Gudauta district, Abkhazia /car tire | 43.473889 | 40.538056 | 24.07.2021 | 16 | - | - | - | - | - | 16 (100) | - |
No., location number; AT – An. atroparvus; CL – An. claviger; HY – An. hyrcanus; MA – An. maculipennis; DA – An. daciae; PL – An. plumbeus; ML – An. melanoon. *1 – sequences accession numbers GenBank ID PQ510968–PQ511024 (57); *2 – sequences accession numbers PQ554994–PQ555014 (21); *3 – sequences accession numbers PQ550752–PQ550804 (53); *4 – sequences accession numbers PQ526514–PQ526598 (85); *5 – sequences accession numbers PQ740514–PQ740518 (5); *6 – sequences accession number ID PQ740513 (1).
Table 2.
Ecological characteristics of breeding places of malaria mosquitoes of the Crimean Peninsula and the Black Sea coast of the Caucasus.
Table 2.
Ecological characteristics of breeding places of malaria mosquitoes of the Crimean Peninsula and the Black Sea coast of the Caucasus.
| No. | Location / breeding place | Latitude | Longitude | Date of sampling | Density of larvae (1-4 instars/sq. m) | Ecological characteristics of habitats | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| h (m) | pH | T (°C) | ppt | O₂ (mg/L) | ||||||
| Crimean Peninsula | ||||||||||
| 1 | Pirogovka village, Nakhimov district of Sevastopol /water storage | 44.685296 | 33.739026 | 11.09.2016 | 45 | 63 | 8,05 | 30,0 | 0,34 | 6,0 |
| 2 | Bakhchisaray town / pond | 44.763889 | 33.853056 | 10.09.2016 | 28 | 160 | 8,05 | 18,0 | 0,59 | 7,7 |
| 3 | Bakhchisaray town /dried-up creek | 44.763889 | 33.853611 | 10.09.2016 | 160 | 160 | 7,70 | 20,7 | 0,70 | 4,5 |
| 4 | Simferopol city, botanical garden / pond | 44.939167 | 34.133056 | 10.09.2016 | 76 | 255 | 6,48 | 22,6 | 0,16 | 4,5 |
| 5 | Mazanka village, Simferopol district / pond | 45.014861 | 34.235861 | 12.07.2016 | - | 298 | 8,15 | 24,5 | 0,26 | 7.0 |
| 6 | Konstantinovka village, Simferopol district / lake | 44.856389 | 34.123333 | 20.08.2016 | 3 | 421 | 7,84 | 24,8 | 2,56 | - |
| 7 | Mramornoye village, Simferopol district / lake | 44.813889 | 34.237222 | 20.06.2016 | 7 | 493 | 7,62 | 16,5 | 2,14 | - |
| 8 | Mezhgorye village,Belogorsky district / river | 44.970556 | 34.416111 | 20.06.2016 | 9 | 385 | 7,22 | 21,6 | 1,21 | - |
| 9 | Krasnosyolovka village, Belogorsky district / river spill | 44.917778 | 34.633333 | 20.06.2016 | 2 | 401 | 7,62 | 18,3 | 1,94 | - |
| 10 | Tylovoye village, Balaklava district of Sevastopol / pond | 44.441389 | 33.728056 | 13.09.2016 | 15 | 295 | 8,15 | 22,1 | 0,22 | 7,8 |
| 11 | Rodnikovoye village, Balaklava district of Sevastopol / puddle | 44.453611 | 33.862222 | 20.07.2016 | 3 | 657 | 7,32 | 32,3 | 0,14 | - |
| 12 | Rodnikovoye village, Balaklava district of Sevastopol /tree hollow | 44.457222 | 33.872778 | 20.07.2016 | 7 | 434 | 7,14 | 24,7 | 0,04 | - |
| 13 | Simeiz Settlement, Yalta District / mountain puddle | 44.403611 | 33.991667 | 20.06.2016 | 5 | 149 | 8,32 | 18,0 | 0,17 | - |
| 14 | Yalta district / forest puddle | 44.516389 | 34.143889 | 20.07.2016 | 7 | 212 | 7,24 | 19,7 | 0,15 | - |
| 15 | Gaspra settlement, Yalta District / water in rock cracks | 44.433611 | 34.130000 | 20.06.2016 | 3 | 24 | 7,52 | 24,8 | 0,31 | - |
| 16 | Gaspra settlement, Yalta district / tree hollow | 44.445278 | 34.118889 | 20.06.2016 | 2 | 338 | 7,16 | 25,1 | 0,03 | - |
| 17 | Voskhod settlement, Yalta district / lake | 44.517417 | 34.219796 | 20.08.2016 | 8 | 330 | 7,30 | 21,5 | 0,24 | - |
| 18 | Nikitsky Botanical Gardens, Yalta district / pond | 44.508831 | 34.233093 | 13.09.2016 | 15 | 110 | 7,37 | 20,2 | 0,31 | 7,5 |
| 19 | Krasnokamenka village, Yalta district / forest puddle | 44.577500 | 34.255556 | 20.06.2016 | 2 | 770 | 7,23 | 14,9 | 0,11 | - |
| 20 | Zaprudnoye village, Alushta district / lake | 44.599444 | 34.305000 | 20.08.2016 | 14 | 614 | 7,14 | 17,5 | 0,21 | - |
| 21 | Nizhnyaya Kutuzovka village, Alushta district / pond | 44.709842 | 34.377544 | 14.09.2016 | 78 | 153 | 7,29 | 25,2 | 0,20 | 10,2 |
| 22 | Alushta district / pond | 44.814167 | 34.657778 | 14.09.2016 | 70 | 70 | 7,05 | 22,4 | 0,98 | 4,4 |
| 23 | Gromovka village, Sudak district / pond | 44.857778 | 34.791389 | 20.06.2016 | 3 | 157 | 7,52 | 16,5 | 2,08 | - |
| 24 | Voron village, Sudak district / spring | 44.892222 | 34.820278 | 20.08.2016 | 3 | 229 | 7,72 | 16,7 | 0,74 | - |
| 25 | Veseloye village, Sudak district / water reserve | 44.849722 | 34.883611 | 15.09.2016 | - | 131 | 7,28 | 23,2 | 0,81 | 2,3 |
| 26 | Sudak district / pond | 44.868611 | 34.900556 | 15.09.2016 | - | 125 | 8,31 | 22,6 | 0,25 | - |
| 27 | Veseloye village, Sudak district / lake | 44.851944 | 34.883333 | 20.08.2016 | 2 | 101 | 7,42 | 23,7 | 0,24 | - |
| 28 | Dachnoye village, Sudak district / river spill | 44.888889 | 34.990278 | 20.08.2016 | - | 76 | 7,47 | 22,4 | 3,02 | - |
| 29 | Dachnoye village, Sudak district / lake | 44.897362 | 35.040173 | 20.08.2016 | 4 | 239 | 7,54 | 23,8 | 0,92 | - |
| 30 | Mindalnoye village, Sudak district / lake | 44.831756 | 35.082243 | 20.07.2016 | 3 | 33 | 8,24 | 26,3 | 1,18 | - |
| 31 | Grushevka village, Sudak district / lake | 45.010570 | 34.971796 | 15.09.2016 | 29 | 223 | 6,93 | 22,0 | 0,27 | 8,0 |
| 32 | Feodosia city / pond | 45.063792 | 35.341071 | 16.09.2016 | 26 | 20 | 7,14 | 24,3 | 1,46 | 16,5 |
| Black Sea coast of the Caucasus | ||||||||||
| 33 | Krasnogvardeyskoye village, Stavropol Krai / dried up river | 45.850476 | 41.482381 | 12.08.2015 | 1 | 60 | 9,10 | 28,0 | 5,99 | 11,3 |
| 34 | Stavropol city / pond | 45.013332 | 41.974723 | 12.08.2015 | - | 484 | 8,85 | 26,5 | 0,20 | 7,9 |
| 35 | Malevanyi settlement, Krasnodar Krai / river | 45.531517 | 39.461591 | 21.08.2015 | 1 | 40 | 8,00 | 24,0 | 1,58 | 6,9 |
| 36 | Razdolnaya village, Korenovsky district,Krasnodar Krai / lake | 45.383469 | 39.537257 | 01.08.2024 | 10 | 47 | 8,06 | 25,6 | 1,01 | 10,0 |
| 37 | Shengzhiy settlement, Republic of Adygeya / channel | 44.883810 | 39.075139 | 05.08.2009 | - | 44 | 7,00 | 33,3 | 0,81 | - |
| 38 | Novonikolayevskaya village, Krasnodar Krai / pond | 45.581165 | 38.369233 | 04.08.2019 | 29 | 2 | 8,50 | 25,5 | 0,19 | 8,0 |
| 39 | Tamanskoye Settlement, Temryuksky district, Krasnodar Region / lake | 45.144803 | 36.700849 | 07.07.2016 | 11 | 155 | 8,00 | 20,5 | - | 8,0 |
| 40 | Abinsk town, Krasnodar Krai / pond | 44.862380 | 38.183556 | 14.08.2018 | - | 29 | 7,60 | 30,0 | 0,27 | 5,0 |
| 41 | Gaiduk village, Novorossiysk district, Krasnodar Krai / pond | 44.781486 | 37.679653 | 03.08.2018 | 16 | 98 | 7,40 | 26,2 | 0,27 | 4,0 |
| 42 | Novorossiysk District, Krasnodar Krai / water storage | 44.780000 | 37.815833 | 09.07.2016 | 14 | 162 | 8,40 | 21,5 | - | 5,0 |
| 43 | Pshada village, Gelendzhik District, Krasnodar Krai / river | 44.452257 | 38.346501 | 19.08.2015 | 56 | 27 | 7,30 | 21,8 | 0,48 | 8,1 |
| 44 | Community Zarya, Tuapse district, Krasnodar Krai /car tire | 44.082778 | 39.131667 | 31.07.2021 | - | 172 | 5,50 | 26,5 | 1,45 | 2,5 |
| 45 | Novomikhailovsky settlement, Tuapse district, Krasnodar Krai / drainage ditch | 44.247752 | 38.844524 | 18.08.2015 | 32 | 5 | 7,54 | 23,4 | 0,45 | 2,5 |
| 46 | Agui-Shapsug village, Tuapse district, Krasnodar Krai / river | 44.174722 | 39.066944 | 11.07.2016 | 67 | 31 | 7,80 | 22,0 | - | 8,0 |
| 47 | Zubova Shchel village, Sochi district, Krasnodar Krai / river | 43.837451 | 39.441109 | 13.07.2016 | 72 | 175 | 7,80 | 24,0 | - | 8,0 |
| 48 | Sochi city, Krasnodar Krai / tree hollow | 43.675833 | 39.608889 | 30.07.2021 | - | 74 | 6,00 | 24,0 | 2,04 | 3,0 |
| 49 | Adler town, Krasnodar Krai / swamp | 43.432222 | 39.947222 | 17.07.2016 | 1,5 | 45 | 8,20 | - | - | 6 |
| 50 | Verkhneveseloye village, Sochi district, Krasnodar Krai / drainage ditch | 43.426306 | 39.973515 | 04.08.2024 | 3 | 31 | 9,26 | 27,8 | 0,54 | 7,6 |
| 51 | Sochi city, Krasnodar Krai / stream | 43.410321 | 39.983947 | 07.08.2024 | - | 14 | 7,60 | - | - | - |
| 52 | Sirius settlement, Krasnodar Krai / fire pond | 43.412778 | 39.937778 | 16.07.2016 | 18 | 15 | 8,40 | 27,0 | - | 4 |
| 53 | Vesyoloye microdistrict, Sochi city, Krasnodar Krai / car tire | 43.409722 | 40.008330 | 11.08.2018 | - | 13 | 7,40 | 27,3 | 0,25 | - |
| 54 | Krasnaya Polyana Resort, Sochi district, Krasnodar Krai / hollow tree | 43.711944 | 40.209167 | 25.07.2021 | - | 1693 | 5,50 | 22,6 | 2,30 | 3,5 |
| 55 | Rosa Khutor resort, Sochi district, Krasnodar Krai / hollow tree | 43.638978 | 40.307983 | 29.07.2021 | - | 1708 | 5,20 | 21,5 | 1,50 | 3,0 |
| 56 | Ritsinsky National Park, Gudauta district, Abkhazia /car tire | 43.473889 | 40.538056 | 24.07.2021 | - | 1041 | 6,00 | 21,3 | 2,78 | 2,5 |
No., location number; h (m), altitude above sea level in meters; pH, hydrogen index; T (°C), water temperature in Celsius degrees; ppt, total dissolved solids in grams per liter (parts per thousand); O₂ (mg/L), quantity of dissolved oxygen in water.
Table 3.
The frequencies of chromosomal variants in populations of An. daciae in the Crimean Peninsula and the Black Sea coast of the Caucasus.
Table 3.
The frequencies of chromosomal variants in populations of An. daciae in the Crimean Peninsula and the Black Sea coast of the Caucasus.
| Inversion homo- and heterozygotes | Frequencies of chromosomal variants, f ± Sf, % | |||
|---|---|---|---|---|
| Crimean Peninsula | Black Sea coast of the Caucasus | |||
| Location 10 | Location 36 | Location 40 | Location 42 | |
| Males, n | 30 | 16 | 49 | 46 |
| XL0 | 73,3±8,1 | 56,2±12,4 | 61,2±7,0 | 37,0±7,1 |
| XL1 | 26,7±8,1 | 43,8±12,4 | 38,8±7,0 | 63,0±7,1 |
| Females, n | 36 | 14 | 59 | 61 |
| XL00 | 41,7±8,2 | 35,7±12,8 | 32,2±6,1 | 23,0±5,4 |
| XL01 | 50,0±8,3 | 42,9±13,2 | 45,8±6,5 | 36,0±6,1 |
| XL11 | 8,3±4,6 | 21,4±11,0 | 22,0±5,4 | 41,0±6,3 |
| Both sexes, n | 66 | 30 | 108 | 107 |
| 2R00 | 100 | 100 | 100 | 99,1±0,9 |
| 2R05 | 0 | 0 | 0 | 0,9±0,9 |
| 2L00 | 100 | 100 | 100 | 100 |
| 3R00 | 100 | 90,0±5,5 | 92,6±2,5 | 91,6±2,7 |
| 3R01 | 0 | 10,0±5,5 | 7,4±2,5 | 7,5±2,5 |
| 3R11 | 0 | 0 | 0 | 0,9±0,9 |
| 3L00 | 98,5±1,5 | 83,3±6,8 | 89,8±2,9 | 81,3±3,8 |
| 3L01 | 1,5±1,5 | 13,3±6,2 | 10,2±2,9 | 15,9±3,5 |
| 3L11 | 0 | 3,3±3,3 | 0 | 2,8±1,6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



