
Submitted:
28 January 2025
Posted:
30 January 2025
You are already at the latest version
Abstract
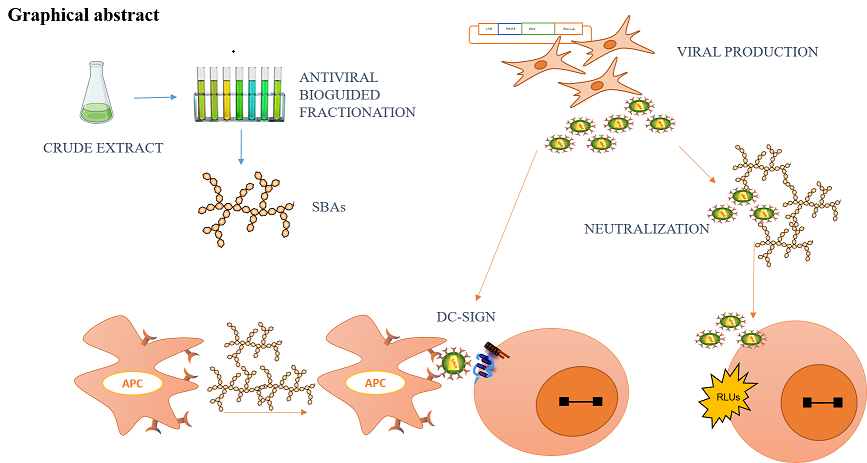
Medicinal plants are widely used in bolivian folk medicine for the treatment of infectious diseases. We have selected one, Clinopodium bolivianum (Benth.) Kuntze, known as “Khoa”, to investigate its potential anti-HIV activity, since traditionally it has been used to treat other viral infectious diseases. We have carried out an antiviral bioassay-guided fractionation of different extracts of C. bolivianum. An antiviral crude polysaccharide was obtained, SBAS, rich in glucose, galactose, mannose, arabinose, xylose, rhamnose and only traces of galacturonic acid. SBAS exhibited antiviral activity with a mechanism of action unrelated to the mannose-lectin DC-SIGN receptors but with a strong viral neutralization activity. In summary, a purified polysaccharide from C. bolivianum have been identified as the main compound responsible of its antiviral activity. SBAs proved to be a neutralizing agent with high antiviral capacity in vitro, so it could be part of new microbicide formulations to prevent HIV transmission.
Keywords:
Clinopodium
; Satureja
; antiviral
; HIV
; polysaccharides
; microbicides
1. Introduction
Polysaccharides isolated from natural resources, e.g. plant, animal, fungi or seaweed, are gaining attention due to their wide variety of pharmacological activities, including antitumor, immunomodulatory, anti-oxidant, antiviral or anti-inflammatory effects [1]. Polysaccharides display several advantages as bioactive molecules compared to other chemical types of natural products, as easy to find available sources and low toxicity and side effects.
As antiviral compounds, the first report involving high-molecular-weight polysaccharides was published more than 70 years ago [2]. Currently, research is focused on immune booster applications and anti-infective activity. Numerous studies show that natural and chemically modified polysaccharides have a significant inhibitory effect against human immunodeficiency virus (HIV) [3,4], herpes simplex virus (HSV) [5] and other viruses like influenza A virus (IAV), and hepatitis B virus (HBV) [6] displaying a variety of mechanisms.
HIV antiretroviral therapy involves the combination of several drugs that must be administered throughout life. This life-long therapy with side effects and viral resistances emergence led to the search of preventive HIV infection therapies, including topical microbicides. Microbicides act through several mechanisms, as neutralizing agents, disrupting the viral particle integrity before it can reach the target cell. Another type of microbicides includes DC-SIGN trans-infection inhibitors, preventing the presentation of the virus to CD4+ T cells. Finally, antiretroviral drugs targeting viral proteins, as tenofovir or cabotegravir, are among the most studied microbicides [7]. In all the cases, the aim is to avoid viral DNA integration into the cell host genome, since, when this occurs, it leads to the establishment of viral reservoirs and lifelong infections.
Clinopodium bolivianum (Benth.) Kuntze (syn. Satureja boliviana (Benth.) Briq.) is native to Bolivia, Perú and Argentina [8]. Popularly known as khoa, muña or inca muña, belongs to the Lamiaceae family and it is used in bolivian folk medicine to treat influenza and as an antiseptic, anthelmintic, insecticide and anti-inflammatory drug [9,10]. Other species of Clinopodium have previously shown a variety of biological activities [11-17], and one of them, S. montana showed antiviral activity due to water soluble polar substances [18].
On the other side, the major constituents identified in C. bolivianum were phenolic compounds, such as kaempferol 3-O-glucoside, kaempferol 3-O-xylosylglucoside, kaempferol 7-O-rhamnoside and quercetin 3-O-sophoroside [19]. There are also reports about the isolation of essential oils [20] and diterpenoid-like compounds [21]. These reports hearten the chemical investigation of the active components of this plant species.
As part of a screening program to investigate the pharmacological activity of higher plants, traditional medicinal plants from the bolivian tropical forest used for the treatment of viral diseases, were submitted to antiviral evaluation. Aqueous or ethanolic extracts of seven species, including C. bolivianum, were assayed for inhibition of HSV-1, vesicular stomatitis virus (VSV) and poliovirus type 1 replication in vitro, revealing that the aqueous extract of this plant displayed antiviral activity, possibly due to polysaccharide substances [22].
The present paper describes the isolation of a highly purified polysaccharide from the water-soluble extract of the aerial parts of C. bolivianum, the identification and quantification of their major constituents and the evaluation of its antiviral activity and mechanism of action.
2. Materials and Methods
2.1. Plant material and chemicals
The aerial parts of Clinopodium bolivianum (Benth.) Kuntze (syn. Satureja boliviana (Benth) Briq). 'WFO (2024): Clinopodium bolivianum (Benth.) Kuntze. Published on the Internet; http://www.worldfloraonline.org/taxon/wfo-0000890868. Accessed on: 09 Oct 2024. Plant specimens were collected by Eduardo Gonzales Dávalos in Huarina municipality, Omasuyos province, La Paz, Bolivia (latitude 16°11′39.81′′S and longitude 68°36′16.43′′W) and identified and certified by the Herbario Nacional de Bolivia, Universidad Mayor de San Andrés. A voucher specimen (No. EG-1, Labiatae) was identified by the Herbario Nacional de Bolivia and has been deposited at the Department of Pharmacology at the Instituto de Investigaciones Fármaco Bioquímicas, UMSA, La Paz, Bolivia.
Most of the chemicals used in the experiments, including glucose, saccharose, cellobiose, trehalose, galactose, mannose, arabinose, maltose, xylose, rhamnose, galacturonic acid, glucuronic acid, trifluoroacetic acid (TFA) and dimethyl sulfoxide (DMSO) were purchased from Merck (Lyon, France). Other chemicals used in this study were of analytical grade and were purchased from Panreac (Barcelona, Spain).
2.2. Fractionation of C. bolivianum aerial parts
A bioassay-guided fractionation was carried out with 20 g of aerial parts of C. bolivianum. 10 g were decocted (30 min at 100 0C) to half volume and lyophilized to obtain a hot water extract denominated SBA (yield: 24.2 %). Another 10 g were extracted with ethanol 95 % (100 mL, 30 min at room temperature) and concentrated to obtain an ethanol extract (SBB, yield: 12.1 %). The antiviral activity of both extracts was evaluated, and SBA was selected as the more active one (data not shown). Thus, SBB was discarded for further analysis, selecting SBA for additional fractionation. SBA was subsequently treated with a mixture of water/EtOH (1:3) and centrifuged to separate the ethanol-soluble material (SBA-0) from the precipitate (SBA-1), that was lyophilized.
Fraction SBA-1 was then treated with 1 M NaOH, the soluble material was recovered and treated with one volume of 96% ethanol (room temperature, overnight), recovering the resulting precipitate by centrifugation. To eliminate sodium hydroxide and other impurities, this material was resuspended in distilled water and dialyzed against water for 4 days using Visking membranes with a molecular cut-off of 12-14 kDa (Medicell International Ltd.®, Islington, London, U.K). The retentate was then centrifuged to separate an insoluble fraction (SBAI) from a soluble fraction (SBAS), whose antiviral activity and composition were studied.
2.3. Determination of total neutral sugars
Total carbohydrates were calorimetrically determined by the previously described phenol-sulfuric acid method [23].
2.4. Analysis of the polysaccharides
2.4.1. Monosaccharide composition analysis
A sample of the SBAS fraction (1 mg) was hydrolyzed with 1 mL of 3 M trifluoroacetic acid (TFA) at 120 0C for 1 h. Immediately, 1 mg of myo-inositol was added as internal standard to each hydrolysate, and the mixture was dried in a rotary evaporator. Prior to GC-MS analysis, the monosaccharides released were derivatized as trimethylsilylated oximes. To do so, 250 µL of a solution of hydroxylamine chloride in 2.5% pyridine were first added to each sample and kept at 70 0C for 30 min, and then 150 µL of bis-trimethylsilyl trifluoroacetamide (BSTFA) were incorporated to the previous mixture and maintained at 80 0C for 10 min [24]. The analysis of the samples was performed by gas chromatography-mass spectrometry (GC-MS) in an Agilent 7980A-5975C instrument (Agilent Technologies, CA, USA), with a column HP-5MS (30 m x 0.25 mm, 0.2 µm) with a temperature program from 180 0C to 2800C with a temperature ramp of 2 0C/min and 1 min at the final temperature. Monosaccharides were identified and quantified using mixtures of commercial monosaccharides with 1 mg of myo-inositol, derivatized and analyzed in identical conditions.
2.4.2. Fourier transform-infrared (FT-IR) spectrometry
The alpha or beta configuration of the monomers, as well as the presence of organic functional groups was assessed by FT-IR. The samples were acidified with hydrochloric acid, dialyzed, dried and use to prepare a solid pellet for FT-IR measurement. Briefly, 1-2 mg of dry sample was mixed with 300 mg of KBr, grinded to fine powder, and then pressed to obtain the solid pellet, that was analyzed in an FTIR 4200 spectrometer (Jasco Corporation®, Tokyo, Japan) in the range of 400-4000 cm-1, using a detector TGS with a resolution of 4 cm-1.
2.4.3. Determination of linkage types in SBAS by methylation analysis
The fraction SBAS was permethylated as previously reported [25,26] and then treated with deuterated sodium borohydride in ethanol 75% to reduce the acid groups of uronic acids to their corresponding neutral monosaccharide before hydrolysis since, otherwise, they are not detected [27]. The monomers sensitive to this treatment carry a mass tag of 2 Da due to the deuterated reagent, which allows differentiating them by mass spectrometry. The product was next hydrolyzed with 2 M TFA, reduced again with deuterated sodium borohydride, and acetylated to obtain a mixture of partially methylated alditol acetates. The final products were analyzed by GC-MS in an Agilent 7980A-5975C (Agilent Technologies, CA, USA), with a HP-5MS column (30 m x 0,25 mm, 0,2µm) and using a temperature program: 1700C (1 min), and 20C/min from 170 to 2100C (1 min). The injector and transfer line were kept at 250 0C. The partially methylated alditol acetates were identified by their retention time and mass spectrum.
2.5. Antiviral activity
2.5.1. Cell culture reagents
MT-2 (obtained from American Type Culture Collection, Manassas, VA, USA, Ref: CRL-2560) and RAJI DC-SIGN cells (kindly provided by Dr. Fernando Arenzana-Seisdedos, Institut Pasteur, Paris, France) were cultured in RPMI-1640 medium while 293T cells (obtained from American Type Culture Collection, Manassas, VA, USA, Ref: CRL-3216) were cultured in DMEM medium, all of them containing 10% (v/v) fetal bovine serum, 2 mmol/L l-glutamine, penicillin (50 IU/ml) and streptomycin (50 μg/mL) (all Whittaker M.A. Bio-Products, Walkerville, Maryland, USA). Cells were maintained at 37°C in a 5% CO2 humidified atmosphere and split twice a week. Buffy Coats from anonymous healthy blood donors were obtained from the Centro de Transfusiones de la Comunidad de Madrid. Proper informed consent was obtained from each subject in accordance with the Spanish legislation on blood donor regulations. Confidentiality and privacy were assured. Sex and gender-based analyses were not performed since the gender of the donor was not disclosed. PBMCs (peripheral blood mononuclear cells) were isolated from Buffy Coats by centrifugation through a Ficoll-Hypaque gradient (Pharmacia Corporation, North Peapack, NJ) and were resuspended in RPMI 1640 medium supplemented with 10% fetal bovine serum, 2 mM L-glutamine and antibiotics (100 mg/L streptomycin and 100 U/mL penicillin) (all Whittaker M.A. Bio-Products, Walkerville, MD, USA) before culture at a concentration of 2 × 106 cells/mL. PBMCs viability was maintained by the addition of Interleukin-2 (IL-2) (Emeryville, CA. USA) every 48 hours.
2.5.2. Plasmids and virus
Plasmid pNL4.3-Ren has been previously described by our group [28]. Viral supernatants were obtained from 293T cells transfection by calcium phosphate method using the plasmid above-mentioned. These supernatants were titrated in serial dilution infections of the same cells used for the experiments by RLUs obtained in a luminometer (Berthodl Detection Systems, Pforzheim, Germany).
2.5.3. Anti-HIV assays
Infectious supernatants were used to infect cells in the presence or absence of different concentrations of the fractions or the polysaccharides obtained. The evaluation of the anti-HIV activity was performed by two different methods, neutralization assay and trans-infection assays.
2.5.3.1. Neutralization assay
Neutralization assays were performed using recombinant HIV supernatants treated with different concentrations of polysaccharides or controls prior to infection. This assay is useful to detect those compounds acting through inactivation of viral particles. Briefly, recombinant HIV-1 supernatants (NL4.3-Ren) were incubated with active concentrations of the samples for 1 h prior to the infection of MT-2 cells. The neutralization process of the HIV particle was developed at four different concentrations (500, 50, 5 and 0.5 μg/ml) to allow the calculation of neutralizing IC50. Then, treated supernatants were diluted to the final infection concentration and added to the MT-2 cell culture. A dilution with the same concentration of the diluted remnant polysaccharides in the viral supernatants were used as control. After 48 hours, cells were lysed with 100 μL of Renilla lysis buffer system (Promega, Madison, Wisconsin, USA) and relative luminescence units (RLUs) were obtained in a luminometer (Berthold Detection Systems, Pforzheim, Germany) after the addition of substrate to cell extracts following the specifications of the manufacturer (Promega, Madison, Wisconsin, USA).
2.5.3.2. Transinfection assays
Antigen presenting cells (APCs) mediated trans-infection was tested using a cell line expressing the DC-SIGN receptor (RAJI DC-SIGN), one of the main receptors used by APCs to deliver the virus to CD4+ T cells in lymph nodes. Primary CD4+T cells from PBMCs obtained from healthy patients were used as target cells. Briefly, RAJI or RAJI-DC-SIGN cells were incubated with polysaccharides and mannan (inhibition control) at final concentrations of 10 or 100 μg/mL, or PBS as vehicle control (100%). An hour later, viral supernatants (NL4.3-Renilla) were added to the culture and cell adsorption allowed in rotation for two hours. Afterwards, cells were collected, washed thoroughly and cultured in RPMI, with IL-2 pre-activated PBMCs, to assess the inhibition of viral transfer by RAJI-DC-SIGN cells [29]. After 48 hours in culture, cells were lysed with 100 μl of Renilla lysis buffer system (Promega, Madison, Wisconsin, USA) and relative luminescence units (RLUs) were obtained in a luminometer (Berthold Detection Systems, Pforzheim, Germany). RAJI cells were used as DC-SIGN negative trans-infection control.
2.5.4. Cell Viability
Cell viability in treated mock-infected cells was measured in parallel with the same conditions as in the antiviral assays following the manufacturer instructions of CellTiter Glo assay system (Promega, Madison, Wisconsin, USA).
2.5.5. Statistics
Inhibitory concentrations 50% (CI50), cytotoxic concentrations 50% (CC50) calculations and ANOVA analysis were performed using GraphPad Prism software v 8.0 (La Jolla, California, USA).
3. Results and discussion
3.1. Bioguided assay fractionation of C. bolivianum aerial parts
C. bolivianum aerial parts were extracted with hot water obtaining the SBA fraction, that represented 24.2% of the starting dry material, with anti-HIV activity (data not shown). The total carbohydrates of the water extract (SBA), determined by the phenol-sulfuric acid, amounted 141 mg/L. To get purest fractions with enhanced antiviral activity, the water extract SBA was sequentially fractionated based on its anti-HIV activity (data not shown). First, treatment with ethanol gave an ethanol-soluble fraction (SBA-0, yield: 15.7%) and an abundant precipitate (SBA-1, yield: 84.3%) that kept the antiviral activity. Then, SBA-1 was recovered and treated with 1 M NaOH to solubilize polysaccharides, simultaneously inducing degradation of proteins and esters to smaller components. The alkali-soluble extract was recovered, and the polysaccharides were precipitated from this solution by adding one volume of 96% ethanol. The precipitate was dialyzed against water and, during dialysis, some water-insoluble polysaccharides precipitate inside the membrane, that were separated from hydrosoluble polysaccharides by centrifugation. At the end of the fractionation process, a minor insoluble fraction (SBAI, yield: 0.9%) was separated from a fourfold more abundant soluble fraction (SBAS, yield: 4.6%), highly enriched in polysaccharides.
3.2. Antiviral activity and cell viability assay
Purified polysaccharides SBAS and SBAI were submitted to antiviral activity evaluation. The experimental approach chosen was a neutralization assay since polysaccharide are high molecular weight molecules ant its activity is more likely to be produced in an extracellular environment. Actually, antiviral activity of polysaccharides is thought to be due to glycan-glycan interactions with the more external surface of the virus. HIV is an enveloped virus with a glycan shield known to be crucial for immune evasion and attachment to target cells (CD4+ T cells). In fact, it is highly improbable that the first contact between HIV and CD4+ T cells would occur via gp120 viral protein (Env) and cellular receptors (CD4, CXCR4 or CCR5) since viral infection could depend on glycan-to-glycan interactions [28]. Therefore, a known strategy to inhibit HIV infection intends to coat gp120 viral protein with compounds that turn the viral particle ineffective to infect new cells. This strategy is especially important for the development of microbicides, drug formulations used to prevent infections through sexual intercourse. Polysaccharide based microbicides will inactivate viral particles before the interaction with target cells would occur, in a process called neutralization.
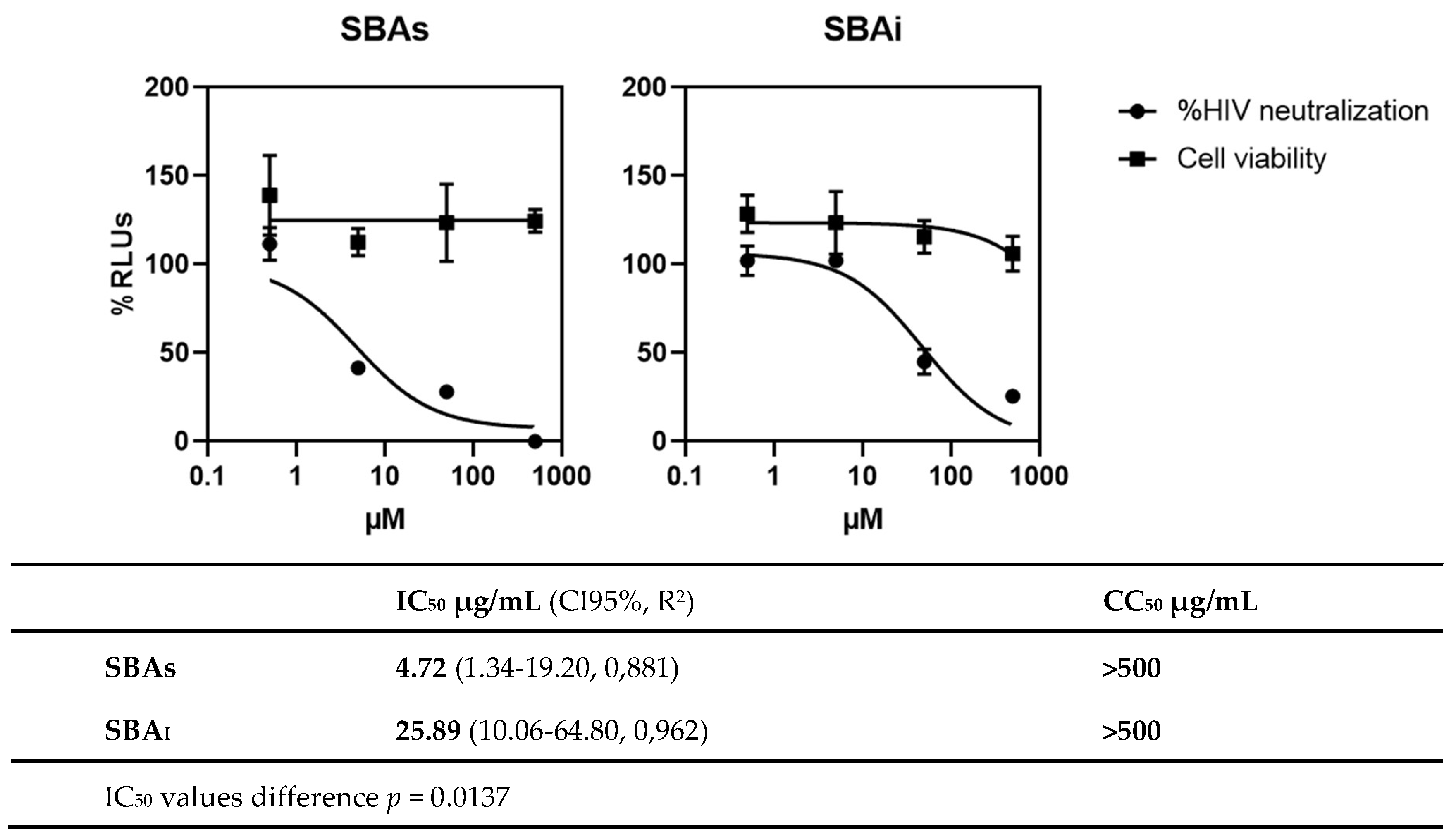
Consequently, the purified polysaccharides were tested in a neutralization assay, in which HIV supernatants were treated for 1 h with different concentrations of the compounds. These treated supernatants were then used to infect target cells and neutralization activity was measured. In figure 1, the activity of the purified SBAS and SBAI is shown. Both fractions inhibited HIV infection and showed no toxicity even at the maximum concentration tested (500 µg/mL). However, SBAs was more powerful, displaying an IC50 of 4.72 µg/mL. Thus, this fraction was selected to deepen into its chemical features.
Figure 1.
HIV-1 neutralization assay of SBAS or SBAI. Results are shown as percentage of RLUs (% of viral replication) in comparison to an untreated control (100%). IC50, CC50 and p values were calculated with GraphPad Prism 8.0 software.
Figure 1.
HIV-1 neutralization assay of SBAS or SBAI. Results are shown as percentage of RLUs (% of viral replication) in comparison to an untreated control (100%). IC50, CC50 and p values were calculated with GraphPad Prism 8.0 software.
3.Carbohydrate analysis of the fraction SBAS
To identify and quantify the main sugars in SBAs, the sample was first hydrolyzed with acid and then converted into a derivative appropriate for GC-MS analysis. Here, the monomers released were transformed into their corresponding trimethylsilyl oxymes to allow detecting uronic acids, if present.
The molar ratio of the monosaccharides identified is given in Table 1, showing a pool of neutral monosaccharides, as well as traces of galacturonic acid. Glucose is the most abundant monosaccharide, which in addition to arabinose, galactose, rhamnose and mannose are typical components of hemicelluloses and, some of them, of the carbohydrate component of arabinogalactan-proteins [30].
In order to obtain information about the configuration α or β of glycosidic bonds, as well as the presence of N-acetyl, carboxyl, phosphate or sulphate substituents, the FT-IR analysis was carried out (Figure 2). The spectrum showed bands at 3100-3600 cm-1 (stretching of hydroxyl groups of alcohols), at 800 and 1430 cm-1, due to stretching of monosaccharides (pyranoses or furanoses), and a band at 1723 cm-1, characteristic of C=O stretching. However, the lack of bands around 1017 and 1100 cm-1, characteristic of stretching vibration of the C-O single bond in carboxylic acids [31], pointed to a low abundance of uronic acids. The band detected at 892 cm-1 is a clear indication of the presence of β-linkages, and that observed at 818 cm-1 is related to α-glycosidic linkages, indicating that both anomeric forms coexist in SBAs) [30].
3.4. Methylation / GC-MS analysis
The type of glycosidic linkages of SBAS were analyzed by methylation analysis (Table 2). The results showed a high complexity in this sample (17 different units were identified). Arabinose units are all found in furanose configuration and linked at O-3, O-5, O-3,5, O-2,5 positions or as non-reducing terminal units. The rest of the detected monosaccharides are in pyranose configuration. Galactose is substituted at its O-3,6, O-6, O-3 positions and as non-reducing terminals. This suggests a (1→3)-linked Galp backbone with (1→6)-linked Galp side chains.
Dissecting this intricate pattern, the linkages deduced for galactose and arabinose units revealed branched chains and structures compatible with type-II arabinogalactans, recognized as extremely complex polysaccharides [32]. The degree of branching and the specific sugar residues present in type-II arabinogalactans can vary widely depending on the plant species, developmental stage and tissue. However, they share the backbone of β-1,3-linked-galactopyranose, decorated with side chains of β-1,6-linked galactopyranose residues. These side chains can vary in length, and the galactan core can also be substituted by short α-arabinofuranose chains (mostly 1,5 and 1,3-linked, as detected in SBAs), rhamnose, glucose, mannose, and uronic acids [33]. All these monosacharides have also been identified in SBAs. On the other hand, the (1,4)-linked glucose detected by methylation analysis probably belongs to a different polysaccharide. Starch, an α-(1,4)-glucan has been described as a component of aqueous plant extracts. However, no starch was observed under the microscope in a preliminary assay after iodine staining powdered aerial parts of C. bolivianum (data not shown). Recent publications describe these units in a complex polysaccharide purified from the infusion of aerial parts from Artemisia absinthium [34] and from another one from leaves of Penthorum chinense Pursh [35].
In summary, the analysis of SBAs fits with the presence of the polysaccharide part of arabinogalactan-proteins. It is probably accompanied by a branched (1,4)-glucan, since these structurally complex molecules are sometimes part of larger complexes with other polysaccharides, to which they can be cross-linked. Due to their structural variability, several structures have been described for type II arabinogalactans, and more structural studies are required to determine the fine structure of the one found in leaves of C. bolivianum. However, based on the linkage types deduced from methylation analysis a hypothetical simplified model of this arabinogalactan is presented in Figure 3.
3.5. Inhibition of trans infection mediated by Raji DC-SIGN cells.
HIV can infect CD4+ T cells directly, via the interaction of viral and cellular receptors, as in the neutralization assay, or might be presented to target cells by antigen presenting cells (APCs), as dendritic cells (DCs), in lymph nodes. This pathway of viral infection, called trans-infection, depends on lectin type receptors, as DC-SIGN receptors, expressed in the membrane of dendritic cells. Thus, the interference in HIV transmission in the DC–lymphocyte synapse is a potential target for the identification of new compounds with anti-HIV activity. HIV–DC interaction is mediated by the glycans of gp120 and the C-type lectin DC-SIGN (dendritic cell-specific ICAM 3-grabbing nonintegrin) expressed on DCs. DC-SIGN is tetrameric and specifically recognizes N-linked high-mannose oligosaccharides (Man9GlcNAc2) through multivalent and Ca2+-dependent protein–carbohydrate interactions [36]. Therefore, DC-SIGN receptor has been described to recognize and bind to mannose rich glycans with relative high affinity. Moreover, the inhibition of trans-infection has been described for mannan rich polysaccharides [37]. In this sense, our group have previously described the antiviral activity of new glyconanoparticles (GNPs) coated with sets of different structural motifs of the N-linked high-mannose undecasaccharide Man9(GlcNAc)2 [37]. Therefore, the potential activity of purified polysaccharides from C. bolivianum has been tested in this context, using RAJI DC-SIGN cells as APCs and primary PBMCs as target cells (Figure 4).
As shown in figure 4, SBAs did not inhibit DC-SIGN mediated trans-infection of HIV, suggesting the lectin-gp120 interaction was not blocked by this compound. The few mannose residues in the SBAs could be related to the lack of activity in this assay.
In summary, this work suggests a dose-dependent antiviral activity, where a mechanism of action is described in the phases prior to infection, that is, before virus come into contact to the cell, generating a blockade at the level of some receptor of the viral membrane.
4. Conclusions
In this study, the antiviral activity of a purified heterogeneous polysaccharide from the aerial parts of Clinopodium bolivianum, SBAS, not previously described in this species, has been evaluated. After adequate extraction and subsequent analysis (FTIR, methylation, GC-MS) its structure was partially deduced, finding a polysaccharide fraction rich in galactose, rhamnose and arabinose, but poor in mannoses and uronic acids. The mechanisms of action of polysaccharides as antivirals include their interaction with the glycoprotein gp120 via acidic interactions or the block of antigen presenting cells (APS) mediated infection through interaction of mannan rich regions with DC-SIGN receptors. Our experiments rule out the involvement of DC-SIGN and acidic interactions as mechanisms of action, due to the low mannose and uronic acid residues found in the polysaccharide structure. However, our results show that SBAS targets the viral membrane, turning the viral particles non infective and thus, displaying a powerful neutralizing activity. This activity could be a result of the interaction between the glycan shield of the viral glycoprotein gp120 and the neutral structure of SBAS, disrupting the viral particle or inactivating its infectivity by a polymer-to-polymer interaction. In summary, we have purified a heterogeneous polysaccharide from C. bolivianum, SBAs, that inhibit HIV-1 infection in vitro through a neutralizing mechanism independent of DC-SIGN receptors or acidic interactions, with could explain, at least partially, the antiviral activity of C. bolivianum. Further in vivo and clinical experimentation would be needed to explore a new potential use as topical microbicide to prevent HIV infection.
Author Contributions
Conceptualization, Paulina Bermejo, Alicia Prieto and Luis Miguel Bedoya; Data curation, Estefania Hernández, Alicia Prieto and Luis Miguel Bedoya; Formal analysis, Estefania Hernández, María José Abad and Luis Miguel Bedoya; Funding acquisition, Paulina Bermejo, José Alcami, Alicia Prieto, José Antonio Guerra and Luis Miguel Bedoya; Investigation, Estefania Hernández, María José Abad, Manuela Beltrán, Alicia Prieto and Luis Miguel Bedoya; Methodology, Paulina Bermejo, Manuela Beltrán, Alicia Prieto and Luis Miguel Bedoya; Resources, Paulina Bermejo, José Alcami, Alicia Prieto and Luis Miguel Bedoya; Software, José Antonio Guerra; Validation, Estefania Hernández, Alicia Prieto and José Antonio Guerra; Writing – original draft, Estefania Hernández, María José Abad, Alicia Prieto and Luis Miguel Bedoya; Writing – review & editing, Estefania Hernández, Paulina Bermejo, María José Abad, Manuela Beltrán, José Alcami, Alicia Prieto, José Antonio Guerra and Luis Miguel Bedoya. All authors have read and agreed to the published version of the manuscript.
Funding
This work was partially supported by Instituto de Salud Carlos III and the Infectious Diseases Networking Biomedical Research Center (CIBERINF) and co-funded by European Regional Development Fund (ERDF) "A way to build Europe" (projects AIDS Research Network RD16CIII/0002/0001 to J.A.).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
We would like to thank Dr. Eduardo Gonzales Dávalos from Universidad Mayor de San Andrés in La Paz, Bolivia, for the collection of plant specimens.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Yu, Y.; Shen, M.; Song, Q.; Xie, J. Biological activities and pharmaceutical applications of polysaccharide from natural resources: A review. Carbohyd Polym 2018, 183, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Ginsberg, H.S.; Goebel, W.F.; Horsfall, F.L. Inhibition of Mumps Virus Multiplication by a Polysaccharide. P Soc Exp Biol Med 1947, 66, 99–100. [Google Scholar] [CrossRef] [PubMed]
- Martinez, M.J.A.; Olmo, L.M.B.D.; Benito, P.B. Antiviral Activities of Polysaccharides from Natural Sources. Stud Nat Prod Chem 2005, 30, 393–418. [Google Scholar]
- Sanniyasi, E.; Venkatasubramanian, G.; Anbalagan, M.M.; Raj, P.P.; Gopal, R.K. In vitro anti-HIV-1 activity of the bioactive compound extracted and purified from two different marine macroalgae (seaweeds) (Dictyota bartayesiana J.V.Lamouroux and Turbinaria decurrens Bory). Sci Rep 2019, 9, 12185. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.H.; Niu, F.J.; Xie, Y.X.; Xie, S.M.; Liu, Y.N.; Yang, Y.Y.; Zhou, C.Z.; Wan, X.H. A review: Natural polysaccharides from medicinal plants and microorganisms and their anti-herpetic mechanism. Biomed Pharmacother 2020, 129, 110469. [Google Scholar] [CrossRef]
- Lu, W.; Yang, Z.; Chen, J.; Wang, D.; Zhang, Y. Recent advances in antiviral activities and potential mechanisms of sulfated polysaccharides. Carbohyd Polym 2021, 272, 118526. [Google Scholar] [CrossRef]
- Obiero, J.; Ogongo, P.; Mwethera, P.G.; Wiysonge, C.S. Topical microbicides for preventing sexually transmitted infections. Cochrane Database Syst Rev 2021, 3, Cd007961. [Google Scholar] [CrossRef]
- IPNI. International Plant Names Index. 2024.
- Girault, L. Kallawaya : curanderos itinerantes de los Andes : investigacion sobre practicas medicinales y magicas, 1ª ed.; UNICEF: La Paz, 1987. [Google Scholar]
- Fournet, A.; Barrios, A.A.; Muñoz, V. Leishmanicidal and trypanocidal activities of Bolivian medicinal plants. J Ethnopharmacol 1994, 41, 19–37. [Google Scholar] [CrossRef]
- Noriega, P.; Calderón, L.; Ojeda, A.; Paredes, E. Chemical Composition, Antimicrobial and Antioxidant Bioautography Activity of Essential Oil from Leaves of Amazon Plant Clinopodium brownei (Sw.). Molecules 2023, 28. [Google Scholar] [CrossRef]
- Amirova, K.M.; Dimitrova, P.; Marchev, A.S.; Aneva, I.Y.; Georgiev, M.I. Clinopodium vulgare L. (wild basil) extract and its active constituents modulate cyclooxygenase-2 expression in neutrophils. Food Chem Toxicol 2019, 124, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Fierascu, I.; Dinu-Pirvu, C.E.; Fierascu, R.C.; Velescu, B.S.; Anuta, V.; Ortan, A.; Jinga, V. Phytochemical Profile and Biological Activities of Satureja hortensis L.: A Review of the Last Decade. Molecules 2018, 23. [Google Scholar] [CrossRef] [PubMed]
- Jafari, F.; Ghavidel, F.; Zarshenas, M.M. A Critical Overview on the Pharmacological and Clinical Aspects of Popular Satureja Species. J Acupunct Meridian Stud 2016, 9, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Wang, S.; Luan, H.; Tuerhong, D.; Lin, Y.; Liang, J.; Xiong, Y.; Rui, L.; Wu, F. Clinopodium chinense Attenuates Palmitic Acid-Induced Vascular Endothelial Inflammation and Insulin Resistance through TLR4-Mediated NF- κ B and MAPK Pathways. Am J Chin Med 2019, 47, 97–117. [Google Scholar] [CrossRef]
- Tepe, B.; Cilkiz, M. A pharmacological and phytochemical overview on Satureja. Pharm Biol 2016, 54, 375–412. [Google Scholar] [CrossRef]
- Mohanty, S.; Kamolvit, W.; Zambrana, S.; Sandström, C.; Gonzales, E.; Östenson, C.G.; Brauner, A. Extract of Clinopodium bolivianum protects against E. coli invasion of uroepithelial cells. J Ethnopharmacol 2017, 198, 214–220. [Google Scholar] [CrossRef]
- Yamasaki, K.; Nakano, M.; Kawahata, T.; Mori, H.; Otake, T.; Ueda, N.; Oishi, I.; Inami, R.; Yamane, M.; Nakamura, M.; et al. Anti-HIV-1 Activity of Herbs in Labiatae. Biol Pharm Bull 1998, 21, 829–833. [Google Scholar] [CrossRef]
- Lizarraga, E.; Abdala, L.R. Compuestos fenólicos mayoritarios en Satureja boliviana (Benth.) Briq. (Lamiaceae). Acta Farm Bon 2004, 23, 198–200. [Google Scholar]
- Velasco-Negueruela, A.; Abarca, G.E.; Pérez-Alonso, M.J.; Esteban, J.L. Essential Oil of Satureja boliviana Briq. from Peru. J Essent Oil Res 1994, 6, 641–642. [Google Scholar] [CrossRef]
- Apaza Ticona, L.; Martínez Noguerón, A.; Sánchez Sánchez-Corral, J.; Montoto Lozano, N.; Ortega Domenech, M. Anti-Inflammatory, Antibacterial, Anti-Biofilm, and Anti-Quorum Sensing Activities of the Diterpenes Isolated from Clinopodium bolivianum. Pharmaceutics 2024, 16. [Google Scholar] [CrossRef]
- Abad, M.J.; Bermejo, P.; Gonzales, E.; Iglesias, I.; Irurzun, A.; Carrasco, L. Antiviral activity of Bolivian plant extracts. Gen Pharmacol 1999, 32, 499–503. [Google Scholar] [CrossRef] [PubMed]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric Method for Determination of Sugars and Related Substances. Anal Chem 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Rojas-Escudero, E.; Alarcón-Jiménez, A.L.; Elizalde-Galván, P.; Rojo-Callejas, F. Optimization of carbohydrate silylation for gas chromatography. J Chromatogr A 2004, 1027, 117–120. [Google Scholar] [CrossRef]
- Ciucanu, I.; Kerek, F. A simple and rapid method for the permethylation of carbohydrates. Carbohyd Res 1984, 131, 209–217. [Google Scholar] [CrossRef]
- Needs, P.W.; Selvendran, R.R. Avoiding oxidative degradation during sodium hydroxide/methyl iodide-mediated carbohydrate methylation in dimethyl sulfoxide. Carbohyd Res 1993, 245, 1–10. [Google Scholar] [CrossRef]
- Hollingsworth, R.I.; Abe, M.; Sherwood, J.E.; Dazzo, F.B. Bacteriophage-induced acidic heteropolysaccharide lyases that convert the acidic heteropolysaccharides of Rhizobium trifolii into oligosaccharide units. J Bacteriol 1984, 160, 510–516. [Google Scholar] [CrossRef]
- Garcia-Perez, J.; Sanchez-Palomino, S.; Perez-Olmeda, M.; Fernandez, B.; Alcami, J. A new strategy based on recombinant viruses as a tool for assessing drug susceptibility of human immunodeficiency virus type 1. J Med Virol 2007, 79, 127–137. [Google Scholar] [CrossRef]
- Martínez-Avila, O.; Bedoya, L.M.; Marradi, M.; Clavel, C.; Alcamí, J.; Penadés, S. Multivalent manno-glyconanoparticles inhibit DC-SIGN-mediated HIV-1 trans-infection of human T cells. Chembiochem 2009, 10, 1806–1809. [Google Scholar] [CrossRef]
- Ghosh, K.; Takahashi, D.; Kotake, T. Plant type II arabinogalactan: Structural features and modification to increase functionality. Carbohyd Res 2023, 529, 108828. [Google Scholar] [CrossRef]
- Robert, P.; Marquis, M.; Barron, C.; Guillon, F.; Saulnier, L. FT-IR investigation of cell wall polysaccharides from cereal grains. Arabinoxylan infrared assignment. J Agric Food Chem 2005, 53, 7014–7018. [Google Scholar] [CrossRef]
- Kikuchi, A.; Hara, K.; Yoshimi, Y.; Soga, K.; Takahashi, D.; Kotake, T. In vivo structural modification of type II arabinogalactans with fungal endo-β-1, 6-galactanase in Arabidopsis. Front Plant Sci 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Saeidy, S.; Petera, B.; Pierre, G.; Fenoradosoa, T.A.; Djomdi, D.; Michaud, P.; Delattre, C. Plants arabinogalactans: From structures to physico-chemical and biological properties. Biotechnol Adv 2021, 53, 107771. [Google Scholar] [CrossRef] [PubMed]
- Corrêa-Ferreira, M.L.; Noleto, G.R.; Oliveira Petkowicz, C.L. Artemisia absinthium and Artemisia vulgaris: A comparative study of infusion polysaccharides. Carbohyd Polym 2014, 102, 738–745. [Google Scholar] [CrossRef]
- Feng, S.; Xu, X.; Li, X.; Deng, G.; Xia, C.; Zhou, L.; Chen, T.; Gao, T.; Yuan, M.; Cao, X.; Ding, C. Structure elucidation and antioxidant activity of a polysaccharide from Penthorum chinense Pursh. Int J Biol Macromol 2024, 264, 130720. [Google Scholar] [CrossRef]
- Feinberg, H.; Mitchell, D.A.; Drickamer, K.; Weis, W.I. Structural basis for selective recognition of oligosaccharides by DC-SIGN and DC-SIGNR. Science 2001, 294, 2163–2166. [Google Scholar] [CrossRef]
- McReynolds, K.D.; Gervay-Hague, J. Chemotherapeutic interventions targeting HIV interactions with host-associated carbohydrates. Chem Rev 2007, 107, 1533–1552. [Google Scholar] [CrossRef]
Figure 2.
Fourier transform-infrared (FT-IR) spectrometry of SBAS.
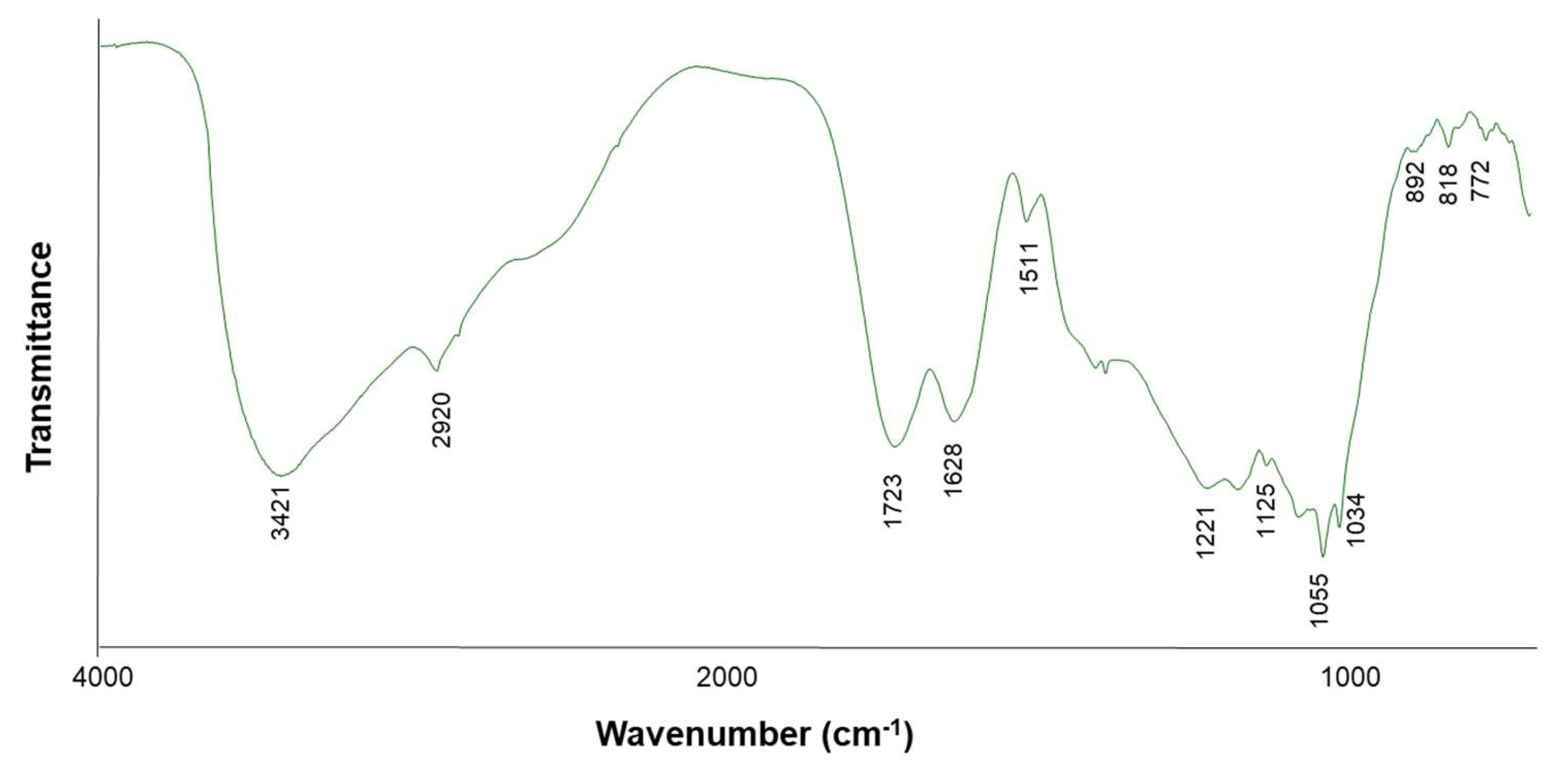
Figure 3.
Figure 3. Simplified model of the possible structure of SBAS arabinogalactan, based on linkage analysis.
Figure 3.
Figure 3. Simplified model of the possible structure of SBAS arabinogalactan, based on linkage analysis.
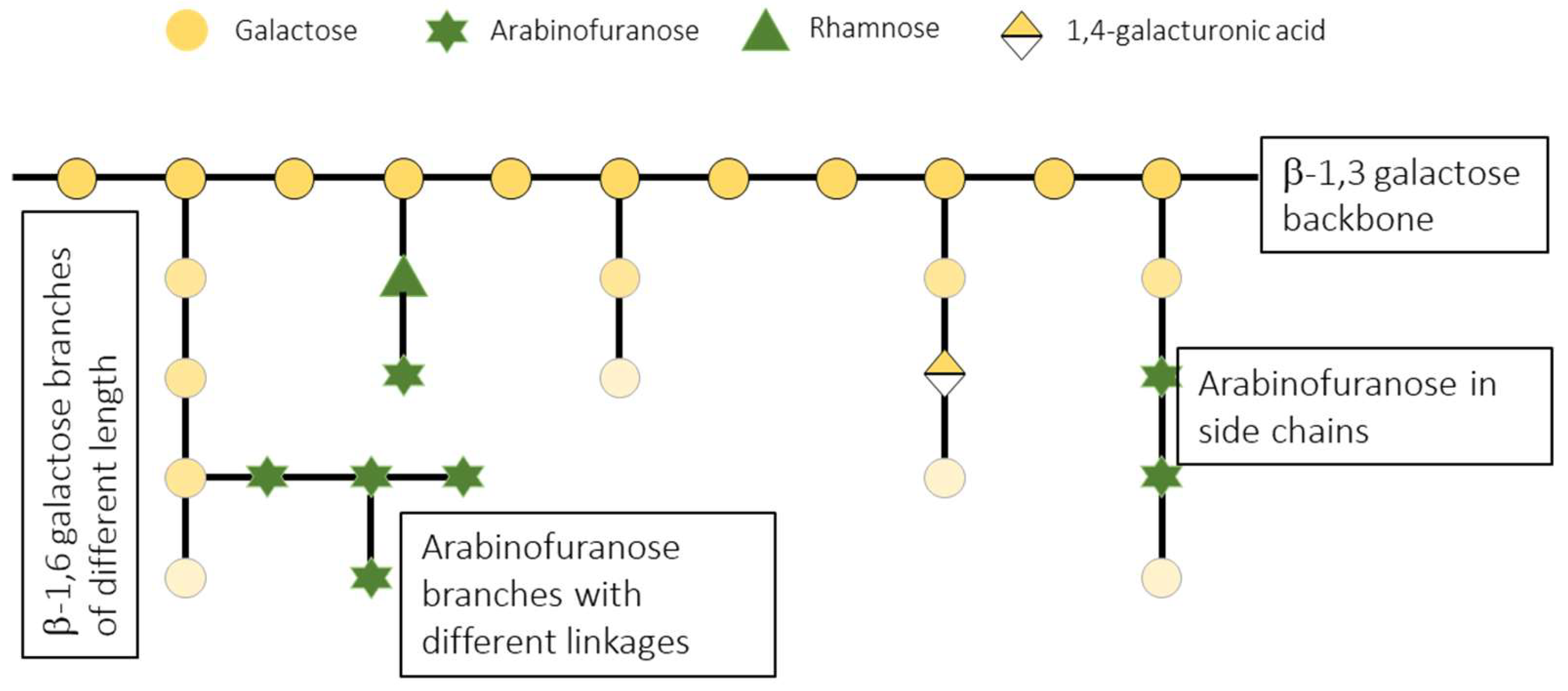
Figure 4.
Figure 4. Trans-infection assay. Results are shown as percentage of RLUs in comparison to an untreated control (100%). ANOVA analysis (Turkey’s multiple comparison test) was performed to calculate the values of p.
Figure 4.
Figure 4. Trans-infection assay. Results are shown as percentage of RLUs in comparison to an untreated control (100%). ANOVA analysis (Turkey’s multiple comparison test) was performed to calculate the values of p.
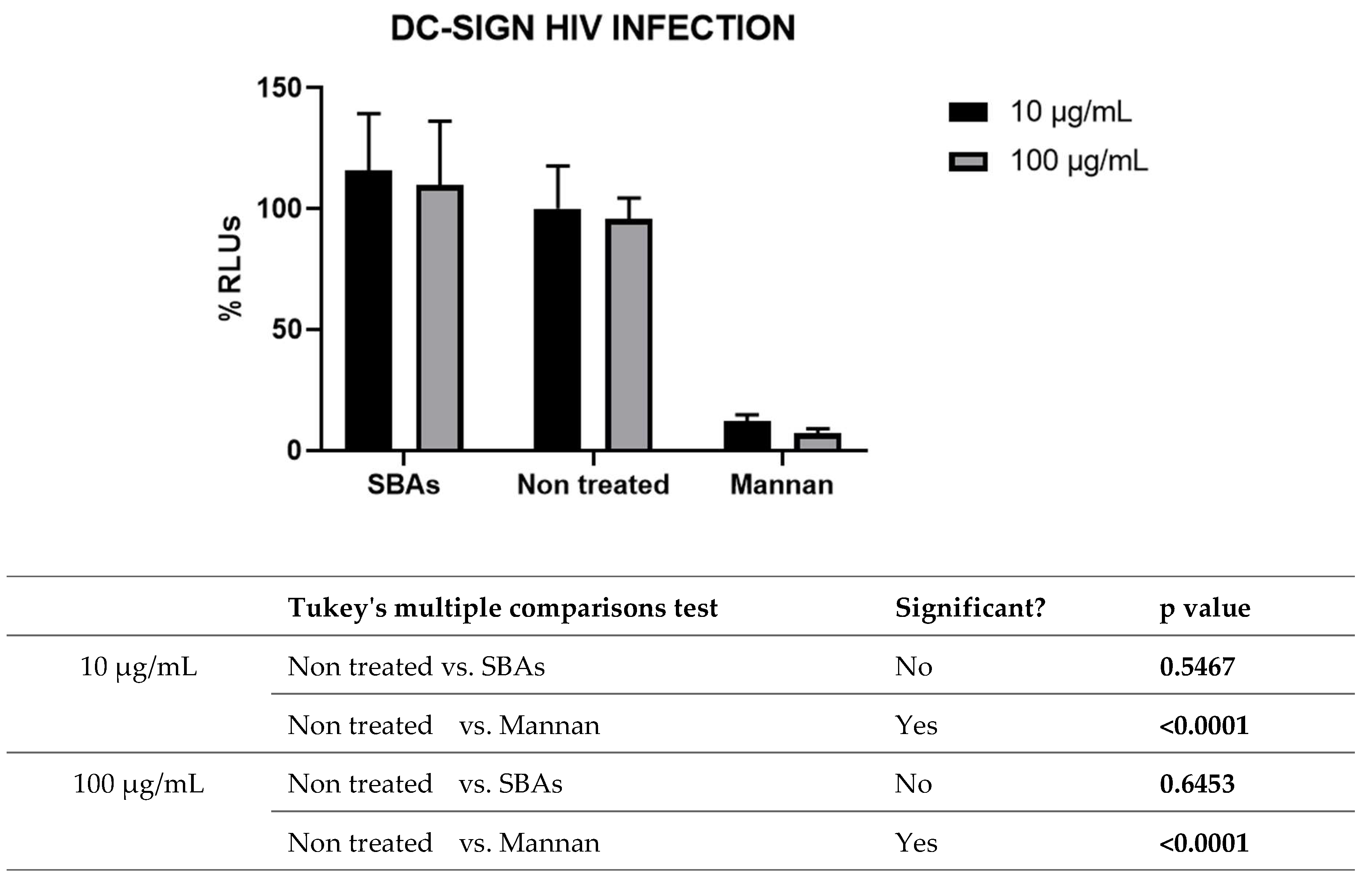

Table 1.
Monosaccharide composition of SBAS.
| Monosaccharide | Molar ratio |
|---|---|
| Rhamnose | 0.4 |
| Arabinose | 0.6 |
| Mannose | 0.1 |
| Glucose | 1.0 |
| Galactose | 0.6 |
| Galacturonic acid | tr. |
Table 2.
Linkage types deduced from methylation analysis of SBAS.
| Linkage | Rt1 (min) |
Major mass fragments (m/z) |
Molar ratio2 % |
|---|---|---|---|
| Araf-(1→ | 4.0 | 87, 101, 118, 129, 161 | 5.7 |
| →3)-Araf-(1→ | 5.2 | 118, 173, 233 | 6.5 |
| →5)-Araf-(1→ | 5.7 | 87,88,102,118,129,130,162,173,189 | 7.9 |
| →3,5)-Araf-(1→ | 7.2 | 85, 118, 127, 159, 201, 261 | 3.8 |
| →2,5)-Araf-(1→ | 7.5 | 87, 88, 129, 130, 190 | 4.0 |
| Galp-(1→ | 7.0 | 88, 101, 102, 118, 129, 130, 205, 206 | 5.9 |
| →4)-Galp-(1→ | 8.5 | 88, 102, 113, 118, 130, 162, 173, 233 | 4.5 |
| →3)-Galp-(1→ | 8.9 | 87, 101, 118, 129, 161, 174, 234, 277 | 4.0 |
| →6)-Galp-(1→ | 9.9 | 88, 102, 118, 129, 130, 162, 189, 233 | 8.5 |
| →3,6)-Galp-(1→ | 12.5 | 87, 118, 129, 174, 189, 234, 305 | 14.4 |
| Glcp-(1→ | 6.5 | 88, 101, 102, 118, 129, 130, 205, 206 | 3.6 |
| →4)-Glcp-(1→ | 8.7 | 88, 102, 113, 118, 130, 162, 173, 233 | 19.3 |
| →4,6)-Glcp-(1→ | 11.3 | 87, 102, 118, 127, 129, 201, 261 | 4.5 |
| →4)-GalAcp-(1→ | 11.5 | 87, 102, 118, 127, 129, 203, 263 | 3.6 |
| →4)-Rhap-(1→ | 4.3 | 88, 101, 102, 131, 162 | 1.9 |
| →2)-Manp-(1→ | 8.3 | 87, 88, 101, 129, 130, 161, 190, 205 | 1.0 |
| →4,6)-Manp-(1→ | 8.3 | 102, 118, 162, 201, 261 | 0.9 |
| 1 Retention time. 2 Relative molar ratios, calculated from peak areas. | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



