
Submitted:
27 January 2025
Posted:
28 January 2025
You are already at the latest version
Abstract
The presence of cadmium (Cd) in agricultural soils poses a serious risk to crop growth and food safety. Cd uptake and transport in plants occur through various transporters of nutrient ions which have the similar physical and chemical properties to Cd, indicating genetic manipulation of these transporters and agronomic improvement of the Cd-antagonistic nutrients could be a good approach for reducing Cd uptake and accumulation in crops. In this review, we discussed the interaction of Cd and some micronutrients, including zinc (Zn) and manganese (Mn), focusing on their influence on the expression of genes encoding Cd related transporters, including ZIP7, NRAMP3 and NRAMP4. Genetic improvement in enhancing the specificity and efficiency of transporters, and agronomic improvement in optimizing micronutrient nutrition can inhibit Cd uptake and transporters by these transporters. This comprehensive review provides a deep insight into genetic and agronomic improvement for fighting against Cd contamination and enhancing sustainable agricultural production.
Keywords:
Cadmium
; Genetic improvement
; Ion interaction
; Manganese
; Transpoter
; Zinc
1. Introduction
Cadmium (Cd) is an unessential metal with no biological function and toxic to plants even at low concentrations (Haider et al. 2021). Many physiological dysfunctions will occur when plants are exposed to Cd stress, mainly leading to oxidative stress (Moradi et al. 2019). Oxidative stress is injurious and even lethal to plants, as it inhibits or disturbs physiological activities by altering membrane integrity and permeability at the organelle and molecular levels. Membrane damage will occur due to excessive generation of reactive oxygen species (ROS), such as superoxide anion (O2−), hydroxyl radical (OH−), and hydrogen peroxide (H2O2) under Cd stress (Anjum et al. 2015). Moreover, Cd contamination in agricultural soils bring a huge threat to human health via a food chain, because Cd can induce several illnesses, including cancer, heart disease, vascular issues, kidney and liver damage, and disruptions to male reproductive system (Kumar and Sharma 2019) (Figure 1).
It is well documented that Cd uptake by roots from soils and transportation from roots to shoots (above ground organs) are dependent on the transporters of some nutrient ions that have the similar chemical and physical properties as Cd, such as Zn and Mn (Huang et al. 2020b). There is a dramatic difference in Cd accumulation among crop species, and genotypes within a species (Lin et al. 2022), which provides the possibility for developing crop cultivars with low Cd accumulation through genetic improvement or gene engineering. The physiological and molecular mechanisms of Cd uptake and transportation in plants have been intensively assessed to explain the distinct differences in Cd accumulation found across various species or genotypes. Cd and several micro-elements, including Mn, Zn, Cu, and Fe, interact antagonistically and synergistically in their plant uptake and transportation (Figure 1). Understanding the relationship between Cd and nutrient ions may offer evidence to explain the nature of Cd accumulation in crops (Mapodzeke et al. 2021).
Cd contamination has become a serious factor affecting sustainable crop production and human health via food chain. Hence, it is necessary to make major efforts to control Cd contamination in soil by cutting off entrance of Cd into agricultural ecosystems, phytoremediation of Cd-contaminated soil and reducing Cd bio-availability in soil. Meanwhile it is more important to develop the crop cultivars with high tolerance and low accumulation of Cd. In the past 20 years, numerous studies have examined the physiological and molecular mechanisms of Cd accumulation and detoxification in plants using genomics, transcriptomics, proteomics, and metabolomics, with an emphasis on identifying the genes responsible for Cd uptake, translocation, sequestration, and tolerance in plant tissues (Sterckeman and Thomine 2020). On the other hand, the plants with high tolerance and Cd accumulation can be used in phytoremediation of Cd contaminated soil. Naturally, certain plant species can accumulate a significant amount of Cd without experiencing any toxicity. It is well known that these plants have special tolerance mechanisms, such as compartmentalization, redox homeostasis maintenance, reducing rhizosphere Cd activity and Cd transfer to aboveground plant parts (Shiyu et al. 2020).
This review explores the complex interactions between cadmium (Cd) and essential micronutrients in plants, focusing on Cd uptake, transport, and detoxification mechanisms. By highlighting the roles of key transporters like OsZIP7, ATPase2, AtNRAMP3, and AtNRAMP4, the review provides insights into how Cd interacts antagonistically or synergistically with nutrients such as zinc (Zn), manganese (Mn), copper (Cu), and iron (Fe). Additionally, it examines genetic and agronomic strategies for reducing Cd accumulation in crops, emphasizing the potential for molecular and biochemical interventions to enhance crop tolerance to Cd toxicity.
2. Cd Uptake and Transport in Plants
2.1. Mechanisms of Cd Uptake by Roots
Cadmium (Cd) in soil is generally insoluble and not easily absorbed by plants. However, its bioavailability significantly increases when soil pH decreases, meaning that soil acidification can enhance Cd contamination. Plants can affect the bioavailability of cadmium (Cd) by releasing root exudates that alter the rhizosphere’s pH, which can strengthen Cd uptake. In addition, passive diffusion occurs for Cd entering plants through the apoplastic pathway, while the symplastic pathway, an active transport process, relies on electrochemical potential gradients and concentration differences across the plasma membrane (Bali et al. 2020, Thakur et al. 2016). For Cd absorbed by plant roots, it must be available for uptake, which is contingent upon the species of the plants, the physicochemical conditions of the soil, and the speciation of the metals (Rizwan et al. 2018). This metal is readily taken up by and delivered to the aerial portions of the plants (Shanmugaraj et al. 2019). The transfer of cadmium (Cd) from soil to grains is a complex and multi-stage process involving sequential mechanisms: initial uptake by roots, sequestration within root vacuoles to mitigate toxicity, subsequent translocation through the vascular system to aerial parts, and ultimate partitioning and accumulation in grains (Huang et al. 2020a). (Figure 2).
2.2. Role of Metal Transporters
In plants, metal transporters such as ZIP, NRAMP, and HMA families play pivotal roles in the uptake of essential micronutrients. However, these transporters can also facilitate the entry of non-essential and potentially toxic metals like cadmium (Cd) into root cells (Li et al. 2022). For instance, members of the ZIP family, including OsIRT1 and OsIRT2 in rice, are primarily responsible for iron and zinc uptake, but can inadvertently transport Cd due to its chemical similarity to these micronutrients. Similarly, NRAMP transporters, such as TcNRAMP3, are involved in the uptake of divalent cations like Fe²⁺ and Mn²⁺, yet they also permit Cd²⁺ entry into plant roots (Takahashi et al. 2011). The HMA family, particularly transporters like HMA2 and HMA4, is crucial for translocating essential metals; however, they can also contribute to Cd movement within the plants. This dual functionality underscores a significant challenge in plant nutrition and heavy metal detoxification, as the mechanisms that enable the acquisition of vital nutrients can simultaneously increase Cd accumulation, posing risks to plant health and food safety (Wong and Cobbett 2009).
2.3. Transporter Families Facilitating Cd Uptake and Transport
The transport of cadmium in rice (OsNramp5) and barley (HvNramp5) is mediated by Nramp5 homologs, which are also present in wheat (TaNramp5A, TaNramp5D) and maize (ZmNramp5). Nramp5, a Natural Resistance-associated Macrophage Protein family member, transports manganese and cadmium in plants (Palali Delen et al. 2024, Sui et al. 2018, Wu et al. 2016). OsNramp5 is localized at the distal edges of the exodermis in rice roots, facilitating Cd uptake, which is then either sequestered in vacuoles by OsHMA3 or transported to the shoots via the xylem. Cd competes with other divalent cations (Ca²⁺, Fe²⁺, Mg²⁺, Cu²⁺, Zn²⁺) for transport through root cell membranes, and its absorption varies significantly across plant species and genotypes due to morphological and physiological differences (Feng et al. 2021, Gupta et al. 2019, Ismael et al. 2019, Satoh-Nagasawa et al. 2012). Cd can enter plant roots from the soil solution through cell walls via passive transport. Additionally, active transport mechanisms involve nonspecific membrane transport proteins, such as iron transporters (IRT), zinc transporters (ZIP), and metal-pumping ATPases, which facilitate Cd movement across the plasma membrane of root cells. Other transporter families, including NRAMP, P-type ATPase, ABC transporters, CAX, LCT, and CE, have also been associated with Cd translocation within plants (Kim et al. 2015, Song et al. 2017). ZNT1 may be involved in the transport of Cd in the low Cd accumulation ecotype. At the same time, a high-affinity Cd transporter may also play a role in the transportation of Cd in the high accumulation ecotype.
2.4. Factors Influencing Cadmium Uptake in Plants
Cadmium (Cd) uptake in plants is intricately influenced by micronutrient nutrition, interactions with other metals, and the genetic and molecular machinery of plants. Micronutrients such as zinc (Zn) and iron (Fe) play pivotal roles in modulating Cd absorption, as they compete for transporters like ZIP proteins, often reducing Cd uptake when present in optimal concentrations (Rai et al. 2021) (Figure 2). Additionally, interactions with other metals, such as calcium (Ca) and magnesium (Mg), can alter membrane permeability and ion exchange, further influencing Cd transport (Rahman et al. 2016). Plant-specific genetic factors, including the diversity of metal transporter families like NRAMPs and HMA, regulate Cd uptake and sequestration in vacuoles or cell walls (Yadav et al. 2021). At the molecular level, transcription factors such as bZIP and MYB are critical in modulating gene expression for metal homeostasis, leading to variability in Cd accumulation among species and cultivars (Li et al. 2023b). Understanding this genetic and molecular diversity is crucial for breeding or engineering plants with reduced Cd uptake, ensuring safer agricultural production and mitigating heavy metal contamination in crops.
3. Genetic Regulation and Micronutrient-Cadmium Interaction
3.1. Genetic Regulation of Cd Uptake
Rice varieties display significantly genetic heterogeneity in Cd accumulation, offering a valuable resource for identifying functional alleles in improving Cd tolerance (Mei et al. 2022). Cd enters vacuoles for sequestration by a Cd transporter that is encoded by OsHMA3 (Ueno et al. 2010). The molecular mechanisms of toxic metal Cd in its transport and accumulation in plant tissues have been extensively studied in recent years, and appreciation of the massive amount of data has been offered by bio-informatic techniques, which have been applied extensively in a variety of plant species (Xu et al. 2020). These studies have shown the five groups into which the differentially expressed genes can be categorized, i.e., transporters, organic acids, metabolic pathways, phytohormones, and ROS generation (Angulo-Bejarano et al. 2021). Transcriptome sequencing revealed that higher Cd accumulation in Nicotiana tabacum leaves is driven by coordinated mechanisms, including reduced cell wall binding, weakened Casparian strip barriers, and enhanced xylem loading. Similarly, in barley, genes regulating ion transport, stress response, cell wall dynamics, and reactive oxygen species metabolism play crucial roles in Cd transport and tolerance (Huang et al. 2021c). Phytometallophores facilitate the transport of metals like Cd by binding to them during their passage through root cells, enabling efficient metal translocation (Singh et al. 2020).
3.2. Micronutrient-Cadmium Interaction in Their Uptake and Transport
Cadmium (Cd) enters plant cells via transporters typically involved in the uptake of essential micronutrients (Song et al. 2017). The uptake of Cd in plants is influenced by several essential elements such as calcium (Ca), copper (Cu), iron (Fe), zinc (Zn), and manganese (Mn) in the rhizosphere solution (Vítková et al. 2018). Cd poisoning often manifests as leaf chlorosis, resembling iron deficiency, as Cd affects iron accumulation essential for chlorophyll synthesis. It is showed that Cd stress reduces Fe uptake and its movement from roots to shoots. In Arabidopsis, increased iron availability reduces Cd uptake, whereas in peanuts, an iron deficit enhances Cd accumulation (He et al. 2017). The competitive relationship between Fe and Cd is further evident when phosphate or sulfate deprivation reduces Cd uptake by boosting Fe uptake (Yang et al. 2016). Moreover, different chelated forms of Fe, such as Fe (III) citrate, result in higher Cd accumulation than Fe (III) ethylenediaminetetraacetic acid (EDTA) (Gul et al. 2021). Mn has been found to reduce Cd uptake, as the two metals share similar absorption and transport pathways. Mn competes with Cd for uptake, and under Mn-replete conditions, plants exhibit increased antioxidant enzyme activity, high leaf Mn content, and improved photosynthetic efficiency, leading to alleviation of Cd toxicity, oxidative stress, and lipid peroxidation, likely due to Mn’s role as a cofactor in Mn-CAT and Mn-SOD enzymes (Leitenmaier and Küpper 2013, Sterckeman and Thomine 2020, Wang et al. 2018a). Zinc (Zn) and Cd interact in several ways due to their similar atomic structures, which complicates efforts to selectively reduce Cd uptake while maintaining Zn transport. Zn inhibits Cd uptake through shared transporters on the root plasma membrane, and it can mitigate oxidative stress and enhance plant growth (Kinnell 2019). Some studies suggest genotype-specific differences in Cd translocation, and Zn may help reduce Cd bioaccumulation in wheat (Zhou et al. 2020). Additionally, Zn Transporter7 (OsZIP7) and heavy metal ATPase2 (OsHMA2) are involved in the xylem loading of Zn and Cu in roots (Tan et al. 2019) (Figure 2).
Taken together, the interactions between essential nutrients and Cd uptake highlight the complexity of managing Cd toxicity in plants. Optimizing the balance of these elements can help mitigate Cd’s harmful effects, especially in crops like rice, wheat, and peanuts. Table 1 and Table 2 summarize with details of genes and their corresponding functions with identified crops.
4. Approaches for Alleviating Cd Toxicity and Accumulation in Crops
Cd toxicity and accumulation in crops are mainly controlled by genetic factors, but also heavily influenced by environmental (soil) conditions and agronomic practices. Hence, approaches of alleviating Cd toxicity and accumulation in crops are multifaceted, involving in genetic improvement for enhancing tolerance and reducing uptake and transport of Cd, soil remediation for reducing Cd content or bio-availability, and improvement of agronomic managements, including irrigation and fertilization for reducing Cd uptake.
4.1. Phytoremediation
An effective strategy for reducing Cd content in soil is bioremediation, a process that leverages the natural capabilities of plants, animals, and microorganisms to restore contaminated environments. Nowadays, phytoremediation of the contaminated soils by heavy metals, including Cd has been particularly highlighted and widely used because of its multiple advantages, such as low cost, no secondary contamination, and environmental friendly (Karami and Shamsuddin 2010, Raza et al. 2020). The application of Cd phytoremediation is mainly dependent on the availability of Cd hyperaccumulators, the plants with high Cd tolerance and root-to-shoot translocation of Cd. For instance, Noccaea caerulescens, is an extremophile heavy metal hyperaccumulator with a high capacity of Cd and Zn accumulation in shoots. It may be able to effectively phytoextract Cd from soils contaminated with Cd (Li et al. 2021). One factor contributing to Cd tolerance is the increased capacity of rhizosphere microorganisms to generate organic acid to chelate Cd2+ (Hou et al. 2018).
The effectiveness of Cd phytoremediation is significantly influenced by the balance and availability of essential soil micronutrients. Elements like Fe, Zn, and Mn affect Cd absorption by competing for uptake sites or modifying the bioavailability of metals (Li et al. 2023b, Raza et al. 2020). Factors such as soil pH, organic matter content, and microbial interactions further regulate the accessibility of these nutrients and Cd. Plants capable of efficiently absorbing and utilizing micronutrients tend to demonstrate higher tolerance and accumulation of Cd. Enhancing the levels and interactions of soil micronutrients is key to improving the success of phytoremediation efforts (Bali et al. 2020, Shahid et al. 2017).
4.2. Genetic Improvement
Genetic improvement represents a promising approach for mitigating cadmium (Cd) stress and toxicity in crop plants, addressing this critical issue from multiple angles. Advances in plant genetics allow for the development of cultivars with enhanced tolerance to Cd, achieved through various strategies such as the manipulation of Cd uptake and translocation pathways, increased sequestration and detoxification mechanisms, and enhanced repair and stress response systems. Key genetic modifications include the alteration of genes involved in Cd transport and binding, as well as the integration of traits that boost the plant’s ability to handle and detoxify heavy metals. Such improvements not only reduce Cd accumulation in edible plant parts but also enhance overall plant health and yield under contaminated conditions. These genetic strategies are crucial for developing crops that can thrive in Cd-contaminated soils, ensuring food safety and sustainability in agriculture (Li et al. 2017, Zhang et al. 2022).
In view of the fact that Cd enters roots and transports from roots to shoots via the transporters of some nutrient ions which have the similar physical and chemical properties with Cd, regulating expression of the genes encoding these ion transporters might be efficient to control Cd uptake and transport. Zinc (Zn), a crucial microelement for various plant enzymes, competes with Cd for binding sites on root surfaces and in soil, influencing Cd uptake in plants. Zinc-specific transporters, such as ZIP (zinc/iron-regulated transporter-like proteins), may inadvertently facilitate Cd entry into root cells and its subsequent redistribution throughout the plant. These Zn transporters, along with others from the IRT1, HMA2, HMA3, ZIP, and NRAMP families, play a key role in the uptake and translocation of Zn, Cd, and other ions (Zare et al. 2018). Modulating metal transporters provides an effective strategy to reduce Cd accumulation while preserving essential nutrient uptake. Silencing IRT1, which facilitates both Fe and Cd uptake, can limit Cd entry, but requires careful regulation to avoid disrupting Fe homeostasis (Ghorbani et al. 2024, Loix et al. 2017, Seth 2012). Overexpression of HMA3 enhances Cd sequestration into vacuoles, reducing its translocation to edible plant parts. These approaches balance nutrient acquisition with minimized Cd accumulation, offering potential for safer crop development and improved phytoremediation (Dendena 2012, Upadhyaya et al. 2023, Zhu et al. 2023).
There are multiple steps, from soil to grain, and a number of transporters involved. Thus, it is thought that one efficient way to lessen the amount of Cd that crops absorb is to breed low-grain cultivars that accumulate less Cd. This can be achieved through the use of transgenic technology. The main advantage of transgenic technology is the ability to efficiently express target genes in plants, giving them the correct genetic and biological traits, all without affecting crop quality or yield. For instance, low-Cd rice that accumulates less Cd can be produced without yield penalty by utilizing the CRISPR/Cas9 system to delete OsNramp5 (Songmei et al. 2019).
4.3. Agronomic Practices
A promising and cost-effective method to prevent cadmium (Cd) contamination in food involves using plant nutrients to mitigate Cd toxicity in crops (Nazar et al. 2012). Plants require a balanced supply of essential nutrients at optimal levels and timings to thrive and reduce Cd’s adverse effects. Farmers often enhance soil fertility to boost crop yields, but effective management of these nutrients is crucial for minimizing soil Cd toxicity. Understanding the interplay between essential plant nutrients and soil Cd is key to achieving this (Shiyu et al. 2020). Essential soil nutrients influence Cd availability and toxicity through both direct and indirect mechanisms (Zhang et al. 2019). These mechanisms include Cd sequestration in plant tissues, adsorption and precipitation in the soil, competition for membrane transporters, and prevention of Cd accumulation in grains and fruits, all of which work together to reduce Cd solubility and its impact on plant health (Dharma-Wardana 2018). The application of N and other nutrients can acidify the soil, which increases the solubility of Cd (Ning et al. 2017). Similarly, application of P fertilizer may have an impact on Cd solubility in soil (Noh et al. 2017). Fertilizers also determine the speciation of Cd, affecting Cd transport to roots and rhizosphere incorporation (Qaswar et al. 2017). The rhizosphere’s nutritional composition, root development, and overall plant growth are all impacted by fertilizer application (Wang et al. 2018b). Selenium (Se) applied at low concentrations to contaminated soil helps reduce Cd stress and promotes healthy plant growth (Gao et al. 2018).
Manganese (Mn) supplementation has been shown to mitigate cadmium (Cd) toxicity in plants by enhancing nutrient homeostasis and reducing Cd uptake. In maize seedlings, exposure to Mn alleviated Cd-induced root growth inhibition, with the degree of improvement correlating with Mn concentration (Choppala et al. 2014, Li et al. 2023a). Similarly, in rice seedlings, Mn application reduced Cd uptake and translocation, thereby improving growth and chlorophyll content under Cd stress. These findings suggest that Mn supplementation can enhance plant tolerance to Cd stress by modulating nutrient uptake and distribution (Huang et al. 2021a, Hussain et al. 2020, Shahzad et al. 2024).
Silicon (Si) also impacts Cd accumulation by modifying gene expression related to Cd uptake and sequestration. Si treatment is associated with the downregulation of Cd transporters, such as OsNRAMP5 and OsHMA2, and enhances the phytochelatin-driven compartmentalization of Cd in the vacuoles of rice roots. This interplay between Si and Cd management underscores Si’s role in mitigating Cd toxicity through complex biochemical and genetic mechanisms in agricultural practices(Bari et al. 2020). (Figure 3)
The chemical approach to reducing cadmium (Cd) uptake and transport in crops involves the application of foliar sprays or direct soil amendments, which utilize chelation or competition to inhibit Cd absorption. Silicon (Si) and selenium (Se) play a synergistic role in mitigating Cd toxicity by regulating gene expression, immobilizing Cd within root cell walls and organelles, and reducing its translocation to shoots (Huang et al. 2021b). The use of silicon-rich biochar or Si-fertilizers shows strong potential in further decreasing Cd uptake and movement within plants. Additionally, various other compounds and hormones have been identified as effective in minimizing Cd accumulation, offering a multifaceted strategy to combat Cd contamination in crops (Kapoor et al. 2023).
Applying micronutrient fertilizers, particularly those containing Zn and Mn, can effectively mitigate Cd uptake and accumulation in plants (Lv et al. 2019, Zhou et al. 2020). Foliar application of Mn and Zn has been shown to increase their concentrations in plant tissues, thereby enhancing nutrient content and potentially reducing Cd translocation to edible parts. Additionally, the interaction between Cd and Zn in the soil can influence their uptake and translocation within plants (Gupta et al. 2019, Riaz et al. 2021). By carefully balancing the application of these micronutrients, it is possible to enhance plant growth and reduce Cd accumulation, thereby improving food safety and crop quality.
5. Conclusions and Prospects
Cadmium (Cd) contamination poses a significant threat to crop productivity and food safety. While substantial progress has been made in understanding Cd absorption, transport, and resistance mechanisms in rice, critical gaps remain, particularly in the molecular regulation of vacuolar compartmentalization and cell wall sequestration of Cd. Future studies should focus on elucidating metabolic pathways altered under Cd stress and their interaction with plant nutrition.
Developing crop varieties with reduced Cd accumulation and enhanced tolerance, such as through the manipulation of transporters like OsNramp5 and OsHMA3, is a promising strategy. However, the functional roles of these transporters remain largely unexplored in crops other than rice and Arabidopsis. Bioremediation techniques, including phytoremediation, microbial remediation, and the use of biochar and organic amendments, also offer sustainable approaches for mitigating Cd toxicity in agricultural soils.
Emerging challenges, such as the impact of climate change on soil organic matter breakdown and Cd bioavailability, warrant further investigation. Additionally, identifying novel genes and understanding their roles in Cd uptake, transport, and detoxification will be critical for advancing sustainable agricultural practices and ensuring food safety in Cd-contaminated areas. Comprehensive research and innovative remediation methods are essential to mitigate Cd toxicity and improve crop yields in the face of environmental challenges.
Acknowledgments
This study was supported by the Key Research Foundation of the Science and Technology Department of Zhejiang Province of China (2021C02064-3) and the Jiangsu Collaborative Innovation Center for Modern Crop Production (JCIC-MCP).
References
- Angulo-Bejarano P I, Puente-Rivera J, Cruz-Ortega R. 2021. Metal and metalloid toxicity in plants: An overview on molecular aspects. Plants, 10, 635.
- Anjum S A, Tanveer M, Hussain S, Bao M, Wang L, Khan I, Ullah E, Tung S A, Samad R A, Shahzad B. 2015. Cadmium toxicity in Maize (Zea mays L.): consequences on antioxidative systems, reactive oxygen species and cadmium accumulation. Environmental Science and Pollution Research, 22, 17022-17030.
- Bali A S, Sidhu G P S, Kumar V. 2020. Root exudates ameliorate cadmium tolerance in plants: a review. Environmental Chemistry Letters, 18, 1243-1275.
- Bari M A, Prity S A, Das U, Akther M S, Sajib S A, Reza M A, Kabir A H. 2020. Silicon induces phytochelatin and ROS scavengers facilitating cadmium detoxification in rice. Plant Biology, 22, 472-479.
- Choppala G, Saifullah, Bolan N, Bibi S, Iqbal M, Rengel Z, Kunhikrishnan A, Ashwath N, Ok Y S. 2014. Cellular mechanisms in higher plants governing tolerance to cadmium toxicity. Critical Reviews in Plant Sciences, 33, 374-391.
- Dendena B. 2012. Cadmium exclusion from rice grains: development of molecular and physiological markers.
- Dharma-Wardana M. 2018. Fertilizer usage and cadmium in soils, crops and food. Environmental Geochemistry and Health, 40, 2739-2759.
- Feng J, Shen R F, Shao J F. 2021. Transport of cadmium from soil to grain in cereal crops: A review. Pedosphere, 31, 3-10.
- Gao M, Zhou J, Liu H, Zhang W, Hu Y, Liang J, Zhou J. 2018. Foliar spraying with silicon and selenium reduces cadmium uptake and mitigates cadmium toxicity in rice. Science of the Total Environment, 631, 1100-1108.
- Ghorbani A, Emamverdian A, Pehlivan N, Zargar M, Razavi S M, Chen M. 2024. Nano-enabled agrochemicals: mitigating heavy metal toxicity and enhancing crop adaptability for sustainable crop production. Journal of Nanobiotechnology, 22, 91.
- Gul I, Manzoor M, Hashim N, Shah G M, Waani S P T, Shahid M, Antoniadis V, Rinklebe J, Arshad M. 2021. Challenges in microbially and chelate-assisted phytoextraction of cadmium and lead–A review. Environmental Pollution, 287, 117667.
- Gupta N, Yadav K K, Kumar V, Kumar S, Chadd R P, Kumar A. 2019. Trace elements in soil-vegetables interface: translocation, bioaccumulation, toxicity and amelioration-a review. Science of the Total Environment, 651, 2927-2942.
- Haider F U, Liqun C, Coulter J A, Cheema S A, Wu J, Zhang R, Wenjun M, Farooq M. 2021. Cadmium toxicity in plants: Impacts and remediation strategies. Ecotoxicology and Environmental Safety, 211, 111887.
- He B-Y, Yu D-P, Chen Y, Shi J-L, Xia Y, Li Q-S, Wang L-L, Ling L, Zeng E Y. 2017. Use of low-calcium cultivars to reduce cadmium uptake and accumulation in edible amaranth (Amaranthus mangostanus L.). Chemosphere, 171, 588-594.
- Hou D, Wang R, Gao X, Wang K, Lin Z, Ge J, Liu T, Wei S, Chen W, Xie R. 2018. Cultivar-specific response of bacterial community to cadmium contamination in the rhizosphere of rice (Oryza sativa L.). Environmental Pollution, 241, 63-73.
- Huang G, Changfeng D, Yibing M, Yurong W, Zhigao Z, Xingxiang W. 2021a. Rice (Oryza sativa L.) seedlings enriched with zinc or manganese: Their impacts on cadmium accumulation and expression of related genes. Pedosphere, 31, 849-858.
- Huang H, Li M, Rizwan M, Dai Z, Yuan Y, Hossain M M, Cao M, Xiong S, Tu S. 2021b. Synergistic effect of silicon and selenium on the alleviation of cadmium toxicity in rice plants. Journal of Hazardous Materials, 401, 123393.
- Huang S, Wang P, Yamaji N, Ma J F. 2020a. Plant nutrition for human nutrition: hints from rice research and future perspectives. Molecular Plant, 13, 825-835.
- Huang W X, Zhang D M, Cao Y Q, Dang B J, Jia W, Xu Z C, Han D. 2021c. Differential cadmium translocation and accumulation between Nicotiana tabacum L. and Nicotiana rustica L. by transcriptome combined with chemical form analyses. Ecotoxicology and Environmental Safety, 208, 111412.
- Huang X, Duan S, Wu Q, Yu M, Shabala S. 2020b. Reducing cadmium accumulation in plants: structure–function relations and tissue-specific operation of transporters in the spotlight. Plants, 9, 223.
- Hussain B, Li J, Ma Y, Tahir N, Ullah A. 2020. Effects of Fe and Mn cations on Cd uptake by rice plant in hydroponic culture experiment. PLoS One, 15, e0243174.
- Ismael M A, Elyamine A M, Moussa M G, Cai M, Zhao X, Hu C. 2019. Cadmium in plants: uptake, toxicity, and its interactions with selenium fertilizers. Metallomics, 11, 255-277.
- Kapoor B, Kumar P, Gill N S, Sharma R, Thakur N, Irfan M. 2023. Molecular mechanisms underpinning the silicon-selenium (Si-Se) interactome and cross-talk in stress-induced plant responses. Plant and Soil, 486, 45-68.
- Karami A, Shamsuddin Z H. 2010. Phytoremediation of heavy metals with several efficiency enhancer methods. African Journal of Biotechnology, 9, 3689-3698.
- Kim R-Y, Yoon J-K, Kim T-S, Yang J E, Owens G, Kim K-R. 2015. Bioavailability of heavy metals in soils: definitions and practical implementation—a critical review. Environmental geochemistry and health, 37, 1041-1061.
- Kinnell P. 2019. Comment on “Evaluating and modelling splash detachment capacity based on laboratory experiments” by Wu et al.(2019) Catena 176, 189–196. Catena, 183, 104189.
- Kumar S, Sharma A. 2019. Cadmium toxicity: effects on human reproduction and fertility. Reviews on Environmental Health, 34, 327-338.
- Leitenmaier B, Küpper H. 2013. Compartmentation and complexation of metals in hyperaccumulator plants. Frontiers in Plant Science, 4, 374.
- Li D, He T, Saleem M, He G. 2022. Metalloprotein-specific or critical amino acid residues: Perspectives on plant-precise detoxification and recognition mechanisms under cadmium stress. International Journal of Molecular Sciences, 23, 1734.
- Li H, Luo N, Li Y W, Cai Q Y, Li H Y, Mo C H, Wong M H. 2017. Cadmium in rice: transport mechanisms, influencing factors, and minimizing measures. Environmental Pollution, 224, 622-630.
- Li Y, Ding L, Zhou M, Chen Z, Ding Y, Zhu C. 2023a. Transcriptional regulatory network of plant cadmium stress response. International Journal of Molecular Sciences, 24, 4378.
- Li Y, Rahman S U, Qiu Z, Shahzad S M, Nawaz M F, Huang J, Naveed S, Li L, Wang X, Cheng H. 2023b. Toxic effects of cadmium on the physiological and biochemical attributes of plants, and phytoremediation strategies: A review. Environmental Pollution, 325, 121433.
- Li Y, Wang Y, Khan M A, Luo W, Xiang Z, Xu W, Zhong B, Ma J, Ye Z, Zhu Y. 2021. Effect of plant extracts and citric acid on phytoremediation of metal-contaminated soil. Ecotoxicology and Environmental Safety, 211, 111902.
- Lin K, Williams D V, Zeng M, Ahmed I M, Dai H, Cao F, Wu F. 2022. Identification of low grain cadmium accumulation genotypes and its physiological mechanism in maize (Zea mays L.). Environmental Science and Pollution Research, 29, 20721-20730.
- Loix C, Huybrechts M, Vangronsveld J, Gielen M, Keunen E, Cuypers A. 2017. Reciprocal interactions between cadmium-induced cell wall responses and oxidative stress in plants. Frontiers in Plant Science, 8, 1867.
- Lv G, Wang H, Xu C, Shuai H, Luo Z, Zhang Q, Zhu H, Wang S, Zhu Q, Zhang Y. 2019. Effectiveness of simultaneous foliar application of Zn and Mn or P to reduce Cd concentration in rice grains: a field study. Environmental Science and Pollution Research, 26, 9305-9313.
- Mapodzeke J M, Adil M F, Wei D, Joan H I, Ouyang Y, Shamsi I H. 2021. Modulation of key physio-biochemical and ultrastructural attributes after synergistic application of zinc and silicon on rice under cadmium stress. Plants, 10, 87.
- Mei S, Lin K, Williams D V, Liu Y, Dai H, Cao F. 2022. Cadmium accumulation in cereal crops and tobacco: A review. Agronomy, 12, 1952.
- Moradi R, Pourghasemian N, Naghizadeh M. 2019. Effect of beeswax waste biochar on growth, physiology and cadmium uptake in saffron. Journal of cleaner production, 229, 1251-1261.
- Nazar R, Iqbal N, Masood A, Khan M I R, Syeed S, Khan N A. 2012. Cadmium toxicity in plants and role of mineral nutrients in its alleviation. American Journal of Plant Science 3,1476–1489.
- Ning C C, Gao P D, Wang B Q, Lin W P, Jiang N H, Cai K Z. 2017. Impacts of chemical fertilizer reduction and organic amendments supplementation on soil nutrient, enzyme activity and heavy metal content. Journal of Integrative Aagriculture, 16, 1819-1831.
- Noh Y D, Park H J, Kim K R, Kim W I, Jung K Y, Kim S U, Owens V N, Moon J S, Yun S W, Kim S Y. 2017. Contrasting effect of phosphate on phytoavailability of arsenic and cadmium in soils supporting medicinal plants. Applied Biological Chemistry, 60, 119-128.
- Palali Delen S, Lee J, Yang J. 2024. Improving the metal composition of plants for reduced Cd and increased Zn content: molecular mechanisms and genetic regulations. Cereal Research Communications, 52, 901-918.
- Qaswar M, Hussain S, Rengel Z. 2017. Zinc fertilisation increases grain zinc and reduces grain lead and cadmium concentrations more in zinc-biofortified than standard wheat cultivar. Science of the Total Environment, 605, 454-460.
- Rahman A, Nahar K, Hasanuzzaman M, Fujita M. 2016. Manganese-induced cadmium stress tolerance in rice seedlings: Coordinated action of antioxidant defense, glyoxalase system and nutrient homeostasis. Comptes Rendus Biologies, 339, 462-474.
- Rai S, Singh P K, Mankotia S, Swain J, Satbhai S B. 2021. Iron homeostasis in plants and its crosstalk with copper, zinc, and manganese. Plant Stress, 1, 100008.
- Raza A, Habib M, Kakavand S N, Zahid Z, Zahra N, Sharif R, Hasanuzzaman M. 2020. Phytoremediation of cadmium: physiological, biochemical, and molecular mechanisms. Biology, 9, 177.
- Riaz M, Kamran M, Rizwan M, Ali S, Parveen A, Malik Z, Wang X. 2021. Cadmium uptake and translocation: selenium and silicon roles in Cd detoxification for the production of low Cd crops: a critical review. Chemosphere, 273, 129690.
- Rizwan M, Ali S, ur Rehman M Z, Rinklebe J, Tsang D C, Bashir A, Maqbool A, Tack F, Ok Y S. 2018. Cadmium phytoremediation potential of Brassica crop species: a review. Science of the Total Environment, 631, 1175-1191.
- Satoh-Nagasawa N, Mori M, Nakazawa N, Kawamoto T, Nagato Y, Sakurai K, Takahashi H, Watanabe A, Akagi H. 2012. Mutations in rice (Oryza sativa) heavy metal ATPase 2 (OsHMA2) restrict the translocation of zinc and cadmium. Plant and Cell Physiology, 53, 213-224.
- Seth C S. 2012. A review on mechanisms of plant tolerance and role of transgenic plants in environmental clean-up. The Botanical Review, 78, 32-62.
- Shahid M, Dumat C, Khalid S, Niazi N K, Antunes P M. 2017. Cadmium bioavailability, uptake, toxicity and detoxification in soil-plant system. Reviews of environmental Contamination and Toxicology 241, 73-137.
- Shahzad M, Peng D, Khan A, Ayyaz A, Askri S M H, Naz S, Huang B, Zhang G. 2024. Sufficient manganese supply is necessary for OsNramp5 knockout rice plants to ensure normal growth and less Cd uptake. Ecotoxicology and Environmental Safety, 288, 117386.
- Shanmugaraj B M, Malla A, Ramalingam S. 2019. Cadmium stress and toxicity in plants: an overview. Science Direct. 10.1016/B978-0-12-814864-8.00001-2, 1-17.
- Shiyu Q, Hongen L, Zhaojun N, Rengel Z, Wei G, Chang L, Peng Z. 2020. Toxicity of cadmium and its competition with mineral nutrients for uptake by plants: A review. Pedosphere, 30, 168-180.
- Singh P K, Pratap S G, Tandon P K. 2020. The mechanisms of trace element uptake and transport up to grains of crop plants. Sustainable Solutions for Elemental Deficiency and Excess in Crop Plants, 119-133.
- Song Y, Jin L, Wang X. 2017. Cadmium absorption and transportation pathways in plants. International Journal of Phytoremediation, 19, 133-141.
- Songmei L, Jie J, Yang L, Jun M, Shouling X, Yuanyuan T, Youfa L, Qingyao S, Jianzhong H. 2019. Characterization and evaluation of OsLCT1 and OsNramp5 mutants generated through CRISPR/Cas9-mediated mutagenesis for breeding low Cd rice. Rice Science, 26, 88-97.
- Sterckeman T, Thomine S. 2020. Mechanisms of cadmium accumulation in plants. Critical Reviews in Plant Sciences, 39, 322-359.
- Sui F-Q, Chang J-D, Tang Z, Liu W-J, Huang X-Y, Zhao F-J. 2018. Nramp5 expression and functionality likely explain higher cadmium uptake in rice than in wheat and maize. Plant and Soil, 433, 377-389.
- Takahashi R, Ishimaru Y, Senoura T, Shimo H, Ishikawa S, Arao T, Nakanishi H, Nishizawa N K. 2011. The OsNRAMP1 iron transporter is involved in Cd accumulation in rice. Journal of Experimental Botany, 62, 4843-4850.
- Tan L, Zhu Y, Fan T, Peng C, Wang J, Sun L, Chen C. 2019. OsZIP7 functions in xylem loading in roots and inter-vascular transfer in nodes to deliver Zn/Cd to grain in rice. Biochemical and BiophysicalResearch Communications, 512, 112-118.
- Thakur S, Singh L, Wahid Z A, Siddiqui M F, Atnaw S M, Din M F M. 2016. Plant-driven removal of heavy metals from soil: uptake, translocation, tolerance mechanism, challenges, and future perspectives. Environmental Monitoring and Assessment, 188, 1-11.
- Ueno D, Yamaji N, Kono I, Huang C F, Ando T, Yano M, Ma J F. 2010. Gene limiting cadmium accumulation in rice. Proceedings of the National Academy of sciences, USA 107, 16500-16505.
- Upadhyaya G, Mondal S, Roychoudhury A. 2023. Arsenic and cadmium toxicity in plants: Mitigation and remediation strategies. In: Emerging Contaminants and Plants: Interactions, Adaptations and Remediation Technologies. Springer. pp. 249-285.
- Vítková M, Puschenreiter M, Komárek M. 2018. Effect of nano zero-valent iron application on As, Cd, Pb, and Zn availability in the rhizosphere of metal (loid) contaminated soils. Chemosphere, 200, 217-226.
- Wang M, Yang Y, Chen W. 2018a. Manganese, zinc, and pH affect cadmium accumulation in rice grain under field conditions in southern China. Journal of Environmental Quality, 47, 306-311.
- Wang W, Wang Y, Hoch G, Wang Z, Gu J. 2018b. Linkage of root morphology to anatomy with increasing nitrogen availability in six temperate tree species. Plant and Soil, 425, 189-200.
- Wong C K E, Cobbett C S. 2009. HMA P-type ATPases are the major mechanism for root-to-shoot Cd translocation in Arabidopsis thaliana. New phytologist, 181, 71-78.
- Wu D, Yamaji N, Yamane M, Kashino-Fujii M, Sato K, Feng Ma J. 2016. The HvNramp5 transporter mediates uptake of cadmium and manganese, but not iron. Plant Physiology, 172, 1899-1910.
- Xu X, Zhang S, Cheng Z, Li T, Jia Y, Wang G, Yang Z, Xian J, Yang Y, Zhou W. 2020. Transcriptome analysis revealed cadmium accumulation mechanisms in hyperaccumulator Siegesbeckia orientalis L. Environmental Science and Pollution Research, 27, 18853-18865.
- Yadav B, Dubey R, Gnanasekaran P, Narayan O P. 2021. OMICS approaches towards understanding plant’s responses to counterattack heavy metal stress: an insight into molecular mechanisms of plant defense. Plant Gene, 28, 100333.
- Yang Y, Chen R, Fu G, Xiong J, Tao L. 2016. Phosphate deprivation decreases cadmium (Cd) uptake but enhances sensitivity to Cd by increasing iron (Fe) uptake and inhibiting phytochelatins synthesis in rice (Oryza sativa). Acta Physiologiae Plantarum, 38, 1-13.
- Zare A, Khoshgoftarmanesh A, Malakouti M, Bahrami H, Chaney R. 2018. Root uptake and shoot accumulation of cadmium by lettuce at various Cd: Zn ratios in nutrient solution. Ecotoxicology and Environmental Safety, 148, 441-446.
- Zhang F, Liu M, Li Y, Che Y, Xiao Y. 2019. Effects of arbuscular mycorrhizal fungi, biochar and cadmium on the yield and element uptake of Medicago sativa. Science of the Total Environment, 655, 1150-1158.
- Zhang J, Zhu Y, Yu L, Yang M, Zou X, Yin C, Lin Y. 2022. Research advances in cadmium uptake, transport and resistance in rice (Oryza sativa L.). Cells, 11, 569.
- Zhou J, Zhang C, Du B, Cui H, Fan X, Zhou D, Zhou J. 2020. Effects of zinc application on cadmium (Cd) accumulation and plant growth through modulation of the antioxidant system and translocation of Cd in low-and high-Cd wheat cultivars. Environmental Pollution, 265, 115045.
- Zhu Y X, Zhuang Y, Sun X H, Du S T. 2023. Interactions between cadmium and nutrients and their implications for safe crop production in Cd-contaminated soils. Critical Reviews in Environmental Science and Technology, 53, 2071-2091.
Figure 1.
This schematic graphic illustrates the impact of cadmium (Cd) uptake and translocation on crop health and human well-being. It highlights the molecular mechanisms underlying Cd absorption, translocation, and sequestration within plants, while also presenting effective mitigation strategies to minimize Cd toxicity and its entry into the food chain.
Figure 1.
This schematic graphic illustrates the impact of cadmium (Cd) uptake and translocation on crop health and human well-being. It highlights the molecular mechanisms underlying Cd absorption, translocation, and sequestration within plants, while also presenting effective mitigation strategies to minimize Cd toxicity and its entry into the food chain.
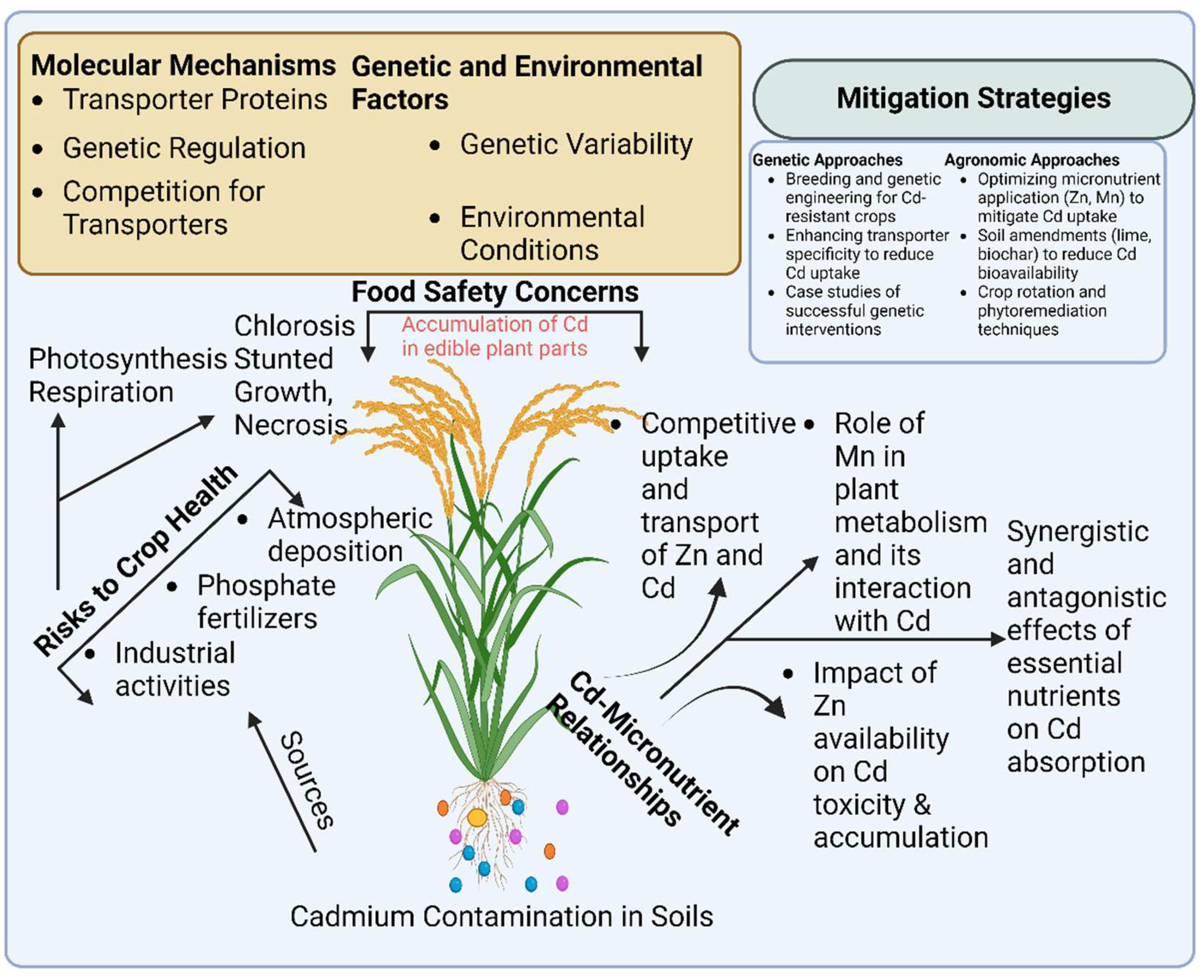
Figure 2.
This graphical representation illustrates the interaction between cadmium (Cd) and essential micronutrients in plants, highlighting its impact on metabolic processes and resistance mechanisms. Cd disrupts physiological functions, while micronutrients like Fe, Mn, and Zn modulate its uptake pathways, affecting toxicity levels. Key transporters, including NRAMPs, ZIP, CDF family, HMAs, and YSL proteins, are pivotal in facilitating Cd uptake, translocation through the xylem-phloem pathway, and sequestration, as elaborated in Section 4.
Figure 2.
This graphical representation illustrates the interaction between cadmium (Cd) and essential micronutrients in plants, highlighting its impact on metabolic processes and resistance mechanisms. Cd disrupts physiological functions, while micronutrients like Fe, Mn, and Zn modulate its uptake pathways, affecting toxicity levels. Key transporters, including NRAMPs, ZIP, CDF family, HMAs, and YSL proteins, are pivotal in facilitating Cd uptake, translocation through the xylem-phloem pathway, and sequestration, as elaborated in Section 4.
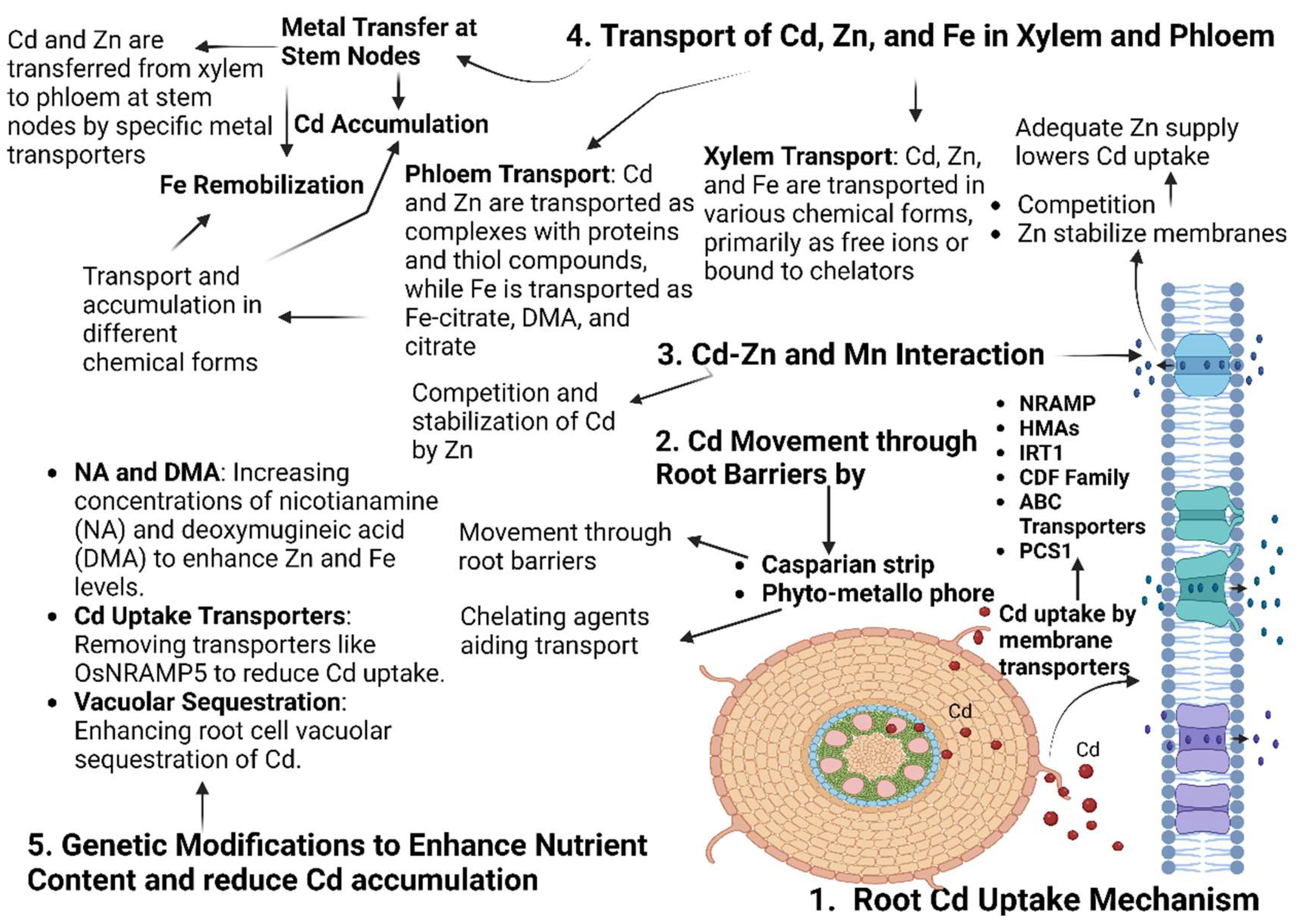
Figure 3.
This illustration elucidates the mechanisms by which plants absorb cadmium (Cd) via apoplastic and symplastic pathways, with root exudates significantly enhancing bioavailability. Key transport proteins, including OsHMA3 and CAL1, govern Cd uptake and translocation, while genotypic variations dictate the efficiency of these processes. Additionally, the figure highlights the intricate interplay between Cd and essential micronutrients, as detailed in Section 3, shedding light on their roles in modulating Cd dynamics and plant responses.
Figure 3.
This illustration elucidates the mechanisms by which plants absorb cadmium (Cd) via apoplastic and symplastic pathways, with root exudates significantly enhancing bioavailability. Key transport proteins, including OsHMA3 and CAL1, govern Cd uptake and translocation, while genotypic variations dictate the efficiency of these processes. Additionally, the figure highlights the intricate interplay between Cd and essential micronutrients, as detailed in Section 3, shedding light on their roles in modulating Cd dynamics and plant responses.
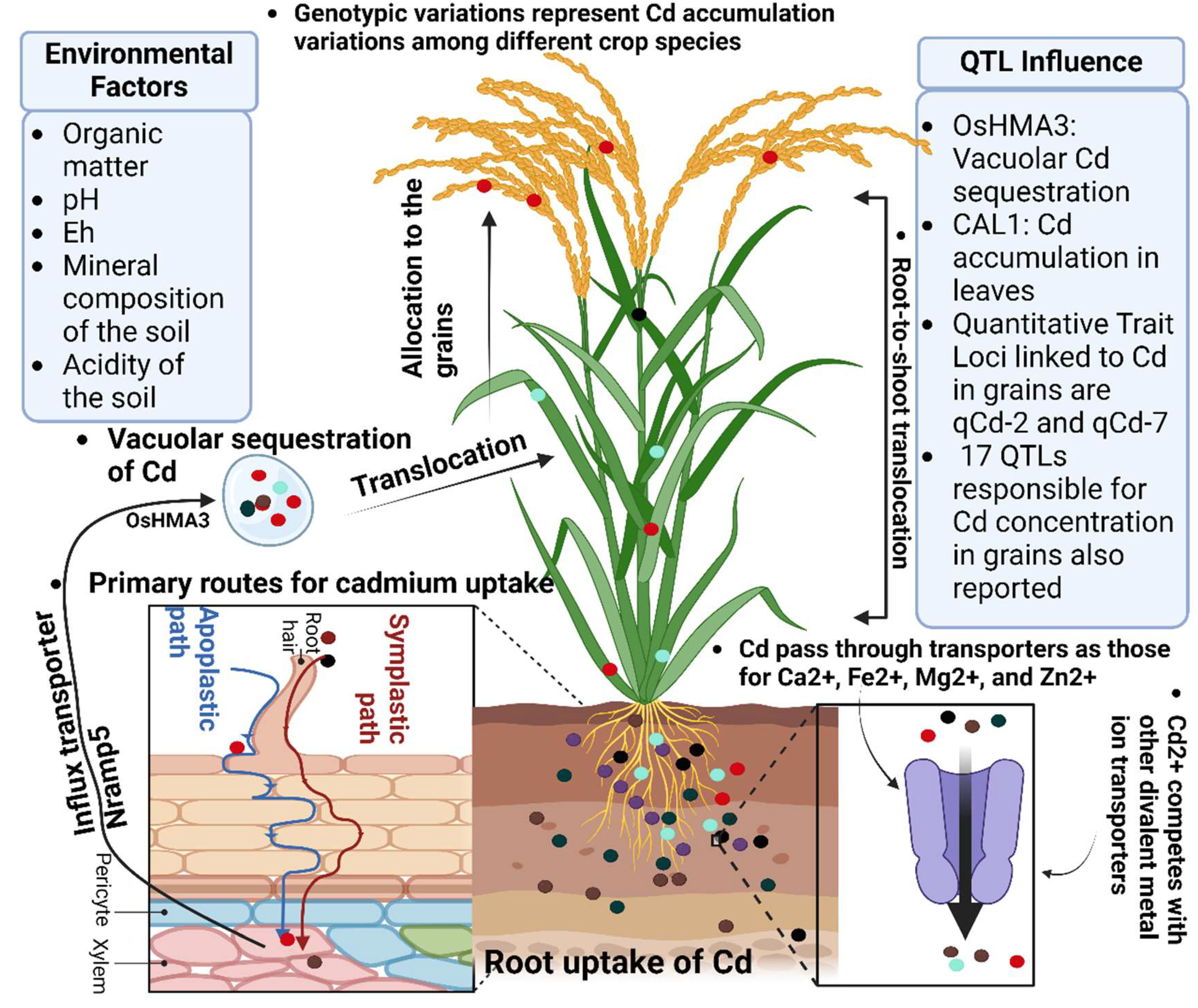

Table 1.
Key genes involved in cadmium (Cd) uptake and transport, along with their respective protein functions in rice and references. .
Table 1.
Key genes involved in cadmium (Cd) uptake and transport, along with their respective protein functions in rice and references. .
| Gene | Gene ID | Protein | Function | Reference |
| OsIRT1 | LOC_Os03g46470 | Iron-regulated transporter | Cd uptake | Ishimaru et al., 2011 |
| OsNramp1 | LOC_Os07g15460 | Natural resistance-associated macrophage protein | Cd uptake | Chang et al., 2020 |
| OsZIP3 | LOC_Os04g52310 | Zinc- and iron-regulated transporter | Cd uptake | Ondrasek et al., 2021 |
| OsHMA2 | LOC_Os06g48720 | P-type heavy metal ATPase | Cd transport | Yamaji et al., 2013b |
| OsZIP6 | LOC_Os05g07210 | Zinc- and iron-regulated transporter | Cd transport | Kavitha et al., 2015 |
| OsCCX2 | LOC_Os03g45370 | Cation/calcium exchanger | Cd transport | Hao et al., 2018 |
| OsCLT1 | LOC_Os01g72570 | CRT-like transporter | Antioxidation | Yang et al., 2016a |
Table 2.
Genes related to cadmium (Cd) transport and their roles across different plant species, including subcellular localization, functions, and references. This table provides insights into Cd uptake, efflux, and translocation in various tissues and species.
Table 2.
Genes related to cadmium (Cd) transport and their roles across different plant species, including subcellular localization, functions, and references. This table provides insights into Cd uptake, efflux, and translocation in various tissues and species.
| Plant species | Genes | Tissue | Subcellular Location | Function | Reference |
| Arabidopsis thaliana | AtIRT1 | Roots | Plasma membrane | Cd uptake | Lin et al., 2016 |
| Oryza sativa L. | OsZIP1 | Roots | Endoplasmic reticulum and plasma membrane | Cd efflux | Liu et al., 2019c |
| Nicotiana tabacum var Xanthi | NtZIP4A/B | Leaves and roots | Plasma membrane | Cd translocation | Maślińska-Gromadka et al., 2021 |
| Miscanthus sacchariflorus | MsYSL1 | Stems | Plasma membrane | Cd translocation | Houming et al., 2018 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



