
Submitted:
23 January 2025
Posted:
24 January 2025
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
Dengue is the most prevalent arthropod-borne disease, causing 100-400 million infections annually. The clinical symptoms range from asymptomatic to severe, involving hemorrhage and organ failure. Severe dengue often occurs in secondary infections with different serotypes due to antibody-dependent enhancement (ADE). During the SARS-CoV-2 pandemic, cross-reactions between dengue and SARS-CoV-2 antibody tests have been reported, due to antibody cross-reactivity between the dengue envelope protein and SARS-CoV-2 S1-RBD protein. This study analyzed the serum proteome of dengue patients with and without SARS-CoV-2 IgG antibodies. We conducted a bottom-up label-free proteomic analysis of whole serum samples from healthy donors and dengue patients, with or without SARS-CoV-2 IgG antibodies. We identified 1,122 protein sequences, 296 of which were gene-associated. Gene ontology analysis revealed that dengue patients with both SARS-CoV-2 and dengue IgG antibodies had more genes involved in immune regulation, humoral response, blood coagulation, and platelet degranulation, which may increase severe dengue risk. Unique proteins linked to platelet activation, such as coagulation factor XIII B chain, thrombospondin, Antigen KI-67, bone marrow proteoglycan, and colorectal cancer mutant protein may explain thrombocytopenia mechanisms. Our results indicate that SARS-CoV-2 IgG antibodies in dengue infection cause differential protein expression compared with healthy donors or other groups.
Keywords:
Dengue
; SARS-CoV-2
; Protein expression
; Disease severity
; Antibody-dependent enhancement (ADE)
1. Introduction
Dengue is an arthropod-borne disease, with a global prevalence, affecting 100-400 million individuals annually. The clinical spectrum encompasses asymptomatic cases, febrile manifestations, and severe presentations associated with hemorrhagic complications and organ dysfunction [1]. Severe dengue frequently occurs in secondary infections involving heterologous serotypes, wherein pre-existing antibody titers correlate with the exacerbation of severe dengue pathogenesis [2]. This phenomenon, known as Antibody-Dependent Enhancement (ADE), involves the interaction of non-neutralizing antibodies with the envelope proteins of the dengue virus, resulting in the formation of virus-antibody complexes. These complexes are subsequently recognized by immune cells, including macrophages, dendritic cells, and monocytes via Fcγ receptors [3,4]. ADE can be classified into two primary types: extrinsic and intrinsic. Extrinsic ADE refers to the enhancement of mast cell infection by virus-antibody complexes, leading to increased viral entry. In contrast, intrinsic ADE involves intricate molecular mechanisms within the host cell that modulate the innate immune response and promote viral replication and release [3].
During primary dengue infection, the dengue virus enters host cells primarily through the canonical receptor DC-SIGN, facilitating endocytosis. Once internalized, pathogen recognition receptors (PRRs), such as Toll-like receptors (TLR) 3 and 7, detect the viral presence, resulting in the secretion of interleukin-8 (IL-8) and interferon-alpha (IFN-α). These cytokines activate the Janus kinase-signal transducer and activator of transcription (JAK-STAT) pathway, leading to the production of interleukin-12 (IL-12) and interferon-gamma (IFN-γ). Collectively, these mediators orchestrate a T helper type 1 (Th1) antiviral immune response to eradicate infection. However, during intrinsic ADE, the recognition of virus-antibody complexes through Fcγ receptors leads to the upregulation of negative regulators of TLR signaling pathways, including the TAF family associated NF-κB activator (TANK) and sterile-alpha and armadillo motif-containing proteins (SARM). This upregulation results in the inhibition of interferon regulatory factors IRF1 and IRF3 [5]. Additionally, there is an increased production of IL-10 and IL-6, which further induces the expression of suppressor of cytokine signaling 3 (SOCS3), a known inhibitor of the JAK-STAT pathway, and IFN-γ signaling. These alterations collectively promote a T helper type 2 (Th2) immune response that is ineffective against viral infections, thereby exacerbating disease severity [6].
RNA viruses such as MERS-CoV, SARS-CoV, HIV, and dengue virus are commonly associated with ADE [7,8]. Notably, ADE can be triggered by antibodies targeting related viruses including Zika virus [9]. This phenomenon stems from structural similarities or molecular mimicry among viral proteins, leading to the generation of cross-reactive antibodies, often categorized as non-neutralizing antibodies. These antibodies fail to effectively opsonize the entire viral particle, enabling the virus to interact with cellular receptors and circumvent immune detection, thereby facilitating cellular entry and replication [10]. During ADE, the interaction between suboptimal antibodies and the virus enhances the viral infectivity. Specifically, when the virus-antibody complex binds to Fc receptors on immune cells, it promotes complex internalization. Paradoxically, rather than neutralizing the virus, this process may allow viral evasion of degradation pathways and support replication within immune cells.
Cross-reactive antibodies can lead to false-positive results in serological diagnostic assays. Notably, during the early stages of the SARS-CoV-2 outbreak, several patients who were later diagnosed with COVID-19 initially tested positive for dengue antibodies. This raises significant concerns regarding the specificity and reliability of serological diagnostics, particularly in areas where multiple arboviruses are endemic. Such limitations pose considerable challenges for accurate disease identification and effective public health strategies [11,12,13].
A study conducted in 2020 revealed the capacity of anti-dengue antibodies to recognize SARS-CoV-2 antigens [14]. This cross-reactivity is attributed to the structural similarities observed between the 6LVN chains, components of the SARS-CoV-2 spike protein H2R domain, and various chains of the dengue virus envelope protein, such as 4CBF, 4UIF, and 5A1Z. Subsequently, researchers performed serological tests for anti-dengue antibodies in COVID-19 patients and SARS-CoV-2 serological assays in individuals with dengue. The results indicated that patients with COVID-19 tested positive for dengue-specific IgG and IgM antibodies, whereas dengue patients tested positive for antibodies targeting the SARS-CoV-2 spike protein [15].
In a 2023 study, Cheng et al. extended the previous research by elucidating the cross-reactivity between anti-dengue antibodies and SARS-CoV-2 proteins. Their findings revealed that antibodies targeting the envelope (E) protein, precursor membrane (PrM), and non-structural protein 1 (NS1) of the dengue virus can recognize both the spike protein and S1 receptor-binding domain (S1-RBD) of SARS-CoV-2. Researchers identified specific amino acid sequences involved in this cross-reactivity: residues 343-347 on SARS-CoV-2 S1-RBD and residues 64-69 on the dengue E protein. Additionally, Cheng et al. explored the role of anti-S1-RBD antibodies in antibody-dependent enhancement (ADE) of dengue infections. Experiments in mouse models have demonstrated that these antibodies protect against hemorrhage and decrease NS1 production [16].
The structural homology between SARS-CoV-2 and dengue virus may account for false-positive outcomes in serological assays for both infections. Concerning the potential for antibody-dependent enhancement (ADE) in dengue infections triggered by anti-SARS-CoV-2 antibodies, a single study has documented this phenomenon [17], whereas another investigation employing anti-S1-RBD antibodies in animal models has dismissed it. Thus, additional validation through clinical trials and diverse research approaches is crucial to thoroughly elucidate the implications of ADE and cross-reactivity in regions where both diseases are endemic.
Proteomic analysis serves as a sophisticated method for unraveling protein dynamics during viral infections, and in dengue research, it has been used to identify potential biomarkers for prognosticating disease outcomes [18,19,20]. We employed proteomic analysis to investigate the potential role of anti-SARS-CoV-2 antibodies in antibody-dependent enhancement (ADE) during dengue infection. This study commenced with an evaluation of dengue severity markers in patients diagnosed with dengue who also demonstrated seropositivity for SARS-CoV-2 IgG antibodies. Following this, we examined serum proteome alterations. The primary focus of our investigation was to elucidate whether anti-SARS-CoV-2 antibodies could potentially influence the immune response, as is typically observed in ADE.
2. Materials and Methods
Serum samples from dengue patients were provided by the State Laboratory of Oaxaca in 2023. Serum samples from healthy donors were obtained by venipuncture. Nineteen patients were enrolled in this study. Patients were categorized into distinct groups, including the control group (healthy donors). To classify the patients, we conducted Ab tests for anti-SARS-CoV-2 IgG, Ab anti-dengue IgG/IgM, Ab anti NS1 protein, protein concentration using the Bradford method, and RT-PCR for viral identification.
2.1. Antibodies Detection
The anti-SARS-CoV-2 Ab tests were conducted using commercial ELISA immunoassay kits (Euroimmune, Germany) for IgG according to the manufacturer's protocol. Initially, a 1:101 dilution was prepared using the buffer solution and the patient serum sample. Subsequently, 100 µL of this dilution was added to each test well and incubated for 60 min at 37°C. After incubation, three washes were performed using the washing solution. Thereafter, 100 µL of enzyme conjugate was added to the test wells and incubated for 30 min at 37°C. Three additional washes were performed, followed by the addition of 100 µL of chromogenic solution. The solution was incubated for 30 min at room temperature, after which 100 µL of the stop solution was added. Absorbance was measured at a wavelength of 450-630 nm using a Chromate reader (AwarenessTechnology INC). Anti-dengue IgG-IgM Ab tests were conducted using the rapid test for detection of IgG and IgM antibodies from Dengue Duo (Standard Q), in accordance with the manufacturer's protocol.
2.2. RNA Extraction and Detection of Dengue Serotypes Using qRT-PCR
Viral RNA was extracted from 140 μL of each clinical sample in accordance with the manufacturer's instructions using the QIAamp Viral RNA Mini Kit (Qiagen, Venlo, Netherlands). RNA extraction was performed using an automated protocol (Qiagen, Hilden, Germany), with an elution volume of 60 μL. The RNA intended for use as a template was stored at − 80°C until testing. qRT-PCR assays were conducted following the Viasure Dengue Serotyping Real-Time PCR Detection Kit protocol, as specified by the manufacturer. A total of 5 μL template RNA samples (4–140 ng/μL) and negative and positive controls were added to each well. A Fast ABI 7500 thermocycler (Thermo Fisher Scientific, Waltham, MA, USA) was used. The PCR procedure was performed with one cycle at 45°C for 15 min (RT) and one cycle at 95°C for 2 min (initial denaturation), followed by 45 cycles at 95°C for 10 s (denaturation) and 60°C for 50 s (annealing/extension (data collection)). A positive control and a negative control of molecular biology grade were used concurrently in all experiments. A sample was considered positive if its Ct value was less than 40.
2.3. Protein Integrity
Serum proteins were analyzed using 12% acrylamide SDS-PAGE gel. A 5 µg serum sample mixed with 1X Laemmli loading buffer was loaded into each well. Electrophoresis was conducted for 90 min at 110 V, 3.0 A, and 300 watts in an electrophoresis chamber (Mini-Protean, Bio-Rad). The gel was subsequently stained with 0.02% Coomassie Blue G-250. Only samples with standard protein bands ranging from 10 kDa in the human serum were used.
2.4. Label-Free Based Quantitative Proteomic Analysis of Serum Proteome
After classification, a pool was established for proteomic analysis, using serum samples from each individual. Bottom-up label-free quantitative proteomic analysis was performed using a minimum of 300 µg of protein per patient pool. Pooled samples were loaded onto an acrylamide electrophoresis gel, followed by band excision and trypsin digestion (Promega, Madison, WI, USA). The resulting peptides were resuspended in 20 μL 0.1% formic acid for LC-MS/MS analysis.
Chemicals and Instrumentation. Sigma (St. Louis, MO, USA), dithiothreitol (DTT), iodoacetamide (IAA), formic acid (FA), and acetonitrile (ACN). Trypsin derived from bovine pancreas was acquired from Promega (Madison, WI). Ultrapure water was prepared using a Millipore purification system (Billerica, MA). The experimental assays were conducted using an Ultimate 3000 nano UHPLC system in conjunction with a Q Exactive HF mass spectrometer (Thermo Fisher Scientific, USA) fitted with an ESI nanospray source.
Nano LC-MS/MS Analysis. Proteomic analyses were conducted using Creative Proteomics services (Shirley, NY, USA) with an Ultimate 3000 nano UHPLC system (Thermo Fisher Scientific, USA), which incorporated a nanocolumn trapping column (PepMap C18, 100 Å, 100 μm × 2 cm, 5 μm) and an analytical column (PepMap C18, 100 Å, 75 μm × 50 cm, 2 μm). The sample loading was set to 1 μg. The mobile phase consisted of 0.1% formic acid in water (A) and 0.1% formic acid in 80% acetonitrile (B). The total flow rate was maintained at 250 nl/min. The LC linear gradient was as follows: 2–8% buffer B for 3 min, 8–20% buffer B for 56 min, 20–40% buffer B for 37 min, and 40–90% buffer B for 4 min.
Mass spectrometric analysis. A comprehensive scan was performed across a 300-1,650 m/z range with a 60,000 resolution at 200 m/z. A full-scan automatic gain control target was established at 3e6. For the MS/MS scan, the top 20 modes were employed, utilizing the following settings: 15,000 resolution at 200 m/z, 1e5 automatic gain control target, 19 ms maximum injection time, 28% normalized collision energy; 1.4 Th isolation window. Charge state exclusion was applied to unassigned, 1, and states exceeding 6. Dynamic exclusion was performed for 30 s.
2.5. Data Analysis
Four raw MS files were analyzed and searched against a human protein database based on the species of the samples using Maxquant (1.6.2.6). The parameters were established as follows: protein modifications were carbamidomethylation (C) (fixed) and oxidation (M) (variable); enzyme specificity, trypsin; maximum missed cleavage 2, precursor ion mass tolerance, 10 ppm; and MS/MS tolerance, 0.5 Da.
The ratio of abundance (RA) was calculated by comparing protein abundance in the control/groups of interest. Proteins with RA > 2 were considered to be relatively more abundant in the acute dengue infection state. Enrichment analysis of biological processes was conducted using the ShinyGO bioinformatic website 0.77 (Ge SX, Jung D & Yao R, Bioinformatics 36:2628–2629, 2020). Protein IDs were obtained for each protein using the UniProt ID number.
3. Results
Patients were classified into four groups: control group (healthy donors), patients with Ab anti SARS-CoV-2 IgG and NS1 protein (dengue infection, group one); patients with Ab anti SARS-CoV-2 IgG and Ab anti-dengue IgG (group two); and patients with Ab anti-dengue IgG and NS1 protein (Group three) (Table 1).
3.1. Serum Proteome Analysis
We identified 1,121 protein sequences in the samples. Sequences without an associated gene and those identified solely by site were eliminated along with uncharacterized proteins, duplicates, and gender-related proteins. A total of 296 proteins were identified with high confidence levels. Protein expression differed among the groups as follows: control group, 236 proteins; group one, 256 proteins; group two, 261 proteins; and group three, 255 proteins. Gene enrichment analysis of proteins expressed in each group revealed differences in the biological processes occurring during acute dengue infection under the various evaluated conditions compared to healthy donors, as illustrated in Figure 1. Regulation of the immune response, humoral immune response, blood coagulation, and platelet degranulation are processes that exhibit the most notable changes.
3.1.1. Differentially Expressed Proteins (DEPs)
Twelve of the 296 proteins were identified as DEPs across all groups. Three DEPs were exclusive to the control group: syntaxin-binding protein 5, Galectin-1, and Glucose-6-phosphate isomerase.
Three DEPs were identified in group 1: Coagulation factor IX, Ribosomal protein L19, and Peptidyl-prolyl cis-trans isomerase B. Five DEPs were found in group 2: Thrombospondin-1, Colorectal mutant cancer protein, Coagulation factor XIII B chain, Antigen KI-67, and Bone marrow proteoglycan. Only one DEP was identified in group 3: putative uncharacterized zinc finger protein 814. The functions of most of these proteins have been elucidated, with the exception of the putative uncharacterized zinc finger protein 814. Expression of Galectin-1, Ribosomal protein L19, Peptidyl-prolyl cis-trans isomerase B, and Antigen KI-67 has been described during viral infection, although their specific roles are not fully understood. Proteins such as Coagulation factor IX and Coagulation factor XIII B chain are altered during dengue infection. The involvement of syntaxin-binding protein 5, Glucose-6-phosphate isomerase, Thrombospondin-1, Colorectal mutant cancer protein, and bone marrow proteoglycan in viral infection is unknown. However, based on their functions, it is reasonable to hypothesize that they may play a role in infection. Notably, the bone marrow proteoglycan found in group two was secreted by activated eosinophils, which are key mediators of the Th2 immune response. The protein ID and most relevant functions of each DEP are described in Table 2.
3.2. Protein Abundance Index
We determined the abundance ratio of proteins (AR) relative to the control group and identified 30 proteins among the groups with an AR > 2.0, as presented in Table 3). Most of these proteins have been previously identified during viral infections, whereas others play crucial roles in the innate immune response. Notably, group two exhibited the highest number of proteins with an AR > 2, totaling 23, followed by group one with 18 and group three with 14.
4. Discussion
Severe dengue is associated with several risk factors, including age, comorbidities, and subsequent infections with different serotypes. The potential role of anti-SARS-CoV-2 antibodies as non-neutralizing antibodies in dengue infections and their capacity to trigger ADE is a critical consideration of the cross-reactivity observed between these two viruses.
In this study, we conducted a comprehensive proteomic analysis of the serum of patients with acute dengue infection, stratified based on the presence of anti-SARS-CoV-2 IgG, anti-dengue IgG/IgM antibodies, or both. Our high-throughput analysis identified 1,196 protein sequences, of which 296 were associated with known protein genes. Comparative analysis of protein expression profiles revealed distinct differences between healthy donors and infected patients, indicating significant alterations in the serum proteome during dengue infection. We observed a notable trend in protein expression among the patient groups. Patients with either anti-SARS-CoV-2 IgG (group 1) or anti-dengue antibodies (group 3) exhibited similar numbers of expressed proteins (256 and 255, respectively). However, the most striking observation was in group 2, comprising patients with both anti-SARS-CoV-2 IgG and anti-dengue IgG/IgM antibodies, which demonstrated the highest number of expressed proteins (261).
This differential protein expression pattern aligns with the established understanding of antibody-dependent enhancement (ADE) in dengue infections. Heterologous antibodies, particularly those from different dengue serotypes, can exacerbate ADE clinical manifestations. Recent evidence has extended this concept to antibodies against other flaviviruses, such as Zika [9] and even anti-SARS-CoV-2 antibodies, as evidenced by a fatal case of dengue attributed to ADE induced by anti-SARS-CoV-2 IgG antibodies [17]. The observed increase in protein expression in group 2 supports our hypothesis that the presence of both anti-SARS-CoV-2 and anti-dengue antibodies may synergistically enhance ADE, potentially leading to more severe dengue manifestations. This finding is particularly significant, as it suggests that individuals with prior SARS-CoV-2 exposure may be at a higher risk for severe dengue outcomes, a critical consideration in regions where both diseases are endemic. Furthermore, the elevated protein expression in Group 2 may reflect a more robust and potentially dysregulated immune response, a characteristic of ADE. This heightened protein expression could be indicative of increased viral replication, enhanced inflammatory responses, and altered cellular signaling pathways, all of which are hallmarks of ADE-mediated severe dengue infections.
Through comprehensive enrichment analysis of gene expression patterns, we identified significant alterations in key biological processes associated with dengue pathogenesis. The most striking findings emerged in four critical domains: immune response regulation, humoral immune response, blood coagulation, and platelet degranulation. These processes were markedly upregulated in dengue patients compared to healthy controls, with particularly pronounced effects in patients harboring both anti-SARS-CoV-2 and anti-dengue antibodies (Group 2).
Enrichment analysis revealed a robust immune response signature, with the immune effector process and the response to external biotic stimuli showing the highest number of associated genes. This pattern was consistently elevated across all patient groups but reached its peak in group 2, suggesting potentially excessive immune activation in patients with dual antibodies.
The blood coagulation pathway showed significant enrichment, with multiple genes involved in hemostasis and differential expression of platelet function. This finding aligns with the clinical manifestations of severe dengue, in which coagulopathy plays a central role in disease progression. Platelet degranulation-related genes showed marked upregulation, particularly in Group 2, indicating enhanced platelet activation and consumption. This observation provides a molecular basis for the increased risk of bleeding complications in patients with severe dengue.
The most compelling finding was the consistently higher gene enrichment scores in Group 2 across all analyzed biological processes. This pattern suggests that the presence of both anti-SARS-CoV-2 and anti-dengue antibodies may create a synergistic effect, potentially amplifying the immune response and associated pathological processes.
The enhanced activation of these pathways, particularly in group 2, provides molecular evidence for the increased risk of disease severity in patients with dual antibodies. The simultaneous upregulation of the immune response and coagulation pathways suggests a complex interplay between immune activation and hemostatic dysfunction, potentially contributing to more severe clinical manifestations. This provides crucial molecular insights into the mechanisms underlying severe dengue manifestations and suggests that the presence of anti-SARS-CoV-2 antibodies may contribute to disease severity through enhanced immune activation and coagulation dysfunction. This understanding has important implications for the clinical management and risk assessment of dengue patients with prior SARS-CoV-2 exposure.
Analysis of differentially expressed proteins (DEP) in each group revealed significant findings. In healthy groups, the presence of Galectin-1/LGALS1 has been shown to suppress viral replication in both cell culture and murine models. Its absence in our patients facilitated dengue replication [23], underscoring its crucial role during infection, although the underlying mechanism remains elusive. The syntaxin-binding protein 5/STXBP5, which is synthesized in endothelial cells and platelets, is an intriguing case. While its involvement in dengue infection is not well documented, studies have shown that endothelial cells lacking this protein exhibit increased P-selectin expression and elevated vWF levels in the plasma [21], promoting platelet adherence and agglutination [48]. This phenomenon has also been observed in dengue infections. Furthermore, STXBP5 knockout in mice results in reduced PF4/CXCL4 secretion from platelets [22]. Given that low PF4/CXCL4 levels correlate with dengue severity, although the cause remains unclear [49], the absence of STXBP5 in groups 1, 2, and 3 may suggest a link between dengue virus and STXBP5 downregulation, which may explain the decreased PF4 synthesis in platelets and warrants further investigation. Glucose-6-phosphate isomerase/GPI, a critical glycolytic enzyme, plays an unclear role in viral infection. However, considering that microorganisms often utilize the Warburg pathway for efficient energy production during replication [50], the absence of this protein in dengue patients may represent a host defense mechanism to impede this metabolic pathway and combat infection. Further research is needed to substantiate this hypothesis.
Differentially expressed proteins (DEPs) in group one, specifically ribosomal protein L19/RPL19 and Peptidyl-prolyl cis-trans isomerase B/PPIB, have been identified as crucial host factors in the replication of flaviviruses such as Yellow Fever Virus (YFV) and West Nile Virus (WNV) [27,28]. The detection of coagulation factor IX indicates an ongoing coagulation process. Disruptions in coagulation during dengue infection have been previously reported [24,25]. The exact mechanisms underlying this phenomenon remain incompletely understood; however, they may involve platelet aggregation, leading to microthrombus formation, as well as increased tissue factor (TF) expression by endothelial cells or activated platelets [51], which subsequently triggers the coagulation cascade.
In the second group, a high number of differentially expressed proteins (DEPs) were identified, suggesting an accelerated rate of viral replication facilitated by the simultaneous presence of antibodies targeting both dengue and SARS-CoV-2. Most DEPs are associated with mechanisms underlying severe clinical manifestations of dengue, potentially driven by the antibody-dependent enhancement (ADE) phenomenon. Thrombospondin-1/THBS1 serves as an indicator of an elevated platelet activation rate and is released from alpha granules during this process. Enhanced platelet activation leads to diminished platelet levels due to increased consumption and destruction, correlating with severe dengue cases, in which NS1 proteins [52] and immunocomplexes are primary activation triggers [53]. Reduced platelet counts are typically noted in the critical phase of dengue, commencing three–four days post-infection and manifesting as petechiae on the skin. These low levels continue through the convalescent phase, generally normalizing within two weeks post-infection. While the progression of complications is influenced by host factors (age and comorbidities), earlier platelet activation, such as that during the acute phase in our study subjects, increases the likelihood of complications. Platelets not only contribute to bleeding, plasma leakage, and organ failure but also play a crucial role in immune response modulation, stimulating immune cell activation, and pro-inflammatory molecule release [54].
The presence of coagulation factor XIII B chain/FIIIB in this group suggests potential coagulation pathway dysfunction, as this protein is crucial for FXIII-A2B2 complex formation. FXIII plays a vital role in stabilizing fibrin clots at the termination of the coagulation pathway. Despite high FIIIB levels in normal serum, its absence in other groups, including healthy controls, is noteworthy. We postulate that in group two, the absence of STXBP5, as seen in Groups 1 and 3, resulted in elevated immature vWF levels, facilitating platelet adherence and microthrombus formation, thus activating the coagulation cascade. Early coagulation cascade activation in dengue infection is also linked to severe cases owing to the rapid consumption and depletion of coagulation factors, which become unavailable in later infection stages to prevent bleeding and plasma leakage [24,25].
Furthermore, the third DEP, Bone marrow proteoglycan/PRG2, which is secreted by eosinophils, also activates platelets. Eosinophil activation is induced by IL4 and IL10, which are produced in large quantities during the ADE phenomenon, shifting the immune response from Th1 to Th2 [6]. Consequently, the detection of PRG2, FXIIIB, and THBS1 strongly indicated, based on their functions, the potential presence of an ADE phenomenon in group 2.
The final DEPs identified in this group, Antigen KI-67/MKI67 and Colorectal mutant cancer protein/MCC, were involved in cellular replication. Given that these processes are dysregulated in other infections, it would be valuable to investigate their potential roles in dengue. Finally, Putative uncharacterized zinc finger protein 814/ZNF814, the only DEPs expressed in group 3, was not associated with any function, but other proteins that contain the zinc finger domain participate in the regulation of the immune response during viral infection [55]; therefore, future evaluation of this protein should be performed.
The final analysis, examining the abundance ratio among the groups, revealed that 30 of the 296 proteins were overexpressed during dengue infection, as demonstrated by AR > 2.0. Upon examination of each group, it was observed that group two contained the highest number of overexpressed proteins (23), followed by groups 1 (18 and group three (14. The majority of the overexpressed proteins have been previously identified in viral infections, including Alpha-2-macroglobulin, Inter-alpha-trypsin inhibitor heavy chain H2, Alpha-1B-glycoprotein, Leucine-rich-alpha-2 glycoprotein, Insulin-like growth factor-binding protein complex acid labile subunit, N-acetylmuramoyl-L-alanine amidase, 60 kDa heat shock protein, mitochondrial, retinol-binding protein 4/ RBP4, Beta-Ala-His dipeptidase/ CNDP1, Tropomyosin alpha-1 chain/TPM1, monocyte differentiation antigen CD14, Triosephosphate isomerase/ TPI1, Nucleophosmin/ NPM1 and Prothymosin alpha/PTMA. Others, such as General transcription factor IIF subunit/ GTF2F1, Serum amyloid A-4 protein, Leucine-rich repeat protein 1, Attractin, L-lactate dehydrogenase B chain and Biotinidase/ BTD, currently lack evidence of participation during viral infection. The remaining proteins are involved in the innate immune response, including C-reactive protein/CRP, complement components C9, C7, C1q subcomponent subunit B, Complement C1q subcomponent subunit A, C4b-binding protein alpha chain, and Complement C1r subcomponent-like protein, or in blood coagulation, such as Kallistatin and Fibrinogen gamma chain/FGG. Dysregulation of proteins in the complement system during dengue infection has been previously described [18], and it is anticipated that these proteins were also overexpressed in the patients in this study.
Proteomic analysis revealed a compelling association between the presence of both anti-dengue and anti-SARS-CoV-2 antibodies and enhanced immune activation patterns, suggesting ADE. This finding has significant implications for populations in dengue-endemic regions with prior SARS-CoV-2 exposure, whether through natural infection or vaccination, as it may contribute to increased disease severity. The observed increase in severe dengue cases and associated mortality in the Americas following the COVID-19 pandemic, as reported by the WHO/PAHO, adds epidemiological support to this molecular evidence [56].
However, it is crucial to note that ADE development involves multiple variables in addition to the presence of antibodies alone. Our findings suggest that anti-SARS-CoV-2 antibodies may function as an additional risk factor for severe dengue manifestations, acting in concert with established risk factors including host health status and viral characteristics. This proteomic approach may raise concerns in biological interpretation, particularly as the differential expression of proteoforms and protein species across groups of interest underscores the intricate nature of the immune response during dengue infection, particularly in the context of ADE. The proteins identified in our analysis not only reflect alterations in immune regulation and coagulation processes but also highlight the diverse functional roles that different proteoforms may play in pathophysiological contexts.
The comprehensive proteomic approach employed in this study not only elucidated the potential mechanisms of disease severity but also identified candidate biomarkers for disease progression and diagnosis. However, the limitations of our investigation include a potential lack of comprehensive characterization of all proteoforms present in serum samples, which may influence our understanding of protein interactions and their biological significance. An additional limitation is that reliance on bottom-up proteomics techniques may obscure critical post-translational modifications and variations among protein species, thereby complicating the interpretation of results.
Despite our findings require validation through subsequent studies, aimed to explore these aspects further, employing advanced proteomic techniques to elucidate the full complexity of the dengue proteome and its interactions with antibodies from SARS-CoV-2, this bottom-up proteomic analysis provides a crucial foundation for understanding the complex interplay between SARS-CoV-2 and dengue virus infections.
5. Conclusions
Proteomic analysis of serum samples from dengue-infected patients with varying antibody profiles has revealed significant alterations in key biological processes. Enrichment analysis demonstrated the upregulation of proteins involved in platelet activation, coagulation cascades, and Th2 immune responses, particularly in patients with both anti-SARS-CoV-2 IgG and anti-dengue IgG/IgM antibodies. These molecular signatures parallel those typically observed in secondary dengue infections characterized by the ADE phenomenon, as evidenced by differential gene enrichment patterns across immune effector processes and hemostatic pathways. While these findings provide compelling molecular evidence for the potential ADE-mediated pathogenesis in the context of dual-antibody presence, further clinical studies are warranted to validate these observations and establish their therapeutic implications.
Author Contributions
Conceptualization, Investigation, and Writing - Original Draft, ECA; Visualization, Project administration; Investigation, Resources, Visualization, Writing - Review & Editing, and Funding acquisition, MMC; Investigation, Formal Analysis, Methodology, and Validation, LRP; Conceptualization, Supervision, Data Curation, Resources, Writing - Original Draft, Writing - Review & Editing, Visualization, Funding acquisition, and Project administration, JA. All authors have read and agreed to the published version of the manuscript.”.
Funding
This Project was supported by the Consejo Nacional de Humanidades Ciencia y Tecnologia, CONAHCYT (scholarship #1070651), and Autonomous University Benito Juárez of Oaxaca.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki and was approved by the Committee of the Institutional Review Board of the Experimental Biomedicine program.
Informed Consent Statement
Informed consent was obtained from all the subjects involved in the study.
Data Availability Statement
The original contributions presented in this study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author(s)..
Acknowledgments
The authors thank all the instructors of the Undergraduate Biochemical Laboratory FMC-UABJO for technical assistance during this research.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Dengue and Severe Dengue. Available online: https://www.who.int/news-room/fact-sheets/detail/dengue-and-severe-dengue (accessed on 9 January 2025).
- Katzelnick, L.C.; Gresh, L.; Halloran, M.E.; Mercado, J.C.; Kuan, G.; Gordon, A.; Balmaseda, A.; Harris, E. Antibody-Dependent Enhancement of Severe Dengue Disease in Humans. Science 2017, 358, 929–932. [Google Scholar] [CrossRef]
- Chan, C.Y.Y.; Low, J.Z.H.; Gan, E.S.; Ong, E.Z.; Zhang, S.L.-X.; Tan, H.C.; Chai, X.; Ghosh, S.; Ooi, E.E.; Chan, K.R. Antibody-Dependent Dengue Virus Entry Modulates Cell Intrinsic Responses for Enhanced Infection. mSphere 2019, 4, e00528–19. [Google Scholar] [CrossRef]
- Rodrigo, W.W.S.I.; Jin, X.; Blackley, S.D.; Rose, R.C.; Schlesinger, J.J. Differential Enhancement of Dengue Virus Immune Complex Infectivity Mediated by Signaling-Competent and Signaling-Incompetent Human FcγRIA (CD64) or FcγRIIA (CD32). J. Virol. 2006, 80, 10128–10138. [Google Scholar] [CrossRef] [PubMed]
- Narayan, R.; Tripathi, S. Intrinsic ADE: The Dark Side of Antibody Dependent Enhancement During Dengue Infection. Front. Cell. Infect. Microbiol. 2020, 10, 580096. [Google Scholar] [CrossRef] [PubMed]
- Ubol, S.; Phuklia, W.; Kalayanarooj, S.; Modhiran, N. Mechanisms of Immune Evasion Induced by a Complex of Dengue Virus and Pre-existing Enhancing Antibodies. J. Infect. Dis. 2010, 201, 923–935. [Google Scholar] [CrossRef]
- Wen, J.; Cheng, Y.; Ling, R.; Dai, Y.; Huang, B.; Huang, W.; Zhang, S.; Jiang, Y. Antibody-Dependent Enhancement of Coronavirus. Int. J. Infect. Dis. 2020, 100, 483–489. [Google Scholar] [CrossRef] [PubMed]
- Sawant, J.; Patil, A.; Kurle, S. A Review: Understanding Molecular Mechanisms of Antibody-Dependent Enhancement in Viral Infections. Vaccines 2023, 11, 1240. [Google Scholar] [CrossRef] [PubMed]
- Bonheur, A.N.; Thomas, S.; Soshnick, S.H.; McGibbon, E.; Dupuis, A.P.; Hull, R.; Slavinski, S.; Del Rosso, P.E.; Weiss, D.; Hunt, D.T.; et al. A Fatal Case Report of Antibody-Dependent Enhancement of Dengue Virus Type 1 Following Remote Zika Virus Infection. BMC Infect. Dis. 2021, 21, 749. [Google Scholar] [CrossRef] [PubMed]
- Jain, D.; Salunke, D.M. Antibody Specificity and Promiscuity. Biochem. J. 2019, 476, 433–447. [Google Scholar] [CrossRef] [PubMed]
- Yan, G.; Lee, C.K.; Lam, L.T.M.; Yan, B.; Chua, Y.X.; Lim, A.Y.N.; Phang, K.F.; Kew, G.S.; Teng, H.; Ngai, C.H.; et al. Covert COVID-19 and False-Positive Dengue Serology in Singapore. Lancet Infect. Dis. 2020, 20, 536. [Google Scholar] [CrossRef]
- Kembuan, G.J. Dengue Serology in Indonesian COVID-19 Patients: Coinfection or Serological Overlap? IDCases 2020, 22, e00927. [Google Scholar] [CrossRef]
- Spinicci, M.; Bartoloni, A.; Mantella, A.; Zammarchi, L.; Rossolini, G.M.; Antonelli, A. Low Risk of Serological Cross-Reactivity between Dengue and COVID-19. Mem. Inst. Oswaldo Cruz 2020, 115, e200225. [Google Scholar] [CrossRef]
- Nath, H.; Mallick, A.; Roy, S.; Sukla, S.; Biswas, S. Computational Modelling Supports That Dengue Virus Envelope Antibodies Can Bind to SARS-CoV-2 Receptor Binding Sites: Is Pre-Exposure to Dengue Virus Protective against COVID-19 Severity? Comput. Struct. Biotechnol. J. 2021, 19, 459–466. [Google Scholar] [CrossRef] [PubMed]
- Lustig, Y.; Keler, S.; Kolodny, R.; Ben-Tal, N.; Atias-Varon, D.; Shlush, E.; Gerlic, M.; Munitz, A.; Doolman, R.; Asraf, K.; et al. Potential Antigenic Cross-Reactivity Between Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) and Dengue Viruses. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2021, 73, e2444–e2449. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.-L.; Chao, C.-H.; Lai, Y.-C.; Hsieh, K.-H.; Wang, J.-R.; Wan, S.-W.; Huang, H.-J.; Chuang, Y.-C.; Chuang, W.-J.; Yeh, T.-M. Antibodies against the SARS-CoV-2 S1-RBD Cross-React with Dengue Virus and Hinder Dengue Pathogenesis. Front. Immunol. 2022, 13, 941923. [Google Scholar] [CrossRef] [PubMed]
- Beig, I.; Talwar, D.; Kumar, S.; Hulkoti, V. Post Covid Fatal Antibody Dependent Enhancement of Dengue Infection in a Young Male: Double Trouble. 2021.
- Kumar, Y.; Liang, C.; Bo, Z.; Rajapakse, J.C.; Ooi, E.E.; Tannenbaum, S.R. Serum Proteome and Cytokine Analysis in a Longitudinal Cohort of Adults with Primary Dengue Infection Reveals Predictive Markers of DHF. PLoS Negl. Trop. Dis. 2012, 6, e1887. [Google Scholar] [CrossRef]
- Han, L.; Ao, X.; Lin, S.; Guan, S.; Zheng, L.; Han, X.; Ye, H. Quantitative Comparative Proteomics Reveal Biomarkers for Dengue Disease Severity. Front. Microbiol. 2019, 10, 2836. [Google Scholar] [CrossRef]
- Garishah, F.M.; Boahen, C.K.; Vadaq, N.; Pramudo, S.G.; Tunjungputri, R.N.; Riswari, S.F.; Van Rij, R.P.; Alisjahbana, B.; Gasem, M.H.; Van Der Ven, A.J.A.M.; et al. Longitudinal Proteomic Profiling of the Inflammatory Response in Dengue Patients. PLoS Negl. Trop. Dis. 2023, 17, e0011041. [Google Scholar] [CrossRef]
- Zhu, Q.; Yamakuchi, M.; Ture, S.; De La Luz Garcia-Hernandez, M.; Ko, K.A.; Modjeski, K.L.; LoMonaco, M.B.; Johnson, A.D.; O’Donnell, C.J.; Takai, Y.; et al. Syntaxin-Binding Protein STXBP5 Inhibits Endothelial Exocytosis and Promotes Platelet Secretion. J. Clin. Invest. 2014, 124, 4503–4516. [Google Scholar] [CrossRef] [PubMed]
- Ye, S.; Huang, Y.; Joshi, S.; Zhang, J.; Yang, F.; Zhang, G.; Smyth, S.S.; Li, Z.; Takai, Y.; Whiteheart, S.W. Platelet Secretion and Hemostasis Require Syntaxin-Binding Protein STXBP5. J. Clin. Invest. 2014, 124, 4517–4528. [Google Scholar] [CrossRef] [PubMed]
- Toledo, K.A.; Fermino, M.L.; Andrade, C.D.C.; Riul, T.B.; Alves, R.T.; Muller, V.D.M.; Russo, R.R.; Stowell, S.R.; Cummings, R.D.; Aquino, V.H.; et al. Galectin-1 Exerts Inhibitory Effects during DENV-1 Infection. PLoS ONE 2014, 9, e112474. [Google Scholar] [CrossRef]
- Bhamarapravati, N. Hemostatic Defects in Dengue Hemorrhagic Fever. Clin. Infect. Dis. 1989, 11, S826–S829. [Google Scholar] [CrossRef] [PubMed]
- Funahara, Y. ; Sumarmo; Wirawan, R. Features of DIC in Dengue Hemorrhagic Fever. In Current Studies in Hematology and Blood Transfusion; Abe, T., Yamanaka, M., Eds.; S. Karger AG, 1983; Vol. 49, pp. 201–211 ISBN 978-3-8055-3726-1.
- Yang, E.-J.; Seo, J.-W.; Choi, I.-H. Ribosomal Protein L19 and L22 Modulate TLR3 Signaling. Immune Netw. 2011, 11, 155. [Google Scholar] [CrossRef] [PubMed]
- Petrova, E.; Gracias, S.; Beauclair, G.; Tangy, F.; Jouvenet, N. Uncovering Flavivirus Host Dependency Factors through a Genome-Wide Gain-of-Function Screen. Viruses 2019, 11, 68. [Google Scholar] [CrossRef]
- Qing, M.; Yang, F.; Zhang, B.; Zou, G.; Robida, J.M.; Yuan, Z.; Tang, H.; Shi, P.-Y. Cyclosporine Inhibits Flavivirus Replication through Blocking the Interaction between Host Cyclophilins and Viral NS5 Protein. Antimicrob. Agents Chemother. 2009, 53, 3226–3235. [Google Scholar] [CrossRef] [PubMed]
- Isenberg, J.S.; Romeo, M.J.; Yu, C.; Yu, C.K.; Nghiem, K.; Monsale, J.; Rick, M.E.; Wink, D.A.; Frazier, W.A.; Roberts, D.D. Thrombospondin-1 Stimulates Platelet Aggregation by Blocking the Antithrombotic Activity of Nitric Oxide/cGMP Signaling. Blood 2008, 111, 613–623. [Google Scholar] [CrossRef]
- Pimanda, J.E.; Ganderton, T.; Maekawa, A.; Yap, C.L.; Lawler, J.; Kershaw, G.; Chesterman, C.N.; Hogg, P.J. Role of Thrombospondin-1 in Control of von Willebrand Factor Multimer Size in Mice. J. Biol. Chem. 2004, 279, 21439–21448. [Google Scholar] [CrossRef] [PubMed]
- Sigglekow, N.D.; Pangon, L.; Brummer, T.; Molloy, M.; Hawkins, N.J.; Ward, R.L.; Musgrove, E.A.; Kohonen-Corish, M.R.J. Mutated in Colorectal Cancer Protein Modulates the NFκB Pathway. ANTICANCER Res. 2012.
- Edwards, S.K.; Baron, J.; Moore, C.R.; Liu, Y.; Perlman, D.H.; Hart, R.P.; Xie, P. Mutated in Colorectal Cancer (MCC) Is a Novel Oncogene in B Lymphocytes. J. Hematol. Oncol.J Hematol Oncol 2014, 7, 56. [Google Scholar] [CrossRef]
- Maqueda-Alfaro, R.A.; Marcial-Juárez, E.; Calderón-Amador, J.; García-Cordero, J.; Orozco-Uribe, M.; Hernández-Cázares, F.; Medina-Pérez, U.; Sánchez-Torres, L.E.; Flores-Langarica, A.; Cedillo-Barrón, L.; et al. Robust Plasma Cell Response to Skin-Inoculated Dengue Virus in Mice. J. Immunol. Res. 2021, 2021, 5511841. [Google Scholar] [CrossRef]
- Rohrbach, M.S.; Wheatley, C.L.; Slifman, N.R.; Gleich, G.J. Activation of Platelets by Eosinophil Granule Proteins. J. Exp. Med. 1990, 172, 1271–1274. [Google Scholar] [CrossRef] [PubMed]
- Kunder, M.; Lakshmaiah, V.; Moideen Kutty, A.V. Plasma Neutrophil Elastase, A1-Antitrypsin, A2-Macroglobulin and Neutrophil Elastase–A1-Antitrypsin Complex Levels in Patients with Dengue Fever. Indian J. Clin. Biochem. 2018, 33, 218–221. [Google Scholar] [CrossRef]
- Sahin, A.T.; Yurtseven, A.; Dadmand, S.; Ozcan, G.; Akarlar, B.A.; Kucuk, N.E.O.; Senturk, A.; Ergonul, O.; Can, F.; Tuncbag, N.; et al. Plasma Proteomics Identify Potential Severity Biomarkers from COVID-19 Associated Network. PROTEOMICS – Clin. Appl. 2023, 17, 2200070. [Google Scholar] [CrossRef]
- Kimura, Y.; Nakai, Y.; Shin, J.; Hara, M.; Takeda, Y.; Kubo, S.; Jeremiah, S.S.; Ino, Y.; Akiyama, T.; Moriyama, K.; et al. Identification of Serum Prognostic Biomarkers of Severe COVID-19 Using a Quantitative Proteomic Approach. Sci. Rep. 2021, 11, 20638. [Google Scholar] [CrossRef] [PubMed]
- di Flora, D.C.; Dionizio, A.; Pereira, H.A.B.S.; Garbieri, T.F.; Grizzo, L.T.; Dionisio, T.J.; Leite, A. de L. ; Silva-Costa, L.C.; Buzalaf, N.R.; Reis, F.N.; et al. Analysis of Plasma Proteins Involved in Inflammation, Immune Response/Complement System, and Blood Coagulation upon Admission of COVID-19 Patients to Hospital May Help to Predict the Prognosis of the Disease. Cells 2023, 12, 1601. [Google Scholar] [CrossRef]
- Padwad, Y.S.; Mishra, K.P.; Jain, M.; Chanda, S.; Karan, D.; Ganju, L. RNA Interference Mediated Silencing of Hsp60 Gene in Human Monocytic Myeloma Cell Line U937 Revealed Decreased Dengue Virus Multiplication. Immunobiology 2009, 214, 422–429. [Google Scholar] [CrossRef] [PubMed]
- Vollenberg, R.; Tepasse, P.-R.; Fobker, M.; Hüsing-Kabar, A. Significantly Reduced Retinol Binding Protein 4 (RBP4) Levels in Critically Ill COVID-19 Patients. Nutrients 2022, 14, 2007. [Google Scholar] [CrossRef]
- Bramer, L.M.; Hontz, R.D.; Eisfeld, A.J.; Sims, A.C.; Kim, Y.-M.; Stratton, K.G.; Nicora, C.D.; Gritsenko, M.A.; Schepmoes, A.A.; Akasaka, O.; et al. Multi-Omics of NET Formation and Correlations with CNDP1, PSPB, and L-Cystine Levels in Severe and Mild COVID-19 Infections. Heliyon 2023, 9, e13795. [Google Scholar] [CrossRef]
- Soe, H.J.; Yong, Y.K.; Al-Obaidi, M.M.J.; Raju, C.S.; Gudimella, R.; Manikam, R.; Sekaran, S.D. Identifying Protein Biomarkers in Predicting Disease Severity of Dengue Virus Infection Using Immune-Related Protein Microarray. Medicine (Baltimore) 2018, 97, e9713. [Google Scholar] [CrossRef] [PubMed]
- Upasani, V.; Ter Ellen, B.M.; Sann, S.; Lay, S.; Heng, S.; Laurent, D.; Ly, S.; Duong, V.; Dussart, P.; Smit, J.M.; et al. Characterization of Soluble TLR2 and CD14 Levels during Acute Dengue Virus Infection. Heliyon 2023, 9, e17265. [Google Scholar] [CrossRef]
- Yin, G.-Q.; Zeng, H.-X.; Li, Z.-L.; Chen, C.; Zhong, J.-Y.; Xiao, M.-S.; Zeng, Q.; Jiang, W.-H.; Wu, P.-Q.; Zeng, J.-M.; et al. Differential Proteomic Analysis of Children Infected with Respiratory Syncytial Virus. Braz. J. Med. Biol. Res. 2021, 54, e9850. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.-H.; Liu, C.-C.; Wang, S.-T.; Lei, H.-Y.; Liu, H.-S.; Lin, Y.-S.; Wu, H.-L.; Yeh, T.-M. Activation of Coagulation and Fibrinolysis during Dengue Virus Infection. J. Med. Virol. 2001, 63, 247–251. [Google Scholar] [CrossRef]
- Abraham, R.; Mudaliar, P.; Jaleel, A.; Srikanth, J.; Sreekumar, E. High Throughput Proteomic Analysis and a Comparative Review Identify the Nuclear Chaperone, Nucleophosmin among the Common Set of Proteins Modulated in Chikungunya Virus Infection. J. Proteomics 2015, 120, 126–141. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.-L.; Lin, C.-L.; Chen, Y.-C.; Lu, I.-A.; Su, B.-H.; Chen, Y.-H.; Liu, K.-T.; Wu, C.-L.; Shiau, A.-L. Prothymosin α Accelerates Dengue Virus-Induced Thrombocytopenia. iScience 2024, 27, 108422. [Google Scholar] [CrossRef] [PubMed]
- Riswari, S.F.; Tunjungputri, R.N.; Kullaya, V.; Garishah, F.M.; Utari, G.S.R.; Farhanah, N.; Overheul, G.J.; Alisjahbana, B.; Gasem, M.H.; Urbanus, R.T.; et al. Desialylation of Platelets Induced by Von Willebrand Factor Is a Novel Mechanism of Platelet Clearance in Dengue. PLOS Pathog. 2019, 15, e1007500. [Google Scholar] [CrossRef] [PubMed]
- Trugilho, M.R. de O. ; Hottz, E.D.; Brunoro, G.V.F.; Teixeira-Ferreira, A.; Carvalho, P.C.; Salazar, G.A.; Zimmerman, G.A.; Bozza, F.A.; Bozza, P.T.; Perales, J. Platelet Proteome Reveals Novel Pathways of Platelet Activation and Platelet-Mediated Immunoregulation in Dengue. PLoS Pathog. 2017, 13, e1006385. [Google Scholar] [CrossRef]
- Pouysségur, J.; Marchiq, I.; Parks, S.K.; Durivault, J.; Ždralević, M.; Vucetic, M. ‘Warburg Effect’ Controls Tumor Growth, Bacterial, Viral Infections and Immunity – Genetic Deconstruction and Therapeutic Perspectives. Semin. Cancer Biol. 2022, 86, 334–346. [Google Scholar] [CrossRef]
- Panes, O.; Matus, V.; Sáez, C.G.; Quiroga, T.; Pereira, J.; Mezzano, D. Human Platelets Synthesize and Express Functional Tissue Factor. Blood 2007, 109, 5242–5250. [Google Scholar] [CrossRef]
- García-Larragoiti, N.; Kim, Y.C.; López-Camacho, C.; Cano-Méndez, A.; López-Castaneda, S.; Hernández-Hernández, D.; Vargas-Ruiz, Á.G.; Vázquez-Garcidueñas, M.S.; Reyes-Sandoval, A.; Viveros-Sandoval, M.E. Platelet Activation and Aggregation Response to Dengue Virus Non-structural Protein 1 and Domains. J. Thromb. Haemost. JTH 2021, 19, 2572–2582. [Google Scholar] [CrossRef] [PubMed]
- Ojha, A.; Nandi, D.; Batra, H.; Singhal, R.; Annarapu, G.K.; Bhattacharyya, S.; Seth, T.; Dar, L.; Medigeshi, G.R.; Vrati, S.; et al. Platelet Activation Determines the Severity of Thrombocytopenia in Dengue Infection. Sci. Rep. 2017, 7, 41697. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Bisht, P.; Bhattacharya, S.; Guchhait, P. Role of Platelet Cytokines in Dengue Virus Infection. Front. Cell. Infect. Microbiol. 2020, 10, 561366. [Google Scholar] [CrossRef]
- Esposito, S.; D’Abrosca, G.; Antolak, A.; Pedone, P.V.; Isernia, C.; Malgieri, G. Host and Viral Zinc-Finger Proteins in COVID-19. Int. J. Mol. Sci. 2022, 23, 3711. [Google Scholar] [CrossRef] [PubMed]
- Dengue - OPS/OMS | Organización Panamericana de la Salud. Available online: https://www.paho.org/es/arbo-portal/dengue (accessed on 9 January 2025).
Figure 1.
Enrichment analysis of the biological processes in the evaluated groups. DV (Dengue virus infection).
Figure 1.
Enrichment analysis of the biological processes in the evaluated groups. DV (Dengue virus infection).
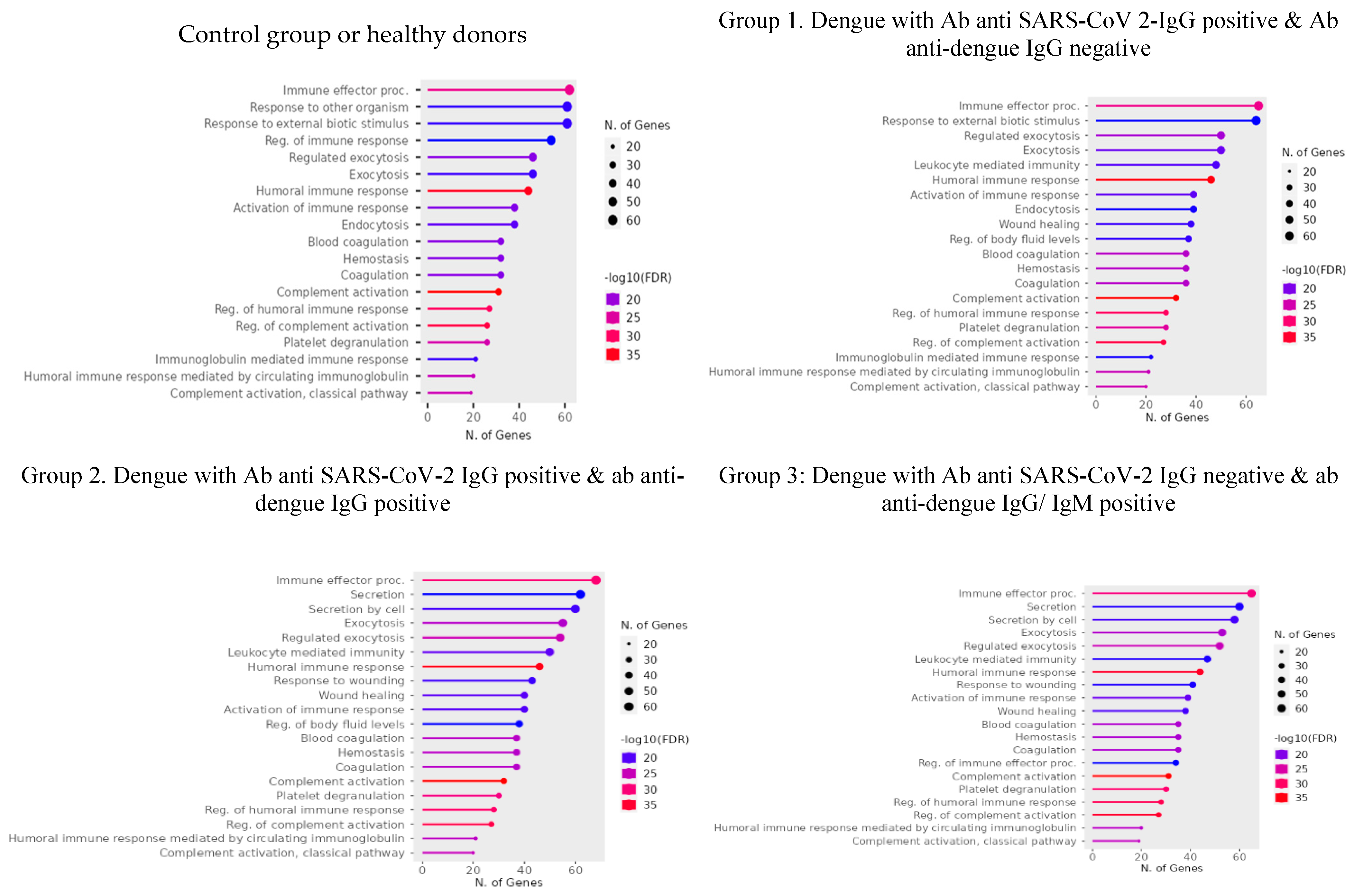

Table 1.
Serological and Molecular Profiles of Study Participants Across Groups Based on Anti-SARS-CoV-2 and Anti-dengue Antibody Status and Dengue Diagnostic Markers.
Table 1.
Serological and Molecular Profiles of Study Participants Across Groups Based on Anti-SARS-CoV-2 and Anti-dengue Antibody Status and Dengue Diagnostic Markers.
| Group | ID | Ab anti SARS-CoV-2 IgG | Ab anti dengue IgG | Ab anti dengue IgM | Dengue NS1 | Dengue RT PCR cT |
|---|---|---|---|---|---|---|
| Control Group | 1 | Pos | Neg | Neg | Neg | - |
| 2 | Pos | Pos | Neg | Neg | - | |
| 3 | Pos | Pos | Neg | Neg | - | |
| Group 1. SARS-CoV 2-IgG & Dengue NS1 | 5162 | Pos | Neg | Neg | Pos | 23.2 |
| 4810 | Pos | Neg | Neg | Pos | 19.3 | |
| 4658 | Pos | Neg | Neg | Pos | 27 | |
| 4972 | Pos | Neg | Neg | Pos | 30.5 | |
| 5088 | Pos | Neg | Neg | Pos | 32.3 | |
| Group 2. SARS-CoV 2-IgG, Dengue NS1 & IgG | 5063 | Pos | Pos | Neg | Pos | 24.8 |
| 4947 | Pos | Pos | Pos | Pos | 30.7 | |
| 4548 | Pos | Pos | Neg | Pos | 25 | |
| 4375 | Pos | Pos | Pos | Pos | 26.2 | |
| 4452 | Pos | Pos | Neg | Pos | 26.6 | |
| 4556 | Pos | Pos | Neg | Pos | 23 | |
| Group 3. Dengue NS1 & IgG | 5098 | Neg | Pos | Neg | Pos | 32.9 |
| 5134 | Neg | Pos | Neg | Pos | 22.1 | |
| 4376 | Neg | Pos | Neg | Pos | 20.2 | |
| 5062 | Neg | Pos | Pos | Pos | 24.8 |
Table 2.
Differential Expression of Proteins (DEP) Across Groups of Interest. Each protein is listed alongside its UniProt ID, gene symbol, and functional description, highlighting their roles in the immune response, coagulation, and viral interactions.
Table 2.
Differential Expression of Proteins (DEP) Across Groups of Interest. Each protein is listed alongside its UniProt ID, gene symbol, and functional description, highlighting their roles in the immune response, coagulation, and viral interactions.
| Protein | Uniprot ID | Gen | Function |
|---|---|---|---|
| Control group of healthy donors | |||
| Syntaxin-binding protein 5 | Q5T5C0 | STXBP5 | STXBP5 inhibits endothelial exocytosis and promotes platelet secretion. In KO STXBP5 cells increase vWF & P-selectin secretion [21]. In platelets it is important for cargo release from dense granules, α granules, and lysosomes as well as for packaging of some cargo into granules [22]. |
| Galectin-1 | P09382 | LGALS1 | Gal-1 binds to DENV-1 and inhibits its adsorption and internalization processes in ECV-304 cells [23]. |
| Glucose-6-phosphate isomerase | P06744 | GPI | In the cytoplasm, catalyzes the conversion of glucose-6-phosphate to fructose-6-phosphate, the second step in glycolysis. |
| Group 1. Dengue with Ab anti SARS-CoV 2-IgG positive & Ab anti dengue IgG negative | |||
| Coagulation factor IX | P00740 | F9 | Participates in the intrinsic pathway of blood coagulation. Alteration of coagulation in dengue infection [24,25]. |
| Ribosomal protein L19 | P84098 | RPL19 | Component of the large ribosomal subunit. Ribosomal Protein L19 and L22 Modulate TLR3 Signaling [26]. RPL19 is required for efficient YFV and WNV protein production [27]. |
| Peptidyl-prolyl cis-trans isomerase B | P23284 | PPIB | Catalyze the isomerization of peptide bonds from trans to cis form at proline residues and facilitates protein folding. Knockdown of CYP isoforms reduces the replication of flavivirus VLPS [28]. |
| Group 2. Dengue with Ab anti SARS-CoV-2 IgG positive & ab anti dengue IgG positive | |||
| Thrombospondin-1 | P07996 | THBS1 | TSP1 is a major protein component of Alfa platelet-granules from which it is rapidly released during platelet activation. It stimulates platelet aggregation [29].TSP1 binds to A3 domain of vWF and may compete with ADAMTS13 for interaction with vWF [30]. |
| Colorectal mutant cancer protein | P23508 | MCC | Play role in cell proliferation. Colorectal Cancer Protein Modulates the NF-κB Pathway [31]. Knockdown of MCC induced apoptosis and inhibited proliferation in human MM cells [32]. |
| Coagulation factor XIII B chain | P05160 | F13B | The B chain of factor XIII is not catalytically active, but is thought to stabilize the A subunits. Alteration of coagulation in dengue infection [24,25]. |
| Antigen KI-67 | P46013 | MKI67 | Widely used as a marker to assess cell proliferation, as is detected in the nucleus of proliferating cells only. Expressed in plasmatic cells infected with dengue [33]. |
| Bone marrow proteoglycan | P13727 | PRG2 | Cytotoxin and helminthotoxin. It causes platelet activation [34]. |
| Group 3. Dengue with Ab anti SARS-CoV-2 IgG negative & ab anti dengue IgG/ IgM positive | |||
| Putative uncharacterized zinc finger protein 814 | B7Z6K7 | ZNF814 | Unknow function |
Table 3.
Serum Proteome Changes in Dengue Patients Across Diverse Groups of Interest. This table presents the abundance ratios and functional roles of various proteins identified in the sera of dengue patients, categorized into three distinct groups. Group 1 consisted of patients with positive anti-SARS-CoV-2 IgG and negative anti-dengue IgG; Group 2 included patients with both positive anti-SARS-CoV-2 IgG and anti-dengue IgG; and Group 3 comprised patients with negative anti-SARS-CoV-2 IgG and positive anti-dengue IgG/IgM. Bold data show that proteins with RA>2.
Table 3.
Serum Proteome Changes in Dengue Patients Across Diverse Groups of Interest. This table presents the abundance ratios and functional roles of various proteins identified in the sera of dengue patients, categorized into three distinct groups. Group 1 consisted of patients with positive anti-SARS-CoV-2 IgG and negative anti-dengue IgG; Group 2 included patients with both positive anti-SARS-CoV-2 IgG and anti-dengue IgG; and Group 3 comprised patients with negative anti-SARS-CoV-2 IgG and positive anti-dengue IgG/IgM. Bold data show that proteins with RA>2.
| Uniprot ID | Abundance ratio | Protein | Function | ||
| Group 1 | Group2 | Group 3 | |||
| P01023 | 0.67 | 2.49 | 0.28 | Alpha-2-macroglobulin | Antiprotease plays a role in hemostatic balance. Decreased levels during dengue infection as part of a normal immune response [35]. |
| P19823 | 0.55 | 2.10 | 0.63 | Inter-alpha-trypsin inhibitor heavy chain H2 | Decreased expression in patients with COVID 19 [36]. |
| P04217 | 2.70 | 2.66 | 2.44 | Alpha-1B-glycoprotein | Increased expression during dengue infection [18]. |
| P02750 | 2.28 | 2.18 | 2.51 | Leucine-rich-alpha-2glycoprotein* | Increased expression during dengue infection [18]. |
| P02748 | 2.04 | 4.75 | 0.91 | Complement component C9 | Elevated during dengue infection [18]. |
| P35542 | 2.17 | 0.90 | 1.01 | Serum amyloid A-4 protein | Unknow function in viral infection. |
| P02746 | 1.90 | 2.13 | 1.53 | Complement C1q subcomponent subunit B | First component of the serum complement system. |
| P35858 | 2.60 | 2.48 | 3.31 | Insulin-like growth factor-binding protein complex acid labile subunit | Involved in protein-protein interactions. Reduced levels in patients with severe COVID 19 [37]. |
| P04003 | 0.66 | 2.48 | 0.61 | C4b-binding protein alpha chain | Decreased in patients with severe COVID 19 [38]. |
| P03952 | 1.57 | 2.29 | 1.94 | Plasma kallikrein. | Participates in blood coagulation. |
| Q96PD5 | 1.56 | 1.41 | 2.24 | N-acetylmuramoyl-L-alanine amidase. | Decreased in patients with severe COVID 19 [38]. |
| P10643 | 2.39 | 4.36 | 3.43 | Complement component C7 | Constituent of the membrane attack complex (MAC). |
| P10809 | 1.19 | 3.09 | 0.14 | 60 kDa heat shock protein, mitochondrial | elevated levels of expression during dengue infection in U937 cells [39]. |
| Q96L50 | 7.51 | 4.20 | 8.03 | Leucine-rich repeat protein 1 | Involve in protein modification and ubiquitination. Unknow function in viral infection. |
| P02753 | 3.61 | 1.91 | 2.81 | Retinol-binding protein 4/ RBP4 | Low level in acute phase of patients with critical COVID 19 [40]. |
| P29622 | 0.86 | 3.01 | 1.92 | Kallistatin | Inhibits human kininogenase activities of tissue kallikrein. |
| P35269 | 3.70 | 0.54 | 1.25 | General transcription factor IIF subunit/ GTF2F1 | Promote transcription elongation. Unknow function in viral infection. |
| P02745 | 1.69 | 3.45 | 1.33 | Complement C1q subcomponent subunit A | C1q is associated with the proenzymes C1r and C1s to yield C1. |
| P02741 | 2.10 | 6.97 | 0.00 | C reactive protein | Elevated during acute phase in patients with DHF [18]. |
| Q96KN2 | 2.59 | 0.52 | 1.03 | Beta-Ala-His dipeptidase/ CNDP1 | Up regulation in mild COVID patients as a protective mechanism of oxidative stress [41]. |
| O75882 | 2.41 | 1.13 | 0.61 | Attractin | Involved in the initial immune cell clustering during inflammatory response and may regulate chemotactic activity of chemokines. Unknow function in viral infection |
| Q9NZP8 | 2.08 | 2.39 | 1.25 | Complement C1r subcomponent-like protein | Mediates the proteolytic cleavage of HP/haptoglobin in the endoplasmic reticulum. |
| P09493 | 2.48 | 2.69 | 2.66 | Tropomyosin alpha-1 chain/TPM1 | Binds to actin filaments. Hight levels in patients with severe dengue [42]. |
| P08571 | 6.01 | 4.27 | 4.22 | Monocyte differentiation antigen CD14 | Elevated levels in plasma during acute phase of dengue fever [43]. |
| P07195 | 2.24 | 2.18 | 1.89 | L-lactate dehydrogenase B chain | Interconverts simultaneously and stereospecifically pyruvate and lactate. Unknown function in viral infection. |
| P60174 | 1.78 | 2.84 | 2.05 | Triosephosphate isomerase/ TPI1 | Participation in glycolysis and gluconeogenesis. Up regulated during acute phase of infection with RSV [44]. |
| P02679 | 0.69 | 0.95 | 2.06 | Fibrinogen gamma chain/ FGG | Major function in hemostasis. Alteration of coagulation in dengue infection[45]. |
| P43251 | 1.71 | 3.44 | 2.47 | Biotinidase/ BTD | Catalytic release of biotin from biocytin. Unknow function during viral infection. |
| P06748 | 4.46 | 6.20 | 3.16 | Nucleophosmin/ NPM1 | Involved in diverse cellular processes. Enhanced expression in HEK293cells infected with CHIKV [46]. |
| P06454 | 3.89 | 5.17 | 5.75 | Prothymosin alpha/ PTMA | May mediate immune function by conferring resistance to certain opportunistic infections. DENV infection upregulates PTMA expression, leading to suppressing megakaryopoiesis. Plasma levels are elevated in dengue patients and DENV-infected megakaryoblasts [47]. |
| Proteins with AR>2 | 18 | 23 | 14 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



