Submitted:
22 January 2025
Posted:
24 January 2025
Read the latest preprint version here
Abstract
Assessing cognitive abilities is crucial in educational contexts to inform student selection processes. Presently, academic metrics are widely used for grading, evaluation, selection and placement decisions. This study investigates the correlation between P300 latency, reaction time, and fluid intelligence in children, utilizing Raven’s Standard Progressive Matrices (RSPM) for intelligence measurement. Participants were divided into two groups based on their RSPM scores, reflecting "high mental abilities" and "average mental abilities." We hypothesized that children with higher RSPM scores would demonstrate shorter P300 latency and faster reaction times, indicative of more efficient cognitive processing. Electrophysiological data were collected through Event Related Potentials (ERPs), specifically analyzing the P300 component. Results confirmed that higher intelligence is associated with shorter P300 latencies and faster reaction times, supporting theories of neural efficiency and cognitive speed's role in intelligence. This study enhances understanding of the neurophysiological correlates of intelligence in children and informs educational strategies tailored to individual cognitive profiles.
Keywords:
1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Electrophysiological Assessment
2.2.1. Electrode Placement and Data Recording
2.2.2. P300 Component Detection
2.2.3. Data Preprocessing and Artifact Removal
2.3. Implementation
2.4. Statistical Analysis
3. Results
4. Discussion
4.1. Psychoeducational Implications of the Study Using ERPs and RSPM Results in Identifying Children’s Mental Abilities
4.2. Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, T., Ren, X., Altmeyer, M., & Schweizer, K. (2013). An account of the relationship between fluid intelligence and complex learning in considering storage capacity and executive attention. Intelligence, 41(5), 537-545. [CrossRef]
- Amin, H. U., Malik, A. S., Kamel, N., Chooi, W. T., & Hussain, M. (2015). P300 correlates with learning & memory abilities and fluid intelligence. Journal of neuroengineering and rehabilitation, 12, 1-14. [CrossRef]
- Neubauer, A. C., & Fink, A. (2009). Intelligence and neural efficiency. Neuroscience & Biobehavioral Reviews, 33(7), 1004-1023.
- Liu, T., Xiao, T., Shi, J., Zhao, D., & Liu, J. (2011). Conflict control of children with different intellectual levels: an ERP study. Neuroscience Letters, 490(2), 101-106. [CrossRef]
- Raven, J. C. (1938). Raven standard progressive matrices. Journal of Cognition and Development.
- Raven, J. (2008). The Raven progressive matrices tests: their theoretical basis and measurement model. Uses and abuses of Intelligence. Studies advancing Spearman and Raven’s quest for non-arbitrary metrics, (Part I).
- Duncan, J., Chylinski, D., Mitchell, D. J., & Bhandari, A. (2017). Complexity and compositionality in fluid intelligence. Proceedings of the National Academy of Sciences, 114(20), 5295-5299. [CrossRef]
- Tanaka, F., Kachi, T., Yamada, T., & Sobue, G. (1998). Auditory and visual event-related potentials and flash visual evoked potentials in Alzheimer's disease: correlations with Mini-Mental State Examination and Raven's Coloured Progressive Matrices. Journal of the neurological sciences, 156(1), 83-88.
- Jung, R. E., & Haier, R. J. (2007). The Parieto-Frontal Integration Theory (P-FIT) of intelligence: converging neuroimaging evidence. Behavioral and brain sciences, 30(2), 135-154. [CrossRef]
- Zacks, J. M. (2008). Neuroimaging studies of mental rotation: a meta-analysis and review. Journal of cognitive neuroscience, 20(1), 1-19.
- Haier, R. J., Siegel, B., Tang, C., Abel, L., & Buchsbaum, M. S. (1992). Intelligence and changes in regional cerebral glucose metabolic rate following learning. Intelligence, 16(3-4), 415-426. [CrossRef]
- Neubauer, A. C., Grabner, R. H., Fink, A., & Neuper, C. (2005). Intelligence and neural efficiency: Further evidence of the influence of task content and sex on the brain–IQ relationship. Cognitive Brain Research, 25(1), 217-225. [CrossRef]
- Zygouris, N. C. (2024). Differences in children and adolescents with depression before and after a remediation program: an event-related potential study. Brain sciences, 14(7), 660. [CrossRef]
- Karapetsas, A. V., & Zygouris, N. C. (2011). Event Related Potentials (ERPs) in prognosis, diagnosis and rehabilitation of children with dyslexia. Encephalos, 48(3), 118-127.
- Howe, A. S., Bani-Fatemi, A., & De Luca, V. (2014). The clinical utility of the auditory P300 latency subcomponent event-related potential in preclinical diagnosis of patients with mild cognitive impairment and Alzheimer’s disease. Brain and cognition, 86, 64-74. [CrossRef]
- Demirayak, P., Kıyı, İ., İşbitiren, Y. Ö., & Yener, G. (2023). Cognitive load associates prolonged P300 latency during target stimulus processing in individuals with mild cognitive impairment. Scientific Reports, 13(1), 15956. [CrossRef]
- Zhong, R., Li, M., Chen, Q., Li, J., Li, G., & Lin, W. (2019). The P300 event-related potential component and cognitive impairment in epilepsy: a systematic review and meta-analysis. Frontiers in neurology, 10, 943. [CrossRef]
- Sternberg, R. J. (Ed.). (2020). The Cambridge handbook of intelligence. Cambridge University Press.
- Walhovd, K. B., Nyberg, L., Lindenberger, U., Amlien, I. K., Sørensen, Ø. et al. (2022). Brain aging differs with cognitive ability regardless of education. Scientific reports, 12(1), 13886. [CrossRef]
- Warchoł, Ł., & Zając-Lamparska, L. (2023). The Relationship of N200 and P300 Amplitudes With Intelligence, Working Memory, and Attentional Control Behavioral Measures In Young Healthy Individuals. Advances in Cognitive Psychology, 19(4). [CrossRef]
- Walhovd, K. B., & Fjell, A. M. (2002). One-year test–retest reliability of auditory ERPs in young and old adults. International Journal of Psychophysiology, 46(1), 29-40. [CrossRef]
- Polich, J. (2011). Neuropsychology of P300. In E. S. Kappenman & S. J. Luck (Eds.), The Oxford handbook of event-related potential components (pp. 160-188). Oxford University Press.
- Sternberg, R. J. (2019). A theory of adaptive intelligence and its relation to general intelligence. Journal of Intelligence, 7(4), 23. [CrossRef]
- Lozano-Blasco, R., Quílez-Robres, A., Usán, P., Salavera, C., & Casanovas-López, R. (2022). Types of intelligence and academic performance: A systematic review and meta-analysis. Journal of Intelligence, 10(4), 123. [CrossRef]
- Ren, X., Schweizer, K., Wang, T., Chu, P., & Gong, Q. (2017). On the relationship between executive functions of working memory and components derived from fluid intelligence measures. Acta Psychologica, 180, 79-87. [CrossRef]
- Ren, X., Wang, T., Sun, S., Deng, M., & Schweizer, K. (2018). Speeded testing in the assessment of intelligence gives rise to a speed factor. Intelligence, 66, 64-71. [CrossRef]
- Bazana, P. G., & Stelmack, R. M. (2002). Intelligence and information processing during an auditory discrimination task with backward masking: an event-related potential analysis. Journal of personality and social psychology, 83(4), 998.
- De Pascalis, V. A., Varriale, V., & Matteoli, A. (2008). Intelligence and P3 components of the event-related potential elicited during an auditory discrimination task with masking. Intelligence, 36(1), 35-47. [CrossRef]
- Troche, S. J., Houlihan, M. E., Stelmack, R. M., & Rammsayer, T. H. (2009). Mental ability, P300, and mismatch negativity: Analysis of frequency and duration discrimination. Intelligence, 37(4), 365-373. [CrossRef]
- Teixeira-Santos, A. C., Pinal, D., Pereira, D. R., Leite, J., Carvalho, S., & Sampaio, A. (2020). Probing the relationship between late endogenous ERP components with fluid intelligence in healthy older adults. Scientific Reports, 10(1), 11167. [CrossRef]
- Polich, J. (2007). Updating P300: an integrative theory of P3a and P3b. Clinical neurophysiology, 118(10), 2128-2148. [CrossRef]
- Der, G., & Deary, I. J. (2017). The relationship between intelligence and reaction time varies with age: Results from three representative narrow-age age cohorts at 30, 50 and 69 years. Intelligence, 64, 89-97. [CrossRef]
- Kannen, K., Aslan, B., Boetzel, C., Herrmann, C. S., Lux, S. et al. (2022). P300 modulation via transcranial alternating current stimulation in adult attention-deficit/hyperactivity disorder: a crossover study. Frontiers in Psychiatry, 13, 928145. [CrossRef]
- Jungeblut, H. M., Hagemann, D., Löffler, C., & Schubert, A. L. (2021). An investigation of the slope parameters of reaction times and P3 latencies in the Sternberg memory scanning task–A fixed-links model approach. Journal of Cognition, 4(1). [CrossRef]
- Schubert, A. L., Löffler, C., Hagemann, D., & Sadus, K. (2023). How robust is the relationship between neural processing speed and cognitive abilities? Psychophysiology, 60(2), e14165.
- Schubert, A. L. (2019). A meta-analysis of the worst performance rule. Intelligence, 73, 88-100. [CrossRef]
- Regel, S., Meyer, L., & Gunter, T. C. (2014). Distinguishing neurocognitive processes reflected by P600 effects: Evidence from ERPs and neural oscillations. PloS one, 9(5), e96840. [CrossRef]
- Beldzik, E., & Ullsperger, M. (2024). A thin line between conflict and reaction time effects on EEG and fMRI brain signals. Imaging Neuroscience, 2, 1-17. [CrossRef]
- Raven, J. (2003). Raven progressive matrices. In Handbook of nonverbal assessment (pp. 223-237). Boston, MA: Springer US.
- Jasper, H. H. (1958). Ten-twenty electrode system of the international federation. Electroencephalogr Clin Neurophysiol, 10, 371-375.
- Sadus, K., Schubert, A. L., Löffler, C., & Hagemann, D. (2024). An explorative multiverse study for extracting differences in P3 latencies between young and old adults. Psychophysiology, 61(2), e14459. [CrossRef]
- Picton, T. W., Bentin, S., Berg, P., Donchin, E., Hillyard, S. A., Johnson, R., et al., (2000). Guidelines for using human event-related potentials to study cognition: recording standards and publication criteria. Psychophysiology, 37(2), 127-152. [CrossRef]
- Zygouris, N. C., Avramidis, E., Karapetsas, A. V., & Stamoulis, G. I. (2018). Differences in dyslexic students before and after a remediation program: A clinical neuropsychological and event related potential study. Applied Neuropsychology: Child, 7(3), 235-244. [CrossRef]
- Zygouris, N. C., Vlachos, F., & Stamoulis, G. I. (2022). ERPs in Children and Adolescents with Generalized Anxiety Disorder: Before and after an Intervention Program. Brain Sciences, 12(9), 1174. [CrossRef]
- Cohen, J. (1988). Set correlation and contingency tables. Applied psychological measurement, 12(4), 425-434. [CrossRef]
- Beauchamp, C. M., & Stelmack, R. M. (2006). The chronometry of mental ability: An event-related potential analysis of an auditory oddball discrimination task. Intelligence, 34(6), 571-586. [CrossRef]
- Wongupparaj, P., Sumich, A., Wickens, M., Kumari, V., & Morris, R. G. (2018). Individual differences in working memory and general intelligence indexed by P200 and P300: A latent variable model. Biological psychology, 139, 96-105. [CrossRef]
- McGarry-Roberts, P. A., Stelmack, R. M., & Campbell, K. B. (1992). Intelligence, reaction time, and event-related potentials. Intelligence, 16(3-4), 289-313.
- Sur, S., & Sinha, V. K. (2009). Event-related potential: An overview. Industrial psychiatry journal, 18(1), 70-73. [CrossRef]
- Gmaj, B., Januszko, P., Kamiński, J., Drozdowicz, E., Kopera, M., Wołyńczyk-Gmaj, D., & Wojnar, M. (2016). EEG source activity during processing of neutral stimuli in subjects with anxiety disorders. Acta Neurobiologiae Experimentalis, 76(1), 75-85. [CrossRef]
- Doebler, P., & Scheffler, B. (2016). The relationship of choice reaction time variability and intelligence: A meta-analysis. Learning and Individual Differences, 52, 157-166. [CrossRef]
- Tsai, Y. C., Lu, H. J., Chang, C. F., Liang, W. K., Muggleton, N. G., & Juan, C. H. (2017). Electrophysiological and behavioral evidence reveals the effects of trait anxiety on contingent attentional capture. Cognitive, Affective, & Behavioral Neuroscience, 17, 973-983. [CrossRef]
- Jensen, A. R. (1998). The factor. Westport, CT: Prager.
- Sanz, M., Molina, V., Martin-Loeches, M., Calcedo, A., & Rubia, F. J. (2001). Auditory P300 event related potential and serotonin reuptake inhibitor treatment in obsessive-compulsive disorder patients. Psychiatry research, 101(1), 75-81. [CrossRef]
- Deary, I. J., Cox, S. R., & Okely, J. A. (2024). Inspection time and intelligence: A five-wave longitudinal study from age 70 to age 82 in the Lothian Birth Cohort 1936. Intelligence, 105, 101844. [CrossRef]
- Coles, M. G. H., Smid, H. G. O. M., Scheffers, M. K., & Otten, L. J. (1995). Mental Chronometry and the study of human information processing. In Electrophysiology of Mind. (pp. 86-127). Oxford University Press.
- Shaw, P., Greenstein, D., Lerch, J., Clasen, L., Lenroot, R., et al. (2006). Intellectual ability and cortical development in children and adolescents. Nature, 440(7084), 676-679. [CrossRef]
- Kutas, M., & Federmeier, K. D. (2011). Thirty years and counting: finding meaning in the N400 component of the event-related brain potential (ERP). Annual review of psychology, 62, 621-647. [CrossRef]
- Wronka, E., Kaiser, J., & Coenen, A. M. (2013). Psychometric intelligence and P3 of the event-related potentials studied with a 3-stimulus auditory oddball task. Neuroscience Letters, 535, 110-115. [CrossRef]
- Luck, S. J. (2014). An introduction to the event-related potential technique. MIT press.
- Zygouris, N. C., Dermitzaki, I., & Karapetsas, A. V. (2015). Differences in brain activity of children with higher mental abilities. An Event Related Potentials study using the latency of P300 and N100 waveforms. International Journal of Developmental Neuroscience, (47), 118-119. [CrossRef]
- Gagné, F. (2005). From gifts to talents: The DMGT as a devalopmental model. U: RJ Sternberg & JE Davidson (Eds.), Conceptions of giftedness (str. 98-120).
- Greer, K. (2022). Neural Assemblies as Precursors for Brain Function. NeuroSci, 3(4), 645-655. [CrossRef]
- Liu, X., Yang, S., & Liu, Z. (2021). Predicting Fluid Intelligence via Naturalistic Functional Connectivity Using Weighted Ensemble Model and Network Analysis. NeuroSci, 2(4), 427-442. [CrossRef]

| Electro/ Encephalographic Sites |
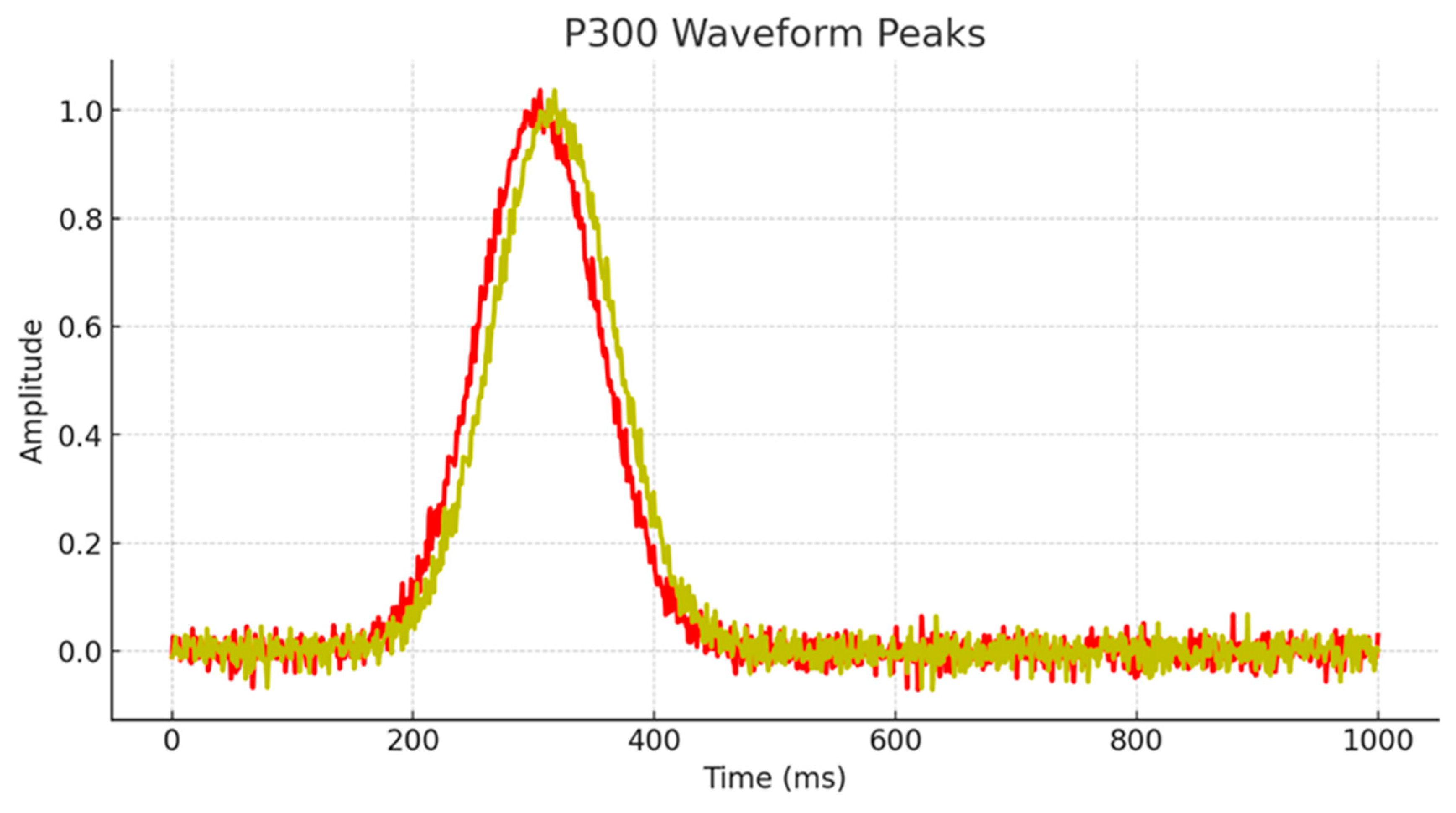
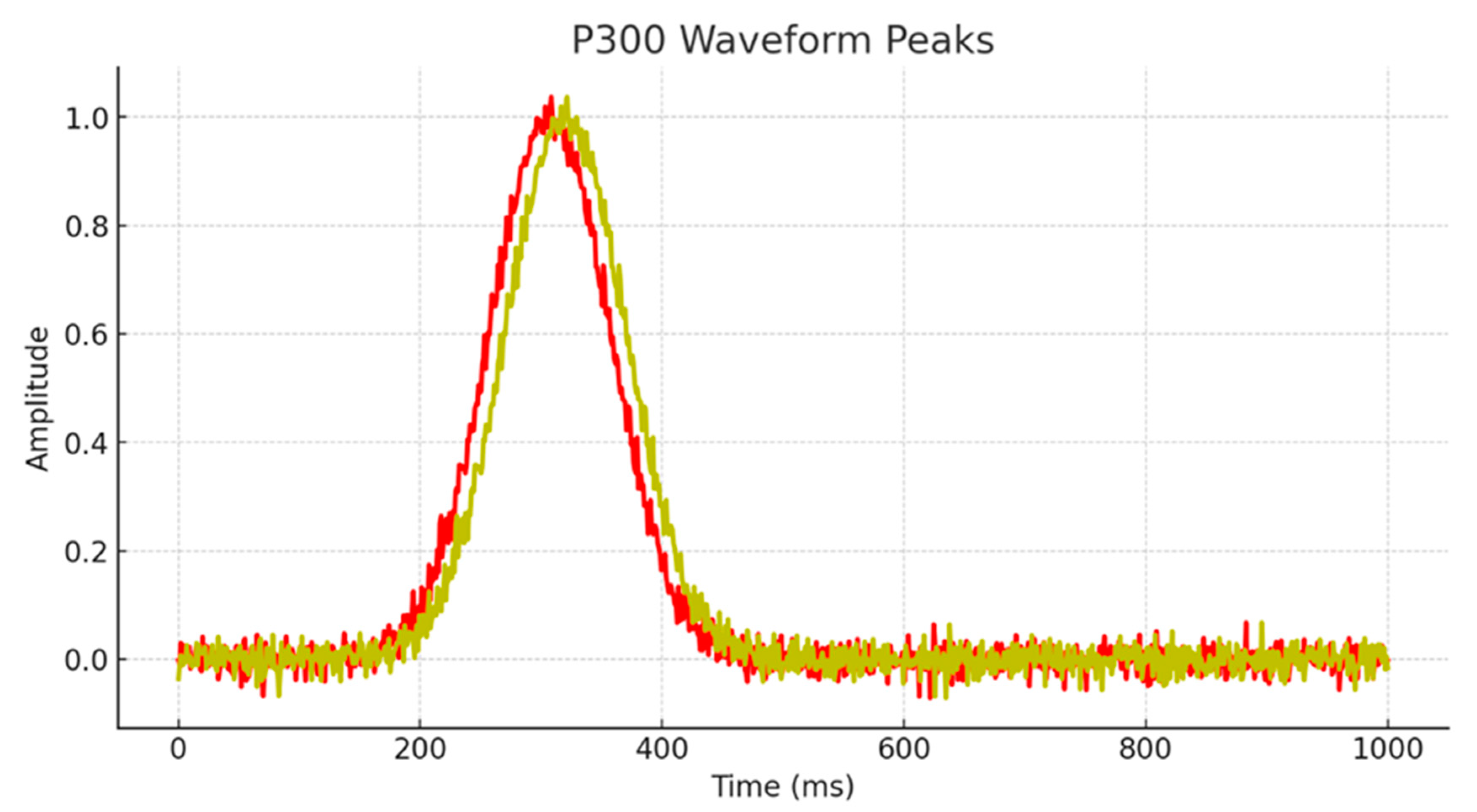
P300 Latency of Children with high mental abilities |
SD | P300 Latency of Children with average mental abilities |
SD | F | p | Cohen's d |
|---|---|---|---|---|---|---|---|
| Fp1 | 304.48 | 6.21 | 316.58 | 3.46 | 34.80 | <0.001 | 2.41 |
| FPz | 305.44 | 7.64 | 318.41 | 4.63 | 25.33 | <0.001 | 2.05 |
| Fp2 | 307.55 | 6.33 | 320.53 | 5.92 | 26.89 | <0.001 | 2.12 |
| F3 | 307.00 | 7.14 | 326.65 | 7.56 | 42.84 | <0.001 | 2.67 |
| Fz | 307.87 | 6.05 | 325.53 | 2.21 | 90.16 | <0.001 | 3.88 |
| F4 | 313.15 | 10.31 | 336.38 | 1.99 | 58.74 | <0.001 | 3.13 |
| T3 | 306.28 | 10.03 | 329.00 | 4.82 | 50.03 | <0.001 | 2.89 |
| T4 | 308.32 | 11.85 | 325.85 | 2.45 | 25.16 | <0.001 | 2.05 |
| C3 | 306.78 | 13.10 | 330.45 | 6.37 | 31.67 | <0.001 | 2.30 |
| Cz | 311.69 | 15.84 | 337.53 | 3.22 | 30.65 | <0.001 | 2.26 |
| C4 | 317.77 | 9.41 | 338.19 | 2.89 | 51.63 | <0.001 | 2.93 |
| P3 | 309.51 | 10.04 | 329.96 | 5.21 | 39.19 | <0.001 | 2.56 |
| Pz | 311.98 | 14.98 | 336.37 | 5.45 | 28.10 | <0.001 | 2.16 |
| P4 | 313.39 | 17.56 | 339.68 | 3.67 | 25.79 | <0.001 | 2.07 |
| Oz | 316.91 | 10.38 | 337.87 | 5.71 | 37.58 | <0.001 | 2.50 |
| Electro/ Encephalographic Sites |
Lower Bound (p) | Upper Bound (p) |
|---|---|---|
| Fp1 | 6.17 × 10⁻⁶ | 9.26 × 10⁻⁵ |
| FPz | 4.87 × 10⁻⁵ | 7.30 × 10⁻⁴ |
| Fp2 | 3.37 × 10⁻⁵ | 5.05 × 10⁻⁴ |
| F3 | 1.39 × 10⁻⁶ | 2.09 × 10⁻⁵ |
| Fz | 3.07 × 10⁻⁹ | 4.61 × 10⁻⁸ |
| F4 | 1.19 × 10⁻⁷ | 1.79 × 10⁻⁶ |
| T3 | 4.28 × 10⁻⁷ | 6.42 × 10⁻⁶ |
| T4 | 5.07 × 10⁻⁵ | 7.60 × 10⁻⁴ |
| C3 | 1.17 × 10⁻⁵ | 1.75 × 10⁻⁴ |
| Cz | 1.45 × 10⁻⁵ | 2.18 × 10⁻⁴ |
| C4 | 3.34 × 10⁻⁷ | 5.02 × 10⁻⁶ |
| P3 | 2.66 × 10⁻⁶ | 3.99 × 10⁻⁵ |
| Pz | 2.55 × 10⁻⁵ | 3.83 × 10⁻⁴ |
| P4 | 4.36 × 10⁻⁵ | 6.54 × 10⁻⁴ |
| Oz | 3.60 × 10⁻⁶ | 5.40 × 10⁻⁵ |
| Reaction Time | High Mental Abilities | Average Mental Abilities | |||||
|---|---|---|---|---|---|---|---|
| M | SD | M | SD | F | p | Cohen’s d | |
| 319.70 | 6.54 | 352.30 | 11.76 | 70.42 | <0.001 | 3.43 | |
| Electro/ encephalographic sites |
Correlation ρ | Sign |
|---|---|---|
| FP1 | -0.844 | 0.001 |
| FPZ | -0.804 | 0.001 |
| FP2 | -0.742 | 0.001 |
| F3 | -0.862 | 0.001 |
| FZ | -0.889 | 0.001 |
| F4 | -0.813 | 0.001 |
| T3 | -0.818 | 0.001 |
| T4 | -0.893 | 0.001 |
| C3 | -0.844 | 0.001 |
| CZ | -0.865 | 0.001 |
| C4 | -0.770 | 0.001 |
| P3 | -0.853 | 0.001 |
| PZ | -0.803 | 0.001 |
| P4 | -0.790 | 0.001 |
| OZ | -0.781 | 0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).



