Submitted:
22 January 2025
Posted:
23 January 2025
You are already at the latest version
Abstract
The role of the sand prawn, Callichirus kraussi (Stebbing, 1900), as an ecosystem engineer was evaluated using a three-month-long caging experiment during a prolonged drought in the lower reaches of the temporarily open/closed Kasouga Estuary along the eastern seaboard of South Af-rica. The current study indicates that at intermediate densities, the burrowing activities of the sand prawn contributed to a significant increase in the microphytobenthic algal concentrations and a concurrent increase in the abundance and biomass of the macrofauna (P < 0.05 in all cases). The increase in macrofauna abundance and biomass could largely be ascribed to the elevated densities of the gastropod, Nassarius kraussianus which were sustained by the elevated microphytobenthic biomass. As such, the sand prawn Callichirus kraussi was established to be an important ecosystem engineer as the results indicate that intermediate densities of the sand prawn may locally con-tribute to increases in the biology even during those periods when the system is stressed, in this case, the prolonged drought.
Keywords:
Ecosystem engineers
; bioturbation
; sand prawn
; drought
1. Introduction
Ecosystem engineers are organisms that change the state of matter, both biotic and abiotic, by altering their environments through bioturbation [1,2,3,4]. The mixing and alteration of sediment structure in coastal habitats is thought to be the primary drivers of biodiversity as they have a fundamental effect on the environment [5]. Bioturbation facilitates the dispersal of particles in sediments, modifies geochemical cycling alters food sources, and alleviates the transport of water and gaseous exchange in soils [6]. Because bioturbators modify the availability of resources to other species, they are viewed as ecosystem engineers [5]. Bioturbation, therefore, affects the physical (by constructing burrows, mound construction, ploughing of the surface, and trampling) and chemical (the mixing of water and solutes) characteristics of soft-sediment habitats [7,8] which promotes food availability, species recruitment, increases oxygenation and mineralization and modifies the distribution of microalgae and species composition of macrofaunal species [2,4,9,10,11,12].
The South African coastline is numerically dominated by temporarily open/closed estuaries (TOCEs) that under natural conditions, are periodically separated from the marine environment by a sandbar at the mouth [13,14,15,16]. The duration of this separation is largely a function of rainfall within the catchment area and the extent of the sandbar [17]. During periods of drought, TOCEs may be separated from the marine environment for periods exceeding 12 months [13,16]. Mouth status plays an important role in determining the physical properties of the water column and consequently, the biology within these systems [18]. Temporarily open/closed estuaries are generally characterized by low species richness [19] due to the lack of recruitment opportunities for marine breeding species into these systems and the fact that many marine species are incapable of completing their life cycles in these systems [12]. Moreover, extreme variability in physico-chemical factors such as temperature and salinity, typically recorded within these systems can pose physiological constraints on many species, particularly freshwater and marine representatives [20,21].
The sand prawn, Callichirus kraussi (Stebbing, 1900), has been identified as a key component of the macrobenthic fauna in soft sediment nearshore marine environments and estuaries, including TOCEs along the southern African coastline. [1,22]. Densities of sand prawn are highly variable and may attain levels of up to 100 ind m-2. Sand prawns create burrows extending over 1m into the sediment, bringing the sediment to the surface where it is deposited in volcano-like mounds [ 23, 2]. The burrowing activity of the sand prawn modifies the sediment's biogeochemical properties, particularly pore-water, nutrients, and gaseous exchange between the sediment and water column and sediment granulometry and erodability [2,4,12]. Sand prawns modify nutrient cycles in the sediments and water column [24] and also contribute to the alteration of sediment texture (digenesis), mixing of water and solutes (bio-irrigation), and the displacement of non-living organisms [25]. Bioturbation fuels plankton primary production by releasing nutrients from sediments into the water column [9]. The direct release of nutrients, as well as nutrient cycling, are both enhanced by bioturbation through sediment oxygenation and an increase in the surface area available for microbial production and activity [7]. Therefore, bioturbation changes the dispersion deepness of organic matter and can raise the food stock and quality in sediments for deposit feeders. In addition to having a noticeable impact on bacteria, microalgae, meiofauna, macrofauna, and seagrasses in estuarine environments, sand prawns play a significant role in the structuring of soft-bottom communities through their sediment reworking activities [10,24,26,27]. Studies have shown that the presence of sand prawns can lead to a decrease in the abundance of macrofauna due to direct physical disturbances or competition for space and resources [28]. However, in other cases, they can promote biodiversity by creating habitats that are more favorable to certain species [29]. For instance, the presence of sandprawn burrows can provide shelter for small invertebrates or offer an oxygenated refuge in otherwise anoxic sediments [10].
Global climate change along the southern African coastline has been implicated in the increased incidence of extreme weather events including storm activity, increased wave height, and prolonged droughts [32,33]. These events are likely to have a far-reaching effects on the ecosystem functioning of TOCEs within the subregion [32,34] Much of the Eastern Cape province of South Africa experienced a severe drought from 2015 to 2019 which contributed to the prolonged mouth closure of TOCEs within the region [33]. The current investigation was conducted to assess the role of the sand prawn, C. kraussi, as an ecosystem engineer under conditions of prolonged mouth closure in a temperate TOCE located along the southeastern coastline of South Africa. The study was conducted employing a 16-week-long caging experiment set up in the lower reach of a temperate temporarily/open closed estuary in the Eastern Cape province of South Africa
2.Study Site
This study was conducted in the lower reach of the medium-sized warm-temperate Kasouga Estuary located on the south-eastern coastline of South Africa which enters the Indian Ocean at 330 39’ 17’’ S 260 44’ 08’’ E [35,36]. Its surface area and width are estimated at 22 hectares and 150m, respectively and the estuary is navigable for about 2.5 km with its main channel depth varying between 0.5 m and 2 m [37]. This system is considered to be pristine and in good ecological condition due to limited agricultural activities within its 39 km2 catchment area [38,39]. Due to limited freshwater inflow because of a small catchment area, the Kasouga Estuary receives limited freshwater which consequently results in low levels of macronutrients entering the system [40]. Compared to permanently open estuaries in the region, Kasouga Estuary supports low levels of phytoplankton (< 2.0mg Chl-a m-3) and zooplankton biomass (< 15 mg dwt m-3) [39]. The zooplankton community is numerically and dominated by copepods such as the Pseudodiaptomus hessei and Paracartia longipatella [41,42]. Additionally, benthic communities in TOCEs typically include diverse macrofaunal and meiofaunal assemblages such as polychaetes, amphipods, mollusks (e.g., gastropods like Nassarius kraussianus), and crustaceans, including the burrowing sand prawn (Callichirus kraussi) [10,11]. Microphytobenthic communities, such as benthic diatoms, also play a significant role in primary production nutrient cycling through bioturbation and organic matter decomposition [12].
Figure 1.
Geographic location of Kasouga Estuary along the southern African coast (after Froneman, 2013). The caging experiment was conducted in the lower reach of the system.
Figure 1.
Geographic location of Kasouga Estuary along the southern African coast (after Froneman, 2013). The caging experiment was conducted in the lower reach of the system.
3. Materials and Methods
Caging experiments were conducted in the lower reach of the temporarily open/closed Kasouga Estuary. Manipulations were conducted over a period of three months from 1 January to 15 March 2020. The densities of the sand prawn in the lower reach of the system where the experiment was conducted ranged from 5 to 36 and m-2 (determined by counting the number of active burrows- data not shown). Stainless steel open-topped cages with dimensions of 50 cm × 50 cm × 30 cm, with 1 mm mosquito netting on the bottom and sides, were buried in the sediment, leaving ≈10 cm protruding above the sediment, in the lower reaches of the estuary and were filled with sun-dried sand collected from the edge of the estuary 48 hours before the study to ensure faunal absence at the onset of the study. The cages were not roofed, since this would affect their colonization from the water column and would alter the light environment. Three weeks after installing the cages, sand prawns of between 43 to 54 mm standard length (mean = 44.3 ± 5.35 mm), were collected from the lower reach of the estuary using a prawn pump. Four treatments were prepared. For the control, prawns were absent from the cages. For Treatment 1, 5 prawns were added, Treatment 2, 10 prawns, and for Treatment 3, 20 prawns were added to each of the cages. Three replicates were prepared for each treatment. Cages were monitored weekly to ensure that prawn densities in the various treatments remained constant. Densities of the prawn within the cages were assumed to correspond to the number of burrows [9]. At the end of the experiment, bioturbation by the sand prawn, microphytobenthic algal concentrations, and the abundance, biomass and community composition of the macrobenthos (epifauna and infauna) were determined as described below.
3.1. Bioturbation
Centrifuge tubes, 10 cm long with a mouth of 0.78 cm2, were employed as sediment traps to estimate bioturbation by C. kraussi. The aspect ratio (length: diameter) of each tube was 10:1, considered the optimum value to prevent re-suspension of trapped sediments [44]. Tubes were buried vertically, leaving ~2 cm protruding above the sediment surface. Tubes were sealed after 1 h and the sediment in each was extracted and dried at 60 °C for 24 h and weighed using a Sartorius microbalance. The amount of sediment within each tube was employed as an index of sediment erodability and expressed as g cm–2 h–1 [44]
3.1.1. Microphytobenthic Algal Concentrations
Microphytobenthic algal concentrations were determined by collecting a sediment surface sample in the upper 1 cm of the sediment from each cage using a polycarbonate tube. The tubes were sealed and kept in the dark in a cooler box until their return to the laboratory and were stored at −40 °C freezer in the dark with 30 ml 90% acetone until processing. Total microphytobenthics chlorophyll a (Chl-a) concentrations were then determined fluorometrically using a Turner 10-AU fluorometer before and after acidification [45]. Microphytobenthic Chl-a concentrations were expressed as μg Chl a cm–2.
3.1.2. Epifauna and Infauna Community Structure
To compare the macrobenthic community structure of the epifauna and infauna in the different treatments, the sediment in the cages was collected at the end of the experiment and sifted through a 1 mm mesh on site. The organisms collected were preserved in 70% alcohol and transported to the laboratory. Species composition was identified to the lowest taxon using identification keys [46]. Epifaunal and infaunal abundances were expressed as ind. m–2 (epifauna) or ind. m–3 (infauna), wet biomass per cage, expressed as g wwt m–2 (epifauna) or g wwt m–3 (infauna). Wet weight was determined by removing excess water from the animals using blotting paper and weighing to the nearest 0.1g using an Sartorius electronic balance. .
4. Statistical Analysis
To test for significant differences between the various treatments, a one-way ANOVA was employed. Post hoc Tukey test was then used to determine the sources of variation. Species diversity was calculated using Simpson`s diversity index (1-D). Non-metric Multi-Dimensional Scaling (MDS) was then employed to compare the macrofaunal community structure in the various treatments. Finally, Analysis of Similarity (ANOSIM) test was employed to detect the degree of similarity between the various treatments and to identify which species accounted for the differences between the various groupings identified in the MDS. Data analysis was conducted using the statistical packages, Statistica version 8, Past version 4, Minitab, and Microsoft Excel.
5. Results
The estuary remained closed throughout the duration of the caging experiment because of the protracted drought within the region. Due to the presence of an extensive sandbank at the mouth of the system, no overtopping events were recorded over the duration of the caging experiment.
5.1. Bioturbation
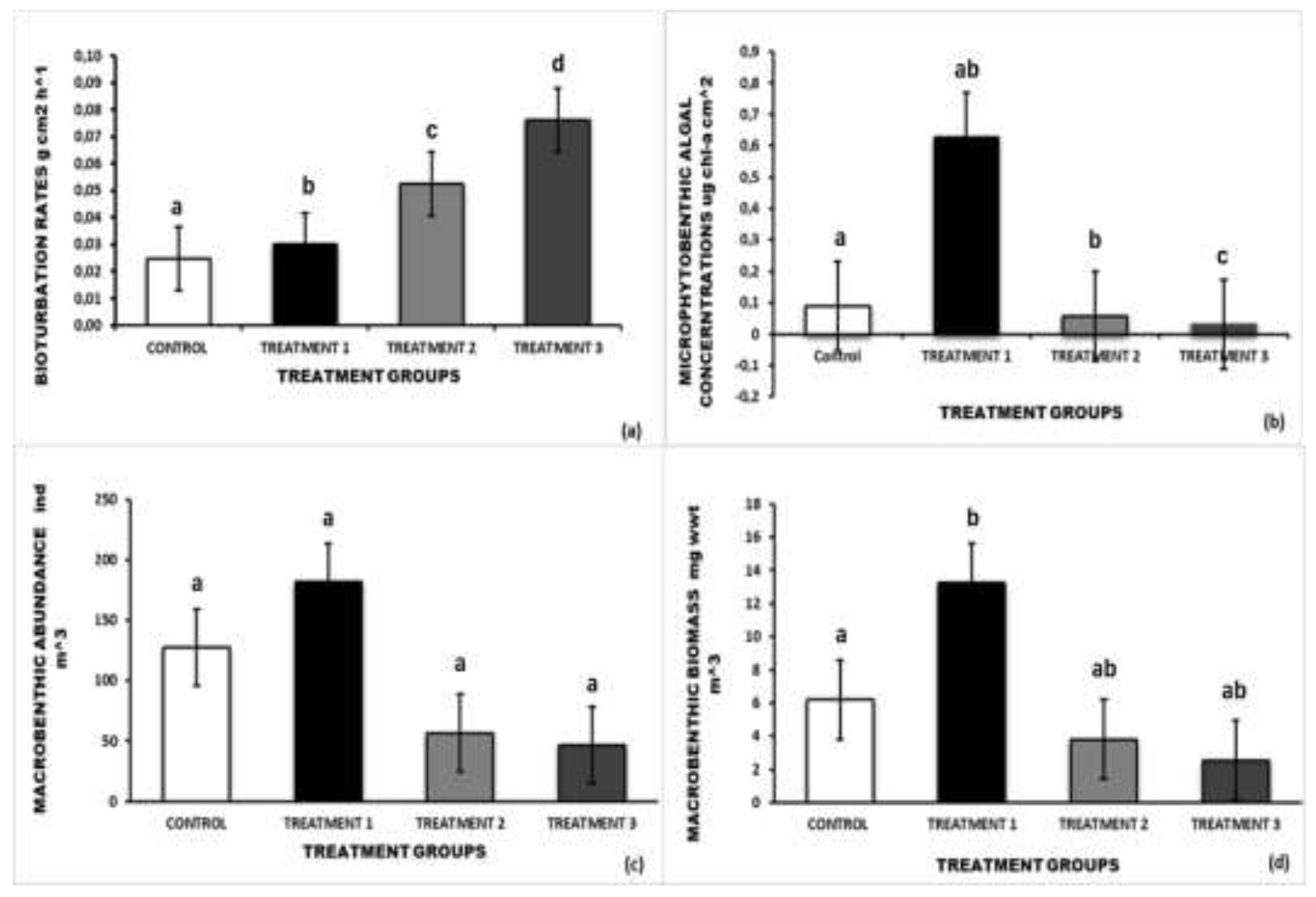
Bioturbation rates in the control cages (prawns excluded) ranged between 0.018 and 0.031 g cm2 h-1 (mean = 0,025 g cm2 h-1, SD=0.006). By contrast, the bioturbation rates in Treatment 1 ranged from 0.018 to 0.039 g cm2 h-1 (mean = 0,030 g cm2 h-1, SD= 0.009) and between 0.047 and 0.058 g cm2 h-1 (mean = 0,053 g cm2 h-1, SD= 0.005) in Treatment 2. The mean bioturbation rate in Treatment 3 was 0.076 g cm2 h-1 (range 0.065- 0.093 g cm2 h-1 , SD = 0.013, Figure 2(a)). ANOVA indicated that the bioturbation rates in Treatments 2 and 3 were significantly higher than those recorded in the Control and Treatment 1 (F= 29.78, P < 0.001). There were no significant differences in the bioturbation rates between Treatment 2 and Treatment 3 (P> 0.05). Similarly, the bioturbation rates in the Control and Treatment 1 were not significantly different from one another (P > 0.05).
5.1.1. Microphytobenthic Algal Concentrations
Total microphytobenthic algal concentrations in the control cages ranged from 0.06 to 0.104 µg chl-a cm-2 (mean = 0,089 µg chl-a cm-2 , SD = 0.016) (Figure 2b). Microphytobenthic algal concentrations in Treatment 1, ranged from 0.094 - 1.203 µg chl-a cm-2 (mean = 0,627 µg chl-a cm-2 , SD= 0.61) from 0.047 to 0.058 µg chl-a cm-2 (mean = 0,058 µg chl-a cm-2 , SD= 0.018) in Treatment 2 and from 0.065 to 0.093 µg chl-a cm-2 (mean = 0,032 µg chl-a cm-2 , SD= 0.007) in Treatment 3, (Figure 2(b)). ANOVA revealed that there was a significant difference between all treatment groups (F= 3,48, P< 0.05,).
5.1.2. Macrobenthic Abundances
Total macrobenthic abundances in the control cages varied from 68 to 203 ind m-3 (mean = 127 ind m-3 ,SD=67.8), while in Treatment 1 the values ranged between 98 and 254 ind m-3 (mean = 18 ind m-3 1, SD=68.8). In Treatment 2, the total macrobenthic abundances ranged between 38 to 72 ind m-3 with (mean =56.7 ind m-3 ,SD=14.7) and between 38 and 53 ind m-3 ( mean = 46.5 ind m-3, SD=6.7) in Treatment 3 (Figure 2c). One-way ANOVA indicated that the total macrobenthic abundances did not differ significantly between the Control and the various treatments (F=6.7, P=0.006, figure 2(c).
5.1.3. Macrobenthic Biomass
Macrobenthic biomass in the Controls and Treatment 1 ranged from 4.92 to 8.14 mg wwt m-3 (mean =6.20 mg wwt m-, SD = 1.48) and between 9.18 and 18.5 mg wwt m-3, (mean = 13.2 mg wwt m-, SD = 3.88), respectively. In Treatment 2 the macrobenthic biomass ranged from 3.06 to 4.18 mg wwt m-3 (mean = 3.80 mg wwt m-, SD = 0.508) while in Treatment 3 it ranged between 1.95 and 3.18 mg wwt m-3 (mean =2.56 mg wwt m-, SD = 0.642) (Figure 2(d)). ANOVA indicated that the total macrobenthic biomass in Treatment 1 was significantly higher than that recorded in Treatments 2 and 3 ( F=21.4, P< 0.001).
Figure 2.
(a) Sediment bioturbation rate, (b) macrophytobenthic algal concentrations, (c) macrobenthic abundance, (d) macrobenthic biomass in various treatments in Kasouga estuary. Error bars denote SD. Different letters indicate a significant difference (ANOVA, P < 0.05).
Figure 2.
(a) Sediment bioturbation rate, (b) macrophytobenthic algal concentrations, (c) macrobenthic abundance, (d) macrobenthic biomass in various treatments in Kasouga estuary. Error bars denote SD. Different letters indicate a significant difference (ANOVA, P < 0.05).
5.1.4. Epifaunal and Infaunal Community Structure
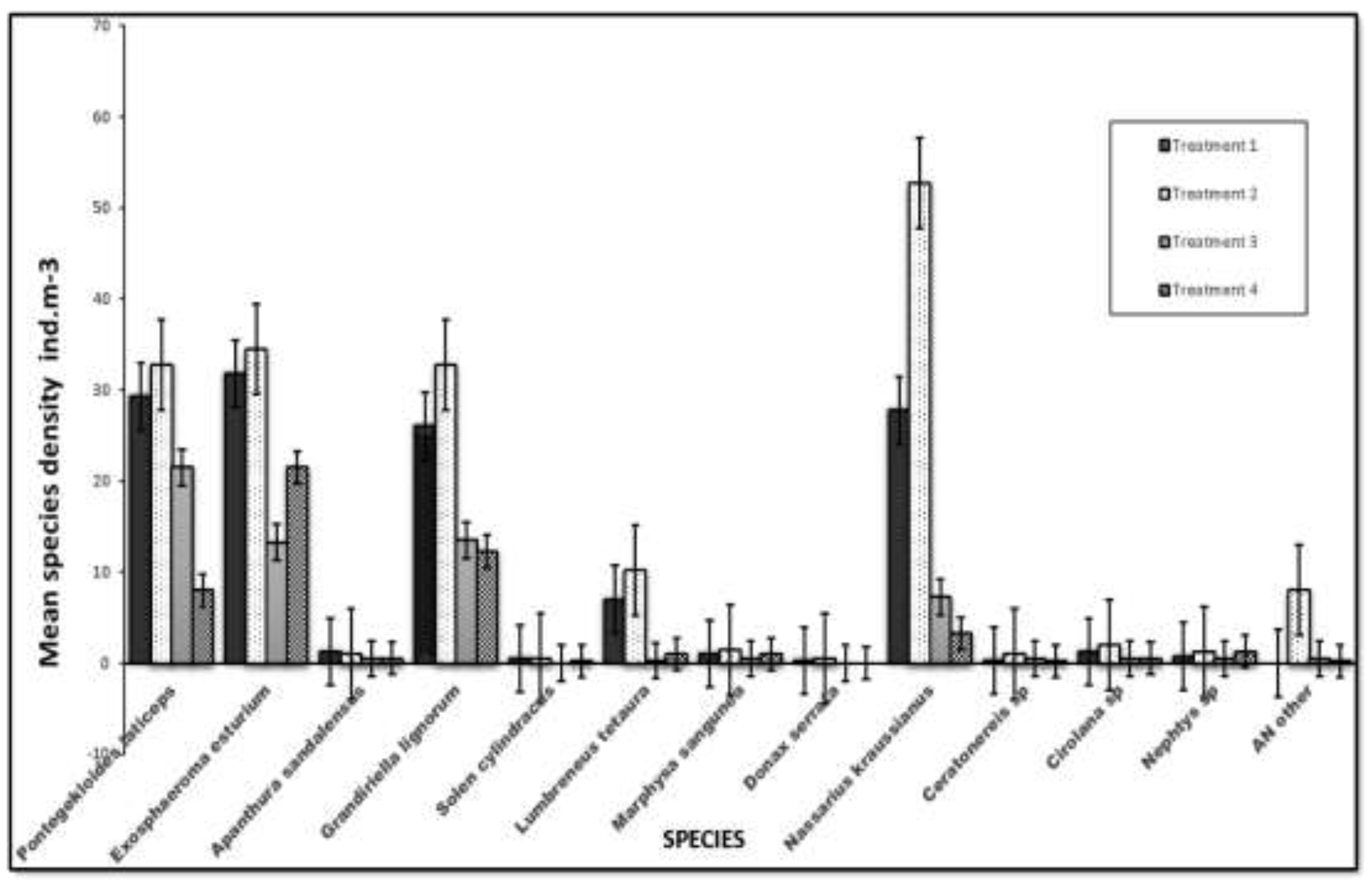
The epifaunal species recorded in the cages included three species of isopods: Pontogeloides latipes, Apanthura sandalensis, and Exosphaeroma estuarium, one species of amphipod, Grandidierella lignorum, two bivalve species, Solen capensis and Donax serra, and two polychaete species, Lumbreneus tetaura and Marphysa sanguinea. The epifaunal species included Nassarius kraussianus, Ceratonereis species, Cirolana species, and Nephtys species. The most common infaunal species were the isopods E. estuarium and P. latipes, and the amphipod G. lignorum, while the most common epifaunal species was N. kraussianus. The least common species were the bivalves Donax serra and Solen capensis. There were no significant differences in the community structure of epifaunal and infaunal species between the control and treatment groups (F = 16.29, P = 1.13E-23).
Figure 3.
Mean species density in four treatments ((T1- Control- 0 ind/m2, T2- 5 ind/m2, T3- 10 ind/m2, T4- 20 ind/m2) in the Kasouga Estuary. Error bars denote SD.
Figure 3.
Mean species density in four treatments ((T1- Control- 0 ind/m2, T2- 5 ind/m2, T3- 10 ind/m2, T4- 20 ind/m2) in the Kasouga Estuary. Error bars denote SD.
5.1.5. Bray Curtis Cluster Analysis
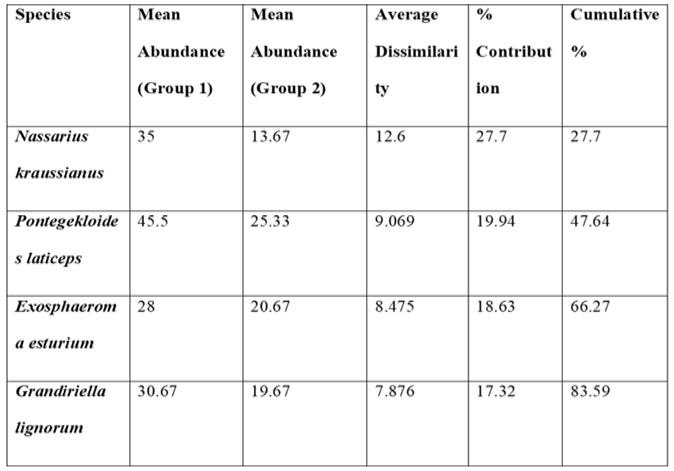
At the 70% similarity level, hierarchical cluster analysis identified two distinct groupings, designated Groups 1 and 2. Group 1 largely comprised the Control and Treatment 1 samples, while Group 2 comprised samples from Treatments 2 and 3 (Figure 5). One of the replicates for Treatment 2 was identified as an outlier. ANOSIM indicated that the differences between the two groups was significant (R=0.37, P= 0.008). The most prevalent epifaunal species was Nassarius. kraussianus, whereas the most prevalent infaunal species were the isopods Exosphaeroma estuarium and Pontegekloides latipes, as well as the amphipod Grandiriella. lignorum. The two bivalves, Donax serra and Solen capensis, were the least common species among the infauna.
Figure 4.
Multi-dimensional scaling (MDS) hierarchical cluster plot, showing Bray-Curtis similarity for the different treatments. Blue lines on the dendrogram indicate different groups and the outlier is circled.
Figure 4.
Multi-dimensional scaling (MDS) hierarchical cluster plot, showing Bray-Curtis similarity for the different treatments. Blue lines on the dendrogram indicate different groups and the outlier is circled.
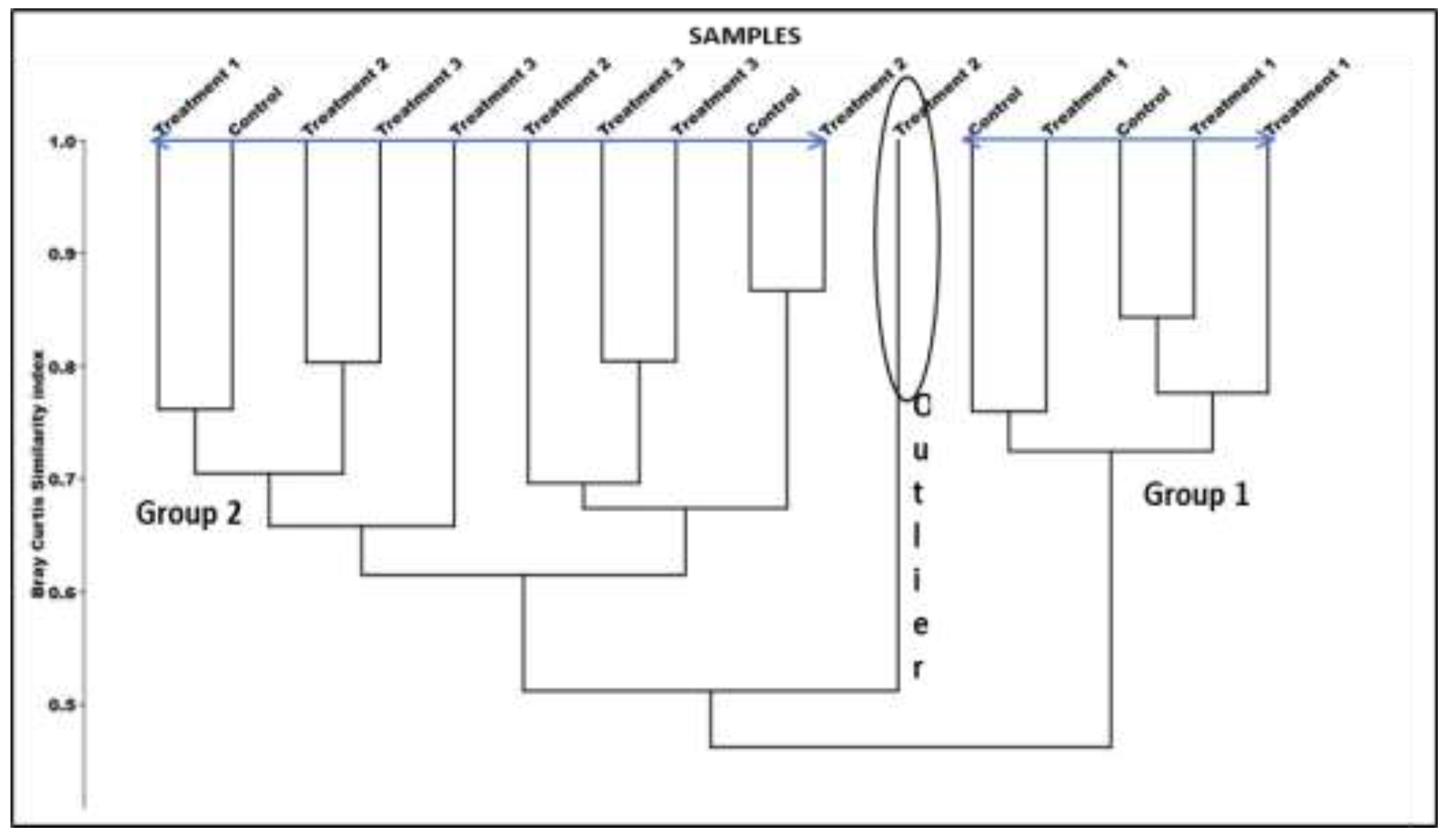

Table 1.
Showing SIMPER analysis for the most abundant infauna and Epifaunal species with their contributions to the Bray Curtis cluster groupings.
Table 1.
Showing SIMPER analysis for the most abundant infauna and Epifaunal species with their contributions to the Bray Curtis cluster groupings.
 |
6. Discussion
Historically, studies on estuarine ecosystem functioning of temporarily open/closed South African estuaries have largely focused on the importance of freshwater inflow, sediment characteristics, and mouth status in determining biological community structure within these systems [16,43,44,45]. There is now, however, a growing appreciation of the importance of non-trophic interactions in driving community dynamics within these systems [47]. Ecosystem engineers have emerged as one of the forms of non-trophic interactions responsible for shaping communities and ecosystems within shallow water marine ecosystems [26,48,49,50,51]. The current study was conducted to assess the role of the sand prawn, Calianassa kraussi as an ecosystem engineer in a TOCE during a prolonged drought which contributed to the Kasouga Estuary along the south-east coastline of South Africa being separated from the marine environment for a period exceeding 12 months.
Studies carried out in several southern African TOCEs have demonstrated that the maximum estimates of biology are typically attained during the closed phase of the system following freshwater inflow [4,26,52,53,54]. The estimates of total microphytobenthic alga biomass and macrobenthos abundance and biomass recorded during the current study were reduced compared to previous studies conducted in TOCEs in the same region [26,53]. The lower estimates can likely be attributed to the prolonged mouth closure which limited the recruitment of marine breeding species into the system and low macronutrient availability conferred by the low freshwater inflow resulting from the prolonged drought within the region [54,55]. The macrobenthic community structure observed during the present study is in agreement with previous investigations both within the same estuary and indeed, inTOCE’s within the same biogeographic region [4,47,49,52,54]
The increased rates of bioturbation recorded with the increase in the density of C. kraussi observed during the caging experiment is not unexpected and likely contributed to the decline in the total microphytobenthic algal concentrations within the cages (Figure 1(b)). A notable exception was recorded in Treatment 1, where a significant increase in microphytobenthic algal concentration was recorded over the duration of the caging experiment. The reduced microphytobenthic algal concentrations observed in Treatments 2 and 3, can likely be ascribed to the re-suspension of sediments by the burrowing activities of the prawn which creates an unfavourable light environment and ultimately bury the microphytobenthic algae [4,8,53]. The burrowing activities of sand prawn have been demonstrated to modify nutrient cycles in the sediments and water column of shallow water ecosystems [24]. Thus, the observed increase in microphytobenthic algal concentrations in Treatment 1 can possibly be attributed to the increased availability of macronutrients due to the burrowing activities of the sand prawn.
Total macrofaunal abundances and biomass within the different treatments generally decreased with increased sand prawn density (Figure 3and Figure 4). Again, a notable exception was recorded in Treatment 1 where the values increased. The observed patterns in Treatments 2 and 3 can possibly be ascribed to reduced food availability (microphytobenthic algae) or an attempt of macrofauna to avoid being buried by settling sediment due to the bioturbating activities of sand prawns [2,11]. Indeed, it is worth noting that differences in the groupings identified with the numerical analyses could largely be attributed to changes in the abundances of mobile components of the macrobenthos including the tick shell, Nassarius kraussianus and the isopod, Exospheroma hylocoetes. The decrease in macrofaunal abundance and biomass due to increased sand prawn densities during this investigation corresponds with similar studies conducted in other shallow water ecosystems and indeed within the same system [4,28,53,57]. For instance, [10] proposed that the reduction in surface microalgal development brought about by C. kraussi's reworking of sediments resulted in enhanced food intake by N. kraussianus and decreased food uptake by E. paupercula. This is because the physical disturbance of the sediment surface, direct consumption, and competition for nutrients create an environment that is less favorable for microalgal growth, which ultimately results in a reduction in microalgal biomass.
The elevated concentrations of microphytobenthic algae in the Control and Treatment 1 coincided with elevated abundances and biomass of the macrofauna (figure 1 a, b, c, and d). (Figure 5 and Figure 6). Bioturbation generated by sand prawns at low densities has been demonstrated to enhance macrofaunal abundances and species richness [58]. The increase in abundances and biomass of macrofauna observed in Treatment 1 could mainly be attributed to an increase in the abundance of N. kraussianus which were likely sustained by the elevated biomass of microphytobenthos [4,10].This result suggests that the effect of C. kraussi burrowing activities in shaping macrofauna within the study estuary was density-dependent [46]. Therefore, at higher sandprawn densities, production declines which in turn decreases the macrofauna in the experimental cages [59,60,61].
7. Conclusions
In agreement with previous studies, the results of the caging experiment indicate that the sand prawn can be considered a key ecological engineer in TOCE along the South African coastline. Importantly the caging experiments suggest that the burrowing activities of the sand prawn at low/medium densities may locally mitigate the adverse effects of prolonged mouth closure of TOCE by contributing to increased concentrations of microphytobenthic algae which sustain elevated abundances and biomass of macrofauna.
Funding
This work was funded by the National Research Foundation (NRF) of South Africa.
Conflicts of Interest
There is no conflict of interest to declare.
References
- Moyo, R. 2014. Ecology and behaviour of burrowing prawns and their burrow symbionts. University of Cape Town. MSc thesis.
- Siebert, T., & Branch, G.M. 2006. Ecosystem engineers: interactions between eelgrass Zostera capensis and the sandprawn Callianassa kraussi and their indirect effects on the mudprawn Upogebia africana. Journal of Experimental Marine Biology and Ecology 338: 253–270.
- Meysman, F. J. R., Middelburg, J. J., & Heip, C. H. R. (2006a). Bioturbation: a fresh look at Darwin’s last idea. In Trends in Ecology and Evolution (Vol. 21, Issue 12, pp. 688–695). [CrossRef]
- Henninger, T. O., & Froneman, P. W. (2013). Role of the sandprawn Callichirus kraussi as an ecosystem engineer in a South African temporarily open/closed estuary. African Journal of Aquatic Science, 38(1), 101–107. [CrossRef]
- Richter, R. (1952). Fluidal-texture in Sediment-Gesteinen und ober Sedifluktion überhaupt. Notizbl. Hess. L.-Amt. Bodenforsch. 3, 67-81.
- Dauwe, B., Herman P. M. J., & Heip C. H. R. (1998). Community structure and bioturbation potential of macrofauna at four North Sea stations with contrasting food supply. Marine Ecology Progress Series. 173: 67-83.
- Wilkinson, M. T., Richards, P. J., & Humphreys, G. S. (2009). Breaking ground: Pedological, geological, and ecological implications of soil bioturbation. In Earth-Science Reviews (Vol. 97, Issues 1–4). [CrossRef]
- Vasquez-Cardenas, D., Quintana, C. O., Meysman, F. J. R., Kristensen, E & Henricus, T. S., Boschker, H. T. S. (2016), Species-specific effects of two bioturbating polychaetes on sediment chemoautotrophic bacteria. Marine Ecology Series. 549: 55–68. [CrossRef]
- Queiros, A. M., Stephens, N., Cook, R., Ravaglioli C., Nunes, J., Dashfield, S., Harris, C., Tilstone, G. H., Fishwick, J., Braeckman, U., Somerfield, P. J., & Widdicombe, S.S. (2015), Can benthic community structure be used to predict the process of bioturbation in real ecosystems? Progress in Oceanography. 137: 559–569.
- Biles, C. and Paterson, David and Ford, Rich and Solan, Martin and Raffaelli, David. (2002). Bioturbation, ecosystem functioning and community structure. Hydrology and Earth System Sciences. 6. 10.5194/hess-6-999-2002.
- Pillay, D., Branch, G. M., & Forbes, A. T. (2007a). Experimental evidence for the effects of the thalassinidean sandprawn Callianassa kraussi on macrobenthic communities. Mar. Biol. 152, 611–618.
- Pillay, D., Branch, G. M., & Forbes, A. T. (2007). Effects of Callianassa kraussi on microbial biofilms and recruitment of macrofauna: A novel hypothesis for adult-juvenile interactions. Marine Ecology Progress Series, 347, 1–14. [CrossRef]
- Hanekom, N., & Russell, I. (2015). Temporal changes in the macrobenthos of sandprawn (Callichirus kraussi) beds in Swartvlei Estuary, South Africa. African Zoology. 50. 41-51. [CrossRef]
- Whitfield, A.K. (1998) Biology and Ecology of Fishes in Southern African Estuaries. Ichthyol. Monogr. Smith Inst. Ichthyol. No. 2.223 pp.
- Whitfield, A.K. (2022). Fishes of Southern African estuaries: from species to systems. Available at: http://hdl.handle.net/10962/97933.
- Gama, P. T., Adams J. B., Schael D.M., & Skinner T. (2005). Phytoplankton chlorophyll a concentration and community structure of two temporarily open/closed estuaries. WRC Report No. 1255/1/05. ISBN: 1-77005-326-3.
- Whitfield, A.K., & Bate G.C., (eds). 2007. A review of information on temporary open/closed estuaries in the warm and cool temperate biogeographic region of South Africa, with particular emphasis on the influence of river flow on these systems. WRC Report 1581/1/07. Pretoria: Water Research Commission.
- Whitfield, A. K., Adams, J. B., Bate, G. C., Bezuidenhout, K., Bornman, T. G, Cowley, P. D., Froneman, P. W., Gama, P. T., James, N. C., Mackenzie, B., Riddin T., Snow, G. C., Strydom, N. A., Taljaard, S., Terörde, A.I., Theron, A. K., Turpie, J. K., van Niekerk, L., Vorwerk, P. D., & Wooldridge, T. H. (2008). A multidisciplinary study of a small, temporarily open/closed South African estuary, with particular emphasis on the influence of mouth state on the ecology of the system, African Journal of Marine Science. Volume 30:3, 453-473 pp. [CrossRef]
- Whitfield, A. K., Bate, G. C., Adams, J. B., Cowley, P. D., Froneman, P. W., Gama, P. T., Strydom, N. A., Taljaard, S., Theron, A. K., Turpie, J. K., van Niekerk, L., & Wooldridge, T. H. (2012). A review of the ecology and management of temporarily open/closed estuaries in South Africa, with particular emphasis on river flow and mouth state as primary drivers of these systems. In African Journal of Marine Science (Vol. 34, Issue 2, pp. 163–180). [CrossRef]
- Harrison, T. (2004). Physico-chemical characteristics of South African estuaries in relation to the zoogeography of the region. Estuarine, Coastal and Shelf Science. 61. 73-87. [CrossRef]
- 21. Harrison,T.D. Whitfield,A.K.2006. Temperature and salinity as primary determinants influencing the biogeography of fishes in South African estuaries,Estuarine, Coastal and Shelf Science. 66,(1–2):335-345. ISSN 0272-7714(https://www.sciencedirect.com/science/article/pii/S0272771405003161). [CrossRef]
- Teske, P., & Wooldridge, T. (2001). A comparison of the benthic faunas of permanently open and temporarily open/closed South African estuaries. Hydrobiologia. 464. 227-243. 10.1023/A:1013995302300.
- Teske P.R., & Wooldridge T.H. (2003). What limits the distribution of subtidal macrobethos in permanently open and temporarily open/closed South African estuaries? Salinity vs sediment particle size. Estuarine, Coastal and Shelf Science 85: 407–421.
- Branch G.M., & Pringle A. (1987). The impact of the sand prawn Callianassa kraussi Stebbing on sediment turnover and on bacteria, meiofauna, and benthic microflora. Journal of Experimental Marine Biology and Ecology 107: 219–235.
- Humphreys G. S., & Mitchell P. B. (1983), A preliminary assessment of the role of bioturbation and rain wash on sandstone hillslopes in the Sydney Basin, in Australian and New Zealand Geomorphology Group. 66-80pp.
- Njozela, C. (2012). The role of the sandprawn, Callichirus kraussi, as an ecosystem engineer in a temporarily open/closed Eastern Cape estuary, South Africa.
- Wyness A.J., Fortune I., Blight AJ., Browne P., Hartley M., Holden M., Paterson D.M. (2021). Ecosystem engineers drive differing microbial community composition in intertidal estuarine sediments. PLoS One.16(2):e0240952. PMID: 33606695, PMCID: PMC7895378. [CrossRef]
- Pillay D, and Branch G.M. (2011). Bioengineering effects of burrowing thalassinidean shrimps on marine soft-bottom ecosystems. Oceanogr. Mar. Biol. Ann. Rev. ,49:137–192.
- Goedefroo, N., Braeckman, U., Hostens, K., Vanaverbeke, J., Moens, T., & De Backer, A. (2023). Understanding the impact of sand extraction on benthic ecosystem functioning: A combination of functional indices and biological trait analysis. Frontiers in Marine Science, 10, 1268999. [CrossRef]
- Emmerson, M.C., Solan, M., Emes, C., Paterson, D.M. andRaffaelli., D. (2001). Consistent patterns and the idiosyncraticeffects of biodiversity in marine systems. Nature, 411: 73–77.
- IPCC (Intergovernmental Panel on Climate Change). Summary for policymakers. In: T.F., Stocker, D.Q., Plattner, G.K., Tignor, M., Allen, S.K., Boschung J. (eds), Climate change (2013). The physical science basis. Contribution of working group I to the fifth assessment report of the Intergovernmental Panel on Climate Change. Cambridge, UK and New York: Cambridge University Press.
- Ziervogel, G., New, M., van Garderen, A.E., Midgley, G., Taylor, A., Hamann, R., Stuart-Hill, S., Myers, J., & Warburton, M. (2014). Climate change impacts and adaptation in South Africa. Wiley Interdisciplinary Reviews: Climate Change, 5(5): 605–620. [CrossRef]
- Mahlalela P.T., Blamey R.C., Hart N.C.G., Reason C.J.C. (2020). Drought in the Eastern Cape region of South Africa and trends in rainfall characteristics. Clim Dyn.,55(9-10):2743-2759. PMID: 32836893, PMCID: PMC7428292. [CrossRef]
- Froneman, P. W. (2018). The Ecology and Food Web Dynamics of South African Intermittently Open Estuaries. In Estuary. InTech. [CrossRef]
- Whitfield AK. 2000. Available scientific information on individual South African estuarine systems. Water Research Commission Report No. 577/3/00: 217 pp. Pretoria, South Africa: Water Research Commission.
- Wasserman, R. J., Kramer, R., Vink, T. J. F. & Froneman, P. W. (2014). Conspecific alarm cue sensitivity by the estuarine calanoid copepod Paracartia longipatella. Austral Ecology 39(6): 732–738.
- Froneman, P. W. (2002b). Seasonal changes in selected physico-chemical and biological variables in the temporarily open/closed kasouga estuary, eastern cape, south africa. African Journal of Aquatic Science, 27(2), 117–123. [CrossRef]
- Whitfield A.K. (1992). A characterization of southern African estuarine systems. South African Journal of Science 18:89-103.
- Froneman P.W. (2002). Response of the biology to three different hydrological phases in the temporarily open/closed Kasouga estuary. Estuarine, Coastal and Shelf Science., 55:535-546. ISSN 0272-7714. [CrossRef]
- Wooldridge TH, McGwynne L. (1996) The estuarine environment. Report No. C31. Institute of Coastal Research, Port Elizabeth, South Africa. 91pp.
- Froneman, P., W. (2004). In situ grazing rates of the copepods, Pseudodiaptomus hessei and Acartia longipatella in a temperate, temporarily open/closed estuary. South African Journal of Science. 100: 577-583.
- Wasserman, R. J., Froneman P. W., (2013). Risk effects on copepods: preliminary experimental evidence for the suppression of clutch size by predatory early life-history fish, Journal of Plankton Research. 35: 421–426. [CrossRef]
- Henninger, T., Froneman, P.,W., Richoux, N., & Hodgson, A,. N. (2009). Role of submerged macrohytes and a refuge and food source for the estuarine isopod, Exospheroma hylocoetes. Estuarine, Coastal and Shelf Science. 82:285-293.
- Ellis, J., Cumming V., Hewitt, J., Thrush, S., & Norkko, A. (2002). Determining effects of suspended sediment on condition of suspension bivalve (Atrina zelanica): results from a survey, a laboratory experiment and field transplant experiment. Journal of Experimental Marine Biology and Ecology 267: 147-174.
- Whitfield, A., & Baliwe, N. (2012). A numerical assessment of research outputs on South African estuaries. South African Journal of Science. 109. 01-04. [CrossRef]
- Day, J.H. (1969). A guide to marine life on South African shores. AA Bakema, Cape Town. South Africa.
- Menge, B., Berlow, E., Blanchette, C., Navarrete, S. and Yamada, S. (1994). The keystone species concept: variation in interactionstrength in a rocky intertidal habitat. Ecol. Monogr., 64: 249–286.
- Marenco, K., & Bottjer, D. (2007). 8 Ecosystem engineering in the fossil record: Early examples from the Cambrian Period. Theoretical Ecology Series. 4. [CrossRef]
- Berke, S. (2010). Functional Groups of Ecosystem Engineers: A Proposed Classification with Comments on Current Issues. Integrative and comparative biology. 50. 147-57. [CrossRef]
- Ellison, A., M. (2019). Foundation species, non-trophic interactions, and the value of being common. iScience 13: 254–268.
- Almeida G.M., Valente N. F., Pacheco, E.O., Ganci, C. L, Mathew A. M, Adriano S. P., & Diogo, B. (2020). How Does the Landscape Affect Metacommunity Structure? A Quantitative Review for Lentic Environments. Curr Landscape Ecol Rep 5, 68–75.
- Nozias, C., Persinotto R., Mundree, S. (2001). Annual cycle of microalgal biomass in a South African temporarily-open estuary: Nutrient versus light limitation. Marine Ecology Progress Series.,223:39-48.
- Perissinotto, R., Iyer, K., and Nozais, C. (2006). Response of microphytobenthos to flow and trophic variation in two South African temporarily open/closed estuaries. Botanica Marina, 49(1), 10–22. [CrossRef]
- Scharler, U.M., Lechman, K., Radebe, T., Jerling, H.L., (2020). Effects of prolonged mouth closure in a temporarily open/closed estuary : a summary of the responses of invertebrate communities in the uMdloti Estuary , South Africa. Afr. J. Aquat. Sci. 45, 121–130. [CrossRef]
- Adams, J.B & Van Niekerk, L. (2020). Ten Principles to Determine Environmental Flow Requirements for Temporarily Closed Estuaries. Water 12, 1944. [CrossRef]
- Pillay, D. (2010). Expanding the envelope: linking invertebrate bioturbators with micro-evolutionary change. Mar Ecol Prog Ser 409:301-303. [CrossRef]
- Queiros, A.M., Stephens N., Cook R., Ravaglioli C., Nunes J., Dashfield S., Harris C., Tilstone G. H., Fishwick J., Braeckman U., Somerfield P. J., & Widdicombe S. (2015). Can benthic community structure be used to predict the process of bioturbation in real ecosystems? Progress in Oceanography. 137: 559–569.
- Pillay, D., Branch, G.M., Dawson, J., & Henry, D. (2011). Contrasting effects of ecosystem engineering by cordgrass Spartima maritima and the sandprawn Callianassa kraussi in a marine-dominated lagoon. Estuarine, Coastal and Shelf Science 91: 169-176.
- Venter, O., Pillay, D., & Prayag, K. (2020). Water filtration by burrowing sandprawns provides novel insights on endobenthic engineering and solutions for eutrophication. Scientific Reports, 10(1). [CrossRef]
- Pillay, D., Williams, C., & Whitfield, A.K. (2012). Indirect effects of bioturbation by the burrowing sandprawn Callichirus kraussi on a benthic foraging fish, Liza richardsonii. Mar. Ecol. Prog. Ser. 453, 151–158.
- McIlroy, D., & Logan, G.A., (1999). The impact of bioturbation on infaunal ecology and evolution during the Proterozoic–Cambrian transition: Palaios, v. 14, p. 58–72.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



