
Submitted:
21 January 2025
Posted:
22 January 2025
You are already at the latest version
Abstract
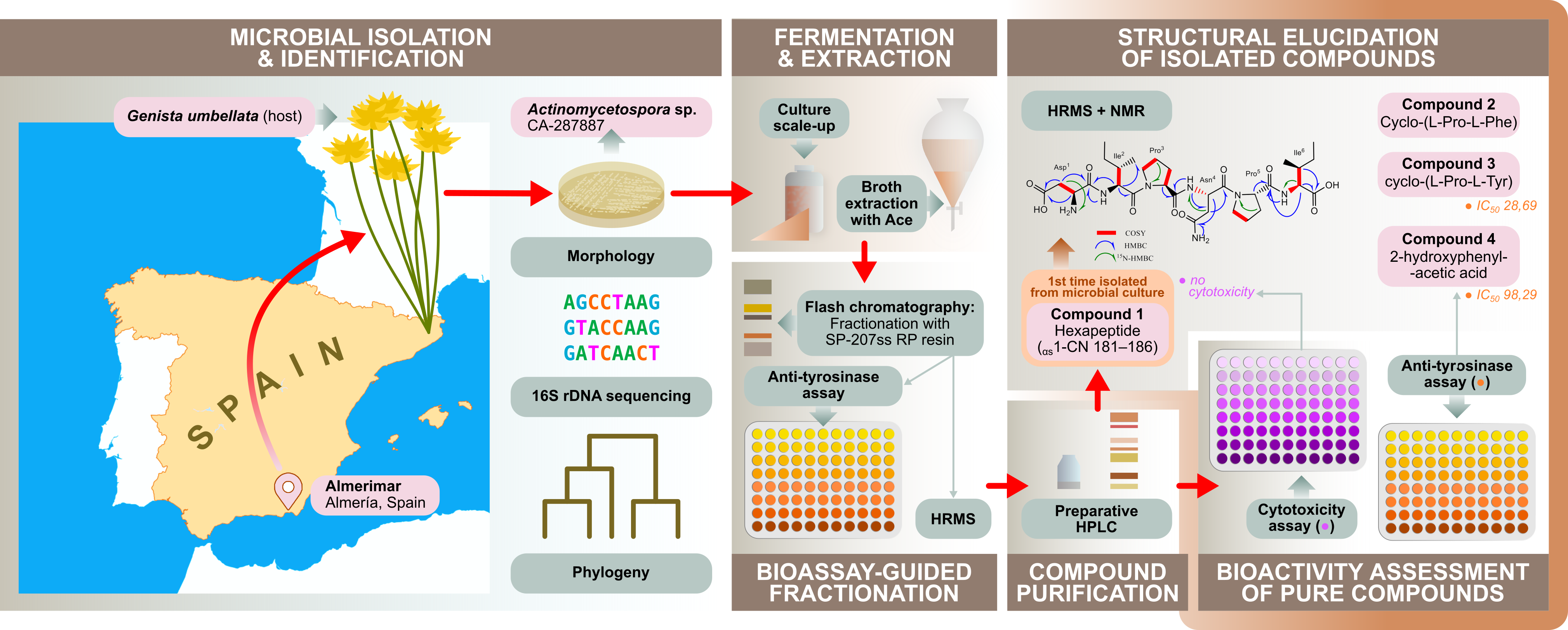
A high proteolytic resistant hexapeptide (αs1-CN 181–186) (1) along with two known 2,5-diketopiperazines, namely cyclo-(L-Pro-L-Phe) (2) and cyclo-(L-Pro-L-Tyr) (3), and the carboxylic acid 2-hydroxyphenylacetic acid (4) were isolated from the actinomycete strain CA287887. The morphological, 16S rRNA gene sequence and phylogenetic data of the strain exhibited a high similarity with members of the genus Actinomycetospora. The structure of 1 was thoroughly investigated for the first time by the extensive use of 1D and 2D NMR experiments while its absolute configuration was determined by Marfey’s analysis. The anti-tyrosinase effects of aforementioned compounds were investigated in vitro by using kojic acid as positive control (IC50 14.07μΜ). Compound 3 exhibited the highest activity (IC50 28.69), followed by compound 4 (IC50 98.29). Compound 1 was further evaluated for cytotoxicity against HepG2, A2058, A549 and MiaPaca-2 cell lines. At all the tested concentrations (0.01-200 μg/ml) no cytotoxic effect was observed.
Keywords:
Endophytic actinomycetes
; Pseudonocardiaceae
; Actinomycetospora sp.
; tyrosinase inhibition
; peptide
1. Introduction
The genus Actinomycetospora, belonging to the family Pseudonocardiacae, was first proposed in 2008 by Jiang et al. [1]. Most of the Actinomycetospora species have been isolated from soil or marine environments, while a few of them have been reported from lichen samples or as endophytes [1,2,3,4,5,6,7,8]. Although many different species have been described, their chemical content remains practically unknown. In fact, three thiazole derivatives are the only reported metabolites isolated from Actinomycetospora spp. [6]. In recent years, in-depth researches have focused on the diversity of endophytic actinomycetes, and on their ability to produce a wide range of bioactive secondary metabolites, characterized mainly as alkaloids, flavonoids, steroids, terpenoids, phenolics, quinones, and peptides, as well as on their applications in medicine, agriculture, and industry [3,5,7,8]. However, to the best of our knowledge, no studies have been conducted yet about the potential of endophytic actinomycetes to produce metabolites that could be used as whitening agents by inhibiting tyrosinase, the key enzyme of melanin synthesis [9]. As part of our screening program for the discovery of bioactive natural products from microorganisms of the global biodiversity with potential cosmeceutical applications, the fermentation extract of Actinomycetospora sp. CA-287887 strain cultivated in the medium M016 was selected for further investigation, due to its major impact on tyrosinase enzyme. The tyrosinase inhibition was observed both in cell-free (34.72 ± 8.46% tested at 0.02×WBE (whole broth equivalent)) and cell-based assays (70 ± 3.51% tested at 0.02×WBE, with statistical significance of P<0.01 vs. cells treated with the vehicle). Herein, we report for the first time the NMR assignments and the absolute configuration of the high proteolytic resistant hexapeptide, αs1-CN 181–186 (1) (Figure 1), along with the isolation of three known compounds (2-4) that demonstrated moderate to strong tyrosinase inhibitory activity.
2. Materials and Methods
2.1. General Experimental Procedures
IR spectra were recorded using a JASCO FT/IR-4100 spectrometer (JASCO Corporation) equipped with a PIKE MIRacleTM single reflection ATR accessory. 1H and 13C NMR spectra were obtained at 500 MHz, using a Bruker Avance III 500 MHz spectrometer (500 MHz and 125 MHz for 1H and 13C NMR respectively) equipped with a low volume 1.7 mm inverse detection microcryoprobe (Bruker Biospin, Fällanden, Switzerland). HRESIMS and LC-UV-MS data were measured using a Bruker maXis QTOF mass spectrometer (Bruker Daltonics GmbH, Bremen, Germany) coupled to an Agilent 1200 HPLC system (Agilent Technologies, Waldbronn, Germany) and on an Agilent 1100 single quadrupole LC-MS system (Santa Clara, CA, USA), as previously described [10]. Preparative HPLC was performed on a Gilson 322 System (Gilson Technologies, USA) using a Xbridge™ C18 (19 × 250mm, 5 μm) column at a flow rate of 20 ml/min. Semipreparative HPLC was performed on the same system using a Xbridge™ C18 (10 × 150mm, 5 μm) column at a flowrate of 2 ml/min. Evaporation of solvents was performed on a vacuum rotatory evaporator (Rotavapor R-3000r, Buchi, Postfach, Switzerland). The acetone employed for extraction, as well as the solvents used for isolation were of analytical and HPLC grade, respectively.
2.2. Isolation and Identification of the Strain CA-287887
2.2.1. Microbial Source
The producing strain CA-287887 is an endophyte isolated from the aerial part of a specimen of the plant Genista umbellata, collected in Almerimar (Almería, Spain). The original colony was isolated from YECD agar medium (yeast extract-casein hydrolysate agar) [11], containing nalidixic acid (20 µg/ml) and purified on Yeast Extract Malt Extract Glucose medium (ISP2) before its preservation as frozen agar plugs in 10% glycerol in MEDINA’s culture collection.
2.2.2. DNA Extraction and 16S rDNA Sequencing
Total genomic DNA was recovered and purified as previously described [12] from the strain grown in ATCC-2 liquid medium (0.5% yeast extract (Difco, Franklin Lakes, NJ, USA), 0.3% beef extract (Difco), 0.5% peptone (Difco), 0.1% dextrose (Difco), 0.2% starch from potato (Panreac, Barcelona, Spain), 0.1% CaCO3 (E. Merck, Darmstadt, Germany) and 0.5% NZ amine E (Sigma, St Louis, MO, USA). DNA preparations were used as template DNA for Taq Polymerase. PCR primers fD1 and 1100r were used for amplifying the 16S ribosomal RNA gene of the strain [12]. Reactions were performed in a final volume of 50 µl containing 0.4 µM of each primer, 0.2 mM of each of the four deoxyribonucleotide triphosphates (Roche, Indianapolis, IN, USA), 5 µl of extracted DNA, 1U Taq polymerase (Appligene, Watford, UK) with its recommended reaction buffer. PCR amplifications were performed in a Peltier Thermal Cycler PTC-200, according to the following profile: 5 min at 95°C and 40 cycles of 30s at 94°C, 30s at 52°C for and 1min at 72°C, followed by 10 min at 72°C. The amplification products were analyzed by electrophoresis in 2% (w/v) pre-cast agarose gels stained with ethidium bromide (E-gel 2%, 48 wells, Invitrogen, Carlsbad, CA, USA). PCR products were sent to Secugen (http://www.secugen.es/) for sequencing and were purified and used as a template in sequencing reactions using the primers fD1 and 1100r [12]. Amplified DNA fragments were sequenced using the ABI PRISMDYE Terminator Cycle sequencing kit and fragments were resolved using the ABI3130 genetic analyzer (Applied Biosystems, Foster City, CA, USA). Partial sequences were assembled and edited using the Assembler contig editor component of Bionumerics (ver 6.6) analysis software (Applied Maths NV, Sint-Martens-Latem, Belgium).
2.2.3. Phylogenetic Analysis
The nearly complete 16S rRNA gene sequence (1361 nucleotides) of strain CA-287887 was compared with those deposited in public databases and the EzBiocloud server [13,14]. The strain exhibited the highest similarity (99.56%) with Actinomycetospora atypica NEAU-st4T (KC412867), using EzBiocloud and GenBank sequence similarity searches and homology analysis.
Phylogenetic and molecular evolutionary analyses were conducted using MEGA version X [15]. Multiple alignment was carried out using CLUSTALX [16], integrated in the software. The phylogenetic analysis, based on the Neighbor-Joining method [17] using matrix pairwise comparisons of sequences corrected with Jukes and Cantor algorithm [18], shows that the strain is closely related to the type strain Actinomycetospora atypica NEAU-st4T and this relatedness is supported in the analysis by the bootstrap value (76) (Figure S17).
The morphological, 16S rRNA gene sequence and phylogenetic data were indicative that strain CA-287887 was representative of members of the genus Actinomycetospora and the strain was referred to as Actinomycetospora sp. CA-287887.
2.3. Scale-Up Fermentation, Extraction and Isolation of Compounds 1-4
To scale-up the microfermentation to 2 L, 0.5 ml of the frozen inoculum stocks were added into a sterile colony tube (150 × 24 mm with a cover slip inside) filled with 10 ml of ATCC-2 seed medium (starch 20 g, dextrose 10 g, NZ Amine type E 5 g, Difco Beef extract 3 g, Bacto Peptone 5 g, Yeast extract 5 g, CaCO3 1 g, distilled H2O 1 L) and incubated in an orbital Kühner shaker for 5 days, at 28 °C, 70% humidity, 225 rpm. After the end of the incubation, 2.5 ml were transferred into two 250 ml baffled-flasks filled with 50 ml of seed medium and were incubated following the same conditions. 7.5 ml of the second seed medium was transferred to twenty 500 ml flasks containing 150 ml of the fermentation medium M016 [19] and incubated for 7 days, under the same conditions.
The scale up fermentation broth (2 L) was extracted with acetone (2 L) under continuous shaking at 220 rpm for 1 h and centrifugation was followed. The remaining mixture (ca. 4 L) was concentrated to ca. 2 L under a nitrogen flow. The solution was loaded, with continuous 1:1 water dilution, keeping the flow-through (aqueous fraction, AQ) on a column packed with SP-207ss reversed-phase resin (brominated styrenic polymer, 65 g) previously equilibrated with water. The loaded column was further washed with water (2 L) and eluted at 10 ml min−1 on an automatic flash-chromatography system (CombiFlash Rf, Teledyne Isco) using a linear gradient from 5% to 100% acetone in water (in 30 min) with a final 100% acetone step (for 20 min) collecting 32 fractions of 20 ml. Fractions were concentrated to dryness on a centrifugal evaporator, tested for their tyrosinase inhibitory activity and analyzed by HRMS. Fractions 12, 13, 14 and AQ exhibited tyrosinase inhibitory effect and thus they were forwarded to a detailed chemical investigation.
Purification of fraction 12, containing mainly compound 1, was performed by preparative HPLC (UV detection at 210 nm) using a linear gradient of H2O−CH3CN from 5% to 35% CH3CN over 36 min, followed by a gradient to 100% CH3CN in 2 min to obtain 86 fractions, that were collected every 0.5 min. Fraction 44 (13.2 mg, eluted at 22.2 min), that contained compound 1, was further purified by semipreparative HPLC (UV detection at 210 nm) using a linear gradient of H2O−CH3CN from 5% to 60% CH3CN over 20 min, followed by a gradient to 100% CH3CN in 2 min, to yield compound 1 in high purity (1.7 mg) with a retention time of 10.4 min.
Compounds 2, 3 and 4 were obtained from fractions 13, 14 and AQ respectively. Compound 2 (2.6 mg) was obtained by using semipreparative HPLC (UV detection at 210 nm), using isocratic elution with H2O−CH3CN: 25-75% for 20 min, followed by a gradient to 100% CH3CN in 15 min (retention time of 2: 2.6 min).
Compound 3 (5.6 mg) was obtained by using first a preparative HPLC (UV detection at 210 nm), using a linear gradient from H2O−CH3CN: 95-5% to H2O−CH3CN: 30-70% over 36 min, followed by a gradient to 100% CH3CN in 2 min (elution time of fraction containing 3: 10.1 min). Further purification of the fraction containing 3 was performed by semipreparative HPLC using a linear gradient system from H2O−CH3CN: 95-5% to H2O−CH3CN: 78-22% over 18 min, followed by a gradient to 100% CH3CN in 2 min (retention time of 3: 9.9 min).
Compound 4 (1.2 mg) was eluted at 4.7 min from the AQ fraction, using an isocratic method with H2O−CH3CN: 98-2% for 10 min, followed by a gradient to 100 % CH3CN in 6 min (retention time of 4: 4.7 min).
Compound (1)
Brownish solid; UV (MeOH) λmax only end absorption detected; IR vmax 3293, 3286, 2965, 1722, 1644, 1632, 1537, 1448, 1314, 1254, 1220, 1024, 953 cm-1; 1H (500 MHz) and 13C (125 MHz) NMR spectral data in DMSO-d6, see Table S1; HRESIMS m/z: 668.3585 [M + H]+ (calcd for C30H50N7O10, 668.3619).
2.4. Marfey’s Analysis
Compound 1 (0.5 mg, 0.5 mg/ml) was treated with 6 N HCl in a sealed vial at 110 °C for 16 h. After concentration of the hydrolyzed sample to dryness, it was reconstituted in H2O (100 μl) and 50 μl of the sample were treated with 150 μl of L-FDVA (1% in acetone) and 20 μl of 1 M NaHCO3 in a sealed vial at 40 °C for 1 h. The neutralization of the reaction mixture was performed with 30 μl of 1 N HCl [20]. For the LC-MS analysis a 10 μl aliquot was diluted with 40 μl of CH3CN. For each standard amino acid the same process was followed starting with a stock solution of 50 mM. The resulting solutions were analyzed by LC-MS using a Zorbax SB-C8 column (21 x 300 mm, 3.5 μm) and the solvents used were A (10% CH3CN, 90% H2O, 1.3 mM TFA, 1.3 mM ammonium formiate) and B (90% CH3CN, 10% H2O, 1.3 mM TFA, 1.3 mM ammonium formiate)/90%). For the hydrolysate of 1 and amino acids Ile and allo-Ile, an isocratic system of 10% B for 2 min, followed by a linear gradient to 60% B in 33 min at flow of 1 ml/min was used. For the hydrolysate of 1 and amino acids Pro and Asp, an isocratic system of 10% B for 2 min, followed by a linear gradient to 20% B in 33 min at flow of 1 ml/min was used. The hydrolysate of 1 contained L-Ile (12.76 min), L-Pro (13.27 min) and L-Asp (6.75 min). The retention time of the L-FDVA derivatives of the authentic amino acids were as follows: L-Ile (12.77 min), D-Ile (17.83 min), L-Allo-Ile (12.55 min), D-Allo-Ile (17.66 min), L-Pro (13.21 min), D-Pro (18.94 min), L-Asp (6.52 min) and D-Asp (9.08 min).
2.5. Biological Evaluation
2.5.1. Tyrosinase Inhibitory Assay
Enzymatic Assay
The ability of the tested extracts and compounds to inhibit the catalytic action of tyrosinase in the oxidation of L-DOPA to dopachrome was performed by using mushroom tyrosinase, a lyophilized powder, ≥ 1000 units/mg solid (EC Number: 1.14.18.1) [21].
Cell-Based Assays
Cell lines and cell culture conditions
Mouse skin melanoma cells (B16-F10) were obtained from the American Tissue Culture Collection (ATTC). B16-F10 cells were cultured in DMEM containing 10% (v/v) FBS and 2 mM glutamine. Cells were maintained in a humidified environment of 5% CO2 and 37°C. They were subcultured using a trypsin/EDTA solution (ThermoFisher Scientific).
Tyrosinase activity
Mouse melanocytes B16F10 were planted in 60-mm dishes. After 24h of treatment with 1 μg/ml or 10 μg/ml for each extract, cells were lysed with 0.2% Nonidet P-40 buffer. L-DOPA (Sigma Chemical) was used as a substrate for the tyrosinase activity assay. A total of 20 μg of proteins from each sample (diluted in 100 μl of phosphate buffer) were placed in 96 well micro plate. L-DOPA (final concentration 5 mM) was added to each sample. The mixture was incubated at 37°C for 60 min. The absorbance was measured at 475 nm. Each sample was prepared in triplicate. The percentage of relative tyrosinase activity was calculated as follows: % Activity = [(A1-Bl)/ (A0-Bl)] x100. Where A0 = Control absorbance, Bl = Blank absorbance (L-DOPA only), A1 = Sample absorbance.
2.5.2. Cytotoxicity
Cytotoxicity was tested against HepG2, A2058, A549, MCF-7 and MIA PaCa-2 cell lines by the ΜΤΤ method and on CCD25sk cell line by the HOECHST assay, following an already described process [12].
3. Results and Discussion
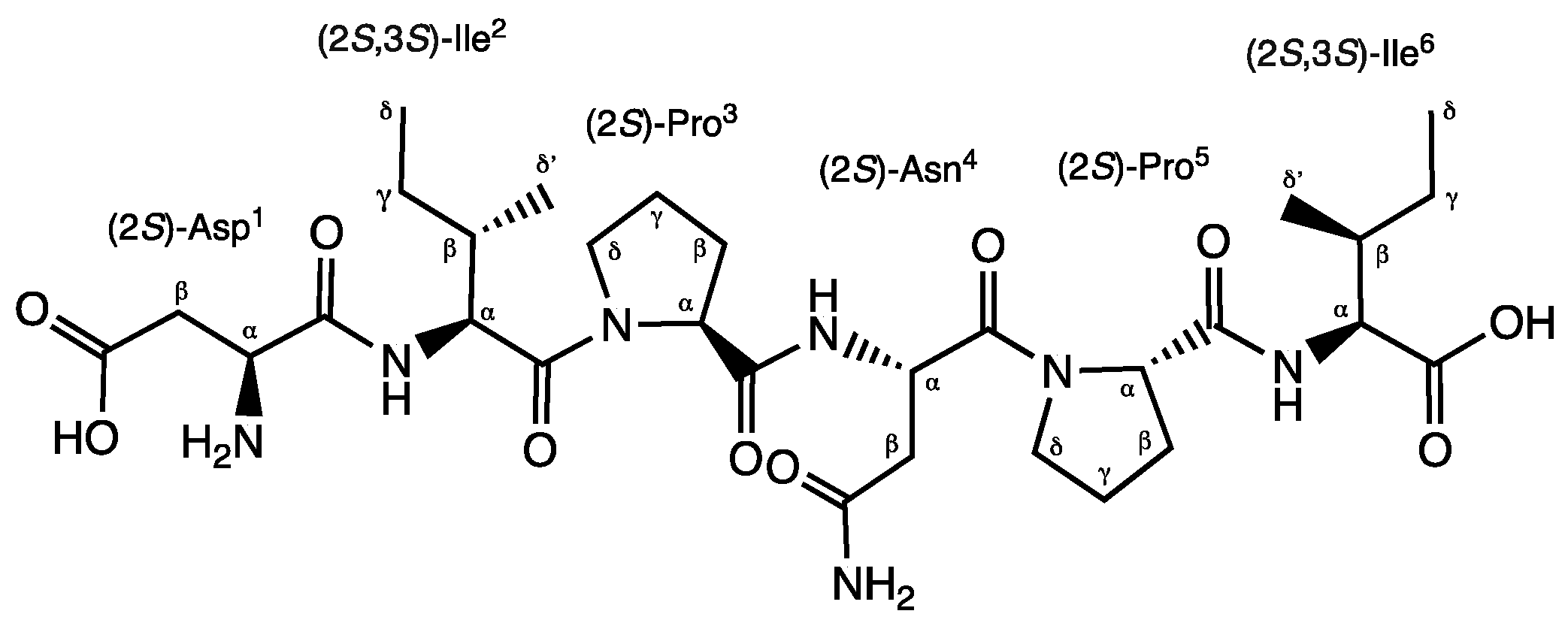
Compound 1 was obtained as a brownish solid. On the basis of 1D and 2D- Nuclear Magnetic Resonance (NMR) experiments, along with High Resolution Electrospray Ionization Mass Spectroscopy (HRESIMS) data, its molecular formula, implying ten degrees of unsaturation, was deduced to be C30H49N7O10 (m/z 666.3454 [M-H]-, m/z 668.3585 [M+H]+). The infrared spectroscopy (IR) spectrum of compound 1 showed characteristic absorption bands for secondary amines (3286 cm−1) and amide carbonyls (1632 cm−1), while the absence of any UltraViolet (UV) absorption above 201 nm excluded the presence of conjugated π (pi) bond systems. Analysis of the 13C NMR spectrum revealed 30 signals. On the basis of a Distortionless Enhancement by Polarization Transfer (DEPT) and of a Heteronuclear Single-Quantum Correlation Spectroscopy (HSQC)-DEPT experiment, aforementioned signals were assigned to eight methine, ten methylene, four methyl, and eight quaternary carbons, while the analysis of the 15N-HSQC and 15N-Heteronuclear Multiple Bond Correlation (15N-HMBC) spectra revealed additional nitrogen signals relative to one primary, three secondary and two tertiary amide groups. In the ¹H-¹H Correlation Spectroscopy (COSY) experiment, NH signals of each secondary amide showed cross-peaks with proton resonances at δH 4.07, 4.34 and 4.74 in the 1H NMR spectrum, which is characteristic of the α-methine protons of amino acidic residues. The analysis of the different spin systems by 2D- Total Correlation Spectroscopy (2D-TOCSY) resolved and assigned the α-methine protons at δH 4.07 and 4.34 to two isoleucine residues (Ile2 and Ile6), while the HMBC correlation of the α-methine proton at δH 4.74 with the carbonyl of a primary amide group at δC 171.8 suggested the presence of Asparagine (Asn4). In addition, the HMBC correlations of Ile2 and Ile6 Hα protons with their respective Cγ, Cδ’ protons, as well the hetero correlations between the primary amide protons at δH 7.40 and 6.88 and the carbonyl carbon at δC 171.8 of the side chain of Asn4, further confirmed the above hypothesis.
Three additional α-methine protons at δH 3.75, 4.31, 4.38 were identified in the 1HNMR spectrum. However, unlike Hα protons of Ile2, Ile6 and of Asn4, they didn’t show any COSY correlation with amino carbonyl protons. Regarding downshifted α-protons at δH 4.31 and 4.38, this was a clear evidence of an adjacent tertiary amide group instead of a primary amine. Analysis of the HMBC spectrum evidenced the correlation of the α-proton at δH 4.31 and 4.38 with methylene carbons at δC 29.1, 24.2, 47.2 and 28.9, 24.4, 46.6, respectively. Aforementioned chemical shifts in combination to the presence of tertiary amide groups suggested the presence of two proline residues in the molecule (Pro3 and Pro5). Further analysis of COSY and TOCSY spectra confirmed the above findings.
Finally, the upfield shift of the α-methine proton at δH 3.75 in combination to its COSY correlation with methylene β-protons at δH 2.45 and 2.27, which in turn give 15N HMBC correlations to an aliphatic ammine at δC 32.9, suggested the presence of a N-terminal aminoacidic residue. Further analysis of the HMBC spectrum showed the 2J and 3J long range correlation between the α- and β-protons and the quaternary carbon at δC 172.6, suggesting an acetyl side chain relative to aspartic acid (Asp1).
The exact sequence was established on the basis of HMBC and Nuclear Overhauser Effect Spectroscopy (NOESY) experiments (Figure S1 & Figure S2). The HMBC correlation between the Hα of Ile2 and the carbonyl amide at δC 170.9 of Asp1 confirmed the first two amino acids of the hexapeptide molecule, while the NOESY correlations Hα, Hδ΄ (Ile2)/ Hδ (Pro3), Hα (Pro3)/ NH(Asn4), Hα (Pro3)/ NH(Asn4) in combination to the HMBC correlation Hα(Pro3), NH(Asn4)/ CO(Pro3) unambiguously assigned the position of the residues Pro3 and Asn4. Finally, the positions of Pro5 and of Ile6 were defined on the basis of the NOESY correlations between Hα (Asn4)/ Hδ (Pro5), Hα (Pro5)/ NH(Ile6) and of the HMBC correlations between Hα, NH (Ile6)/ CO(Pro5). Consequently, the presence of the linear peptide N-Asp-Ile-Pro-Asn-Pro-Ile was assumed. Further ESI-MS/MS analysis confirmed our hypothesis. Indeed, it showed characteristic ions at m/z 551.3, 438.2, 324.1, 341.1, 227.1, and 130.9 corresponding to the fragmentation of the five peptide bonds composing the hexapeptide. (Figure S3).
The absolute configuration of the amino acid residues was determined by performing Marfey’s analysis [20]. The hydrolysis of compound 1 with hydrochloric acid (HCl) 6N and the derivatization of the hydrolysate with 1-fluoro-2,4-dinitrophenyl-5-L-valinamide (L-FDVA) disclosed that the absolute configuration of all residues was L. Amino acid derivatives were identified by comparison of their retention time with Nα-(2,4- dinitro-5-fluorophenyl)-L-leucinylamide (FDLA) derivatized D- and L-amino acid standards. Accordingly, the absolute configuration of compound 1 was elucidated as L-Asp-L-Ile-L-Pro-L-Asn-L-Pro-L-Ile.
In addition, bioassay-guided fractionation led to the isolation of three known compounds, namely cyclo-(L-Pro-L-Phe) (2) [22], cyclo-(L-Pro-L-Tyr) (3) [23] and 2-hydroxyphenylacetic acid (4) [24]. Their structures were established on the basis of NMR and HRMS data and by comparison of their spectroscopic and physical data with the literature.
All isolated compounds were tested for their ability to inhibit tyrosinase. Compound 3 (IC50 =28.69 μΜ) exhibited the highest inhibitory activity with an IC50 value comparable to that of the positive control kojic acid (IC50 =14.07 μΜ). Its potential use as a tyrosinase inhibitor was further supported by its lack of any cytotoxic effect [25,26,27]. A moderate activity was also found for compound 4 (IC50 =98.29 μΜ) while no inhibitory effect was confirmed for compounds 1 and 2 (IC50 >300 μM). The inhibitory capacity of compounds 3 and 4 was statistically significant (P<0.01) vs. control samples. To our knowledge, this is the first report on investigating the anti-tyrosinase effect of compounds 3 and 4 using L-3,4-dihydroxyphenylalanine (L-DOPA) as substrate [28]. Compound 1 represent a class of peptides with high resistance to proteolysis and capable of immune modulation and of other therapeutic activities, including but not limited to the protection against viral infection, in growth stimulation and in normalizing serum cholesterol levels [29,30,31]. During our analysis, we screened compound 1 against a panel of cancer cell lines. No cytotoxic effect against the cancer cell lines HepG2, A2058, A549 and MiaPaca-2 was detected at the highest concentration used. Consideration that compound 1 belongs to a class of peptides that are capable of surviving gastrointestinal digestion, the lack of cytotoxicity can be interpreted as a positive outcome in view of its potential therapeutic activities.
Compound (1)
Brownish solid; UV (MeOH) λmax only end absorption detected; IR vmax 3293, 3286, 2965, 1722, 1644, 1632, 1537, 1448, 1314, 1254, 1220, 1024, 953 cm-1; 1H (500 MHz) and 13C (125 MHz) NMR spectral data in DMSO-d6, see Table S1; NMR homo- and hetero-correlations in DMSO-d6, see Table S2; HRESIMS m/z: 668.3605 [M + H]+ (calcd for C30H50N7O10, 668.3619).
4. Conclusions
In conclusion, the strain CA-287887, identified as Actinomycetospora sp., demonstrated the most significant tyrosinase inhibitory activity among other extracts of endophytic actinomycetes. The bio-guided fractionation led to isolation and the proteolytic resistant hexapeptide αs1-CN 181–186 (1), described for the first time in microbial cultures, along with three known compounds, namely cyclo-(L-Pro-L-Phe) (2), cyclo-(L-Pro-L-Tyr) (3) and 2-hydroxyphenylacetic acid (4). A complete structural elucidation was carried out for compound 1, including NMR and HRMS/MS analyses, along with the establishment of the absolute configuration. A wide cytotoxic screening against a panel of cancer cell lines was also caried out. Regarding the whitening activity, compounds 3 and 4 exhibited the highest inhibitory activity against tyrosinase enzyme with IC50 values of 28.69 and 98.29 μΜ, respectively, explaining the significant whitening effect that the extract demonstrated during the high throughput screening.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Formal analysis, K. Georgousaki, N. Tsafantakis, I. Gonzalez, J.Martín, T. Mackenzie and S.Gumeni; Investigation, K. Georgousaki, I. Gonzalez and F. Reyes; Methodology, K. Georgousaki, T. Mackenzie and F. Reyes; Writing – original draft, K.Georgousaki and N.Tsafantakis; Writing – review & editing, N. Tsafantakis, I.Trougakos, O. Genilloud and N. Fokialakis. Supervision, O. Genilloud and N. Fokialakis; Conceptualization, I. Trougakos, O. Genilloud and N. Fokialakis; Funding acquisition, N. Fokialakis;.
Acknowledgments
This work has been financially supported by the Hellenic Foundation for Research and Innovation (HFRI) and the General Secretariat for Research and Technology (GSRT), under the HFRI PhD Fellowship grant (GA. no. 2369); Part of this work has been supported by EU in the frame of MICROSMETICS project (FP7-PEOPLE- Industry-Academia Partnerships and Pathways), Grant agreement No. 612276.
Conflicts of Interest
The authors declare that they have no conflict of interest.
References
- Jiang, Y.; Wiese, J.; Tang, S.-K.; Xu, L.-H.; Imhoff, J.F.; Jiang, C.-L. Actinomycetospora chiangmaiensis gen. nov., sp. nov., a New Member of the Family Pseudonocardiaceae. Int. J. Syst. Evol. Microbiol. 2008, 58, 408–413. [Google Scholar] [CrossRef] [PubMed]
- Yamamura, H.; Ashizawa, H.; Nakagawa, Y.; Hamada, M.; Ishida, Y.; Otoguro, M.; Tamura, T.; Hayakawa, M. Actinomycetospora iriomotensis sp. nov., a Novel Actinomycete Isolated from a Lichen Sample. J. Antibiot. 2011, 64, 289–292. [Google Scholar] [CrossRef] [PubMed]
- Tamura, T.; Ishida, Y.; Hamada, M.; Otoguro, M.; Yamamura, H.; Hayakawa, M.; Suzuki, K.-I. Description of Actinomycetospora chibensis sp. nov., Actinomycetospora chlora sp. nov., Actinomycetospora cinnamomea sp. nov., Actinomycetospora corticicola sp. nov., Actinomycetospora lutea sp. nov., Actinomycetospora straminea sp. nov., and Actinomycetospora succinea sp. nov. and Emended Description of the Genus Actinomycetospora. Int. J. Syst. Evol. Microbiol. 2011, 61, 1275–1280. [Google Scholar] [PubMed]
- Zhang, Y.; Liu, C.; Zhang, J.; Shen, Y.; Li, C.; He, H.; Wang, X.; Xiang, W. Actinomycetospora atypica sp. nov., a Novel Soil Actinomycete and Emended Description of the Genus Actinomycetospora. Antonie van Leeuwenhoek 2014, 105, 891–897. [Google Scholar] [CrossRef]
- He, H.; Zhang, Y.; Ma, Z.; Li, C.; Liu, C.; Zhou, Y.; Li, L.; Wang, X.; Xiang, W. Actinomycetospora rhizophila sp. nov., an Actinomycete Isolated from Rhizosphere Soil of a Peace Lily (Spathiphyllum Kochii). Int. J. Syst. Evol. Microbiol. 2015, 65, 1520–1524. [Google Scholar] [CrossRef]
- Fu, P.; MacMillan, J.B. Thiasporines A–C, Thiazine and Thiazole Derivatives from a Marine-Derived Actinomycetospora chlora. J. Nat. Prod. 2015, 78, 548–551. [Google Scholar] [CrossRef]
- Sakdapetsiri, C.; Ngaemthao, W.; Suriyachadkun, C.; Duangmal, K.; Kitpreechavanich, V. Actinomycetospora endophytica sp. nov., Isolated from Wild Orchid (Podochilus microphyllus Lindl.) in Thailand. Int. J. Syst. Evol. Microbiol. 2018, 68, 3017–3021. [Google Scholar] [CrossRef]
- Kaewkla, O.; Franco, C.M.M. Actinomycetospora callitridis sp. nov., an Endophytic Actinobacterium Isolated from the Surface-Sterilized Root of an Australian Native Pine Tree. Antonie van Leeuwenhoek 2019, 112, 331–337. [Google Scholar] [CrossRef]
- Pillaiyar, T.; Manickam, M.; Namasivayam, V. Skin Whitening Agents: Medicinal Chemistry Perspective of Tyrosinase Inhibitors. J. Enzyme Inhib. Med. Chem. 2017, 32, 403–425. [Google Scholar] [CrossRef]
- Martín, J.; Crespo, G.; González-Menéndez, V.; Pérez-Moreno, G.; Sánchez-Carrasco, P.; Pérez-Victoria, I.; Ruiz-Pérez, L.M.; González-Pacanowska, D.; Vicente, F.; Genilloud, O.; et al. MDN-0104, an Antiplasmodial Betaine Lipid from Heterospora chenopodii. J. Nat. Prod. 2014, 77, 2118–2123. [Google Scholar] [CrossRef]
- Coombs, J.T.; Franco, C.M.M. Isolation and Identification of Actinobacteria from Surface-Sterilized Wheat Roots. Appl. Environ. Microbiol. 2003, 69, 5603–5608. [Google Scholar] [CrossRef]
- Georgousaki, K.; Tsafantakis, N.; Gumeni, S.; Gonzalez, I.; Mackenzie, T.A.; Reyes, F.; Lambert, C.; Trougakos, I.P.; Genilloud, O.; Fokialakis, N. Screening for Tyrosinase Inhibitors from Actinomycetes; Identification of Trichostatin Derivatives from Streptomyces sp. CA-129531 and Scale Up Production in Bioreactor. Bioorg. Med. Chem. Lett. 2020, 30, 126952. [Google Scholar] [CrossRef]
- Yoon, S.-H.; Ha, S.-M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A Taxonomically United Database of 16S rRNA Gene Sequences and Whole-Genome Assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef]
- EzBioCloud. Available online: https://www.ezbiocloud.net/ (accessed on 1 June 2019).
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X Windows Interface: Flexible Strategies for Multiple Sequence Alignment Aided by Quality Analysis Tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The Neighbor-Joining Method: A New Method for Reconstructing Phylogenetic Trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Jukes, T.H.; Cantor, C.R. Evolution of Protein Molecules. In Mammalian Protein Metabolism [Internet]; Elsevier: [place unknown], 1969; pp 21–132.
- Santos, J.D.; Vitorino, I.; De la Cruz, M.; Díaz, C.; Cautain, B.; Annang, F.; Pérez-Moreno, G.; Gonzalez Martinez, I.; Tormo, J.R.; Martín, J.M.; et al. Bioactivities and Extract Dereplication of Actinomycetales Isolated from Marine Sponges. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef]
- Marfey, P. Determination of D-Amino Acids. II. Use of a Bifunctional Reagent, 1,5-Difluoro-2,4-Dinitrobenzene. Carlsberg Res. Commun. 1984, 49, 591. [Google Scholar] [CrossRef]
- Chaita, E.; Lambrinidis, G.; Cheimonidi, C.; Agalou, A.; Beis, D.; Trougakos, I.; Mikros, E.; Skaltsounis, A.-L.; Aligiannis, N. Anti-Melanogenic Properties of Greek Plants. A Novel Depigmenting Agent from Morus alba Wood. Molecules 2017, 22, 514. [Google Scholar] [CrossRef]
- Ye, F.; Cai, M.-H.; Chen, B.; Xiao, W.; Li, X.-W.; Guo, Y.-W. Absolute Configuration of (2R,3R,6S,8R)-Methyl Homononactate, a Polyketide from Actinomycetes Streptomyces sp. R-527F of the Arctic Region. Chem. Nat. Compd. 2018, 54, 821–825. [Google Scholar] [CrossRef]
- Jayatilake, G.S.; Thornton, M.P.; Leonard, A.C.; Grimwade, J.E.; Baker, B.J. Metabolites from an Antarctic Sponge-Associated Bacterium, Pseudomonas aeruginosa. J. Nat. Prod. 1996, 59, 293–296. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Khan, A.L.; Ali, L.; Rizvi, T.S.; Khan, S.A.; Hussain, J.; Hamayun, M.; Al-Harrasi, A. Enzyme Inhibitory Metabolites from Endophytic Penicillium citrinum Isolated from Boswellia sacra. Arch. Microbiol. 2017, 199, 691–700. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.; Ku, S.-K.; Choi, H.; Bae, J.-S. Three Diketopiperazines from Marine-Derived Bacteria Inhibit LPS-Induced Endothelial Inflammatory Responses. Bioorg. Med. Chem. Lett. 2016, 26, 1873–1876. [Google Scholar] [CrossRef] [PubMed]
- Buedenbender, L.; Robertson, L.P.; Lucantoni, L.; Avery, V.M.; Kurtböke, D.İ.; Carroll, A.R. HSQC-TOCSY Fingerprinting-Directed Discovery of Antiplasmodial Polyketides from the Marine Ascidian-Derived Streptomyces sp. (USC-16018). Marine Drugs 2018, 16, 189. [Google Scholar] [CrossRef]
- Solecka, J.; Rajnisz-Mateusiak, A.; Guspiel, A.; Jakubiec-Krzesniak, K.; Ziemska, J.; Kawęcki, R.; Kaczorek, D.; Gudanis, D.; Jarosz, J.; Wietrzyk, J. Cyclo(Pro-DOPA), a Third Identified Bioactive Metabolite Produced by Streptomyces sp. 8812. J. Antibiot. 2018, 71, 757. [Google Scholar] [CrossRef]
- Lee, J.; Hwang, I.H.; Kim, J.H.; Kim, M.-A.; Hwang, J.S.; Kim, Y.H.; Na, M. Quinoxaline-, Dopamine-, and Amino Acid-Derived Metabolites from the Edible Insect Protaetia brevitarsis seulensis. Arch. Pharm. Res. 2017, 40, 1064–1070. [Google Scholar] [CrossRef]
- Sidelman, Z. Casein Derived Peptides and Therapeutic Uses Thereof (US54840104P·2004-03-01).
- Picariello, G.; Ferranti, P.; Fierro, O.; Mamone, G.; Caira, S.; Di Luccia, A.; Monica, S.; Addeo, F. Peptides Surviving the Simulated Gastrointestinal Digestion of Milk Proteins: Biological and Toxicological Implications. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2010, 878, 295–308. [Google Scholar] [CrossRef]
- Zhang, Q.; Ren, J.; Zhao, M.; Zhao, H.; Regenstein, J.M.; Li, Y.; Wu, J. Isolation and Characterization of Three Novel Peptides from Casein Hydrolysates that Stimulate the Growth of Mixed Cultures of Streptococcus thermophilus and Lactobacillus delbrueckii subsp. bulgaricus. J. Agric. Food Chem. 2011, 59, 7045–7053. [Google Scholar] [CrossRef]
Figure 1.
Structure of compound 1.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



