
Submitted:
18 January 2025
Posted:
20 January 2025
You are already at the latest version
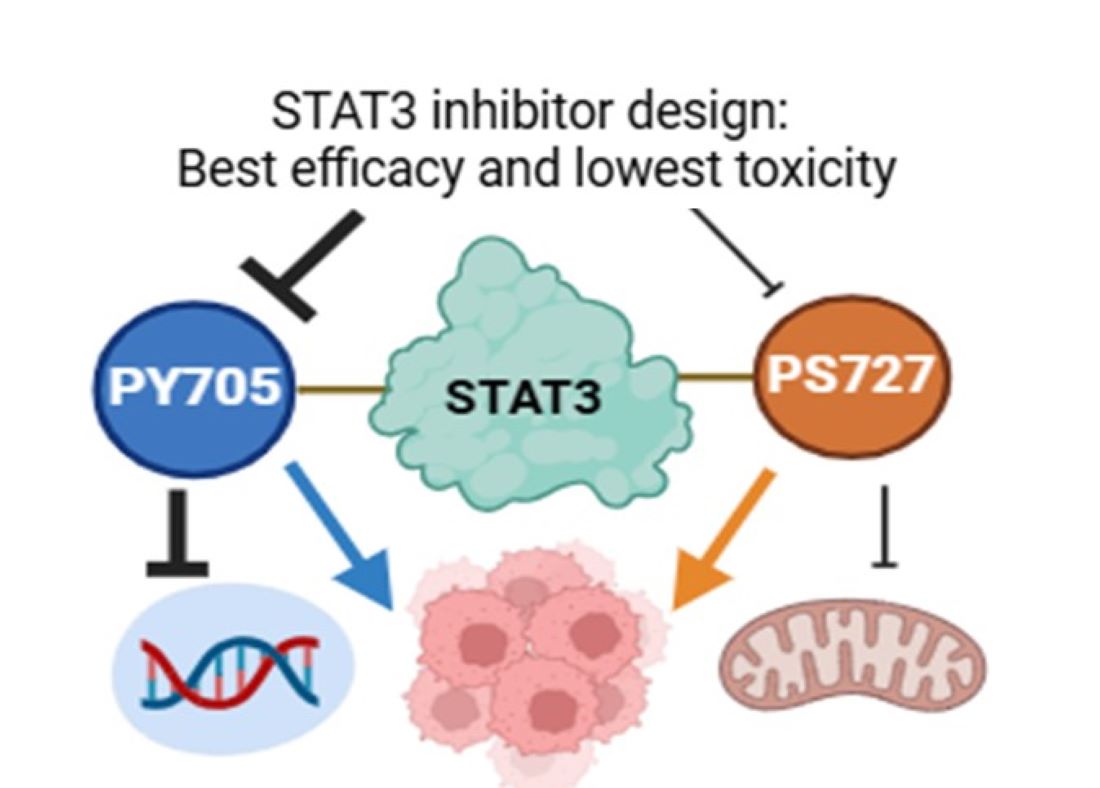
Abstract
Background/Objectives: Signal Transducer and Activator of Transcription 3 (STAT3) is a transcription factor strongly implicated in various cancers. In its canonical signaling pathway, Janus kinases (JAKs) phosphorylate STAT3 at the Y705 residue in response to cytokines or growth factors, with pY705 serving as a key marker of STAT3 oncogenic ac-tivity. Elevated pY705 levels correlate with poor prognosis, and numerous small-molecule inhibitors have been developed to block this phosphorylation site. More recently, phos-phorylation at the S727 residue (pS727) has emerged as a critical contributor to STAT3-mediated oncogenesis, particularly due to its role in mitochondrial translocation. Evidence suggests that pS727 may even surpass pY705 in driving oncogenic activity. These findings prompt an important question: Which residue should be prioritized for ef-fective STAT3 inhibition in cancer therapy? Methods: This review compiles and critically analyzes current literature on STAT3 inhibitors targeting pY705 and/or pS727, evaluating their therapeutic efficacy in vitro, in vivo, and in clinical trials. We assess the unique ef-fects of targeting each residue on downstream signaling, toxicity, and clinical outcomes. Results: Our analysis indicates that inhibitors targeting both pY705 and pS727 achieve the greatest therapeutic effectiveness. However, pS727 targeting is associated with higher tox-icity risks. Conclusions: Comprehensive evaluation of STAT3 inhibitors underscores the importance of targeting both pY705 and pS727 for maximum therapeutic benefit. None-theless, pS727 inhibition should be approached with lower affinity to minimize toxic side effects and enhance the clinical feasibility of dual-targeting strategies.
Keywords:
STAT3
; cancer therapy
; STAT3 inhibitor
; Y705
; S727
1. Introduction
The Signal transducer and activator of transcription 3 (STAT3) is a transcription factor that plays a pivotal role in various cellular processes, including cell growth, differentiation, survival, and immune responses [1,2]. Normally, STAT3 activation is transient and tightly regulated in healthy cells; however, in many cancers, STAT3 is constitutively active, thus promoting cell proliferation, invasion, drug resistance, and immune cell evasion [1,3,4,5,6]. Basal-level STAT3 signaling is necessary for regular cell maintenance, as is evidenced by the fact that STAT3 knockouts proved lethal in embryonic mouse studies [7,8]. However, in cancer, hyperphosphorylated STAT3 activates oncogenic transcription factors such c-Myc and Oct-4, leading to enhanced cell proliferation and survival as well as aggressive cancer cell stemness [9,10,11]. This aberrant activation has established STAT3 as a critical target for cancer therapy due to its involvement in tumor progression and association with poor disease prognosis.
Inactive STAT3 proteins are dispersed in the cytoplasm until being recruited to the plasma membrane in response to cytokine or growth factor signals where they are phosphorylated by upstream protein kinases such as Janus kinase proteins (JAKs), proto-oncogene tyrosine-protein kinase Src (Src), and mitogen-activated protein kinases (MAPKs) (Figure 1) [12,13]. Once phosphorylated at the Y705 site, STAT3 dimerizes with itself or other STAT family members and travels to the nucleus where it initiates gene transcription [14]. Recently, studies have shown that S727 also undergoes phosphorylation and is necessary for mitochondrial translocation, but whether or not pS727 should be inhibited to improve cancer therapy is debated [15,16,17,18,19].
Due to its central role in oncogenesis, STAT3 has garnered significant interest as a therapeutic target. Numerous efforts have been made over the last twenty years to develop small molecules to inhibit the STAT3 signaling axis; however, none have reached FDA approval for cancer treatment, underscoring the need for a better understanding of oncogenic STAT3 signaling mechanisms in order to develop an effective therapeutic approach [1,6,9,20]. Small molecule inhibitors, such as napabucasin and OPB-51602, aim to block STAT3 activation directly or indirectly. Additionally, antisense oligonucleotides, peptides, and other modalities are being investigated for STAT3 inhibition. Current research also suggests selectively targeting Y705 or S727, or both, could differentially affect downstream oncogenic signaling. The dual activation of STAT3 at pY705 and pS727 allows STAT3 to promote oncogenic signaling both in the nucleus and in mitochondria. However, there have been mixed results as to whether both pY705 and pS727 should be targeted for effective cancer treatment [21,22]. Thus, the purpose of this review is to synthesize the current findings in this field to inform current STAT3 therapeutic development with respect to pY705 versus pS727 inhibition.
2. STAT3 Signaling Mechanisms
2.1. Canonical STAT3 Signaling and the Role of Y705 Phosphorylation
To initiate downstream signaling, STAT3 is activated in response to extracellular signals, such as cytokines (e.g., IL-6, IL-10) and growth factors (e.g., EGF), through the Janus kinase (JAK) pathway. Latent STAT3 molecules are dispersed in the cytoplasm prior to activation [11]. Upon ligand binding to cytokine receptors, JAK proteins phosphorylate STAT3 at the tyrosine-705 (Y705) residue, prompting dimerization, nuclear translocation, and subsequent binding to DNA sequences to initiate transcription of target genes involved in cell cycle progression and survival. While STAT3 activation can occur in response to various cytokines and growth factors, interleukin-6 (IL-6) is the most well-known activator of STAT3 signaling [23,24]. IL-6 binds to the IL-6 receptor-α (IL-6Rα) on the cell surface which induces conformational changes, leading to the formation of a hexameric signaling complexing including a gp130 homodimer and two IL-6-IL-6Rα heterodimers [23]. This results in Janus kinase (JAK) activation, which are tyrosine kinases that are constitutively associated with the proline-rich cytoplasmic domain of gp130 near the plasma membrane.[13,23] In total, there are four members of the JAK family: JAK1, JAK2, JAK3, and TYK2.[25] Then, activated JAKs cause gp130 to become phosphorylated which recruits cytosolic STAT3 for phosphorylation. This STAT3 signaling cascade can also be initiated by epidermal growth factor (EGF) and interleukin-11 (IL-11)[24,26]. In all, the JAK-STAT pathway can be activated by over 50 different cytokine receptors leading to modulation of cell cycle progression, the immune response, and inflammation [25,27].
While JAK-mediated signaling can affect any of the STAT protein family members (STAT1, STAT2, STAT3, STAT4, STAT5α, STAT5β, or STAT6), STAT3 has been the most heavily implicated in oncogenesis and cancer progression [4,11,23]. STAT3 signaling is necessary for regular cell maintenance, as is evidenced by the fact that STAT3 knockouts proved lethal in embryonic mouse studies [7]. However, in cancer cells, hyperphosphorylated STAT3 activates oncogenic transcription factors such c-Myc, Cyclin-D1, Bcl-xL, VEGF, and Oct-4, leading to enhanced cell proliferation and survival as well as aggressive cancer cell stemness [9,10,11]. STAT3 also regulates transcription factors Sox2 and NANOG which contribute to cancer cell stemness and TWIST1 and Vimentin which promote the epithelial-mesenchymal (EMT) transition to occur [28,29]. EMT is a hallmark of cancer metastasis and is a necessary transition for cancer cells to undergo in order to travel to distant sites [28,30].
2.2. Noncanonical STAT3 Signaling and the Role of S727 Phosphorylation
Even though pY705 has traditionally been measured as the marker of oncogenic STAT3 activities, accumulating studies have highlighted the importance of serine-727 (S727) phosphorylation, which can enhance STAT3 transcriptional activity and influence mitochondrial functions [15]. S727 phosphorylation occurs through various signaling pathways, including the MAPK, mTOR, and CDK pathways, often in response to stress signals or mitochondrial activity. Unlike Y705 phosphorylation, which primarily promotes transcriptional activity within the nucleus, S727 phosphorylation has been shown to enhance STAT3's association with mitochondria, where it regulates cellular respiration and metabolic functions. Since pS727 allows STAT3 translocation to the mitochondria, pS727 affects the electron transport chain and contributes to the Warburg effect on cell metabolism [12,31]. The Warburg effect describes how cancer cells can rewire their metabolism to support high rates of growth and proliferation by relying on high glucose uptake that results in the production of lactate [32].
Recent studies indicate that pS727 may influence STAT3's oncogenic potential by supporting mitochondrial biogenesis and modulating reactive oxygen species (ROS) production, which can contribute to tumor progression under hypoxic conditions. This mitochondrial role of STAT3, termed "mitochondrial STAT3" or mitoSTAT3, affects cellular bioenergetics, which is essential for the survival and growth of cancer cells in stressful microenvironments. In some cancers, mitoSTAT3 activity driven by pS727 has been linked to increased tumor invasiveness and resistance to apoptosis, particularly in response to chemotherapy. Despite the fact that recent studies have shown that pS727 is necessary for mitochondrial translocation, it is debated whether or not pS727 should be inhibited to improve cancer therapy due to clinical effectiveness and possible toxicities [15,16,17,18,19].
2.3. Interplay Between pY705 and pS727 in Cancer
Initially, it was believed that phosphorylation at both the Y705 and S727 sites were necessary for STAT3 activation, and that S727 phosphorylation occurred sequentially after Y705 phosphorylation [33]. However, accumulating evidence has shown that S727 can be phosphorylated independently of Y705 and is capable of driving oncogenic activities by itself, thus validating it as an independent therapeutic target [15,16,17,34].
Additionally, phosphorylation of both residues has demonstrated both complementary and antagonistic effects. For, it has been shown that pY705 is necessary for import inside of the mitochondria while pS727 is needed for mitochondrial transcriptional activity [17]. In this case, phosphorylation at both residues works together to yield downstream effects such as stem cell proliferation. In a separate study on pancreatic cancer, it was discovered that pS727 decreases in response to increased phosphorylation at Y705 [35]. This interplay is of great interest for cancer therapeutics inhibition of either Y705 or S727 yields distinct biological outcomes. Targeting Y705 alone may reduce STAT3’s nuclear transcriptional activity, while inhibiting S727 phosphorylation could impair mitochondrial functions critical for cancer cell metabolism. Consequently, dual-targeting strategies, which inhibit both phosphorylation sites, are being explored to comprehensively block STAT3’s oncogenic functions.
3. Clinical Relevance of STAT3 in Cancer and Rational for Therapeutic Development
STAT3 is highly implicated in the progression and survival of various cancers, establishing it as a key target in oncology. The signal transducer and activator of transcription (STAT) family consists of six members, of which STAT3 has been the most implicated in cancer progression [4,11,23]. Aberrant activation of STAT3 has been reported in numerous cancers, including breast, lung, liver, colorectal, pancreatic, and hematological malignancies [9,36]. STAT3's role in promoting cell proliferation, inhibiting apoptosis, and modulating the immune response makes it central to oncogenesis and tumor progression, particularly in cancers where STAT3 activation correlates with poor prognosis, treatment resistance, and increased metastasis [14,23].
3.1. STAT3 Hyperactivation in Various Cancers Is Associated with Poor Prognosis
In many cancers, STAT3 is constitutively activated, either through cytokine receptor signaling or through mutations in upstream pathways [21,36]. For example, in breast cancer, STAT3 is frequently activated by interleukin-6 (IL-6) through the Janus kinase (JAK) pathway, promoting an inflammatory microenvironment that fosters tumor growth [3,4,11]. Zhong et al. showed that STAT3 was upregulated in gastric tumors as well as triple negative breast cancer (TNBC) and that this upregulation correlated with lower survival overall [18]. Several other studies have supported the observation that increased STAT3 activity corresponds with poorer breast cancer prognosis, especially in TNBC [30,37,38,39].
In lung cancer, STAT3 activation is associated with enhanced metastatic potential, aiding in epithelial-to-mesenchymal transition (EMT), which allows cancer cells to spread to distant organs [40]. Due to its role in EMT, constitutively activated STAT3 has been correlated with poor prognosis in non-small cell lung cancer [41]. In glioblastoma and hepatocellular carcinoma, constitutive STAT3 signaling has been shown to support angiogenesis through the upregulation of vascular endothelial growth factor (VEGF) [42]. Several studies have also shown that upregulated STAT3 contributes to poor prognosis for patients with glioblastoma, especially due to the role STAT3 plays in immune evasion [43,44]. Hepatocellular carcinoma progression has also been shown to be worsened by STAT3 overactivation [45,46]. Additionally, STAT3 hyperactivation has been implicated in cancer progression and poor prognosis in pancreatic cancer,[47,48] prostate cancer,[49] colorectal,[50] and esophageal cancer [51,52]. In all, STAT3 has been shown to be an appealing pan-cancer therapeutic target [53].
3.2. Rationale for Targeting STAT3 in Cancer
There are three main signaling axis implicated in cancer progression which have been heavily targeted for therapeutic development: PI3K/AKT, RAS/RAF/MEK, and JAK/STAT. While numerous PI3K inhibitors have been developed, they have been met with various complications upon reaching clinical trials. For instance, pan-PI3K inhibitors, which act on all four PI3K isoforms, have demonstrated a lack of clinical effectiveness with only copanlisib showing significant effect in clinical trials [54,55]. However, pan-PI3K inhibitors also greatly increase the risk of off-target toxicity. Capivasertib, a pan-AKT inhibitor was approved by the FDA in November 2023 for treating HR-positive, HER2-negative metastatic breast cancer with at least one alteration in PIK3/CA/AKT1/PTEN genes in combination with fulvestrant [56,57].
In general, PI3K-AKT inhibitors are prone to side effects, especially hyperglycemia (high blood sugar), which occurs in up to 80% of patients in clinical trials.[58] Other side effects of PI3K-AKT inhibitors include diarrhea, colitis, nausea, and rash [58,59,60]. Thus, further development in the field of PI3K-AKT inhibitors is needed to mitigate these debilitating side effects.
Similar to the PI3K/AKT story, many inhibitors have been developed targeting the RAS/RAF/MEK pathway. Two examples are Sorafenib, which mainly inhibits RAF but has since been shown to inhibit other targets as well, and Trametinib, an MEK inhibitor [61]. One of the challenges to targeting this pathway is the prevalence of mutations, and a large breakthrough in this area occurred in 2022 with the development of a covalent inhibitor of KRAS(G12C) [62]. It has been found that RAF inhibitors combined with MEK inhibitors give the most promising clinical results, but drug resistance remains a large problem in this field of inhibitor development [63].
With respect to the JAK/STAT signaling pathway, STAT3 stands out as a therapeutic target since it is downstream of the JAK proteins, which means targeting STAT instead of JAK should cause less off-target side effects. Additionally, STAT3's involvement in promoting tumorigenesis and resistance to therapy also supports the hypothesis that targeting STAT3 is a promising therapeutic approach for cancer therapy [20,36]. STAT3 modulates numerous pathways critical for tumor survival, including apoptosis inhibition, immune evasion, angiogenesis, and EMT [21,29]. Therefore, inhibiting STAT3 could provide multifaceted benefits by disrupting these cancer-supportive mechanisms simultaneously. First of all, due to its role as a transcription factor, targeting STAT3 also inhibits downstream oncogenic gene expression including targets such as c-Myc, cyclin D1, and survivin. By targeting STAT3, these downstream oncogenic pathways can be suppressed, potentially reducing cell proliferation and inducing apoptosis across cancer types.
STAT3 not only drives oncogenic signaling but also contributes to chemoresistance, a major obstacle in cancer therapy. In TNBC and esophageal squamous cell carcinoma, for instance, STAT3 activation has been correlated with resistance to chemotherapy agents mediated by STAT3's regulation of anti-apoptotic proteins such as Bcl-2 and Bcl-xL [64]. Similarly, in head and neck cancers, STAT3 signaling confers resistance to platinum-based therapies, diminishing the efficacy of standard treatments [65]. The association between STAT3 activation and chemoresistance highlights its importance as a therapeutic target, as inhibiting STAT3 could enhance cancer cell sensitivity to existing treatments. These findings indicate that STAT3's oncogenic activities play a significant role in cancer progression, invasion, and metastatic potential, supporting its status as a prognostic marker and a therapeutic target.
STAT3 plays a significant role in shaping the immunosuppressive tumor microenvironment (TME), contributing to immune evasion by upregulating immune-suppressive cytokines (e.g., IL-10, TGF-β) and downregulating pro-inflammatory cytokines [66]. Inhibiting STAT3 could restore immune function within the TME, making it more susceptible to immune-mediated cell death [67].
Additionally, STAT3 is critical for the maintenance of CSCs, which are implicated in cancer recurrence and metastasis. In cancers such as glioblastoma and pancreatic cancer, STAT3 activation supports CSC properties, including self-renewal and resistance to conventional therapies [10,68,69,70]. Targeting STAT3 in these cells could reduce tumor recurrence and improve patient outcome. For example, he small molecule STAT3 inhibitor napabucasin has been shown to attenuate tamoxifen resistance of breast cancer cells by reducing CSC properties [71].
Finally, STAT3 inhibitors are attractive therapeutics because STAT3 inhibition have been shown to synergize with other therapeutic modalities, such as immunotherapy, chemotherapy, and target therapies to enhance their effectiveness [52,72,73]. Combining STAT3 inhibitors with immune checkpoint inhibitors, for example, has shown promise in preclinical studies, where STAT3 inhibition sensitizes tumors to immune cell infiltration and attack [74].
4. Evaluation of STAT3 Inhibitors of Y705 and/or S727 in Clinical Development
Stattic was the first developed STAT3 inhibitor and prevents STAT-STAT dimerization, however its high toxicity prevented it from progressing to clinical trials of any kind [75]. Despite the diversity of STAT3 inhibitors developed to date, many candidates have shown poor bioavailability, structural instability, and off-target toxicity upon reaching clinical trials (Figure 2) [9,76,77,78,79].
Napabucasin (BBI608) is a small molecule STAT3 inhibitor that has been shown to reduce breast cancer [80,81] as well as glioblastoma [69] progression by mediating STAT3 signaling. Napabucasin has been said to bind to the STAT3 SH2 domain but currently there is lack of definitive crystallographic evidence [82]. That being said, napabucasin has been shown to block both pY705 and S727 [71]. Napabucasin was granted orphan drug status in 2016 by the FDA for gastric cancer while it was also being explored for several other cancers [83]. A Phase Ib/II study in TNBC showed that napabucasin combined with paclitaxel was well-tolerated and showed promising results indicating anticancer-activity [81]. Conversely, napabucasin was discontinued in Phase III clinical trials for pancreatic cancer due to futility with no additional safety concerns listed [84].
SLSI-1216 has been developed for targeting STAT3 in TNBC and has been shown SH2 domain, specifically inhibiting EMT and reducing both pY705 and pS727 [30]. SLSI-1216 has not yet progressed to clinical trials.
C188-9 (TTI-101) has been studied as a STAT3 inhibitor in head and neck squamous cell carcinoma that specifically inhibits pY705 to reduce migration and invasion as well as increase chemosensitivity in vitro [76]. C188-9 targets the SH2 domain and has been shown to sensitize pancreatic cells to 5-Aza-2′-deoxycytidine (DAC) treatment in mice [85]. Activity against pY705 has been measured in acute myeloid leukemia, and there is little evidence for the effect on pS727 [86]. C188-9 was studied in a Phase 1b/2 clinical trial in advanced hepatocellular carcinoma in which it was well-tolerated and displayed clinical benefits in about 60% of patients as a monotherapy [87].
InS3-54 is a small molecule STAT3 inhibitor that targets the DNA-binding domain; as such, it did not inhibit pY705 activation or total STAT3 expression, yet reduced STAT3 activity by disrupting DNA binding, leading to decreased expression of downstream genes [88]. N4 targets the STAT3 SH2 domain in pancreatic cancer and selectively blocks pY705 over pS727 [89]. N4 effectively inhibited STAT3 dimerization as well as cross-talk with EGFR and NF-κB, and animal models showed that N4 was well tolerated and effectively suppressed tumor growth and metastasis. There is currently no clinical trial data available for N4.
YY002 is another small molecule therapeutics for pancreatic cancer that targets the STAT3 SH2 domain, but unlike N4, it blocks both pY705 and pS727 [90]. This dual blockage disrupts both nuclear and mitochondrial functions of STAT3. YY002 displayed good pharmacokinetics as well as an acceptable safety profile and displayed better antitumor efficacy compared to napabucasin. LLY17 is a small molecule inhibitor developed for TNBC targeting the STAT3 SH2 domain and decreasing pY705, but pS727 was not examined in the initial study [91]. The small molecule WZ-2-033 has been studied in TNBC and gastric cancer; it targets the STAT3 SH2 domain and specifically affects pY705 and not pS727 [18]. Currently, WZ-2-033 is still in the preclinical phase. Another small molecule inhibitor, OPB-31121, interacts with the STAT3 SH2 domain and decreases both pY705 and pS727 [92]. A phase 1 clinical trial was completed but failed in subjects with advanced solid tumors due to adverse effects and therapeutic resistance [93].
OPB-51602 is a small molecule inhibitor that binds to the SH2 domain and disrupts mitochondrial STAT3 [94]. Specific inhibition of pY705 and pS727 were not measured, but the disruption of mitochondria-STAT3 implies that pS727 is inhibited. OPB-51602 was tested in a Phase I clinical study of non-small cell lung cancer, and the results showed promising antitumor activity coupled with a poor tolerability of continuous dosing.[19] A separate Phase I study in hematological malignancies displayed no clear therapeutic response, and while it was well-tolerated at low dosages, long-term administration at higher dosages produced adverse effects [95].
Bayer’s orally available STAT3 inhibitor (VVD-130850) is currently being tested in phase I clinical trials in conjunction with or without a checkpoint inhibitor Pembrolizumab in hematologic tumors [96]. VVD-130850 binds to an allosteric pocket of STAT3 which prevents STAT3 binding to target genes.
A unique type of STAT3 inhibitor is the Novo Nordisk small interfering RNA inhibitor, DCR-STAT3 [97]. Since this inhibitor ubiquitously knocks down STAT3, it affects both pY705 and pS727. DCR-STA3T is currently being evaluated in a first-in-human study for safety and tolerability assessment in adults with refractory solid tumors. A benefit of this type of inhibitor is that it also disrupts unphosphorylated-STAT3 (U-STAT3), which has also been shown to play an oncogenic role in STAT3 signaling.[98,99,100] U-STAT3 is capable of binding to DNA, thus regulating transcription, similar to phosphorylated STAT3 [99]. Additionally, U-STAT3 has also been shown to indirectly affect NF-kB regulated gene expression by binding to NF-kb and regulating its nuclear transport, aiding the constitutive activation of NF-kB and promoting oncogenic progression [101]. Thus, especially in cancer types with hyperactive NF-kB, it may be necessary to fully remove STAT3 in order to combat U-STAT3 oncogenic pathways.
In addition to small interfering RNA inhibitors like DCR-STAT3, another novel approach to STAT3 inhibitor design that mitigates U-STAT3 signaling effects is proteolysis-targeting chimeras (PROTACs). PROTACs offer unique advantages over traditional small molecule inhibitors with respect to difficult drug targets because they can target any part of the protein for degradation, rather than a specific binding pocket.[102] Utilizing PROTACs may improve STAT3 inhibitor efficacy due to the mechanism of action which allows for PROTAC molecules to be reused within the cell following protein degradation by the cell’s own proteasomal system [103].
Overall, STAT3 inhibitor development has encountered many challenges, especially with respect to clinical translation. Despite the diversity of STAT3 inhibitors developed to date, many candidates have shown poor bioavailability, structural instability, and off-target effects upon reaching clinical trials in addition to therapeutic resistance an ineffectiveness [9,76,77,78,79].
Based on the inhibitors discussed above, toxicity is one of the greatest challenges faced in STAT3 inhibitor development. This observation is not unsurprising given that it has been shown that STAT3 is a necessary gene for embryonic development in mice, indicating that STAT3 plays crucial regulatory roles in regular cell growth and homeostasis [7]. Thus, any inhibitor of nuclear STAT3 risks targeting healthy cells in addition to cancer cells and inhibiting cell growth and proliferation. Global inhibition of STAT3 negatively affects mitochondrial and transcriptional functions across cell types, causing severe side effects. Genini et al. showed that OPB-51602 caused mitochondrial dysfunction via accumulated STAT3 aggregates leading to cell death, and that this effect was increased under glucose starvation conditions [22]. However, this demonstrated toxicity via mitochondrial-STAT3 could occur in healthy cells as well. One consideration for mitigating these toxicities is to lower the effective concentration needed to reduce the effect on healthy cells. However, this approach must not sacrifice overall drug efficacy. Another possible way of addressing these toxicities is to improve cancer cell specific targeting of STAT3 inhibitors, such as through the use of antibody-drug conjugates. Antibody-drug conjugates allow for more specific targeting of cancer cells through an antibody targeting a cancer cell specific antigen in order to deliver the drug needed to kill the cancer cells, such as a STAT3 inhibitor [104].
While significant progress has been made with respect to STAT3 inhibitors for cancer applications, more mechanistic data is needed on the role of STAT3 in cancer in order to improve the translation of pre-clinical STAT3 inhibitors to clinically-meaningful therapeutics.
5. Comparative Analysis: Targeting Y705 vs. S727 in STAT3 Inhibitor Design
As discussed in Section 2, downstream STAT3 signaling is influenced by both pY705 and pS727. In general, inhibition of pY705 leads to decreased STAT3 nuclear translocation while pS727 inhibition reduces mitochondrial STAT3, but there is also a significant amount of cross-talk between these two pathways [51,105]. Additionally, there have been mixed results as to whether these pathways occur independently or concurrently with one another [15,16,17].
Based on the results detailed in Section 4, compounds that specifically target pY705 such as N4 and WZ-2-033 have been well-tolerated in preclinical studies and have also shown good efficacy in preclinical models, but there is a lack of evidence suggesting effective clinical applications (Table 1). It is unclear whether some small molecules, such as C188-9 and LLy17, depend on inhibiting both phosphorylation sites or only the tyrosine residue due to lack of experimental investigation of pS727. If C188-9 is pY705 specific, then there is evidence of clinical benefits in about 60% of patients with advanced hepatocellular carcinoma.
To date, there is no clear inhibitor that solely targets pS727. OPB-51602 may be pS727 specific since it is implicated in blocking mitochondrial DNA, but as discussed earlier, whether or not pS727 in combination with or independently of Y705 phosphorylation is context dependent, so further mechanistic studies would need to be performed to clarify this point. That being said, OPB-51602 was discontinued in clinical trials due to adverse effects, suggesting a link between targeting pS727 and inducing toxicities in patients.
In general, compounds targeting both pY705 and pS727 have shown the most clinical success as far as efficacy, but there are discrepancies with respect to safety. For instance, napabucasin and C188-9 have been well tolerated in clinical trials, but OPB-31121 was discontinued due to adverse effects. Both compounds displayed some promising anti-tumor activity.
6. Conclusions
Our analysis highlights the critical need for preclinical studies to investigate the effects of potential therapeutics on both pY705 and pS727. Without such data, it remains challenging to compare inhibitors across various stages of development and fully evaluate the advantages and disadvantages of targeting each residue. These foundational mechanistic insights would be instrumental in predicting both toxicities and therapeutic effectiveness as the repertoire of STAT3 inhibitors expands.
Current evidence suggests that while pS727 plays a pivotal role in mitochondrial STAT3 translocation and cancer progression, excessive inhibition of pS727 could lead to undesirable toxic effects in clinical trials. Based on these findings, we propose that future STAT3 inhibitor development should prioritize small molecules targeting pY705 to minimize off-target toxicity. However, the most effective inhibitors might combine selective pY705 inhibition with moderate pS727 inhibition (at a higher IC50) to harness the therapeutic benefits of suppressing mitochondrial STAT3 activity while maintaining a favorable safety profile.
In addition, future therapeutic strategies should consider the role of unphosphorylated STAT3, particularly in cases where blocking pY705 and/or pS727 proves insufficient for achieving meaningful clinical outcomes. Exploring alternative approaches, such as small interfering RNA (siRNA), could be advantageous if these technologies can be made cancer-specific to minimize effects on healthy cells.
While STAT3 is well-established as a promising therapeutic target across multiple cancer types, significant gaps in our understanding remain. These include differences in cancer-specific STAT3 signaling and post-translational modifications. Addressing these gaps is essential for designing safe and effective STAT3 inhibitors that can achieve clinical success.
Author Contributions
A.S. and K.B. and J.Z. defined the review scope, perspective, and purpose of this study. J.Z. and K.B. conceived and crafted illustrative figures.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| EMT | Epithelieal-to-mesenchymal transition |
| JAK | Janus kinase |
| MAPK | Mitogen-activated protein kinase |
| Src | Proto-oncogene tyrosine-protein kinase Src |
| STAT3 | Signal transducer and activator of transcription 3 |
| VEGF | Vascular endothelial growth factor |
| EMT | Epithelieal-to-mesenchymal transition |
References
- Huang, Q.; Zhong, Y.; Dong, H.; Zheng, Q.; Shi, S.; Zhu, K.; Qu, X.; Hu, W.; Zhang, X.; Wang, Y. Revisiting Signal Transducer and Activator of Transcription 3 (STAT3) as an Anticancer Target and Its Inhibitor Discovery: Where Are We and Where Should We Go? European Journal of Medicinal Chemistry 2020, 187, 111922. [Google Scholar] [CrossRef]
- Li, Y.-J.; Zhang, C.; Martincuks, A.; Herrmann, A.; Yu, H. STAT Proteins in Cancer: Orchestration of Metabolism. Nat Rev Cancer 2023, 23, 115–134. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, K.; Resat, H. Constitutive Activation of STAT3 in Breast Cancer Cells: A Review: Constitutive STAT3 Activation in Breast Cancer. Int. J. Cancer 2016, 138, 2570–2578. [Google Scholar] [CrossRef] [PubMed]
- Dinakar, Y.H.; Kumar, H.; Mudavath, S.L.; Jain, R.; Ajmeer, R.; Jain, V. Role of STAT3 in the Initiation, Progression, Proliferation and Metastasis of Breast Cancer and Strategies to Deliver JAK and STAT3 Inhibitors. Life Sciences 2022, 309, 120996. [Google Scholar] [CrossRef]
- Qin, J.-J.; Yan, L.; Zhang, J.; Zhang, W.-D. STAT3 as a Potential Therapeutic Target in Triple Negative Breast Cancer: A Systematic Review. J Exp Clin Cancer Res 2019, 38, 195. [Google Scholar] [CrossRef]
- Gharibi, T.; Babaloo, Z.; Hosseini, A.; Abdollahpour-alitappeh, M.; Hashemi, V.; Marofi, F.; Nejati, K.; Baradaran, B. Targeting STAT3 in Cancer and Autoimmune Diseases. European Journal of Pharmacology 2020, 878, 173107. [Google Scholar] [CrossRef] [PubMed]
- Takeda, K.; Noguchi, K.; Shi, W.; Tanaka, T.; Matsumoto, M.; Yoshida, N.; Kishimoto, T.; Akira, S. Targeted Disruption of the Mouse Stat3 Gene Leads to Early Embryonic Lethality. Proc Natl Acad Sci U S A 1997, 94, 3801–3804. [Google Scholar] [CrossRef] [PubMed]
- Bharadwaj, U.; Kasembeli, M.M.; Robinson, P.; Tweardy, D.J. Targeting Janus Kinases and Signal Transducer and Activator of Transcription 3 to Treat Inflammation, Fibrosis, and Cancer: Rationale, Progress, and Caution. Pharmacol Rev 2020, 72, 486–526. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.-Q.; Man, Q.-W.; Huo, F.-Y.; Gao, X.; Lin, H.; Li, S.-R.; Wang, J.; Su, F.-C.; Cai, L.; Shi, Y.; et al. STAT3 Pathway in Cancers: Past, Present, and Future. MedComm 2022, 3, e124. [Google Scholar] [CrossRef]
- Cheng, C.-C.; Shi, L.-H.; Wang, X.-J.; Wang, S.-X.; Wan, X.-Q.; Liu, S.-R.; Wang, Y.-F.; Lu, Z.; Wang, L.-H.; Ding, Y. Stat3/Oct-4/c-Myc Signal Circuit for Regulating Stemness-Mediated Doxorubicin Resistance of Triple-Negative Breast Cancer Cells and Inhibitory Effects of WP1066. Int J Oncol 2018. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Qin, L.; Li, X. Role of STAT3 Signaling Pathway in Breast Cancer. Cell Commun Signal 2020, 18, 33. [Google Scholar] [CrossRef] [PubMed]
- Gough, D.J.; Koetz, L.; Levy, D.E. The MEK-ERK Pathway Is Necessary for Serine Phosphorylation of Mitochondrial STAT3 and Ras-Mediated Transformation. PLoS ONE 2013, 8, e83395. [Google Scholar] [CrossRef]
- O’Shea, J.J.; Schwartz, D.M.; Villarino, A.V.; Gadina, M.; McInnes, I.B.; Laurence, A. The JAK-STAT Pathway: Impact on Human Disease and Therapeutic Intervention. Annu. Rev. Med. 2015, 66, 311–328. [Google Scholar] [CrossRef]
- Johnston, P.A.; Grandis, J.R. STAT3 SIGNALING: Anticancer Strategies and Challenges. Mol Interv 2011, 11, 18–26. [Google Scholar] [CrossRef]
- Dimri, S.; Malhotra, R.; Shet, T.; Mokal, S.; Gupta, S.; De, A. Noncanonical pS727 Post Translational Modification Dictates Major STAT3 Activation and Downstream Functions in Breast Cancer. Experimental Cell Research 2020, 396, 112313. [Google Scholar] [CrossRef]
- Arévalo, J.; Campoy, I.; Durán, M.; Nemours, S.; Areny, A.; Vall-Palomar, M.; Martínez, C.; Cantero-Recasens, G.; Meseguer, A. STAT3 Phosphorylation at Serine 727 Activates Specific Genetic Programs and Promotes Clear Cell Renal Cell Carcinoma (ccRCC) Aggressiveness. Sci Rep 2023, 13, 19552. [Google Scholar] [CrossRef]
- Peron, M.; Dinarello, A.; Meneghetti, G.; Martorano, L.; Betto, R.M.; Facchinello, N.; Tesoriere, A.; Tiso, N.; Martello, G.; Argenton, F. Y705 and S727 Are Required for the Mitochondrial Import and Transcriptional Activities of STAT3, and for Regulation of Stem Cell Proliferation. Development 2021, 148, dev199477. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Deng, L.; Shi, S.; Huang, Q.; Ou-Yang, S.; Mo, J.; Zhu, K.; Qu, X.; Liu, P.; Wang, Y.; et al. The Novel STAT3 Inhibitor WZ-2-033 Causes Regression of Human Triple-Negative Breast Cancer and Gastric Cancer Xenografts. Acta Pharmacol Sin 2022, 43, 1013–1023. [Google Scholar] [CrossRef] [PubMed]
- Wong, A.L.; Soo, R.A.; Tan, D.S.; Lee, S.C.; Lim, J.S.; Marban, P.C.; Kong, L.R.; Lee, Y.J.; Wang, L.Z.; Thuya, W.L.; et al. Phase I and Biomarker Study of OPB-51602, a Novel Signal Transducer and Activator of Transcription (STAT) 3 Inhibitor, in Patients with Refractory Solid Malignancies. Annals of Oncology 2015, 26, 998–1005. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Wang, L.; Guan, X.; Qin, J.-J. Inhibiting STAT3 Signaling Pathway by Natural Products for Cancer Prevention and Therapy: In Vitro and in Vivo Activity and Mechanisms of Action. Pharmacological Research 2022, 182, 106357. [Google Scholar] [CrossRef] [PubMed]
- Beebe, J.D.; Liu, J.-Y.; Zhang, J.-T. Two Decades of Research in Discovery of Anticancer Drugs Targeting STAT3, How Close Are We? Pharmacology & Therapeutics 2018, 191, 74–91. [Google Scholar] [CrossRef]
- Genini, D.; Brambilla, L.; Laurini, E.; Merulla, J.; Civenni, G.; Pandit, S.; D’Antuono, R.; Perez, L.; Levy, D.E.; Pricl, S.; et al. Mitochondrial Dysfunction Induced by a SH2 Domain-Targeting STAT3 Inhibitor Leads to Metabolic Synthetic Lethality in Cancer Cells. Proceedings of the National Academy of Sciences 2017, 114, E4924–E4933. [Google Scholar] [CrossRef]
- Yu, H.; Lee, H.; Herrmann, A.; Buettner, R.; Jove, R. Revisiting STAT3 Signalling in Cancer: New and Unexpected Biological Functions. Nat Rev Cancer 2014, 14, 736–746. [Google Scholar] [CrossRef] [PubMed]
- Stat3: A STAT Family Member Activated by Tyrosine Phosphorylation in Response to Epidermal Growth Factor and Interleukin-6 | Science Available online: https://www.science.org/doi/10.1126/science.8140422 (accessed on 20 September 2024).
- Agashe, R.P.; Lippman, S.M.; Kurzrock, R. JAK: Not Just Another Kinase. Molecular Cancer Therapeutics 2022, 21, 1757–1764. [Google Scholar] [CrossRef]
- Putoczki, T.L.; Thiem, S.; Loving, A.; Busuttil, R.A.; Wilson, N.J.; Ziegler, P.K.; Nguyen, P.M.; Preaudet, A.; Farid, R.; Edwards, K.M.; et al. Interleukin-11 Is the Dominant IL-6 Family Cytokine during Gastrointestinal Tumorigenesis and Can Be Targeted Therapeutically. Cancer Cell 2013, 24, 257–271. [Google Scholar] [CrossRef] [PubMed]
- Morris, R.; Kershaw, N.J.; Babon, J.J. The Molecular Details of Cytokine Signaling via the JAK/STAT Pathway. Protein Sci 2018, 27, 1984–2009. [Google Scholar] [CrossRef]
- Wu, Y.; Diab, I.; Zhang, X.; Izmailova, E.S.; Zehner, Z.E. Stat3 Enhances Vimentin Gene Expression by Binding to the Antisilencer Element and Interacting with the Repressor Protein, ZBP-89. Oncogene 2004, 23, 168–178. [Google Scholar] [CrossRef]
- Carpenter, R.L.; Lo, H.-W. STAT3 Target Genes Relevant to Human Cancers. Cancers 2014, 6, 897–925. [Google Scholar] [CrossRef] [PubMed]
- Park, S.K.; Byun, W.S.; Lee, S.; Han, Y.T.; Jeong, Y.-S.; Jang, K.; Chung, S.-J.; Lee, J.; Suh, Y.-G.; Lee, S.K. A Novel Small Molecule STAT3 Inhibitor SLSI-1216 Suppresses Proliferation and Tumor Growth of Triple-Negative Breast Cancer Cells through Apoptotic Induction. Biochemical Pharmacology 2020, 178, 114053. [Google Scholar] [CrossRef]
- Garama, D.J.; White, C.L.; Balic, J.J.; Gough, D.J. Mitochondrial STAT3: Powering up a Potent Factor. Cytokine 2016, 87, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Liberti, M.V.; Locasale, J.W. The Warburg Effect: How Does It Benefit Cancer Cells? Trends in biochemical sciences 2016, 41, 211. [Google Scholar] [CrossRef] [PubMed]
- Wen, Z.; Zhong, Z.; Darnell, J.E. Maximal Activation of Transcription by Statl and Stat3 Requires Both Tyrosine and Serine Phosphorylation. Cell 1995, 82, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Gough, D.J.; Corlett, A.; Schlessinger, K.; Wegrzyn, J.; Larner, A.C.; Levy, D.E. Mitochondrial STAT3 Supports Ras-Dependent Oncogenic Transformation. Science 2009, 324, 1713–1716. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Weng, S.; Zhu, Y.; Chen, H.; Pan, J.; Qiu, S.; Liu, Y.; Wei, D.; Zhu, T. PKCι Induces Differential Phosphorylation of STAT3 to Modify STAT3-Related Signaling Pathways in Pancreatic Cancer Cells. J Cell Commun Signal 2023, 17, 1417–1433. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Zhong, Y.; Dong, H.; Zheng, Q.; Shi, S.; Zhu, K.; Qu, X.; Hu, W.; Zhang, X.; Wang, Y. Revisiting Signal Transducer and Activator of Transcription 3 (STAT3) as an Anticancer Target and Its Inhibitor Discovery: Where Are We and Where Should We Go? European Journal of Medicinal Chemistry 2020, 187, 111922. [Google Scholar] [CrossRef] [PubMed]
- Byun, W.S.; Bae, E.S.; Cui, J.; Park, H.J.; Oh, D.-C.; Lee, S.K. Antitumor Activity of Pulvomycin via Targeting Activated-STAT3 Signaling in Docetaxel-Resistant Triple-Negative Breast Cancer Cells. Biomedicines 2021, 9, 436. [Google Scholar] [CrossRef] [PubMed]
- Morrow, E.; Pennel, K.; Hatthakarnkul, P.; Leslie, H.; Mallon, E.; Andersen, D.; Jamieson, N.; McMillan, D.; Roseweir, A.; Edwards, J. High Expression of STAT3 within the Tumour-Associated Stroma Predicts Poor Outcome in Breast Cancer Patients. Cancer Medicine 2023, 12, 13225–13240. [Google Scholar] [CrossRef] [PubMed]
- Jiang, R.; Zhu, J.; Zhang, H.; Yu, Y.; Dong, Z.; Zhou, H.; Wang, X. STAT3: Key Targets of Growth-Promoting Receptor Positive Breast Cancer. Cancer Cell International 2024, 24, 356. [Google Scholar] [CrossRef]
- Wang, L.; Cao, L.; Wang, H.; Liu, B.; Zhang, Q.; Meng, Z.; Wu, X.; Zhou, Q.; Xu, K. Cancer-Associated Fibroblasts Enhance Metastatic Potential of Lung Cancer Cells through IL-6/STAT3 Signaling Pathway. Oncotarget 2017, 8, 76116. [Google Scholar] [CrossRef]
- Parakh, S.; Ernst, M.; Poh, A.R. Multicellular Effects of STAT3 in Non-Small Cell Lung Cancer: Mechanistic Insights and Therapeutic Opportunities. Cancers (Basel) 2021, 13, 6228. [Google Scholar] [CrossRef]
- Kim, J.E.; Patel, M.; Ruzevick, J.; Jackson, C.M.; Lim, M. STAT3 Activation in Glioblastoma: Biochemical and Therapeutic Implications. Cancers 2014, 6, 376–395. [Google Scholar] [CrossRef] [PubMed]
- Fu, W.; Hou, X.; Dong, L.; Hou, W. Roles of STAT3 in the Pathogenesis and Treatment of Glioblastoma. Front Cell Dev Biol 2023, 11, 1098482. [Google Scholar] [CrossRef] [PubMed]
- Liang, B.; Li, S.-Y.; Gong, H.-Z.; Wang, L.-X.; Lu, J.; Zhao, Y.-X.; Gu, N. Clinicopathological and Prognostic Roles of STAT3 and Its Phosphorylation in Glioma. Dis Markers 2020, 2020, 8833885. [Google Scholar] [CrossRef] [PubMed]
- Hashemi, M.; Sabouni, E.; Rahmanian, P.; Entezari, M.; Mojtabavi, M.; Raei, B.; Zandieh, M.A.; Behroozaghdam, M.; Mirzaei, S.; Hushmandi, K.; et al. Deciphering STAT3 Signaling Potential in Hepatocellular Carcinoma: Tumorigenesis, Treatment Resistance, and Pharmacological Significance. Cellular & Molecular Biology Letters 2023, 28, 33. [Google Scholar] [CrossRef]
- Lee, C.; Cheung, S.T. STAT3: An Emerging Therapeutic Target for Hepatocellular Carcinoma. Cancers (Basel) 2019, 11, 1646. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, A.; Wang, S.C.; Morris, J.P.; Folias, A.E.; Liou, A.; Kim, G.E.; Akira, S.; Boucher, K.M.; Firpo, M.A.; Mulvihill, S.J.; et al. Stat3 and MMP7 Contribute to Pancreatic Ductal Adenocarcinoma Initiation and Progression. Cancer Cell 2011, 19, 441–455. [Google Scholar] [CrossRef] [PubMed]
- Nagathihalli, N.S.; Castellanos, J.A.; Shi, C.; Beesetty, Y.; Reyzer, M.L.; Caprioli, R.; Chen, X.; Walsh, A.J.; Skala, M.C.; Moses, H.L.; et al. STAT3 Mediated Remodeling of the Tumor Microenvironment Results in Enhanced Tumor Drug Delivery in a Mouse Model of Pancreatic Cancer. Gastroenterology 2015, 149, 1932–1943.e9. [Google Scholar] [CrossRef]
- Sadrkhanloo, M.; Paskeh, M.D.A.; Hashemi, M.; Raesi, R.; Motahhary, M.; Saghari, S.; Sharifi, L.; Bokaie, S.; Mirzaei, S.; Entezari, M.; et al. STAT3 Signaling in Prostate Cancer Progression and Therapy Resistance: An Oncogenic Pathway with Diverse Functions. Biomedicine & Pharmacotherapy 2023, 158, 114168. [Google Scholar] [CrossRef]
- Morikawa, T.; Baba, Y.; Yamauchi, M.; Kuchiba, A.; Nosho, K.; Shima, K.; Tanaka, N.; Huttenhower, C.; Frank, D.A.; Fuchs, C.S.; et al. STAT3 Expression, Molecular Features, Inflammation Patterns and Prognosis in a Database of 724 Colorectal Cancers. Clin Cancer Res 2011, 17, 1452–1462. [Google Scholar] [CrossRef] [PubMed]
- Ma, R.-J.; Ma, C.; Hu, K.; Zhao, M.-M.; Zhang, N.; Sun, Z.-G. Molecular Mechanism, Regulation, and Therapeutic Targeting of the STAT3 Signaling Pathway in Esophageal Cancer (Review). Int J Oncol 2022, 61, 105. [Google Scholar] [CrossRef]
- Zheng, Z.-Y.; Chu, M.-Y.; Lin, W.; Zheng, Y.-Q.; Xu, X.-E.; Chen, Y.; Liao, L.-D.; Wu, Z.-Y.; Wang, S.-H.; Li, E.-M.; et al. Blocking STAT3 Signaling Augments MEK/ERK Inhibitor Efficacy in Esophageal Squamous Cell Carcinoma. Cell Death Dis 2022, 13, 1–14. [Google Scholar] [CrossRef]
- He, Z.; Song, B.; Zhu, M.; Liu, J. Comprehensive Pan-Cancer Analysis of STAT3 as a Prognostic and Immunological Biomarker. Sci Rep 2023, 13, 5069. [Google Scholar] [CrossRef] [PubMed]
- Glaviano, A.; Foo, A.S.C.; Lam, H.Y.; Yap, K.C.H.; Jacot, W.; Jones, R.H.; Eng, H.; Nair, M.G.; Makvandi, P.; Geoerger, B.; et al. PI3K/AKT/mTOR Signaling Transduction Pathway and Targeted Therapies in Cancer. Mol Cancer 2023, 22, 138. [Google Scholar] [CrossRef]
- Cerma, K.; Piacentini, F.; Moscetti, L.; Barbolini, M.; Canino, F.; Tornincasa, A.; Caggia, F.; Cerri, S.; Molinaro, A.; Dominici, M.; et al. Targeting PI3K/AKT/mTOR Pathway in Breast Cancer: From Biology to Clinical Challenges. Biomedicines 2023, 11, 109. [Google Scholar] [CrossRef] [PubMed]
- Capivasertib. Available online: https://go.drugbank.com/drugs/DB12218 (accessed on 17 September 2024).
- Truqap (Capivasertib) plus Faslodex Approved in the US for Patients with Advanced HR-Positive Breast Cancer. Available online: https://www.astrazeneca.com/media-centre/press-releases/2023/truqap-approved-in-us-for-hr-plus-breast-cancer.html (accessed on 17 September 2024).
- Liu, D.; Weintraub, M.A.; Garcia, C.; Goncalves, M.D.; Sisk, A.E.; Casas, A.; Harding, J.J.; Harnicar, S.; Drilon, A.; Jhaveri, K.; et al. Characterization, Management, and Risk Factors of Hyperglycemia during PI3K or AKT Inhibitor Treatment. Cancer Med 2022, 11, 1796–1804. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.G.; Barrios, D.M.; Blinder, V.S.; Bromberg, J.F.; Drullinsky, P.R.; Funt, S.A.; Jhaveri, K.L.; Lake, D.E.; Lyons, T.; Modi, S.; et al. Dermatologic Adverse Events Related to the PI3Kα Inhibitor Alpelisib (BYL719) in Patients with Breast Cancer. Breast Cancer Res Treat 2020, 183, 227–237. [Google Scholar] [CrossRef]
- Gasmi, A.; Roubaud, G.; Dariane, C.; Barret, E.; Beauval, J.-B.; Brureau, L.; Créhange, G.; Fiard, G.; Fromont, G.; Gauthé, M.; et al. Overview of the Development and Use of Akt Inhibitors in Prostate Cancer. J Clin Med 2021, 11, 160. [Google Scholar] [CrossRef]
- Chappell, W.H.; Steelman, L.S.; Long, J.M.; Kempf, R.C.; Abrams, S.L.; Franklin, R.A.; Bäsecke, J.; Stivala, F.; Donia, M.; Fagone, P.; et al. Ras/Raf/MEK/ERK and PI3K/PTEN/Akt/mTOR Inhibitors: Rationale and Importance to Inhibiting These Pathways in Human Health. Oncotarget 2011, 2, 135–164. [Google Scholar] [CrossRef]
- Zhang, Z.; Rohweder, P.J.; Ongpipattanakul, C.; Basu, K.; Bohn, M.-F.; Dugan, E.J.; Steri, V.; Hann, B.; Shokat, K.M.; Craik, C.S. A Covalent Inhibitor of K-Ras(G12C) Induces MHC Class I Presentation of Haptenated Peptide Neoepitopes Targetable by Immunotherapy. Cancer Cell 2022, 40, 1060–1069.e7. [Google Scholar] [CrossRef]
- Bahar, M.E.; Kim, H.J.; Kim, D.R. Targeting the RAS/RAF/MAPK Pathway for Cancer Therapy: From Mechanism to Clinical Studies. Sig Transduct Target Ther 2023, 8, 1–38. [Google Scholar] [CrossRef]
- Yang, P.-L.; Liu, L.-X.; Li, E.-M.; Xu, L.-Y. STAT3, the Challenge for Chemotherapeutic and Radiotherapeutic Efficacy. Cancers (Basel) 2020, 12, 2459. [Google Scholar] [CrossRef]
- Geiger, J.L.; Grandis, J.R.; Bauman, J.E. The STAT3 Pathway as a Therapeutic Target in Head and Neck Cancer: Barriers and Innovations. Oral Oncol 2016, 56, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Kortylewski, M.; Yu, H. Role of Stat3 in Suppressing Anti-Tumor Immunity. Current Opinion in Immunology 2008, 20, 228–233. [Google Scholar] [CrossRef]
- Jin, J.; Li, Y.; Zhao, Q.; Chen, Y.; Fu, S.; Wu, J. Coordinated Regulation of Immune Contexture: Crosstalk between STAT3 and Immune Cells during Breast Cancer Progression. Cell Commun Signal 2021, 19, 50. [Google Scholar] [CrossRef]
- Doheny, D.; Sirkisoon, S.; Carpenter, R.L.; Aguayo, N.R.; Regua, A.T.; Anguelov, M.; Manore, S.G.; Arrigo, A.; Jalboush, S.A.; Wong, G.L.; et al. Combined Inhibition of JAK2-STAT3 and SMO-GLI1/tGLI1 Pathways Suppresses Breast Cancer Stem Cells, Tumor Growth, and Metastasis. Oncogene 2020, 39, 6589–6605. [Google Scholar] [CrossRef] [PubMed]
- Han, D.; Yu, T.; Dong, N.; Wang, B.; Sun, F.; Jiang, D. Napabucasin, a Novel STAT3 Inhibitor Suppresses Proliferation, Invasion and Stemness of Glioblastoma Cells. Journal of Experimental & Clinical Cancer Research 2019, 38, 289. [Google Scholar] [CrossRef]
- Weng, Y.-S.; Tseng, H.-Y.; Chen, Y.-A.; Shen, P.-C.; Al Haq, A.T.; Chen, L.-M.; Tung, Y.-C.; Hsu, H.-L. MCT-1/miR-34a/IL-6/IL-6R Signaling Axis Promotes EMT Progression, Cancer Stemness and M2 Macrophage Polarization in Triple-Negative Breast Cancer. Mol Cancer 2019, 18, 42. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Huang, J.; Xie, Y.; Zhou, Y.; Wang, R.; Lou, J. Napabucasin Attenuates Resistance of Breast Cancer Cells to Tamoxifen by Reducing Stem Cell-Like Properties. Medical Science Monitor : International Medical Journal of Experimental and Clinical Research 2019, 25, 8905. [Google Scholar] [CrossRef]
- Lue, H.-W.; Cole, B.; Rao, S.A.M.; Podolak, J.; Van Gaest, A.; King, C.; Eide, C.A.; Wilmot, B.; Xue, C.; Spellman, P.T.; et al. Src and STAT3 Inhibitors Synergize to Promote Tumor Inhibition in Renal Cell Carcinoma. Oncotarget 2015, 6, 44675–44687. [Google Scholar] [CrossRef] [PubMed]
- Furtek, S.L.; Backos, D.S.; Matheson, C.J.; Reigan, P. Strategies and Approaches of Targeting STAT3 for Cancer Treatment. ACS Chem. Biol. 2016, 11, 308–318. [Google Scholar] [CrossRef] [PubMed]
- Zou, S.; Tong, Q.; Liu, B.; Huang, W.; Tian, Y.; Fu, X. Targeting STAT3 in Cancer Immunotherapy. Mol Cancer 2020, 19, 145. [Google Scholar] [CrossRef] [PubMed]
- Schust, J.; Sperl, B.; Hollis, A.; Mayer, T.U.; Berg, T. Stattic: A Small-Molecule Inhibitor of STAT3 Activation and Dimerization. Chemistry & Biology 2006, 13, 1235–1242. [Google Scholar] [CrossRef]
- Di, J.-X.; Zhang, H.-Y. C188-9, a Small-Molecule STAT3 Inhibitor, Exerts an Antitumor Effect on Head and Neck Squamous Cell Carcinoma. Anti-Cancer Drugs 2019, 30, 846–853. [Google Scholar] [CrossRef]
- Zhang, X.; Yue, P.; Page, B.D.G.; Li, T.; Zhao, W.; Namanja, A.T.; Paladino, D.; Zhao, J.; Chen, Y.; Gunning, P.T.; et al. Orally Bioavailable Small-Molecule Inhibitor of Transcription Factor Stat3 Regresses Human Breast and Lung Cancer Xenografts. Proceedings of the National Academy of Sciences 2012, 109, 9623–9628. [Google Scholar] [CrossRef] [PubMed]
- Poria, D.K.; Sheshadri, N.; Balamurugan, K.; Sharan, S.; Sterneck, E. The STAT3 Inhibitor Stattic Acts Independently of STAT3 to Decrease Histone Acetylation and Modulate Gene Expression. Journal of Biological Chemistry 2021, 296, 100220. [Google Scholar] [CrossRef]
- Ren, X.; Duan, L.; He, Q.; Zhang, Z.; Zhou, Y.; Wu, D.; Pan, J.; Pei, D.; Ding, K. Identification of Niclosamide as a New Small-Molecule Inhibitor of the STAT3 Signaling Pathway. ACS Med. Chem. Lett. 2010, 1, 454–459. [Google Scholar] [CrossRef]
- Shao, Z.; Wang, H.; Ren, H.; Sun, Y.; Chen, X. The Anticancer Effect of Napabucasin (BBI608), a Natural Naphthoquinone. Molecules 2023, 28, 5678. [Google Scholar] [CrossRef]
- Becerra, C.; Braiteh, F.S.; Spira, A.I.; Langleben, A.; Panasci, L.C.; Vukelja, S.J.; Hinshaw, I.M.; Goodwin, R.A.; Panella, T.J.; Edenfield, W.J.; et al. A Phase Ib/II Study of Cancer Stemness Inhibitor Napabucasin (BB608) Combined with Weekly Paclitaxel in Advanced Triple Negative Breast Cancer. JCO 2016, 34, 1094–1094. [Google Scholar] [CrossRef]
- Dong, J.; Cheng, X.-D.; Zhang, W.-D.; Qin, J.-J. Recent Update on Development of Small-Molecule STAT3 Inhibitors for Cancer Therapy: From Phosphorylation Inhibition to Protein Degradation. J. Med. Chem. 2021, 64, 8884–8915. [Google Scholar] [CrossRef] [PubMed]
- FDA Grants Napabucasin Orphan Status for Gastric Cancer | Sandra and Edward Meyer Cancer Center. Available online: https://meyercancer.weill.cornell.edu/news/2016-06-29/fda-grants-napabucasin-orphan-status-gastric-cancer (accessed on 1 December 2024).
- Napabucasin Phase III Trial Discontinued in Pancreatic Cancer. Available online: https://www.onclive.com/view/napabucasin-phase-iii-trial-discontinued-in-pancreatic-cancer (accessed on 1 December 2024).
- Kong, R.; Sun, G.; Li, X.; Wu, L.; Li, L.; Li, Y.; Wang, F.; Xuan, P.; Yang, S.; Sun, B.; et al. Small Molecule Inhibitor C188-9 Synergistically Enhances the Demethylated Activity of Low-Dose 5-Aza-2′-Deoxycytidine Against Pancreatic Cancer. Front Oncol 2020, 10, 612. [Google Scholar] [CrossRef]
- Redell, M.S.; Ruiz, M.J.; Alonzo, T.A.; Gerbing, R.B.; Tweardy, D.J. Stat3 Signaling in Acute Myeloid Leukemia: Ligand-Dependent and -Independent Activation and Induction of Apoptosis by a Novel Small-Molecule Stat3 Inhibitor. Blood 2011, 117, 5701–5709. [Google Scholar] [CrossRef]
- Dayyani, F.; Baretti, M.; Lee, S.S.; He, A.R.; Kim, R.D.; Lin, B.S.-L.; Enzler, T.; Al Hallak, M.N.; Ulahannan, S.V.; Davis, S.L.; et al. A Phase 1b/2 Study to Evaluate the Safety and Efficacy of TTI-101 as Monotherapy and in Combination in Advanced Hepatocellular Carcinoma. JCO 2024, 42, TPS577–TPS577. [Google Scholar] [CrossRef]
- Huang, W.; Dong, Z.; Wang, F.; Peng, H.; Liu, J.-Y.; Zhang, J.-T. A Small Molecule Compound Targeting STAT3 DNA-Binding Domain Inhibits Cancer Cell Proliferation, Migration, and Invasion. ACS Chem. Biol. 2014, 9, 1188–1196. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Bian, A.; Yang, L.; Yin, X.; Wang, J.; Ti, C.; Miao, Y.; Peng, S.; Xu, S.; Liu, M.; et al. Targeting STAT3 by a Small Molecule Suppresses Pancreatic Cancer Progression. Oncogene 2021, 40, 1440–1457. [Google Scholar] [CrossRef]
- Chen, H.; Bian, A.; Zhou, W.; Miao, Y.; Ye, J.; Li, J.; He, P.; Zhang, Q.; Sun, Y.; Sun, Z.; et al. Discovery of the Highly Selective and Potent STAT3 Inhibitor for Pancreatic Cancer Treatment. ACS Cent. Sci. 2024, 10, 579–594. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.; Chen, X.; Fu, S.; Yu, W.; Li, C.; Wang, T.; Lo, H.-W.; Lin, J. LLY17, a Novel Small Molecule STAT3 Inhibitor Induces Apoptosis and Suppresses Cell Migration and Tumor Growth in Triple-Negative Breast Cancer. Breast Cancer Res Treat 2020, 181, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Brambilla, L.; Genini, D.; Laurini, E.; Merulla, J.; Perez, L.; Fermeglia, M.; Carbone, G.M.; Pricl, S.; Catapano, C.V. Hitting the Right Spot: Mechanism of Action of OPB-31121, a Novel and Potent Inhibitor of the Signal Transducer and Activator of Transcription 3 (STAT3). Mol Oncol 2015, 9, 1194–1206. [Google Scholar] [CrossRef] [PubMed]
- Bendell, J.C.; Hong, D.S.; Burris, H.A.; Naing, A.; Jones, S.F.; Falchook, G.; Bricmont, P.; Elekes, A.; Rock, E.P.; Kurzrock, R. Phase 1, Open-Label, Dose-Escalation, and Pharmacokinetic Study of STAT3 Inhibitor OPB-31121 in Subjects with Advanced Solid Tumors. Cancer Chemother Pharmacol 2014, 74, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Brambilla, L.; Lahiri, T.; Cammer, M.; Levy, D.E. STAT3 Inhibitor OPB-51602 Is Cytotoxic to Tumor Cells Through Inhibition of Complex I and ROS Induction. iScience 2020, 23, 101822. [Google Scholar] [CrossRef]
- Ogura, M.; Uchida, T.; Terui, Y.; Hayakawa, F.; Kobayashi, Y.; Taniwaki, M.; Takamatsu, Y.; Naoe, T.; Tobinai, K.; Munakata, W.; et al. Phase I Study of OPB-51602, an Oral Inhibitor of Signal Transducer and Activator of Transcription 3, in Patients with Relapsed/Refractory Hematological Malignancies. Cancer Sci 2015, 106, 896–901. [Google Scholar] [CrossRef]
- Vividion Therapeutics, Inc. A Phase 1, Open-Label, 2-Part, Multicenter, First-in-Human Dose Escalation and Dose Expansion Study to Evaluate the Safety, Tolerability, Pharmacokinetics, Pharmacodynamics, and Preliminary Anti-Tumor Activity of the STAT3 Inhibitor VVD-130850 as Single Agent and in Combination With Checkpoint Inhibition in Participants With Advanced Solid and Hematologic Tumors, 2024.
- Dicerna Pharmaceuticals, Inc., a Novo Nordisk company An Open-Label, Phase 1, Dose-Ranging Study to Evaluate the Safety, Tolerability, and Pharmacokinetics of Intravenous DCR-STAT3 in Adults With Refractory Solid Tumors; clinicaltrials. 2024.
- Yang, J.; Liao, X.; Agarwal, M.K.; Barnes, L.; Auron, P.E.; Stark, G.R. Unphosphorylated STAT3 Accumulates in Response to IL-6 and Activates Transcription by Binding to NFκB. Genes Dev 2007, 21, 1396–1408. [Google Scholar] [CrossRef] [PubMed]
- Timofeeva, O.A.; Chasovskikh, S.; Lonskaya, I.; Tarasova, N.I.; Khavrutskii, L.; Tarasov, S.G.; Zhang, X.; Korostyshevskiy, V.R.; Cheema, A.; Zhang, L.; et al. Mechanisms of Unphosphorylated STAT3 Transcription Factor Binding to DNA. J Biol Chem 2012, 287, 14192–14200. [Google Scholar] [CrossRef]
- Nkansah, E.; Shah, R.; Collie, G.W.; Parkinson, G.N.; Palmer, J.; Rahman, K.M.; Bui, T.T.; Drake, A.F.; Husby, J.; Neidle, S.; et al. Observation of Unphosphorylated STAT3 Core Protein Binding to Target dsDNA by PEMSA and X-Ray Crystallography. FEBS Letters 2013, 587, 833–839. [Google Scholar] [CrossRef]
- Liu, Z.; Hazan-Halevy, I.; Harris, D.M.; Li, P.; Ferrajoli, A.; Faderl, S.; Keating, M.J.; Estrov, Z. STAT-3 Activates NF-κB in Chronic Lymphocytic Leukemia Cells. Molecular Cancer Research 2011, 9, 507–515. [Google Scholar] [CrossRef] [PubMed]
- Gu, S.; Cui, D.; Chen, X.; Xiong, X.; Zhao, Y. PROTACs: An Emerging Targeting Technique for Protein Degradation in Drug Discovery. BioEssays 2018, 40, 1700247. [Google Scholar] [CrossRef]
- Berkley, K.; Zalejski, J.; Sharma, N.; Sharma, A. Journey of PROTAC: From Bench to Clinical Trial and Beyond. Biochemistry 2025. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.; Li, S.; Han, S.; Shi, C.; Zhang, Y. Antibody Drug Conjugate: The “Biological Missile” for Targeted Cancer Therapy. Sig Transduct Target Ther 2022, 7, 1–25. [Google Scholar] [CrossRef]
- Wang, H.; Man, Q.; Huo, F.; Gao, X.; Lin, H.; Li, S.; Wang, J.; Su, F.; Cai, L.; Shi, Y.; et al. STAT3 Pathway in Cancers: Past, Present, and Future. MedComm (2020) 2022, 3, e124. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schematic representation of STAT3 activation in cells.
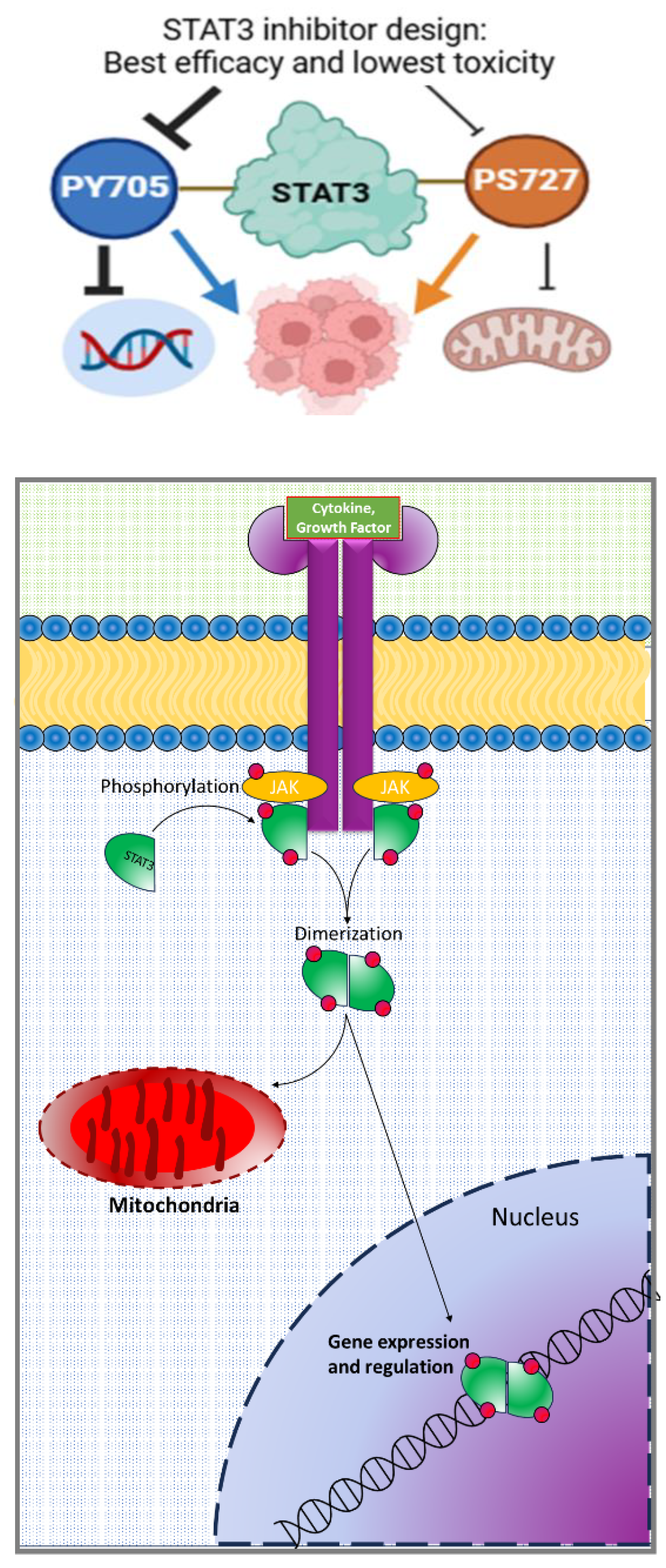
Figure 2.
Representation of drug target sites. Single STAT3 protein, folding done by AlphaFold (AF-P40763-F1-v4). Blue region indicates Src Homology 2 domain (SH2). Magenta indicates DNA binding domain. Red spheres indicate Ser727 residue. Orange spheres indicate Tyr705.
Figure 2.
Representation of drug target sites. Single STAT3 protein, folding done by AlphaFold (AF-P40763-F1-v4). Blue region indicates Src Homology 2 domain (SH2). Magenta indicates DNA binding domain. Red spheres indicate Ser727 residue. Orange spheres indicate Tyr705.
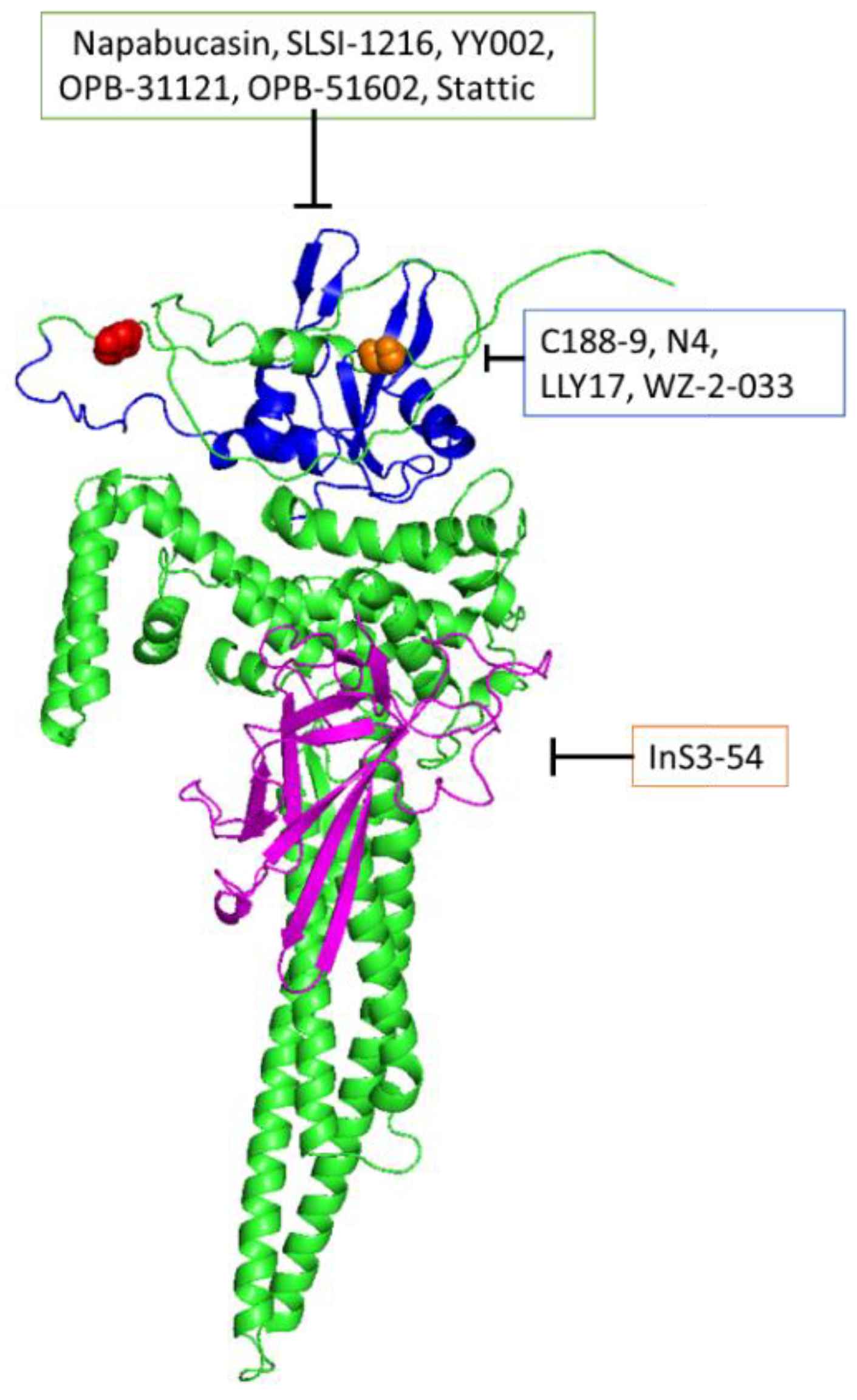

Table 1.
List of STAT3 inhibitors including the target site, affected phospho-residues, clinical efficacy, and toxicity.
Table 1.
List of STAT3 inhibitors including the target site, affected phospho-residues, clinical efficacy, and toxicity.
| Compound | Target Domain | pY705 | pS727 | Clinical Efficacy | Toxicity |
| Stattic | SH2 | Yes | Yes | N/A | High |
| Napabucasin (BBI608) | SH2 | Yes | Yes | Low | Low |
| SLSI-1216 | SH2 | Yes | Yes | N/A | N/A |
| C188-9 (TTI-101) |
SH2 | Yes | N/A | High | Low |
| InS3-54 | DNA-binding | No | No | N/A | N/A |
| N4 | SH2 | Yes | No | N/A | Low |
| YY002 | SH2 | Yes | Yes | N/A | Low |
| LLY17 | SH2 | Yes | N/A | N/A | N/A |
| WZ-2-033 | SH2 | Yes | No | N/A | Low |
| OPB-31121 | SH2 | Yes | Yes | Low | High |
| OPB-51602 | SH2 | N/A | N/A | High | High |
| VVD-130850 | Allosteric | N/A | N/A | N/A | N/A |
| DCR-STAT3 | RNA | Yes | Yes | N/A | N/A |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



