
Submitted:
15 January 2025
Posted:
15 January 2025
You are already at the latest version
Abstract
Multiple sclerosis (MS) is a chronic inflammatory neurodegenerative disease of the central nervous system. The manifestation of MS is related to steroid changes during the menstrual cycle and pregnancy. As data focusing on the effect of anti-MS drug treatment on steroidome are scarce, we evaluated steroidomic changes (79 steroids) in 60 female MS patients of reproductive age 39 (29, 47) years (median with quartiles) after treatment with anti-MS drugs on the GC-MS/MS platform and immunoassays (cortisol and estradiol). The changes were assessed using steroid levels and steroid molar ratios that may reflect activities of steroidogenic enzymes (SMRs). A repeated measures ANOVA followed by multiple comparisons and OPLS models were used for statistical analyses. The anti-MS treatment decreased steroid levels in follicular phase. Anti-CD20 monoclonal antibodies (mAb) such as ofatumumab and ocrelizumab, inhibitors of the sphingosine-1-phosphate receptor (S1PRI), and IFNβ-1a decreased circulating 17-hydroxy-pregnanes and shifted the CYP17A1 functioning from the hydroxylase- toward the lyase step. Decreased conjugated/unconjugated steroid ratios were found after treatment with anti-MS drugs, especially for glatiramer acetate and anti-CD20 mAb. In the luteal phase, IFN-β1a treatment increased steroidogenesis, and both IFN-β1a and ocrelizumab increased AKR1D1 and S1PRI increased SRD5A functioning. Anti-CD20 mAb reduced the functioning of enzymes catalyzing the synthesis of immunomodulatory 7α/β and 16α-hydroxy-androgens, which may affect the severity of MS. The above findings may be important concerning the alterations in bioactive steroids such as cortisol, active androgens and estrogens, neuroactive, neuroprotective and immunomodulatory steroids in terms of optimization of anti-MS treatment.
Keywords:
multiple sclerosis
; steroidomics
; GC-MS/MS
; multivariate statistics
; anti-MS drugs
1. Introduction
Multiple sclerosis (MS) is a chronic inflammatory disease of the central nervous system (CNS) that causes demyelination and neurodegeneration [1]. MS is associated with varying degrees of inflammation, with the inflammatory reaction being promoted by the secretion of autoantigens, and is therefore an autoimmune inflammatory response. This hypothesis suggests that primary degeneration is present several years before the first overt clinical symptoms and progresses over the course of the disease (see review [2]).
1.1. Multiple Sclerosis and Steroids
Hormones play a key role in the physiology and pathology of the immune system, specifically in adaptive immunity. Exacerbation of MS is closely related to the menstrual cycle and occurs at very low levels of sex steroids [3]. In addition, the MS symptoms decrease in the last three months of pregnancy and increase after delivery, probably due to hormonal and immunological changes [4].
1.1.1. Cortisol
Cortisol is a potent endogenous anti-inflammatory steroid hormone that is released into the circulation after physical and mental stimuli. With the loss of adequate response to stress, the hypothalamic pituitary adrenal axis (HPAA) does not produce sufficient quantities of this glucocorticoid (see review [5]). Cortisol is produced in the zona fasciculata of the adrenal cortex and increases the body's overall alertness in stressful situations by raising blood sugar levels via gluconeogenesis [6]. Genomic effects of cortisol are mediated by binding to high affinity mineralocorticoid receptors (MRs) and low affinity glucocorticoid receptors. Non-genomic cortisol effects are mediated by membrane receptors and G-protein coupled signaling [6].
Circulating cortisol follows a circadian rhythm, reaching a peak in early morning and having their nadir in late evening (see review [5]). Chronic inflammation is associated with altered HPAA in patients with autoimmune diseases (see review [5]) and cortisol suppresses levels of several pro-inflammatory cytokines, including IL-2, IL-3, IL-6, TNF, and interferon (IFN)-γ and influences activity and survival of immune system cells (see review [5]). Reduced cortisol secretion in response to chronic inflammation has been reported in patients with autoimmune diseases compared to healthy subjects (see review [5]). Some authors consider that the relative adrenal insufficiency in these patients may be secondary to impaired hepatic metabolism of steroid hormones. The key enzyme involved in the conversion of inactive cortisone to active cortisol, 11β-hydroxysteroid dehydrogenase type 1 (HSD11B1), is significantly stimulated by TNF and other pro-inflammatory cytokines, and the conversion of inactive cortisone to active hormone takes place primarily in the liver. According to this hypothesis, the stimulation of HSD11B1 and the subsequent increased concentration of the active glucocorticoid cortisol could induce a negative feedback loop and thus dysfunction of the HPAA [11] (see review [5]).
In T cells, cortisol decreases IL-2 expression, suppressing T cell receptor function via membrane-bound receptors. Cortisol binds to the GC receptor on T-cell mitochondria and disrupts their function, leading to cell death. Cortisol also suppresses the production of cytokines, chemokines, arachidonic acid derivatives in mast cells, as well as the number of basophils and histamine production. In dendritic cells (DCs), cortisol reduces their maturation potential and ability to activate T cells which results in attenuated IFN-production. While low cortisol levels induce activation of macrophages, high concentrations have an immunosuppressive effect (see review [5]). With prolonged exposure to high concentrations of cortisol, its effects on B cells prevail (see review [5]).
Cortisol also affects the activity of several neurotransmitter systems that influence reward processing, attention regulation, executive function, mood and emotion such as type A GABA receptors (GABAARs), glutamate, dopamine and acetylcholine receptors. Moreover, cortisol inhibits the synthesis and secretion of serotonin, which increases the risk of depression [6] and also affects attention, perception, memory and emotions. Chronically elevated cortisol may result in cognitive impairment [7-9].
1.1.2. Δ5 Steroids
While the adrenal zona fasciculata produces all the cortisol circulating in the body, the adrenal zona reticularis produces approximately 80% of dehydroepiandrosterone (DHEA) and the testes, ovaries, and brain synthesize the rest. Although adrenocorticotropic hormone (ACTH) stimulates both cortisol and DHEA and its sulfate (DHEAC) synthesis, the feedback mechanisms typical for cortisol are absent in DHEA/C [6]. In contrast to the diabetogenic effect of cortisol, DHEAC is an anti-glucocorticoid that protects the hippocampus from the damaging effect of cortisol. [7,9-11].
The formation of another neuroprotective Δ5 steroid pregnenolone (Preg) is catalyzed by cholesterol desmolase, which activity is rate-limiting for adrenal steroidogenesis [6]. Preg, DHEA, Preg sulfate (PregC) and DHEAC cross the blood-brain barrier (BBB) and their adrenal production affects their levels in the CNS [12-14].
The aforementioned steroids protects from glutamate-induced neurotoxicity, stabilizes microtubules, activates neurite growth and promotes myelination [15,16] and also modulate various ionotropic receptors, such as N-methyl-D-aspartate receptors (NMDARs), α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptors (AMPARs), melastatin receptors (TRPM3s), short transient receptor potential receptors 5 (TRPC5s) or vanilloid receptors (TRPV1s) and may thus improve cognitive function, and counteract pain transmission and fear (see [7] and [17]). Furthermore, DHEAC and PregC act as agonists, whereas progesterone (P) functions as an antagonist on σ1 receptors (σ1Rs), which are abundant in the brain and are potent modulators of excitatory glutamatergic and cholinergic systems [15,18].
DHEA/DHEAC has anti-inflammatory effects reducing the levels of interleukin 1 (IL-1), interleukin 4 (IL-4) and interleukin 6 (IL-6), interleukin 12 (IL-12), tumor necrosis factor (TNF) [16]. Interestingly, higher TNF levels, which are typical for autoimmune diseases, inhibit the conversion from the DHEAC to the DHEA, which may induce disequilibrium between anti- and pro-inflammatory factors (see review [5]).
1.1.3. Active Androgens
Androgens play a positive role in the development and function of the innate immune response by inhibiting the adaptive immune system, thereby preventing autoimmunity [1]. Lower androgen levels we observed in some autoimmune diseases and the protective effects of androgens were attributed to suppression of DCs activation and presentation of co-stimulatory molecules, decreased secretion of type I IFNγ, diminished secretion of pro-inflammatory cytokines from macrophages, and reduced T-helper 1 (Th1) T cell response. Moreover, androgens suppress B cell differentiation and antibody production, stimulate T-helper 2 (Th2) pathway and IL-10 secretion (see review [5]).
1.1.4. Estrogens
Although estradiol (E2) is a key female sex hormone, its synthesis is not limited to the ovaries. E2 is also produced in significant amounts in extragonadal tissues, especially in adipocytes, which express enzymes required for its synthesis, such as aromatase (CYP19A1), converting androstenedione (A) to estrone (E1), and subfamily AKR1C type 3 aldoketoreductase (AKR1C3), converting inactive estrone to active E2 http://biogps.org/#goto=genereport&id=8644) (also see reviews [19] and [20]). E2 is pro-inflammatory at low concentrations and anti-inflammatory at high concentrations (see review [2]). While at low levels E2 stimulates DCs to produce IL-1, IFNγ and promotes T-cell and TNF production, at high levels it inhibits IL-1, IL-6 and TNF production and in turn stimulates IL-4 and IL-10 formation (see review [7]]). Since TNF activates CYP19A1, low levels of E2 increase the local conversion of androgens to estrogens (see review [5]). E2 in almost any concentration stimulates the production of antibodies from B cells (see review [5]).
1.1.5. Progesterone and Its Metabolites
Progesterone binds to different nuclear receptors, i.e., the P receptor and the glucocorticoid receptor (GC), both of which have suppressive effects on IFN function. While binding to progesterone receptors occurs at physiological levels and mediates a reduction in Th1 cell differentiation, binding to GC receptors occurs at higher P levels, for example in pregnancy, and is mediated by a reduction in type I IFN secretion from plasmacytoid DCs (see review [5]). During pregnancy, the severity of MS decreases as P suppresses nitric oxide production and toll-like receptor expression by macrophages. Progesterone, via binding to the P receptor, also stimulates the differentiation of regulatory T cells, which are involved in immunosuppression (see review [5]).
The P metabolite allopregnanolone (3α,5α-THP) has various neuroprotective effects including alleviation of neurobehavioral deficits and attenuation of neuropathology and inflammation in animal models of autoimmune demyelination [21,22]. Moreover, recent studies demonstrated that 3α,5α-THP suppresses neuroinflammation through activation of TLR4 protein in macrophages and in the brain [51].
However, apart from the above protective effects, P increases the vulnerability of neurons to apoptotic damage in experimental autoimmune encephalomyelitis (EAE) (see review [23]).
1.2. Effect of Anti-MS Drugs on Circulating Steroids and Steroid Molar Ratios (SMRs)
Data in the literature focusing on the effect of steroid treatment on anti-MS are rare but a few relevant reports were found [24-29].
1.2.1. Glatiramer Acetate (GA)
GA is an immunomodulatory drug used for the treatment of relapsing-remitting MS (RRMS). GA blocks peptide-major histocompatibility complex and exerts its anti-inflammatory effect by affecting antigen presenting cells (APCs) to promote the development of Th2 and production of anti-inflammatory cytokines (IL-4, IL-13, IL-10), and consequently, suppressing the development of pathogenic T cells (Th1, Th17) [30,31]. As to effect of GA therapy on steroidome, Cil et al. did not found changes in follitropin (FSH) and E2 levels in MS patients treated by a combined therapy with interferon β and GA but found diminished ovarian volume in MS patients compared to age matched controls [24].
1.2.2. Interferons β1a (IFN-β1a)
Interferon β-1a (IFN-β1a), which is produced by mammalian cells is a cytokine used for (MS) treatment. IFN-β1a balances pro- and anti-inflammatory cytokines, and primarily decreases pro-inflammatory Th17 cells [31-33]. Griffiths and Wang [25] proposed that cholesterol 25-hydroxylase (CH25H) is an interferon-stimulated gene and 25-hydroxycholesterol can be considered as an immunoregulatory oxysterol that affects the adaptive immune system.
1.2.3. Inhibitors of the Sphingosine-1-Phosphate Receptor (S1PRI)
S1PRI decrease T and B cells, activates S1P signaling in CNS and reduce Th1 cytokines [31,32]. E2 rapidly stimulates sphingosine kinase activity in endothelial cells (EC), promoting sphingosine-1-phosphate (S1P) production in EC. Decreased S1P1R levels impair estrogen-induced EC migration and tube formation [26]. S1P rapidly increases cortisol biosynthesis and the expression of genes participating in cholesterol uptake and transport in H295R adrenocortical cells [27]. Luteotropin (LH) and FSH promote S1P synthesis in follicular cells via activation of protein kinase C, while S1P mediates FSH and LH proliferation, survival and viability in follicular cells [28].
1.2.4. Ofatumumab (Kesimpta) and Ocrelizumab (Ocrevus)
Ofatumumab is a humanized anti-CD20 mAb developed as an immunosuppressive drug for the treatment of MS [34]. Regarding ofatumumab, there is no information on its effect on the steroidome to date. Ocrelizumab is another humanized anti-CD20 mAb frequently used for the treatment of RRMS [35] and primary progressive multiple sclerosis (PPMS) [36]. In a recent study, suppression of the production of various androstane and pregnane steroids by ocrelizumab was reported [29].
2. Results
2.1. Pooled Trends of Changes in Circulating Steroid Levels and Steroid Molar Ratios (SMR) After Treatment with Anti-MS Drugs
The significance of the changes after the anti-MS treatment (stage or treatment effect) were evaluated for Stage factor in ANOVA model (before treatment vs. after treatment) and for least significant difference (LSD) multiple comparisons separately for follicular (FP) and luteal (LP) phases of the menstrual cycle (PMCs). The pooled steroidomic changes (without differentiation of individual anti-MS drugs) after treatment are shown in Table S1. The results of the Wilcoxon one-sample test with correction for continuity, which was used to assess the prevailing trend of changes in steroid levels or SMRs in MS patients after treatment with anti-MS drugs, are summarized in Table 1, separately for FP and LP.
Generally, significant trends in steroidomic changes after anti MS-treatment are obvious in the FP, changes in LP are mostly insignificant with the exception of borderline trend to lower values of SMRs that may reflect the activity of CYP11B1. Therefore, the following results apply to FP. For a total of 79 investigated steroids, a significant trend towards a decrease in their levels after treatment was found. A borderline trend to decrease after treatment was observed for unconjugated steroids but this trend reached significance for steroid conjugates. Regarding the groups of steroids, while the C21 steroids (pregnanes) showed a significant downward trend after treatment, the C19 steroids (androstanes) showed only an insignificant tendency in this direction. Also, 17α-hydroxy-pregnanes tended to decrease after treatment, but their 17-deoxy-counterparts did not. Furthermore, the 20α-dihydro-pregnanes showed a borderline trend to decrease in FP, but their 20-oxo counterparts did not. Finally, the GABAergic 3α-hydroxy-5α/β-steroids showed a significant downward trend after treatment; however, this was not the case for the 3-oxo-/3β -hydroxy-5α/β-steroids.
2.1.1. Corticoids (C21 Δ4 Steroids) and 11β-Hydroxy-Androstanes (C19 Δ4 and 5α/β Steroids)
The corticoids and 11β-hydroxy-androstanes tended to a borderline decrease after treatment in FP but not in LP. Specifically, cortisol measured by both GC-MS/MS and RIA showed a significant decrease in FP like cortisone and corticosterone. The changes in circulating cortisol, cortisone and corticosterone are also illustrated in Figure 1.
2.1.2. Δ5 and Δ4 Steroids
The changes after treatment did not show significant trend neither for Δ5 steroids nor for 11-deoxy Δ4 steroids. Nevertheless, the neuroprotective steroids pregnenolone and DHEA significantly increased when evaluating the factor Stage in ANOVA model (Figure 2). However, the levels of circulating metabolites of DHEA and androstenediol (Adiol) such as 7α-hydroxy-DHEA and 5-androstene-3β,7β,17β-triol (3β,7β,17β-AT) significantly decreased in FP after anti-MS-treatment.
2.1.3. Active Androgens and Estrogens
From active androgens, testosterone after treatment significantly increased in LP but not in FP, conjugated 5α-dihydrotestosterone significantly decreased (Figure 4) and unconjugated 5α-dihydrotestosterone (5α-DHT) was not significantly influenced by anti-MS treatment. From estrogens, E1 sulfate showed a significant decrease after treatment while E2 did not change.
2.1.4. Progesterone and Its Metabolites
As expected, P and its C21 Δ4 metabolites strongly correlated with PMC but did not exhibit a pronounced trend in changes after anti-MS treatment. Nevertheless, the levels of GABAergic 3α-hydroxy-5α/β-pregnanes showed a borderline trend to decrease in FP while their 3-oxo-/3β-hydroxy-counterparts did not.
2.1.5. C17-Hydroxylase, C17-20-Lyase (CYP17A1) and 3β-Hydroxysteroid Dehydrogenases (HSD3Bs)
There was no significant trend in SMRs that could reflect summary CYP17A1 (including both hydroxylase and lyase steps) activity and CYP17A1 activity separately in the hydroxylase and lyase steps. Otherwise, all of the conjugated 17-hydroxy-5α/β-pregnanes decreased significantly in FP (Figure 4), but none of their unconjugated counterparts, whereas of the 17-hydroxy- Δ5 and Δ4 steroids, only cortisol, measured by both RIA and GC-MS/MS, decreased significantly in FP.
The SMRs, which may reflect HSD3B activity, did not show a significant trend toward changes after treatment with anti-MS drugs.
2.1.6. A Balance Between Steroid Sulfotransferase (SULT2A1) and Steroid Sulfatase (STS)
The ratios of conjugated steroids to their unconjugated counterparts that may reflect a balance between steroid sulfotransferase SULT2A1 and steroid sulfatase STS tended to decrease in FP. The same was valid for the levels of conjugated steroids.
2.1.7. 11β-Hydroxylase (CYP11B1) and Type 1 11β-Hydroxysteroid Dehydrogenase (HSD11B1)
SMRs that may reflect CYP11B1 activity did not reach significance (probably due to insufficient data) but showed a borderline trend to decrease after treatment with anti-MS drugs in LP. A low number of SMRs, which may reflect HSD11B1 activity, were also observed to assess the trend of changes after treatment. However, these data did not even suggest such a trend.
2.1.8. 7α-, 7β-, and 16α-Hydroxylating Enzymes (CYP7B1, CYP3A4, CYP3A7)
The SMRs that may reflect the activities of pluripotent 7α/β- and 16α-hydroxylating enzymes showed a borderline trend to decrease after treatment in FP. Due to low number of indices (n=8) this trend did not reach significance but still, 4 of them decreased after treatment with anti-MS drugs and 4 showed no significant change.
2.1.9. 5α-Reductases (SRD5As), 5β-Reductase (AKR1D1) and Aldoketoreductases Subfamily1c (AKR1C1-4))
The SMRs that may reflect the activities of SRD5As, AKR1C1, AKR1C2, and AKR1C3 did not show consistent trends in changes after treatment.
2.2. Effect of Anti-MS Drugs on Circulating Steroids and Steroid Molar Ratios (SMRs)
In addition to the aggregate evaluation of the effects of treatment on the steroidome, the effects of individual anti-MS drugs were evaluated as well. The following drugs were used: GA (Copaxone) FP n=6, LP n=2; dimethyl fumarate (Tecfidera) FP n=2, LP n=1; natalizumab (Tysabri) FP n=1, LP n=0; interferons β1a (Avonex, Rebif, Plegridy FP n=6, LP n=5); inhibitors of the sphingosine-1-phosphate receptor (S1PR) (Ponvory, Zeposia) FP n=5, LP n=1; teriflunomide (Aubagio) FP n=3, LP n=0; ofatumumab (Kesimpta) FP n=7, LP n=1; ocrelizumab (Ocrevus) FP n=11, LP n=5; MS patients in Stage 1 without medication FP n=2, LP n=2. Due to the low numbers of applications of these drugs, the effects of some of them were monitored in both PMCs, while others were monitored only in the follicular phase of the menstrual cycle (FP) and some drugs were not included in the evaluation at all.
2.2.1. Glatiramer Acetate (GA) in the Follicular Menstrual Phase
GA treatment did not show a significant trend in changes for steroid levels, levels of corticoids and 11β-hydroxy-androgens, Δ5 and Δ4 steroids, P 5α/β-reduced metabolites, summary CYP17A1 activity, activities in CYP17A1 hydroxylase step, CYP17A1 lyase step, HSD3Bs, conjugated/unconjugated steroids ratio, CYP11B1, CYP7B1, HSD11B1, SRD5As, and in balances AKR1C1 vs. HSD17B2, AKR1C2 vs. HSD17B2 and HSD17B6, and AKR1C3 vs. HSD17B2.
GA therapy induced a trend to decreased ratios of conjugated steroids to their unconjugated counterparts. Also, the SMRs that may reflect the balance between reductive AKR1C1 on one hand and oxidative HSD17B2 enzymes showed a trend towards a shift to the latter one. Although the trend in SMRs that may reflect the activity of CYP17A1 in the lyase step was insignificant, specifically the DHEA/17-OH-Preg and A/17-OH-P ratios were higher after GA treatment. Concerning the effects of GA treatment on the levels of androstenedione and active androgens (T, 5α-DHT and 5α-DHTC), their number was low to assess their pooled trend after anti-MS treatment. Nevertheless, GA did not show any effect on the levels of the aforementioned steroids.
2.2.2. Interferons β1a (IFN-β1a)
In FP, the treatment with IFN-β1a did not show a significant trend in changes after treatment for steroid levels, and specifically for corticoids and 11β-hydroxy-androgens, HSD3Bs, conjugated/unconjugated steroids ratios, CYP11B1, CYP7B1, HSD11B1, SRD5As, AKR1D1. The balance AKR1C3 vs. HSD17B2, showed indications of significant trends to decrease in the levels of Δ5 and Δ4 steroids and of P 5α/β-reduced metabolites. In FP, the treatment with IFN-β1a displayed a significant trend to higher values for of CYP17A1 in the lyase step but an insignificant indication of trend to lower values for CYP17A1 in the hydroxylase step. From the biologically most important steroids also the levels of T, 5α-DHT, and cortisol measured by GC-MS/MS (but not by RIA) showed significant increase after IFN-β1.
In the LP, the treatment with IFN-β1a did not show significant trends in changes after treatment for summary CYP17A1 activity, CYP17A1 activity in the hydroxylase step, CYP17A1 activity in the lyase step, HSD3Bs, conjugated/unconjugated steroids ratio, CYP11B1, CYP7B1, HSD11B1, SRD5As, AKR1D1, and balances AKR1C2 vs. HSD17B2, HSD17B6, AKR1C2 vs. HSD17B2 and HSD17B6, and AKR1C3 vs. HSD17B2. However, there was a pronounced pooled trend to higher steroid levels, and specifically higher levels of corticoids and 11β-hydroxy-androgens, Δ5 and Δ4 steroids, P 5α/β-reduced metabolites, and also a significant trend to higher SMRs that may reflect AKR1D1 activity.
2.2.3. Inhibitors of the Sphingosine-1-Phosphate Receptor (S1PRI) in the Follicular Menstrual Phase
S1PRI treatment did not significantly change the levels of corticoids and 11β-hydroxy-androgens, Δ5 and Δ4 steroids, CYP17A1 activity in the lyase step, HSD3Bs, conjugated/unconjugated steroid ratios, CYP11B1, CYP7B1, HSD11B1, AKR1D1, and balances AKR1C1 vs. HSD17B2, AKR1C2 vs. HSD17B2 and HSD17B6.
S1PRI treatment showed an indication of insignificant trend to lower levels of Δ5 and Δ4 steroids, lower summary CYP17A1 activity in the hydroxylase step and a borderline trend to lower summary CYP17A1 activity. On one hand, S1PRI treatment resulted in a significant pooled trend to decrease of steroid levels, and specifically for the levels of P 5α/β-reduced metabolites. On the other, there was a significant trend to increase for SMSs that may reflect SRD5As activity. S1PRI did not show any effect on the levels of androstenedione and active androgens (T, 5α-DHT and 5α-DHTC).
2.2.4. Ofatumumab (Kesimpta) in the Follicular Menstrual Phase
Ofatumumab treatment did not show a significant trend in changes of the levels of corticoids and 11β-hydroxy-androgens, summary activity of CYP17A1, activities of CYP17A1 in the hydroxylase and lyase steps, conjugated/unconjugated steroid ratios, CYP11B1, HSD11B1, SRD5As, AKR1D1, balances between activities of AKR1C2 vs. HSD17B2 and HSD17B6.
Ofatumumab treatment resulted in a borderline trend to decrease for HSD3Bs, significant trend to decrease of circulating steroids, and specifically Δ5 and Δ4 steroids and P 5α/β-reduced metabolites, SMRs that may reflect activities of CYP7B1, CYP3A4 and CYP3A7 enzymes, and a significant trend to decrease for the balance between reductive AKR1C3 and oxidative HSD17B2 activities.
2.2.5. Ocrelizumab (Ocrevus)
In the FP, ocrelizumab treatment did not show a significant trend regarding the levels of corticoids and 11β-hydroxy-androgens, SMRs that may reflect summary CYP17A1 activity, activities of HSD3Bs, CYP11B1, HSD11B1, SRD5As, AKR1D1, SMRs that may reflect balances between activities of reductive AKR1C1 vs. oxidative HSD17B2, reductive AKR1C2 vs. oxidative HSD17B2 and HSD17B6 (also possessing 3α/β-hydroxy-isomerase activity), and also reductive AKR1C3 vs. oxidative HSD17B2. Despite the absence of a significant trend in corticoid and 11β-hydroxy-androgen levels, of the most important of these, cortisol, increased significantly in FP after ocrelizumab treatment.
In FP, ocrelizumab treatment, also resulted in a borderline trend to decrease for SMRs that may reflect CYP7B1 activity, and significant trend to decrease for steroid levels and specifically for the levels of P 5α/β-reduced metabolites and also a significant trend to decrease for SMRs that may reflect activities of CYP17A1 in the hydroxylase step and at the same time, a significant trend to increase for SMRs that may reflect activities of CYP17A1 in the lyase step, and a significant trend to decrease for conjugated/unconjugated steroid ratios.
In the LP, ocrelizumab treatment did not show a significant trend in the levels of P 5α/β-reduced metabolites, SMRs that may reflect activities of CYP17A1 in the lyase step, HSD3Bs, CYP11B1, HSD11B1, SMRs that may reflect balances between activities of reductive AKR1C1 vs. oxidative HSD17B2, reductive AKR1C2 vs. oxidative HSD17B2 and oxidative HSD17B6 (also possessing 3α/β-hydroxy-isomerase activity).
In the LP, ocrelizumab treatment showed an indication of insignificant trend to lower SMRs that may reflect a balance between activities of reductive AKR1C3 vs. oxidative HSD17B2. In the LP, ocrelizumab treatment also showed a borderline trend to decrease for the levels of Δ5 and Δ4 steroids and for the SMRs that may reflect activities of CYP7B1, CYP3A4, CYP3A7 enzymes, a significant trend to decrease for the levels of steroids and specifically for corticoids and 11β-hydroxy-androgens, a significant trend to decrease for SMRs that may reflect summary CYP17A1 activity and activities of CYP17A1 in the hydroxylase step, conjugated/unconjugated steroid ratios, and for SMRs that may reflect activity of AKR1D1. It should also be noted that from individual circulating steroids in the LP, P, 5α-DHP, and 3β,5α-THP positively correlated with ocrelizumab treatment.
3. Discussion
3.1. Changes of Steroid Levels and Steroid Molar Ratios (SMRs) in MS Patients After Treatment with Anti-MS Drugs
While our previous results evaluated steroidomic alterations in drug-naïve female MS patients in FP in comparison with age-matched female controls in FP [37], our present data showed the steroidomic changes in MS-patients after treatment with anti-MS drugs. While the aggregated results from our previous study showed a significant trend towards lower steroid levels in patients in FP [37], this study demonstrated a significant trend to further decrease of steroid levels in MS patients after anti-MS treatment in FP.
Female sex hormones in FP are synthesized either directly in the adrenal glands or in peripheral tissues from adrenal precursors, while steroids in mid-cycle and LP are predominantly of ovarian origin [38]. The results from our previous study suggested an overall trend toward impaired adrenal activity in patients, which may affect the synthesis of most steroids (including active steroid hormones and neuroactive, neuroprotective and immunoprotective steroids) found downstream in the metabolic pathway [37]. However, our previous data demonstrated a trend towards positive correlations of steroid levels with the MS severity. This was interpreted that although lower steroid levels could be related to the onset of the disease, there could be a counter-regulation towards an increase during the progression of the disease. In addition, lower steroid levels were suggested as an indicator of a predisposition to MS [37]. Nevertheless, during the development of MS, the differences between steroid levels in patients and controls diminished, which could hinder the differentiation of patients with more serious MS from controls [37]. Our current data show that anti-MS treatment tends to further decrease the levels of predominantly adrenal steroids in FP, but the general trend is absent in LP. In the latter the steroidogenesis is more dependent on the ovarian activity.
In terms of changes in steroid levels after treatment with individual anti-MS drugs, significant general trends to decrease were found for S1PRI, ofatumumab in FP and also ocrelizumab in both PMCs but a trend to increase for IFNβ-1a in LP.
The steroidogenesis is mainly under adrenal control in the FP. Alternatively, in the LP, the steroidogenesis is predominantly controlled by the ovarian activity. The attenuation of steroidogenesis in FP after therapy with anti-MS drugs may be explained as weakening of protective steroid-dependent counterregulatory mechanisms depending on adrenal activity due to a beneficial effect of anti-MS therapy. In addition, in play could be the anti-steroidogenic effect of key anti-MS drugs on adrenal steroidogenesis. The trend toward increased steroid levels after IFN-β1a in LP included important bioactive steroids as the neuroprotective pregnenolone and the steroid hormones as testosterone, 5α-DHT, and cortisol, which indicated a beneficial effect of IFN-β1a treatment in LP in terms of MS severity.
Regarding the effect of GA on steroidogenesis, Cil et al. did not found changes in FSH and E2 levels in MS patients treated by a combined therapy with interferon β and GA, however the authors reported diminished ovarian volume and follicular reserve in MS patients compared to age matched healthy controls [24]. Our current results showing no effect of GA on circulating steroids in FP are in line with the data from Cil et al. [24].
Regarding the effect of IFN-β1a treatment on steroidogenesis, the only reference in the literature was from Griffiths and Wang [25], who stated that cholesterol 25-hydroxylase (CH25H) is an interferon-stimulated gene and 25-hydroxycholesterol can be considered as an immunoregulatory oxysterol that has a role in the adaptive immune system.
As to the steroidomic effect of S1PRI, Lucki and Sewer reported that S1P rapidly increased cortisol biosynthesis and the expression of genes participating in cholesterol uptake and transport in H295R adrenocortical cells [27]. Also, LH and FSH promoted S1P synthesis in the follicular cells via activation of protein kinase C. Moreover, S1P is an important mediator of FSH and LH proliferation, survival and viability effects in follicular cells [28]. However, a borderline trend to decrease in circulating corticoids and 11β-hydroxy-androstanes in association with S1PRI treatment was observed in our current data similarly like for the general trend in circulating steroids.
Although there is still no information on the effect ofatumumab on the steroidome, another anti-MS drug, which is also based on the anti-CD20 monoclonal antibody ocrelizumab, suppressed the production of a number of androstane and pregnane steroids [29], which is consistent with the attenuation of steroidogenesis after treatment with the above drugs observed in this study.
3.1.1. Corticoids (C21 Δ4 Steroids) and 11β-Hydroxy-Androgens (C19 Δ4 and 5α/β Steroids), 11β-Hydroxylase (CYP11B1) and Type 1 11β-Hydroxysteroid Dehydrogenase (HSD11B1)
MS is an autoimmune disease induced by autoreactive T-lymphocytes that is characterized by an imbalance of pro-inflammatory cytokines, such as TNF, interferon γ (IFN-γ), IL-2 and lymphotoxin, and regulatory cytokines (e.g., IL-4 and IL-10). These substances stimulate pituitary gland via hypothalamus to produce adrenocorticotropic hormone (ACTH), which in turn stimulates cortisol production in the zona fasciculata [39-41]. The secretion of ACTH is controlled by hypothalamic corticotropin-releasing hormone (CRH), pituitary ACTH, as well as by cytokines. The bioavailability of cortisol in target tissues depends on its interconversion to cortisone, which is inactive. The balance between bioactive cortisol and inactive cortisone is regulated by the reductive enzyme HSD11B1and the oxidative enzyme HSD11B2. In contrast to HSD11B1 that may influence cortisol circulating levels, the latter enzyme is primarily expressed in selective tissues acting mainly in paracrine and autocrine manner and does not substantially influence the circulating cortisol [42]. Glucocorticoids play a decisive role in the regulation of the immune system and act through binding to the low affinity glucocorticoid receptors. Although glucocorticoids are mainly a product of the adrenal zona fasciculata, they can be produced extra-adrenally, for example in cells of the immune system, intestine, skin, or brain [42]. Our current data did not find a significant trend in SMRs that may reflect the HSD11B1 activity (n=4) but this result cannot be considered as conclusive due to the small number of relevant SMRs and thus the low statistical power of the test.
In addition to the response to emotional and cognitive stress, HPAA also regulates the interaction between peripheral inflammatory processes and cortisol secretion [43,44]. Most authors have reported that MS patients have overactive HPAA with elevated cortisol levels (summarized in [45]) but others found unchanged circulating cortisol levels in patients or even reduced cortisol levels in the cerebrospinal fluid of patients compared with controls [46,47]. Foroughipour et al. [47] indicated that chronic activation of HPAA occurs in patients. However, Wei and Lightman suggested that HPAA is unlikely to play a major role in the initial pathogenesis of MS, but the overactivation of the HPAA in MS patients is secondary to an active inflammatory stimulus [48].
Our current data showed that the anti-MS treatment reduced the levels of active glucocorticoid cortisol and corticosterone, which is a precursor of the mineralocorticoid aldosterone pathway, as well as the inactive metabolite of cortisol, cortisone. As the SMRs that may reflect CYP11B1 did not show significant change, this trend may be ascribed to the attenuated adrenal activity after anti-MS treatment instead of blocked CYP11B1 activity. However, the number of above SMRs was low (n=7) and therefore a definite conclusion could be not be made.
Our previous study indicated an absence of serious blocks in the pathway of cortisol synthesis, which was indicated by the lower levels of 17-OH-P at elevated levels of cortisol and cortisone [37]. Our current data also suggested an absence of specific enzyme blocks in cortisol pathway associated with anti-MS treatment (unaffected MS treatment CYP17A1, HSD3B, CYP11B1 and HSD11B1) activities). The only change associated with anti-MS treatment on the cortisol pathway was an increase in pregnenolone levels, but this occurred with a parallel increase in DHEA levels, again indicating an absence of enzyme blocks.
Corticoid and 11β-hydroxy-androgen levels showed no significant trend after GA treatment in FP. However, IFNβ-1a treatment in LP increased the levels of these steroids, whereas ocrelizumab suppressed them in this menstrual phase. These changes in LP were consistent with trends found for steroids in general and like the pooled trends of steroids in LP for IFN-β1a and ocrelizumab treatments. From the biologically most important steroids also the levels of cortisol showed significant increase after IFN-β1 and ocrelizumab treatment in LP.
No changes were observed in corticosteroid levels after S1PRI treatment in our present data. Our data are not consistent with the results of Lucki and Sewer, who reported that S1P rapidly increased cortisol biosynthesis [27].
Of note, our data for cortisol (Figure 1) were in line with data from Hamidović et al. [49]who proved using meta-analysis that the cortisol levels were lower in the follicular phase.
3.1.2. Δ5 and Δ4 Steroids
Our previous study did not find significant associations of MS with these steroids except of lower levels of 7β-hydroxy-DHEA (7β-OH-DHEA). Concerning the changes of Δ5 and Δ4 steroids in association with anti-MS treatment, there was a parallel increase in pregnenolone and DHEA levels as mentioned above, which may be of importance as both steroids are neuroprotective. They cross the BBB and therefore their adrenal production and/or therapeutic supplementation may affect their concentration in the brain [13,14]. Pregnenolone is pro-cognitive and neuroprotective against glutamate-induced neurotoxicity [15]. Furthermore, sulfates of pregnenolone and DHEA positively modulate NMDARs and σ1 receptors (σ1R) [6,15,50]. The protective effects of DHEA and its sulfate can be attributed to their modulation of type A GABA receptors (GABAAR) and protection of mitochondria from intracellular Ca2+ overload [15]. In addition, DHEA and its sulfate have antioxidant and anti-inflammatory potential also outside the CNS [16,51]. Therefore, specific increases in pregnenolone and DHEA can be considered one of the beneficial effects of anti-MS therapy. From the anti-MS drugs, GA did not show an effect on Δ5 and Δ4 steroids. In the FP, the treatment with IFNβ-1and with S1PRI showed an insignificant indication of decrease of these steroids, treatment with ocrelizumab showed a borderline trend to their decrease, and the treatment with ofatumumab exhibit a trend to decrease the levels of the Δ5 and Δ4 steroids. In the LP, ocrelizumab treatment had the same effect as in the FP. These trends were also similar for all steroids.
3.1.3. Active Androgens and Estrogens
Androstenedione metabolite T, like T metabolite 5α-DHT are important neuromodulators. However, their neuromodulatory effects are sex dependent (see reviews [23,52]). T is a modulator of the mesocorticolimbic system affecting the density of dopaminergic neurons [51]. Nevertheless, only T, not 5α-DHT has a direct neuroprotective effect, suggesting that T and 5α-DHT may have independent effects on the hippocampal immune cells (see review [52]).
The experiments with EAE, induced by helper T-lymphocytes showed that MS patients have a predilection for Th1, which may be associated with low T levels (see review [52]). The latter finding is in accordance with our previous study [37].
Surprisingly, RRMS patients with higher T levels were more likely to have tissue damage [53]. Based on correlations with MRI data, the authors [53]considered the increase of protective steroids E and T to be a response to brain damage [53]. The active androgens shift from Th1 to Th2 phenotype. Increased IFN-γ secretion due to this change may contribute to the known susceptibility of female experimental animals to induction of autoimmune diseases., including EAE [1].
Other effects of T include proliferation and differentiation of lymphocytes and inhibition of immunoglobulin production. Furthermore, T can penetrate the BBB and protect neuronal cells from glutamate toxicity, increase neurite outgrowth, and brain-derived neurotrophic factor expression [1]. Androgens also indirectly increase T-cell thymocyte apoptosis and thus exert neuroprotective effects in EAE (see review [21]). In addition, T reduces reactive gliosis and astrocyte proliferation, which promotes axon regeneration and also promotes brain lesion repair in MS patients and animal models [54-57]. However, sex hormones can not only protect brain tissue, but also they can have excitotoxic and apoptotic effects [58] (see also review [23]).
Data from our previous study showed lower levels of A and T compared to controls, but non-significantly different levels of conjugated and unconjugated 5α-DHT [37].
Data of the current study show an increase in circulating T after anti-MS treatment for LP with unchanged A levels, but show a decrease in conjugated 5α-DHT levels. The observed increase in T levels after treatment could be related to the progression of the disease, but also to the effect of anti-MS treatment itself. The increase in T levels with unchanged A levels is consistent with the results for the T/A ratio, which could reflect a shift in the balance between the activities of the reductive AKR1C3 and oxidative HSD17B2 to the former enzyme. However, in the aggregate, no significant trend was found for this balance in terms of treatment effect. Since the literature data point to predominantly positive effects of T on the condition of MS patients, it can be assumed that anti-MS treatment also has a positive effect in this respect, at least in LP.
With regard to the active estrogen E2, unlike testosterone, no changes in circulating levels were detected after anti-MS treatment, regardless of the PMC. As for the decrease in E1 sulfate levels, this result is consistent with the pooled trend toward a decrease in circulating steroid levels after anti-MS treatment.
Concerning the effects of anti-MS treatment on androstenedione and active androgens (T, 5α-DHT and 5α-DHTC), the number of variables was insufficient to assess their pooled trend after anti-MS treatment. Nevertheless, while GA and IFNβ-1, and S1PRI in FP did not show any effect, T and 5α-DHT were higher after IFNβ-1a therapy in LP. Androstenedione and 5α-DHT decreased after ofatumumab therapy and 5α-DHT decreased after ocrelizumab treatment in LP but not in FP.
3.1.4. Progesterone and Its 5αβ-Reduced Metabolites
The P metabolite 3α,5α-THP has various neuroprotective effects including alleviation of neurobehavioral deficits and attenuation of neuropathology and inflammation in animal models of autoimmune demyelination [21,22]. Moreover, recent studies demonstrated that 3α,5α-THP suppresses neuroinflammation through activation of TLR4 protein in macrophages and in the brain [51].
Although there were no consistent changes in circulating P 5α/β-reduced metabolites, levels of 3α-hydroxy-5α/β-steroids tended to decrease in FP after anti-MS treatment, whereas their 3β-hydroxy counterparts did not. Thus, the decline in the previously mentioned steroids that are neuroprotective could also be related to the attenuated counterregulatory activity against the effects of MS due to the beneficial effects of anti-MS treatment.
From the anti-MS drugs, GA did not show an effect on P 5α/β-reduced metabolites. IFNβ-1a treatment in the FP indicated an insignificant tendency to decrease of these steroids, whereas the treatment with S1PRI, ofatumumab, and ocrelizumab reached significance. In the LP, IFNβ-1a treatment showed a significant trend to increase, while for ocrelizumab there was no significant trend. Progesterone, 5α-DHP, and 3β, positively correlated with ocrelizumab treatment in LP. Progesterone has promyelinating effects on the CNS, acts against glutamate toxicity, normalizes functional defects of injured neurons, enhances proliferation and differentiation of oligodendrocyte precursor cells. In addition, P is an immunomodulator that shifts the pro-inflammatory Th1 response to an anti-inflammatory Th2 response and promotes a reduction in IFN-γ production by natural killer cells. [59,60] (see also review [23]). Therefore, the increase in P levels in LP after treatment with ocrelizumab could have a beneficial effect on the severity of MS.
3.1.5. C17-Hydroxylase, C17-20-Lyase (CYP17A1), Hydroxylase and 3β-Hydroxysteroid Dehydrogenases (HSD3Bs)
The SMRs, which may reflect summary CYP17A1 activity in the hydroxylase and lyase steps, did not show a significant trend for change after anti-MS treatment. However, the SMRs, which may reflect CYP17A1 activity separately in the hydroxylase step, showed a borderline trend toward a decrease in FP, but a borderline trend toward an increase in CYP17A1 activity in the lyase step. In addition, all conjugated 17-hydroxy-5α/β-pregnanes showed a significant decrease in the FP (Figure 4).
In this context Gupta et al. reported that 17-hydroxyallopregnanolone (3α,5α17-PD) was rapidly converted to androsterone (3α,5α-THA) under CYP17A1 catalysis in a lyase step, which occurred even in the absence of cytochrome B5 (CYB5) [61]. The authors found that 3α,5α17-PD was a better substrate for CYP17A1 than 17-OH-Preg. It should be mentioned that CYB5, which activates the CYP17A1 lyase step, is expressed about four times more in the adrenal cortex than in other tissues (http://biogps.org/#goto=genereport&id=80777, accessed on 26 December 2024). However, CYP17A1 is more than 1500 times expressed in the adrenal cortex compared to the other tissues and approximately 70 and 40 times more in the kidneys and testes, respectively. (http://biogps.org/#goto=genereport&id=1586, accessed on 26 December 2024).
These data suggest that the adrenal cortex forms mainly 17-hydroxy C21 steroids and C19 Δ5 steroids from their 17-deoxy C21 precursors, whereas 5α/β-reduced steroids are formed mainly extra-adrenally, independent of the enzyme CYB5, which is weakly expressed outside the zona reticularis of the adrenal gland. [62]. This may be the reason why in the synthesis of Δ5 androgens the adrenal cortex prefers the Δ5 pathway, while in other tissues the synthesis of 5α/β-reduced androstanes occurs and it is independent of CYB5. Our results showed very low circulating levels of intermediates such as 17-hydroxyallopregnanolone sulfate (3α,5α,17-PDC), 3α,5β,17-PD, and 3α,5β,17-PD) in the pathway from 17-deoxy-5α/β-reduced pregnanes to 5α/β-reduced androstanes, which indicated their rapid conversion to the corresponding androstanes.
There was no significant trend in SMRs that could reflect summary CYP17A1 activity and CYP17A1 activities separately in the hydroxylase and lyase steps. Otherwise, all of the conjugated 17-hydroxy-5α/β-pregnanes decreased significantly in FP (Figure 4), but none of their unconjugated counterparts. Whereas of the 17-hydroxy- Δ5 and Δ4 steroids only cortisol decreased significantly in FP. In the present study, the trend toward a decrease in 17-hydroxy-pregnanes after treatment with anti-MS drugs in FP, but not the one in SMRs, suggests a possible downward shift in CYP17A1 activity in the hydroxylase step. This may prompt a weakening of one of the steps in the pathway toward cortisol. Our data also indicate that monoclonal anti-CD20 mAbs, S1PRI and IFNβ-1a may be responsible for attenuating CYP17A1 activity while enhancing the lyase step. In LP, only ocrelizumab, but not IFNβ-1a, promoted total CYP17A1 activity and CYP17A1 activity at the hydroxylase step, with no effect at the lyase step. To summarize, the aforementioned findings could have implications for the synthesis and metabolism of bioactive steroids, including cortisol, active androgens and estrogens, or neuroactive and neuroprotective substances.
In the case of HSD3Bs (involved in the synthesis of cortisol as well as active androgens and estrogens), the anti-MS drug treatment does not seem to affect their activity.
3.1.6. A Balance Between Steroid Sulfotransferase (SULT2A1) and Steroid Sulfatase (STS)
Regarding the neuroactive steroids, unconjugated steroids and their corresponding sulfates are often antagonists at the same receptors or may act antagonistically in relation to neuronal activity at different receptors [63,64,65]. Preg and DHEA sulfates are known to modulate several types of ionotropic receptors, such as NMDARs, AMPARs, nicotinic receptors, TRPM3s, TRPC5s or TRPV1s, and may improve cognitive function while counteracting pain and fear transmission [7,17].
The aggregated data in our previous study showed a significant trend towards lower levels of unconjugated steroids but not in steroid conjugates in MS-patients compared to controls. This finding was ascribed to increased SULT2A1 activity in MS patients compared to controls. Moreover, the SMRs that may reflect the balance between SULT2A1 and STS showed a significant trend towards elevated values in patients compared to controls, which indicated an increased SULT2A1 activity in MS patients.
Our current data showed a significant trend to decrease of the ratios of conjugated steroids to their unconjugated counterparts in FP after anti-MS therapy, which indicated either suppression of SULT2A1 activity and/or stimulation of STS activity by anti-MS drugs. In addition to a significant pooled trend toward a decrease in the ratios of conjugated steroids to their unconjugated counterparts after treatment with anti-MS drugs, a significant trend toward a decrease in these ratios was found for GA in FP, while for ocrelizumab this trend was observed in both PMCs. Moreover, our current data showed a significant trend towards a decrease in circulating conjugated steroids in FP.
These data may be of relevance for regulation of neuronal excitability. Unconjugated steroids act through binding to nuclear receptors while their sulfates are inactive. Nevertheless, steroids may be accumulated in the sulfate form and may be consequently hydrolyzed to active hormones [66]. Furthermore, sulfation of unconjugated neuroinhibitory and neuroprotective GABAergic steroids leads to their inactivation or even to the formation of their antagonists [64,67]. Conversely, some sulfated steroids can be positive or negative modulators of excitatory glutamate receptors, whereas all their unconjugated analogues are inactive in this context [68,69]. In addition, sulfated androgens may serve as a reservoir of substrates for the synthesis of bioactive androgens acting as sex hormones, neuroprotective and immunomodulatory agents.
3.1.7. 11β-Hydroxylase (CYP11B1) and 11β-Hydroxysteroid Dehydrogenase Type 1 (HSD11B1)
11β-Hydroxylase (CYP11B1) and 11β-hydroxysteroid dehydrogenase type 1 (HSD11B1) are important enzymes participating in cortisol synthesis and homeostasis. CYP11B1 catalyzes the final step in cortisol synthesis converting 11-deoxycortisol to cortisol as well as 11-deoxy-androstanes to 11β-hydroxy-androstanes. 11β-Hydroxy-androstanes cannot be formed from cortisol and its 5α/β-reduced metabolites under CYP17A1 catalysis in the lyase step [70]. Our present data show a borderline trend toward a decrease in CYP11B1 after anti-MS therapy, which could lead to a decrease in cortisol synthesis in this last metabolic step. However, the effect of individual anti-MS drugs on SMRs that may reflect CYP11B1 activity was not observed, probably due to the low number of relevant SMRs combined with insufficient number of patients.
HSD11B1 is an important diabetogenic enzyme that converts inactive cortisone to the active glucocorticoid cortisol [71]. However, our current data do not even suggest that anti-MS treatment could affect the activity of HSD11B1, which otherwise increases the level of the circulating glucocorticoid cortisol at the expense of inactive cortisone.
3.1.8. 7α-,7β-, and 16α-Hydroxylating Enzymes (CYP7B1, CYP3A4, CYP3A7) and 11β-Hydroxysteroid Dehydrogenase, Type 1 (HSD11B1)
In our previous study [37], 7α-hydroxy-DHEA and 3β,7β,17β-AT showed reduced levels in MS patients versus controls, despite DHEA levels being significantly higher in patients. Concerning our current data, although the levels of 7α/β- and 16α-hydroxy-steroids did not show a significant trend towards changes after treatment with anti-MS drugs, the SMRs of 7α/β- and 16α-hydroxy-steroids to the parent steroids suggested a reduction in the activities of the enzymes catalyzing their synthesis (CYP7B1, CYP3A4 and CYP3A7) of borderline significance despite the low number of corresponding SMRs (n=8). Regarding individual anti-MS drugs, there was a trend towards lower values of the above SMRs reaching significance with ofatumumab, a non-significant indication of this trend with ocrelizumab in FP and the same trend of borderline significance with ocrelizumab in LP, suggesting which anti-MS drugs contribute to the pooled effect of anti-MS therapy.
It should be pointed out that reduced activity of 7α/β- and 16α-hydroxylation enzymes may have a significant impact not only on the etiopathogenesis of the disease, but also on its course and further development. It is clear from the abundant data in the literature that Δ5 androstanes (and particularly their immunoprotective and antidiabetic 7α/β- and 16α-hydroxy-metabolites) alleviate the severity of autoimmune diseases, while these diseases can suppress the synthesis of adrenal Δ5 androstanes [72,73,74,75,76,77,78]. Thus, the latter finding could be related to the reduced activities of 7α/β- and 16α-hydroxylation enzymes in MS patients compared to controls. Interestingly, there is a further reduction of these enzymes in MS-patients as a result of treatment with anti-MS drugs.
DHEA is known to control the Th1/Th2 balance and either favors the Th1 component or attenuates the production of both components [76,79]. The Δ5 androstanes also suppress cellular immunity, autoantibody production [74,75,76,77,80] and are capable of forming a Th1-dominated cytokine profile. The Δ5 androstanes and their 7α/β-,16α-hydroxylated metabolites may also prevent suppression of the primary immune response by glucocorticoids [81].
Two mechanisms have been proposed to explain the immunomodulatory effects of 7α/β- and 16α-hydroxy-metabolites of Δ5 androstanes. The first mechanism has suggested a link between the immunomodulatory effects of 7α/β-hydroxy-∆5-androstanes and their competition for active sites on 11β-hydroxysteroid dehydrogenase (HSD11B1), an enzyme that catalyzes the conversion of inactive cortisone to immuno-suppressive cortisol [82,83].
The second mechanism has suggested that E2 may induce the autoimmune response via estrogen receptors and catabolism of estrogen precursors such as DHEA and Adiol to their 7α/β- and 16α-hydroxylated metabolites reduces the concentration of these estrogen precursors and consequently E2 [84]. In addition, E2 promotes the catalytic activity of CYP7B1 and may thus have a feedback effect on the regulation of DHEA, Adiol and its own levels [85].
Moreover, the Adiol catabolite 3β,7β,17β-AT, which can be synthesized either by interconversion from 5-androstene-3β,7α,17β-triol or directly from Adiol by the catalytic activity of CYP3A4 and CYP3A7, is itself immunoprotective, despite its very low concentrations and high metabolic turnover [86].
Based on our previous data [37], we hypothesized that increased 7α-, 7β-, and 16α-hydroxylation play a role in the transition from adaptive immunity involving autoimmunity to the innate immune system involving inflammatory processes [7]. Nonetheless, synthetic anti-inflammatory derivatives of 3β,7β,17β-AT also suppress the production of inflammatory markers and pro-inflammatory cytokines [87].
Thus, the above data suggest that impaired 7α/β-hydroxylation of Δ5 androstanes may be involved in the pathophysiology of MS, impeding the formation of more potent immunomodulatory metabolites.
To summarize, the impaired 7α/β-hydroxylation of Δ5 androstanes, which appears to be primarily associated with anti-CD20 mAb may on the one hand be associated with an increase in autoimmune response. On the other hand, a shift in the balance from the Th1 pattern towards the Th2 pattern resulting from the aforementioned impairment could attenuate acute inflammatory processes, so that decreased 7α/β-hydroxylation of Δ5 androstanes may not be a clearly undesirable phenomenon in terms of MS severity.
3.1.9. 5α-Reductases (SRD5As), 5β-Reductase (AKR1D1) and Aldoketoreductases Subfamily1c (AKR1C1-4)
The role of SRD5As, AKR1D1 and aldoketoreductases subfamily1C (AKR1C1-4) in relation to MS was discussed in detail in our previous study [37]. In summary, our current data mostly did not show significant trends in SMRs changes after treatment with anti-MS drugs, which may reflect the activity of these enzymes. Nevertheless, the SMRs that may reflect the activities of steroidogenic enzymes or balances between them indicate that S1PRI stimulates the activity of SDR5As in the FP. Also, the transcriptional analyses in the study by Noorbakhsh et al. showed significantly lower SRD5A1 transcripts in the white matter of MS-patients postmortem compared to corresponding control samples [21]. Unfortunately, the authors did not specify the medication used.
Otherwise, our previous study reported that all indices of MS severity tended to correlate positively with 5α-reduced steroids [37], suggesting that these findings may also be related to the counter-regulatory effects of 5α-reduced steroids, which include a number of neuroprotective agents [21,22,23,64,67], (see also reviews [52,88,89]). Nevertheless, our previous study also showed that in contrast to trend to positive correlations of 5α-reduced steroids with indices of MS severity, the SMRs that may reflect the functioning of SRD5As were unrelated to MS presence as well as to indices of MS severity. These results suggested that changes of SRD5As activities were not directly associated with MS pathophysiology. Therefore, the aforementioned attenuation of SDR5As activities may be related to the effect of treatment with S1PRI.
Various 5α/β-reduced metabolites of P and A are neuroprotective substances [90,91,92,93] that have anti-inflammatory effects, stimulate myelination and remyelination of Schwann cells in the peripheral nervous system, protect mitochondria, regulate neurogenesis, affect mood, memory, cognition (see reviews [94,95]) and protect nervous system cells from hyperexcitation. The 5α/β-reduced C21 and C19 steroids are active at a number of receptors including GABAA (γ-aminobutyric acid type A) and glutamate receptors (see review [95]).
On one hand, S1PRI treatment resulted in a significant overall trend to decrease of steroid levels, and specifically for the levels of P 5α/β-reduced metabolites, but on the other, there was a significant trend to increase for SMSs that may reflect SRD5As and activity. Therefore, the tendency to stimulate the activity of both SRD5A by S1PRI may partially mitigate the adverse consequences of the overall trend of decreasing steroid levels after S1PRI medication, including 5α/β P metabolites.
SMRs suggest that AKR1D1 activity in LP may be stimulated by IFNβ-1a as well as by ocrelizumab. Therefore, the tendency for stimulation of both SRD5As and AKR1D1 activities by the abovementioned anti-MS drugs may be beneficial for MS patients.
Concerning the balance between the activities of reductive AKR1C1 and oxidative HSD17B2 (as assessed from SMRs), the only effect was found for GA shifting the balance from the former to the latter enzyme. Our previous data did not found trends in correlations with neither indices of MS severity nor with MS presence [37], which was consistent with the results of transcriptional analysis of AKR1C isoforms in the study by Noorbakhsh et al. that also show no significant alteration for the AKR1C1 transcripts in the white matter of patients when compared with controls [21]. Therefore, the aforementioned shift observed in our present data may be considered as the effect of GA therapy.
The relationship between GA therapy and a shift in the balance from the AKR1C1 to the HSD17B2 may be important as AKR1C1 suppresses lipid peroxidation of byproducts with catalytic efficiency comparable to that for 20α-hydroxysteroids, and provides an inducible cytosolic barrier for 4-hydroxy-2-nonenal formed during lipid peroxidation after exposure to reactive oxygen species (ROS). In addition, AKR1C1 stimulation induces the activity of the transcription factor NF-E2-related factor-2 (NRF2), a key player in the regulation of antioxidant pathways [96]. Therefore, in terms of its effect on the steroidome, GA treatment may be adverse because it may be associated with the promotion of inflammatory processes accompanying MS.
The balance between reductive AKR1C2 on one hand and oxidative HSD17B2, and oxidative and at the same time 3α/β isomerizingHSD17B6 on the other is of particularly importance for neuroactive steroid biosynthesis, as AKR1C2 catalyzes the last step in the reduction of P to pregnanolone isomers including the most important of them 3α,5α-THP (allopregnanolone) and 3α,5β -THP (pregnanolone), which are a neuro-inhibitory and neuroprotective GABAergic steroids [21,22,23,64,67], (see also reviews [52,88,89]).
Noorbakhsh et al. found that although there was no difference between MS patients and controls regarding pregnenolone, there was significantly lower 3α,5α-THP in the white matter of patients compared to controls [21]. However, these alterations were specific for 3α,5α-THP, while the remaining pregnanolone isomers, including GABAergic 3α,5β-THP did not differ between MS patients and controls [21]. These observations were in accordance with data from our previous study [37] and were also consistent with data from transcriptional analysis of the same authors who found significantly lower levels of AKR1C2 transcript in the brains of the MS patients compared to controls [21]. In addition, Noorbakhsh et al also reported significantly lower mRNA immunoreactivity for AKR1C2 in MS patients compared to controls [61]. Concerning our present results, no significant effect of anti-MS treatment on SMRs reflecting the balance between activity of AKR1C2 and activities of HSD17B2 and HSD17B6 was found.
CNS inflammation and immune dysfunction play a key role in the pathogenesis of MS [1,52,89,97]. AKR1C3 (further enzyme from the AKR1C subfamily) is not only involved in steroidogenesis, but it is also effective as prostaglandin (PG) F2α synthase. At the same time, a highly active metabolite of PGF2α, 8-iso-prostaglandin F2α (8-iso-PGF2α) stimulates oxidative stress and contributes to the inflammatory process [98,99,100]. AKR1C3 is highly expressed in immunocompetent cells, adipose tissue, intestine, smooth muscle, bronchial cells, colon, and liver, but its expression has also been detected in adrenal zona reticularis and in a variety of other tissues [101,102,103], http://biogps.org/#goto=genereport&id=8644, accessed on 26 December 2024) (see also reviews [104], [19] and [105]).
Our previous study showed a borderline trend towards higher values of the SMRs that may reflect the balance between reductive AKR1C3 and oxidative HSD17B2 in MS patients. This may contribute to the increased incidence of inflammatory responses in MS patients [37]. Our current data suggested that ofatumumab in the FP (observed significant trend to decrease after treatment) and ocrelizumab in the LP (insignificant decrease after treatment) may shift a balance from the reductive proinflammatory AKR1C3 to oxidative HSD17B2. This data may indicate an attenuation of proinflammatory milieu associated with lower AKR1C3 activity after treatment with humanized anti-CD20 monoclonal antibodies.
4. Potential Clinical Implications of the Findings
This study can be considered the first attempt to systematically analyze the relationship between endogenous steroids and anti-MS drug treatment. The findings may be important concerning alterations in circulating bioactive steroids such as cortisol, active androgens and estrogens, neuroactive, neuroprotective and immunomodulatory substances in terms of an optimization of the MS treatment.
5. Future Directions
Respecting the use of SMRs as surrogate markers for estimation of functioning of steroidogenic enzymes, further research should focus on direct assessment of the enzyme activities and expression of respective steroidogenic enzymes. Also, in respect of insufficient data for some anti-MS drugs in our study future studies should focus on individual anti-MS drugs with sufficient numbers of volunteers.
6. Limitations of the Study
The reported study results were focused on steroidomics only. Thus, the assessments of changes in steroidogenic enzyme functioning after anti-MS used only the steroidomic surrogate markers based on SMRs and the changes in activities of steroidogenic enzymes were not directly tested.
Although this study includes a number of steroids and covers most of the steroid metabolic pathways, the study does not include 11-deoxycortisol and 11-deoxy-corticosterone.
In addition, the number of patients included in the present study was satisfactory for the estimation of pooled trends, but there was insufficient data concerning some of the individual anti MS-drugs and therefore the respective results are preliminary. Nevertheless, despite the limited sample size, our patient population was homogeneous, i.e., included only Caucasian patients due to the demography of the Czech Republic.
7. Materials and Methods
7.1. Subjects
A total of 60 female MS patients of reproductive age 39 (29, 47) years was included in this study (shown as median with quartiles). The diagnosis multiple sclerosis was confirmed by cerebrospinal fluid analysis and by magnetic resonance imaging. All of the MS patients fulfilled the revised 2017 McDonald criteria [106]. The MS patients included in this study had just been diagnosed and had not yet been treated. Patients and controls who had experienced COVID were not included in the present study.
The study was approved by the Ethics Committee of the General University Hospital, Prague, Czech Republic (Approval number: 74/19 Grant AZV VES 2020 VFN, 20 June 2019), and all procedures involving human subjects were conducted following ethical standards set by national and institutional committees on human experimentation and the Helsinki Declaration of 1975, as updated in 2008. The authors guarantee that all research procedures were carried out with the utmost respect for the participant's safety, well-being, and confidentiality. Participants were examined after signing an informed consent approved by the aforementioned Ethics Committee. For the evaluation of steroidome, the peripheral blood was withdrawn on fasting in the morning. Blood samples were centrifuged and stored at -20˚C until analyzed.
7.2. Steroid Analyses
Steroids and their polar conjugates were measured using our previously described validated GC-MS/MS method [107] with the exception of E2, which was quantified using Electro-chemiluminescence immunoassays (ECLIA), performed on Cobas® Pro, Roche Diagnostics International Ltd. (Rotkreuz, Switzerland). In addition to the use of GC-MS/MS, cortisol was also measured using the RIA cortisol kit from Orion, Espoo, Finland.
7.3. Statistical Analyses
In the first step, the power transformation parameters were found for each metric variable so that its distribution was as close as possible to the Gaussian distribution. The steroidomic data were evaluated using an ANOVA model as well as multivariate regression with reduced dimensionality known as orthogonal projections to latent structure (OPLS) model. The ANOVA consisted of subject factor separating the inter-individual variability, factor of menstrual phase (PMC, follicular vs. luteal), Stage factor (Stage 0, before treatment vs. Stage 1, after treatment) and PMC × Stage interaction. Statgraphics Centurion v. XVIII statistical software from Statgraphics Technologies, Inc. (The Plains, Virginia, USA) was used for power transformations of the original data and for evaluation using the ANOVA model, while SIMCA-P v.12.0 statistical software from Umetrics AB (Umeå, Sweden) was used for OPLS analysis.
The OPLS models were focused on the differentiation between naïve MS patients and those treated with individual anti-MS drugs. In terms of diagnosing MS based on highly intercorrelated steroidomic data, it was appropriate to use OPLS models that examined the correlation of MS with multiple parameters simultaneously. The OPLS model, which is a multivariate regression with dimensionality reduction, permits the evaluation of relationships between explanatory variables and a number of explanatory variables that may be highly correlated, which is also the case for steroids in metabolic pathways. The use of the given anti-MS drug is expressed in the OPLS model as the logarithm of the likelihood ratio (the ratio of the probability of its use p to the probability of its absence (1-p)), i.e. the logarithm of the likelihood ratio is calculated, which then ranges from -infinity to +infinity. This approach ensures that the prediction of the probability of the presence of pathology is between 0 and 1 (after using a recurrent formula that converts the logarithm of the likelihood ratio to the probability of the presence of pathology).
The variability of the explaining and explained variables is separated into two independent components in the OPLS. The former contains the variability in explaining variables that were shared with the probability of pathology (predictive component), while the orthogonal components express the variability shared in between highly correlated explaining variables (orthogonal components). OPLS identifies significant explanatory variables and their best linear combination to estimate the probability of the presence of pathology. After standardizing the variables, the OPLS model can be expressed as follows:
where X is the matrix with predictors and subjects, Y is the vector of dependent variable and subjects; Tp is the vector of component scores from the single predictive component and subjects extracted from Y; To is the vector of component scores from the single orthogonal component and subjects extracted from X; Pp is the vector of component loadings for the predictive component extracted from Y; Po is the vector of component loadings for the orthogonal component extracted from X and independent variables; and E and F are the error terms.
Significant predictors were selected using the variable importance statistics (VIP). The statistical software SIMCA-P v.12.0 from Umetrics AB (Umeå, Sweden), which was used for OPLS analysis, enabled finding the number of relevant components, the detection of multivariate non-homogeneities, and testing the multivariate normal distribution and homoscedasticity (constant variance).
The algorithm for obtaining the predictions were as follows:
- Transformation of the original data to obtain the values with symmetric distribution and constant variance
- Checking the data homogeneity in predictors using Hotelling’s statistics and the eventual elimination of non-homogeneities
- Testing the relevance of predictors using variable importance statistics and the elimination of irrelevant predictors
- Calculating component loadings for individual variables to evaluate their correlations with the predictive component
- Calculating regression coefficients for the ordinary multiple regression model (OMR) to evaluate the mutual independence of predictors after comparison with the corresponding component loadings from the OPLS model
- Calculating predicted values of the logarithm of the ratio of the probability of pathology presence to the probability of pathology absence (LLR)
- Calculating the probability of the pathology presence for individual subjects
- Calculating the sensitivity and specificity of the prediction
The ratio between significantly positive, missing and significantly negative correlations with MS was evaluated using the one-sample Wilcoxon test with correction for continuity.
8. Conclusions
In conclusion:
- 1)
- A comprehensive steroidomic analysis of steroidomic changes after treatment with anti-MS drugs was performed. The study participants were MS newly diagnosed female patients who met the 2017 revised McDonald criteria and had not yet been treated.
- 2)
- Almost all steroids have been studied for the first time in terms of the effects of treatment with anti-MS drugs.
- 3)
- MS patients showed a general tendency towards a decrease in steroid levels in follicular menstrual phase due to treatment with anti-MS drugs but absence of this trend in luteal phase, which could be interpreted as a weakening of counterregulatory mechanisms associated with adrenal steroids, which may mitigate adverse effects of MS. Another explanation could be the anti-steroidogenic effect of key anti-MS drugs on adrenal steroidogenesis.
- 4)
- Decreased 17-hydroxy-pregnane levels after treatment with anti-MS drugs in FP indicate a decreasing trend of CYP17A1 activity, suggesting a weakening of one of the steps of the cortisol pathway and, together with the analysis for individual anti-MS drugs, a strengthening of the step towards immunomodulatory adrenal androgens. These results also suggest that among the anti-MS drugs, the monoclonal anti-CD20, S1PRI and IFNβ-1a antibodies may be responsible for the above findings. These findings could have implications for the synthesis and metabolism of bioactive steroids, whether they are cortisol, active androgens and estrogens, or neuroactive and neuroprotective substances acting through modulation of ionotropic receptors.
- 5)
-
A significant trend to decreased ratios of conjugated steroids to their unconjugated counterparts was observed indicating either suppression of SULT2A1 activity and/or stimulation of STS activity by anti-MS drugs, from which GA and ofatumumab in FP, and ocrelizumab in both menstrual phases contributed this pooled trend. Moreover, our current data show a significant trend towards a decrease in conjugated steroids level in FP after anti-MS drug therapy. The above data may be relevant to the regulation of neuronal excitability because:
- a. Unconjugated steroids act through binding to nuclear receptors while their sulfates are inactive.
- b. Sulfation of unconjugated neuroinhibitory GABAergic steroids leads to their inactivation or even to the formation of their antagonists.
- c. Some sulfated steroids can be positive or negative modulators of excitatory glutamate receptors, whereas all their unconjugated analogues are inactive in this context.
- d. Sulfated androgens may serve as a reservoir of substrates for the synthesis of bioactive androgens acting as sex hormones, neuroprotective and immunomodulatory agents
- 6)
- A borderline downward trend was observed in CYP11B1 after anti-MS therapy, which may downregulate cortisol synthesis in the last metabolic step.
As for further changes in steroid levels after treatment with individual anti-MS drugs:
- 1)
- GA treatment in FP also shifted the balance from reductive AKR1C1 to oxidative HSD17B2 which may contribute to the inflammatory milieu.
- 2)
-
In addition to affecting CYP17A1 activities, IFN-β1a also showed:
- a. A significant trend toward increased steroidogenesis in LP, which is predominantly controlled by ovarian activity but no effect in FP, which is predominantly controlled by adrenal activity.
- b. A significant trend toward increased steroid levels after IFN-β1a in LP included significant increases in some important bioactive steroids such as the neuroprotective pregnenolone and steroid hormones such as testosterone, 5α-DHT, and cortisol. These observations indicate a beneficial effect of IFN-β1a treatment in terms of MS severity.
- c. A significant trend to higher SMRs that may reflect AKR1D1 activity which could be of importance in terms of the synthesis of neuroinhibitory 5β-pregnanes.
- 3)
- S1PRI (in addition to affecting a general trend in steroid levels and SMRs that may reflect CYP17A1 activities) also showed a trend to lower SMRs that may reflect SRD5A activity.
- 4)
-
Anti-CD20 mAb (ofatumumab, ocrelizumab) treatment (in addition to affecting a general trend in steroid levels and SMRs that may reflect CYP17A1 activities and shift in the balance conjugated/unconjugated steroid) showed:
- a. A borderline trend to decrease for the SMRs that may reflect the activities of HSD3Bs (ofatumumab).
- b. A significant (ofatumumab) or borderline (ocrelizumab in LP) trend, or at least indication of the trend (ocrelizumab in LP) to lower values of SMRs that may reflect activities of CYP7B1, CYP3A4 and CYP3A7 enzymes catalyzing the synthesis of immunomodulatory 7α/β and 16α-hydroxy-androgens. Moreover, there was a pooled borderline downward trend in their levels after therapy with anti-MS drugs. These data are relevant concerning the immunomodulatory effect of these steroids as they affect Th1/Th2 balance and thus may influence the severity of MS.
- c. An indication of the trend to decrease in SMRs that may reflect the activity of reductive AKR1C3 (primarily converting 17-oxo androstanes and estrogens to their 17 β-hydroxy counterparts) to oxidative HSD17B2 activity acting in the opposite direction for ocrelizumab in the LP and even the significant trend for ofatumumab.
- d. A significant trend to lower values of the SMRs that may reflect the activity of AKR1D1 (ocrelizumab in the LP).
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, R.K., E.K.-H.; M.H.; methodology, M.H., M.Ve., M.Vo.; formal analysis, R.K., M.H., J.Vc., E.K.-H.; investigation, R.K., E.K.-H., M.H., R.A.; L.Ka.; J.Vc.; JVi, L.Ko., T.S., M.Z., I.S., J.L.; resources, R.K., E.K.-H.; data curation, R.K., E.K.-H., M.H., R.A.; L.K. M.Z., I.S., J.L.; writing—original draft preparation, M.H., R.K.; writing—review and editing, R.K., M.H.; visualization, R.K.; supervision, E.K.-H., R.K., M.H.; project administration, R.K., E.K.-H., M.H., J.V.; funding acquisition, R.K., E.K.-H.
Funding
This research was funded by “Czech Research Health Council, grant number NU20-04-00450”.
Institutional Review Board Statement
The study was approved by the Ethics Committee of the General University Hospital, Prague, Czech Republic (Approval number: 74/19 Grant AZV VES 2020 VFN, 20 June 2019), and all procedures involving human subjects were conducted following ethical standards set by national and institutional committees on human experimentation and the Helsinki Declaration.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Acknowledgments
The excellent technical assistance of Mrs. Ivona Králová is gratefully acknowledged.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
11β-OH-3α,5α-THA 11β-Hydroxyandrosterone
11β-OH-A 11β-hydroxyandrostenedione
16α-OH-P 16α-Hydroxyprogesterone
16α-OH-Preg 16α-Hydroxypregnenolone
17-OH-P 17-Hydroxyprogesterone
17-OH-Preg 17-Hydroxypregnenolone
17-OH-20α-DHP 17α-Hydroxy-20α-dihydroprogesterone
5α-DHA 5α-Androstane-3,17-dione
5α-DHT 5α-Dihydrotestosterone
3α,5α,17-PD 17-Hydroxyallopregnanolone
3α,5α,17,20α-PT 5α-Pregnane-3α,17,20α-triol
3α,5α,20α-PD 5α-Pregnane-3α, 20α-diol
3α,5α-THA Androsterone
3α,5α-THP 3α,5α-Tetrahydroprogesterone, allopregnanolone
3β,5α,17,20α-PT 5α-Pregnane-3β,17,20α-triol
3β,5α-THA Epiandrosterone
3α,5β,17,20α-PT 5β-Pregnane-3α,17,20α-triol
3α,5β,17-PD 17-Hydroxypregnanolone
3α,5β-THA Etiocholanolone
3α,5β-THA Epietiocholanolone
8-iso-PGF2α 8-iso-Prostaglandin F2α
σ1Rs Sigma-1 receptors
A Androstenedione
ACTH Adrenocorticotropic hormone
Adiol Androstenediol
AKR1C1 Aldo-keto reductase family 1 member C1
AKR1C2 Aldo-keto reductase family 1 member C2
AKR1C3 Aldo-keto reductase family 1 member C3
AKR1D1 5β-Reductase
AMPARs α-Amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptors
ANOVA Analysis of variance
AT Androstenetriol (5-androstene)
BBB Blood-brain barrier
CNS Central nervous system
CRH Corticotropin-releasing hormone
CYP11B1 11β-Hydroxylase
CYP17A1 C17-hydroxylase-C17,20-lyase
CYP19A1 Aromatase
CYP21A2 21-Hydroxylase
CYP3A4 Cytochrome P450 3A4
CYP3A7 Cytochrome P450 3A7
CYP7B1 7α-hydroxylase
DHEA Dehydroepiandrosterone
DHEAC Dehydroepiandrosterone sulfate
DHP Dihydroprogesterone
EDSS Expanded Disability Status Scale
FSH Follicle-stimulating hormone
GABAARs Type A γ-aminobutyric acid receptors
GR Glucocorticoid receptors
GC-MS/MS Gas chromatography tandem mass spectrometry
HPAA Hypothalamic-pituitary-adrenal axis
HSD3B1 Type 1 3β-Hydroxysteroid dehydrogenase
HSD3B2 Type 2 3β-Hydroxysteroid dehydrogenase
HSD3Bs Type 1 and 2 3β-Hydroxysteroid dehydrogenases
HSD11B1 Type 1 11β-Hydroxysteroid dehydrogenase
HSD11B2 Type 2 11β-Hydroxysteroid dehydrogenase
HSD17B2 Type 2 17β-Hydroxysteroid dehydrogenase
HSD17B6 Type 6 17β-Hydroxysteroid dehydrogenase (3α/β epimerase)
HPT9_R 9-Hole Peg Test for right hand
HPT9_L 9-Hole Peg Test for left hand
IFN-γ Interferon γ
IL-1 Interleukin 1
IL-2 Interleukin 2
IL-4 Interleukin 4
IL-6 Interleukin 6
IL-10 Interleukin 10
IL-12 Interleukin 12
IL-17 Interleukin 17
LLR Logarithm of likelihood ratio
LSD Least significant difference
mRNA Messenger ribonucleic acid
NMDARs N-methyl-D-aspartate receptors
OPLS Orthogonal predictions to latent structure
P Progesterone
PD Pregnanediol
PGF2α Prostaglandin F2α
PMC Phase of menstrual cycle
PPMS Primary progressive multiple sclerosis
Preg Pregnenolone
PregC Pregnenolone sulfate
PT Pregnanetriol
ROS Reactive oxygen species
RRMS Relapsing remitting multiple sclerosis
SRD5A1 Type 1 5α-reductase
SRD5As 5α-Reductases
SULT2A1 Steroid sulfotransferase type 2A1
STS Steroid sulfatase
T Testosterone
T25FWT Timed 25-foot walk
THP Tetrahydroprogesterone
TNF Tumor necrosis factor
TRPC5s Short transient receptor potential channels 5
TRPV1s Vanilloid receptors
TRPM3s Melastatin receptors
The letter C after the abbreviation of a steroid indicates its conjugated form
The ", C" after the molar ratio of a steroid indicates that they are its conjugated forms.
References
- Ysrraelit, M.C.; Correale, J. Impact of sex hormones on immune function and multiple sclerosis development. Immunology 2019, 156, 9–22. [Google Scholar] [CrossRef] [PubMed]
- Sparaco, M.; Bonavita, S. The role of sex hormones in women with multiple sclerosis: From puberty to assisted reproductive techniques. Front Neuroendocrinol 2021, 60, 100889. [Google Scholar] [CrossRef]
- Smith, R.; Studd, J.W. A pilot study of the effect upon multiple sclerosis of the menopause, hormone replacement therapy and the menstrual cycle. J R Soc Med 1992, 85, 612–613. [Google Scholar] [CrossRef] [PubMed]
- Argyriou, A.A.; Makris, N. Multiple sclerosis and reproductive risks in women. Reprod Sci 2008, 15, 755–764. [Google Scholar] [CrossRef] [PubMed]
- Trombetta, A.C.; Meroni, M.; Cutolo, M. Steroids and Autoimmunity. Front Horm Res 2017, 48, 121–132. [Google Scholar] [PubMed]
- Kamin, H.S.; Kertes, D.A. Cortisol and DHEA in development and psychopathology. Horm Behav 2017, 89, 69–85. [Google Scholar] [CrossRef]
- Mikulska, J.; Juszczyk, G.; Gawronska-Grzywacz, M.; Herbet, M. HPA Axis in the Pathomechanism of Depression and Schizophrenia: New Therapeutic Strategies Based on Its Participation. Brain Sci 2021, 11. [Google Scholar] [CrossRef]
- Cherian, K.; Schatzberg, A.F.; Keller, J. HPA axis in psychotic major depression and schizophrenia spectrum disorders: Cortisol, clinical symptomatology, and cognition. Schizophr Res 2019, 213, 72–79. [Google Scholar] [CrossRef]
- Misiak, B.; Piotrowski, P.; Chec, M.; Samochowiec, J. Cortisol and dehydroepiandrosterone sulfate in patients with schizophrenia spectrum disorders with respect to cognitive performance. Compr Psychoneuroendocrinol 2021, 6, 100041. [Google Scholar] [CrossRef] [PubMed]
- Ritsner, M.; Maayan, R.; Gibel, A.; Strous, R.D.; Modai, I.; Weizman, A. Elevation of the cortisol/dehydroepiandrosterone ratio in schizophrenia patients. Eur Neuropsychopharmacol 2004, 14, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Ritsner, M.; Gibel, A.; Maayan, R.; Ratner, Y.; Ram, E.; Modai, I.; Weizman, A. State and trait related predictors of serum cortisol to DHEA(S) molar ratios and hormone concentrations in schizophrenia patients. Eur Neuropsychopharmacol 2007, 17, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Kancheva, R.; Hill, M.; Novak, Z.; Chrastina, J.; Kancheva, L.; Starka, L. Neuroactive steroids in periphery and cerebrospinal fluid. Neuroscience 2011, 191, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Begemann, M.J.; Dekker, C.F.; van Lunenburg, M.; Sommer, I.E. Estrogen augmentation in schizophrenia: a quantitative review of current evidence. Schizophr Res 2012, 141, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Qaiser, M.Z.; Dolman, D.E.M.; Begley, D.J.; Abbott, N.J.; Cazacu-Davidescu, M.; Corol, D.I.; Fry, J.P. Uptake and metabolism of sulphated steroids by the blood-brain barrier in the adult male rat. J Neurochem 2017, 142, 672–685. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.; Cao, T.; Zhou, X.; Yao, J.K. Neurosteroids in Schizophrenia: Pathogenic and Therapeutic Implications. Front Psychiatry 2018, 9, 73. [Google Scholar] [CrossRef] [PubMed]
- Powrie, Y.S.L.; Smith, C. Central intracrine DHEA synthesis in ageing-related neuroinflammation and neurodegeneration: therapeutic potential? J Neuroinflammation 2018, 15, 289. [Google Scholar] [CrossRef] [PubMed]
- Honcu, P.; Hill, M.; Bicikova, M.; Jandova, D.; Velikova, M.; Kajzar, J.; Kolatorova, L.; Bestak, J.; Macova, L.; Kancheva, R. , et al. Activation of Adrenal Steroidogenesis and an Improvement of Mood Balance in Postmenopausal Females after Spa Treatment Based on Physical Activity. Int J Mol Sci 2019, 20. [Google Scholar] [CrossRef] [PubMed]
- MacKenzie, E.M.; Odontiadis, J.; Le Melledo, J.M.; Prior, T.I.; Baker, G.B. The relevance of neuroactive steroids in schizophrenia, depression, and anxiety disorders. Cell Mol Neurobiol 2007, 27, 541–574. [Google Scholar] [CrossRef] [PubMed]
- Luu-The, V. Assessment of steroidogenesis and steroidogenic enzyme functions. J Steroid Biochem Mol Biol 2013, 137, 176–182. [Google Scholar] [CrossRef]
- Labrie, F. All sex steroids are made intracellularly in peripheral tissues by the mechanisms of intracrinology after menopause. J Steroid Biochem Mol Biol 2015, 145, 133–138. [Google Scholar] [CrossRef]
- Noorbakhsh, F.; Ellestad, K.K.; Maingat, F.; Warren, K.G.; Han, M.H.; Steinman, L.; Baker, G.B.; Power, C. Impaired neurosteroid synthesis in multiple sclerosis. Brain 2011, 134, 2703–2721. [Google Scholar] [CrossRef] [PubMed]
- Noorbakhsh, F.; Baker, G.B.; Power, C. Allopregnanolone and neuroinflammation: a focus on multiple sclerosis. Front Cell Neurosci 2014, 8, 134. [Google Scholar] [CrossRef] [PubMed]
- Tomassini, V.; Pozzilli, C. Sex hormones, brain damage and clinical course of Multiple Sclerosis. J Neurol Sci 2009, 286, 35–39. [Google Scholar] [CrossRef]
- Cil, A.P.; Leventoglu, A.; Sonmezer, M.; Soylukoc, R.; Oktay, K. Assessment of ovarian reserve and Doppler characteristics in patients with multiple sclerosis using immunomodulating drugs. J Turk Ger Gynecol Assoc 2009, 10, 213–219. [Google Scholar]
- Griffiths, W.J.; Wang, Y. Sterols, Oxysterols, and Accessible Cholesterol: Signalling for Homeostasis, in Immunity and During Development. Front Physiol 2021, 12, 723224. [Google Scholar] [CrossRef] [PubMed]
- Sukocheva, O.; Wadham, C.; Gamble, J.; Xia, P. Sphingosine-1-phosphate receptor 1 transmits estrogens' effects in endothelial cells. Steroids 2015, 104, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Lucki, N.C.; Li, D.; Sewer, M.B. Sphingosine-1-phosphate rapidly increases cortisol biosynthesis and the expression of genes involved in cholesterol uptake and transport in H295R adrenocortical cells. Mol Cell Endocrinol 2012, 348, 165–175. [Google Scholar] [CrossRef]
- Guzman, A.; Rosales-Torres, A.M.; Medina-Moctezuma, Z.B.; Gonzalez-Aretia, D.; Hernandez-Coronado, C.G. Effects and action mechanism of gonadotropins on ovarian follicular cells: A novel role of Sphingosine-1-Phosphate (S1P). A review. Gen Comp Endocrinol 2024, 357, 114593. [Google Scholar] [CrossRef] [PubMed]
- Siavoshi, F.; Ladakis, D.C.; Muller, A.; Nourbakhsh, B.; Bhargava, P. Ocrelizumab alters the circulating metabolome in people with relapsing-remitting multiple sclerosis. Ann Clin Transl Neurol 2024, 11, 2485–2498. [Google Scholar] [CrossRef] [PubMed]
- Ito, K.; Ito, N.; Yadav, S.K.; Suresh, S.; Lin, Y.; Dhib-Jalbut, S. Effect of switching glatiramer acetate formulation from 20 mg daily to 40 mg three times weekly on immune function in multiple sclerosis. Mult Scler J Exp Transl Clin 2021, 7, 20552173211032323. [Google Scholar] [CrossRef] [PubMed]
- Dargahi, N.; Katsara, M.; Tselios, T.; Androutsou, M.E.; de Courten, M.; Matsoukas, J.; Apostolopoulos, V. Multiple Sclerosis: Immunopathology and Treatment Update. Brain Sci 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Murdoch, D.; Lyseng-Williamson, K.A. Spotlight on subcutaneous recombinant interferon-beta-1a (Rebif) in relapsing-remitting multiple sclerosis. BioDrugs 2005, 19, 323–325. [Google Scholar] [CrossRef] [PubMed]
- Giovannoni, G.; Munschauer, F.E., 3rd; Deisenhammer, F. Neutralising antibodies to interferon beta during the treatment of multiple sclerosis. J Neurol Neurosurg Psychiatry 2002, 73, 465–469. [Google Scholar] [CrossRef] [PubMed]
- Hauser, S.L.; Kappos, L.; Bar-Or, A.; Wiendl, H.; Paling, D.; Williams, M.; Gold, R.; Chan, A.; Milo, R.; Das Gupta, A. , et al. The Development of Ofatumumab, a Fully Human Anti-CD20 Monoclonal Antibody for Practical Use in Relapsing Multiple Sclerosis Treatment. Neurol Ther 2023, 12, 1491–1515. [Google Scholar] [CrossRef] [PubMed]
- McGinley, M.P.; Moss, B.P.; Cohen, J.A. Safety of monoclonal antibodies for the treatment of multiple sclerosis. Expert Opin Drug Saf 2017, 16, 89–100. [Google Scholar] [CrossRef]
- Lamb, Y.N. Ocrelizumab: A Review in Multiple Sclerosis. Drugs 2022, 82, 323–334. [Google Scholar] [CrossRef]
- Kancheva, R.; Hill, M.; Velikova, M.; Kancheva, L.; Vcelak, J.; Ampapa, R.; Zido, M.; Stetkarova, I.; Libertinova, J.; Vosatkova, M. , et al. Altered Steroidome in Women with Multiple Sclerosis. Int J Mol Sci 2024, 25. [Google Scholar] [CrossRef] [PubMed]
- Labrie, F.; Martel, C.; Belanger, A.; Pelletier, G. Androgens in women are essentially made from DHEA in each peripheral tissue according to intracrinology. J Steroid Biochem Mol Biol 2017, 168, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Angeli, A.; Masera, R.G.; Sartori, M.L.; Fortunati, N.; Racca, S.; Dovio, A.; Staurenghi, A.; Frairia, R. Modulation by cytokines of glucocorticoid action. Ann N Y Acad Sci 1999, 876, 210–220. [Google Scholar] [CrossRef] [PubMed]
- de Kloet, E.R.; Joels, M.; Holsboer, F. Stress and the brain: from adaptation to disease. Nat Rev Neurosci 2005, 6, 463–475. [Google Scholar] [CrossRef] [PubMed]
- Hildebrandt, H.; Stachowiak, R.; Heber, I.; Schlake, H.P.; Eling, P. Relation between cognitive fatigue and circadian or stress related cortisol levels in MS patients. Mult Scler Relat Disord 2020, 45, 102440. [Google Scholar] [CrossRef] [PubMed]
- Slominski, R.M.; Tuckey, R.C.; Manna, P.R.; Jetten, A.M.; Postlethwaite, A.; Raman, C.; Slominski, A.T. Extra-adrenal glucocorticoid biosynthesis: implications for autoimmune and inflammatory disorders. Genes Immun 2020, 21, 150–168. [Google Scholar] [CrossRef] [PubMed]
- Tucha, L.; Fuermaier, A.B.; Koerts, J.; Buggenthin, R.; Aschenbrenner, S.; Weisbrod, M.; Thome, J.; Lange, K.W.; Tucha, O. Sustained attention in adult ADHD: time-on-task effects of various measures of attention. J Neural Transm (Vienna) 2017, 124, 39–53. [Google Scholar] [CrossRef]
- Vedhara, K.; Hyde, J.; Gilchrist, I.D.; Tytherleigh, M.; Plummer, S. Acute stress, memory, attention and cortisol. Psychoneuroendocrinology 2000, 25, 535–549. [Google Scholar] [CrossRef] [PubMed]
- Pereira, G.M.; Soares, N.M.; Souza, A.R.; Becker, J.; Finkelsztejn, A.; Almeida, R.M.M. Basal cortisol levels and the relationship with clinical symptoms in multiple sclerosis: a systematic review. Arq Neuropsiquiatr 2018, 76, 622–634. [Google Scholar] [CrossRef]
- Heidbrink, C.; Hausler, S.F.; Buttmann, M.; Ossadnik, M.; Strik, H.M.; Keller, A.; Buck, D.; Verbraak, E.; van Meurs, M.; Krockenberger, M. , et al. Reduced cortisol levels in cerebrospinal fluid and differential distribution of 11beta-hydroxysteroid dehydrogenases in multiple sclerosis: implications for lesion pathogenesis. Brain Behav Immun 2010, 24, 975–984. [Google Scholar] [CrossRef]
- Foroughipour, A.; Norbakhsh, V.; Najafabadi, S.H.; Meamar, R. Evaluating sex hormone levels in reproductive age women with multiple sclerosis and their relationship with disease severity. J Res Med Sci 2012, 17, 882–885. [Google Scholar] [PubMed]
- Wei, T.; Lightman, S.L. The neuroendocrine axis in patients with multiple sclerosis. Brain 1997, 120 ( Pt 6) Pt 6, 1067–1076. [Google Scholar] [CrossRef]
- Hamidovic, A.; Karapetyan, K.; Serdarevic, F.; Choi, S.H.; Eisenlohr-Moul, T.; Pinna, G. Higher Circulating Cortisol in the Follicular vs. Luteal Phase of the Menstrual Cycle: A Meta-Analysis. Front Endocrinol (Lausanne) 2020, 11, 311. [Google Scholar] [CrossRef] [PubMed]
- Marx, C.E.; Keefe, R.S.; Buchanan, R.W.; Hamer, R.M.; Kilts, J.D.; Bradford, D.W.; Strauss, J.L.; Naylor, J.C.; Payne, V.M.; Lieberman, J.A. , et al. Proof-of-concept trial with the neurosteroid pregnenolone targeting cognitive and negative symptoms in schizophrenia. Neuropsychopharmacology 2009, 34, 1885–1903. [Google Scholar] [CrossRef] [PubMed]
- Matuszewska, A.; Kowalski, K.; Jawien, P.; Tomkalski, T.; Gawel-Dabrowska, D.; Merwid-Lad, A.; Szelag, E.; Blaszczak, K.; Wiatrak, B.; Danielewski, M. , et al. The Hypothalamic-Pituitary-Gonadal Axis in Men with Schizophrenia. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef] [PubMed]
- Gubbels Bupp, M.R.; Jorgensen, T.N. Androgen-Induced Immunosuppression. Front Immunol 2018, 9, 794. [Google Scholar] [CrossRef]
- Tomassini, V.; Onesti, E.; Mainero, C.; Giugni, E.; Paolillo, A.; Salvetti, M.; Nicoletti, F.; Pozzilli, C. Sex hormones modulate brain damage in multiple sclerosis: MRI evidence. J Neurol Neurosurg Psychiatry 2005, 76, 272–275. [Google Scholar] [CrossRef]
- Garcia-Estrada, J.; Del Rio, J.A.; Luquin, S.; Soriano, E.; Garcia-Segura, L.M. Gonadal hormones down-regulate reactive gliosis and astrocyte proliferation after a penetrating brain injury. Brain Res 1993, 628, 271–278. [Google Scholar] [CrossRef]
- Bovolenta, P.; Wandosell, F.; Nieto-Sampedro, M. CNS glial scar tissue: a source of molecules which inhibit central neurite outgrowth. Prog Brain Res 1992, 94, 367–379. [Google Scholar] [PubMed]
- Burda, J.E.; Sofroniew, M.V. Reactive gliosis and the multicellular response to CNS damage and disease. Neuron 2014, 81, 229–248. [Google Scholar] [CrossRef]
- Dalal, M.; Kim, S.; Voskuhl, R.R. Testosterone therapy ameliorates experimental autoimmune encephalomyelitis and induces a T helper 2 bias in the autoantigen-specific T lymphocyte response. J Immunol 1997, 159, 3–6. [Google Scholar] [CrossRef] [PubMed]
- Caruso, A.; Di Giorgi Gerevini, V.; Castiglione, M.; Marinelli, F.; Tomassini, V.; Pozzilli, C.; Caricasole, A.; Bruno, V.; Caciagli, F.; Moretti, A. , et al. Testosterone amplifies excitotoxic damage of cultured oligodendrocytes. J Neurochem 2004, 88, 1179–1185. [Google Scholar] [CrossRef]
- Ghoumari, A.M.; Ibanez, C.; El-Etr, M.; Leclerc, P.; Eychenne, B.; O'Malley, B.W.; Baulieu, E.E.; Schumacher, M. Progesterone and its metabolites increase myelin basic protein expression in organotypic slice cultures of rat cerebellum. J Neurochem 2003, 86, 848–859. [Google Scholar] [CrossRef] [PubMed]
- Bodhankar, S.; Wang, C.; Vandenbark, A.A.; Offner, H. Estrogen-induced protection against experimental autoimmune encephalomyelitis is abrogated in the absence of B cells. Eur J Immunol 2011, 41, 1165–1175. [Google Scholar] [CrossRef] [PubMed]
- Gupta, M. K.; Guryev, O. L.; Auchus, R. J. , 5alpha-reduced C21 steroids are substrates for human cytochrome P450c17. Arch Biochem Biophys 2003, 418, (2), 151–60. [Google Scholar] [CrossRef]
- Rege, J.; Nakamura, Y.; Wang, T.; Merchen, T.D.; Sasano, H.; Rainey, W.E. Transcriptome profiling reveals differentially expressed transcripts between the human adrenal zona fasciculata and zona reticularis. J Clin Endocrinol Metab 2014, 99, E518–527. [Google Scholar] [CrossRef] [PubMed]
- Park-Chung, M.; Wu, F.S.; Purdy, R.H.; Malayev, A.A.; Gibbs, T.T.; Farb, D.H. Distinct sites for inverse modulation of N-methyl-D-aspartate receptors by sulfated steroids. Mol Pharmacol 1997, 52, 1113–1123. [Google Scholar] [CrossRef]
- Park-Chung, M.; Malayev, A.; Purdy, R.H.; Gibbs, T.T.; Farb, D.H. Sulfated and unsulfated steroids modulate gamma-aminobutyric acidA receptor function through distinct sites. Brain Res 1999, 830, 72–87. [Google Scholar] [CrossRef]
- Smejkalova, T.; Korinek, M.; Krusek, J.; Hrcka Krausova, B.; Candelas Serra, M.; Hajdukovic, D.; Kudova, E.; Chodounska, H.; Vyklicky, L. Endogenous neurosteroids pregnanolone and pregnanolone sulfate potentiate presynaptic glutamate release through distinct mechanisms. Br J Pharmacol 2021, 178, 3888–3904. [Google Scholar] [CrossRef] [PubMed]
- Labrie, F. Adrenal androgens and intracrinology. Semin Reprod Med 2004, 22, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Majewska, M.D. Steroid regulation of the GABAA receptor: ligand binding, chloride transport and behaviour. Ciba Found Symp 1990, 153, 83–97. [Google Scholar] [PubMed]
- Petrovic, M.; Sedlacek, M.; Horak, M.; Chodounska, H.; Vyklicky, L., Jr. 20-oxo-5beta-pregnan-3alpha-yl sulfate is a use-dependent NMDA receptor inhibitor. J Neurosci 2005, 25, 8439–8450. [Google Scholar] [CrossRef] [PubMed]
- Johansson, T.; Le Greves, P. The effect of dehydroepiandrosterone sulfate and allopregnanolone sulfate on the binding of [(3)H]ifenprodil to the N-methyl-d-aspartate receptor in rat frontal cortex membrane. J Steroid Biochem Mol Biol 2005, 94, 263–266. [Google Scholar] [CrossRef]
- Barnard, L.; Gent, R.; van Rooyen, D.; Swart, A.C. Adrenal C11-oxy C(21) steroids contribute to the C11-oxy C(19) steroid pool via the backdoor pathway in the biosynthesis and metabolism of 21-deoxycortisol and 21-deoxycortisone. J Steroid Biochem Mol Biol 2017, 174, 86–95. [Google Scholar] [CrossRef]
- do Nascimento, F.V.; Piccoli, V.; Beer, M.A.; von Frankenberg, A.D.; Crispim, D.; Gerchman, F. Association of HSD11B1 polymorphic variants and adipose tissue gene expression with metabolic syndrome, obesity and type 2 diabetes mellitus: a systematic review. Diabetol Metab Syndr 2015, 7, 38. [Google Scholar] [CrossRef] [PubMed]
- Bottasso, O.; Bay, M.L.; Besedovsky, H.; del Rey, A. The immuno-endocrine component in the pathogenesis of tuberculosis. Scand J Immunol 2007, 66, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Du, C.; Khalil, M.W.; Sriram, S. Administration of dehydroepiandrosterone suppresses experimental allergic encephalomyelitis in SJL/J mice. J Immunol 2001, 167, 7094–7101. [Google Scholar] [CrossRef]
- Rontzsch, A.; Thoss, K.; Petrow, P.K.; Henzgen, S.; Brauer, R. Amelioration of murine antigen-induced arthritis by dehydroepiandrosterone (DHEA). Inflamm Res 2004, 53, 189–198. [Google Scholar] [PubMed]
- Tan, X.D.; Dou, Y.C.; Shi, C.W.; Duan, R.S.; Sun, R.P. Administration of dehydroepiandrosterone ameliorates experimental autoimmune neuritis in Lewis rats. J Neuroimmunol 2009, 207, 39–44. [Google Scholar] [CrossRef]
- Choi, I.S.; Cui, Y.; Koh, Y.A.; Lee, H.C.; Cho, Y.B.; Won, Y.H. Effects of dehydroepiandrosterone on Th2 cytokine production in peripheral blood mononuclear cells from asthmatics. Korean J Intern Med 2008, 23, 176–181. [Google Scholar] [CrossRef] [PubMed]
- Sudo, N.; Yu, X.N.; Kubo, C. Dehydroepiandrosterone attenuates the spontaneous elevation of serum IgE level in NC/Nga mice. Immunol Lett 2001, 79, 177–179. [Google Scholar] [CrossRef] [PubMed]
- Kasperska-Zajac, A.; Brzoza, Z.; Rogala, B. Dehydroepiandrosterone and dehydroepiandrosterone sulphate in atopic allergy and chronic urticaria. Inflammation 2008, 31, 141–145. [Google Scholar] [CrossRef] [PubMed]
- Romagnani, S.; Kapsenberg, M.; Radbruch, A.; Adorini, L. Th1 and Th2 cells. Res Immunol 1998, 149, 871–873. [Google Scholar] [CrossRef]
- Pratschke, S.; von Dossow-Hanfstingl, V.; Dietz, J.; Schneider, C.P.; Tufman, A.; Albertsmeier, M.; Winter, H.; Angele, M.K. Dehydroepiandrosterone modulates T-cell response after major abdominal surgery. J Surg Res 2014, 189, 117–125. [Google Scholar] [CrossRef]
- Sterzl, I.; Hampl, R.; Sterzl, J.; Votruba, J.; Starka, L. 7Beta-OH-DHEA counteracts dexamethasone induced suppression of primary immune response in murine spleenocytes. J Steroid Biochem Mol Biol 1999, 71, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Hennebert, O.; Chalbot, S.; Alran, S.; Morfin, R. Dehydroepiandrosterone 7alpha-hydroxylation in human tissues: possible interference with type 1 11beta-hydroxysteroid dehydrogenase-mediated processes. J Steroid Biochem Mol Biol 2007, 104, 326–333. [Google Scholar] [CrossRef] [PubMed]
- Le Mee, S.; Hennebert, O.; Ferrec, C.; Wulfert, E.; Morfin, R. 7beta-Hydroxy-epiandrosterone-mediated regulation of the prostaglandin synthesis pathway in human peripheral blood monocytes. Steroids 2008, 73, 1148–1159. [Google Scholar] [CrossRef] [PubMed]
- Pettersson, H.; Lundqvist, J.; Norlin, M. Effects of CYP7B1-mediated catalysis on estrogen receptor activation. Biochim Biophys Acta 2010, 1801, 1090–1097. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.; Eggertsen, G.; Chiang, J.Y.; Norlin, M. Estrogen-mediated regulation of CYP7B1: a possible role for controlling DHEA levels in human tissues. J Steroid Biochem Mol Biol 2006, 100, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Ahlem, C.N.; Page, T.M.; Auci, D.L.; Kennedy, M.R.; Mangano, K.; Nicoletti, F.; Ge, Y.; Huang, Y.; White, S.K.; Villegas, S. , et al. Novel components of the human metabolome: the identification, characterization and anti-inflammatory activity of two 5-androstene tetrols. Steroids 2011, 76, 145–155. [Google Scholar] [CrossRef]
- Reading, C.L.; Frincke, J.M.; White, S.K. Molecular targets for 17alpha-ethynyl-5-androstene-3beta,7beta,17beta-triol, an anti-inflammatory agent derived from the human metabolome. PLoS One 2012, 7, e32147. [Google Scholar] [CrossRef]
- Reddy, D.S. Neurosteroids: endogenous role in the human brain and therapeutic potentials. Prog Brain Res 2010, 186, 113–137. [Google Scholar] [PubMed]
- Balan, I.; Beattie, M.C.; O'Buckley, T.K.; Aurelian, L.; Morrow, A.L. Endogenous Neurosteroid (3alpha,5alpha)3-Hydroxypregnan-20-one Inhibits Toll-like-4 Receptor Activation and Pro-inflammatory Signaling in Macrophages and Brain. Sci Rep 2019, 9, 1220. [Google Scholar] [CrossRef] [PubMed]
- Lapchak, P.A. The neuroactive steroid 3-alpha-ol-5-beta-pregnan-20-one hemisuccinate, a selective NMDA receptor antagonist improves behavioral performance following spinal cord ischemia. Brain Res 2004, 997, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Kudova, E.; Mares, P.; Hill, M.; Vondrakova, K.; Tsenov, G.; Chodounska, H.; Kubova, H.; Vales, K. The Neuroactive Steroid Pregnanolone Glutamate: Anticonvulsant Effect, Metabolites and Its Effect on Neurosteroid Levels in Developing Rat Brains. Pharmaceuticals (Basel) 2021, 15. [Google Scholar] [CrossRef]
- Abramova, V.; Leal Alvarado, V.; Hill, M.; Smejkalova, T.; Maly, M.; Vales, K.; Dittert, I.; Bozikova, P.; Kysilov, B.; Hrcka Krausova, B. , et al. Effects of Pregnanolone Glutamate and Its Metabolites on GABA(A) and NMDA Receptors and Zebrafish Behavior. ACS Chem Neurosci 2023, 14, 1870–1883. [Google Scholar] [CrossRef]
- Munawar Cheema, M.; Macakova Kotrbova, Z.; Hrcka Krausova, B.; Adla, S.K.; Slavikova, B.; Chodounska, H.; Kratochvil, M.; Vondrasek, J.; Sedlak, D.; Balastik, M. , et al. 5beta-reduced neuroactive steroids as modulators of growth and viability of postnatal neurons and glia. J Steroid Biochem Mol Biol 2024, 239, 106464. [Google Scholar] [CrossRef] [PubMed]
- Akwa, Y. Steroids and Alzheimer's Disease: Changes Associated with Pathology and Therapeutic Potential. Int J Mol Sci 2020, 21. [Google Scholar] [CrossRef] [PubMed]
- Hill, M.; Parizek, A.; Simjak, P.; Koucky, M.; Anderlova, K.; Krejci, H.; Vejrazkova, D.; Ondrejikova, L.; Cerny, A.; Kancheva, R. Steroids, steroid associated substances and gestational diabetes mellitus. Physiol Res 2021, 70, S617–S634. [Google Scholar] [CrossRef]
- Burczynski, M.E.; Sridhar, G.R.; Palackal, N.T.; Penning, T.M. The reactive oxygen species--and Michael acceptor-inducible human aldo-keto reductase AKR1C1 reduces the alpha,beta-unsaturated aldehyde 4-hydroxy-2-nonenal to 1,4-dihydroxy-2-nonene. J Biol Chem 2001, 276, 2890–2897. [Google Scholar] [CrossRef]
- Murgia, F.; Giagnoni, F.; Lorefice, L.; Caria, P.; Dettori, T.; D'Alterio, M.N.; Angioni, S.; Hendren, A.J.; Caboni, P.; Pibiri, M. , et al. Sex Hormones as Key Modulators of the Immune Response in Multiple Sclerosis: A Review. Biomedicines 2022, 10. [Google Scholar] [CrossRef]
- Xu, C.; Liu, W.; You, X.; Leimert, K.; Popowycz, K.; Fang, X.; Wood, S.L.; Slater, D.M.; Sun, Q.; Gu, H. , et al. PGF2alpha modulates the output of chemokines and pro-inflammatory cytokines in myometrial cells from term pregnant women through divergent signaling pathways. Mol Hum Reprod 2015, 21, 603–614. [Google Scholar] [CrossRef]
- Zheng, L.; Fei, J.; Feng, C.M.; Xu, Z.; Fu, L.; Zhao, H. Serum 8-iso-PGF2alpha Predicts the Severity and Prognosis in Patients With Community-Acquired Pneumonia: A Retrospective Cohort Study. Front Med (Lausanne) 2021, 8, 633442. [Google Scholar] [CrossRef] [PubMed]
- Sharma, I.; Dhaliwal, L.K.; Saha, S.C.; Sangwan, S.; Dhawan, V. Role of 8-iso-prostaglandin F2alpha and 25-hydroxycholesterol in the pathophysiology of endometriosis. Fertil Steril 2010, 94, 63–70. [Google Scholar] [CrossRef]
- Tchernof, A.; Mansour, M.F.; Pelletier, M.; Boulet, M.M.; Nadeau, M.; Luu-The, V. Updated survey of the steroid-converting enzymes in human adipose tissues. J Steroid Biochem Mol Biol 2015, 147, 56–69. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, Y.; Hornsby, P.J.; Casson, P.; Morimoto, R.; Satoh, F.; Xing, Y.; Kennedy, M.R.; Sasano, H.; Rainey, W.E. Type 5 17beta-hydroxysteroid dehydrogenase (AKR1C3) contributes to testosterone production in the adrenal reticularis. J Clin Endocrinol Metab 2009, 94, 2192–2198. [Google Scholar] [CrossRef]
- Ostinelli, G.; Vijay, J.; Vohl, M.C.; Grundberg, E.; Tchernof, A. AKR1C2 and AKR1C3 expression in adipose tissue: Association with body fat distribution and regulatory variants. Mol Cell Endocrinol 2021, 527, 111220. [Google Scholar] [CrossRef] [PubMed]
- Rizner, T.L.; Penning, T.M. Role of aldo-keto reductase family 1 (AKR1) enzymes in human steroid metabolism. Steroids 2014, 79, 49–63. [Google Scholar] [CrossRef] [PubMed]
- Miller, W.L.; Auchus, R.J. The molecular biology, biochemistry, and physiology of human steroidogenesis and its disorders. Endocr Rev 2011, 32, 81–151. [Google Scholar] [CrossRef] [PubMed]
- Thompson, A.J.; Banwell, B.L.; Barkhof, F.; Carroll, W.M.; Coetzee, T.; Comi, G.; Correale, J.; Fazekas, F.; Filippi, M.; Freedman, M.S. , et al. Diagnosis of multiple sclerosis: 2017 revisions of the McDonald criteria. Lancet Neurol 2018, 17, 162–173. [Google Scholar] [CrossRef] [PubMed]
- Hill, M.; Hana jr., V.; Velikova, M.; Parizek, A.; Kolatorova, L.; Vitku, J.; Skodova, T.; Simkova, M.; Simjak, P.; Kancheva, R. , et al. A method for determination of one hundred endogenous steroids in human serum by gas chromatography-tandem mass spectrometry. Physiol Res 2019, 68, 179–207. [Google Scholar] [CrossRef]
Figure 1.
Changes of cortisol (measured by RIA) and cortisone levels (measured by GC-MS/MS) in patients with MS after treatment with anti-MS drugs. White, black and grey circles with error bars represent the retransformed means with 95% confidence intervals for follicular menstrual phase, luteal menstrual phase and changes in main factor Stage of ANOVA model consisting of subject factor (Subj), effects of treatment (Stage) and menstrual phase (PMC) and Stage × PMC interaction. Panels A, B, and C show the effects of Stage, PMC, and Stage × PMC interactions for cortisol, panels D, E, and F for cortisone, and panels G, H, and I for corticosterone. The symbols F, p, and ηp2 represent the F-statistics, p-values and effect size for factors and interaction. The statistics for factors and interaction are as follows: Stage: F=6.6, p=0.013, ηp2=0.11; PMC: F=1.9, p=0.171, ηp2=0.0351; Stage × PMC: F=0, p=0.925, ηp2=0.000167; Subj: F=2.1, p=0.004, ηp2=0.7 for cortisol (measured by RIA), Stage: F=4, p=0.051, ηp2=0.0702; PMC: F=3.3, p=0.074, ηp2=0.059; Stage × PMC: F=0.9, p=0.351, ηp2=0.0164; Subj: F=1.5, p=0.07, ηp2=0.621 for cortisone and Stage: F=2.5, p=0.12, ηp2=0.0442; PMC: F=2.2, p=0.144, ηp2=0.0392; Stage × PMC: F=2, p=0.16, ηp2=0.0363; Subj: F=2.5, p<0.001, ηp2=0.725. Non-overlapping error bars indicate a significant difference (p<0.05, least significant multiple comparisons).
Figure 1.
Changes of cortisol (measured by RIA) and cortisone levels (measured by GC-MS/MS) in patients with MS after treatment with anti-MS drugs. White, black and grey circles with error bars represent the retransformed means with 95% confidence intervals for follicular menstrual phase, luteal menstrual phase and changes in main factor Stage of ANOVA model consisting of subject factor (Subj), effects of treatment (Stage) and menstrual phase (PMC) and Stage × PMC interaction. Panels A, B, and C show the effects of Stage, PMC, and Stage × PMC interactions for cortisol, panels D, E, and F for cortisone, and panels G, H, and I for corticosterone. The symbols F, p, and ηp2 represent the F-statistics, p-values and effect size for factors and interaction. The statistics for factors and interaction are as follows: Stage: F=6.6, p=0.013, ηp2=0.11; PMC: F=1.9, p=0.171, ηp2=0.0351; Stage × PMC: F=0, p=0.925, ηp2=0.000167; Subj: F=2.1, p=0.004, ηp2=0.7 for cortisol (measured by RIA), Stage: F=4, p=0.051, ηp2=0.0702; PMC: F=3.3, p=0.074, ηp2=0.059; Stage × PMC: F=0.9, p=0.351, ηp2=0.0164; Subj: F=1.5, p=0.07, ηp2=0.621 for cortisone and Stage: F=2.5, p=0.12, ηp2=0.0442; PMC: F=2.2, p=0.144, ηp2=0.0392; Stage × PMC: F=2, p=0.16, ηp2=0.0363; Subj: F=2.5, p<0.001, ηp2=0.725. Non-overlapping error bars indicate a significant difference (p<0.05, least significant multiple comparisons).
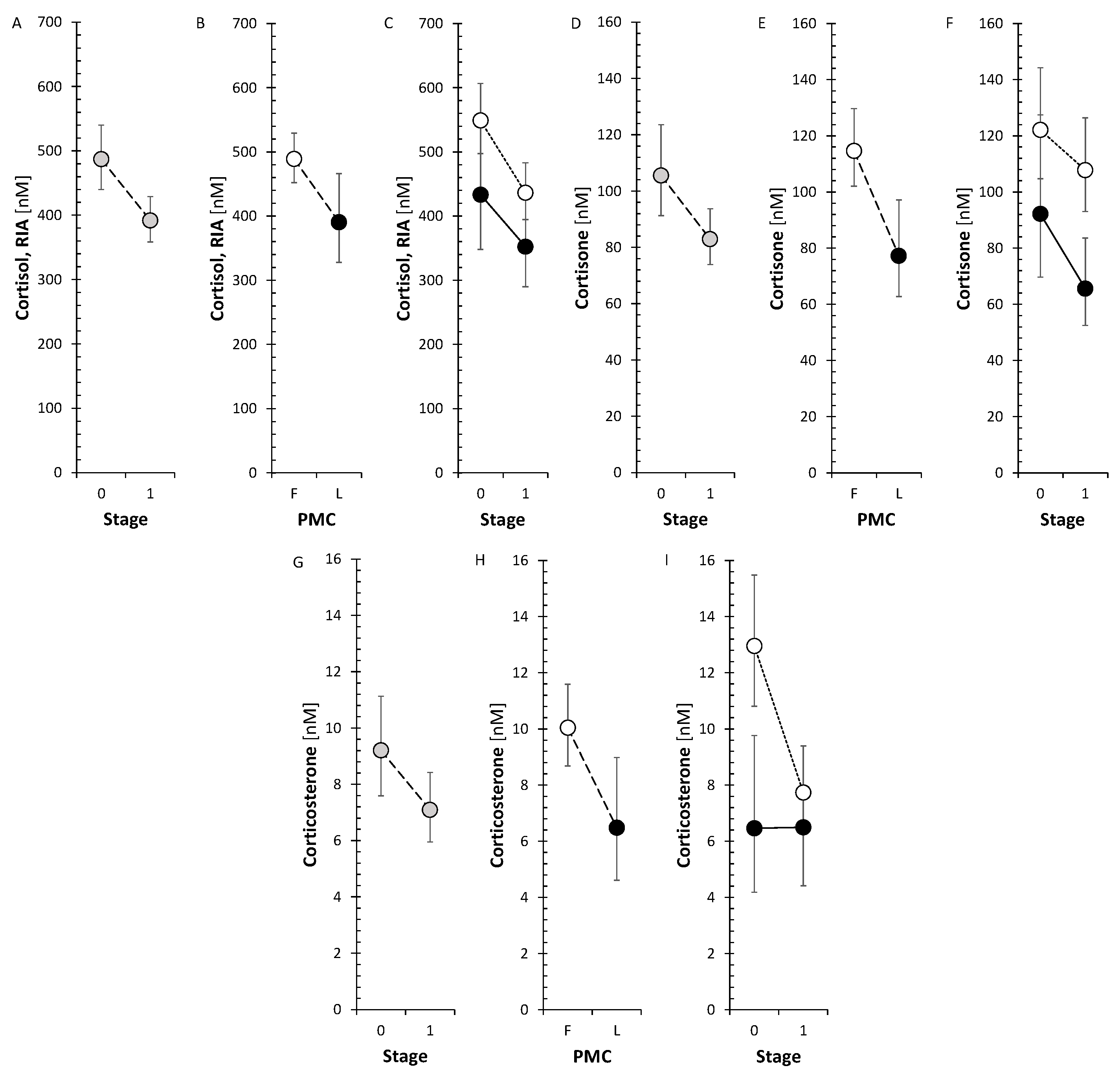
Figure 2.
Changes of pregnenolone and DHEA levels in patients with MS after treatment with anti-MS drugs. The drawings and symbols are the same as for Figure 1. The statistics for factors and interaction are as follows: Stage: F=5.1, p=0.028, ηp2=0.0848; PMC: F=0.8, p=0.376, ηp2=0.0143; Stage × PMC: F=0.4, p=0.543, ηp2=0.00675; Subj: F=2.3, p=0.001, ηp2=0.709 for pregnenolone and Stage: F=7.5, p=0.009, ηp2=0.119; PMC: F=0.2, p=0.656, ηp2=0.00364; Stage × PMC: F=0.8, p=0.374, ηp2=0.0144; Subj: F=2.8, p<0.001, ηp2=0.751 for DHEA.
Figure 2.
Changes of pregnenolone and DHEA levels in patients with MS after treatment with anti-MS drugs. The drawings and symbols are the same as for Figure 1. The statistics for factors and interaction are as follows: Stage: F=5.1, p=0.028, ηp2=0.0848; PMC: F=0.8, p=0.376, ηp2=0.0143; Stage × PMC: F=0.4, p=0.543, ηp2=0.00675; Subj: F=2.3, p=0.001, ηp2=0.709 for pregnenolone and Stage: F=7.5, p=0.009, ηp2=0.119; PMC: F=0.2, p=0.656, ηp2=0.00364; Stage × PMC: F=0.8, p=0.374, ηp2=0.0144; Subj: F=2.8, p<0.001, ηp2=0.751 for DHEA.
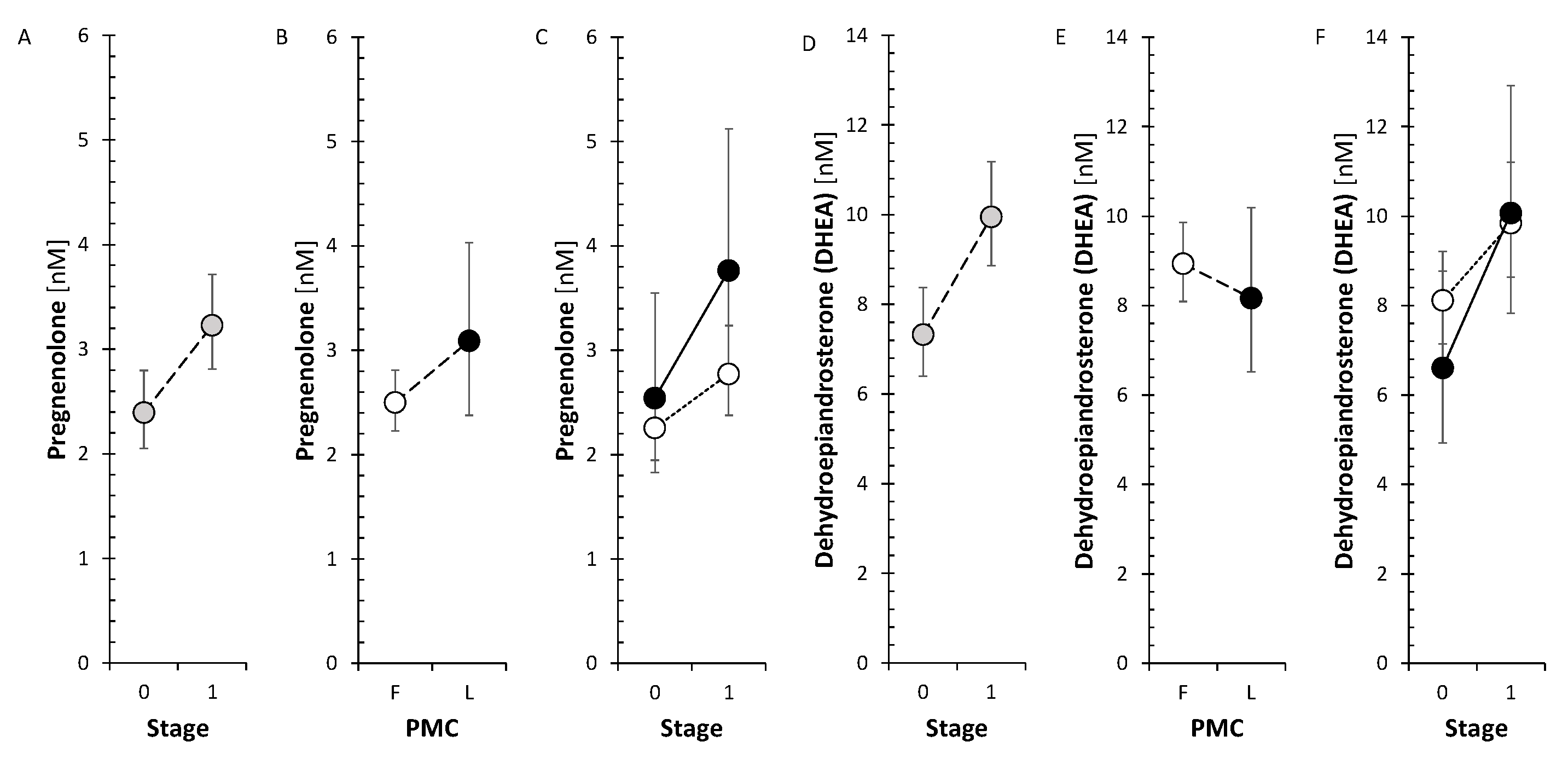
Figure 3.
Changes of 7α-hydroxy-DHEA and 5-androstene-3β,7β,17β-triol levels in patients with MS after treatment with anti-MS drugs. The drawings and symbols are the same as for Figure 1. The statistics for factors and interaction are as follows: Stage: F=3.3, p=0.074, ηp2=0.0578; PMC: F=1.4, p=0.249, ηp2=0.0245; Stage × PMC: F=0.3, p=0.602, ηp2=0.00506; Subj: F=4, p<0.001, ηp2=0.809 for 7α-hydroxy-DHEA and Stage: F=0.8, p=0.377, ηp2=0.0148; PMC: F=0.5, p=0.468, ηp2=0.00998; Stage × PMC: F=4.2, p=0.045, ηp2=0.0736; Subj: F=3.6, p<0.001, ηp2=0.794 for 5-androstene-3β,7β,17β-triol.
Figure 3.
Changes of 7α-hydroxy-DHEA and 5-androstene-3β,7β,17β-triol levels in patients with MS after treatment with anti-MS drugs. The drawings and symbols are the same as for Figure 1. The statistics for factors and interaction are as follows: Stage: F=3.3, p=0.074, ηp2=0.0578; PMC: F=1.4, p=0.249, ηp2=0.0245; Stage × PMC: F=0.3, p=0.602, ηp2=0.00506; Subj: F=4, p<0.001, ηp2=0.809 for 7α-hydroxy-DHEA and Stage: F=0.8, p=0.377, ηp2=0.0148; PMC: F=0.5, p=0.468, ηp2=0.00998; Stage × PMC: F=4.2, p=0.045, ηp2=0.0736; Subj: F=3.6, p<0.001, ηp2=0.794 for 5-androstene-3β,7β,17β-triol.
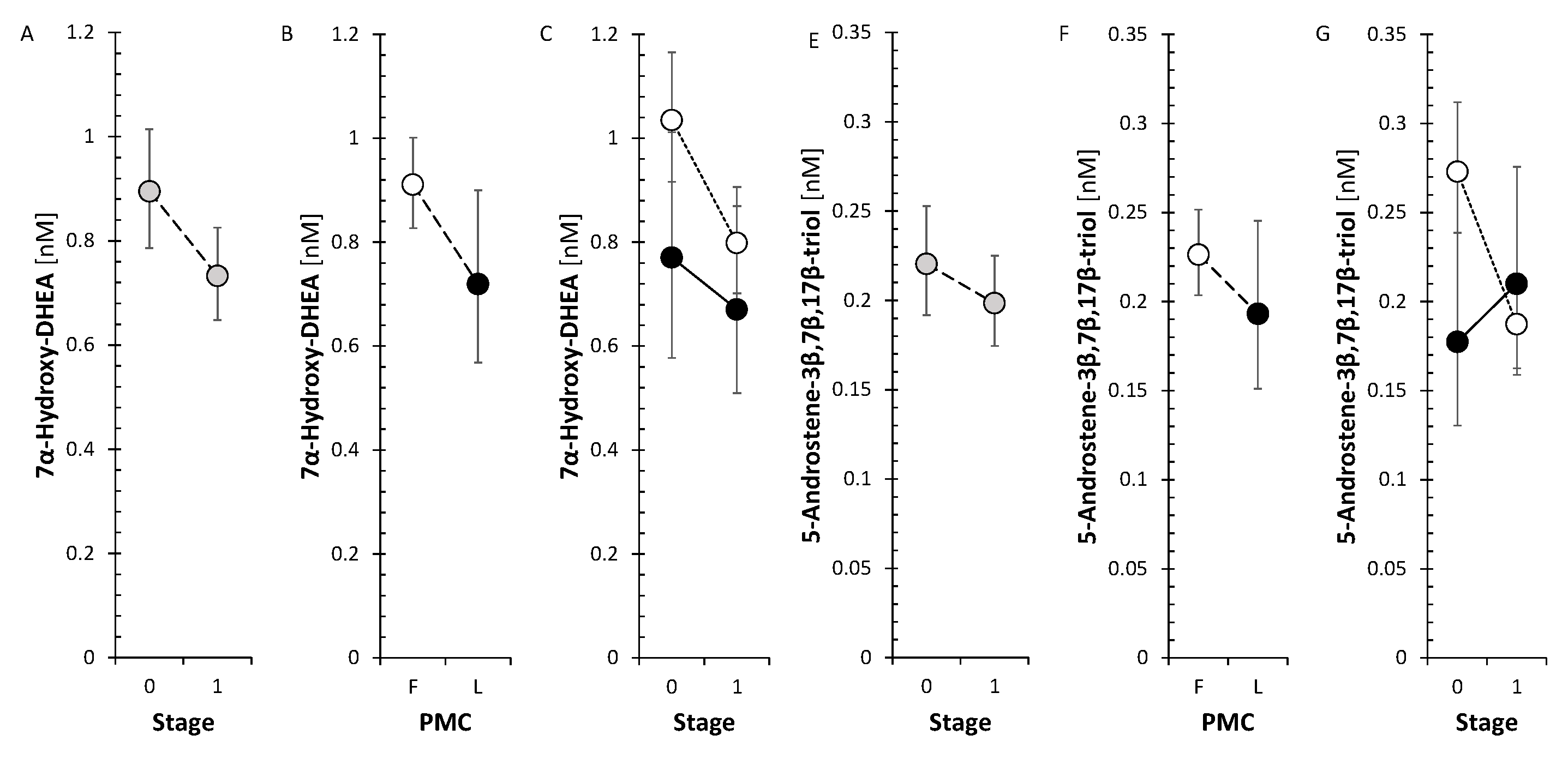
Figure 4.
Changes of testosterone and conjugated 5α-dihydrotestosterone levels in patients with MS after treatment with anti-MS drugs. The drawings and symbols are the same as for Figure 1. The statistics for factors and interaction are as follows: Stage: F=9.8, p=0.003, ηp2=0.154; PMC: F=0, p=0.945, ηp2=0.0000883; Stage × PMC: F=3.3, p=0.077, ηp2=0.0568; Subj: F=3, p<0.001, ηp2=0.765 for testosterone and Stage: F=4.8, p=0.033, ηp2=0.0863; PMC: F=0.9, p=0.361, ηp2=0.0164; Stage × PMC: F=1.5, p=0.226, ηp2=0.0286; Subj: F=1.4, p=0.1, ηp2=0.618 for 5α-dihydrotestosterone.
Figure 4.
Changes of testosterone and conjugated 5α-dihydrotestosterone levels in patients with MS after treatment with anti-MS drugs. The drawings and symbols are the same as for Figure 1. The statistics for factors and interaction are as follows: Stage: F=9.8, p=0.003, ηp2=0.154; PMC: F=0, p=0.945, ηp2=0.0000883; Stage × PMC: F=3.3, p=0.077, ηp2=0.0568; Subj: F=3, p<0.001, ηp2=0.765 for testosterone and Stage: F=4.8, p=0.033, ηp2=0.0863; PMC: F=0.9, p=0.361, ηp2=0.0164; Stage × PMC: F=1.5, p=0.226, ηp2=0.0286; Subj: F=1.4, p=0.1, ηp2=0.618 for 5α-dihydrotestosterone.
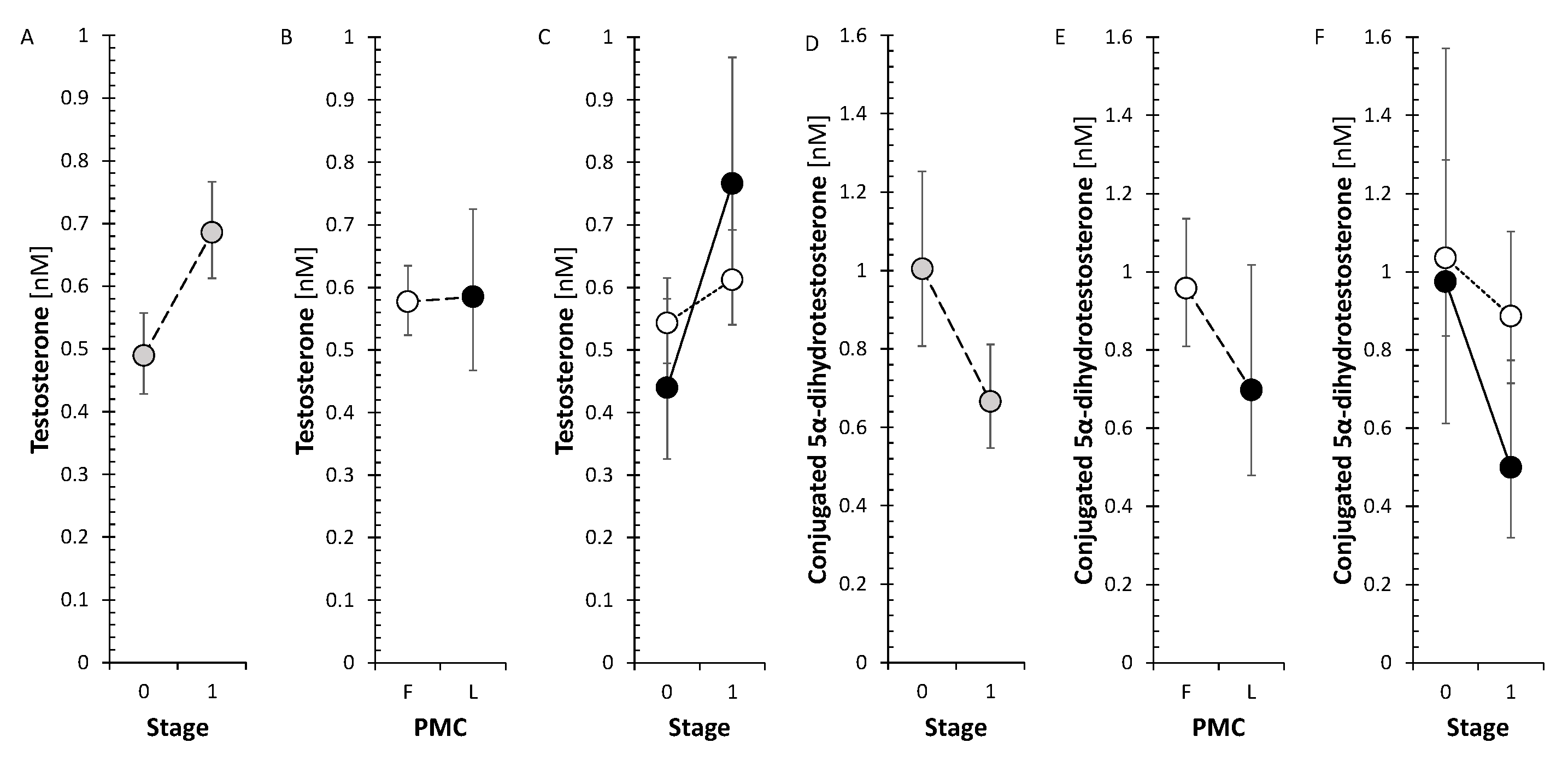
Figure 4.
Changes of 17-hydroxyallopregnanolone sulfate, conjugated 17-hydroxypregnanolone, conjugated 5α-pregnane-3α,17,20α-triol, and conjugated 5β-pregnane-3α,17,20α-triol levels in patients with MS after treatment with anti-MS drugs. The drawings and symbols are the same as for Figure 1. The statistics for factors and interaction are as follows: Stage: F=5.2, p=0.026, ηp2=0.087; PMC: F=5, p=0.029, ηp2=0.0836; Stage × PMC: F=0.1, p=0.831, ηp2=0.000837; Subj: F=3.5, p<0.001, ηp2=0.791 for 17-hydroxyallopregnanolone sulfate, Stage: F=8, p=0.007, ηp2=0.127; PMC: F=3.2, p=0.082, ηp2=0.0541; Stage × PMC: F=0, p=0.961, ηp2=0.0000441; Subj: F=2.4, p<0.001, ηp2=0.721 for conjugated 17-hydroxypregnanolone, Stage: F=9.1, p=0.004, ηp2=0.142; PMC: F=0.2, p=0.634, ηp2=0.00415; Stage × PMC: F=1.6, p=0.205, ηp2=0.029; Subj: F=1.9, p=0.011, ηp2=0.666 for 5α-pregnane-3α,17,20α-triol, and Stage: F=11.7, p=0.001, ηp2=0.178; PMC: F=0.1, p=0.742, ηp2=0.00202; Stage × PMC: F=2.6, p=0.114, ηp2=0.0455; Subj: F=3.4, p<0.001, ηp2=0.787 for conjugated 5β-pregnane-3α,17,20α-triol.
Figure 4.
Changes of 17-hydroxyallopregnanolone sulfate, conjugated 17-hydroxypregnanolone, conjugated 5α-pregnane-3α,17,20α-triol, and conjugated 5β-pregnane-3α,17,20α-triol levels in patients with MS after treatment with anti-MS drugs. The drawings and symbols are the same as for Figure 1. The statistics for factors and interaction are as follows: Stage: F=5.2, p=0.026, ηp2=0.087; PMC: F=5, p=0.029, ηp2=0.0836; Stage × PMC: F=0.1, p=0.831, ηp2=0.000837; Subj: F=3.5, p<0.001, ηp2=0.791 for 17-hydroxyallopregnanolone sulfate, Stage: F=8, p=0.007, ηp2=0.127; PMC: F=3.2, p=0.082, ηp2=0.0541; Stage × PMC: F=0, p=0.961, ηp2=0.0000441; Subj: F=2.4, p<0.001, ηp2=0.721 for conjugated 17-hydroxypregnanolone, Stage: F=9.1, p=0.004, ηp2=0.142; PMC: F=0.2, p=0.634, ηp2=0.00415; Stage × PMC: F=1.6, p=0.205, ηp2=0.029; Subj: F=1.9, p=0.011, ηp2=0.666 for 5α-pregnane-3α,17,20α-triol, and Stage: F=11.7, p=0.001, ηp2=0.178; PMC: F=0.1, p=0.742, ηp2=0.00202; Stage × PMC: F=2.6, p=0.114, ηp2=0.0455; Subj: F=3.4, p<0.001, ηp2=0.787 for conjugated 5β-pregnane-3α,17,20α-triol.
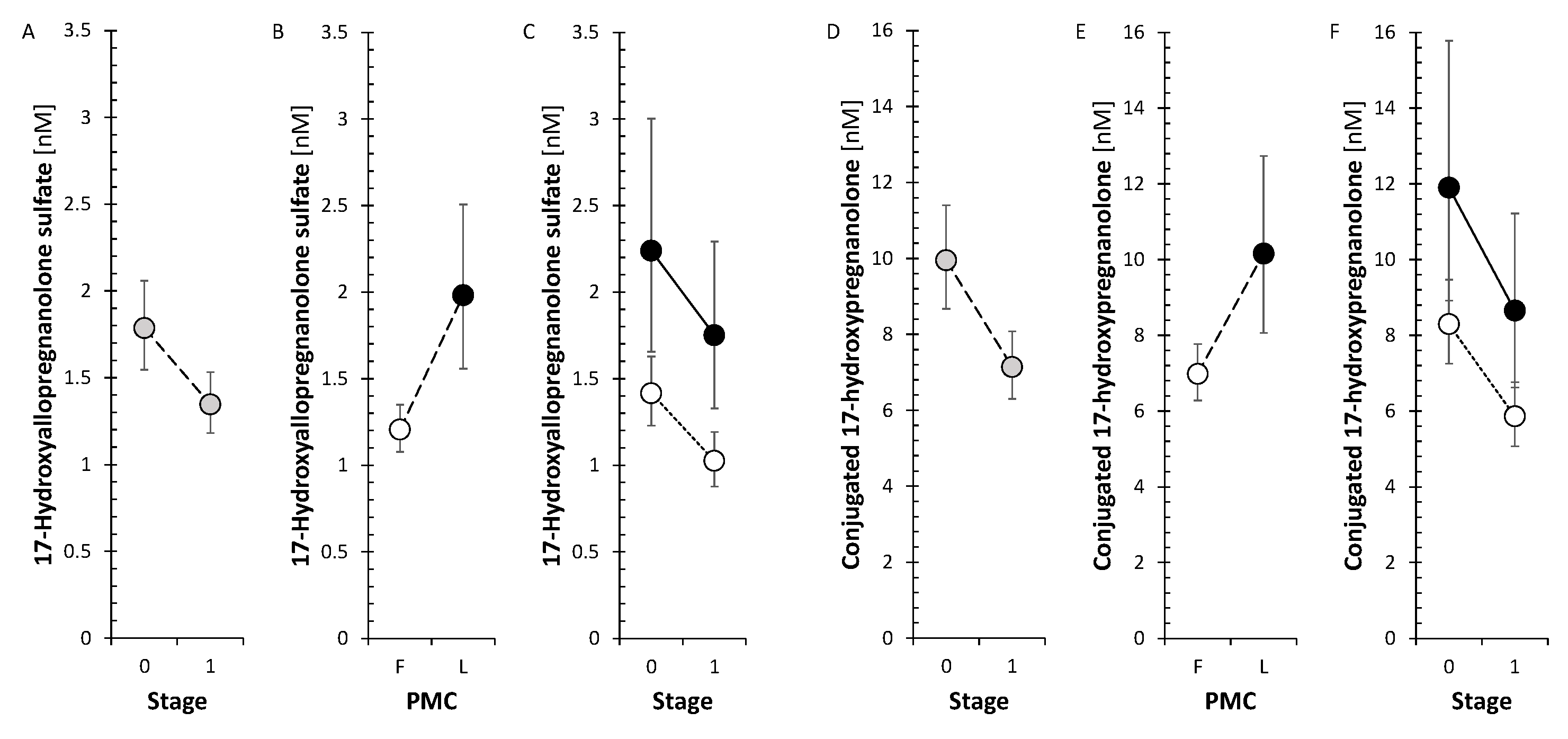
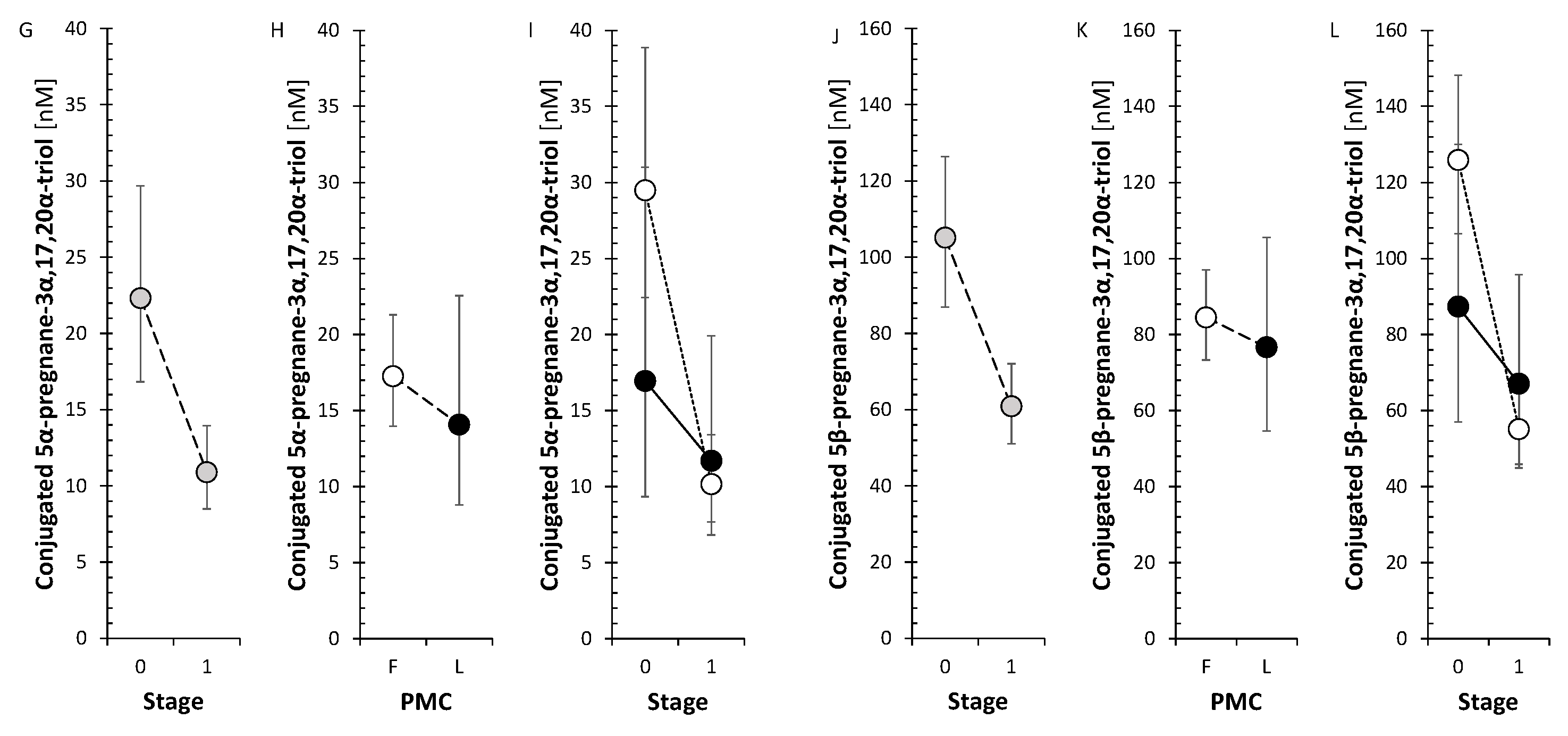
Figure 5.
Scheme illustrating alterations in circulating steroids in patients with early MS, advanced MS and controls and in patients treated with anti-MS drugs.
Figure 5.
Scheme illustrating alterations in circulating steroids in patients with early MS, advanced MS and controls and in patients treated with anti-MS drugs.
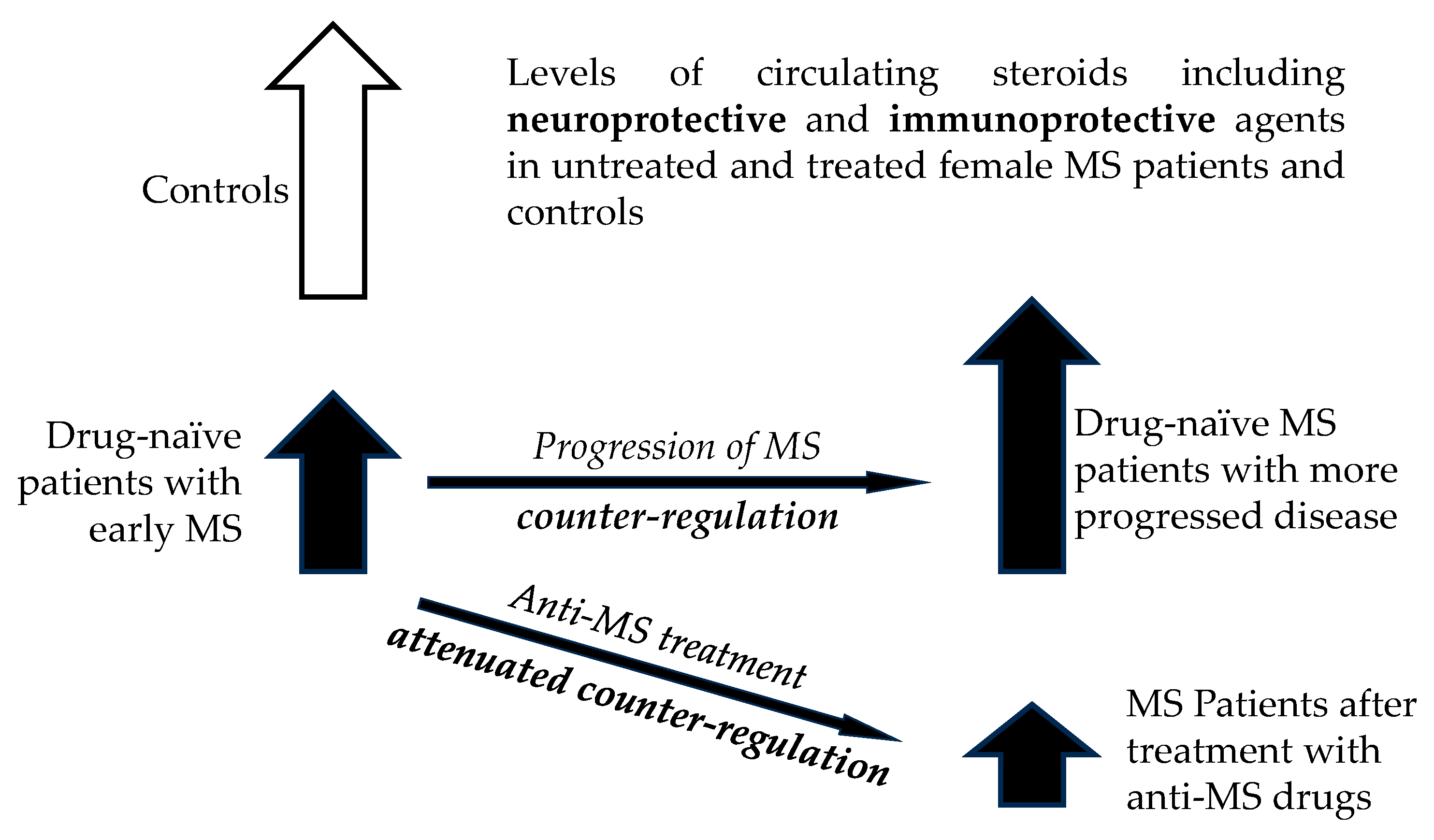
Figure 6.
Scheme illustrating changes in circulating steroids in patients treated with individual anti-MS drugs.
Figure 6.
Scheme illustrating changes in circulating steroids in patients treated with individual anti-MS drugs.

Table 1.
Trends in steroids and steroid molar ratios (that may reflect activities of steroidogenic enzymes and balances between them) after treatment with anti-MS drugs.
Table 1.
Trends in steroids and steroid molar ratios (that may reflect activities of steroidogenic enzymes and balances between them) after treatment with anti-MS drugs.
| Steroids and steroid molar ratios | Trend | Follicular phase | Luteal phase | Steroids and steroid molar ratios | Trend | Follicular phase | Luteal phase | Steroids and steroid molar ratios | Trend | Follicular phase | Luteal phase | Steroids and steroid molar ratios | Trend | Follicular phase | Luteal phase | |||
| Steroids | ↓ | 12 | 0 | 17-oxo- Androstanes/#break#estrogens | ↓ | 2 | 0 | CYP17A1,#break#hydroxylase | ↓ | 4 | 1 | AKR1D1 | ↓ | 1 | 0 | |||
| ~ | 66 | 78 | ~ | 19 | 21 | ~ | 8 | 12 | ~ | 10 | 13 | |||||||
| ↑ | 1 | 1 | ↑ | 0 | 0 | ↑ | 1 | 0 | ↑ | 2 | 0 | |||||||
| p | 0.002 | 0.323 | p | 0.168 | 1 | p | 0.193 | 0.356 | p | 0.176 | 1 | |||||||
| Unconjugated steroids | ↓ | 6 | 0 | 17β-hydroxy Androstanes + estrogens | ↓ | 1 | 0 | CYP17A1,#break#lyase | ↓ | 1 | 0 | AKR1C1 vs. HSD17B2 | ↓ | 5 | 0 | |||
| ~ | 38 | 44 | ~ | 14 | 14 | ~ | 7 | 13 | ~ | 9 | 16 | |||||||
| ↑ | 1 | 1 | ↑ | 0 | 1 | ↑ | 5 | 0 | ↑ | 3 | 1 | |||||||
| p | 0.06 | 0.328 | p | 0.351 | 0.351 | p | 0.111 | 1 | p | 0.496 | 0.347 | |||||||
| Conjugated steroids | ↓ | 6 | 0 | 20-oxo-Pregnanes | ↓ | 5 | 0 | HSD3B | ↓ | 1 | 0 | AKR1C2 vs. HSD17B2 and HSD17B6 | ↓ | 2 | 2 | |||
| ~ | 28 | 34 | ~ | 17 | 23 | ~ | 10 | 11 | ~ | 18 | 18 | |||||||
| ↑ | 0 | 0 | ↑ | 1 | 0 | ↑ | 0 | 0 | ↑ | 0 | 0 | |||||||
| p | 0.015 | 1 | p | 0.107 | 1 | p | 0.363 | 1 | p | 0.168 | 0.168 | |||||||
| C21 steroids (pregnanes) | ↓ | 9 | 0 | 20α-Hydroxy- pregnanes | ↓ | 4 | 0 | Conjugated/#break#unconjugated steroids | ↓ | 7 | 0 | AKR1C2 vs. HSD17B2 and HSD17B6, 3α/β-hydroxy steroids | ↓ | 2 | 2 | |||
| ~ | 33 | 44 | ~ | 16 | 20 | ~ | 19 | 26 | ~ | 11 | 11 | |||||||
| ↑ | 1 | 0 | ↑ | 0 | 0 | ↑ | 0 | 0 | ↑ | 0 | 0 | |||||||
| p | 0.012 | 1 | p | 0.049 | 1 | p | 0.009 | 1 | p | 0.175 | 0.175 | |||||||
| C19 steroids (androstanes) | ↓ | 3 | 0 | 3α-Hydroxy-5α/β-pregnanes | ↓ | 4 | 0 | CYP11B1 | ↓ | 2 | 4 | AKR1C2 vs. HSD17B2 and HSD17B6, 3α-hydroxy/3-oxo steroids | ↓ | 0 | 0 | |||
| ~ | 31 | 33 | ~ | 11 | 15 | ~ | 5 | 3 | ~ | 7 | 7 | |||||||
| ↑ | 0 | 1 | ↑ | 0 | 0 | ↑ | 0 | 0 | ↑ | 0 | 0 | |||||||
| p | 0.086 | 0.332 | p | 0.05 | 1 | p | 0.192 | 0.056 | p | 1 | 1 | |||||||
| Corticoids and 11β-hydroxy-androstanes | ↓ | 4 | 0 | 3-oxo/3β-Hydroxy-5α/β-pregnanes | ↓ | 1 | 0 | CYP7B1, 3A4 | ↓ | 4 | 0 | AKR1C3 | ↓ | 2 | 0 | |||
| ~ | 7 | 11 | ~ | 12 | 14 | ~ | 4 | 8 | ~ | 8 | 12 | |||||||
| ↑ | 0 | 0 | ↑ | 1 | 0 | ↑ | 0 | 0 | ↑ | 2 | 0 | |||||||
| p | 0.051 | 1 | p | 1 | 1 | p | 0.054 | 1 | p | 1 | 1 | |||||||
| 17-Hydroxy-pregnanes | ↓ | 6 | 0 | 7α/β- and 16α-Hydroxy-steroids | -1 | 2 | 0 | HSD11B1 | ↓ | 0 | 0 | |||||||
| ~ | 8 | 14 | 0 | 6 | 8 | ~ | 4 | 4 | ||||||||||
| ↑ | 0 | 0 | 1 | 0 | 0 | ↑ | 0 | 0 | ||||||||||
| p | 0.016 | 1 | p | 0.187 | 1 | p | 1 | 1 | ||||||||||
| 17-Deoxy-pregnanes | ↓ | 3 | 0 | CYP17A1,#break#hydroxylase + lyase | ↓ | 3 | 0 | SRD5A | ↓ | 1 | 0 | |||||||
| ~ | 25 | 29 | ~ | 18 | 23 | ~ | 9 | 13 | ||||||||||
| ↑ | 1 | 0 | ↑ | 2 | 0 | ↑ | 3 | 0 | ||||||||||
| p | 0.326 | 1 | p | 0.67 | 1 | p | 0.339 | 1 |
Table 2.
Effect of anti-MS drugs on steroidomic data including steroid molar ratios (that may reflect activities of steroidogenic enzymes and balances between them).
Table 2.
Effect of anti-MS drugs on steroidomic data including steroid molar ratios (that may reflect activities of steroidogenic enzymes and balances between them).
| Steroids and steroid molar ratios | Trend | GA, follicular phase | IFNβ-1a, follicular phase | IFNβ-1a,#break#luteal phase | S1PR inhibitors,#break#follicular phase | Ofatumumab, follicular phase | Ocrelizumab, follicular phase | Ocrelizumab,#break#luteal phase |
| Steroids | ↓ | 0 | 6 | 0 | 11 | 17 | 16 | 20 |
| ~ | 79 | 71 | 58 | 67 | 62 | 62 | 56 | |
| ↑ | 0 | 2 | 21 | 1 | 0 | 1 | 3 | |
| p | 1 | 0.159 | <0.001 | 0.004 | <0.001 | <0.001 | <0.001 | |
| Corticoids and 11β-hydroxy-androgens | ↓ | 0 | 1 | 0 | 2 | 1 | 0 | 5 |
| ~ | 11 | 10 | 6 | 9 | 10 | 10 | 6 | |
| ↑ | 0 | 0 | 5 | 0 | 0 | 1 | 0 | |
| p | 1 | 0.363 | 0.029 | 0.178 | 0.363 | 0.363 | 0.029 | |
| Δ5 and Δ4 steroids | ↓ | 0 | 3 | 0 | 3 | 8 | 6 | 6 |
| ~ | 11 | 27 | 20 | 27 | 22 | 23 | 23 | |
| ↑ | 0 | 0 | 10 | 0 | 0 | 1 | 1 | |
| p | 1 | 0.087 | 0.002 | 0.087 | 0.005 | 0.061 | 0.061 | |
| Progesterone 5α/β-reduced metabolites | ↓ | 0 | 3 | 0 | 7 | 6 | 7 | 5 |
| ~ | 26 | 23 | 20 | 18 | 20 | 19 | 19 | |
| ↑ | 0 | 0 | 6 | 1 | 0 | 0 | 2 | |
| p | 1 | 0.087 | 0.015 | 0.035 | 0.015 | 0.009 | 0.264 | |
| CYP17A1 | ↓ | 0 | 1 | 0 | 1 | 0 | 0 | 8 |
| ~ | 23 | 18 | 23 | 16 | 23 | 23 | 15 | |
| ↑ | 0 | 4 | 0 | 6 | 0 | 0 | 0 | |
| p | 1 | 0.187 | 1 | 0.061 | 1 | 1 | 0.005 | |
| CYP17A1, hydroxylase | ↓ | 0 | 3 | 1 | 3 | 5 | 6 | 7 |
| ~ | 13 | 10 | 11 | 10 | 6 | 7 | 6 | |
| ↑ | 0 | 0 | 1 | 0 | 2 | 0 | 0 | |
| p | 1 | 0.092 | 1 | 0.092 | 0.273 | 0.016 | 0.009 | |
| CYP17A1, lyase | ↓ | 0 | 0 | 0 | 0 | 1 | 0 | 0 |
| ~ | 11 | 8 | 13 | 13 | 8 | 6 | 13 | |
| ↑ | 2 | 5 | 0 | 0 | 4 | 7 | 0 | |
| p | 0.175 | 0.028 | 1 | 1 | 0.193 | 0.009 | 1 | |
| HSD3Bs | ↓ | 0 | 0 | 0 | 0 | 4 | 0 | 0 |
| ~ | 11 | 11 | 11 | 11 | 7 | 11 | 9 | |
| ↑ | 0 | 0 | 0 | 0 | 0 | 0 | 2 | |
| p | 1 | 1 | 1 | 1 | 0.051 | 1 | 0.178 | |
| Conjugated/#break#unconjugated steroids | ↓ | 6 | 3 | 0 | 4 | 5 | 4 | 6 |
| ~ | 20 | 22 | 26 | 21 | 20 | 22 | 20 | |
| ↑ | 0 | 1 | 0 | 1 | 1 | 0 | 0 | |
| p | 0.015 | 0.327 | 1 | 0.186 | 0.106 | 0.048 | 0.015 | |
| CYP11B1 | ↓ | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| ~ | 7 | 7 | 7 | 7 | 7 | 7 | 7 | |
| ↑ | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| p | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |
| CYP7B1, CYP3A4, CYP3A7 | ↓ | 0 | 0 | 0 | 2 | 5 | 3 | 4 |
| ~ | 8 | 8 | 8 | 5 | 3 | 5 | 4 | |
| ↑ | 0 | 0 | 0 | 1 | 0 | 0 | 0 | |
| p | 1 | 1 | 1 | 0.621 | 0.031 | 0.099 | 0.054 | |
| HSD11B1 | ↓ | 0 | 0 | 0 | 0 | 2 | 1 | 0 |
| ~ | 4 | 4 | 4 | 4 | 1 | 2 | 4 | |
| ↑ | 0 | 0 | 0 | 0 | 1 | 1 | 0 | |
| p | 1 | 1 | 1 | 1 | 0.7 | 1 | 1 | |
| Steroids and steroid molar ratios | Trend | GA, follicular phase | IFNβ-1a, follicular phase | IFNβ-1a,#break#luteal phase | S1PR inhibitors,#break#follicular phase | Ofatumumab, follicular phase | Ocrelizumab, follicular phase | Ocrelizumab,#break#luteal phase |
| SRD5As | ↓ | 0 | 0 | 0 | 0 | 1 | 1 | 0 |
| ~ | 13 | 13 | 13 | 8 | 9 | 9 | 13 | |
| ↑ | 0 | 0 | 0 | 5 | 3 | 3 | 0 | |
| p | 1 | 1 | 1 | 0.028 | 0.339 | 0.339 | 1 | |
| AKR1D1 | ↓ | 0 | 0 | 0 | 4 | 1 | 0 | 7 |
| ~ | 13 | 13 | 8 | 8 | 8 | 13 | 6 | |
| ↑ | 0 | 0 | 5 | 1 | 4 | 0 | 0 | |
| p | 1 | 1 | 0.028 | 0.193 | 0.193 | 1 | 0.009 | |
| AKR1C1 vs. HSD17B2 | ↓ | 6 | 3 | 2 | 5 | 3 | 0 | 2 |
| ~ | 11 | 12 | 12 | 10 | 13 | 17 | 15 | |
| ↑ | 0 | 2 | 3 | 2 | 1 | 0 | 0 | |
| p | 0.015 | 0.676 | 0.676 | 0.268 | 0.333 | 1 | 0.17 | |
| AKR1C2 vs. HSD17B2 and HSD17B6 | ↓ | 3 | 2 | 3 | 2 | 3 | 2 | 0 |
| ~ | 15 | 16 | 17 | 14 | 15 | 15 | 21 | |
| ↑ | 3 | 3 | 1 | 5 | 3 | 4 | 0 | |
| p | 1 | 0.672 | 0.33 | 0.266 | 1 | 0.427 | 1 | |
| AKR1C2 vs. HSD17B2 and HSD17B6, 3α/β-hydroxy steroids | ↓ | 3 | 1 | 3 | 2 | 3 | 2 | 0 |
| ~ | 9 | 10 | 11 | 10 | 10 | 9 | 14 | |
| ↑ | 2 | 3 | 0 | 2 | 1 | 3 | 0 | |
| p | 0.682 | 0.337 | 0.091 | 1 | 0.337 | 0.682 | 1 | |
| AKR1C2 vs. HSD17B2 and HSD17B6, 3α-hydroxy/3-oxo ster | ↓ | 0 | 1 | 0 | 0 | 0 | 0 | 0 |
| ~ | 6 | 6 | 6 | 4 | 5 | 6 | 7 | |
| ↑ | 1 | 0 | 1 | 3 | 2 | 1 | 0 | |
| p | 0.391 | 0.391 | 0.391 | 0.102 | 0.192 | 0.391 | 1 | |
| AKR1C3 vs. HSD17B2 | ↓ | 0 | 0 | 0 | 0 | 5 | 0 | 3 |
| ~ | 12 | 12 | 12 | 12 | 7 | 12 | 9 | |
| ↑ | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| p | 1 | 1 | 1 | 1 | 0.028 | 1 | 0.093 | |
| p = p-value, ↓ = trend to decrease, ~ = absent trend, ↑ = trend to increase, p < 0.05 (bold), p < 0.1 (normal), and p > 0.1 (italics) | ||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



