
Submitted:
13 January 2025
Posted:
14 January 2025
You are already at the latest version
Abstract
The role of natural killer (NK) cells in the management of prostate cancer (PCa) remains incompletely understood. Some have proposed that measuring NK cells in blood samples could serve as a reliable, minimally invasive tool for screening, assessing treatment effects, and predicting survival outcomes in PCa patients. Existing studies support the hypothesis that NK cell assays may be useful for screening, diagnosing, staging, and predicting patient outcomes. However, the significance of different NK cell phenotypes remains unclear. Given the interplay between NK cells and the microbiome, we hypothesize that a combined signature of NK cell phenotypes derived from blood, along with microbiome profiles from oral, urine, and stool samples, could serve as a surrogate marker for NK cell activity. Such an approach provides a practical alternative to invasive tumor biopsies by enabling the indirect assessment of NK cell function in tumors. Additionally, profiling NK cell phenotypes and their interactions with the microbiota has the potential to enhance prognostic accuracy and guide the development of personalized therapeutic strat-egies. Prospective studies are needed to validate the utility of NK cell and microbiome assays in personalized PCa management, with a focus on minimally invasive procedures and predictive signatures for treatment outcomes.
Keywords:
Prostate Cancer
; Biomarker
; NK cells
; Microbiota
; Triple NK cell biomarker approach
1. Introduction
Prostate Cancer (PCa) is the second most common noncutaneous neoplasm and the fifth most common cause of cancer related deaths in men worldwide [1]. PCa is a heterogeneous disease, with most cases progressing slowly [2], but the prognosis can vary significantly. Early detection and monitoring of PCa are critical for effective management. Measurement of serum Prostate-Specific Antigen (PSA) is the cornerstone of current diagnostic practices [3]. Since its introduction in the late 1960s, PSA testing has dramatically shifted the staging of PCa, with most cases diagnosed as prostate-confined tumors [4]. Despite its widespread use, the optimal cutoff for elevated PSA remains debated [5]. Using a threshold of 4 ng/mL, the positive predictive value of PSA in diagnosing PCa varies widely from 25% to 40% [6], with a sensitivity and specificity of around 20% and 65% [7].
While PSA testing has increased early detection, it has also led to overdiagnosis, contributing to a rise in PCa incidence, particularly in the mid-1980s. Recent recommendations for more conservative PSA testing have reduced the diagnosis of new cases [8]. The conventional diagnostic tool for PCa is Transrectal Ultrasound (TRUS)-guided biopsy [9], but it has limitations, including invasiveness, side effects, and a risk of false-negative results [10]. To improve diagnostic accuracy, Magnetic Resonance Imaging (MRI)-targeted biopsy is being explored as a more effective alternative [11].
The risk of recurrence after curative management of PCa primarily depends on the disease stage at initial diagnosis, which is typically determined by clinical Tumor, Node, and Metastasis (TNM) staging, PSA levels, and biopsy Gleason Score (GS) [2]. Radical prostatectomy and radiotherapy are the primary treatments for localized and locally advanced PCa [12], while Androgen Deprivation Therapy (ADT) remains the standard medical treatment for both high-risk localized PCa and metastatic disease [13]. Unfortunately, nearly all patients with incurable PCa eventually progress to the Castration-Resistant. Prostate Cancer (CRPC) stage, with 35% developing metastases and a survival rate of just 2-4 years post-progression [14].
Accumulating evidence suggests that impaired immune response plays a critical role in the pathogenesis of PCa [15,16]. Both preclinical and clinical studies have highlighted the involvement of inflammation in PCa development and progres- sion [17,18]. Immune biomarkers are being increasingly recognized as valuable tools for the early diagnosis and management of various cancers, including PCa [19,20]. Understanding the role of the immune system, particularly the innate immune system, is crucial for improving therapeutic outcomes.
The immune system’s defense mechanisms are primarily mediated by the innate and acquired immune systems, which distinguish between “self” and “non-self” antigens [21]. Natural Killer (NK) cells are a key component of the innate immune response and differ from B and T lymphocytes. Representing 10-15% of all circulating lymphocytes, NK cells are identified by the absence of CD3 and the presence of CD56 [22,23]. NK cells are found throughout the tumor stroma and around the glandular epithelium, indicating their potential involvement in both tumor progression and the immune response [24]. Inflammation in the tumor microenvironment (TME) can further compromise NK cell function, limiting their ability to mount an effective anti-tumor response. These cells are further subdivided into two populations based on CD16 and CD56 expression: CD56dim CD16+cytotoxic) and CD56brighCD16- (cytokine producers). CD56bright NK cells are less mature but have the potential to differentiate into CD56dim cells with strong cytotoxic activity [25]. CD56bright NK cells are less mature but have the potential to differentiate into CD56dim cells with strong cytotoxic activity [21].
As part of the innate immune system, NK cells can recognize the absent or low expression of class I Human Leukocyte Antigen (HLA) molecules on target cells and lyse them directly. The NK cells can also recognize and bind to the NK recognition structure of the tumor cells, which leads to the releasing of NK Cytotoxic Factor (NKCF) and tumor cell lysis [26]. NK cell activity also depends on the recycling capacity of NK cells after lysis of the tumor cell and can be augmented by interferon [22]. The functional activity of NK cells is regulated by a balance of activating and inhibitory receptors. Activating receptors, such as NKG2D, and inhibitory receptors, such as Killer Immunoglobulin-like Receptors (KIR), dictate NK cell function when ligated with stress ligands or Major Histocompatibility Complex (MHC) class I molecules [18,25,27,28,29].
In addition to the direct cytotoxicity mediated by perforin and granzymes, NK cells can also kill target cells through Antibody-Dependent Cell-Mediated Cytotoxicity (ADCC) [30]. In this process, NK cells express the CD16 receptor, which binds to the Fc region of antibodies that have been bound to target tumor cells. This interaction activates NK cells to release cytotoxic granules and induce tumor cell lysis [15].
Furthermore, NK cell activity can be enhanced by cytokines such as Interleukin-2 (IL-2) and interferons (IFNs) but can also be suppressed by factors such as suppressor T cells, monocytes, and prostaglandins [31]. The NK cell recognition of tumor cells can be compromised by immunosuppressive cytokines like Transforming Growth Factor-beta (TGF-β), which downregulate activating receptors and impair NK cell function [32].
NK cell activity may have potential applications as both a diagnostic marker, complementing PSA testing, and a prognostic factor in PCa patients [13,15,33]. The immune response, including NK cell function, can also be influenced by therapies such as ADT, which increase the number of circulating lymphocytes, including NK cells [34]. However, for better diagnostic accuracy, NK cell activity in blood samples must be matched with the phenotype of infiltrating NK cells in tumors.
Despite their potential, relying solely on NK cell biomarkers has limitations, particularly the challenge of monitoring phenotypic changes during treatment via biopsies. To overcome this, combining NK cell biomarkers with additional markers—such as microbiome profiles from oral, stool and urine samples—could facilitate the design of large-scale prospective studies to identify a combinatorial signature of NK cells and the microbiome. Recent studies have underscored the important role of the microbiota in PCa pathogenesis [35], highlighting how specific microorganisms in Pca tissue, such as Vibrio parahaemolyticus and Shewanella, can alter immune responses, potentially aiding tumor evasion [36]. Additionally, the presence of microbes like Delftia acidovorans and Gardnerella vaginalis is linke to down-regulation of immune-related genes, suggesting an immune-suppressive TME in PCa [37].
Given that NK cells interact with the microbiome, we hypothesize that a combined signature of NK cell phenotypes and microbiome profiles from stool, urine and oral samples could serve as a surrogate marker for NK cell activity. This approach offers a feasible alternative to directly monitoring NK cell phenotyping through biopsies, enabling indirect assessment of NK cell function in tumors through NK cell activity in blood tests and microbiome signatures. The role of NK cells in the management of PCa remains incompletely understood. Measuring NK cells in blood samples has been proposed as a reliable, minimally invasive tool for screening, assessing treatment effects, and predicting survival outcomes in PCa patients. This review explores current evidence on the potential role of NK cells in PCa management, with a particular focus on the interaction between NK cells and the microbiome.
2. NK Cells and PCa
2.1. NK cells: Pathogenesis Significance in PCa
2.1.1. Impaired Proliferation of NK Cells
Several studies have highlighted that the suppression of NK cell function, both at the neoplastic and pre-neoplastic stages of tumorigenesis, as well as during the induction and progression of cancer, plays a critical role in disease development [38]. Mechanisms of NK cell dysfunction, such as impaired proliferation, reduced cytotoxicity, and limited tumor infiltration, are central to the pathogenesis of cancer [39] including PCa. These mechanisms can overlap, as a reduced frequency of NK cells in peripheral blood often reflects decreased NK cell activity. For example, a lower frequency of NK cells can result in decreased production of IFN-γ, a key marker of NK cell cytotoxic activity. However, it’s important to note that NK cell activity in peripheral blood may not necessarily correspond to the same function or phenotype within the tumor and its microenvironment (TME) [40]. Therefore, it is essential to measure both peripheral blood NK cells and tumor-infiltrating NK cells to accurately assess their true correlation. Only by doing so can we determine whether blood tests can serve as reliable surrogates for the actual function of NK cells in the tumor and its TME.
Elevated NK cell expression within tumors has been linked to a lower risk of recurrence, suggesting that NK cells play a protective role in controlling tumor growth [41,42,43]. A reduction in the frequency of the cytotoxic CD56dim NK cell population—both in peripheral blood and within tumors—appears to be a critical factor in cancer progression. While Sotosec et al. found no significant differences in NK cell percentages or subsets in peripheral blood between patients with localized, locally advanced, or metastatic PCa, Koo et al. demonstrated a significantly higher CD56dim -to-CD56bright ratio in the peripheral blood of PCa patients compared to healthy controls. This ratio increased progressively with cancer stage [3,15].
The impact of cancer therapies on NK cell proliferation has been explored in several studies. For example, androgen deprivation therapy (ADT), or low testosterone levels associated with ADT, may contribute to decreased NK cell counts in peripheral blood, potentially indicating a shift of NK cells from circulation to tumor sites [44,45,46]. This could impair NK cell function in peripheral blood but increase NK cell infiltration into the tumor and its microenvironment [47]. In fact, prostate cancer patients undergoing ADT have shown higher interferon γ (IFN-γ) expression. Perforin production was lower in PCa group comparing to control which can be attributed to low perforin production in cancerous tissues compared to healthy contorol.
Radiotherapy reduces leukocyte and lymphocyte counts in peripheral blood, [48] but it may also alter TME to a more favorable to NK cells, potentially enhancing their infiltration into solid tumors, including prostate cancer (PCa) [49]. Docetaxel, a chemotherapy agent, has been shown to enhance NK cell cytotoxicity and may promote NK cell proliferation through immune signaling pathways like cGAS/STING-IFN. However, the effect of docetaxel on NK cell infiltration into tumors is less clear, and changes in the TME may influence tumor infiltration indirectly [50].
The mobilization and egress of NK cells during exercise have also been extensively discussed in the literature [51]. Although acute exercise induces NK cell mobilization into circulation, lymphocyte numbers tend to decline sharply during the first few hours of recovery [52,53,54]. Interestingly, CD56bright cells, a less mature and less frequent NK subset primarily responsible for cytokine production, show a smaller response to exercise compared to their CD56dim counterparts [55,56]. Studies on PCa survivors have confirmed that acute exercise induces a temporary increase in NK cell numbers [57,58,59,60]. However, the subsequent sharp decline in NK cell counts during the recovery phase raises concerns about the potential long-term impact of this reduction, such as increased susceptibility to infections or diminished NK cell function [61] In addition, these fluctuations in NK cell numbers may influence immune cell infiltration and tumor viability [62].
2.1.2. Decreased Cytotoxicity
CD56dim NK cells, particularly those expressing CD57 (a marker of NK cell maturation), are the primary subset responsible for cytotoxic function [63,64]. These cells exhibit greater cytotoxic activity than CD57-negative NK cells and have a higher capacity for mobilization and egress compared to other NK subsets [65]. The PCa TME is characterized by an enrichment of exhausted CD56dim NK cells. These NK cells, despite their cytotoxic potential, often exhibit reduced activity due to a combination of immune evasion mechanisms, including the dysregulation of activating and inhibitory receptors [66]. Additionally, NK cells produce interferon γ (IFN-γ) upon activation, which may further enhance their cytotoxic function [55,67]. However, in PCa, immune escape mechanisms are often linked to the low killing capability of NK cells. For example, the Hypoxia-Inducible Factor 1 (HIF-1α) pathway can inhibit the NCR1/NKp46 signaling pathway via miR-224, reducing NK cell activity [68].
A study of peripheral blood NK cells from PCa patients revealed an exhausted NK phenotype with significantly reduced cytotoxicity. These exhausted NK cells release cytokines and chemokines that recruit monocytes, which then polarize into M2-like tumor-associated macrophages (TAMs). These TAMs promote angiogenesis and immune suppression within the TME, facilitating further immune evasion by the tumor. Profiling peripheral NK cells in PCa patients could provide valuable insights into immune dysfunction and help inform the development of targeted therapies [69].
One significant mechanism contributing to NK cell dysfunction in PCa is the downregulation of NKG2D, an activating receptor crucial for NK cell-mediated tumor recognition. Gallazi et al. proposed that downregulation of NKG2D on circulating NK cells in patients with localized or locally advanced PCa is associated with increased markers of NK cell exhaustion, such as PD-1 and Tim-3 [69]. Moreover, Lundholm et al. demonstrated significant downregulation of NKG2D on circulating NK cells in castration-resistant prostate cancer (CRPC) patients compared to healthy controls [70]. In de novo metastatic PCa, tumor recognition by NK cells involves the activation of NKp46, NKG2D, and NKp30 receptors [44]. However, in patients with advanced PCa, the maximal cytotoxic potential and recycling capacity of NK cells are reduced, leading to suppressed NK cell activity [71]. Additionally, cytokine dysregulation is thought to contribute to reduced NK cell cytotoxicity in advanced PCa. Lahat’s group found that patients with advanced PCa exhibited the lowest NKA and IL-2 secretion compared to those with localized PCa and healthy controls [72]. The high levels of soluble MICA (sMICA) observed in advanced PCa patients were linked to reduced NKA [73].
Inhibitory receptors, such as KIRs, may also contribute to immune escape by limiting NK cell cytotoxicity. While some studies have suggested a link between KIR gene expression and various cancers, including hepatocellular carcinoma [74] and bladder cancer [75] , Portela et al. found no significant difference in KIR and HLA gene frequencies between healthy controls and untreated localized PCa patients [76]. This suggests that KIRs may not play a major role in immune evasion in localized PCa, though they may be relevant in other cancer types or advanced stages.
Interestingly, a preclinical study has highlighted an unexpected role for NK cells in the PCa TME. It showed that NK cells preferentially migrate to CRPC cells rather than normal prostate cells and may suppress the expression of androgen receptor splicing variant 7 (ARv7), a marker of enzalutamide resistance. This suppression leads to reduced CRPC cell growth and invasion [77]. NK cell adoptive transfer therapy could therefore represent an effective treatment for enzalutamide resistant CRPC.
TGFβ1 and prostaglandin E2 (PGE2) are known to drive NK cell exhaustion, which further dampens immune responses. Additionally, dendritic cells (DCs) can interact with NK cells and play a key role in modulating their function in cancer. DCs can enhance NK cell cytotoxicity, improving their ability to eliminate tumor cells [78]. As a result, DCs are being explored as a therapeutic target, with ongoing efforts to develop autologous DC-based immunotherapies to boost NK cell activity and improve tumor eradication [79].
Several studies have reviewed the mechanisms underlying tumor cell immune escape and NK cell dysfunction within the PCa TME (Table 1). Despite the use of varying methodologies, these studies consistently found reduced NK cell activity in PCa patients, with all agreeing that low NK cell activity is associated with higher tumor incidence and metastasis. These findings suggest that enhancing NK cell activity could serve as a potential adjuvant therapy to prevent relapse, either as a standalone treatment or in combination with other cancer therapies to inhibit disease progression.
A better understanding of the detailed mechanisms behind NK cell dysfunction in the PCa TME may be crucial in developing PCa-specific NK cell-based therapies or prophylactic strategies to improve clinical outcomes.
2.1.3. Diminished Tumor Infiltration
There is some debate regarding the clinical significance of immune cell infiltration, particularly the role of different NK cell subpopulations in tumor tissue. For example, mononuclear cells from peri-prostatic lymph nodes in PCa patients—whether localized, locally advanced, or metastatic—showed significantly lower reactivation by beta-interferon stimulation compared to those from healthy subjects or autologous peripheral mononuclear cells [80]. These findings suggest that the PCa TME may impair NK cell activity by altering the composition of NK subpopulations. Furthermore, while the physical disruption of the tumor capsule possibly leading to tumor invasion in patients with localized and advanced PCa, this theory is still debated [81]. Even within an activatory NK cell subset, there is a lack of consensus on the impact of NK cell tumor infiltration.
In 2016, Pasero et al. reported that NK cell infiltration into localized and metastatic PCa tissues was primarily driven by CD56/Neural Cell Adhesion Molecule 1 (NCAM1)-positive NK cells, a phenotype associated with low or absent cytotoxic potential [18]. This raises further questions about the functional role of NK cell infiltration in the PCa TME.
In the context of immune suppression, the role of M2 tumor-associated macrophages (TAMs) is notable. M2 TAMs promote tumor growth and metastasis by lacking NK cell activity and supporting new blood vessel formation. They are driven by cytokines like IL-6, IL-10, and TGFβ, especially in hypoxic conditions or with high IL-4 levels. These macrophages release factors like TGF-β and IL-10 to aid their own maturation. Additionally, non-coding RNAs and transcription factors, such as c-Myc, contribute to M2 activation and stimulate cancer-associated fibroblasts (CAFs) [82,83].
Interestingly, combining ADT with exercise has shown promise in improving NK cell infiltration and reducing tumor volume in PCa [84]. This highlights the potential of lifestyle interventions in modulating immune responses within the TME.
2.2. NK Cells: Screening/Diagnosis Significance in PCa
Accurate risk prediction is a major clinical challenge in PCa management, as it is critical for determining the most appropriate treatment approach. While men with low-risk PCa may not require active treatment, the mere diagnosis of the disease can have significant social, financial, and psychological implications. Currently, the PSA test is the primary screening tool, but it lacks accuracy and reliability, often leading to unnecessary biopsies. As a result, there is a growing need for alternative, non-invasive approaches—such as circulating biomarkers—that offer greater specificity and sensitivity Table 2.
Recent studies suggest that the detection of NK cells in peripheral blood could play a role in the early diagnosis of PCa. For example, Barkin et al. observed in a small cohort of 43 patients that those with lower levels of NK cells were more likely to have a positive prostate biopsy result 85. Additionally, recent findings indicate that peripheral NK cells in PCa patients are associated with enhanced expression of markers like CD9, CD49a, CXCR4, CXCL8, and MMP-9. These factors are involved in monocyte recruitment and polarization, highlighting their potential role in PCa diagnosis 69.
Furthermore, NK cell assays could serve as valuable tools for detecting residual microscopic disease after prostatectomy, potentially informing personalized adjuvant treatment strategies for high-risk patients. Specific markers such as low IFN-γ levels, decreased CD56bright NK cells, high CD56dim -to- CD56bright NK cell ratios, and elevated sMIC may indicate a higher risk of residual disease. Monitoring NK cell activity during radiotherapy could help ensure that NK cell activity does not decrease, which could otherwise contribute to disease progression.
2.2.1. Radionuclide Labeling Method
Some studies using the 51Cr release assay have suggested that NK cells could serve as biomarkers for both the diagnosis and staging of PCa [81,86]. One 1993 study reported a sharp decline in NK cell activity in patients with tumor lesions in lymph nodes, bones, or soft tissues, although tumor differentiation had no impact on these results. The study proposed that measuring NK cell lytic capacity could supplement routine clinical staging [86].
2.2.2. Flow Cytometry and Machine Learning
A computerized model based on profiling NK cell subsets in the blood of 72 asymptomatic men with PSA levels below 20 ng/ml showed that certain NK cell markers, such as CD56dim CD56bright, CD56+ NKp30+, and CD56+ NKp46+, may help detect the presence of PCa. Furthermore, NK cell profiling demonstrated the ability to differentiate between low, intermediate, and high-risk PCa by analyzing 32 phenotypic features [87].
2.2.3. NK Vue Cytokine Release Method
A 2013 study found NK cell activity to be a potential diagnostic marker for PCa with a sensitivity of 72% and specificity of 74%. The CD56dim -to- CD56bright cell ratio showed a sensitivity of 66% and specificity of 71% [15,88]. Additional findings supported NK cell activity as a predictor of positive biopsy results, with patients exhibiting low NK cell activity having five times the risk of biopsy-verified PCa [89]. A pilot study indicated that patients with NKA levels below 200 pg/mL had an 86% risk of having PCa [90]. However, a cross-sectional study questioned the utility of NK cell activity for detecting PCa or predicting Gleason scores, noting that serial NK cell activity measurements were needed for more conclusive assessments [91].
2.2.4. Secretome Analysis
Unlike the NK Vue test, secretome analysis offers a broader perspective on NK cell behavior by identifying a range of secreted factors that influence tumor progression and immune responses. For example, Gallazzi et al. demonstrated that NK cells in the PCa TME release pro-inflammatory cytokines and chemokines that recruit monocytes and promote M2-like polarization, which may facilitate immune suppression and tumor growth [69]
2.2.5. MICA ELISA
One study found elevated levels of soluble MICA (sMICA, 21 ng/ml) in the serum of nearly all PCa patients with a Gleason score ≥6, suggesting that sMICA could serve as a novel biomarker for PCa [73]. NK cell assays, including those measuring sMICA, may complement routine clinical management of PCa, although methods like 51Cr-release assays are difficult to manage due to their reliance on radioisotopes. Alternative methods like the ATP assay or flow cytometry-based assays offer more practical ways to assess NK cell activity, with the ATP assay measuring both NK cell activity and other cytotoxic activities [20,92].
2.2.6. Molecular Profiling
A comprehensive molecular profiling study, including whole-exome sequencing (WES) and whole-transcriptome sequencing (WTS), involved 3,365 PCa samples. In prostate biopsies, higher NK cells were less common with certain mutations and AR-V7 but more common with immune suppressors, suggesting an anti-inflammatory response [93]. Recent advances in single-cell RNA sequencing have revealed a variety of innate immune cells in normal prostate tissue, with transcriptional changes observed in PCa. Notably, CD16neg NK cell signatures and CCL5 transcripts are found together in the TME, suggesting that these resident-like NK cells may recruit tumor antigens recriuting cells and potentially improve treatment responses [94].
2.3. NK Cells: Prognostic and Predictive Significance in PCa
The prognostic and predictive roles of NK cells in PCa have been widely investigated, with the potential to improve personalized treatment strategies, including the use of adjuvant therapies. This section reviews the findings from multiple studies assessing the significance of NK cells as both prognostic and predictive biomarkers in PCa (Table 3).
2.3.1. NK Cells and Prognostic Significance
For instance, Gannon et al. found that a high number of CD56+ NK cells in localized PCa were linked to a lower risk of disease progression [24]. Similarly, Pasero et al. showed that increased CD56+ NK cells in prostate tumors correlated with reduced tumor invasion in seminal vesicles, translating to improved survival outcomes. Specifically, patients with high NKp30 expression had a 3-year survival rate of 85%, compared to just 38% in those with low NKp30 expression [44].
Other studies further support this finding. For example, research from 1995 highlighted an association between high expression of HNK-1 antigen in well-differentiated PCa and better prognosis [33]. Moreover, radiotherapy-induced gastrointestinal (GI) side effects appeared to improve prognosis by stimulating the release of Heat Shock Proteins (HSPs), which augment immune function and enhance NK cell cytotoxicity [95].
In contrast, lower NK cell activity has been linked to more advanced disease stages. Marumo et al. reported significantly lower NK cell activity in advanced PCa patients compared to localized cases and healthy controls [71]. Similarly, Lu et al. demonstrated that postoperative NKA was higher in lower-stage PCa patients and those with negative surgical margins, suggesting NK cell activity as a marker of better post-surgical outcomes [96]. However, Blomgren et al. found that post-radiotherapy NK cell activity was lower than pre-radiotherapy levels, potentially contributing to disease progression [97].
Moreover, several studies have noted an increased ratio of CD56dim-to- CD56bright NK cells in the blood of patients with more advanced PCa stages, further supporting the link between NK cell function and disease severity [15]. In metastatic cancer, a significant reduction in NK cell cytotoxicity has been observed, correlating with poor prognosis, especially in patients with high circulating tumor cells (CTCs) [19].
Therapeutic interventions can also affect NK cell activity. A Phase I trial combining ipilimumab and PROSTVAC immunotherapy showed an increase in NK cell subsets expressing Tim-3+, correlating with improved survival [98]. However, a study involving ADT found no significant correlation between IL-2 levels (an NK cell activator) and progression-free survival (PFS) in metastatic PCa, suggesting that NK cell activity is not always predictive of overall survivor (OS) [99] , despite ADT induced CD56+ increased NK cells in PCa, was associated with good prognosis [24].
Moreover, NK cell markers such as NKp46 have been identified as predictive of OS and time to castration resistance (TCR) [44]. A study monitoring serial serum NK cell activity during treatment of mCRPC found a significantly lower response rate and shorter PFS in patients with IFN-γ levels below 200 pg/mL [100].
Recent studies also support the role of NK cells in predicting OS across multiple cancer types, including PCa. Higher NK cell infiltration has been linked to improved OS, particularly in tumors with negative immune regulators, such as AR-V7 [93]. Elevated NK cell expression in prostate tumors is also associated with a lower risk of biochemical recurrence after radical prostatectomy [24,44].
However, NK cells infiltrating prostate tissues often display immature characteristics, with decreased expression of activating receptors such as NKG2D and CD16. This phenotypic immaturity may be mediated by factors like TGFβ1 and PGE2, which impair NK cell function, reducing their cytotoxic potential [18].
Furthermore, the expression of various NK cell subpopulations, such as CD56dim has been shown to correlate with exhaustion and poor prognosis [17]. In peripheral blood, high monocyte counts have been linked to aggressive disease features and poor survival in patients with CRPC, although this association is less clear in earlier disease stages [101,102,103].
Interactional factors affecting NK cell activity within the TME have also been shown to be associated with prognosis in many studies [104].
NK cell markers have also been linked to prognosis in patients undergoing ADT. NK cells from metastatic PCa patients who show a longer response to castration therapy exhibit phenotypic and functional patterns associated with high expression of activating receptors and molecules involved in NK cell maturation and degranulation [44].
Sipuleucel-T, a dendritic cell (DC)-based immunotherapy, has shown improved overall survival in PCa patients. However, the results have been controversial due to unfavorable outcomes in the control arm. Subsequent DC vaccines have failed to produce significant benefits, and the utility of this immunotherapy strategy remains unproven [78].
One study showed that radiotherapy-induced GI or GU toxicity led to an increase in NK cell activity, which was linked to a better prognosis, suggesting that treatment-induced immune activation could benefit patients [95].
Only a limited number of studies have focused on specific NK cell markers, such as NKp30 and NKp46, which have been identified as predictive of TCR and OS in mPCa [44] , [68]. Furthermore, one 1995 study focused on HNK-1 antigen expression in tumor tissue [33] , while another study found no association between IL-2 levels and time to castration-resistant prostate cancer (CRPC) or OS, although NK cell activity was not directly investigated in this study[32].
2.3.2. NK Cells and Predictive Significance
The role of NK cell activity as a predictor of treatment response, particularly in immunotherapy, has been widely discussed [22]. However, while NK cell activity shows promise as a biomarker, the presence or number of NK cells alone is not yet considered a reliable indicator of treatment outcomes. For instance, Johnson et al. studied patients with non-metastatic PCa who had biochemical recurrence and were treated with anti-tumor DNA vaccination. They found no significant difference in NK cell levels between immune responders and non-responders, suggesting that NK cell quantity alone may not predict treatment success [105].
A few studies have specifically investigated NK cell activity as a predictive biomarker, discussing that NK cell activity levels could potentially predict longer responses to castration therapy [44,99]. However, another study that assessed the role of NK cells in response to DNA vaccination found no significant correlation between NK cell levels and treatment outcomes [105].
A retrospective study of untreated mPCa patients showed that high expression of NKp46 was associated with a better response to castration therapy, with response rates of 39% in patients with high NKp46 expression compared to just 8% in those with low expression [44].
Madan et al. recently investigated the immunologic effects of enzalutamide in non-metastatic hormone sensitive PCa patients. In their study, 38 patients treated with short-course enzalutamide showed an increase in antigen-specific T-cell levels targeting PSA, along with a rise in NK cell numbers and a decrease in myeloid-derived suppressor cells in the blood. However, no significant correlation was found between these immune changes and clinical responses, which may be due to the small sample size [106].
3. Microbiome and PCa
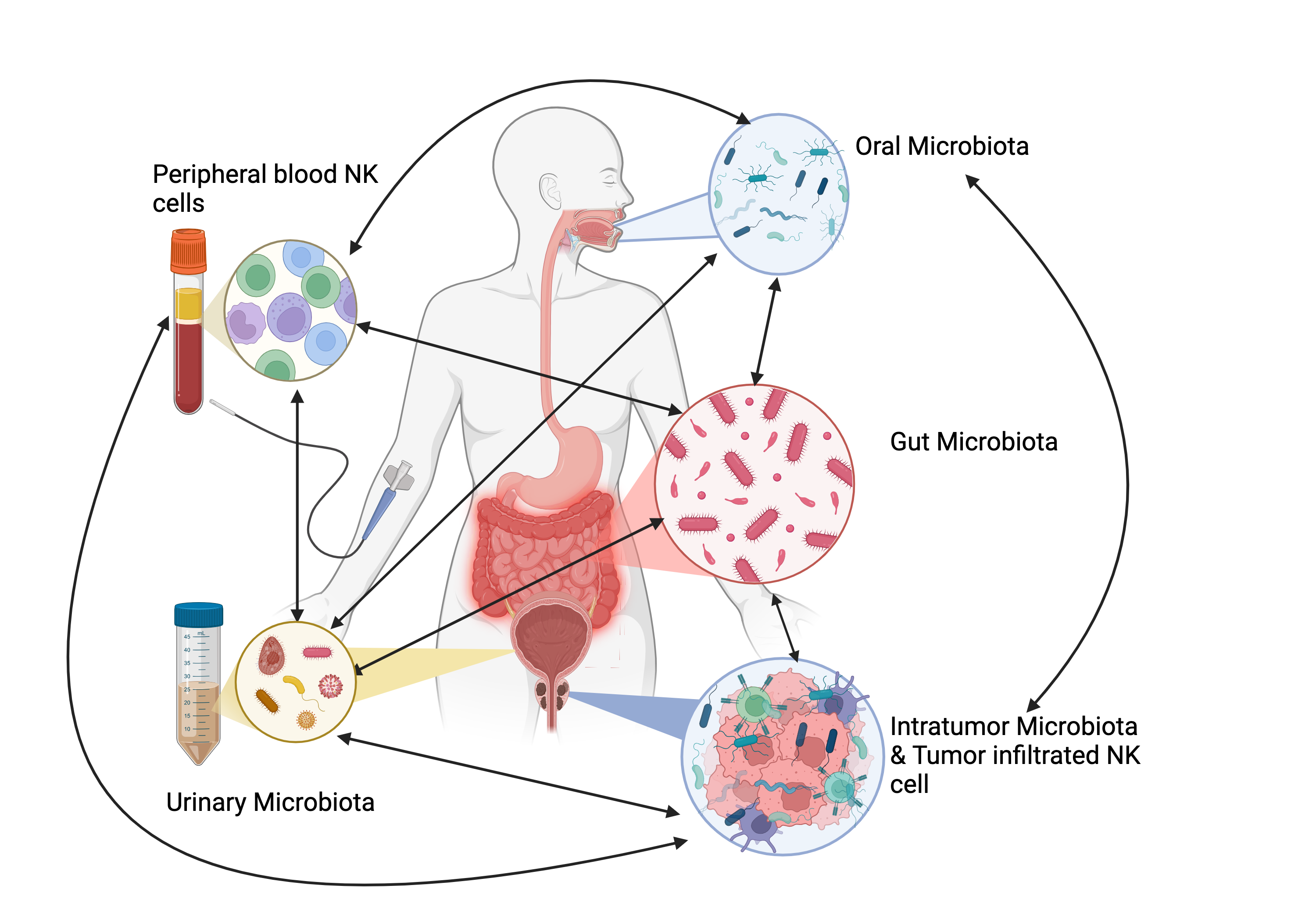
Many studies have examined the link between the microbiome and PCa, exploring its potential as a diagnostic and therapeutic biomarker [107] , Figure 1.
The microbiome plays a critical role in metabolism, immune function, and overall health, with disruptions (dysbiosis) linked to various diseases, including cancer. Notably, the oral microbiome has been shown to influence these processes, and when the oral barrier is compromised, microbes can spread through the gut microbiome axis to other parts of the body, potentially contributing to cancer development [108,109].
In addition, NK cells are involved not only in the antitumor immune response, but also in defending against microbial infection [110]. Their activity can be influenced by changes in the microbiome, further highlighting the interconnectedness between microbial health and cancer. Further research into the “missing self” hypothesis regarding MHC molecules and NK cell recognition may also provide insight into how the microbiome interacts with immune surveillance in the context of PCa [111].
The Prostate, Lung, Colorectal, and Ovarian (PLCO) Cancer Screening Trial, which included about 155,000 participants in the U.S., studied cancer screening methods and collected serum samples over several years [112]. One case-control study from this trial found that elevated levels of specific metabolites, including choline, carnitine, PAGln, and p-cresol sulfate, were linked to a higher risk of lethal prostate cancer, although the associations with other metabolites remain uncertain [113].
While this review does not delve into the methodologies for analyzing the microbiome, these have been extensively covered in prior studies [36].
3.1. Oral Microbiota
While a retrospective study suggests that reliable oral microbial biomarkers for PCa may not yet exist [114] , more recent research has highlighted significant associations between oral microbiota and PCa risk.
A 2020 genome-wide association study (GWAS) conducted in an Asian population, which included 442 PCa cases and 195,745 controls, found strong correlations between PCa and specific bacterial species in the oral cavity. This study identified 27 bacterial species in the tongue, belonging to 17 genera and 14 families, and 42 bacterial species in saliva, spanning 24 genera and 17 families. Notably, nine genera were found in both tongue and saliva samples from PCa patients [35]. Moreover, oral microbial displacement has been linked to the development of prostate cancer [115]. Dysbiosis in the oral microbiota can lead to the colonization of the prostate by oral bacteria, increasing the risk of developing PCa [116].
Interestingly, many studies now recognize that the NK cells’ ability to detect and fight cancer is heavily influenced by the host’s microbial ecosystem [ ref.104].
3.2. Gut Microbiota
Although the prostate is not physically linked to the gut, lifestyle factors that impact the gut microbiome have been found to influence the pathogenesis of PCa [107]. In prostate cancer, certain bacteria, like Alistipes and Lachnospira, are found in higher levels in the gut, suggesting they may influence cancer progression [117]. Additionally, Reichard et al. demonstrated that certain metabolites are linked to a higher risk of fatal prostate cancer, further linking the microbiome to cancer development [118].
Liss et al. found differences in the microbiome between cancer and non-cancer samples, particularly in Bacteroides and Streptococcus species. They also identified 10 altered metabolic pathways linked to prostate cancer, leading to a new microbiome-based risk factor with a promising diagnostic model (AUC = 0.64, p=0.02) [119].
Zhong et al. found that gut dysbiosis, caused by antibiotics and the overgrowth of Proteobacteria, increased gut permeability and lipopolysaccharide (LPS) levels, which promoted PCa in mice. This suggests Proteobacteria could be a biomarker for prostate cancer progression in humans [120].
Other studies found significant differences in the gut microbiota between PCa cases and controls, specifically the higher relative abundance of Bacteroides and Streptococcus species in cancer cases, while Faecalibacterium prausnitzii and Eubacterium rectale were more abundant in controls. Biologically significant differences were also noted in the relative abundance of genes, pathways, and enzymes associated with these bacteria [119,121].
A Mendelian randomization (MR) study identified significant links between 28 gut microbiome taxa and 75 immune cell types with prostate diseases. Notably, the Dorea bacteria may influence immune responses in the prostate, with specific NK cell subsets enhancing its positive effects, potentially lowering the risk of benign prostatic hyperplasia (BPH) [122].
In terms of treatment response, one study showed no significant microbial differences in fecal samples when comparing benign and malignant prostate biopsies [123]. However, Akkermansia muciniphila was found to help regulate changes in the microbiome caused by the drug Abiraterone acetate, suggesting its potential role in modulating treatment responses [124].
There are also distinct differences in the microbiota between hormone-sensitive prostate cancer (HSPC) and castration-resistant prostate cancer (CRPC), highlighting the need for further research. Understanding these differences could help develop strategies to enhance responses to androgen deprivation therapy (ADT) [125]. A preclinical study found that intestinal bacteria and their metabolites, particularly short-chain fatty acids (SCFAs), promoted PCa growth in mouse models [126]. Antibiotic treatment to eliminate the gut microbiota delayed development to castration resistance, and fecal transplants from CRPC patients or mice made tumors resistant to castration treatment[125,127]. In contrast, transplants from hormone-sensitive patients or the addition of Prevotella stercorea controlled tumor growth, suggesting that the gut microbiota contributes to treatment resistance by producing androgens [127].
These findings underscore the complex relationship between the gut microbiome, NK cells and PCa, highlighting its potential role in both cancer progression and treatment response. Further research is needed to fully understand these interactions and identify microbiome-based biomarkers for PCa diagnosis and therapy.
3.3. Urinary Microbiota
In the past, urine was believed to be sterile, which limited research on the urinary microbiome. However, recent studies have revealed its potential role in conditions like lower urinary tract symptoms and inflammatory diseases, and cancerous conditions including PCa. Notably, the act of urination can transfer microbes to the prostate, potentially contributing to prostatic dysbiosis, asymptomatic changes in the prostate environment, and chronic inflammation. [128,129,130,131,132].
In prostate cancer patients, specific bacteria like Bacteroidetes and Firmicutes were found to be more abundant, while others, such as Eubacterium, were less common. Escherichia coli was lower in urine but more prevalent in seminal fluid and epididymal prostate secretions (EPS), while Enterococcus levels increased in seminal fluid [133]. A study of the male urinary microbiome discovered that men with PCa had higher levels of pro-inflammatory bacteria and uropathogens in their urinary tract compared to healthy individuals [134].
A prospective study that examined the link between both urinary and fecal microbiota and PCa diagnosis using 16S rRNA gene analysis found significant differences in the urinary microbiota of prostate cancer patients compared to those without the disease. The study also noted distinct differences in the urinary microbiota of patients with BPH and PCa compared to control subjects [123] , emphasizing the need for further research to identify specific microbes that may contribute to prostate disease. Another research identified several novel bacterial species—Porphyromonas sp. nov., Varibaculum sp. nov., Peptoniphilus sp. nov., and Fenollaria sp. nov.—in the urine of PCa patients. The presence of certain anaerobic bacteria was linked to a higher risk of cancer and metastasis [135]. These findings support the idea that the urinary microbiome could influence the inflammatory environment of the prostate, potentially impacting NK cell activity and the immune response [129].
Moreover, changes in the urinary microbiome, particularly the decreased abundance of certain bacterial species, have been associated with PCa, especially in patients with a Gleason score of 7 or higher. [123] These alterations highlight the potential of the urinary microbiome as a biomarker for PCa and its role in shaping the prostate’s immune landscape. Although no studies have specifically investigated the interaction between NK cells and the urinary microbiome in PCa, this remains an area of interest for future research.
3.4. Intraprostatic/Intratumoral Microbio
The idea that infections may play a role in PCa was first proposed by Ravich et al. in 1951 [136]. Subsequent research has reinforced this hypothesis, with a meta-analysis of 29 studies linking PCa risk to a history of sexually transmitted infections, such as Mycoplasma genitalium and human papillomavirus (HPV) [137,138]. In addition, chronic inflammation is believed to contribute to PCa development by inducing oxidative stress and genetic mutations [129]. Cavarretta et al. investigated the prostate microbiome and identified bacterial species associated with PCa. However, their study had limitations, as it did not assess the role of inflammation or consider non-bacterial microorganisms, such as mycoplasmas and viruses, that may also influence disease progression [139]. Interestingly, bacterial patterns detected in extracellular vesicles and cancerous tissues have been linked to PCa progression, though the precise role of these bacteria in NK cell suppression and cancer development remains unclear [135].
4. Discussion
Understanding the molecular mechanisms underlying NK cell function, microbiota and their interaction in immune surveillance is critical for advancing PCa therapies. This review provides an overview of the role of NK cells and the microbiome in the management of PCa, highlighting their potential as biomarkers for diagnosis, prognosis, and treatment response. The studies reviewed underscore initially the value of NK cells in guiding clinical decisions and improving patient outcomes as a single biomarker. Initial studies proposed NKA as a promising marker for prognosis and treatment response in PCa [Table 1-3]. Some studies, in particular, highlighted that low NK cell activity levels at diagnosis were associated with shorter TCR and OS [44] Additionally, one study demonstrated that radiotherapy-induced gastrointestinal or genitourinary toxicity could upregulate NK cell activity, resulting in better prognosis [95]. Monitoring NK cell function during radiotherapy in patients without radiotherapy-induced side effects could ensure that NK cell activity levels do not decline, thus informing treatment decisions. NK cell assays in PCa could be utilized to detect microscopic residual disease after prostatectomy, helping to design personalized adjuvant therapies for high-risk patients. For example, low levels of IFN-γ, HNK-1 antigen, alongside high CD56dim -to- CD56bright NK cell ratios and sMIC may indicate a high risk of undetected microscopic disease[15,33]. Furthermore, some studies investigated NK cell activity as a predictive biomarker. One study found NK cell activity as a potential predictive biomarker to stratify patients likely to have longer castration responses arguing for therapies aimed at NK cell function augmentation in mPCa patients [44]. Although a small study could not confirm an association between the amount of NK cells and treatment response in PCa patients after DNA vaccination, it was suggested that different measures of antigen-specific tolerance, or regulation, might help predict the immunological outcome of DNA vaccination [105]. However, there are several important limitations to consider. Most of the studies included had relatively small sample sizes, with only a few containing more than 25 participants, which may limit the generalizability of the findings. Additionally, methodological differences across studies—such as the use of outdated technologies, inconsistent patient selection criteria, and variations in statistical analyses—further complicate the interpretation of results. While NK cells show promise as biomarkers, it is essential to recognize that NK cell alone may not be optimal marker for managing PCa. The integration with other biomarkers could significantly enhance diagnostic accuracy and prognostication. Recent studies have highlighted the potential role of the microbiome, particularly in the oral, stool, and urinary microbiota, in shaping the prostate’s immune environment. For instance, specific microbial patterns have been linked to PCa risk, inflammation, and disease progression. Microbial dysbiosis could influence the inflammatory microenvironment, subsequently affecting NK cell function. Thus, combining NK cell phenotypes with microbiome profiles, or utilizing approaches such as the triple NK cell biomarker strategy described earlier [ref. 143], which focuses on the interaction between the tumor, TME, and NK cells, could enhance PCa management. This approach targets three mechanisms of NK cell suppression: low proliferation, low toxicity, and low infiltration, offering a more comprehensive and non-invasive diagnostic and therapeutic method. This approach allows for monitoring NKA and the TME using easily accessible biological samples, such as blood, urine, stool, or saliva. By combining NK cell assays with microbiome analysis, it may be possible to track both infiltrating NK cells and circulating NK cell phenotypes as reliable biomarkers for PCa. Non-invasive microbiome testing in oral, urine, or stool samples could be used to assess the tumor’s immune environment, identifying potential therapeutic targets and improving patient outcomes. Once large-scale studies validate the correlation between circulating NK cells and their tumor-infiltrating counterparts, we may be able to rely on circulating NK cells as a surrogate marker for tumor-infiltrating NK cells, combined with microbiome status, for monitoring and managing PCa.
The findings of this review also emphasize the need for larger, more robust clinical trials. A more evidence-based approach, incorporating well-designed prospective cohort studies combining NK cell combined to microbiome as a linked biomerker is essential to validate the clinical utility of NK cell assays and microbiome profiling as predictive and prognostic biomarkers in PCa. Furthermore, future studies should adhere to established reporting guidelines to ensure their findings are clinically relevant and can be reliably integrated into clinical practice [104]. The limitations of small sample sizes and varying methodologies suggest that a more standardized approach will be key in confirming the role of NK cells and the microbiome in PCa management.
5. Conclusions
NK cells have significant potential as complementary biomarkers in the personalized management of PCa. NK cell assays could enhance diagnostic accuracy, prognostication, and treatment monitoring by improving screening, staging, and predicting treatment outcomes. To address the limitations of traditional biopsies, a combinatorial biomarker approach that integrates NK cell phenotypes (from blood samples) with microbiome profiles from easily accessible samples, such as oral, stool, or urine, could serve as a surrogate marker for NK cell activity in the tumor and its TME. This minimally invasive strategy offers a practical alternative to biopsies, enabling more continuous and feasible monitoring of NK cell function in PCa. However, large-scale, prospective studies are needed to validate the clinical utility of this approach and its potential integration into PCa management.
Author Contributions
S.F contributes to the conceptualization and design of the review content. A.H.Z. and T.F.H. contributes to supervision. S.F. wrote the first draft of the manuscript. All authors contributed to the manuscript revision, read, and approved the final submitted version.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data sharing is not applicable to this review article as it does not contain original datasets. All relevant information and sources are cited within the manuscript.
Acknowledgments
None
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Bray, F., Laversanne, M., Sung, H. et al. Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries.. CA Cancer J Clin. 74, 229–263 (2024). [CrossRef]
- Hernandez, D. J., Nielsen, M. E., Han, M. & Partin, A. W. Contemporary evaluation of the D’amico risk classification of prostate cancer. Urology 70, 931–935 (2007). [CrossRef]
- Sotosek, S. et al. Comparative study of frequency of different lymphocytes subpopulation in peripheral blood of patients with prostate cancer and benign prostatic hyperplasia. Wiener klinische Wochenschrift 123, 718–725 (2011). [CrossRef]
- Neppl-Huber, C. et al. Changes in incidence, survival and mortality of prostate cancer in Europe and the United States in the PSA era: additional diagnoses and avoided deaths. Annals of oncology: official journal of the European Society for Medical Oncology 23, 1325–1334 (2012). [CrossRef]
- Semjonow, A., Brandt, B., Oberpenning, F., Roth, S. & Hertle, L. Discordance of assay methods creates pitfalls for the interpretation of prostate-specific antigen values 3–16 (1996).
- Schröder, F. H. et al. Early Detection of Prostate Cancer in 2007. European Urology 53, 468–477 (2008).
- Qaseem, A., Barry, M. J., Denberg, T. D., Owens, D. K. & Shekelle, P. Screening for Prostate Cancer: A Guidance Statement From the Clinical Guidelines Committee of the American College of Physicians. Annals of Internal Medicine 158, 761–761 (2013).
- Scarpato, K. R. & Albertsen, P. C. Prostate-Specific Antigen Screening Guidelines. Prostate Cancer: Science and Clinical Practice: Second Edition 111–117 (2016).
- Hodge, K. K., Mcneal, J. E. & Stamey, T. A. Ultrasound guided transrectal core biopsies of the palpably abnormal prostate. The Journal of urology 142, 66–70 (1989). [CrossRef]
- Djavan, B. et al. Prospective evaluation of prostate cancer detected on biopsies 1, 2, 3 and 4: when should we stop. The Journal of urology 166, 1679–1683 (2001). [CrossRef]
- Boesen, L. Magnetic resonance imaging-transrectal ultrasound image fusion guidance of prostate biopsies: current status, challenges and future perspectives. Scandinavian Journal of Urology. Taylor and Francis Ltd. 53, 89–96 (2019).
- Graham, J., Baker, M., Macbeth, F. & Titshall, V. Diagnosis and treatment of prostate cancer: summary of NICE guidance. (2008). [CrossRef]
- Aragon-Ching, J. B., Williams, K. M. & Gulley, J. L. (2007).
- Patrikidou, A. et al. Who dies from prostate cancer. Prostate Cancer and Prostatic Disease 17, 348–352 (2014).
- Koo, K. C. et al. Reduction of the CD16-CD56bright NK cell subset precedes NK cell dysfunction in prostate cancer. PLoS One 8, 1–8 (2013). [CrossRef]
- Choe, B. K., Frost, P., Morrison, M. K. & Rose, N. R. Natural killer cell activity of prostatic cancer patients. Cancer investigation 5, 285–291 (1987).
- Hirz, T. et al. Dissecting the immune suppressive human prostate tumor microenvironment via integrated single-cell and spatial transcriptomic analyses. Nat Commun 14, 663–663 (2023). [CrossRef]
- Pasero, C. et al. Inherent and Tumor-Driven Immune Tolerance in the Prostate Microenvironment Impairs Natural Killer Cell Antitumor Activity. Cancer research 76, 2153–2165 (2016). [CrossRef]
- Santos, M. F. et al. Comparative analysis of innate immune system function in metastatic breast, colorectal, and prostate cancer patients with circulating tumor cells. Experimental and molecular pathology 96, 367–374 (2014). [CrossRef]
- Stovgaard, E. S., Nielsen, D., Hogdall, E. & Balslev, E. Triple negative breast cancer - prognostic role of immune-related factors: a systematic review. Acta Oncologica 57, 74–82 (2017).
- Abel, A. M., Yang, C., Thakar, M. S. & Malarkannan, S. Natural Killer Cells: Development, Maturation, and Clinical Utilization. Frontiers in Immunology 9 (2018).
- Hu, W., Wang, G., Huang, D., Sui, M. & Xu, Y. Cancer Immunotherapy Based on Natural Killer Cells: Current Progress and New Opportunities. Frontiers in Immunology 10 (2019). [CrossRef]
- Poli, A. et al. CD56bright natural killer (NK) cells: an important NK cell subset. Immunology 126, 458–465 (2009). [CrossRef]
- Gannon, P. O. et al. Characterization of the intra-prostatic immune cell infiltration in androgendeprived prostate cancer patients. Journal of immunological methods 348, 9–17 (2009). [CrossRef]
- Hanna, J. et al. Novel Insights on Human NK Cells’ Immunological Modalities Revealed by Gene Expression Profiling. The Journal of Immunology 173, 6547–6563 (2004). [CrossRef]
- Yu, C., Young, H. A. & Ortaldo, J. R. Characterization of cytokine differential induction of STAT complexes in primary human T and NK cells. Journal of Leukocyte Biology 64, 245–258 (1998). [CrossRef]
- Moretta, A. et al. Activating receptors and coreceptors involved in human natural killer cell mediated cytolysis. Annual review of immunology 19, 197–223 (2001). [CrossRef]
- Nausch, N. & Cerwenka, A. NKG2D ligands in tumor immunity. Oncogene 27, 5944–5958 (2008). [CrossRef]
- Sheppard, S., Ferry, A., Guedes, J. & Guerra, N. The Paradoxical Role of NKG2D in Cancer Immunity. Frontiers in Immunology 9 (2018). [CrossRef]
- Nigro, C. L. et al. NK-mediated antibody-dependent cell-mediated cytotoxicity in solid tumors: biological evidence and clinical perspectives. Annals of translational medicine 7, 105–105 (2019). [CrossRef]
- Kasai, M. et al. Brief Definitive Report Direct Evidence that Natural Killer Cells in Nonimmune Spleen Cell Populations Prevent Tumor Growth In Vivo (1979).
- Hood, S. P. et al. Phenotype and Function of Activated Natural Killer Cells From Patients With Prostate Cancer: Patient-Dependent Responses to Priming and IL-2 Activation. Frontiers in Immunology 9 (2019). [CrossRef]
- Liu, X., Zhan, B., Tadao, T. & Totsuhiro, M. Immunohistochemical study of HNK-1 (leu-7) antigen in prostate cancer and its clinical significance. Chin Med J (Engl) 108, 516–537 (1995).
- Heidegger, I. et al. A Systematic Review of the Emerging Role of Immune Checkpoint Inhibitors in Metastatic Castration-resistant Prostate Cancer: Will Combination Strategies Improve Efficacy? European urology oncology 4, 745–754 (2020).
- Feng, K., Ren, F. & Wang, X. Association between oral microbiome and seven types of cancers in East Asian population: a two-sample Mendelian randomization analysis.. Front Mol Biosci 10, 1327893–1327893 (2023). [CrossRef]
- Salachan, P. V. & Sørensen, K. D. Dysbiotic microbes and how to find them: a review of microbiome profiling in prostate cancer. J Exp Clin Cancer Res 41, 31–31 (2022). [CrossRef]
- Ma, J. et al. Influence of Intratumor Microbiome on Clinical Outcome and Immune Processes in Prostate Cancer. Cancers (Basel) 12 (2020). [CrossRef]
- Jewett, A. et al. Natural Killer Cells: Diverse Functions in Tumor Immunity and Defects in Pre-neoplastic and Neoplastic Stages of Tumorigenesis. Molecular Therapy Oncolytics 16, 41–52 (2020).https://doi.org/10.1016/j.omto.2019.11.002.
- Fanijavadi, S., Thomassen, M. & Jensen, L. H. (2024).
- Brittenden, J. Natural killer cells and cancer. Cancer 77, 1226–1269 (1996).
- Zhao, S. G. et al. The immune landscape of prostate cancer and nomination of PD-L2 as a potential therapeutic target. J Natl Cancer Inst 111, 301–311 (2019). [CrossRef]
- Liu, G. et al. Perturbation of NK cell peripheral homeostasis accelerates prostate carcinoma metastasis. J Clin Invest 123, 4410–4432 (2013).
- Liu, X. et al. MiRNA-296-3p-ICAM-1 axis promotes metastasis of prostate cancer by possible enhancing survival of natural killer cell-resistant circulating tumour cells. Cell Death Dis 4, 1–15 (2013).
- Pasero, C. et al. Highly effective NK cells are associated with good prognosis in patients with metastatic prostate cancer. Oncotarget 6, 14360–14373 (2015). [CrossRef]
- Morse, M. D. & Mcneel, D. G. Prostate cancer patients on androgen deprivation therapy develop persistent changes in adaptive immune responses. Hum Immunol 71, 496–504 (2010). [CrossRef]
- Singh, J. et al. Levels of plasma cytokine in patients undergoing neoadjuvant androgen deprivation therapy and external beam radiation therapy for adenocarcinoma of the prostate. Ann Transl Med 8, 636–636 (2020). [CrossRef]
- Page, S. T. et al. Effect of medical castration on CD4+ CD25+ T cells, CD8+ T cell IFN-γ expression, and NK cells: A physiological role for testosterone and/or its metabolites. American Journal of Physiology. Endocrinology and Metabolism 290, 856–863 (2006). [CrossRef]
- Dethlefsen, C. et al. Exercise regulates breast cancer cell viability: Systemic training adaptations versus acute exercise responses. Breast Cancer Research and Treatment 159, 469–479 (2016). [CrossRef]
- Nielsen, C. M., White, M. J., Goodier, M. R. & Riley, E. M. Functional significance of CD57 Expression on human NK cells and relevance to disease. Frontiers in Immunology 4, 422–422 (2013). [CrossRef]
- Hojan, K. et al. Physical exercise for functional capacity, blood immune function, fatigue, and quality of life in high-risk prostate cancer patients during radiotherapy: A prospective, randomized clinical study. European Journal of Physical and Rehabilitation Medicine 52, 489–501 (2016).
- Zheng, W., Ling, S., Cao, Y., Shao, C. & Sun, X. Combined use of NK cells and radiotherapy in the treatment of solid tumors.. Front Immunol 14 (2024). [CrossRef]
- Ma, Z. et al. Docetaxel remodels prostate cancer immune microenvironment and enhances checkpoint inhibitor-based immunotherapy. Theranostics 12, 4965–79 (2022). [CrossRef]
- Khosravi, N., Stoner, L., Farajivafa, V. & Hanson, E. D. Exercise training, circulating cytokine levels and immune function in cancer survivors: a meta-analysis. Brain, Behavior, and Immunity 81, 92–104 (2019). [CrossRef]
- Hanson, E. D., Bates, L. C., Moertl, K. & Evans, E. S. Natural Killer Cell Mobilization in Breast and Prostate Cancer Survivors: The Implications of Altered Stress Hormones Following Acute Exercise. Endocrines 2021 2, 121–132. [CrossRef]
- Hanson, E. D., Sakkal, S. & Que, S. Natural killer cell mobilization and egress following acute exercise in men with prostate cancer. Experimental Physiology 105, 1524–1539 (2020). [CrossRef]
- Nieman, D. C. Exercise, infection, and immunity. International Journal of Sports Medicine 15, 131–141 (1994).
- Maria, A. D., Bozzano, F., Cantoni, C. & Moretta, L. Revisiting human natural killer cell subset function revealed cytolytic CD56dimCD16+ NK cells as rapid producers of abundant IFN-γ on activation. Proceedings of the National Academy of Sciences 108, 728–732 (2011).
- Campbell, J. P. et al. Acute exercise mobilises CD8+ T lymphocytes exhibiting an effector-memory phenotype. Brain, Behavior, and Immunity 23, 767–775 (2009). [CrossRef]
- Rooney, B. V. et al. Lymphocytes and monocytes egress peripheral blood within minutes after cessation of steady state exercise: A detailed temporal analysis of leukocyte extravasation. Physiology & Behavior 194, 260–267 (2018). [CrossRef]
- Shephard, R. J. Adhesion molecules, catecholamines and leucocyte redistribution during and following exercise. Sports Med 33, 261–284 (2003).
- Hanson, E. D. et al. Altered stress hormone response following acute exercise during prostate cancer treatment. Scan- dinavian Journal of Medicine and Science in Sports 28, 1925–1933 (2018). [CrossRef]
- Galvao, D. A. et al. Endocrine and immune responses to resistance training in prostate cancer patients. Prostate Cancer and Prostatic Diseases 11, 160–165 (2008). [CrossRef]
- Campbell, J. P. & Turner, J. E. Debunking the myth of exercise-induced immune suppression: redefining the impact of exercise on immunological health across the lifespan. Frontiers in Immunology 9, 648–648 (2018). [CrossRef]
- Kared, H., Martelli, S., Ng, T. P., Pender, S. L. & Larbi, A. CD57 in human natural killer cells and T-lymphocytes 65, 441–452 (2016). [CrossRef]
- Bigley, A. B. et al. Acute exercise preferentially redeploys NK-cells with a highly-differentiated phenotype and aug- ments cytotoxicity against lymphoma and multiple myeloma target cells. Brain, Behavior, and Immunity 39, 160–171 (2014).
- Vivier, E. Innate or adaptive immunity? The example of natural killer cells. Science 331, 44–49 (2011). [CrossRef]
- Wang, R., Jaw, J. J., Stutzman, N. C., Zou, Z. & Sun, P. D. Natural killer cell-produced IFN-γ and TNF-α induce target cell cytolysis through up-regulation of ICAM-1. Journal of Leukocyte Biology 91, 299–309 (2012). [CrossRef]
- Chen, C. et al. (2018).
- Gallazzi, M. et al. Prostate Cancer Peripheral Blood NK Cells Show Enhanced CD9, CD49a, CXCR4, CXCL8, MMP-9 Production and Secrete Monocyte-Recruiting and Polarizing Factors. Frontiers in Immunology 11 (2021). [CrossRef]
- Lundholm, M. et al. Prostate Tumor-Derived Exosomes Down-Regulate NKG2D Expression on Natural Killer Cells and CD8+ T Cells: Mechanism of Immune Evasion. PLoS ONE 9, 108925–108925 (2014). [CrossRef]
- Marumo, K., Ikeuchi, K., Baba, S., Ueno, M. & Tazaki, H. Natural killer cell activity and recycling capacity of natural killer cells in patients with carcinoma of the prostate.. The Keio journal of medicine 38, 27–35 (1989). [CrossRef]
- Lahat, N., Levin, A. B., Moskovitz, R. D. & B. The relationship between clinical stage, natural killer activity and related immunological parameters in adenocarcinoma of the prostate. Cancer Immunology, Immunotherapy 28, 208– 212 (1989).
- Wu, J. D. et al. Prevalent expression of the immunostimulatory MHC class I chain-related molecule is counteracted by shedding in prostate cancer. Journal of Clinical Investigation 114, 560–568 (2004). [CrossRef]
- López-Vázquez, A. et al. Protective effect of the HLA-Bw4I80 epitope and the killer cell immunoglobulin-like receptor 3DS1 gene against the development of hepatocellular carcinoma in patients with hepatitis C virus infection. The Journal of infectious diseases 192, 162–165 (2005). [CrossRef]
- Middleton, D. et al. Analysis of KIR gene frequencies in HLA class I characterised bladder, colorectal and laryngeal tumours. Tissue antigens 69, 220–226 (2007).
- Portela, P. et al. Analysis of KIR gene frequencies and HLA class I genotypes in prostate cancer and control group.International Journal of Immunogenetics 39, 423–428 (2012). [CrossRef]
- Lin, S. et al. Natural killer cells suppress enzalutamide resistance and cell invasion in the castration resistant prostate cancer via targeting the androgen receptor splicing variant 7 (ARv7).. Cancer letters 398, 62–69 (2017). [CrossRef]
- Kantoff, P. W. et al. Sipuleucel-T immunotherapy for castration-resistant prostate cancer. N Engl J Med 363, 411–433 (2010).
- Jähnisch, H. et al. Dendritic cell-based immunotherapy for prostate cancer. Clin Dev Immunol 517493–517493 (2010).
- Wirth, M., Schmitz-Dräger, B. J. & Ackermann, R. Functional properties of natural killer cells in carcinoma of the prostate. The Journal of urology 133, 973–978 (1985). [CrossRef]
- Yuan, H. et al. Destructive impact of t-lymphocytes, NK and mast cells on basal cell layers: implications for tumor invasion. BMC Cancer 13, 258–258 (2012). [CrossRef]
- Pello, O. M. et al. Role of c-MYC in alternative activation of human macrophages and tumor-associated macrophage biology. Blood 119, 411–432 (2012). [CrossRef]
- Comito, G. et al. Cancer-associated fibroblasts and M2-polarized macrophages synergize during prostate carcinoma progression. Oncogene.
- Dufresne, S. Exercise training improves radiotherapy efficiency in a murine model of prostate cancer. FASEB journal : official publication of the Federation of American Societies for Experimental Biology 34, 4984–4996 (2020).
- Barkin, J., Rodriguez-Suarez, R. & Betito, K. Association between natural killer cell activity and prostate cancer: a pilot study. Can J Urol 24, 8708–8721 (2017).
- Tarle, M., Kraljic’, I. & Kaštelan, M. Comparison between NK cell activity and prostate cancer stage and grade in untreated patients: correlation with tumor markers and hormonal serotest data. Urological Research 21, 17–21 (1993). [CrossRef]
- Hood, S. P. et al. Identifying prostate cancer and its clinical risk in asymptomatic men using machine learning of high dimensional peripheral blood flow cytometric natural killer cell subset phenotyping data. 9 (2020).
- Tae, B. S. et al. Can natural killer cell activity help screen patients requiring a biopsy for the diagnosis of prostate cancer? International Brazilian Journal of Urology : official journal of the Brazilian Society of Urology 46, 244–252 (2020). [CrossRef]
- Vidal, A. C. et al. Natural killer cell activity and prostate cancer risk in veteran men undergoing prostate biopsy. Cancer epidemiology 62, 101578–101578 (2019). [CrossRef]
- Barkin, J., Rodriguez-Suarez, R. & Betito, K. immunotherapies can provide opportunities for evaluating human immune responses. neoadjuvant trials with jnci. oxfordjournals.org JNCI | Article 8, 8708–8721 (2017).
- Song, W. et al. The clinical usefulness of natural killer cell activity in patients with suspected or diagnosed prostate cancer: an observational cross-sectional study. OncoTargets and therapy 2018, 3883–3889 (2018). [CrossRef]
- Coca, S. et al. The prognostic significance of intratumoral natural killer cells in patients with colerectal carcinoma. Cancer 79, 2320–2328 (1997).
- Zorko, N. A., Makovec, A. & Elliott, A. Natural Killer Cell Infiltration in Prostate Cancers Predict Improved Patient Outcomes. Prostate Cancer Prostatic Dis (2024). [CrossRef]
- Tuong, Z., Kelvin et al. Resolving the immune landscape of human prostate at a single-cell level in health and cancer. Cell Reports 37. [CrossRef]
- Vranova, J. et al. The evolution of rectal and urinary toxicity and immune response in prostate cancer patients treated with two three-dimensional conformal radiotherapy techniques. Radiation Oncology 6, 87–87 (2011). [CrossRef]
- Lu, Y. et al. Lower postoperative natural killer cell activity is associated with positive surgical margins after radical prostatectomy. Journal of the Formosan Medical Association = Taiwan yi zhi 119, 1673–1683 (2020). [CrossRef]
- Blomgren, H. et al. Natural killer activity in peripheral lymphocyte population following local radiation therapy. Acta radiologica. Oncology 19, 139–143 (1980). [CrossRef]
- Jochems, C. et al. A combination trial of vaccine plus ipilimumab in metastatic castrationresistant prostate cancer patients: immune correlates. Cancer Immunology, Immunotherapy 63, 407–418 (2014). [CrossRef]
- Sharma, J. et al. Elevated IL-8, TNF-α, and MCP-1 in men with metastatic prostate cancer starting androgen-deprivation therapy (ADT) are associated with shorter time to castration-resistance and overall survival. The Prostate 74, 820–828 (2014).
- Hansen, T. et al. Correlation between natural killer cell activity and treatment effect in patients with disseminated cancer. Journal of Clinical Oncology 36, 12029–12029 (2018). [CrossRef]
- Yirga, A. et al. Monocyte counts and prostate cancer outcomes in white and black men: results from the SEARCH database. Cancer Causes Control CCC 32, 189–97 (2021). [CrossRef]
- Hayashi, T. et al. Serum monocyte fraction of white blood cells is increased in patients with high Gleason score prostate cancer. Oncotarget 8, 35255–61 (2017). [CrossRef]
- Shigeta, K. et al. High absolute monocyte count predicts poor clinical outcome in patients with castration-resistant prostate cancer treated with docetaxel chemotherapy. Ann Surg Oncol 23, 4115–4137 (2016). [CrossRef]
- Angel Charles, Ryan M. Thomas,The Influence of the microbiome on the innate immune microenvironment of solid tumors, Neoplasia, Volume 37, 2023, 100878, ISSN 1476-5586. [CrossRef]
- Johnson, L. E., Olson, B. M. & Mcneel, D. G. Pretreatment antigen-specific immunity and regulation - association with subsequent immune response to anti-tumor DNA vaccination. Journal for Immunotherapy of Cancer 5 (2017). [CrossRef]
- Madan, R. A. et al. Clinical and immunologic impact of short-course enzalutamide alone and with immunotherapy in non-metastatic castration sensitive prostate cancer. J Immunother. Cancer 9 (2021). [CrossRef]
- Kim, S. J., Park, M., Choi, A. & Yoo, S. Microbiome and Prostate Cancer: Emerging Diagnostic and Therapeutic Opportunities. Pharmaceuticals 2024. [CrossRef]
- Inamura, K. Oral-Gut Microbiome Crosstalk in Cancer. Cancers 2023, 3396–3396. [CrossRef]
- Park, S. Y. et al. Oral-Gut Microbiome Axis in Gastrointestinal Disease and Cancer. Cancers 2124–2124 (2021).
- Chiesa, D. et al. Human NK cell response to pathogens. Semin Immunol 26, 152–60 (2014).
- Ljunggren, H. G. & Kärre, K. In search of the “missing self”: MHC molecules and NK cell recognition. Immunol Today. 11, 237–281 (1990).
- Black, A. et al. PLCO: Evolution of an Epidemiologic Resource and Opportunities for Future Studies. Rev. Recent. Clin. Trials 10, 238–245 (2015).
- Prakash, P., Verma, S. & Gupta, S. Gut Microbiome and Risk of Lethal Prostate Cancer: Beyond the Boundaries. Cancers 2023. [CrossRef]
- Nearing, J. T., Declercq, V. & Langille, M. G. Investigating the oral microbiome in retrospective and prospective cases of prostate, colon, and breast cancer. Biofilms Microbiomes 9, 23–23 (2023). [CrossRef]
- Silva, A. P. B. D., Alluri, L. S. C., Bissada, N. F. & Gupta, S. Association between oral pathogens and prostate cancer: Building the relationship. Am. J. Clin. Exp. Urol 7, 1–10 (2019).
- Alluri, L. S. C. et al. Presence of Specific Periodontal Pathogens in Prostate Gland Diagnosed with Chronic Inflammation and Adenocarcinoma. 17742–17742 (2021). [CrossRef]
- Zha, C. et al. Potential role of gut microbiota in prostate cancer: Immunity, metabolites, pathways of action? Front. Oncol 1196217–1196217 (2023). [CrossRef]
- Reichard, C. A. et al. Gut Microbiome-Dependent Metabolic Pathways and Risk of Lethal Prostate Cancer: Prospective Analysis of a PLCO Cancer Screening Trial Cohort. Cancer Epidemiol. Biomark. Prev 2022, 192–199. [CrossRef]
- Liss, M. A. et al. Metabolic Biosynthesis Pathways Identified from Fecal Microbiome Associated with Prostate Cancer. Eur. Urol 74, 575–582 (2018). [CrossRef]
- Zhong, W., Wu, K. & Long, Z. Gut dysbiosis promotes prostate cancer progression and docetaxel resistance via activat- ing NF-κB-IL6-STAT3 axis. 10, 94–94 (2022).
- Golombos, D. M. et al. The role of gut microbiome in the pathogenesis of prostate cancer: A prospective, pilot study. Urology 111, 122–128 (2018). [CrossRef]
- Yue, S.-Y. Causality investigation among gut microbiota, immune cells, and prostate diseases: a Mendelian randomiza- tion study. Frontiers in microbiology 15, 1445304–1445304 (2024).
- Alanee, S. et al. A prospective study to examine the association of the urinary and fecal microbiota with prostate cancer diagnosis after transrectal biopsy of the prostate using 16sRNA gene analysis. Prostate 79, 81–87 (2019).
- Daisley, B. A. et al. Abiraterone acetate preferentially enriches for the gut commensal Akkermansia muciniphila in castrate-resistant prostate cancer patients. Nat. Commun 4822–4822 (2020).
- Liu, Y. & Jiang, H. Compositional differences of gut microbiome in matched hormone-sensitive and castration-resistant prostate cancer. Transl. Androl. Urol 2020, 1937–1944. [CrossRef]
- Matsushita, M. et al. The gut microbiota associated with high-Gleason prostate cancer. Cancer Sci 112, 3125–3135 (2021). [CrossRef]
- Pernigoni, N. et al. Commensal bacteria promote endocrine resistance in prostate cancer through androgen biosynthesis. Science 374, 216–224 (2021). [CrossRef]
- Yu, S. H. & Jung, S. I. The potential role of urinary microbiome in benign prostate hyperplasia/lower urinary tract symptoms. Diagnostics 2022 12, 1862–1862. [CrossRef]
- Sfanos, K. S., Yegnasubramanian, S., Nelson, W. G. & Marzo, A. M. D. The inflammatory microenvironment and microbiome in prostate cancer development. Nat. Rev. Urol 15, 11–24 (2018). [CrossRef]
- Yow, M. A. et al. Characterisation of microbial communities within aggressive prostate cancer tissues. Infect. Agent. Cancer 12, 4–4 (2017). [CrossRef]
- Nelson, D. E. et al. Bacterial communities of the coronal sulcus and distal urethra of adolescent males. PLoS ONE 7, 36298–36298 (2012). [CrossRef]
- Jayalath, S. & Magana-Arachchi, D. Dysbiosis of the human urinary microbiome and its association to diseases affecting the urinary system. Indian. J. Microbiol 2022, 153–166. [CrossRef]
- Yu, H. et al. Urinary microbiota in patients with prostate cancer and benign prostatic hyperplasia. Arch. Med. Sci 11, 385–394 (2015). [CrossRef]
- Shrestha, E. et al. Profiling the urinary microbiome in men with positive versus negative biopsies for prostate cancer. J. Urol 199, 161–171 (2018). [CrossRef]
- Hurst, R. et al. Microbiomes of urine and the prostate are linked to human prostate cancer risk groups. Eur. Urol. Oncol. 2022, 412–419.
- Ravich, A. & Ravich, R. A. Prophylaxis of cancer of the prostate, penis, and cervix by circumcision. N Y State J Med. 51 (1951).
- De Martel, S. & Franceschi. Infections and cancer: established associations and new hypotheses. Crit Rev Oncol Hematol (2009).
- Miyake, K. et al. Mycoplasma genitalium Infection and Chronic Inflammation in Human Prostate Cancer: Detection Using Prostatectomy and Needle Biopsy Specimens. Cell (2019).
- Cavarretta, R. et al. Microbiome of the Prostate Tumor Microenvironment. European Urology 72, 625–631 (2017).
- Zedan, A. H., Hansen, T. F. et al. Natural killer cell activity in metastatic castration resistant prostate cancer patients treated with enzalutamide.. Sci Rep (2023). [CrossRef]
- Kastelan, M., Kraljic’, I. & Tarle, M. NK cell activity in treated prostate cancer patients as a probe for circulating tumor cells: hormone regulatory effects in vivo. Prostate 21, 111–131 (199). [CrossRef]
- Riley, R. D. et al. Reporting of prognostic markers: current problems and development of guidelines for evidence-based practice in the future. British Journal of Cancer 88, 1191–1198 (2003). [CrossRef]
- Fanijavadi, S.; Thomassen, M.; Jensen, L.H. Targeting Triple NK Cell Suppression Mechanisms: A Comprehensive Review of Biomarkers in Pancreatic Cancer Therapy. Int. J. Mol. Sci. 2025, 26, 515. [CrossRef]
Figure 1.
NK-Microbiota Interaction in PCa. This figure illustrates the intricate interactions between various microbiota (oral, gut, urinary, and tumor-associated) and their influence on NK cells in prostate cancer (PCa). The different microbiota communities affect each other: for example, oral microbiota can impact the gut, urinary, and tumor microbiota, while tumor-associated microbiota can also alter the other microbiomes. Dysbiosis, or microbial imbalance, can suppress NK cell function by reducing NK cell frequency, cytotoxicity, and tumor infiltration, impairing immune responses and contributing to tumor initiation, progression, and treatment resistance. On the other hand, a balanced microbiota supports NK cell activity, enhancing tumor surveillance and immune response (Biorender).
Figure 1.
NK-Microbiota Interaction in PCa. This figure illustrates the intricate interactions between various microbiota (oral, gut, urinary, and tumor-associated) and their influence on NK cells in prostate cancer (PCa). The different microbiota communities affect each other: for example, oral microbiota can impact the gut, urinary, and tumor microbiota, while tumor-associated microbiota can also alter the other microbiomes. Dysbiosis, or microbial imbalance, can suppress NK cell function by reducing NK cell frequency, cytotoxicity, and tumor infiltration, impairing immune responses and contributing to tumor initiation, progression, and treatment resistance. On the other hand, a balanced microbiota supports NK cell activity, enhancing tumor surveillance and immune response (Biorender).
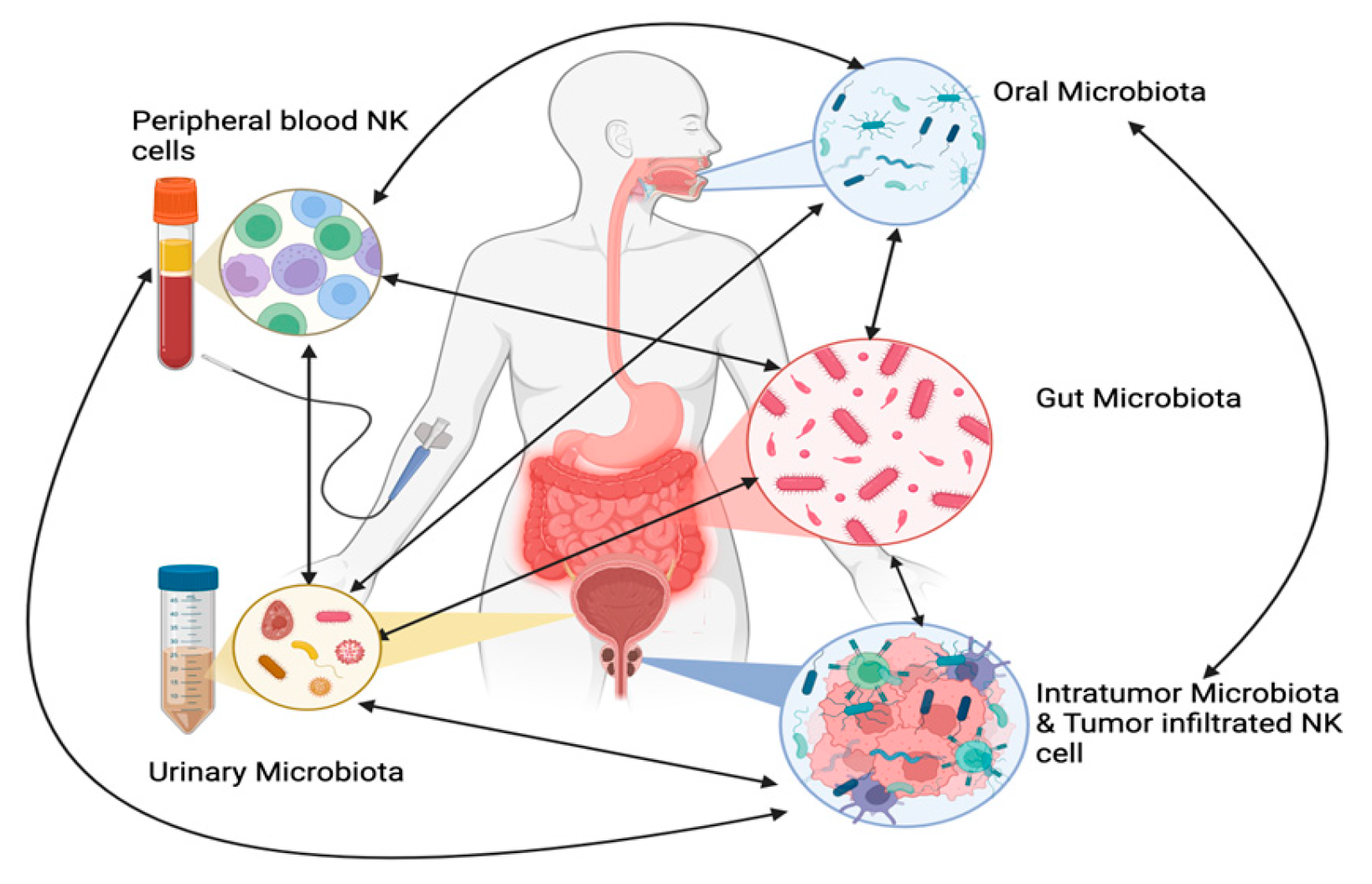

Table 1.
NK cells and Pathogenesis.
| Year/Reference | No/Population | Sample/Methods | Main Findings |
|---|---|---|---|
| 2021/ [69] | 35/Local, Local advanced PCa 27/Control |
PB, Tumor tissue/ Flowcytometry |
In comparison to the control group, tumor-associated NK cells from PCa patients exhibited significantly elevated mRNA expression levels of pro-inflammatory and pro-angiogenic factors. Additionally, these NK cells showed a marked downregulation of NKG2D. Overall, NK cells in PCa patients adopted a pro-inflammatory and pro-angiogenic |
| 2018/ [68] | 10/PCa 10/Control |
Tumor tissue Isolated lymphocyte/ Flowcytometry |
Compared to normal tissue, lymphocytes isolated from PCa showed significantly higher levels of miR-224, which plays a protective role for cancer cells against NK cell-mediated destruction. HIF-1α was found to upregulate miR-224, leading to the inhibition of NCR1/NKp46 and reducing the killing efficiency of NK cells in prostate cancer. |
| 2016/ [18] |
16 /Localized PCa 5 /mPCa 20 /Normal tissue from patients with PCa |
PB, Tumor and normal tissue/ Flowcytomery, 51-Cr release |
A highly immunosuppressive environment in PCa significantly impairs NKA at multiple levels. |
| 2015/ [44] | 18/ Treatment naive PCa, de novo: 10 /SCR 8/ LCR 10 / Control |
PM/ Flow cytometry |
The low expression of activating receptors and reduced cytotoxicity of NK cells contribute to the pathogenesis of Pca. |
| 2014/ [70] | 18/CRPC 8 Control |
PM, Isolated lymphocyte/ Flow cytometry |
Prostate tumor-derived exosomes act as downregulators of the NKG2D-mediated cytotoxic response. |
| 2013/ [15] | 51/ PCa treatment naive 54/ Healthy control |
PMBC/NK Vue |
PCa patients exhibit a higher CD56^dim^ to CD56^bright^ NK cell ratio compared to the control group. |
| 2013/ [81] | 100 PCa 150 BC |
Tumor tissue/Immunohistochemistry | NK cell infiltration in the early stages of PCa is associated with focal disruption of the tumor capsule. |
| 2012/ [76] | 200 Untreated localized Pca 185 Healthy control |
Blood sample (DNA)/PCR | There is no evidence of a potential role for the KIR gene system in PCa. |
| 2011/ [3] | 20 Localized, local advanced Pca 20 BPH 20 Healthy control |
PBMC/Flow cytometry |
The percentage of NK cells and their subsets did not vary among the groups examined. |
| 2004/ [73] | 23/ PCa 10/ Healthy control |
Serum/ Cell line Immunohistochemical staining |
Advanced cancer is associated with high expression of MICs and low NKA. |
| 1989/ [72] | 49 Localized and advanced PCa 15 Healthy controls |
PB, Cell line /Flowcytometry |
Aberrant immune functions in the early stages, with an exacerbation of these immune abnormalities observed in advanced PCa. . |
| 1989/ [71] | 7 Localized PCa 6 Advanced PCa 25 Control |
PB, Cell line/ 51-Cr release | MRC in advanced PCa was lower than in localized PCa and the control group. |
| 1987/ [16] | 10 Localized PCa 49 Advanced PCa 10 BPH 20 Control |
PB, Cell line/ 51-Cr release |
Compared to other groups, patients with advanced PCa demonstrated significantly lower NKA, which was not attributed |
| 1985/ [80] | 8 Pca T2N0M0 8 PCa T3-T4NxM0-1 7 Control |
PB, Cell line/ 51-Cr release | Compared to the control group, beta interferon-induced augmentation of NKA was significantly reduced in the periprostatic lymph nodes of PCa patients. Additionally, the spontaneous cytotoxicity of peripheral NK cells was markedly suppressed in advanced PCa. |
Maximal Killing Potential (Vmax); Maximal Recycling Capacity (MRC); Long Castration Response (LCR); Long Castration Response (LCR); Peripheral Blood (PB); Prostate Cancer (PCa); Metastatic Prostate Cancer (mPCa); Castration-resistant Prostate Cancer (CRPC); Breast Cancer (BC); Benign Prostatic Hyperplasia (BPH); Natural Killer Activity (NKA); Time to Castration Resistance (TCR); Overall Survival (OS); Killer Cell Immunoglobulin- like Receptor (KIR); MHC class I c molecules (MICs).
Table 2.
NK cell and diagnosis/Screening.
| Year/Reference | No/Population | Sample/method | Main Finding |
|---|---|---|---|
| 2020/ [87] | 41 Asymptomatic PCa (PSA<20ng/ml) 31 Control (Benign) |
PB/ Flow cytometric profiling combined to Machine learning |
NK cell profiling identified 8 features of NK cells, which distinguishe between PCa and benign disease. NK cell profiling could serve as a potential screening tool for prostate cancer detection. |
| 2020/ [88] | 18 PCa, GS>7 8 PCa, GS=7 24 PCa, GS=6 52 PCa, Benign |
PB/NK Vue | NKA demonstrated a sensitivity of 68% and a specificity of 73% for detecting PCa, with a cut-off value of 500 pg/mL. |
| 2019/ [89] |
25 Localized PCa 37 Local advanced, advanced 32 Not detected in biopsies. |
PB/NK Vue |
Low NKAcan serve as a predictive tool for positive prostate cancer biopsy, with a cut-off value of 200 pg/mL |
| 2018/ [91] |
221/ Pca 18 GS 9-10 25 GS 8 49 GS 7 43 GS 6 86/ Biopsy negative |
PB/NK Vue | The study could not confirm the usefulness of NKA for detecting PCa or predicting the Gleason grade. |
| 2017/ [90] | 21 /PCa 22/ Biopsy negative |
PB/NK Vue Tissue |
The researchers of the study showed that the absolute risk for PCa is 86% when NKA is low. |
| 2014/ [] |
8/ mPCA 36/ mBC 30/ mCRC |
PB/ Flow Cytometry |
High levels of circulating tumor cells (CTCs) are associated with low NKA levels. |
| 2013/ [15] | 51/ PCa 8 GS=9 8 GS=8 25 GS=7 10 GS=6 54/Biopsy negative |
PB/NK Vue | NKA demonstrates a sensitivity of 72% and specificity of 74% for detecting PCa, with a cut-off value of 530.9 pg/mL. |
| 2004/ [73] | 23/ PCa 10/ Healthy control |
Serum/ ELISA | sMIC is proposed as a novel biomarker for the detection of high-grade PCa. |
| 1993/ [86] | 23/ Localized and advanced PCa 10/Healthy control 6 Chronic diseases |
PB/51-Cr | NK lytic activity in patients with untreated PCa effectively differentiates between tumor dissemination and localized PCa. |
| 1992 | 51 mPCa: 6 DES 11 Estracyt 9 CPA 7 Orchiectomy 12 Orchiectomy +CPA 6 Orchiectomy +Flutamide 7 Healthy control |
PB/51-Cr | Low NKA levels in treated PCa patients suggest the presence of CTCs monitoring for treatment response diagnostic application. |
| 1987/ [16] | |||
| 1985/ [80] |
Prostate Cancer (PCa), Metastatic Prostate Cancer (mPCa), Metastatic Breast Cancer (mBC), Metastatic Colorectal Cancer (mCRC), Peripheral Blood (PB), Castration-resistant prostate cancer (CRPC), Benign Prostatic Hyperplasia (BPH), Gleason sum (GS); Natural Killer Activity (NKA), circulating tumor cells (CTCs), Soluble MHC class I chain–related molecules (MICs).
Table 3.
NK cell and Prognosis/Prediction.
| Year/Reference | No/Population | Sample/Methods | Main Findings |
|---|---|---|---|
| 2024/ [93] | 3365 samples of mPCa 87551 samples of 44 distinct tumor type |
FFPE/ quanTIseq |
NK cell infiltration lead to improved Pca patient outcomes. |
| 2023/ [141] | 87 mCRPC |
PB NK vue |
In the late stage mCRPC patients receiving enzalutamide, higher NKA levels correlated with poorer treatment responses suggesting that NK cells may negatively impact advanced prostate cancer and could serve as predictive markers for treatment efficacy. |
| 2020/ [96] | 24 negative surgical margin 27 positive margin 10 healthy |
PB NK vue |
Higher postoperative NKA levels are associated with an increased likelihood of negative surgical margins, indicating better prognosis. |
| 2018/ [100] | 19 mPCa 23 mOCa 51 mCRC |
PB NK Vue |
Lower NKA levels correlate with reduced response rates and shorter PFS. An increase from low to normal NKA, highlighting its predictive potential. |
| 2017/ [105] | 22 non metastatic non castrate 16 CRPC |
PB FlowCytometry |
The fr The frequency of NK cells does not differ between immune responders and non-responders following vaccination, suggesting limited predictive value. |
| 2015/ [44] | 18 Treatment naive PCa, de novo: 10 SCR 8 LCR 10 Control |
PB FlowCytometry |
NKp30 NKp30 and NKp46 are predictor markers for OS with p-values of 0.0018 and 0.0009, respectively. p-vaof. TCR also demonstrates significance with p-values of 0.007, 0.009, and 0,0001, indicating their prognostic and predictive potential (log-rank test). |
| 2014/ [98] | 30 mCPC | PB Flow Cytometry |
High High lev High levels of Tim-3(+) NK cells after vaccination, compared to before vaccination, are associated with longer OS, indicating their prognostic significance. |
| 2014/ [99] | 72 mPCa before ADT or no longer than 3 months 50 mPCa ADT +/- Risedronate |
Serum Multiplex electrochemiluminescence |
Higher cytokine levels observed 0.5 months after the initiation of ADT are associated with shorter TCR and OS, potentially indicating resistance to treatment, with no relation to IL-2 levels. This highlights no prognostic and predictive value of IL-2 as an NK cell activator |
| 2014/ [19] | 8 mPCa 36 mBC 30 mCRC |
PB CellSearch CTC test |
High CTCs represent low NKA and high risk of poor prognosis. |
| 2013/ [15] | 51 Pca no prior treatment 54 Healthy control |
PB NK Vue |
A higher CD56dim/CD56bright ratio or lower levels of CD56bright cells are associated with poor prognosis. |
| 2012/ [81] | 100 PCa, 150 BC |
Tumor Tissue IHC |
NK cell infiltration in the early stage PCa is linked to focal disruptions of the tumor capsule and poor prognosis. |
| 2011/ [95] | 197/PCa, 3DRT 116 T1-T4 WP 81 T1-T4 PO |
PB Flowcytometry |
A positive correlation exists between NK cell numbers and bone marrow irradiated with 5-25 Gy, as well as with acute GU, late GU, and GI toxicities. This upregulation of NK cytotoxicity is associated with better prognosis and predictive outcomes. |
| 2009/ [1,24] | 35 PCa neoadjuvant ADT+RT 40 Pca RP |
Tumor tissue IHC |
Higher levels of CD56+ NK cells are associated with a lower risk of PCa progression, indicating their potential prognostic application. |
| 2004/ [73] | 23 Localized and advanced PCa 10 Healthy donor |
Serum ELISA |
Higher levels of sMIC are associated with poor prognosis, highlighting its potential prognostic application. |
| 1995 [33] | 11 Localized PCa, RP, TURP 41 Advanced, Castration (medical or surgical) 10 BPH |
Tumor tissue |
Higher expression of HNK-1 antigen is associated with longer survival and PFS, indicating its prognostic and predictive value. |
| 1992/ [140] | |||
| 1989/ [71] | 7 Localized PCa 6 Advanced PCa 25 Healthy control |
PB, cell line K562 | NKA, Vmax, and MRC levels in advanced PCa are lower than those in localized PCa and control groups, indicating potential prognostic significance. |
| 1980/ [97] | 6 PCa 7 UBC 24 BC |
PB/51-Cr release | NKA is positively associated with prognosis, indicating its potential prognostic value. |
Prostate Cancer (PCa); Metastatic Prostate Cancer (mPCa); Breast Cancer (BC); Metastatic Colorectal Cancer (mCRC); Metastatic Ovarian Cancer (mOCa); Urinary Bladder Cancer (UBC); Peripheral Blood (PB); Castration-Resistant Prostate Cancer (CRPC); Natural Killer Activity (NKA); Human Natural kille-1 (HNK-1); Circulating Tumor Cells (CTCs); Soluble MHC class I Chain–related molecules (sMIC); Genitourinary (GU); Gastrointestinal (GI); NKA ratio (postoperative NKA/preoperative NKA); Radiotherapy (RT); Robot-Associated Radical Prostatectomy(RARP); Whole pelvic (WP); Prostate-only (PO); 3-Dimenshional Conformal Radiotherapy (3DRT);Radical prostatectomy(RP); Transurethral Resection of the Prostate (TURP); Overallsurvival(OS).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



