
Submitted:
07 January 2025
Posted:
08 January 2025
You are already at the latest version
Abstract
Rhizobiome is a network of beneficial microbes (fungi, bacteria, nematodes, actino-mycetes, mycorrhizae) and to study their interaction is fascinating in producing higher crop yield and increase resistance against phytopathogens. The interactions could be competitive, neutral, commensal and mutualistic. Beneficial interactions chiefly colo-nize roots and maximize plant growth parameters like plant height, leaves number & fruit/pod/seed number. The positive microinoculants fix nitrogen, solubilize P and Fe, produce phytohormones, improve plant defense chemicals and enzyme machinery in root tissues upon pathogen invasion. The rhizomicroinoculants also increase photo-synthetic and nutritional proficiency by maximizing total chlorophyll, carotenoid, carbohydrates, amino acids and proteins in plants. They play a crucial role in pre-venting disease outbreak under stressful situations like drought, metal stress, salinity, high temperature, soil erosion, change in rainfall and humidity. Understanding rhi-zomicrobiome interaction and preparing novel rhizospheric microbial formulations could be good strategy to improve plant growth parameters, fruit yield and defense ability towards pathogen.
Keywords:
beneficial microbes
; rhizobiome
; root interactions
; plant growth
; plant resistance
1. Introduction
Root is a vital organ which absorb minerals and transport water to aerial parts of the plants. The root structural information is important to know and build beneficial microflora in the vicinity of root activity. The occurrence of soil microbes varies and is influenced by root exudates, chemical components, pH value and edaphic factors. Rhizobiome is a zone where group of microbes interacts with one another. The two main kinds of microbial interactions prevail in rhizobiome include microbe to microbe and microbe to plant. Microbes communicate and compete continuously with each other for space and nutrients to form population and spread colony in rhizobiome [1]. A microbe can antagonise the growth of other microbe to support plant growth or in combination they produce toxins to form infection symptoms in plants. Ustilago maydis (biotrophic pathogen) inhibited wilt pathogen Fusarium verticillioides to support the growth of maize plants [6]. We can observe that, microbe [Albugo laibachii] of one host plant type [A. thaliana] produce infection symptoms and prevent the growth of non-host pathogens [Phytophthora infestans] [2]. Crude elicitors (oligosaccharides, proteins, and sphingolipids) extracted from pathogenic microbe (C. capsici) can induce plant growth and resistance in chilli plants [109]. Communication can be stopped by inhibiting AHL lactone enzyme in pathogenic microbes by rhizobacteria [4]. Trichoderma spp. and Bacillus amyloliquefaciens negatively impact on soil borne pathogen (F. oxysporum) colony to prevent wilt infection and improve growth in tomato plants [108]. Various organic amendments were applied in soil in the form of compost, manure and plant extracts to increase microbial functionality and soil fertility to produce higher crop yield. For example: Phytophthora capsici positively support the B. subtilis, T. gamsii and T. asperellum growth when compost is drenched in rhizobiome and negatively foster fruit rot/ blight disease severity in zucchini plants [107]. Certain microbes particularly viruses whose virulency is much higher that it will overcome the beneficial effects of useful microbes and prevent nitrogen accumulation in plants. Higher application of urea negatively affects virus and bacterial colony growth [81]. It was interestingly observed that, the root exudate trehalose secreted by leguminous plants helps in the colonization of rhizobia and mycorrhizae which increase growth potency and drought tolerance ability in leguminous plants [5]. Useful root resident microbes secrete secondary metabolites/phytohormones to prevent the growth and division of soil borne pathogens [67]. Overall microbe-microbe interaction is dependent on soil factors, habitat, host plant type, root exudates and environmental calamities.
Microbe to plant interaction may be saprophytic or parasitic. Mostly the interactions are saprophytic. The positive or negative interaction is dependent on the amount of rainfall, light intensity, soil moisture, temperature, soil type, mineral availability, root structure and root health [1]. The beneficial fungal colony namely Penicillium, Aspergillus sp., Talaromyces and Trichoderma interacts with plant roots to promote fruit setting in chilli, water melon and cucumber plants [8,10,75]. Certain mycorrhizae and free nitrogen fixing bacteria (Bradyrhizobium japanicum) lies in soil to stimulate tomato plant growth [71]. Priming treatments with Trichoderma sp. prevent over ripening or premature-ripening of melon fruits by declining ethylene concentration and increasing ACC deaminase [11,104]. The information from transcriptome, genomic and proteomic analysis could be useful in identification of organic acids, fatty acids, chemotaxis and stess tolerant gene from beneficial microbes in order to improve yield, height, nutritional parameters and antagonise noxious pathogen growth and division [12,23,24,64,82]. The mechanism behind rhizobacteria and rhizofungi in influencing plant productivity and plant health is by phytohormones production, siderophore production, complex phosphorous, mineral solubilization, lignin, callose and phenol production, PAL, POX, PPO and other antioxidant enzyme production [SOD, CAT] against different biotic, abiotic stressors [7,8,46]. Beneficial microbes were added manually to soil roots chiefly to improve farming methods, increase plant productivity and prevent plants from harmful effects of agrochemicals. The challenging aspect of using biocontrol agents as inducers lies in mass producing and marketing them as biopesticides, bioherbicides and biofertilizers. Henceforth, beneficial microbes’ recognition and their interactions with soil microflora is most important for choosing positive strains to develop novel bioformulations that can stimulate growth and manage diseases in plants.
2. Microbial Interactions with Plants Roots and Their Beneficial Roles
Plant diseases pose serious threat to global food scarcity and cause malnutrition problems. In order to address the problem, beneficial microbial inoculants serve as good source of inducer to achieve sustainable crop yield and manage plant diseases. Microbial colonies vary based on soil type, climatic factor, host plant and kind of interaction. Beneficial interactions of P. fluorescens reduced F. monoliforme infection, induce maize plants growth and improve mineral cycling by phosphate and zinc solubilization [58]. Rhizospheric fungus Trichoderma virens [ZT05] enhances root length (25.11), root surface area (98.19), average root diameter (5.66), root tip number (45.89) and root branch number (74.42%) in Pinus sylvestris var. mongolica plants [15]. Aspergillus ustus promote growth by secondary roots formation and protected A. thaliana plants from infectious pathogens [16]. Different strains of Chaetomium globosum increase IAA, GA production and copper tolerance to improve growth and sustainable capacity in chilli and maize plants [18,19]. Parasitic fungus Sphaerodes mycoparasitica and rhizospheric fungus-T. harzianum treatments increased seed germination %, total plant biomass and suppressed infection of Fusarium graminearum, Pythium ultimum and Colletotrichum graminicola in wheat and maize plants under phytotron conditions [20,35]. Penicillium spp. increase photosynthetic efficiency and nutritional parameters like chlorophyll content, lignans, amino acids and proteins in Sesamum indicum [17]. Application with T. harzianum, Phoma multirostrata, and Penicillium chrysogenum in tomato plants maximized ACC deaminase levels and nitrogen uptake in plants [21]. Fusarium oxysporum prevented nematodes [Pratylenchus goodeyi, Helicotylenchus multicinctus] growth and the priming treatments were also promising in improving banana fruit yield [22]. Mycorrhizae like G. mosseae, G. etunicatum, G. claroideum, G. microaggregatum, G. geosporum and R. irregularis improve fruit yield, plant weight, water uptake and leaf water potential in tomato plants [14]. Funneliformis mosseae application augment growth parameters like fresh weight, dry weight, ca concentration and lowered oxidative damage in tomato plants [13]. Transcriptome studies revealed that mycorrhizal inoculation in Medicago truncatula roots increased aquaporins, ATPases, metal adherence, proteins, mineral transporter, and osmoregulatory proteins in root nodules [24]. Rhizomicrobial interactions improve soil health by scavenging toxic metals and toxic pesticides-Fenvalerate, Chlorpyrifos, Cypermethrin from Stenotrophomonas maltophila, Sphingobacterium thalpophilum plant roots [25]. Talaromyces wortmannii produced terpenoid-like volatile and β-caryophyllene to stimulate turnip plants growth [36]. The flower setting is important for fruit production in all kinds of plants. Penicillium simplicissimum treatment increase size and weight of marigold flower [27]. Free living non-parasitic nematodes improve soil quality by pathogen inhibition, mineral cycling, and controlling soil borne pathogens. The nematodes can sense environmental disturbances and serve as food for microflora to improve soil health [106]. Pochonia chlamydosporia and Piriformospora indica induce premature flowering in A. thaliana, Coleus forskohlii, Lagenaria siceraria and Nicotiana tabacum [30]. T. harzianum, F. equiseti, T. harzianum T22 treatments stimulate ISR in cucumber plants against anthracnose pathogen-C. orbiculare [34]. Certain volatile compounds namely alcohols, alkanes, ketones and terpenoids helps in rhizomicrobial communication in soil [1]. F. oxysporum suppressed Pratylenchus goodeyi and Helicotylenchus multicinctus (nematodes) growth [37]. Saccharothrix yanglingensis induced chitinase production which antagonize Valsa mali reducing apple canker disease infection [40]. Actinomycetales, Microbispora, and Nonomuraea prominently displayed antifungal activity in tomato plants against A. solani, P. parasitica, R. solani, and P. irregulare [42]. Streptomyces albogriseolus produced fungichromin that work competently against nematode infection in banana plants [38]. The actinomycetes and mycorrhizal consortium increased growth parameters like plant height, stem diameter, radical size, leaf area and number and total plant biomass. The bioinoculants antagonise P. capsici pathogen to reduce wilt infection on chilli plants [39]. Bean seedling growth was improved by Cladosporium cladosporioides, Penicillium simplicissimum and Trichoderma sp. [41]. Root resident microbes were potent enough to increase salt tolerance ability by inducing higher level of catalase, PO, SOD, APO, monodehydroascorbate reductase, GR, GST, GPX and ascorbic acid in Brassica juncea plants [26]. Application of Aspergillus versicolor reduced fruit rot symptoms in tomato plants [43] and inoculation of rhizobacteria and rhizofungi prevent toxin accumulation, inhibit root knot pathogen and cyst nematodes [45]. Talc based formulations performed well compared to vermiculite-based formulations in providing better tomato yield [44]. Treatment with Alternaria sp. Aspergillus sp. maximize seedling germination performance of Astragalus utahensis [48]. Treatment with T. longipile and T. tomentosum improved photosynthetic proficiency by 58 to 71% by leaf area expansion in cabbage and lettuce plants [74]. Aspergillus spp. PPA1 and Fusarium spp. PPF1 were amended in soil that maximize chlorophyll accumulation in cucumber and Indian spinach leaves [75,76]. It is suggested that increased in leaf number, area and chlorophyll contents would increase photosynthesis and carbohydrates synthesis in plants [76]. AMF combined treatments with PGPR managed proline and relative water content levels to increase drought stress tolerance level in Melissa officinalis plants [46]. Trichoderma sp., trigger SA, ET and JA signalling defense responses to improve resistance in plants [47,75]. Actinobacteria namely Acidiferrimicrobium, Micrococcus, Arthrobacter sinomonas, Nocardia, Streptomyces, Frankia sp., Rhodococci sp., invade plant roots, expand mycelial hyphae to stimulate growth and defense responses in plants [54]. The cucumber and muskmelon plants sprayed with T. harzianum showed early seed emergence, increase % germination, seedling vigour and % transplantation rate [49,50]. Hypovirulent R. solani strains increased fresh leaf and stem weight in tomato plants under pot culture experiments [51]. Microbispora, Actinomadura, Streptomyces, Mesorhizobium cicero increased grain yield, nitrogen content via nodule formulation and improved root colonization efficiency by flavonoid content synthesis in Cicer aeriatinum plants [55]. Exophiala sp. produced gibberellins to reprogrammed cucumber plants growth under salinity and drought conditions [53]. Streptomyces strains (Rhizobacteria) lowered fungal infection of Dactylonectria sp., Ilyonectria sp., Phaeoacremonium chlamydospora, and Phaeoacremonium sp. in grapevine plants under nursery and field conditions [56]. Micromonospora aurantiaca and Streptomyces griseus increased N, P uptake to promote wheat plants growth and reduce damping-off (Pythium ultimum) infection [85]. Genome analysis of Dermacoccus abyssi identified salt tolerant gene to promote tomato plants growth under halophytic conditions [56]. Thus, beneficial microbes residing in roots help the plant to bear lower water content, scavenge toxic metals from soil and protect plants from adverse natural calamities to finally stimulate higher productivity and flexibility in plants [Figure 1]. The important rhizospheric fungi that promote plant growth and stimulate resistance in various plant crops has been listed in Table 1.
3. Rhizomicrobiome Interaction with Secondary Metabolite, Signalling Proteins and Their Relationship in Plants
Plants secrete secondary metabolites within root region surrounded by soil that can manipulate rhizomicrobiome and have direct role in induction of plant growth promotion and defense responses. To improve soil nutritional efficiency, the survival of beneficial microflora and their colonization ability are the key factors. Root exudates secreted by plants were utilized by beneficial microbes to fulfil nutritional deficiency. Coumarin secreted by plant roots to attract Pseudomonas simiae colony overcome iron deficiency in Arabidopsis plants [59]. Pseudomonas fluorescence JM-1 produced 2,4-Diacetylphloroglucinol to lowered the ear rot infection in Zea mays plants [58]. Certain root secretions help the beneficial microbes to colonize the plant roots and few can affect microbial colonization. For example: Pseudomonas putida residing in rhizophere region secretes benzoxazinoids to establish successful connection with host plants and increase the growth of oxalabacteraceae plant members by making nitrogen available to thrive under stressful conditions [60,61]. Galactoside secretions were utilized by S. meliloti [62]. Interestingly few exudates like Isothiocyanates (ITC) and glucosinolates secretions affect rhizomicrobial growth species namely Pseudomonas syringae, Sphingomonas suberifaciens and Fusarium oxysporum [66]. The plant genotype decides complexity and specificity nature of root exudates which attracts specific beneficial microbes’ interaction. Fatty acid secretion increased Bacillus flexus colonies in Limonium sinense (halophyte) plant [64]. Secreted amino acids help in chemotaxic movement of Pseudomonas protegens [65]. Arthrobacter protophormiae (SA3) and Dietzia natronolimnaea (STR1) improve wheat plants to withstand high salt and low water stress by triggering IAA and ET signalling pathways [67]. During cold stress, rhizobacteria like Funneliformis mosseae and Paraburkholderia graminis lowered ROS generation in tomato plants [68]. Likewise, Trichoderma asperellum induced phytohormone production in cucumber plants that make plants tolerant to saline conditions [69]. Certain rhizobacteria named Bacillus methylotrophicus, Bacillus licheniformis, and Bacillus aryabhattai increase wheat plant capacity to heat stress by reduced RO generation [70]. Bradyrhizobium japonicum (mycorrhizae) activates nod factor that tends to enhance root surface area, root length and lateral root hair number in Arabidopsis thaliana [71]. Caulobacter, Serratia, Flavobacterium, Chromobacterium and Methylobacterium mitigated abiotic stress response in Psoralea corylifolia and Triticum aestivum plants [78]. Funneliformis mosseae increased secondary metabolites, reduce oxidative stress and enzyme activity along with increasing fresh and dry weights. AM fungus inoculation on soybean plants improved salt tolerance ability by targeting StNHX1 gene (Na+ /H+ antiporter gene) [73]. Nicotiana benthamiana produce capsidiol induce defense response by activating ethylene signalling pathway towards Phytophthora infestans [79]. Mesorhizobium ciceri was inoculated with Bacillus sp. and Enterobacter aerogenes increased root nodulation, nitrogen uptake, phosphorous and total protein in chickpea plants [80]. The RNA transcript analysis of Arthrobacter endophyticus and Nocardiopsis alba revealed carotenoid, glycerolipid, phenylalanine production upon salt stress by upregulating chlorophyll a, hydroxyproline rich glycoprotein, cytokinin dehydrogenase and K+ uptake in A. thaliana [82]. An actinomycetes named Streptomyces sp. produced betalain, isoquinoline, alkaloid and zeatin under salt stress in Solanum lycopersicum cv. Jingpeng [83]. The mutation in disease causing gene (mpg1) in M. grisea abrupt appressoria structure and lowered infection symptoms on rice plants [105]. Frankia sp. produced cytosolic glutamine synthetase to metabolise soil nitrogen and promote Datisca glomerata plants growth [84]. The GC-MS analysis helps in secondary metabolites/ root exudates identification and the secretions were linked to functional aspects and variations in A. thaliana plant [29,33]. A proteomic study was performed wherein PR proteins (PR10) helps in the identification of Sinorhizobium meliloti colonization to improve M. truncatula plants roots architecture and the process is regulated by auxin production [23]. Pseudomonas syringae sprayed on Arabidopsis plants induce PR proteins and SA production [32]. RNA transcript analysis revealed that organic acids (fumaric acid) secreted by maize roots was helpful in Bacillus amyloliquefaciens biofilm formation [63]. Utilizing genomic and metabolomic techniques, it was possible to identify antifungal metabolites like surfactin, iturin fengycin, and rhizocticin-A produced from Bacillus cabrialesii and Bacillus amyloliquefaciens effective against spot blotch disease of Triticum turgidum [28,31]. The amino acids, phenol (alanine, arginine, asparagine) and coumarins (cinnamic, salicylic, syringic, ferulic, caffeic acids) play a diverse role in attraction, quorum sensing, symbiotic interaction, rhizomicrobial growth and virulence gene activation. Therefore, identification of root secretions helps in better understanding of root colonization process, secondary metabolite secretions, water and minerals transport, biochemical compounds synthesis, regulatory and stress responsive genes.
4. Saprophytic Microbial Interaction Can Be Beneficial or Parasitic in Rhizosphere
Root exudates released by plants act as signalling molecules which can instigate both pathogenic and mutualistic interactions in rhizosphere. It has been documented that plant roots association helped microbes to establish connection with plants and strengthen plant defense [90]. The root exudates produce sugars (glucose, sucrose, galactose, fructose) and organic acids (malic, lactic, citric and oxalic acids) that serve as C-source which provide energy to increase microbial colonies number. Strigolactones increased plant root length and root hairs. Saprophytic microbes act as decomposers by converting complex organic into smaller simple molecules. They cycled N, P and other micronutrients to make available to plants [91,92]. The saprophytic fungi can act as growth promoters and enhancers of plant defense responses. For eg: Streptomyces ganmycicus, Trichoderma, Gliocladium, Pseudomonas and Bacillus subtilis reduced disease incidence in Rhizoctonia solani in Maize and Soybean plants [92]. Biocontrol agents reduced parasitic fungal infection in roots via antagonistic behaviour and induced ISR in host plants [95]. Beneficial microbes produce nod factors to form nodule in leguminous roots to fix nitrogen and convert into ammonia that can be absorbed by plants [93]. R. leguminosarum (nodule forming bacteria) deliver protection in faba bean plants against mosaic virus infection by decreasing H2O2, MDA contents (oxidative stress indicators) and increasing defense related enzymes [94]. Rhizospheric fungi (T. harzianum, T. asperellum, Aspergillus tubingensis, Penicillium sp., T. funiculosus) increased PAL, POX, Chitinase and β-Glucanase activity. The lignin and callose deposition in chilli plants with C. capsici infection impart resistance towards anthracnose disease. These rhizospheric fungal interactions can increase biochemical parameters like carbohydrates, amino acids, proteins, chlorophyll and chilli fruit yield [8]. Allelopathic plants produce defense chemicals that specifically act on weed plants and this defense strategy is dependent on plant roots secretions. The defense molecules composition, amount and their control mechanism are dependent on plant pathogen interaction [96]. Beneficial microbial diversity in rhizospheric region can be improved by physical methods like crop rotation and intercropping as the plant selects unique bacteria via producing root exudates which interact with plant roots to improve soil fertility and maximum crop yield [97]. AMF mycorrhizae increase phosphorous content via mutualistic association with plants by enzymatic breaking down of insoluble P into soluble form [95]. Thus, the presence of microbial soil flora depends on correct root architecture, root type and niche. The beneficial interactions improve soil health by increasing microbial colony, deep root penetration in soil, protection from soil erosion and adverse climatic factors.
If microbial interaction is incompatible then it will produce infection symptoms that reduce crop yield and resistance towards noxious pathogens. Some saprophytic microbes can be pathogenic/ opportunistic to cause plant diseases [Figure 2]. Aspergillus sp. and Bacillus sp. were plant growth promoters in cotton plants and A. chevalieri, A. egyptiacus were defense enhancer in Vicia faba plants [88,89]. It was observed that rhizomicrobial interaction deliver protection by producing chitinases, glucanases, myrosinases [92]. Alternaria sp., cause early blight, leaf spots, head rot, and black rot disease in plants. But in some plants, some of its species named Alternaria sp. A13 can act as plant growth promoter and improve secondary metabolite [cinnamic acid 4-hydroxylase and lithospermic acid B] contents [86,87]. The root fungi can be parasitic/ saprophytic depending on the presence/absence of host. There will be competition and interference between soil microbes with parasitic and beneficial nature. The type of interaction is decided by root structure, secretions, and living root activities. The mechanism by which microbes interact include vesicle transport, diffusion, and ion channels [91]. Developing variety by designing root architecture with higher root length and more lateral roots to have plasticity towards changing environment conditions. Biochar positively control rhizomicrobial activity and promote good yield [22]. Carbon derived from plant roots can be stored as soil organic matter which depends on the carbon input and decomposition rate. Root resident microbes help in the degradation of organic matter and remove pollutants to improve soil fertility and resilience [87]. Overall, root resident beneficial microbes act as inducers to deliver nutrients for plant growth, tolerance to abiotic stressors and serve as good pathogen inhibitors [98].
5. Conclusions
To achieve sustainable food production, root architecture has to designed in such a way that plants must have long root length, more root hairs and numbers. This could be achieved by artificially inoculating beneficial microflora in soil roots that help the plants to penetrate deeper in soil to absorb vital nutrients chiefly N, P, K and other micro, macronutrients which ultimately lead to higher crop yield. These rhizomicrobiota induce enzymes lignin and callose in roots to improve defensive system and prevent pathogen entry. The beneficial microflora competes naturally with parasitic pathogens for niche and food. The root secretions are dependent on plant species, soil nature and climatic conditions. The soil health can be maintained by increasing soil beneficial flora by adopting crop rotation and intercropping methods. Improving rhizobiome can increase beneficial microflora colony number that help plants to withstand drought, salinity and various stressed conditions. The pathogenicity is decided by low and high molecular weight compounds that play a vital role in plant interaction. Malnutrition and hunger problems are the major issues at present and will increase in more higher rate in the coming years. The main gateway to attain safe, sustainable food production is to develop broad range of rhizomicroformulation that can manage disease causing pathogens, infection symptoms, increase positive soil-root feedback mechanism, improve plants health and productivity. Using scientifically distinct but functionally similar useful strains could be good and promising tactic in near future to design definite disease standards and multi-targeted biocontrol methods.
Author Contributions
The first and corresponding author (Naziya B), came up with the idea, gathered data, and contributed in writing and designing of the review article.
Data Availability Statement
All the information was gathered from authentic research and review articles for the preparation of manuscript.
Acknowledgments
The author would like to thank Department of Collegiate Education, Govt. First Grade College for Women, Vijayanagar, Mysuru for all the assistance in completion of review article. The author specially acknowledges botany department, University of Mysore for all the help and support.
Conflicts of Interest
The author declares that there is no competent interest to publish review article. Also, the review article has not been submitted nor published elsewhere.
Abbreviations
The following abbreviations are used in this manuscript:
| ACC | Aminocyclopropane-1-carboxylate deaminase |
| PGPR | Plant Growth Promoting Rhizobacteria |
| PGPF | Plant Growth Promoting Fungi |
| SOD | Superoxide Dismutase |
| POX/POD | Peroxidase |
| PAL | Phenylalanine lyase |
| PPO | Polyphenol Peroxidase |
| N, P, K | Nitrogen, Phosphorous, Potassium |
| ABA | Abscissic acid |
| SA | Salicylic acid |
| JA | Jasmonic acid |
| H2O2 | Hydrogen peroxide |
| IAA | Indole-3-acetic acid |
| AHLs | Amino homoserine lactones |
| HCN | Hydrogen cyanide |
| CAT | Catalas |
| GA- | Gibberellic acid |
| PR | Pathogenesis related proteins |
| ET | Ethylene |
| MDA | Malondialdehyde |
| APX | Ascorbate peroxidase |
| GR | Glutathione reductase |
| GST | Glutathione-S-transferase |
| GPX | Gaucol peroxidase |
References
- Nadarajah, K.K. Rhizosphere Interactions: Life Below Ground. In Plant-Microbe Interaction: An Approach to Sustainable Agriculture; Choudhary, D.K., Varma, A., Tuteja, N., Eds.; Springer: Singapore, 2016. [Google Scholar] [CrossRef]
- Belhaj, K.; Cano, L.M.; Prince, D.C.; Kemen, A.; Yoshida, K.; Dagdas, Y.F.; Etherington, G.J.; Schoonbeek, H.J.; Van Esse, H.P.; Jones, J.D.; Kamoun, S. Arabidopsis late blight: Infection of a nonhost plant by Albugo laibachii enables full colonization by Phytophthora infestans. Cellular Microbiology 2017, 19, 12628. [Google Scholar] [CrossRef]
- del Carmen Orozco-Mosqueda M, Fadiji AE, Babalola OO, Glick BR, Santoyo G, Rhizobiome engineering: Unveiling complex rhizosphere interactions to enhance plant growth and health. Microbiological Research 2022, 263, 127137. [CrossRef]
- Rajesh, P.; Ravishankar, R. Quorum quenching activity in cell-free-lysate of endophytic bacteria isolated from Pterocarpus santalinus Linn and its effect on quorum sensing regulated biofilm in Pseudomonas aeruginosa PAO. Microbiological Research 2014, 169, 561–569. [Google Scholar] [CrossRef]
- Sharma, M.P.; Grover, M.; Chourasiya, D.; Bharti, A.; Agnihotri, R.; Maheshwari, H.S.; Pareek, A.; Buyer, J.S.; Sharma, S.K.; Schutz, L.; et al. Deciphering the role of trehalose in tripartite symbiosis among rhizobia, arbuscular mycorrhizal fungi, and legumes for enhancing abiotic stress tolerance in crop plants. Frontiers in Microbiology 2020, 11, 2219. [Google Scholar] [CrossRef]
- Pan, J.; Baumgarten, A.; May, G. Effects of host plant environment and Ustilago maydis infection on the fungal endophyte community of maize (Zea mays). New Phytology 2008, 178, 147–156. [Google Scholar] [CrossRef]
- Bhattacharyya, P.N.; Jha, D.K. Plant growth-promoting rhizobacteria (PGPR): Emergence in agriculture. World Journal of Microbiology and Biotechnology 2011, 28, 1327–1350. [Google Scholar] [CrossRef]
- Naziya, B.; Murali, M.; Amruthesh, K.N. Plant growth-promoting fungi (PGPF) instigate plant growth and induce disease resistance in Capsicum annuum L. upon infection with Colletotrichum capsici (Syd. ) Butler & Bisby. Biomolecules 2019, 10, 41. [Google Scholar] [CrossRef]
- Omomowo, I.O.; Adedayo, A.A.; Omomowo, O.I. Biocontrol Potential of Rhizospheric Fungi from Moringa oleifera, their Phytochemicals and Secondary Metabolite Assessment Against Spoilage Fungi of Sweet Orange (Citrus sinensis). Asian Journal of Applied Science 2020, 8, 35–48. [Google Scholar] [CrossRef]
- Martinez-Medina, A.; Del Mar Alguacil, M.; Pascual, J.A.; Van Wees, S.C. Phytohormone profiles induced by Trichoderma isolates correspond with their biocontrol and plant growth-promoting activity on melon plants. Journal of chemical ecology 2014, 40, 804–815. [Google Scholar] [CrossRef] [PubMed]
- Adedayo, A.A.; Babalola, O.O. Fungi that promote plant growth in the rhizosphere boost crop growth. Journal of Fungi 2023, 9, 239. [Google Scholar] [CrossRef] [PubMed]
- Manzoni, C.; Kia, D.; Vandrovcova, J.; Hardy, J.; Wood, N.; Lewis, P.; Ferrari, R. Genome, transcriptome and proteome: The rise of omics data and their integration in biomedical sciences. Briefings in Bioinformatics 2018, 19, 286–302. [Google Scholar] [CrossRef]
- Liu A, Chen S, Wang M, Liu D, Chang R, Wang Z, Lin X, Bai B, Ahammed GJ, Arbuscular mycorrhizal fungus alleviates chilling stress by boosting redox poise and antioxidant potential of tomato seedlings. Journal of Plant Growth Regulation 2016, 35, 109–120. [CrossRef]
- Bakr, J.; Daood, H.; Pek, Z.; Helyes, L.; Posta, K. Yield and quality of mycorrhized processing tomato under water scarcity. Applied Ecology and Environmental Research 2016, 15, 401–413. [Google Scholar] [CrossRef]
- Halifu, S.; Deng, X.; Song, X.; Song, R. Effects of two Trichoderma strains on plant growth, rhizosphere soil nutrients, and fungal community of Pinus sylvestris var. mongolica annual seedlings. Forests 2019, 10, 75. [Google Scholar] [CrossRef]
- Salas-Marina, M.A.; Silva-Flores, M.A.; Cervantes-Badillo, M.G.; Rosales-Saavedra, M.T.; Islas-Osuna, M.A.; Casas-Flores, S. The plant growth promoting fungus Aspergillus ustus promotes growth and induces resistance against different lifestyle pathogens in Arabidopsis thaliana. Journal of Microbiology and Biotechnology 2011, 21, 686–696. [Google Scholar] [CrossRef]
- Radhakrishnan, R.; Kang, S.; Baek, I.; Lee, I. Characterization of plant growth promoting traits of Penicillium species against the effects of high soil salinity and root disease. Journal of Plant Interactions. 2014, 9, 754–762. [Google Scholar] [CrossRef]
- Khan AL, Shinwari ZK, Kim YH, Waqas M, Hamayun M, Kamran M; et al. Role of endophyte Chaetomium globosum LK4 in growth of Capsicum annum by production of gibberellins and indole acetic acid. Pakistan Journal of Botany 2012, 44, 1601–1607. [Google Scholar]
- lhamed, M.F.A.; Shebany, Y.M. Endophytic Chaetomium globosum enhances maize seedling copper stress tolerance. Plant Biology 2012, 14, 859–863. [Google Scholar] [CrossRef]
- Vujanovic, V.; Goh, Y.K. qPCR quantification of Sphaerodes mycoparasitica biotrophic mycoparasite interaction with Fusarium graminearum: In vitro and in planta assays. Archives of Microbiology 2012, 194, 707–717. [Google Scholar] [CrossRef]
- Jogaiah, S.; Abdelrahman, M.; Tran, L.S.P.; Shin-ichi, I. Characterization of rhizosphere fungi that mediate resistance in tomato against bacterial wilt disease. Journal of Experimental Botany 2013, 64, 3829–3842. [Google Scholar] [CrossRef]
- Uzoma, K.; Inoue, M.; Andry, H.; Fujimaki, H.; Zahoor, A.; Nishihara, E. Effect of cow manure biochar on maize productivity under sandy soil condition. Soil Use Management 2011, 27, 205–212. [Google Scholar] [CrossRef]
- van Noorden, G.E.; Kerim, T.; Goffard, N.; Wiblin, R.; Pellerone, F.I.; Rolfe, B.G.; Mathesius, U. Overlap of proteome changes in Medicago truncatula in response to auxin and Sinorhizobium meliloti. Plant Physiology 2007, 144, 1115. [Google Scholar] [CrossRef]
- Manthey, K.; Krajinski, F.; Hohnjec, N.; Firnhaber, C.; Puhler, A.; Perlick, A.; Kuster, H. Transcriptiome profiling in root-nodules and arbuscualr mycorrhiza identifies a collection of novel genes induced during Medicago truncatula root endosymbioses. Molecular Plant-Microbe Interactions 2004, 17, 1063–1077. [Google Scholar] [CrossRef]
- Fulekar, M.H. Rhizosphere bioremediation of pesticides by microbial consortium and potential microorganism. International Journal of Current Microbiology and Applied Sciences 2014, 235–248. [Google Scholar]
- Ahmad P, Hashem A, Abd-Allah EF, Alqarawi AA, John R, Egamberdieva D et al. Role of Trichoderma harzianum in mitigating NaCl stress in Indian mustard (Brassica juncea L.) through antioxidative defense system. Frontiers in Plant Science 2015, 6, 868. [Google Scholar] [CrossRef]
- Hameed, K.; Couch, H.B. Effects of Penicillium simplicissimum on growth, chemical composition, and root exudation of axenically grown marigolds. Phytopathology 1971, 62, 669. [Google Scholar]
- Villa-Rodriguez, E.; Moreno-Ulloa, A.; Castro-Longoria, E.; Parra-Cota, F.I.; de los SantosVillalobos, S. Integrated omics approaches for deciphering antifungal metabolites produced by a novel Bacillus species, B. cabrialesii TE3T, against the spot blotch disease of wheat (Triticum turgidum L. subsp. durum). Microbiological Research 2021, 251, 126826. [Google Scholar] [CrossRef]
- Monchgesang, S.; Strehmel, N.; Schmidt, S.; Westphal, L.; Taruttis, F.; Muller, E.; Herklotz, S.; Neumann, S.; Scheel, D. Natural variation of root exudates in Arabidopsis thaliana-linking metabolomic and genomic data. Scientific Reports 2016, 6, 1–11. [Google Scholar] [CrossRef]
- Zavala-Gonzalez, E.A.; Escudero, N.; Lopez-Moya, F.; Aranda-Martinez, A.; Exposito, A.; Ricano-Rodriguez, J.; Naranjo-Ortiz, M.A.; Ramirez-Lepe, M.; Lopez-Llorca, L.V. Some isolates of the nematophagous fungus Pochonia chlamydosporia promote root growth and reduce flowering time of tomato. Annals of Applied Biology 2015, 166, 472–483. [Google Scholar] [CrossRef]
- Nihorimbere, V.; Cawoy, H.; Seyer, A.; Brunelle, A.; Thonart, P.; Ongena, M. Impact of rhizosphere factors on cyclic lipopeptide signature from the plant beneficial strain Bacillus amyloliquefaciens S499. FEMS Microbiology Ecology 2012, 79, 176–191. [Google Scholar] [CrossRef]
- Palmer, I.A.; Chen, H.; Chen, J.; Chang, M.; Li, M.; Liu, F.; Fu, Z.Q. Novel salicylic acid analogs induce a potent defense response in Arabidopsis. International Journal of Molecular Sciences 2019, 20, 3356. [Google Scholar] [CrossRef]
- Strehmel, N.; Bottcher, C.; Schmidt, S.; Scheel, D. Profiling of secondary metabolites in root exudates of Arabidopsis thaliana. Phytochemistry 2014, 108, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Hyakumachi, M. Plant-growth promoting fungi from turf grass rhizosphere with potential for disease suppression. Soil Microorganisms 1994, 44, 53–68. [Google Scholar]
- Harman, G.E.; Petzoldt, R.; Comis, A.; Chen, J. Interactions between Trichoderma harzianum strain T22 and maize inbred line Mo17 and effects of this interaction on diseases caused by Pythium ultimum and Colletotrichum graminicola. Phytopathology 2004, 94, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Yamagiwa, Y.; Toyoda, K.; Inagaki, Y.; Ichinose, Y.; Hyakumachi, M.; Shiraishi, T. Talaromyces wortmannii FS2 emits β-caryophyllene, which promotes plant growth and induces resistance. Journal of General Plant Pathology 2011, 77, 336–341. [Google Scholar] [CrossRef]
- Waweru, B.; Turoop, L.; Kahangi, E.; Coyne, D.; Dubois, T. Non-pathogenic Fusarium oxysporum endophytes provide field control of nematodes, improving yield of banana (Musa sp.). Biological Control 2014, 74, 82–88. [Google Scholar] [CrossRef]
- Zeng, Q.; Huang, H.; Zhu, J.; Fang, Z.; Sun, Q.; Bao, S. A new nematicidal compound produced by Streptomyces albogriseolus HA10002. Antonie Van Leeuwenhoek 2013, 103, 1107–1111. [Google Scholar] [CrossRef]
- Reyes-Tena, A.; Rincon-Enríquez, G.; Lopez-Perez, L.; Quinones-Aguilar, E.E. Effect of mycorrhizae and actinomycetes on growth and bioprotection of Capsicum annuum L. against Phytophthora capsici. Pakistan Journal of Agricultural Sciences 2017, 54, 4245. [Google Scholar] [CrossRef]
- Lu, Y.; Wang, N.; He, J.; Li, Y.; Gao, X.; Huang, L.; Yan, X. Expression and characterization of a novel chitinase with antifungal activity from a rare actinomycete, Saccharothrix yanglingensis Hhs. 015. Protein Expression and Purification 2018, 143, 45–51. [Google Scholar] [CrossRef]
- Hoyos-Carvajal, L.; Orduz, S.; Bissett, J. Growth stimulation in bean (Phaseolus vulgaris L. ) by Trichoderma. Biological Control 2009, 51, 409–416. [Google Scholar] [CrossRef]
- Inderiati, S.; Franco, C.M. Isolation and identification of endophytic actinomycetes and their antifungal activity. Journal of Tropical Biodiversity and Biotechnology 2008, 1, 1–6. [Google Scholar]
- Chandra-Mohana, N.; Narendra-Kumar, H.K.; Mahadevakumar, S.; Sowmya, R.; Sridhar, K.R.; Satish, S. First Report of Aspergillus versicolor Associated with Fruit Rot Disease of Tomato (Solanum lycopersicum) from India. Plant Disease 2022, 106, 1300. [Google Scholar] [CrossRef]
- Sarma, M.; Kumar, V.; Saharan, K.; Srivastava, R.; Sharma, A.K.; Prakash, A.; Sahai, V.; Bisaria, V.S. Application of inorganic carrier-based formulations of fluorescent pseudomonads and Piriformospora indica on tomato plants and evaluation of their efficacy. Journal of Applied Microbiology 2011, 111, 456–466. [Google Scholar] [CrossRef]
- De Palma, M.; Scotti, R.; D’Agostino, N.; Zaccardelli, M.; Tucci, M. Phyto-Friendly Soil Bacteria and Fungi Provide Beneficial Outcomes in the Host Plant by Differently Modulating Its Responses through (In)Direct Mechanisms. Plants 2022, 11, 2672. [Google Scholar] [CrossRef]
- Eshaghi Gorgi, O.; Fallah, H.; Niknejad, Y.; Barari Tari, D. Effect of Plant growth promoting rhizobacteria (PGPR) and mycorrhizal fungi inoculations on essential oil in Melissa officinalis L. under drought stress. Biologia 2022, 77, 11–20. [Google Scholar] [CrossRef]
- Dutta, P.; Mahanta, M.; Singh, S.B.; Thakuria, D.; Deb, L.; Kumari, A.; Upamanya, G.K.; Boruah, S.; Dey, U.; Mishra, A.K.; Vanlaltani, L. Molecular interaction between plants and Trichoderma species against soil-borne plant pathogens. Frontiers in Plant Science 2023, 14, 1145715. [Google Scholar] [CrossRef]
- Eldredge, S.D.; Geary, B.; Jensen, S.L. Seed isolates of Alternaria and Aspergillus fungi increase germination of Astragalus utahensis. Native Plants Journal 2016, 17, 89–94. [Google Scholar] [CrossRef]
- Yedidia, I.; Srivastva, A.K.; Kapulnik, Y.; Chet, I. Effect of Trichoderma harzianum on microelement concentrations and increased growth of cucumber plants. Plant and Soil 2001, 235, 235–242. [Google Scholar] [CrossRef]
- Kaveh, H.; Vatandoost, S.; Aroiee, H.; Mazhabi, M. Would Trichoderma affect seed germination and seedling quality of two muskmelon cultivars, Khatooni and Qasri and increase their transplanting success? Journal of Biological and Environmental Sciences 2011, 5, 169–175. [Google Scholar]
- Muslim, A.; Horinouchi, H.; Hyakumachi, M. Biological control of Fusarium wilt of tomato with hypovirulent binucleate Rhizoctonia in greenhouse conditions. Mycoscience 2003, 44, 77–84. [Google Scholar] [CrossRef]
- Omotayo, O.P.; Babalola, O.O. Resident rhizosphere microbiome’s ecological dynamics and conservation: Towards achieving the envisioned Sustainable Development Goals, a review. International Soil and Water Conservation Research 2020, 9, 127–142. [Google Scholar] [CrossRef]
- Khan AL, Hamayun M, Ahmad N, Waqas M, Kang SM, Kim YH; et al. Exophiala sp. LHL08 reprograms Cucumis sativus to higher growth under abiotic stresses. Physiologia Plantarum 2011, 143, 329–334. [Google Scholar] [CrossRef] [PubMed]
- Narsing Rao, M.P.; Lohmaneeratana, K.; Bunyoo, C.; Thamchaipenet, A. Actinobacteria–plant interactions in alleviating abiotic stress. Plants 2022, 11, 2976. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Vo, Q.A.T.; Barnett, S.J.; Ballard, R.A.; Zhu, Y.; Franco, C.M.M. Revealing the underlying mechanisms mediated by endophytic actinobacteria to enhance the rhizobia-chickpea (Cicer arietinum L.) symbiosis. Plant Soil. 2022, 474, 299–318. [Google Scholar] [CrossRef]
- Alvarez-Perez, J.M.; Gonzalez-Garcia, S.; Cobos, R.; Olego, M.; Ibanez, A.; Diez-Galan, A.; Garzon-Jimeno, E.; Coque, J.J.R. Use of endophytic and rhizosphere Actinobacteria from grapevine plants to reduce nursery fungal graft infections that lead to young grapevine decline. Applied Environmental Microbiology 2017, 83, e01564–17. [Google Scholar] [CrossRef]
- Rangseekaew, P.; Barros-Rodriguez, A.; Pathom-Aree, W.; Manzanera, M. Plant beneficial deep-sea Actinobacterium, Dermacoccus abyssi MT1.1(T) promote growth of tomato (Solanum lycopersicum) under salinity stress. Biology 2022, 11, 191. [Google Scholar] [CrossRef]
- Mishra, J.; Mishra, I.; Arora, N.K. 2, 4-Diacetylphloroglucinol producing Pseudomonas fluorescens JM-1 for management of ear rot disease caused by Fusarium moniliforme in Zea mays L. Biotech 2022, 12, 1–15. [Google Scholar] [CrossRef]
- Harbort, C.J.; Hashimoto, M.; Inoue, H.; Niu, Y.; Guan, R.; Rombola, A.D.; et al. Root-secreted coumarins and the microbiota interact to improve iron nutrition in Arabidopsis. Cell Host & Microbe 2020, 28, 825–837. [Google Scholar] [CrossRef]
- Neal, A.L.; Ahmad, S.; Gordon-Weeks, R.T.J. Benzoxazinoids in root exudates of maize attract Pseudomonas putida to the rhizosphere. PLoS ONE 2012, 7, e35498. [Google Scholar] [CrossRef]
- Yu, P.; He, X.; Baer, M.; Beirinckx, S.; Tian, T.; Moya, Y.A.; et al. Plant flavones enrich rhizosphere Oxalobacteraceae to improve maize performance under nitrogen deprivation. Nature Plants 2021, 7, 481–499. [Google Scholar] [CrossRef]
- Bringhurst, R.M.; Cardon, Z.G.; Gage, D.J. Galactosides in the rhizosphere: Utilization by Sinorhizobium meliloti and development of a biosensor. Proceedings of the National Academy of Sciences of the United States of America 2001, 98, 4540–4545. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Wang, D.; Liu, Y.; Li, S.; Shen, Q.; Zhang, R. Effects of different plant root exudates and their organic acid components on chemotaxis, biofilm formation and colonization by beneficial rhizosphere-associated bacterial strains. Plant Soil 2014, 374, 689–700. [Google Scholar] [CrossRef]
- Xiong, Y.W.; Li, X.W.; Wang, T.T.; Gong, Y.; Zhang, C.M.; Xing, K.; et al. Root exudates-driven rhizosphere recruitment of the plant growth-promoting rhizobacterium Bacillus flexus KLBMP 4941 and its growth-promoting effect on the coastal halophyte Limonium sinense under salt stress. Ecotoxicology and Environmental Safety 2020, 194, 374. [Google Scholar] [CrossRef]
- Hida, A.; Oku, S.; Miura, M.; Matsuda, H.; Tajima, T.; Kato, J. Characterization of methyl-accepting chemotaxis proteins (MCPs) for amino acids in plant-growth-promoting rhizobacterium Pseudomonas protegens CHA0 and enhancement of amino acid chemotaxis by MCP genes overexpression. Bioscience, Biotechnology, and Biochemistry 2020, 84, 1948–1957. [Google Scholar] [CrossRef]
- Rios, J.; Pascual, J.; Guillen, M.; Lopez-Martinez, A.; Carvajal, M. Influence of foliar Methyl-jasmonate biostimulation on exudation of glucosinolates and their effect on root pathogens of broccoli plants under salinity condition. Scientia Horticulturae 2021, 282, 110027. [Google Scholar] [CrossRef]
- Barnawal, D.; Bharti, N.; Pandey, S.S.; Pandey, A.; Chanotiya, C.S.; Kalra, A. Plant growth-promoting rhizobacteria enhance wheat salt and drought stress tolerance by altering endogenous phytohormone levels and TaCTR1/TaDREB2 expression. Physiol. Plant 2017, 161, 502–514. [Google Scholar] [CrossRef]
- Caradonia, F.; Francia, E.; Morcia, C.; Ghizzoni, R.; Moulin, L.; Terzi, V.; Ronga, D. Arbuscular Mycorrhizal Fungi and Plant Growth Promoting Rhizobacteria Avoid Processing Tomato Leaf Damage during Chilling Stress. Agronomy 2019, 9, 299. [Google Scholar] [CrossRef]
- Zhao, L.; Zhang, Y.Q. Effects of phosphate solubilization and phytohormone production of Trichoderma asperellum Q1 on promoting cucumber growth under salt stress. Journal of Integrative Agriculture 2015, 14, 1588–1597. [Google Scholar] [CrossRef]
- Sukkasem, P.; Kurniawan, A.; Kao, T.C.; Chuang, H.W. A multifaceted rhizobacterium Bacillus licheniformis functions as a fungal antagonist and a promoter of plant growth and abiotic stress tolerance. Environmental and Experimental Botany 2018, 155, 541–551. [Google Scholar] [CrossRef]
- Mabood, F.; Souleimanov, A.; Khan, W.; Smith, D. Jasmonates induce Nod factor production by Bradyrhizobium japonicum. Plant Physiology and Biochemistry 2006, 44, 759–765. [Google Scholar] [CrossRef]
- Chen, S.; Jin, W.; Liu, A.; Zhang, S.; Liu, D.; Wang, F.; Lin, X.; He, C. Arbuscular mycorrhizal fungi (AMF) increase growth and metabolism in cucumber subjected to low temperature. Scientia Horticulturae 2013, 160, 222–229. [Google Scholar] [CrossRef]
- Chen, G.; Yan, W.; Yang, L.; Gai, J.; Zhu, Y. Overexpression of StNHX1, a novel vacuolar Na+ /H+ antiporter gene from Solanum torvum, Enhances salt tolerance in transgenic vegetable Soybean. Horticulture Environment Biotechnology 2014, 55, 213–221. [Google Scholar] [CrossRef]
- Rabeendran, N.; Moot, D.J.; Jones, E.E.; Stewart, A. Inconsistent growth promotion of cabbage and lettuce from Trichoderma isolates. New Zealand Plant Protection 2000, 53, 143–146. [Google Scholar] [CrossRef]
- Hossain, M.M.; Sultana, F. Application and mechanisms of plant growth promoting fungi (PGPF) for phytostimulation. Organic agriculture 2020, 1, 31. [Google Scholar] [CrossRef]
- Islam, S.; Akanda, A.M.; Prova, A.; Sultana, F.; Hossain, M.M. Growth promotion effect of Fusarium spp. PPF1 from Bermuda grass (Cynodon dactylon) rhizosphere on Indian spinach (Basella alba) seedlings are linked to root colonization. Archives of Phytopathology and Plant Protection 2014, 47, 2319–2331. [Google Scholar] [CrossRef]
- Zhang, N.; Wang, D.; Liu, Y.; Li, S.; Shen, Q.; Zhang, R. Effects of different plant root exudates and their organic acid components on chemotaxis, biofilm formation and colonization by beneficial rhizosphere-associated bacterial strains. Plant Soil 2014, 374, 689–700. [Google Scholar] [CrossRef]
- Sorty, A.M.; Meena, K.K.; Choudhary, K.; Bitla, U.M.; Minhas, P.S.; Krishnani, K.K. Effect of Plant Growth Promoting Bacteria Associated with Halophytic Weed (Psoralea corylifolia L.) on Germination and Seedling Growth of Wheat Under Saline Conditions. Applied Biochemistry and Biotechnology 2016, 180, 872–882. [Google Scholar] [CrossRef]
- Rin, S.; Mizuno, Y.; Shibata, Y.; Fushimi, M.; Katou, S.; Sato, I.; Chiba, S.; Kawakita, K.; Takemoto, D. EIN2-Mediated signaling is involved in pre-invasion defense in Nicotiana benthamiana against potato late blight pathogen, Phytophthora infestans. Plant Signaling Behaviour 2017, 12, e13007337474. [Google Scholar] [CrossRef]
- Benjelloun, I.; Alami, I.T.; El Khadir, M.; Douira, A.; Udupa, S. Co-Inoculation of Mesorhizobium ciceri with either Bacillus sp. or Enterobacter aerogenes on Chickpea Improves Growth and Productivity in Phosphate-Deficient Soils in Dry Areas of a Mediterranean Region. Plants 2021, 10, 571. [Google Scholar] [CrossRef]
- Li, Y.; Sun, H.; Yang, W.; Chen, G.; Xu, H. Dynamics of Bacterial and Viral Communities in Paddy Soil with Irrigation and Urea Application. Viruses 2019, 11, 347. [Google Scholar] [CrossRef]
- Dong, Z.Y.; Narsing Rao, M.P.; Wang, H.F.; Fang, B.Z.; Liu, Y.H.; Li, L.; Xiao, M.; Li, W.J. Transcriptomic analysis of two endophytes involved in enhancing salt stress ability of Arabidopsis thaliana. Science of the Total Environment 2019, 686, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.; Chen, L.J.; Pan, S.Y.; Li, X.W.; Xu, M.J.; Zhang, C.M.; Xing, K.; Qin, S. Antifungal potential evaluation and alleviation of salt stress in tomato seedlings by a halotolerant plant growth-promoting actinomycete Streptomyces sp. KLBMP5084. Rhizosphere 2020, 16, 100262. [Google Scholar] [CrossRef]
- Berry, A.M.; Murphy, T.M.; Okubara, P.A.; Jacobsen, K.R.; Swensen, S.M.; Pawlowski, K. Novel Expression Pattern of Cytosolic Gln Synthetase in Nitrogen-Fixing Root Nodules of the Actinorhizal Host, Datisca glomerata. Plant Physiology 2004, 135, 1849–1862. [Google Scholar] [CrossRef]
- Hamdali, H.; Hafidi, M.; Virolle, M.J.; Ouhdouch, Y. Growth promotion and protection against damping-off of wheat by two rock phosphate solubilizing actinomycetes in a P-deficient soil under greenhouse conditions. Applied Soil Ecology 2008, 40, 510–517. [Google Scholar] [CrossRef]
- Zhou, L.S.; Kun, T.; Shun, X.G. The plant growth-promoting fungus (PGPF) Alternaria sp. A13 markedly enhances Salvia miltiorrhiza root growth and active ingredient accumulation under greenhouse and field conditions. International journal of molecular sciences 2018, 19, 270. [Google Scholar] [CrossRef]
- Ito, K.; Mahmood, A.; Kataoka, R.; Takagi, K. Dichlorodiphenyltrichloroethane (DDT) degradation by Streptomyces sp. isolated from DDT contaminated soil. Bioremediation Journal 2021, 25, 148–158. [Google Scholar]
- Daigham, G.E.; Mahfouz, A.Y.; Abdelaziz, A.M.; Nofel, M.M.; Attia, M.S. Protective role of plant growth-promoting fungi Aspergillus chevalieri OP593083 and Aspergillus egyptiacus OP593080 as biocontrol approach against Alternaria leaf spot disease of Vicia faba plant. Biomass Conversion and Biorefinery 2024, 14, 23073–23089. [Google Scholar] [CrossRef]
- Escobar Diaz, P.A.; Gil, O.J.; Barbosa, C.H.; Desoignies, N.; Rigobelo, E.C. Aspergillus spp. and Bacillus spp. as growth promoters in cotton plants under greenhouse conditions. Frontiers in Sustainable Food Systems. 2021, 5, 709267. [Google Scholar] [CrossRef]
- Kudjordjie, E.N.; Sapkota, R.; Steffensen, S.K.; Fomsgaard, I.S.; Nicolaisen, M. Maize synthesized benzoxazinoids affect the host associated microbiome. Microbiome 2019, 7, 59. [Google Scholar] [CrossRef]
- Olanrewaju, O.S.; Ayangbenro, A.S.; Glick, B.R.; Babalola, O.O. Plant health: Feedback effect of root exudates-rhizobiome interactions. Applied Microbiology and Biotechnology 2019, 103, 1155–1166. [Google Scholar] [CrossRef]
- Molefe, R.R.; Amoo, A.E.; Babalola, O.O. Communication between plant roots and the soil microbiome; involvement in plant growth and development. Symbiosis 2023, 90, 231–239. [Google Scholar] [CrossRef]
- Igiehon, N.; Babalola, O. Rhizosphere microbiome modulators: Contributions of nitrogen fixing bacteria towards sustainable agriculture. International Journal of Environment Research Public Health 2018, 15, 574. [Google Scholar] [CrossRef]
- Abdelkhalek, A.; Bashir, S.; El-Gendi, H.; Elbeaino, T.; El-Rahim, W.M.A.; Moawad, H. Protective activity of Rhizobium leguminosarum bv. viciae strain 33504-Mat209 against Alfalfa Mosaic Virus Infection in Faba Bean Plants. Plants 2023, 12, 2658. [Google Scholar] [CrossRef]
- Agbodjato, N.A.; Assogba, S.A.; Babalola, O.O.; Koda, A.D.; Aguegue, R.M.; Sina, H.; Dagbenonbakin, G.D.; Adjanohoun, A.; Baba-Moussa, L. Formulation of biostimulants based on arbuscular mycorrhizal fungi for maize growth and yield. Front Agron 2022, 4, 894489. [Google Scholar] [CrossRef]
- Semchenko, M.; Lepik, A.; Abakumova, M.; Zobel, K. Different sets of belowground traits predict the ability of plant species to suppress and tolerate their competitors. Plant Soil 2017, 424, 157–169. [Google Scholar] [CrossRef]
- Mitter, B.; Brader, G.; Pfaffenbichler, N.; Sessitsch, A. Next generation microbiome applications for crop production-limitations and the need of knowledge-based solutions. Current Opinion in Microbiology 2019, 49, 59–65. [Google Scholar] [CrossRef]
- Dlamini, S.P.; Akanmu, A.O.; Babalola, O.O. Rhizospheric microorganisms: The gateway to a sustainable plant health. Frontiers in Sustainable Food Systems 2022, 6, 92580. [Google Scholar] [CrossRef]
- Djaya, L.; Istifadah, N.; Hartati, S.; Joni, I.M. In vitro study of plant growth promoting rhizobacteria (PGPR) and endophytic bacteria antagonistic to Ralstonia solanacearum formulated with graphite and silica nano particles as a biocontrol delivery system (BDS). Biocatalalysis and Agricultural Biotechnology 2019, 19, 101153. [Google Scholar] [CrossRef]
- Hanif, M.K.; Malik, K.A.; Hameed, S.; Saddique, M.J.; Fatima, K.; Naqqash, T.; et al. Growth stimulatory effect of AHL producing Serratia spp. from potato on homologous and non-homologous host plants. Microbiological Research 2020, 238, 126506. [Google Scholar] [CrossRef]
- Dixit, R.; Agrawal, L.; Singh, S.P.; Singh, P.C.; Prasad, V.; Chauhan, P.S. Paenibacillus lentimorbus induces autophagy for protecting tomato from Sclerotium rolfsii infection. Microbiological Research 2018, 215, 164–174. [Google Scholar] [CrossRef]
- Ali, S.Z.; Sandhya, V.; Grover, M.; Linga, V.R.; Bandi, V. Effect of inoculation with a thermotolerant plant growth promoting Pseudomonas putida strain AKMP7 on growth of wheat (Triticum spp.) under heat stress. Journal of Plant Interaction 2011, 6, 239–246. [Google Scholar] [CrossRef]
- Yin, D.C.; Song, R.Q.; Qi, J.Y.; Deng, X. Ectomycorrhizal fungus enhances drought tolerance of Pinus sylvestris var. mongolica seedlings and improves soil condition. Journal of Forestry Research 2018, 29, 1775–1788. [Google Scholar] [CrossRef]
- Singh, S.B.; Gowtham, H.G.; Murali, M.; Hariprasad, P.; Lakshmeesha, T.R.; Murthy, K.N.; Amruthesh, K.N.; Niranjana, S.R. Plant growth promoting ability of ACC deaminase producing rhizobacteria native to Sunflower (Helianthus annuus L.). Biocatalysis and Agricultural Biotechnology 2019, 18, 101089. [Google Scholar] [CrossRef]
- Talbot, N.J.; Ebbole, D.J.; Hamer, J.E. Identification and characterization of MPG1, a gene involved in pathogenicity from the rice blast fungus Magnaporthe grisea. The Plant Cell 1993, 5, 1575–1590. [Google Scholar]
- Topalovic, O.; Geisen, S. Nematodes as suppressors and facilitators of plant performance. New Phytologist 2023, 238, 2305–2312. [Google Scholar] [CrossRef]
- Cucu, M.A.; Gilardi, G.; Pugliese, M.; Ferrocino, I.; Gullino, M.L. Effects of biocontrol agents and compost against the Phytophthora capsici of zucchini and their impact on the rhizosphere microbiota. Applied Soil Ecology 2020, 154, 103659. [Google Scholar] [CrossRef]
- Bellini, A.; Gilardi, G.; Idbella, M.; Zotti, M.; Pugliese, M.; Bonanomi, G.; Gullino, M.L. Trichoderma enriched compost, BCAs and potassium phosphite control Fusarium wilt of lettuce without affecting soil microbiome at genus level. Applied Soil Ecology 2023, 182, 104678. [Google Scholar] [CrossRef]
Figure 1.
A Graphical Representation of Mutualistic/ Saprophytic Interaction in Plant Rhizobiome.
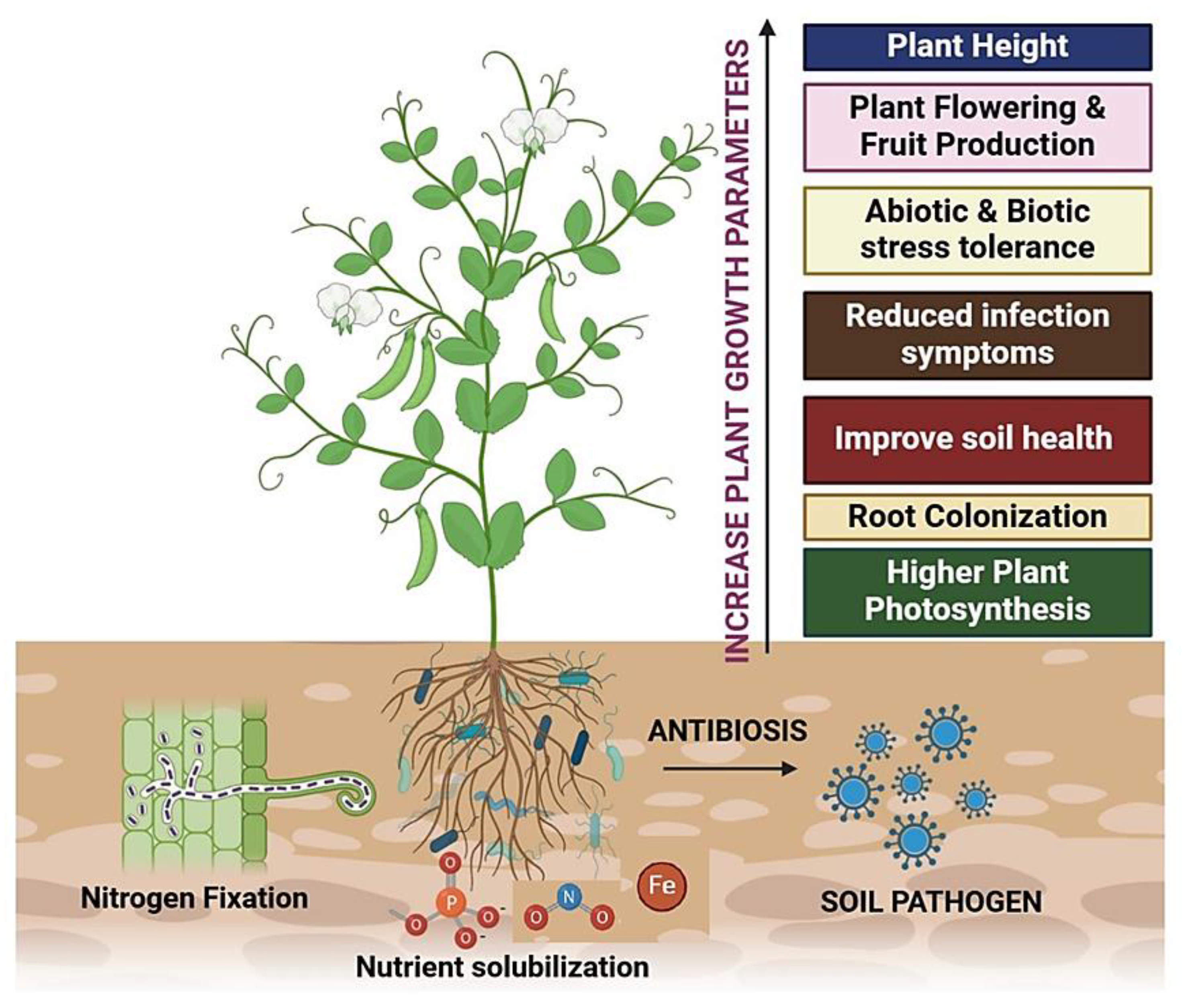
Figure 2.
An overview of Pathogenic/ Parasitic mode of interaction in Plant Rhizobiome.
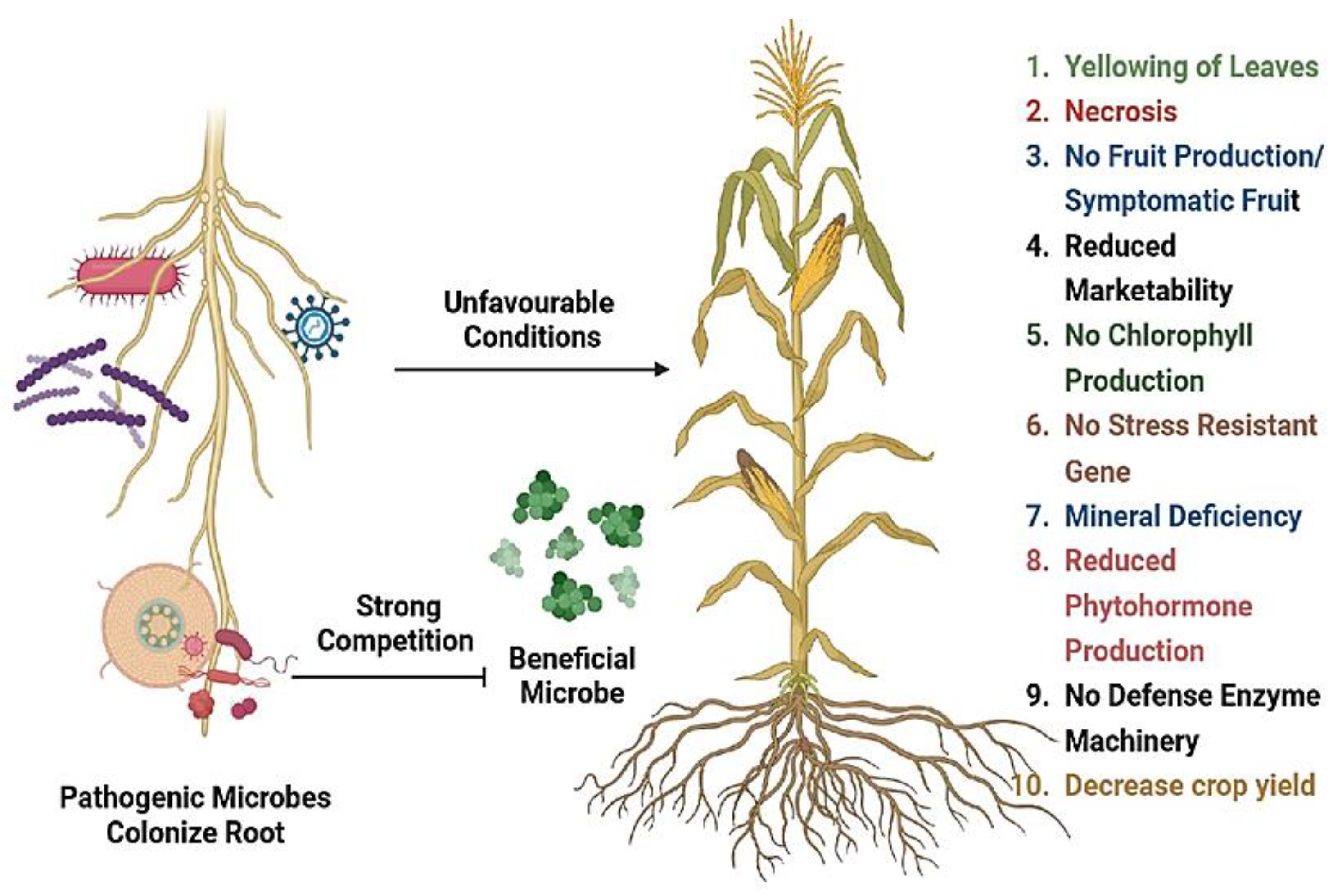

Table 1.
Evaluation of Rhizospheric Microbes in the Promotion of Plant growth and Increased Resistance.
Table 1.
Evaluation of Rhizospheric Microbes in the Promotion of Plant growth and Increased Resistance.
| Plant host | Rhizosphere Microbes | Plant growth Mechanism |
Resistance Mechanism |
Abiotic Pressure | Pathogen | Reference |
|---|---|---|---|---|---|---|
| Growth Promoting Bacteria/ Actinobacteria | ||||||
| Wheat (Triticum aestivum) |
P. putida | Produce more tiller, grain/ spike formation & 100 seed weight | Higher antioxidant activity, Proline, Total proteins, Sugars & Amino acids | Heat Stress | - | [102] |
|
Tomato (Lycopersicon esculentum) |
Pseudomonas strains R62 & R81 | Increase dry root weight, shoot weight & fruit yield under field conditions |
Control wilt disease incidence |
- | Fusarium oxysporum f.sp. lycopersici | [44] |
| - | Streptomyces albogriseolus | Higher 6-methyl-fungichromin | - | - | Meloidogyne incognita & Meloidogyne javanica | [38] |
| Chilli (Capsicum annuum) |
Actinomycetes | Increase plant height, stem diameter, leaves number & leaf area | Increased radical volume & reduce disease severity | - |
Phytophthora capsici |
[39] |
| Wheat (Triticum aestivum) |
Arthrobacter protophormiae (SA3) & Dietzia natronolimnaea (STR1) | Promote IAA content & reduced ABA/ACC content | - | Increase (ABA) & ACC under salt & drought stress | - | [67] |
| Tomato (Solanum lycopersicum) |
Paenibacillus lentimorbus | - | Expression of defense-related & autophagy-related genes |
- |
Sclerotium rolfsi | [101] |
| Cucumber (Cucumis sativus) |
Saccharothrix yanglingensis | Antifungal activity & Chitinase Production |
- |
Valsa mali | [40] | |
| Potato (Solanum tuberosum) | Lysinibacillus sp. | - | Antibiosis |
- |
Ralstonia solanacearum | [99] |
| Helianthus annuus | Bacillus subtilis | Increase Plant Biomass, Phosphate & iron solubilization, | Phytohormone production | - | Fusarium oxysporum |
[104] |
|
Solanum tuberosum Oryza sativa, Zea mays & Phaseolus vulgaris |
Serratia spp. | Higher phytase activity |
AHLs- acyl homoserine lactone & phytohormone production | - | - | [100] |
| Limonium sinense | Bacillus flexus KLBMP 4941 | Increase root length, shoot length & fresh weight | Organic acids production | Mitigated salt stress by producing organic acids (stearic, palmitic, palmitoleic & oleic acids | - | [64] |
| Wheat (Triticum turgidum) | Bacillus cabrialesii | Increase chlorophyll content & Roots colonization | Increased antibacterial metabolites such as Bacillaene, Bacilysin, Bacillibactin & subtilosin-A | - | Bipolaris sorokiniana | [28] |
| Solanum lycopersicum | Dermacoccus abyssi | Phosphate solubilization, Phytohormone (IAA) production, more total soluble sugar & total chlorophyll content |
Mitigated salt stress & osmoregulate by soluble sugars. Increase H2O2 scavenging activity | - |
- | [57] |
| Zea mays | Pseudomonas fluorescens | - | Reduced ear rot disease incidence | Fusarium moniliforme | [58] | |
| Growth Promoting Fungi/ Mycorrhizae | ||||||
| Tagetes erecta |
Penicillium citrinum & Penicillium simplicissimum |
Maximized the carbohydrates & reducing sugars. Increased size, dry matter & flower early. | Produced root exudated & enhanced polygalacturonase, & cellulase enzymes activity |
- | - | [27] |
| Cucumis melo | Trichoderma harzianum Trichoderma ghanense & Trichoderma hamatum. | Maximize leaves number, root and shoot weight, & IAA production | Inhibitory activity, Decrease wilt Incidence, Increase in SA & JA. |
More ACC deaminase |
Fusarium oxysporum | [10] |
| Sesamum indicum | Penicillium sp. NICS01 | Increased plant length, amino acid content & plant weight. | - | Salt stress | - | [17] |
| Tomato (Solanum lycopersicon) |
Funneliformis mosseae. | Increased biomass (Root and Shoot Fresh weight) | Improving Antioxidant Machinery |
Lowered Ascorbate, glutathione, redox ratio & L-galactono-1,4- lactone dehydrogenase ratio | - | [13] |
| Pinus sylvestris var. mongolica |
G. mosseae, G. etunicatum, G. claroideum, G.microaggregatum, G. geosporum & R. irregularis |
Promoted Lycopene & β-Carotene contents |
Increased antioxidant potential by enhancing ascorbic acid content |
Improved Leaf water content (%) under water stress. |
[14] | |
|
Pinus sylvestris var. mongolica |
Trichoderma harzianum E15 Trichoderma virens & S. luteus | Improved root structure by increasing root length & surface area. Increased fresh & dry weight. |
Lower proline content & MDA content | Increase superoxide dismutase-SOD & peroxidase-POX) activity to withstand drought stress |
- |
[15,103] |
| Chilli (Capsicum annuum L.) |
Talaromyces funiculosus | Phosphate solubilization, Siderophore, HCN, IAA, Chitinase & Cellulase production, Increase chlorophyll, fruit weight & plant Biomass. | Higher antagonistic activity, Disease Protection, PAL, POX, Chitinase, ISR, Lignin & Callose production | - | Colletotrichum capsici | [08] |
| Drum stick (Moringa Oleifera) |
P. chrysogenum & T. viride |
- | Maximize Antagonistic behaviour | - |
Fusarium oxysporum, Penicillium digitatum, & Aspergillus wentii |
[09] |
| Rape Mustard (Brassica juncea) | Trichoderma harzianum | Plant height, Root Length & Plant Dry weight | Maximized SOD, POD, APX, GR, GST, GPX, GSH & GSSG activities | Plant tolerant to saline stress [Proline 59.12%, H2O2 69.5% & MDA contents by 36.5%] |
- | [26] |
| Maize (Zea mays) |
Trichoderma harzianum |
Promoted root growth size, area & root hair area | Reduced anthracnose symptoms by maximizing β,1-3 glucanase, exochitinase & endochitinase in roots & shoots. |
- |
Pythium ultimum, & Colletotrichum graminicola |
[35] |
|
Brassica campestris L. var. perviridis. |
Talaromyces wortmannii | Increased growth by high levels of β-caryphyllene contents | Reduce infection symptoms |
- |
Colletotrichum higginsianum | [36] |
| Chilli (Capsicum annuum) | Arbuscular Vesicular Mycorrhizae (AMF) | More plant height, stem diameter, leaves number & leaf area | Increased radical volume & reduced disease severity |
- |
Phytophthora capsici |
[39] |
| Phaseolus vulgaris | Trichoderma sp. | Enhanced root length & aerial parts | Indole-3-acetic acid (IAA) |
- |
- |
[41] |
| Tomato (Solanum lycopersicon) |
Rhizoctonia G1, L2, W1 & W7 | Increase Plant weight (stems & leaves). | Reduced Disease Severity |
- |
Fusarium oxysporum f. sp. lycopersici |
[51] |
| Cucumber (Cucumis melo) |
Funneliformis mosseae | More fresh weight & dry weight was noticed | Increased Phenols, flavonoids, lignin, DPPH activity & phenolic compounds | Increased tolerance to temperature stress by maximizing glucose-6-phosphate dehydrogenase (G6PDH), shikimate dehydrogenase (SKDH), (PAL), cinnamyl alcohol dehydrogenase (CAD), (PPO), guaiacol peroxidase (G-POD), caffeic acid peroxidase (CA-POD) & chlorogenic acid peroxidase (CGA-POD). |
- |
[72] |
| Maize | Pseudomonas putida | - | Benzoxazinoids Allelochemical/ antimicrobial effect |
- | - | [60] |
| Cucumis sativus | Exophiala sp. LHL08 | GA production | Induce defense response by SA production | Increased abscisic acid (ABA) & other phytohormone production under salinity & drought | - | [53] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



