
Submitted:
02 January 2025
Posted:
03 January 2025
You are already at the latest version
Abstract
Porcine respiratory and reproductive syndrome (PRRS) is a viral disease that affects health and profitability in swine farms, largely due to a high variation in vaccination response. The objective was to discover and validate molecular markers associated with the antibody response in gilts after vaccination against PRRS virus. The study included one hundred (n=100) 6-month-old gilts from the breed Yorkshire that were PRRS virus-negative. After 7 days of acclimation period, all gilts were vaccinated with a modified live PRRS virus (day 0). Blood samples collected on day 21 were used to evaluate the antibody response, as the sample-to-positive (S/P) ratio, to PRRS virus after vaccination. DNA was extracted and genotyped using a low-density chip including 10,000 single nucleotide polymorphisms (SNPs). A genome-wide association study (GWAS) was performed to discover candidate SNPs associated with the S/P ratio, which were validated in two independent gilt populations (n=226). The SNPs rs80904326, rs80896559 and rs81358818 in the genes RNF144B, XKR9 and BMAL1, respectively, were associated (p < 0.01) with the S/P ratio and exhibited an additive effect. In conclusion, three SNPs are proposed as candidate markers for an improved immune response to vaccination against PRRS virus and may be useful in genetic selection programs.
Keywords:
antibody response
; DNA
; genetic markers
; polymorphism
; PRRS
1. Introduction
In the last decade, the global pig industry has faced a substantial challenge caused by the PRRS virus. This disease results in economic losses because it impairs feed conversion rate, reduces weight gain, decreases fertility and increases the culling rate in sows, which currently represents a serious risk to global food security [1,2]. New knowledge about the virology, evolution, and host response to PRRS virus has expanded rapidly. At the same time, new variants of the virus constantly emerge from outbreaks that have seriously hampered the effectiveness of PRRS control strategies, including vaccination [3].
Regarding vaccines, specifically those that are modified live virus (MLV), they are widely used to reduce the clinical impact of the PRRS virus and control the dynamics infection within the herd [4]; however, reversion to virulence, recombination between field and MLV strains, and the inability to elicit immunity against heterogeneous viruses compromises the efficacy of their protective effect [5]. Another feature of MLV vaccines is the variability of individual immune responses in vaccinated pigs, suggesting that the host genetic background may be involved in the re-emergence of outbreaks in vaccinated populations [6]. Recently, a genomic region explaining 15% of the genetic variance associated with PRRS virus vaccination was detected on Sus scrofa chromosome 7 (SSC7), which strongly supports the hypothesis of a genetic basis, at least partially, regulating the PRRS vaccination response [7].
Molecular technologies have been proposed as a potential strategy to study the genetic architecture of complex traits in pigs [8]. GWAS are being used to investigate the genomic basis associated with antibody response and postpartum fertility after vaccination against the PRRS virus [9,10]. Likewise, GWAS have been successfully used in pigs to identify chromosomal regions harboring SNPs and candidate genes associated with feed efficiency [11], loin muscular growth [12], intramuscular fat [13], reproductive traits [14], meat quality [15], carcass traits [16], as well as host immune response to PRRS infection and/or vaccination [17,18].
In addition, marker-assisted selection (MAS) has been suggested as a promising strategy to improve host response after PRRS infection or vaccination [19]. Polymorphisms from the CD163 gene were associated with resiliency to PRRS virus infection, suggesting MAS as a strategy to control the widespread of this disease [20]. Similarly, a SNP within the gene LONRF1 was associated with PRRS antibody response [21]. In another study, a SNP from the gene EXOC4 was associated with four reproductive traits in farrowing sows, and this SNP was proposed for MAS in breeding programs [22].
Vaccination against PRRS virus is the main strategy used to control PRRS in commercial swine farms [23]. Genes associated with the Major Histocompatibility Complex (MHC) appear to partially regulate the immune response after PRRS vaccination. The antibody response to PRRS infection, as determined by the sample-to-positive (S/P) ratio, has been linked to genetic variations in the MCH in naturally PRRS-infected sows and F1 replacement gilts [24]. In a similar study, Serão et al. [25] reported a moderately high estimate of heritability for the S/P ratio (0.45) measured at day 46 of a PRRS outbreak. Interestingly, these authors hypothesized that PRRS virus vaccination would yield similar results at the genetic level for S/P ratio, suggesting that this variable would be a reliable indicator of the response to PRRS vaccination.
The aforementioned molecular technologies appear to be potentially useful in deciphering the genetic basis regulating the varied immunological response to PRRS vaccination in gilts. Therefore, our objective was to discover and validate genetic markers associated with the antibody response, measured as S/P ratio, after vaccination against PRRS virus in Yorkshire replacement gilts through genome-wide and marker assisted technologies.
2. Materials and Methods
The Institutional Animal Care and Use Committee of the Instituto Tecnologico de Sonora approved all procedures performed in animals (Approval code: 2020-0043; 01 May 2020).
2.1. Location and Experimental Units
The study was conducted in a full-cycle commercial swine farm located in the Yaqui Valley, Sonora a warm and semiarid region in Northwest Mexico (LN 27°17’, LW 109°56’). The experiment included one hundred gilts (n = 100) from a Yorkshire maternal line, approximately 6-months-old and with initial average weight of 108.23 ± 10.9 kg. Gilts were housed in pens within the quarantine area, where they had free access to a water source. Feed was provided in feed bunks and was formulated according to nutritional requirements of this age and weight of gilts. The gilts were previously tested as PRRS-negative according to serological and molecular analyses performed at the beginning of the study.
2.2. PRRS Vaccination and Blood Sampling
After a period of adaptation (7 days), a commercial vaccine against the PRRS virus was applied intramuscularly (modified active virus strain ATCC-VR-2332, Ingelvac PRRS MLV, Boehringer Ingelheim Laboratory). The day of vaccination was considered day 0 of the study. Blood samples were collected from each female on day -7 (i.e., seven days prior to vaccination) to isolate viral RNA and Real-time PCR analysis was performed to confirm PRRS-negative diagnosis using a commercial kit (Tetracore Nextgen Real-Time QT-PCR). Blood samples were collected again on day 21 of the study and used to measure the total antibody response as the sample-to-positive (S/P) ratio using a commercial ELISA kit (Idexx Labs, Portland, Maine, USA). According to the manufacturer’s instructions, an S/P ratio higher than 0.4 was considered positive, as it indicated the presence of antibodies against the PRRS virus. This serologic assay had 100% sensitivity and 99.5% specificity, and it contained both American and European strains of the PRRS virus.
2.3. Genotyping and Quality Control
An additional blood sample was collected on day 40 after PRRS vaccination and spotted on FTA cards. The cards were then sent to Neogen-Geneseek Laboratory (Lincoln, NE, USA) and processed for DNA extraction and genotyping using a low-density SNP chip containing 10,000 SNPs (Infinium BeadChip, Illumina, San Diego, CA). PLINK v1.07 software [26] was used to implement quality control for the SNPs. For further investigation, only SNPs that matched the following criteria were used in the analyses: a call rate of greater than 95% or a false discovery rate of less than 5%, a missing genotype frequency of less than 5%, a minor allele frequency (MAF) higher than 5%, p-value of Fisher’s exact test for Hardy-Weinberg equilibrium greater than 0.001, and a known position or physical location within an autosomal chromosome. After quality control procedures 8,826 SNPs were retained for genomic analysis.
2.4. Genome-Wide Association Study
The batch effects/stratification of the test input data were corrected using the principal component analysis (PCA) option. A single-locus mixed model was used to perform a GWAS to examine relationships between genotypes of each SNP marker and the variable S/P ratio. The additive model used was y = Xb + Zg + €, where y was a vector of individual S/P ratio, X was the design matrix of fixed effects, Z was the design matrix of random animal additive genetic effects, b was the vector of fixed effects, g was the vector of random effects, and € was the residual effect. The genomic associative analysis was done SNP by SNP using the software SNP Variation Suite version 8.8 (SVSv8).
To account for multiple testing we used the Bonferroni correction, which assumes independency between SNPs, as a criterion to call significant associations (p = α/n = 0.05/8,826 = 5.67 x 10-6). The experiment-wise error was 5% (α = 0.05) and the number of tests (n = 8,826) was taken to be the number of the useful SNPs.
2.5. SNP Validation Genotyping
Two independent populations composed of two hundred and twenty-six (n = 112 and n = 114) 6-month-old gilts from the breed Yorkshire were used to validate the SNPs discovered through GWAS, as candidate markers for PRRS vaccination response (S/P ratio). These SNPs were rs80904326, rs80896559, rs80844350, rs705026086 and rs81358818, within the genes Ring finger protein 144 B (RNF144B), XK related protein 9 (XKR9), Carboxypeptidase Q (CPQ), Forkhead Box P2 (FOXP2) and Basic helix-loop-helix ARNT like 1 (BMAL1), respectively.
The five SNPs selected for the validation study were genotyped using the TaqMan allelic discrimination method and rtPCR (StepOneTM, Applied Biosystems, Foster City CA, USA). The two possible variants at the SNP in a target template sequence were genotyped by using two primer/probe pairs in each reaction. The genotyping PCR reaction was performed by adding the genomic DNA template (2 μl) plus genotyping master mix (5 μl; Thermo Fisher Scientific, Waltham, MA, USA), genotyping custom-made assay mix (0.5 μl; probes and primers; Thermo Fisher Scientific, Waltham, MA, USA) and DNase-free water (2.5 μl).
To prepare the two negative controls, DNase-free water (2.5 μl) was added to each reaction plate instead of genomic DNA for the sample. The cycling parameters were as follows: first, denaturation was performed at 95°C for 10 min, followed by 40 cycles of denaturation at 95°C for 15 s and annealing and extension at 60°C for 60 s. The StepOne Real-Time PCR System (Thermo Fisher Scientific, Waltham, MA, USA) was used to perform the PCR. Finally, the StepOne software v2.3 (Life Technologies Corporation, Carlsbad, CA, USA) was used for PCR data analysis and genotyping.
2.6. Statistical Analysis of the Genotype to Phenotype Validation Study
Descriptive statistics for continuous traits were calculated using PROC MEANS. Assumptions of normality of data distribution and equality of variances were tested using PROC UNIVARIATE and PROC GLM (Levene’s test), respectively. Allele and genotype frequencies were estimated using PROC ALLELE, and the Chi-square test was used to confirm Hardy-Weinberg equilibrium. All analyses were performed with SAS software (Version 9.4; SAS Inst. Inc., Cary, NC).
The SNPs resulting as genomic predictors for the trait S/P ratio were analyzed in a genotype-to-phenotype associative study using a mixed-effects model. This model included the S/P ratio as the response variable, genotype term, female age and herd as fixed effects, body weight as a covariate, and sire as a random effect. If the genotype term was found to be an important (p < 0.05) source of variation in the association analyses for continuous traits, preplanned pairwise comparisons of least squares means were generated with PDIFF. These mean separation tests were executed using the option LSMEANS that included Bonferroni adjustment [27]. The effects of the average allele substitution were also calculated by regressing the phenotype on the number of copies of one SNP allele as a covariate [28].
One-way ANOVA was performed to compare the S/P ratio according to the number of favorable SNP genotypes, whereas the Tukey HSD test was used for pairwise comparisons. Significance between groups was declared at p < 0.05.
3. Results
3.1. Genome-Wide Association Study (GWAS)
The GWAS performed with 8,826 SNPs distributed across the 18 autosomal chromosomes discovered 10 SNPs associated with the S/P ratio, which were located within chromosomes 2, 3, 4, 7, 12, 13 and 18, as presented in Figure 1. All of these SNPs surpassed the correction threshold of the Bonferroni adjustment test (p < 5.67 x 10-6). The SNPs rs81358818, rs705026086, rs343308278, rs80896559, rs80844350, rs80904326 and rs707607708 were intronic variants located within the genes BMAL1, FOXP2, GP9, XKR9, CPQ, RNF144B and SDK1. The SNPs rs331531082 and rs80969120 were intergenic variants located far from the nearest gene (i.e., > 0.1 Mb), whereas the SNP rs3475576322 was a non-coding sequence (Table 1).
3.2. SNP Validation Study
Of the 10 SNPs identified as associated with the S/P ratio in the GWAS, 7 were located within a gene. From these SNPs only 5 met the criteria for minor allele frequency higher than 10% (MAF > 0.10) and no-deviation from Hardy–Weinberg equilibrium (HWE, X2 > 0.05; Table 2). Therefore, these 5 SNPs were considered candidates and suitable for inclusion in a genetic marker validation study.
Least square means for the S/P ratio according to the 5 SNP genotypes are presented in Table 3. The SNPs rs80904326, rs80896559 and rs81358818 were predictors of the S/P ratio in vaccinated gilts, whereas the SNPs rs80844350 and rs705026086 were not associated with this indicator of vaccination response. The genotypes with the highest favorable effect for the SNPs rs80904326, rs80896559 and rs81358818 were GG (2.51 ± 0.09), AA (2.24 ± 0.11) and CC (2.31 ± 0.08), respectively, as they had the greatest antibody response in vaccinated gilts. These results suggested an important (p < 0.01) effect of the genes RNF144B, XKR9 and BMAL1 on the trait S/P ratio observed in gilts after PRRS vaccination.
3.3. SNP Genotype Effects
The favorable genotypes of the SNPs rs80904326, rs80896559 and rs81358818 showed the best antibody response in gilts. Moreover, a significant increment was observed in the trait S/P ratio (p < 0.05) as the number of favorable SNP genotypes increased. The average S/P ratio rises by approximately 0.07 and 0.20 when the number of favorable genotypes increased from 1 to 2 and 2 to 3, respectively (Figure 2).
Interestingly, higher values for the S/P ratio were observed when the favorable genotype of the SNP rs80904326 within the gene RNF144B was present (Figure 3), which indicates the superior contribution of this gene to improve the variable S/P ratio.
Allele substitution and fixed estimated effects for the favorable SNP alleles are presented in Table 4. The SNP rs80904326 in the gene RNF144B had the highest allele contribution (0.301 ± 0.016) for the trait S/P ratio in gilts vaccinated against PRRS virus. However, favorable alleles from the SNPs rs80896559 and rs81358818 also appeared to have a beneficial effect on the evaluated trait. Additionally, an additive fixed effect was confirmed for these SNP markers, suggesting that the sum of their individual effects was equal to their combined allele effects.
4. Discussion
Vaccination against the PRRS virus infection has been the primary strategy to control the virus, but the continuous emergence of new mutant variants, as well as the diversity of the circulating field strains, limits vaccination effectiveness [29]. In addition, the response to vaccination is highly variable among animals, suggesting a genetic approach associated with such a response as a potential alternative strategy to combat and control this disease [19,30].
Recently, genomic technology has been proposed as a tool to explore the genetic architecture associated with the response to vaccination against PRRS virus [7]. GWAS have been successfully used in pigs to identify chromosomal regions and candidate genes associated with complex phenotypes such as response to vaccination [7,18,19]. However, there are no reports on the validation of SNP markers associated with the PRRS vaccination response in gilts through marker-assisted selection.
In the current study, we performed genomic and marker-assisted technologies sequentially to identify and further validate SNPs as potential molecular markers associated with the host immune response in gilts vaccinated against PRRS virus. After using GWAS, we discovered 10 SNPs as predictors of high S/P ratio. Only 5 of these SNPs were selected according to quality criteria and genotyped in two independent gilt populations using TaqMan and rtPCR molecular assays. These SNPs were tested through a genotype-to-phenotype associative statistical analysis that validated 3 of them as molecular markers for the S/P ratio in gilts vaccinated against PRRS virus. Such strategy was also successfully applied by our research group to discover and validate SNPs as markers for heat stress response in ruminants [31,32,33].
Further analyses confirmed the positive and important effect of the favorable alleles and genotypes from the 3 validated SNPs. To our knowledge, this is the first report of genomic SNPs from candidate genes associated with the immune response in gilts after vaccination against PRRS virus, which were further validated as molecular markers in two independent gilt populations. According to Karlsson et al. [34] and Visscher et al. [35], due to the low likelihood that a SNP will be significant in two separate populations, the validation of the SNP effects in independent animal populations appears to be the most reliable method for testing the importance of SNPs as molecular markers.
The whole genome analysis that we performed identified 10 SNPs associated with the S/P ratio, which were located on chromosomes 2, 3, 4, 7, 12, 13 and 18. Of these SNPs, only 5 surpassed the selection criteria (i.e., MAF > 0.10 and HWE > 0.05) and were further tested in a marker associative analysis. Sanglard et al. [10] reported a genomic region on chromosome 7 (∼25 Mb) in the major histocompatibility complex (MHC) area associated with the ratio of positive samples (S/P) in sows vaccinated against PRRS virus. A novel genomic region or quantitative trait loci (QTL) for the S/P ratio was also detected on chromosomes 4, 7, and 9 using haplotype-based GWAS, confirming the QTL previously reported in the MHC region [18]. Similarly, Hickmann et al. [36] identified a QTL within chromosomes 7 and 8 associated with the S/P ratio in Duroc and Landrace sows during a PRRS outbreak.
In our validation study, only 3 SNPs resulted as candidate molecular markers associated with the S/P ratio as they surpassed the Bonferroni correction threshold (p < 5.67 x 10-6). These SNPs were rs80904326, rs80896559 and rs81358818 within the genes RNF144B, XKR9 and BMAL1, respectively. The SNP rs80904326 had an intronic or functional position within the Ring Finger Protein 144B (RNF144B) gene, which expresses ubiquitin activity as its main function. Complex signaling networks are involved in the dynamic activation of the immune system after virus infection, where ubiquitination is an important mechanism modulating both the activation and repression of innate and adaptive immune responses. The RNF144B gene appears to be involved in the innate immune response, which is the first line of defense for the host against invading microbial organisms [37].
The RNF144B gene interacts with TANK binding kinase (TBK1) to inactivate this enzyme, leading to dephosphorylation of interferon regulatory factor 3 (IRF3) and a subsequent reduction in interferon (IFN) synthesis [38]. Then, the RNF144B gene affects the precise control of IFN production, which is required for efficient pathogen clearance without host damage [39], as well as for an effective response to vaccination [40]. IFN is one of the cytokines that plays an important role in the response to viral infections and induces the production of restriction enzymes that suppress the synthesis of viral proteins [41]. IFN also activates signaling pathways responsible for inducing the expression of interferon-stimulated genes (ISGs), which are potent antiviral restriction factors [40,42]. Despite the identification of more than twenty distinct interferon (IFN) genes and proteins, IFN-γ is a notably different member due to its unique receptor activity and distinct intracellular signaling pathway [43]. IFN-γ stimulates immune cell activation, maturation, proliferation, expression of cytokines, and effector function. Interestingly, the PI3K, MAPK/p38, and other cellular pathways have an extensive crosstalk with the IFN-γ signaling pathway [44].
The SNP rs80896559 is in the gene XKR9. This gene encodes apoptosis-inducing protein in cells that exposes phosphatidyl-serine in their membrane as an “eat me” signal. This exposure can occur when cell membrane asymmetry is disrupted, such as in the case of membrane gemmation in enveloped virus release [45]. Phosphatidylserine exposure is one of the general characteristics of PRRS virus-infected cells. A recent study demonstrated that PRRS virus exposes phosphatidyl-serine in its envelope, mimicking apoptosis and infecting cells via the TCR and mucin domain-induced macropinocytosis (TIM), and CD163 as an alternative pathway [46]. It has been reported in studies involving mice that all members of the XKR family, except XKR2, are found in the plasma membrane. Transformed cells expressing XKR9 responded to apoptotic stimuli by phosphatidyl serine exposure and were efficiently engulfed by macrophages. XKR9 possesses a caspase recognition site in the C-terminal region and requires its direct cleavage by caspases for its function [45]. In the current study, we assumed that the SNP rs80896559 in the gene XKR9 was associated with the S/P ratio due to its ability to modulate the immune response after PRRS vaccination, as such a response may be mediated by an apoptotic effect on the PRRS virus induced by the vaccine.
The SNP rs81358818 is within the gene BMAL1, which codes for a protein involved in the regulation of circadian rhythms. This protein is the only component of the mammalian circadian cycle whose deletion in mouse models generates a loss of circadian rhythms. Alterations in circadian rhythms have been found to contribute to the pathogenesis of cardiovascular disease, cancer, metabolic syndromes, aging, and immune response [47]. Silencing of the BMAL1 gene suggested that appropriate functioning of the molecular circadian mechanism is necessary for the immune response to viral infections [48]. The product of the BMAL1 gene has been linked to the control of diurnal oscillations of inflammatory Ly6C monocytes in mice, resulting from diurnal variations of the cytosines involved [49]. The expression patterns of the Ly6C marker in pigs suggested its equivalence to those observed in mice. If the variations in these populations are analogous in pig and mouse, it implies that receptor availability for PRRS virus in pigs may exhibit seasonal and diurnal differences [50].
In the current study, the SNP rs81358818 was detected as a marker for the S/P ratio in PRRS-vaccinated gilts, which appeared to be due to a possible implication of the circadian rhythm in the immune response to immunization against the PRRS virus. This suggested the involvement of seasonal and diurnal variations in the response to PRRS virus vaccination, which appeared to be genetically influenced (i.e., BMAL1 gene).
According to allele substitution effects, the three SNP markers validated in the current study had an important contribution of the favorable allele on the trait S/P ratio. A more detailed analysis of the individual effects of the SNP markers revealed a greater additive contribution of the favorable SNP genotype from the RFN144B gene for the S/P ratio. Interestingly, the average value of this trait improved as the number of favorable genotypes increased, mainly due to the presence of the SNP within the RFN144B gene.
Collectively, our results suggested that the combination of genomic-wide and marker-assisted selection technologies could be an effective and beneficial strategy to identify gilts with high S/P ratios due to an improved response to PRRS vaccination.
A limitation of this study was to use the S/P ratio to evaluate the immune response of gilts vaccinated against PRRS virus; the use of other parameters, such as the evaluation of the neutralizing antibodies or the cellular immune response may be better dependent variables. However, the antibody response evaluated through the S/P ratio can be used as an effective indicator trait of the capacity of gilts to respond against the PRRS virus vaccine. Further studies will probe that these SNP markers are also observed with other immunological parameters.
5. Conclusions
The genetic improvement of the vaccination response against the PRRS virus in gilts could be studied by combining both genomic and marker-assisted technologies. In the current study, three SNPs within the genes RFN144B, XKR9 and BMAL1 were validated as markers for the trait S/P ratio in vaccinated gilts. These genes are involved in regulation of IFN, apoptosis and circadian rhythms, respectively, which appear to modulate the immune host response after vaccination against PRRS virus. We propose these SNPs as molecular markers that should be included in swine selection programs aimed at improving the host immune response to PRRS vaccination. However, further studies including higher-density SNP chips, larger validation populations, as well as others immunological variables are recommended for identifying additional candidate genes and SNP markers within these genes.
Author Contributions
Conceptualization, R.M.E., S.E.S. and P.L.-N; methodology, S.I.-N., X.Z., R.I.R.-A., M.A.S.-C. and M.C.G.-L.; software, X.Z., R.I.R.-A., M.A.S.-C. and G.L.-N.; validation, G.L.-N and P.L.-N.; formal analysis, S.I.-N., R.I.R-A., X.Z., M.A.S.-C. and G.L.-N.; investigation, S.I.-N, J.H. and G.L.-N.; resources, C.M.A.-T., J.R.R.-G. and P.L.-N.; data curation, S.I.-N. and G.L.-N.; writing—original draft preparation, S.I.-N. and P.L.-N.; writing—review and editing, R.M.E., S.E.S., J.H. and P.L.-N.; visualization, S.I.-N. and G.L.-N.; supervision, R.M.E., S.E.S., J.H. and P.L.-N.; project administration, P.L.-N.; funding acquisition, C.M.A.-T., J.R.R.-G. and P.L.-N. All authors have read and agreed to the published version of the manuscript.
Funding
This project was funded by the Instituto Tecnologico de Sonora and the ITSON Swine Diagnostic Laboratory.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, and approved by the Institutional Animal Care and Use Committee of the Instituto Tecnologico de Sonora (Approval code: 2020-0043; 01 May 2020).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author due to they belong to the records of the cooperating swine farms.
Acknowledgments
A very special recognition to Milton G. Thomas, who passed away on February 15, 2024. He was a distinguished and valued collaborator who contributed significantly to this manuscript. We thank the ITSON Swine Diagnostic Laboratory, the cooperating swine farms and the students involved during the project.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- López-Heydecka S.M.; Alonso-Morales, R.A.; Mendieta-Zerón, H.; Vázquez-Chagoyán, J.C. Porcine respiratory and reproductive syndrome: Review. Rev. Mex. Cienc. Pec. 2015, 6(1), 69-68.
- Zimmerman, J.; Locke, A.K.; Ramirez, A.; Schwarts, K.J.; Stevenson, G.W.; Zhang, K. Diseases of swine. First edition. Publisher: John Wiley & Sons, Inc. 2019.
- Nan, Y.; Wu, C.; Gu, G.; Sun, W.; Zhang, Y.; Zhou, E.M. Improved vaccine against PRRSV: Current progress and future perspective. Front. Microbiol. 2017, 8, 1635. [CrossRef]
- Renson, P.; Mahé, S.; Andraud, M.; Le Dimna, M.; Paboeuf, F.; Rose, N.; Bourry, O. Effect of vaccination route (intradermal vs. intramuscular) against porcine reproductive and respiratory syndrome using a modified live vaccine on systemic and mucosal immune response and virus transmission in pigs. BMC Vet Res. 2024, 20(1), 5. [CrossRef]
- Lee, M.A.; Jayaramaiah, U.; You, S.H.; Shin, E.G.; Song, S.M.; Ju, L.; Kang, S.J.; Hyun, B.H.; Lee, H.S. Molecular Characterization of Porcine Reproductive and Respiratory Syndrome Virus in Korea from 2018 to 2022. Pathogens 2023, 12(6), 757. [CrossRef]
- Chae, C. Commercial PRRS modified-live virus vaccines. Vaccines (Basel) 2021, 9(2), 185. [CrossRef]
- Sanglard, L.P.; Hickmann, F.M.W.; Huang, Y.; Gray, K.A.; Linhares, D.C.L.; Dekkers, J.C.M.; Niederwerder, M.C.; Fernando, R.L.; Braccini Neto, J.; Serão, N.V.L. Genomics of response to porcine reproductive and respiratory syndrome virus in purebred and crossbred sows: antibody response and performance following natural infection vs. vaccination. J. Anim. Sci. 2021, 99(5), skab097. [CrossRef]
- Rowland, R.R.; Lunney, J.; Dekkers, J. Control of porcine reproductive and respiratory syndrome (PRRS) through genetic improvements in disease resistance and tolerance. Front. Genet. 2012, 3, 260. [CrossRef]
- Aguilar-Trejo, C.M.; Luna-Nevárez, G.; Reyna-Granados, J.R.; Zamorano-Algandar, R.; Romo-Rubio, J.A.; Sánchez-Castro, M.A.; Enns, R.M.; Speidel, S.E.; Thomas, M.G.; Luna-Nevárez, P. Polymorphisms associated with the number of live-born piglets in sows. Rev. Mex. Cienc. Pec 2020, 11(3), 828-847. [CrossRef]
- Sanglard, L.P.; Fernando, R.L.; Gray, K.A.; Linhares, D.C.L.; Dekkers, J.C.M.; Niederwerder, M.C.; Serão, N.V.L. Genetic analysis of antibody response to porcine reproductive and respiratory syndrome vaccination as an indicator trait for reproductive performance in commercial sows. Front. Genet. 2020, 11, 1011. [CrossRef]
- Li, W.; Wang, Z.; Luo, S.; Wu, J.; Zhou, L.; Liu, J. Genome-wide association analysis and genetic parameters for feed efficiency and related traits in Yorkshire and Duroc pigs. Animals (Basel) 2022, 12(15), 1902. [CrossRef]
- Luan, M.; Ruan, D.; Qiu, Y.; Ye, Y.; Zhou, S.; Yang, J.; Sun, Y.; Ma, F.; Wu, Z.; Yang, J.; Yang, M.; Zheng, E.; Cai, G.; Huang, S. Genome-wide association study for loin muscle area of commercial crossbred pigs. Anim. Biosci. 2023, 36(6), 861-868. [CrossRef]
- Ding, R.; Yang, M.; Quan, J.; Li, S.; Zhuang, Z.; Zhou, S.; Zheng, E.; Hong, L.; Li, Z.; Cai, G.; Huang, W.; Wu, Z.; Yang, J. Single-locus and multilocus genome-wide association studies for intramuscular fat in Duroc pigs. Front. Genet. 2019, 10, 619. [CrossRef]
- Chang Wu, Z.; Wang, Y.; Huang, X.; Wu, S.; Bao, W. A genome-wide association study of important reproduction traits in Large White pigs. Gene, 2022, 838, 146702. [CrossRef]
- Óvilo, C.; Trakooljul, N.; Núñez, Y.; Hadlich, F.; Murani, E.; Ayuso, M.; García-Contreras, C.; Vázquez-Gómez, M.; Rey, A.I.; García, F.; et al. SNP discovery and association study for growth, fatness and meat quality traits in Iberian crossbred pigs. Sci. Rep. 2022, 12(1), 16361. [CrossRef]
- Zhou, F.; Quan, J.; Ruan, D.; Qiu, Y.; Ding, R.; Xu, C.; Ye, Y.; Cai, G.; Liu, L.; Zhang, Z.; et al. Identification of candidate genes for economically important carcass cutting in commercial pigs through GWAS. Animals (Basel), 2023, 13(20), 3243. [CrossRef]
- Boddicker, N.; Waide, E.H.; Rowland, R.R.; Lunney, J.K.; Garrick, D.J.; Reecy, M.; Dekkers, J.C. Evidence for a major QTL associated with host response to porcine reproductive and respiratory syndrome virus challenge. J. Anim. Sci. 2012, 90(6), 1733-1746. [CrossRef]
- Sanglard, L.P.; Huang, Y.; Gray, K.A.; Linhares, D.; Dekkers, J.; Niederwerder, M.C.; Fernando, R.L.; Serão, N. Further host-genomic characterization of total antibody response to PRRSV vaccination and its relationship with reproductive performance in commercial sows: Genome-wide haplotype and zygosity analyses. Gen. Sel. Evol. 2021, 53(1), 91. [CrossRef]
- Dunkelberger, J.R.; Serão, N.V.L.; Weng, Z.; Waide, E.H.; Niederwerder, M.C.; Kerrigan, M.A.; Lunney, J.K.; Rowland, R.R.R.; Dekkers, J.C.M. Genomic regions associated with host response to porcine reproductive and respiratory syndrome vaccination and coinfection in nursery pigs. BMC Genomics 2017, 18(1), 865. [CrossRef]
- Torricelli, M.; Fratto, A.; Ciullo, M.; Sebastiani, C.; Arcangeli, C.; Felici, A.; Giovannini, S.; Sarti, F.M.; Sensi, M.; Biagetti, M. Porcine reproductive and respiratory ryndrome (PRRS) and CD163 resistance polymorphic markers: What is the scenario in naturally infected pig livestock in central Italy?. Animals (Basel), 2023, 13(15), 2477.
- Walker, L.R.; Jobman, E.E.; Sutton, K.M.; Wittler, J.; Johnson, R.K.; Ciobanu, D.C. Genome-wide association analysis for porcine reproductive and respiratory syndrome virus susceptibility traits in two genetic populations of pigs1. J Anim Sci. 2019, 97(8), 3253-3261. [CrossRef]
- He, Y.; Zhou, X.; Zheng, R.; Jiang, Y.; Yao, Z.,; Wang, X.; Zhang, Z.; Zhang, H.; Li, J.; Yuan, X. The Association of an SNP in the EXOC4 gene and reproductive traits suggests its use as a breeding marker in pigs. Animals (Basel), 2021, 11(2), 521. [CrossRef]
- Liu, Z.; Shan, B.; Ni, C.; Feng, S.; Liu, W.; Wang, X.; Wu, H.; Zuofeng, Y.; et al. Optimized protocol for double vaccine immunization against classical swine fever and porcine reproductive and respiratory syndrome. BMC Vet Res. 2023, 19(1), 14. [CrossRef]
- Serão, N.V.; Kemp, R.A.; Mote, B.E.; Willson, P.; Harding, J.C.; Bishop, S.C.; Plastow, G.S.; Dekkers, J.C. Genetic and genomic basis of antibody response to porcine reproductive and respiratory syndrome (PRRS) in gilts and sows. Gen. Sel. Evol. 2016, 48(1), 51. [CrossRef]
- Serão, N.V.; Matika, O.; Kemp, R.A.; Harding, J.C.; Bishop, S.C.; Plastow, G.S.; Dekkers, J.C. Genetic analysis of reproductive traits and antibody response in a PRRS outbreak herd. J. Anim. Sci. 2014, 92(7), 2905-2921. [CrossRef]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.; Daly, M.J.; Sham, P.C. PLINK: a tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 2007, 81(3), 559-575. [CrossRef]
- Weir, B.S. Forensics. Iin Handbook of Statistical Genetics. First Edition. D.J. Balding, M. Bishop, C. Cannings Eds. Publisher: John Wiley and Sons, LTD, New York, NY. 2001. Pp. 275.
- Falconer, D.S.; Mackay, T.F.C. Introduction to quantitative genetics. 4th Edition. Longman Scientific and Technical, New York, NY. 1996. [CrossRef]
- Montaner-Tarbes, S.; Del Portillo, H.A.; Montoya, M.; Fraile, L. Key gaps in the knowledge of the porcine respiratory and reproductive syndrome virus (PRRSV). Front. Vet. Sci. 2019, 6, 38.
- Ko, H.; Sammons, J.; Pasternak, J.A., Hamonic, G.; Starrak, G., MacPhee, D.J., Detmer, S.E., Plastow, G.S., Harding, J.C.S. Phenotypic effect of a single nucleotide polymorphism on SSC7 on fetal outcomes in PRRSV-2 infected gilts. Liv. Sci., 2022, 255, 104800. [CrossRef]
- Luna-Nevárez, G.; Kelly, A.C.; Camacho, L.E.; Limesand, S.W.; Reyna-Granados, J.R.; Luna-Nevárez, P. Discovery and validation of candidate SNP markers associated to heat stress response in pregnant ewes managed inside a climate-controlled chamber. Trop. Anim. Health Prod. 2020, 52, 3457–3466. [CrossRef]
- Luna-Nevárez, G.; Pendleton, A.L.; Luna-Ramirez, R.I.; Limesand, S.W.; Reyna-Granados, J.R.; Luna-Nevárez, P. Genome-wide association study of a thermo-tolerance indicator in pregnant ewes exposed to an artificial heat-stressed environment. J. Therm. Biol. 2021, 101, 103095. [CrossRef]
- Zamorano-Algandar, R.; Medrano, J.F.; Thomas, M.G;, Enns, R.M.; Speidel, S.E.; Sánchez-Castro, M.A.; Luna-Nevárez, G.; Leyva-Corona, J.C.; Luna-Nevárez, P. Genetic markers associated with milk production and thermotolerance in Holstein dairy cows managed in a heat-stressed environment. Biology (Basel), 2023, 12, 679. [CrossRef]
- Karlsson, E.K.; Baranowska, I.; Wade, C.M.; Salmon Hillbertz, N.H.; Zody, M.C.; Anderson, N.; Biagi, T.M.; Patterson, N.; Pielberg, G.R.; Kulbokas, E.J.; et al. Efficient mapping of Mendelian traits in dogs through genome-wide association. Nature Genetics 2007, 39(11), 1321-1328. [CrossRef]
- Visscher, P.M. Sizing up human height variation. Nature Genetics 2008, 40(5), 89-90. [CrossRef]
- Hickmann, F.M.W.; Braccini-Neto, J.; Kramer, L.M.; Huang, Y.; Gray, K.; Dekkers, J.C.M.; Sanglard, L.P.; Serão, N.V.L. Host genetics of response to porcine reproductive and respiratory syndrome in sows: Antibody response as an indicator trait for improved reproductive performance. Front. Gen. 2021, 12, 707873. [CrossRef]
- Liu, X.; Wang, Q.; Chen, W.; Wang, C. Dynamic regulation of innate immunity by ubiquitin and ubiquitin-like proteins. Cytokine & Growth Factor Reviews 2013, 24(6), 559-570. [CrossRef]
- Zhang, Z.; Wang, B.; Zhu, X.; Zhao, L.; Chu, C.; Guo, Q.; Wei, R.; Yin, X.; Zhang, Y.; Li, X. RNF144B inhibits LPS-induced inflammatory responses by binding TBK1. J. Leukoc. Biol. 2019, 106(6), 1303-1311. [CrossRef]
- Crow, M.K. Type I interferon in organ-targeted autoimmune and inflammatory diseases. Arthritis Res. Ther. 2010, 12(Suppl 1), S5. [CrossRef]
- Li, J.; Miller, L.C.; Sang, Y. Current status of vaccines for porcine reproductive and respiratory syndrome: Interferon response, immunological overview, and future prospects. Vaccines (Basel) 2024, 12(6), 606. [CrossRef]
- Budronim, V.; Versteeg, G.A. Negative regulation of the innate immune response through proteasomal degradation and deubiquitination. Viruses 2021, 13(4), 584.
- Ke, H.; Yoo, D. The viral innate immune antagonism and an alternative vaccine design for PRRS virus. Vet. Microbiol. 2017, 209, 75-89. [CrossRef]
- Han, J.; Wu, M.; Liu, Z. Dysregulation in IFN-γ signaling and response: the barricade to tumor immunotherapy. Front. Immunol. 2023, 14, 1190333. [CrossRef]
- Cheon, H.; Wang, Y.; Wightman, S.M.; Jackson, M.W.; Stark, G.R. How cancer cells make and respond to interferon-I. Trends Cancer 2023, 9(1), 83-92. [CrossRef]
- Suzuki, J.; Imanishi, E.; Nagata, S. Exposure of phosphatidylserine by Xk-related protein family members during apoptosis. J. Biol. Chem. 2014, 289(44), 30257-30267. [CrossRef]
- Wei, X.; Li, R.; Qiao, S.; Chen, X.; Xing, G.; Zhanga, G. Porcine reproductive and respiratory syndrome virus utilizes porcine reproductive and respiratory syndrome virus utilizes host cells. J. Virol. 2020, 94(17), e00709-20.
- Li, H.; Li, K.; Zhang, K.; Li, Y.; Gu, H.; Liu, H.; Yang, Z.; Cai, D. The circadian physiology: Implications in livestock health. Int. J. Mol. Sci. 2021, 22(4), 2111. [CrossRef]
- Mazzoccoli, G.; Vinciguerra, M.; Carbone, A.; Relógio, A. The circadian clock, the immune system, and viral infections: The intricate relationship between biological time and host‒virus interaction. Pathogens 2020, 9(2), 83.
- Nguyen, K.D.; Fentress, S.; Qiu, Y.; Yun, K.; Cox, J.S.; Chawla, A. Circadian gene Bmal1 regulates diurnal oscillations of Ly6C(hi) inflammatory monocytes. Science 2013, 341(6153), 1483-1488.
- Fairbairn, L.; Kapetanovic, R;, Beraldi, D.; Sester, D.P.; Tuggle, C.K.; Archibald, A.L.; Hume, D.A. Comparative analysis of monocyte subsets in the pig. J. Immunol. 2013, 190(12), 6389-6396. [CrossRef]
Figure 1.
Manhattan plot showing the SNPs associated with the sample-to-positive (S/P) ratio in gilts vaccinated against PRRS virus (n = 100). The horizontal line corresponds to the Bonferroni adjusted threshold for a significance level of p < 5.67 x 10-6.
Figure 1.
Manhattan plot showing the SNPs associated with the sample-to-positive (S/P) ratio in gilts vaccinated against PRRS virus (n = 100). The horizontal line corresponds to the Bonferroni adjusted threshold for a significance level of p < 5.67 x 10-6.
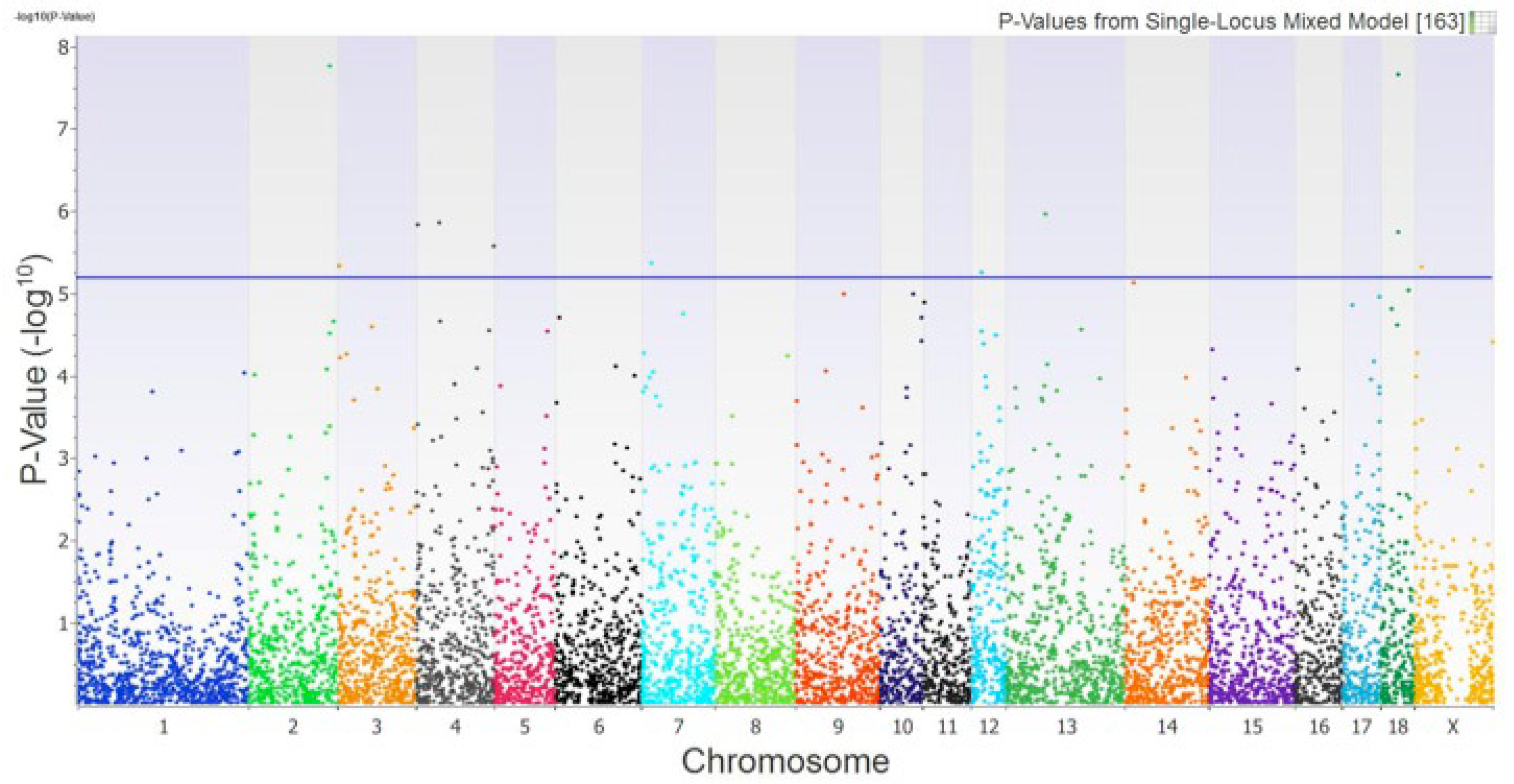
Figure 2.
Average values (± SE) for the variable sample-to-positive (S/P) ratio according to the SNPs with favorable genotype from the genes RNF144B, XKR9 and BMAL1 in two independent gilt populations (n = 226) vaccinated against the PRRS virus.
Figure 2.
Average values (± SE) for the variable sample-to-positive (S/P) ratio according to the SNPs with favorable genotype from the genes RNF144B, XKR9 and BMAL1 in two independent gilt populations (n = 226) vaccinated against the PRRS virus.

Figure 3.
Values for the variable sample-to-positive (S/P) ratio according to the number of favorable SNP genotypes from the candidate genes RNF144B, XKR9 and BMAL1 in two independent gilt populations (n = 226) vaccinated against the PRRS virus.
Figure 3.
Values for the variable sample-to-positive (S/P) ratio according to the number of favorable SNP genotypes from the candidate genes RNF144B, XKR9 and BMAL1 in two independent gilt populations (n = 226) vaccinated against the PRRS virus.
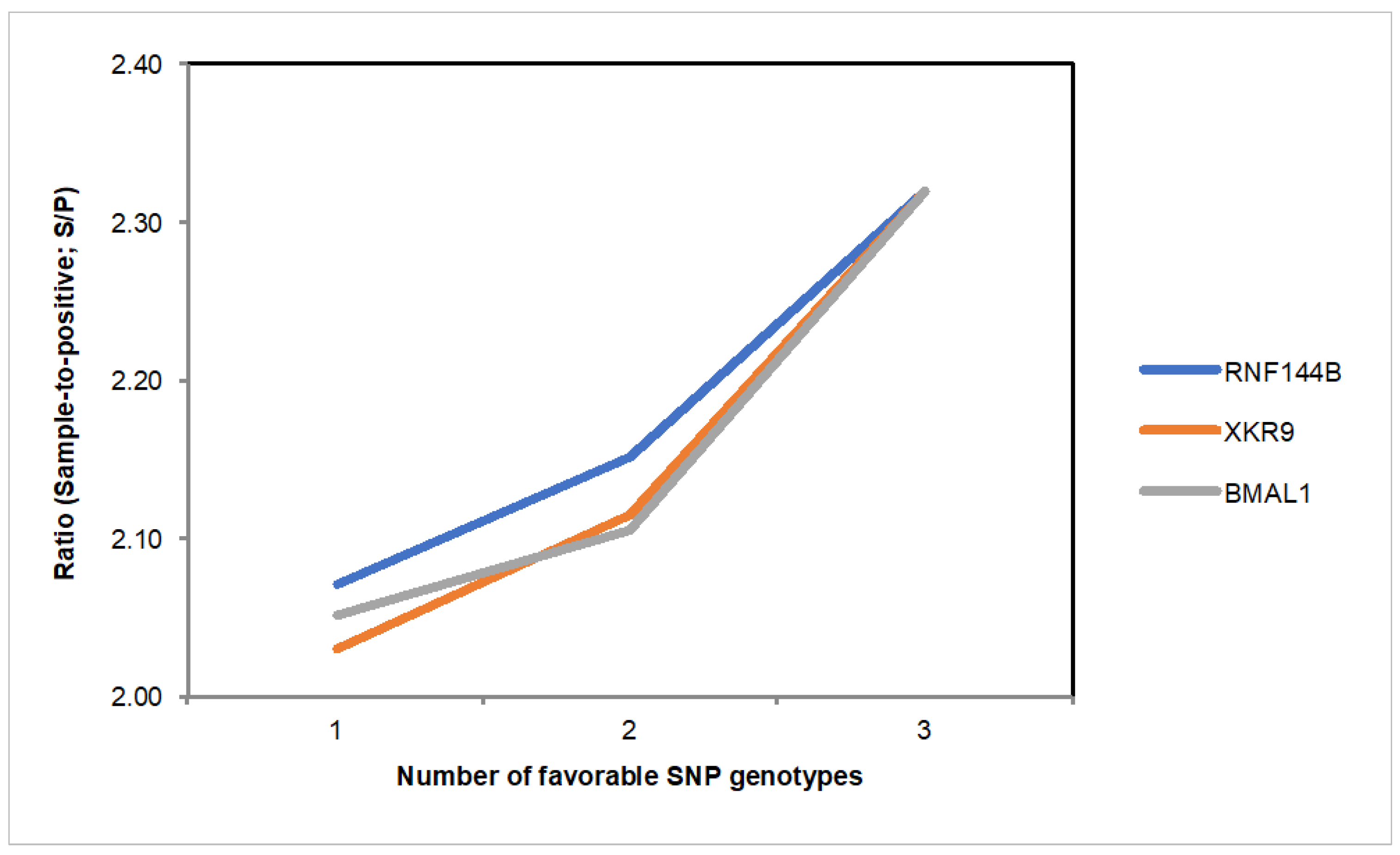

Table 1.
Summary of SNPs after p-Value Bonferroni correction (p < 5.67 x 10-6) resulting from a single-marker genome-wide association study (GWAS) in replacement gilts (n = 100) vaccinated against PRRS virus.
Table 1.
Summary of SNPs after p-Value Bonferroni correction (p < 5.67 x 10-6) resulting from a single-marker genome-wide association study (GWAS) in replacement gilts (n = 100) vaccinated against PRRS virus.
| SNP ID 1 | Variant 2 | BTA 3 | Position 4 | Gene 5 | Alleles 6 | Variance 7 | p-Value 8 |
|---|---|---|---|---|---|---|---|
| rs81358818 | Intronic | 2 | 49’336890 | BMAL1 | C/T | 2.82 | 1.77x10-8 |
| rs705026086 | Intronic | 18 | 31’655298 | FOXP2 | A/G | 2.79 | 2.25x10-8 |
| rs343308278 | 3’UTR | 13 | 71’562221 | GP9 | C/T | 2.19 | 1.12x10-6 |
| rs80896559 | Intronic | 4 | 70’291983 | XKR9 | A/G | 2.16 | 1.40x10-6 |
| rs80844350 | Intronic | 4 | 42’489363 | CPQ | A/G | 2.15 | 1.46x10-6 |
| rs331531082 | Intergenic | 18 | 32’235004 | -------- | C/T | 2.12 | 1.81x10-6 |
| rs80969120 | Intergenic | 4 | 70’092569 | -------- | C/T | 2.06 | 2.67x10-6 |
| rs80904326 | Intronic | 7 | 14’901075 | RNF144B | A/G | 1.98 | 4.36x10-6 |
| rs707607708 | Intronic | 3 | 3’156750 | SDK1 | C/T | 1.96 | 4.73x10-6 |
| rs3475576322 | Non-coding | 12 | 19’663911 | -------- | C/G | 1.85 | 5.63x10-6 |
1 SNP reference of the NCBI; 2 SNP chromosome variant; 3 Sus scrofa autosomal chromosome number; 4 SNP position within the chromosome; 5 candidate gene symbol (BMAL1= Basic Helix-Loop-Helix ARNT Like 1, FOXP2= Forkhead Box P2, GP9= Glycoprotein IX Platelet, XKR9= XK related protein 9, CPQ= Carboxypeptidase Q, RNF144B= Ring finger protein 144 B, SDK1= Sidekick Cell Adhesion Molecule 1); 6 alleles from the SNP; 7 percentage of trait variance explained by the SNP; 8 SNP statistical significance.
Table 2.
Identification, gene name, allele frequency and Hardy-Weinberg equilibrium results for seven SNPs associated with PRRS virus response as S/P ratio in vaccinated gilts (n = 100).
Table 2.
Identification, gene name, allele frequency and Hardy-Weinberg equilibrium results for seven SNPs associated with PRRS virus response as S/P ratio in vaccinated gilts (n = 100).
| SNP ID 1 | Gene 2 | Allele Frequency 3 | HWE Test 4 | HWE p-Value 5 | |
|---|---|---|---|---|---|
| A | G | ||||
| rs80904326 | RNF144B | 0.69 | 0.31 | 0.28 | 0.79 |
| rs80896559 | XKR9 | 0.46 | 0.54 | 1.15 | 0.46 |
| rs80844350 | CPQ | 0.51 | 0.49 | 0.86 | 0.58 |
| rs705026086 | FOXP2 | 0.23 | 0.77 | 0.47 | 0.71 |
| C | T | ||||
| rs81358818 | BMAL1 | 0.47 | 0.53 | 0.75 | 0.64 |
| rs343308278 | GP9 | 0.38 | 0.62 | 24.19 | < 0.01 |
| rs707607708 | SDK1 | 0.97 | 0.03 | 18.73 | < 0.01 |
1 SNP reference of the NCBI; 2 gene symbol name (RNF144B= Ring finger protein 144 B, XKR9= XK related protein 9, CPQ= Carboxypeptidase Q, FOXP2= Forkhead Box P2, BMAL1= Basic Helix-Loop-Helix ARNT Like 1, GP9= Glycoprotein IX Platelet, SDK1= Sidekick Cell Adhesion Molecule 1); 3 frequency of both alleles within gilt population; 4 Hardy–Weinberg equilibrium “χ2” test value; 5 “χ2” test p-value with 1 degree of freedom and α = 0.05.
Table 3.
Least square means ± SE according to SNP marker genotypes for the S/P ratio in two independent gilt populations (n = 226) vaccinated against PRRS virus.
Table 3.
Least square means ± SE according to SNP marker genotypes for the S/P ratio in two independent gilt populations (n = 226) vaccinated against PRRS virus.
| SNP ID 1 | Gene 2 | Least-Square Means by Genotype ± SE 3 | p-Value 4 | ||
|---|---|---|---|---|---|
| AA | AG | GG | |||
| rs80904326 | RNF144B | 1.90 ± 0.03a | 2.36 ± 0.10b | 2.51 ± 0.09b | 0.0009 |
| rs80896559 | XKR9 | 2.24 ± 0.11a | 1.88 ± 0.07b | 1.76 ± 0.08b | 0.0065 |
| rs80844350 | CPQ | 1.96 ± 0.12a | 1.79 ± 0.10a | 1.68 ± 0.14a | 0.4238 |
| rs705026086 | FOXP2B | 1.74 ± 0.06a | 2.01 ± 0.06a | 2.18 ± 0.08a | 0.1875 |
| CC | CT | TT | |||
| rs81358818 | BMAL1 | 2.31 ± 0.08a | 2.14 ± 0.06b | 1.87 ± 0.07c | <0.0001 |
1 SNP reference of the NCBI; 2 symbol name of the candidate gene (RNF144B= Ring finger protein 144 B, XKR9= XK related protein 9, CPQ= Carboxypeptidase Q, FOXP2= Forkhead Box P2, BMAL1= Basic Helix-Loop-Helix ARNT Like 1); 3 least-square means according to SNP genotype ± SE (a,b,c indicate statistical difference among least-square means by genotype in the mixed model at p < 0.05); 4 p-value = statistical significance. .
Table 4.
Allele substitution effect estimates and fixed effect estimates of additive and dominance of the favorable allele for the S/P ratio in two independent gilt populations (n = 226) vaccinated against PRRS virus.
Table 4.
Allele substitution effect estimates and fixed effect estimates of additive and dominance of the favorable allele for the S/P ratio in two independent gilt populations (n = 226) vaccinated against PRRS virus.
| SNP ID 1 | Gene 2 | Allele Substitution Effects | Fixed Estimates Effects | ||||
|---|---|---|---|---|---|---|---|
| F. Allele 3 | p-Value 4 | Estimate ± SE 5 | p-Value 6 | AddE 7 | DomE 8 | ||
| rs80904326 | RNF144B | G | <0.0010 | 0.301 ± 0.016 | <0.0010 | 0.305 | 0.155 |
| rs80896559 | XKR9 | A | <0.0100 | 0.230 ± 0.010 | <0.0080 | 0.240 | 0.120 |
| rs81358818 | BMAL1 | C | <0.0001 | 0.216 ± 0.012 | <0.0001 | 0.220 | 0.050 |
1 SNP reference of the NCBI; 2 symbol name of the candidate gene (RNF144B= Ring finger protein 144 B, XKR9= XK related protein 9, BMAL1= Basic Helix-Loop-Helix ARNT Like 1); 3 SNP allele with a favorable effect on the phenotype; 4 p-values obtained from allele substitution analysis in SAS which included the term genotype as a covariate; 5 estimates of the effect expressed in units of the traits ± standard error; 6 p-values for fixed effects obtained from the substitution of favorable allele analysis that included the genotype term as fixed effect; 7 the additive effect estimated as the difference between the 2 homozygous means divided by 2; 8 the dominance effect calculated as the deviation of the heterozygous from the mean of the 2 homozygous.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



