
Submitted:
31 December 2024
Posted:
31 December 2024
You are already at the latest version
Abstract
In the Caleri lagoon, a coastal lagoon in the Po River Delta, Northern Adriatic, the transplant of the dwarf eelgrass Zostera noltei was used as a nature-based solution to attempt the ecological restoration of a previously depleted lagoon area. A total of 135 15 cm diameter sods were transplanted, with the donor site at the Venice lagoon. Using unmanned aerial vehicles (UAVs), eelgrass transplants have been mapped and monitored with great precision. After 2 years, the area covered by eelgrass increased from the initial 2.5 m2 to 60 m2. Changes in the community structure and on the frequency of biological traits of macrobenthos occurred at the transplant site, with a higher frequency of epifaunal predators and herbivores, of organisms with longer life spans and larger body sizes. Sensitive and indifferent taxa were always higher in the transplant site than in the bare bottom control site, where opportunistic taxa continued to dominate. Ecological quality status measured through M-AMBI and HBFI indices showed a clear improvement in the transplant site. The rapid changes in benthos demonstrate that even relatively small-scale transplantation of dwarf eelgrass can restore faunal communities very rapidly.
Keywords:
Nature-based solution
; Zostera noltei
; Macrobenthos
; Biological traits
; Unmanned aerial vehicles
1. Introduction
The regression and loss of seagrass meadows from coastal and transitional waters due to human activities is a problem of global concern [1]. Seagrass loss has a significant impact on coastal biodiversity, leading to alterations in food webs and depletion of harvestable resources [2]. Seagrass meadows provide complex three-dimensional structures for benthic organisms and fish, counteract erosion by retaining sediment, remove nutrients from the water column, sequester carbon (Blue Carbon) by mitigating atmospheric CO2 and ocean acidification [3]. Seagrass meadows are included among the internationally protected species (Bern Convention and SPAMI Barcelona Convention), and regulatory initiatives (European Habitats Directive and European Water Framework Directive) have been promoting their conservation. In line with the UN Decade on Ecosystem Restoration (2021-2030), and the new European Nature Restoration Law, restoration has been promoted to counteract habitat loss and degradation, and several initiatives have been undertaken to facilitate the reintroduction of seagrass meadows to sites where they were formally present [4, 5].
Effective monitoring actions are essential for understanding the dynamic nature of seagrass meadows and evaluating their response to restoration efforts. Recent advances in remote sensing technologies, particularly the use of Unmanned Aerial Vehicles (UAVs), have enhanced the ability to monitor and map seagrass meadows with unprecedented accuracy [6, 7]. UAVs provide high-resolution spatial data that enable researchers to rapidly and cost-effectively assess species, composition, coverage, and biomass over large areas. As valuable complement to traditional in situ methods, UAV-based monitoring offers repeatability, accessibility to remote areas, and temporal flexibility [8, 9].
The LIFE19 NAT/IT/000264-TRANSFER is a restoration through nature-based solutions project funded by the European Union, whose aim is to favor seagrass recolonization in the lagoons of the Po River Delta (Italy), of the Amvrakikos Gulf (Greece), and in Mar Menor (Spain). Within this framework, seagrass transplantations were carried out in the Caleri lagoon, a waterbody located in the norther sector of the Po Delta (Northwestern Adriatic Sea). In the past, lagoons and ponds of the Po Delta hosted quite extended seagrass meadows [10, 11], but, from the mid-1980s onwards, they have almost disappeared because of increased eutrophication [12]. Probably thanks to the implementation of the management measures provided for by the Water Framework Directive and other relevant directives (e.g. the Nitrate Directive), in turn implemented by national legislative decrees (D.Lgs 152/1999, 152/2006; 260/2010; 172/2015), a gradual decrease in eutrophication has been observed along the coastal area of the Northern Adriatic Sea [13]. Therefore, in a restoration through nature-based solution perspective, we have recently identified limits and parameters of water and sediments that define the suitability of a recipient site for the rooting of seagrasses, in order to increase the chances of success of transplantation operations [14]. An ex-ante monitoring of water and sediment parameters, carried out at the beginning of the TRANSFER project in 2021, allowed us to identify an area in the Caleri lagoon where the restoration actions could be carried out with chances of success. A UAV survey was conducted in the transplant area employing a lightweight drone equipped with a high-resolution RGB camera. Dedicated flights were planned and realized to perform an initial evaluation of seagrass distribution after the sods transplants.
The aim of this work is twofold: i) to assess the potential of Zostera noltei Hornemann, 1832, as a nature-based solution to restore a previously depleted habitat; ii) to assess the contribution of transplanted Z. noltei meadows to the improvement of the ecological quality status (sensu European Water Framework Directive) and local biodiversity. If ecological quality status and local biodiversity resulted improved in the transplanted area, this will also highlight the role of lagoonal seagrasses, which have very different characteristics compared to other seagrass (e.g., Posidonia oceanica) in ecosystem recovery, particularly in the restoration of those habitats.
2. Materials and Methods
2.1. Study Area and Seagrass Transplantation
The Caleri lagoon (Figure 1) has a surface area of about 9.8 km2, an average depth of 1.5 m, and is separated from the sea by a large dune belt interrupted by a short, 120-150 m-wide canal, through which the lagoon communicates with the sea. Fresh water supplies are scarce, and mostly come from the irrigation water used in the area surrounding the lagoon. The transplant area (45.092579°; 12.327251°) is located on a silty-sandy seabed, with an average depth of 0.5 m, a few hundred meters from the marine canal.
The dwarf eelgrass Zostera noltei was transplanted as sods, since the root and rhizome system remains relatively intact and also provides a reserve of the original rooting medium [2, 5]. Transplants were carried out in two periods [2], namely in autumn 2022 and late spring 2023. The donor site was the Venice Lagoon, where seagrass meadows cover a surface of thousands of hectares [15] and, consequently, the removal of some dozen of sods did not constitute damage to the ecosystem [2]. A 15 cm diameter corer was used to collect the seagrass and the underlying sediment to a depth of ~20 cm; after collection, sods were placed in perforated buckets, in turn immersed in larger baskets in order to remain moist, and immediately transported by road to the recipient site, the Caleri lagoon, about 60 km far. Here sods were manually positioned in the seabed with a corer during low tide (Figure 2a), according to a scheme that has proven effective in the previous LIFE SERESTO project [16]: sods were transplanted in 3 groups of 3 sods (triplets), each approx. 1 m from each other, and the 3 groups of sods in turn were spaced approx. 5 m (Figure 2b). Total sods transplanted were 90 (10 plots consisting of 3 triplets each) in November 2022, and 45 (5 plots consisting of 3 triplets each) in June 2023.
2.2. Sampling Environmental Parameters and the Biota
The main physical-chemical parameters of the water column [temperature; pH; Eh; salinity; dissolved oxygen (DO); dissolved inorganic nitrogen (DIN), as sum of nitrite, nitrates and ammonium; reactive phosphorus (RP); reactive silicates (RSi); total suspended solids (TSS); total chlorophyll-a (Chl-a tot), as sum of chlorophyll-a and pheophytin-a] and sediment [pH; Eh; total nitrogen (Ntot), carbon (Ctot) and phosphorus (Ptot); the fine fraction <63 μm (Fines); moisture; density] were analyzed following the procedures described in [17, 18, 19], from January to December 2023, with monthly frequency.
On field monitoring of seagrass growth was carried out during daylight low tides in August 2023 by means of UAV surveys (see paragraph 2.4), and in June 2024 by means of ground surveys, measuring the diameter of each patch covered by the eelgrass meadow (previously consisting of a triplet of sods) with a metric cord.
The biota (macrobenthos and fish fauna) was sampled following a BACI (Before-After, Control-Impact) design, that is sampling, before and after the transplants, in a control site (45.096143°; 12.322183°) approximately 200 m from the transplant area, and in the area subjected to the restoration. Macrobenthos and fish fauna were sampled in June 2021 (Before), and then in January, June, September 2023, and June 2024 (After). Macrobenthos was sampled with a Van Veen grab (4 L volume, 5 replicates at each area), sieved at 0.5 mm and preserved in 96° ethanol (Carlo Erba Reagents). In the laboratory, animals were carefully sorted and identified at the species level. Fish fauna was sampled using a beach seine (internode distance 2 mm in the central bag and 4 mm in the wings), 10 m long and 2 m high in the center. Two replicates per sampling area (transplant and control) were carried out. Fish were anesthetized in ice, then in the laboratory they were identified at the species level, counted and weighed (±0.001 g).
2.3. Data Analysis and Ecological Quality
Shannon-Wiener diversity index on log2 basis (H’) and Pielou index (J’), were calculated for macrobenthic community at each sampling date. The variability in community structure was examined by means of non-metric multidimensional scaling (MDS) ordination on the similarity matrix constructed using the Bray-Curtis index calculated on untrasformed abundance data. Data were analyzed using the PRIMER v.6 software package [20].
Biological trait categories of macrobenthos were chosen related to aspects of life history and habits of the benthic fauna [21]. These categories were feeding mode, adult life habitat, adult body size, and life span. Each trait was subdivided into a number of modalities (the different trait categories displayed by organisms.) ranging from 2 to 4, for a total of 12 variable modalities. The affinity of each species for different modalities was measured using a fuzzy coding method (range 0–3), where 0 denoted full non-correspondence and 3 denoted substantial correspondence. For example, the 4 modalities of the trait feeding mode for macrobenthic species are: predator, herbivorous, deposit-feeder, and filter-feeder. After choosing the functional traits and the modalities to be considered (Table 1) the “taxa by traits” matrix was compiled [21]. We also considered the ecological groups (EG) proposed for applying AMBI [22] as ecological traits, with each group (i.e., sensitive, tolerant, etc.) representing a modality of a particular trait.
The ecological status (ES) through the biological quality element “macrobenthos” was assessed by applying the index M-AMBI [23] on the species/abundance data set. The M-AMBI index is based on a multivariate analysis in which factor analysis combines the values of AMBI [22], with those of Shannon-Wiener diversity (H’) and number of species (S). The M-AMBI is calculated by means of the dedicated software (www.azti.es). According to the Italian D.Lgs 260/2010, ecological quality ratio (EQR) boundaries between ES classes are: High/Good=0.96, Good/Moderate=0.71, Moderate/Poor=0.57, Poor/Bad=0.46.
The ES through the biological quality element “fish fauna” was assessed by applying the index Habitat Fish Bio-Indicator (HFBI) [24], the multi-metric fish index adopted in Italy, whose EQR boundaries between ES classes are: High/Good=0.94, Good/Moderate=0.55, Moderate/Poor=0.33, Poor/Bad=0.11 [24].
2.4. UAV Survey
The UAV survey of the Caleri lagoon targeted a ~0.5 ha area where seagrass sods were transplanted. The objective was to obtain high-resolution imagery to enable georeferenced orthomosaic generation for the localisation of transplanted seagrass. The theoretical Ground Sample Distance (GSD) of ~2 mm/pixel is significantly degraded due to water presence, ripple effects, and transparency, leading to an expected realistic subcentimetric resolution. Flight parameters were configured to ensure an image overlap of around 80%, both vertically and horizontally, necessary for thorough post-processing. The surveys employed a DJI Air 2S UAV equipped with a 1-inch CMOS RGB camera (20 MP resolution, 5472 × 3648 pixels, 22 mm focal length). A polarizing filter was used to reduce water surface reflections. The drone, with an approximate flight autonomy of 20 minutes, operated on automated flight plans designed to optimize coverage and data acquisition.
Flights were conducted on August 6, 2023 (after the second transplant period) during low tide and optimal weather conditions, with a water level less than 50 cm, ensuring good visibility of submerged sods. A preliminary flight, covering an area of ~5 ha, was performed at 50 m of altitude, with a 10 m track spacing and a speed of 5 m/s, to provide a wider overview of the area (Figure 3a; b). Four detailed flights were planned to fully cover the transplant area while adhering to the drone’s battery limitations. The UAV flew at an altitude of 7 m with a track spacing of 2 m and a speed of 1 m/s. Images were captured at 0.5 Hz, achieving a vertical and a horizontal overlap exceeding 70% and 80% respectively.
3. Results
3.1. Environmental Parameters and Biota
Table 2 reports the mean (with standard deviation), minimum and maximum value of water and sediment parameters at the transplant site, from January to December 2023. Values shown in the Table, particularly those of TSS, Chl-a tot and nutrients, confirm that the chosen site was suitable for seagrass transplantation, according to the limits described in [14].
Overall, our transplanting effort added initially about 2.5 m2 of plant cover to the 0.5 ha-wide transplant area. Ground measurements taken in June 2024 provided a rough estimate of the eelgrass meadow extent of 60 m2: in each triplet, the individual sods were no longer recognizable but had formed continuous meadow patches, the diameter of which varied between about 1.5 and 3 m, showing a satisfactory plagiotropic expansion of the rhizomes and the leaf system.
A total of 115 benthic species was collected in the study period (Table S1). Consequently the matrix “taxa by traits” had 115 rows (number of taxa) and 12 columns (total number of modalities). The most abundant group was Annelida (46 taxa), followed by Arthropoda (39 taxa), Bivalvia (20 taxa), Gastropoda (9 taxa), and other less represented groups (Nemertea, Echinodermata, Ascidiacea). Diversity and evenness values are shown in Figure 4. From January 2023 onwards, both indices showed slightly higher values in the transplant site.
The MDS ordination (stress=0.06) for the macrobenthic community at the 2 sites is shown in Figure 5. The plot appears to highlight 3 basins of attraction, i.e. regions in state space in which the system tends to remain [25]: a region corresponding to bare seabed and warm period (right of the plot, red circle), one corresponding to the autumn-winter period (center of the plot, orange circle), and one corresponding to a new partially vegetated state and warm period (left of the plot, green circle).While the community at the control site (C) shows a sort of cyclical trend in its structure and composition, with a certain degree of modification apparent in June 2024, the community at the transplant site (T) shows points conforming rather closely to a linear sequence, suggesting that the community is undergoing directional changes in its structure towards a different basin of attraction that are not dependent (only) on seasonality.
The frequencies of modalities within each biological trait are shown in Figure 6. It is evident that, particularly at the transplant site, the relative frequencies of the traits of the benthic community are changing: greater frequency of epifaunal predators and herbivores, and relative decrease of infaunal deposit-feeders, and a greater frequency of organisms with longer life spans and larger body sizes.
Table 2 shows the distribution of macrobenthos into ecological groups (EG) according to their sensitivity to disturbance [24]. In June 2021, the benthic communities at sites T and C showed the same structure, with the clear dominance of tolerant taxa (EG-III), and the small presence of sensitive taxa (EG-I). Apart from some seasonal fluctuations due to the alternation of species more or less favored by temperature, it is quite evident that, from January 2023 onwards, the percentage of sensitive (EG-I) and indifferent (EG-II) taxa was always greater at the transplant (T) than at the control (C) site, while the amount of second order (EG-IV) and first order (EG-V) opportunistic species was constantly greater at C than at T.

Table 2.
Macrobenthic community composition at the two sites by ecological groups.
| Date | Site | Ecological Groups | ||||
| I(%) | II(%) | III(%) | IV(%) | V(%) | ||
| June | C | 2.4 | 1.7 | 92.9 | 0.7 | 2.3 |
| 2021 | T | 4.1 | 1.6 | 90.8 | 1.2 | 2.4 |
| January | C | 0 | 3.1 | 20.6 | 21.6 | 54.7 |
| 2023 | T | 0.6 | 0.2 | 53.4 | 24.1 | 21.7 |
| June | C | 4.1 | 12.9 | 71 | 3.1 | 8.8 |
| 2023 | T | 25 | 23.7 | 41 | 9.6 | 0.6 |
| September | C | 0.0 | 0.6 | 20.4 | 30.9 | 48.1 |
| 2023 | T | 4.0 | 11.3 | 45.2 | 14.8 | 24.6 |
| June | C | 22.4 | 1.7 | 18.1 | 7.7 | 50.1 |
| 2024 | T | 26.1 | 5.2 | 35.5 | 15.2 | 18.1 |
In Figure 7a the values of the M-AMBI index at the transplant (Cal-Tr) and control (Cal-C) sites are shown. The ecological quality, similar for both sites in June 2021, i.e. before the start of transplants, was, from January 2023 onwards, always better at the Cal-Tr site, with values always above the Moderate/Good threshold (green line on Figure 3) defined by D.Lgs 260/2010.
A total of 17 fish species was collected in the study period (Table S2). Lagoon resident species (gobies, pipefish, blennies, and sand smelts) accounted for a greater proportion of total abundance at both transplant and control sites, followed by marine migrant species (mullets, sea bass, herrings). In Figure 7b the values of the HFBI index at the transplant (Cal-Tr) and control (Cal-C) sites are shown. Although the Good quality status has been achieved only in the summer period, it seemed that the presence of seagrass patches attracted a richer and more diverse fish community than the bare seabed.
3.2. Processing and Analysis of UAV Data
A total of 2380 images were captured during UAV surveys, including 630 from a preliminary flight and 1750 from detailed flights conducted at an altitude of 7 meters. These images were processed using Agisoft Metashape Pro to generate georeferenced orthomosaics aligned with satellite imagery, as underwater target placement was not feasible. Although the theoretical GSD of ~2 mm/pixel was influenced by surface ripples and water clarity, the resulting orthomosaics maintained subcentimetric resolution, enabling the detection, enumeration, and georeferencing of the plants. Visual inspection of the high-resolution orthomosaics, combined with prior knowledge of the transplantation locations, enabled accurate calibration of the method and precise mapping and identification of transplanted seagrass sods, facilitating detailed structural analysis. Of the 135 transplanted sods (90 in November 2022 and 45 in June 2023), 102 (75.5%) were identified and georeferenced (Figure 8). The high-resolution of the acquired images permitted to identify clearly the single sods and to measure the diameters (Figure 9).
The frequency histogram (Figure 10) computed after the measurements, revealed a positive asymmetry of the distribution. The measured diameters ranged from a minimum of 3 cm (indicative of a decrease in leaf density of the sod) to a maximum of 66 cm (indicative of a growth in leaf cover of more than 4 times). Overall, the area covered by Z. noltei in August 2023 was estimated to be 3.8 m2.
4. Discussion
Seagrass meadows form highly productive coastal habitats, and provide, among others, fundamental ecosystem supporting services for biological diversity maintenance. In the Caleri lagoon, the results of a restoration through nature-based solution support this notion for Zostera noltei meadows, as improving effects on the biota appear to occur after less than 2 years. Z. noltei has been recently successfully transplanted in the Venice lagoon [16], and in the NW Atlantic coast of Portugal [26]. The results of the present study are also noteworthy for the contribution they can make to the development of seagrass transplant techniques. In fact, unlike other restoration interventions, such as the SERESTO project [16], the donor site was not in the same water basin as the recipient site, but rather about 60 km away. The sods were collected in the Venice lagoon, transported by car to the Caleri lagoon, and finally transplanted onto the bare seabed. Over 75% of the transplanted sods survived and took root, which proves that the technique used is effective. Prior to this intervention, the site had no vegetation cover: less than 1 year after transplants (August 2023), this increased to 3.8 m2 of seagrass meadow. Two years after transplants (June 2024) the area covered by eelgrass meadows was roughly estimated to be 60 m2: Figure 11 shows the continuous meadow formed by the plagiotropic growth of a triplet of sods. This indicates that the original transplant effort allowed for substantial growth and expansion of eelgrass at the receiving site.
Development of seagrass cover is strongly influenced by physical and ecological processes: insufficient light availability, inappropriate sediment granulometry, nutrient (N, P) and organic matter content, pore water sulfide concentration, and competition with micro- and macroalgae, are all known to affect the growth and expansion of seagrasses beds [14, 27]. The chemical-physical monitoring carried out monthly for a whole year confirmed that the characteristics of the site chosen for the reintroduction of Z. noltei are adequate to support the growth and development of the eelgrass meadow.
The time elapsed since the seagrass transplants is probably still too short to observe substantial changes in the structure and composition of biotic communities. However, although the transplant area is still made up of individual patches of meadow, both the macrobenthic community and the fish community have begun to structure differently where the meadow is present. The macrobenthic community showed changes in its structure and composition, and although some community attributes, such as diversity and evenness, were not much higher than the bare site, the community seems clearly directed towards a different basin of attraction. At the transplant site the community is undergoing a succession from a community dominated by opportunistic (EG-IV and EG-V) and tolerant (EG-III) species, to more sensitive species (EG-I and EG-II) [28]. This translated into an improvement in the ecological quality status measured by M-AMBI at the transplant site, which showed an ES always higher than Good even when at the control site, a couple of hundred meters away, the ES was Moderate.
Despite possible variations due to seasonality and the success or failure in recruiting the various species, at the transplant site some biological traits have shown a trend of change over time from June 2021 to June 2024 (Figure 5). In particular, a general change in the proportion of feeding guilds was observed, with a decrease of the proportion of deposit feeders (DepF, from 92% to 41%), and the increase of the proportion of herbivorous (He, from 0.7% to 35.4%), and predators (P, from 1.2% to 13.4%). A significant recruitment of filter-feeder organisms (mostly Serpulid worms) was observed at both T and C sites in June 2023 (FilF, T: 30.7%; C: 59%), but by September 2023 their proportion had already significantly decreased, particularly at the bare sediment site C (FilF, T: 25.5%; C: 1.3%). At site T, the proportion of epifaunal organisms increased (Epif, from 1.2% to 44.3%), as well as life span (LS/M, from 1.8% to 45.2%), and body size (BS/M, from 1.4% to 48.8%). Habitat heterogeneity can influence the composition of biological traits of macrobenthos, as species are selected for traits suitable for survival in that particular habitat [29]. In the Caleri lagoon the composition of biological traits of macrobenthos in the vegetated and bare bottom habitats was found, after less than 2 years, to be already undergoing diversification. Thus, the observed differences in biological traits of different habitats were a reflection of differences in species composition of macrobenthic communities. Moreover, the differences in the relative distributions of biological traits between the 2 sites (patchily vegetated and bare sediment) habitats are in agreement with the theoretical references of the “Habitat Model” [30], which hypothesizes that trait composition is influenced by environmental conditions that determine species-specific traits in particular habitats, and consequently shape the species composition of local communities [31].
Fish assemblages are arranged in response to cyclic variations mostly driven by seasonality, and this may have partially affected the result of the application of the HBFI index. For marine migrating species, for example, in winter the lagoon mainly exerts the function of feeding ground, while during the other seasons the lagoon exerts both nursery and feeding functions [32]. Shallow-water fish communities at our sites resulted more diversified at the end of spring, when both lagoon resident and marine migrant species coexisted, and were found to be dominated, in terms of fish abundance, by 4 families: Gobiidae, Syngnathidae, Mugilidae, and Atherinidae. These families have already been identified as those that mainly characterize the fish community in temperate coastal lagoons and estuaries [33]. Two of these families, Syngnathidae and Gobiidae, were found to be more present in the area where seagrass transplants were carried out, compared to bare bottoms. Despite seasonal variations in fish species and abundance, the application of the index HFBI resulted, on average, in better ES classification of the transplant area. Habitat heterogeneity contributes to the variability of fish assemblages, and the transplant area was found to host more diversified fish communities, with the presence of species typically associated with seagrass meadows [33], and characterized by higher proportions of lagoon resident species [34].
UAV monitoring provides significant benefits for marine ecosystems, offering high spatial and temporal resolution for precise, non-intrusive data collection. UAVs are cost-effective for small areas (i.e., few hectares), flexible, and can safely access remote locations. They enable frequent, repeatable surveys, enhancing monitoring efficiency in time-sensitive studies. Key advantages include rapid coverage of vast areas, high spatial resolution, and real-time data for early detection of plant stresses such as drought, diseases, and nutrient deficiencies. Their adaptability to remote areas, combined with advancements in imaging and machine learning, facilitates precise analysis and decision-making. UAVs provide high-resolution, georeferenced data, not only on the seagrass but also on the overall context, and frequent monitoring capabilities, ideal for capturing fine-scale temporal changes considering the context of operations. This offers a comprehensive overview which is often not possible with ground surveys. However, UAV monitoring has limitations. It’s less effective for large-scale applications due to battery life and regulatory constraints like altitude and line-of-sight requirements. Weather conditions can disrupt operations, and the high-resolution data need substantial processing capacity. UAVs must operate during low tide for optimal water clarity, and species discrimination depends on water surface conditions. Although cost-effective long-term, UAVs require significant initial investments in equipment and training. Processing large volumes of high-resolution data necessitates expertise and computational resources, making ground-based methods more feasible in some scenarios [35, 36, 37].
Several uncertainties were inherent in the UAV survey methodology. Verification of the GSD through underwater target placement was not feasible, and advanced georeferencing corrections, such as RTK or LiDAR, were not employed. However, based on direct analysis of the UAV imagery and corroborated by findings in the literature, where degradation between theoretical and observed GSD is reported to be approximately a factor of two [38], even accounting for surface ripple, water turbidity, and orthomosaic construction artifacts, our resulting sub-centimeter resolution is sufficient to delineate the characteristic leaf structures of seagrass species. Georeferencing accuracy, constrained to standalone GNSS without ground control points and referenced to satellite-derived imagery, provided an estimated absolute orthomosaic accuracy on the metric order [39]. While this level of absolute accuracy could be optimized, it is sufficient for determining the relative spatial arrangement of vegetation within the study area, thereby meeting the objectives of this research. Advancements in artificial intelligence (AI) offer promising solutions for automated plant recognition and health assessment, particularly for submerged vegetation like seagrass meadows. AI-driven models, such as convolutional neural networks (CNNs), can process high-resolution UAV imagery to identify species, detect stress signals, and evaluate vegetation health with precision. UAVs, as optimal data acquisition platforms, provide extensive datasets critical for CNN training, significantly enhancing classification accuracy. Real-time integration of UAV imagery and AI algorithms could enable rapid analysis and dynamic decision-making in field applications. With over 90% accuracy already demonstrated in precision agriculture for crop disease detection [40, 41], AI shows strong potential for monitoring and managing seagrass ecosystems.
5. Conclusions
The results of this study confirm that habitat restoration through nature-based solutions, such as Zostera noltei transplant, is an effective tool to mitigate previously impoverished habitats, to recover lost biodiversity, and to improve the ecological quality status as required by the European Water Framework Directive. Less than 2 years after the restoration interventions, our results have shown clear positive signs of recovery of benthic community. Despite the encouraging results highlighted in this study, the benthic community of the transplanted area still needs time to reach the level of complexity typical of seagrass meadows, with the food web dominated by key functional groups such as herbivores and predators. Increasing biodiversity will be essential to provide stability in the restored area by increasing functional redundancy [42] and stimulating ecosystem functioning [43].
It is important to highlight that the benefits of restoration through the reintroduction of Z. noltei must be evaluated on a longer time scale: restoration times in estuarine and coastal systems require more time than the one we considered [44], so it would be essential to continue monitoring in the following years to detect trends that were not yet well defined after less than 2 years from transplants.
Supplementary Materials
The following supporting information can be downloaded at: Preprints.org, Table S1: Macrobenthos species list; Table S2: Fish species list.
Author Contributions
Conceptualization, M.M., A.S.; methodology, C.M., F.M., A.S.; formal analysis, C.M., V.S., A.S.; investigation, V.S., F.C., C.C., M.G.P., M.P., M.Pa., A.A.S., M.A., E.C., N.I.E., K.G.C.R.; data curation, C.M., V.S.; writing—original draft preparation, M.M., F.B.; funding acquisition, M.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was partly funded by the European Union’s LIFE financial instrument (grant LIFE19 NAT/IT/00026 TRANSFER), and by the University of Ferrara through the “Fondo per l’Incentivazione alla Ricerca” (FIR-2021).
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
We wish to thank Ms. Maria Pullara and Ms. Virgilia Marcato for valuable work at sea.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Orth, RJ, Carruthers, TJB, Dennison, WC, Duarte, CM, Fourqurean, JW, Heck, KL Jr., Hughes, AR, Kendrick, GA, Kenworthy, WJ, Olyamik, S, et al. A global crisis for seagrass ecosystems. BioScience 2006, 56:987–996.
- Sfriso, A., Buosi, A., Facca, C., Sfriso, A.A., Tomio, Y., Juhmani, A. S., et al. Environmental restoration by aquatic angiosperm transplants in transitional water systems: The Venice Lagoon as a case study. Sci. Total Environ. 2021, 795, 148859. [CrossRef]
- Sousa, A.I., da Silva, J.F., Azevedo, A., Lillebø, A.I. Blue Carbon stock in Zostera noltei meadows at Ria de Aveiro coastal lagoon (Portugal) over a decade. Sci. Rep. 2019, 9, 14387. [CrossRef]
- Curiel, D., Kraljevic Pavelic, S., Kovacev, A., Miotti, C., Rismondo, A. Marine Seagrasses Transplantation in Confined and Coastal Adriatic Environments: Methods and Results. Water 2021, 13, 2289. [CrossRef]
- Costa, V., Flindt, M.R., Lopes, M., Coelho, J.P., Costa, A.F., Lillebø, A.I., Sousa, A.I. Enhancing the resilience of Zostera noltei seagrass meadows against Arenicola spp. bio-invasion: a decision-making approach. J. Environ. Manag. 2022, 302. [CrossRef]
- Hossain, M.S.; Bujang, J.S.; Zakaria, M.H.; Hashim, M. The application of remote sensing to seagrass ecosystems: an overview and future research prospects. Intl. J. Remote Sensing 2014, 36, 61–114. [Google Scholar] [CrossRef]
- Veettil, B.K.; Ward, R.D.; Lima, M.D.A.C.; Stankovic, M.; Hoai, P.N.; Quang, N.X. Opportunities for seagrass research derived from remote sensing: A review of current methods. Ecol. Ind. 2020, 117. [Google Scholar] [CrossRef]
- Elma, E.; Gaulton, R.; Chudley, T.R.; Scott, C.L.; East, H.K.; Westoby, H.; Fitzsimmons, C. Evaluating UAV-based multispectral imagery for mapping an intertidal seagrass environment. Aquat. Conserv. Mar. Freshw. Ecosyst. 2024, 34. [Google Scholar] [CrossRef]
- Duffy, J.P.; Pratt, L.; Anderson, K.; Land, P.E.; Shutler, J.D. Spatial assessment of intertidal seagrass meadows using optical im-aging systems and a lightweight drone. Estuar. Coast. Shelf Sci. 2018, 200, 169–180. [Google Scholar] [CrossRef]
- Poluzzi, A. , Sabelli, B., Taviani, M. Auto-sinecologia del Molluschi dei fondi mobili del Delta settentrionale del Po (Estate 1980). Boll. Soc. Paloeont. It. 1981, 20, 169–178. [Google Scholar]
- Colombo, G., Ferrari, I., Rossi, R., Ceccherelli, V.U., Cavallini, G. Risorse biologiche di una sacca del delta del Po. Conv. Sc. Naz. Ocean. Fondi Mar. 1979, 199-214.
- Viaroli, P., Bartoli, M., Giordani, G., Naldi, M., Orfanidis, S., Zaldivar, J. Community shifts, alternative stable states, biogeochemical controls and feedbacks in eutrophic coastal lagoons: a brief overview. Aquatic Conserv: Mar. Freshw. Ecosyst. 2008, 18, S105–S117. [CrossRef]
- Marini, M.; Grilli, F. The Role of Nitrogen and Phosphorus in Eutrophication of the Northern Adriatic Sea: History and Future Scenarios. Appl. Sci. 2023, 13, 9267. [Google Scholar] [CrossRef]
- Sfriso, A.A., Sciuto, K., Mistri, M., Munari, C., Juhmani, A.-S., Buosi, A., Tomio, Y., Sfriso, A. Where, when, how and what seagrass to transplant for long lasting results in transitional water systems: the cases of Cymodocea nodosa, Zostera marina, Zostera noltei and Ruppia cirrhosa. Front. Mar. Sci. 2023, 1299428. [CrossRef]
- Sfriso, A., Buosi, A, Sciuto, K., Wolf, M., Tomio, Y., Juhmani A.-S., Sfriso,A.A. Effect of Ecological Recovery on Macrophyte Dominance and Production in the Venice Lagoon. Front. Mar. Sci. 2022, 9. [CrossRef]
- Sfriso, A., Bonometto, A., Boscolo, R., Bruno, L., Buosi, A., Facca, C., et al. Trapianto delle piante acquatiche per il ripristino dell’habitat «lagune costiere»—Linee guida dall’esperienza del progetto LIFE Natura SERESTO. 2017. Available at: http://www.lifeseresto.eu.
- Sfriso, A., Buosi, A., Tomio, Y., Juhmani, A.-S., Chiesa, S., Greco, M., et al. Sediment carbon variations in the Venice lagoon and other transitional water systems of the Northern Adriatic Sea. Water 2020, 12, 3430. [CrossRef]
- Sfriso, A., Buosi, A., Mistri, M., Munari, C., Franzoi, P., Sfriso, A.A. (2019b). Long-term changes of the trophic status in transitional ecosystems of the northern Adriatic Sea, key parameters and future expectations: The lagoon of Venice as a study case. Nat. Conserv. 2019, 34, 193–215. [CrossRef]
- Strickland, J.D.H., Parsons, T.R. A Practical Handbook of Seawater Analyses (2nd Ed.) 1984. Ottawa, Canada: Bulletin of Fishery Research Board of Canada.
- Clarke, K.R., Gorley, R.N., 2006. PRIMER V6: User Manual/Tutorial. PRIMER-E, Plymouth, UK.
- Marchini, A., Munari, C., Mistri, M. Functions and ecological status of eight Italian lagoons examined using biological traits analysis (BTA). Mar. Pollut. Bull. 2008, 56, 1076–1085.
- Borja, A., Franco J., Perez V. A marine biotic index to estabilish the ecological quality of soft bottom benthos within European estuarine and coastal environments. Mar. Poll. Bull. 2000, 40, 1100–1114. [CrossRef]
- Muxika, I., Borja, A., Bald, J. Using historical data, expert judgement and multivariate analysis in assessing reference conditions and benthic ecological status, according to the European Water Framework Directive. Mar. Pollut. Bull 2007, 55, 16–29. [CrossRef] [PubMed]
- ISPRA. Manuale per la classificazione dell’Elemento di Qualità Biologica “Fauna Ittica” nelle lagune costiere italiane. 2017. Manuali e Linee Guida 168, 267 pp.
- Holling, C.S. Resilience and stability of ecological systems. Ann. Rev.. Ecol. Syst. 1973, 4, 1–23. [Google Scholar] [CrossRef]
- Crespo D, Faião R, Freitas V, Oliveira VH, Sousa AI, Coelho JP, Dolbeth M. Using seagrass as a nature-based solution: Short-term effects of Zostera noltei transplant in benthic communities of a European Atlantic coastal lagoon. Mar Pollut Bull 2023, 197, 115762. [CrossRef]
- Fonseca, M., Bell, S. Influence of physical setting on seagrass landscapes near Beaufort, North Carolina, USA. Mar. Ecol. Prog. Ser. 1998; 171, 109–121.
- Grall, J., Glémarec, M. Using biotic indices to estimate macrobenthic community perturbations in the Bay of Brest. Estuar. Coast. Shelf Sci. 1997, 44, 43–53. [CrossRef]
- Leung, J.Y.S. Habitat heterogeneity affects ecological functions of macrobenthic communities in a mangrove: implication for the impact of restoration and afforestation. Glob. Ecol. Conserv. 2015, 4, 423–433. [Google Scholar] [CrossRef]
- Southwood, T. Habitat, the templet for ecological strategies? J. Anim. Ecol. 1977, 46, 337–365. [Google Scholar] [CrossRef]
- Townsend, C.R., Hildrew, A.G. Species traits in relation to a habitat templet for river systems. Freshw. Biol. 1994, 31, 265–275. [CrossRef]
- Franco, A.; Franzoi, P.; Malavasi, S.; Riccato, F.; Torricelli, P.; Mainardi, D. Use of shallow water habitats by fish assemblages in a Mediterranean coastal lagoon. Estuar. Coast. Shelf Sci. 2006, 66, 67–83. [Google Scholar] [CrossRef]
- Elliott, M., Dewailly, F. The structure and components of European estuarine fish assemblages. Neth. J. Aquat. Ecol, 1995; 29, 397–417.
- Mathieson, S., Cattrijsse, A., Costa, M.J., Drake, P., Elliott, M., Gardner, J., Marchand, J. Fish assemblages of European tidal marshes: a comparison based on species, families and functional guilds. Mar. Ecol., Prog. Ser, 2000; 204, 225–242.
- Barbedo, J.G.A. A review on the use of unmanned aerial vehicles and imaging sensors for monitoring and assessing plant stresses. Drones 2019, 3(2), 40. [Google Scholar] [CrossRef]
- Cassel, M., Piégay, H., Fantino, G., Lejot, J., Bultingaire, L., Michel, K., & Perret, F. Comparison of ground-based and UAV a-UHF artificial tracer mobility moni-toring methods on a braided river. Earth Surf. Proc. Land. 2020; 45, 5, 1123–1140.
- Casagli, N., Frodella, W., Morelli, S., Tofani, V., Ciampalini, A., Intrieri, E., Lu, P. Spaceborne, UAV and ground-based remote sensing techniques for land-slide mapping, monitoring and early warning. Geoenv. Disasters 2017, 4, 1–23.
- Zeybek, M., Taşkaya, S., Elkhrachy, I., Tarolli, P. Improving the Spatial Accuracy of UAV Platforms Using Direct Georeferencing Methods: An Application for Steep Slopes. Remote Sens 2003, 15, 2700.
- Kim, H., Hyun, C.-U., Park, H.-D., Cha, J. Image Mapping Accuracy Evaluation Using UAV with Standalone, Differential (RTK), and PPP GNSS Positioning Techniques in an Abandoned Mine Site. Sensors, 2003; 2, 5858.
- Agrawal, J., Arafat, M.Y. Transforming Farming: A Review of AI-Powered UAV Technologies in Precision Agriculture. Drones, 2024; 8, 664.
- Zhang, Z., Zhu, L. A Review on Unmanned Aerial Vehicle Remote Sensing: Platforms, Sensors, Data Processing Methods, and Applications. Drones, 2023; 7, 398.
- Mistri, M., Fano E.A., Rossi R. Redundancy of macrobenthos from lagoonal habitats in the Adriatic Sea. Mar. Ecol. Prog. Ser 2001, 215, 289–296. [CrossRef]
- Barbier, E.B., Hacker, S.D., Kennedy, C., Koch, E.W., Stier, A.C., Silliman, B.R. The value of estuarine and coastal ecosystem services. Ecol. Monogr. 2011, 8, 169–193.
- Borja, A., Dauer, D.M., Elliott, M., Simenstad, C.A. Medium-and long-term recovery of estuarine and coastal ecosystems: patterns, rates and restoration effectiveness. Estuar. Coasts 2010, 33, 1249–1260. [CrossRef]
Figure 1.
Intervention area (map from https://www.d-maps.com/conditions.php?lang=it).
Figure 1.
Intervention area (map from https://www.d-maps.com/conditions.php?lang=it).
Figure 2.
a) Transplant operation in the Caleri lagoon; b) transplant scheme of the sods.
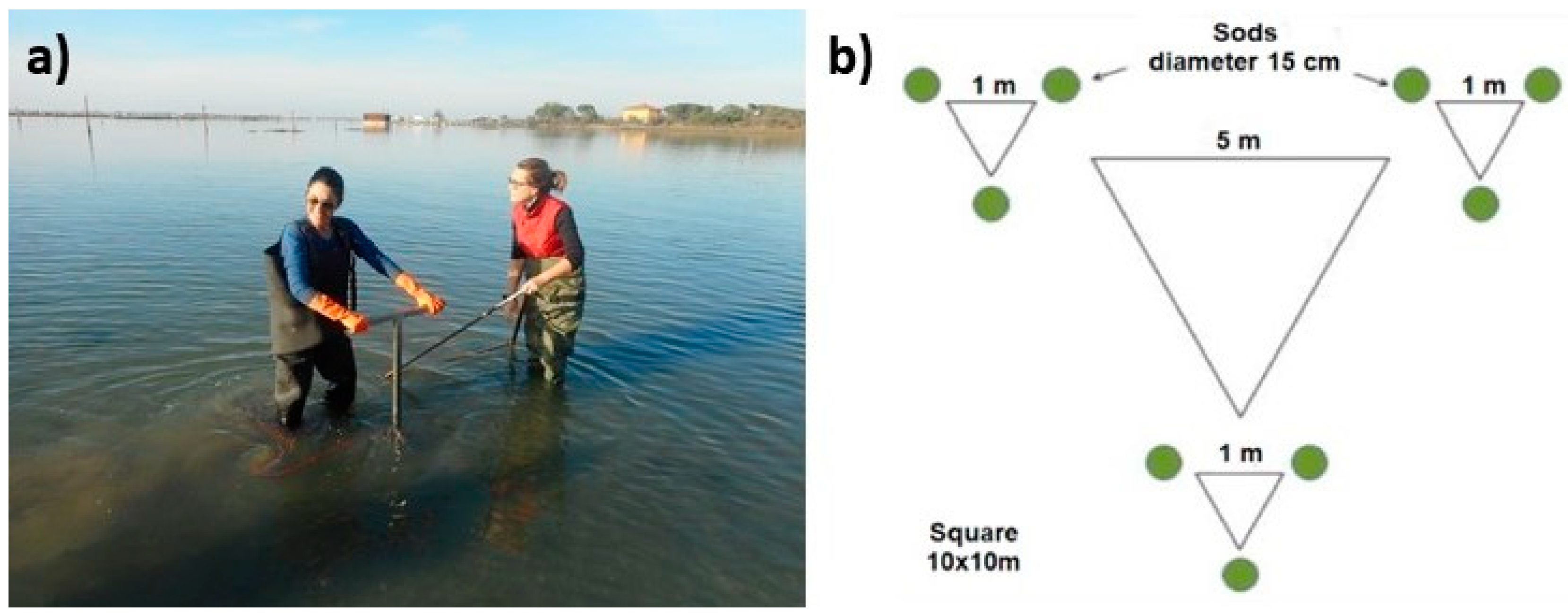
Figure 3.
a) Georeferenced orthomosaic obtained from the preliminary flight; the red line delimits the portion of the transplant area covered by the four detailed flights. b) Aerial image acquired during one of the detailed flights; the field of view is 7 x 10 m.
Figure 3.
a) Georeferenced orthomosaic obtained from the preliminary flight; the red line delimits the portion of the transplant area covered by the four detailed flights. b) Aerial image acquired during one of the detailed flights; the field of view is 7 x 10 m.

Figure 4.
a) Diversity (H’) and b) Evennes (J’) at the transplant (grey bars) and control (white bars) sites.
Figure 4.
a) Diversity (H’) and b) Evennes (J’) at the transplant (grey bars) and control (white bars) sites.
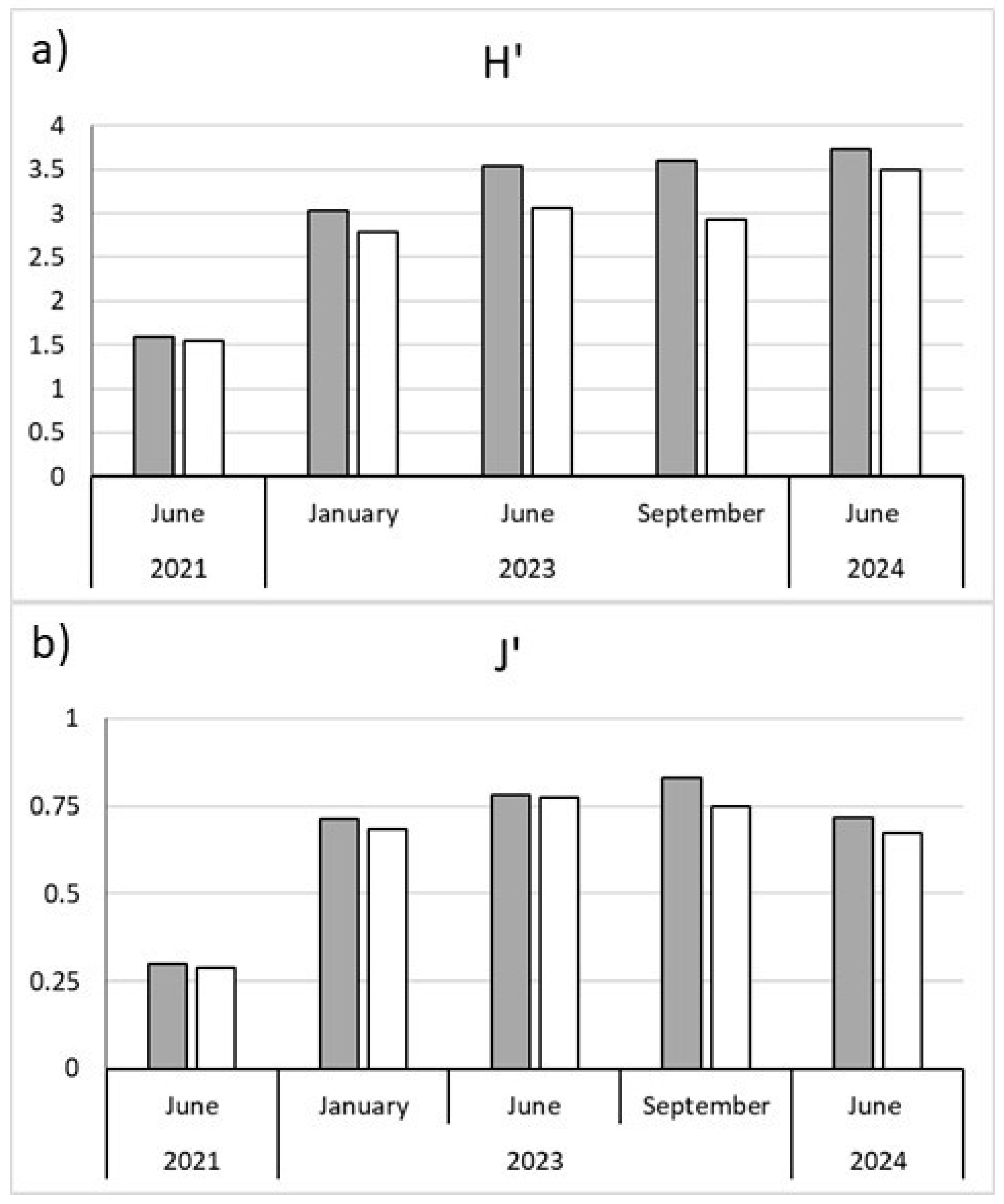
Figure 5.
Ordination by MDS of the untransformed community data for each site. Points are connected in time sequence. Stress value is given on the right corner of the plot.
Figure 5.
Ordination by MDS of the untransformed community data for each site. Points are connected in time sequence. Stress value is given on the right corner of the plot.
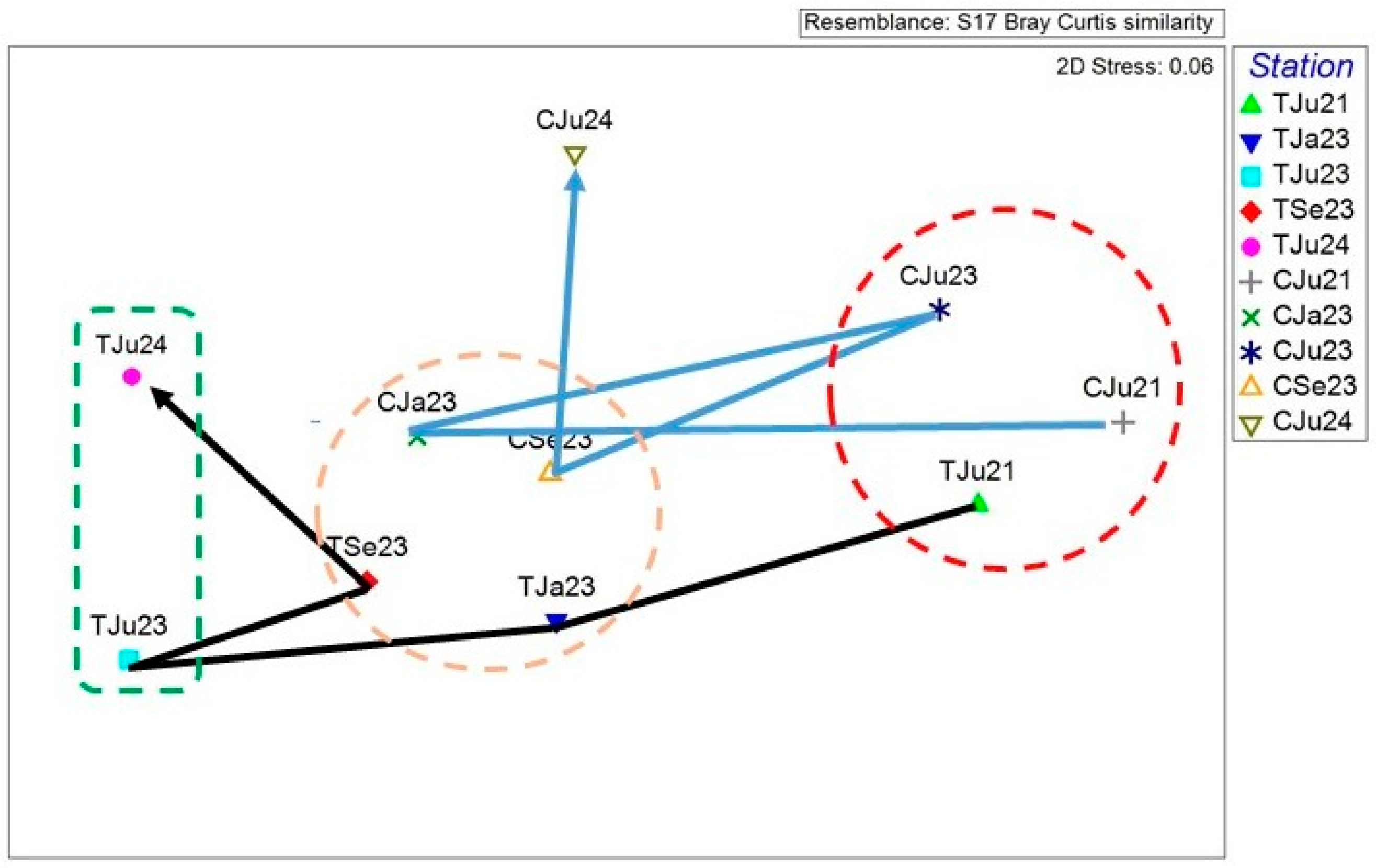
Figure 6.
Percent frequencies of modalities within each trait, weighted for taxa abundance, calculated for each site (T: grey bars; C: white bars) at each sampling date. See Table 1 for labels.
Figure 6.
Percent frequencies of modalities within each trait, weighted for taxa abundance, calculated for each site (T: grey bars; C: white bars) at each sampling date. See Table 1 for labels.
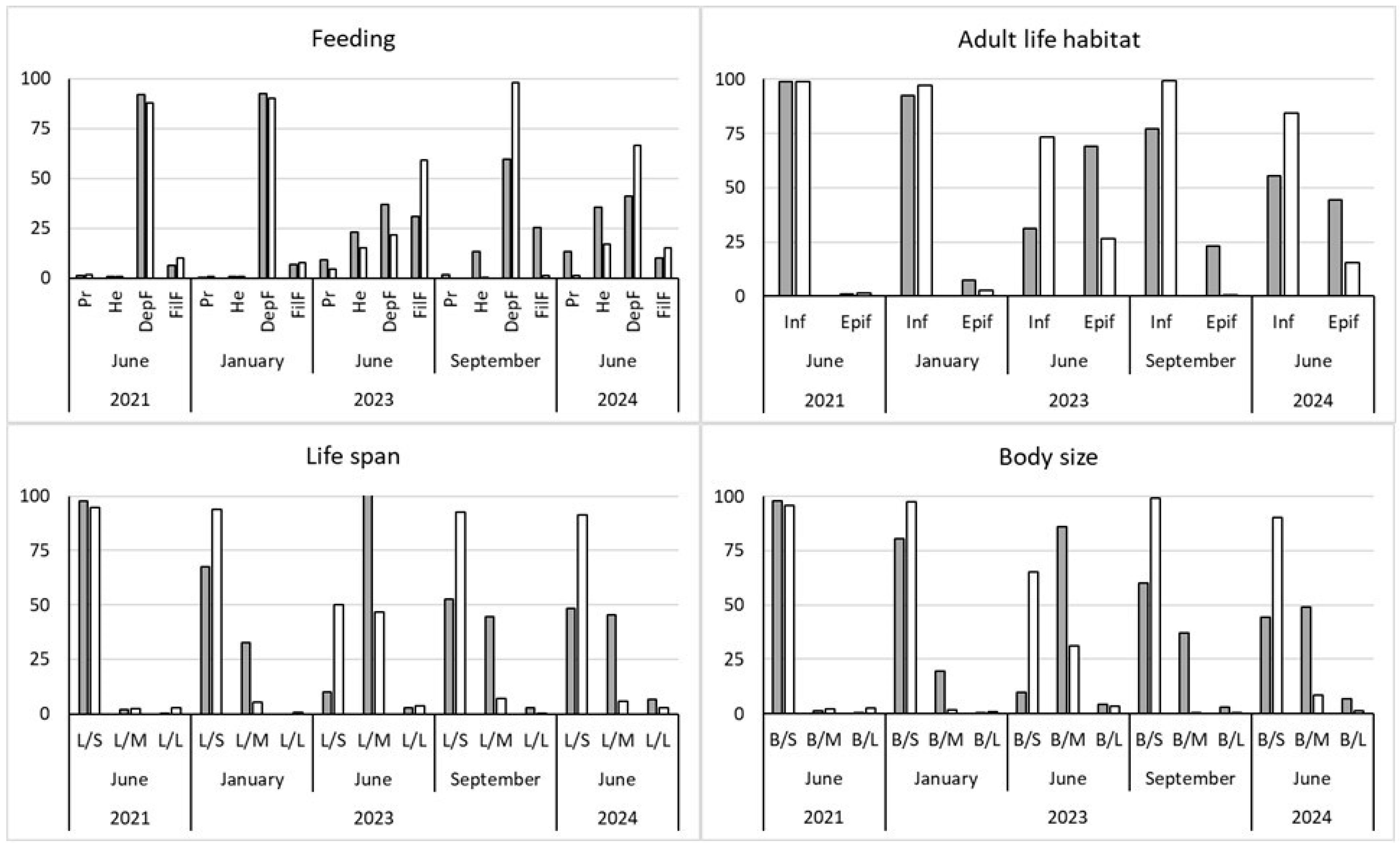
Figure 7.
a) Ecological quality ratio (EQR) at the transplant (T) and control (C) sites through the biotic index M-AMBI. The green line indicates the Moderate/Good ES threshold. b) EQR at T and C sites through the fish index HFBI. The green line indicates the Moderate/Good status threshold.
Figure 7.
a) Ecological quality ratio (EQR) at the transplant (T) and control (C) sites through the biotic index M-AMBI. The green line indicates the Moderate/Good ES threshold. b) EQR at T and C sites through the fish index HFBI. The green line indicates the Moderate/Good status threshold.
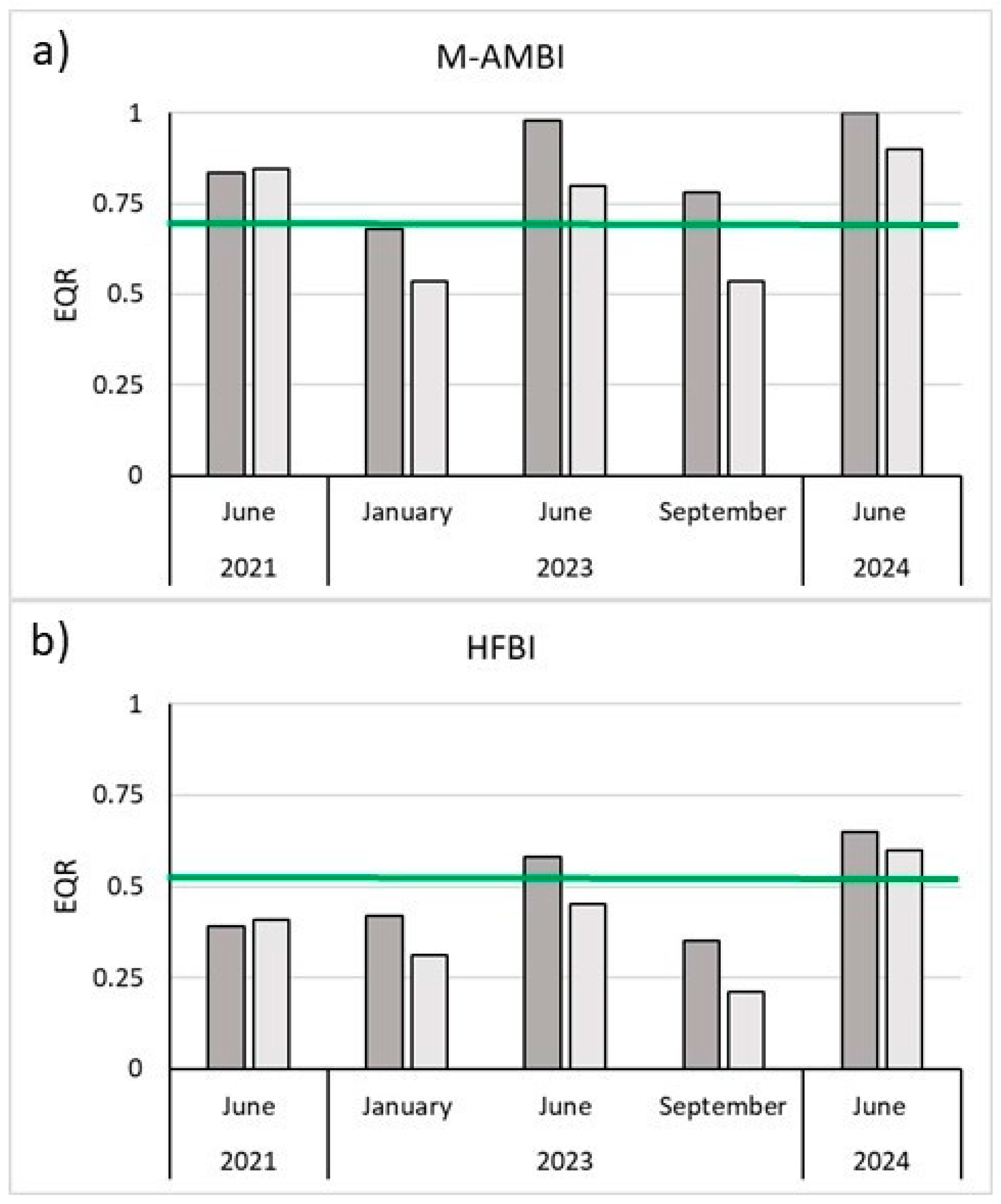
Figure 8.
Geoereferenced orthomosaic obtained from the detailed UAV flights. The red points correspond to the location of the 102 seagrass sods identified in the images.
Figure 8.
Geoereferenced orthomosaic obtained from the detailed UAV flights. The red points correspond to the location of the 102 seagrass sods identified in the images.
Figure 9.
a) b) c) Examples of seagrass sods identified in the images. d) Example of diameter measurements performed on the largest identified seagrass sod.
Figure 9.
a) b) c) Examples of seagrass sods identified in the images. d) Example of diameter measurements performed on the largest identified seagrass sod.

Figure 10.
Frequency histogram of the measured diameters of the seagrass sods identified in the images acquired during the UAV survey.
Figure 10.
Frequency histogram of the measured diameters of the seagrass sods identified in the images acquired during the UAV survey.
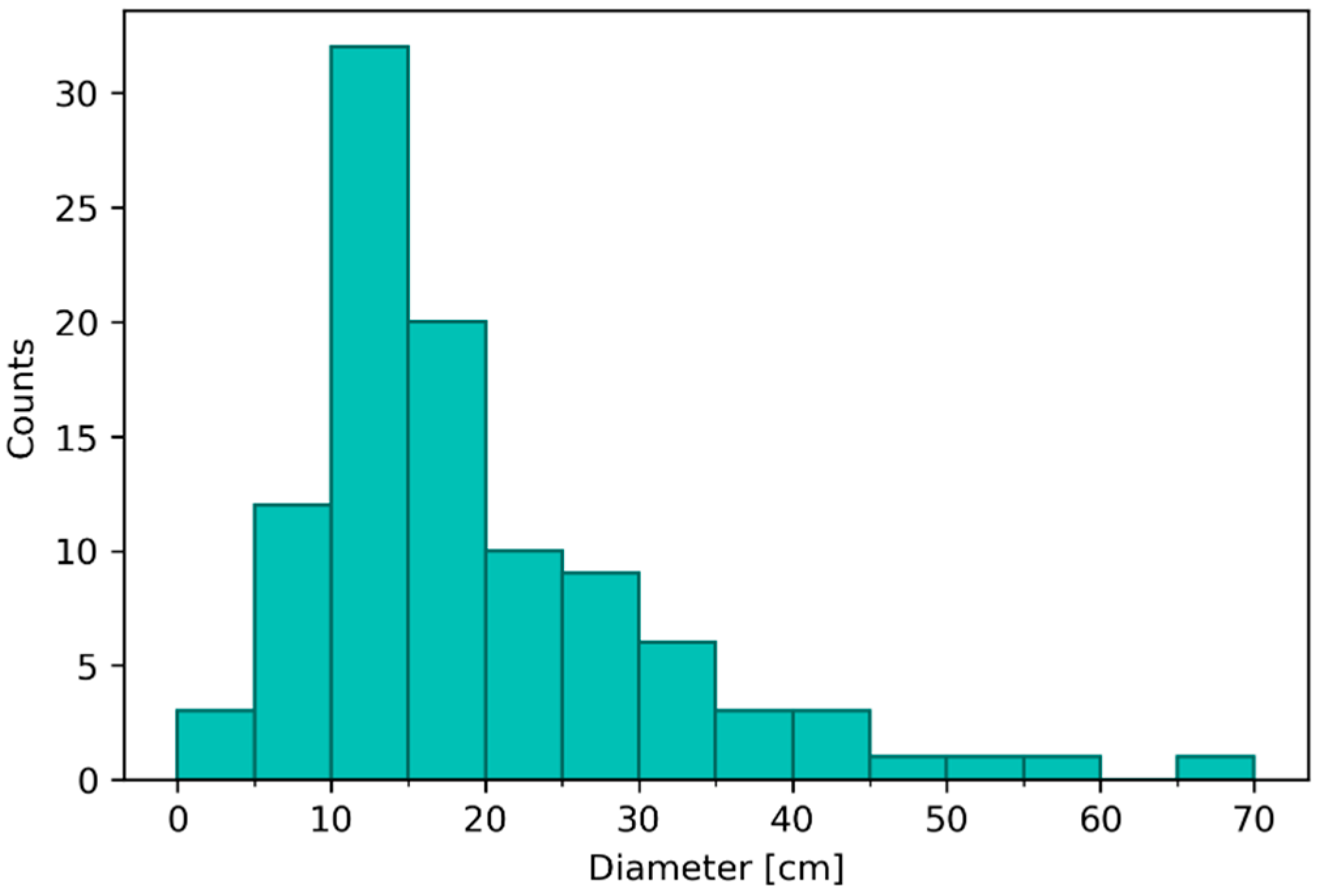
Figure 11.
June 2024: continuous meadow formed by the plagiotropic growth of a triplet of sods (with some thalli of Gracilaria vermiculophylla and Ulvaceae entangled in the leaf layer).
Figure 11.
June 2024: continuous meadow formed by the plagiotropic growth of a triplet of sods (with some thalli of Gracilaria vermiculophylla and Ulvaceae entangled in the leaf layer).

Table 1.
(a) Biological traits and relative modalities and (b) ecological groups of AMBI.
| (a) Biological traits | Traits modalities | Labels |
| Feeding mode | Predator | Pr |
| Herbivorous | He | |
| Deposit-feeder | DepF | |
| Filter-feeder | FilF | |
| Adult life habitat | Infauna | Inf |
| Epifauna | Epif | |
| Life span | Short (<1 year) | L/S |
| Medium (1–5 years) | L/M | |
| Long (>5 years) | L/L | |
| Body size (g) | Small (<0.001 g) | B/S |
| Medium (0.01–0.05 g) | B/M | |
| Large (>0.05 g) | B/L | |
| (b) Ecological groups | Sensitive | EG-I |
| Indifferent | EG-II | |
| Tolerant | EG-III | |
| 2nd order opportunists | EG-IV | |
| 1st order opportunists | EG-V |
Table 2.
Mean (plus standard deviation, SD), minimum and maximum value of water and sediment parameters (January-December 2023) at the transplant site.
Table 2.
Mean (plus standard deviation, SD), minimum and maximum value of water and sediment parameters (January-December 2023) at the transplant site.
| Mean | SD | Min | Max | |||
| Water | Temp | °C | 17.8 | 7.2 | 7.5 | 29 |
| pH | 8.3 | 0.2 | 8.1 | 8.6 | ||
| Eh | mV | 307.1 | 47.5 | 216.0 | 383.0 | |
| Salinity | psu | 18.7 | 2.8 | 15.0 | 23.2 | |
| DO | mg/L | 9.7 | 2.0 | 6.7 | 13.6 | |
| TSS | mg/L | 17.4 | 9.0 | 7.2 | 37.0 | |
| RP | µg/L | 0.4 | 0.1 | 0.2 | 0.6 | |
| DIN | µg/L | 12.5 | 5.7 | 5.4 | 23.9 | |
| RSI | µg/L | 24.0 | 14.5 | 2.6 | 44.2 | |
| Chl-a tot | µg/L | 2.5 | 1.2 | 0.9 | 4.7 | |
| Sediment | pH | 7.6 | 0.1 | 7.4 | 7.8 | |
| Eh | mV | -15.3 | 81.4 | -162.0 | 122.0 | |
| Ptot | µg/g | 616.7 | 82.7 | 507.0 | 734.0 | |
| Ntot | mg/g | 1.4 | 0.3 | 0.9 | 1.8 | |
| Ctot | mg/g | 34.5 | 3.6 | 27.3 | 39.0 | |
| Fines | % | 36.7 | 7.1 | 28.2 | 50.0 | |
| Density | g/cm3 | 0.8 | 0.2 | 0.4 | 1.0 | |
| Moisture | % | 38.0 | 6.4 | 27.1 | 48.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



