
Submitted:
30 December 2024
Posted:
30 December 2024
You are already at the latest version
Abstract
Background: Decathlon is a multimodality sport that requires the combination of endurance, strength, speed, and agility. A polymorphism present in the gene encoding for the protein alpha-actinin-3 (ACTN3) potentially influences sports performance, since αalpha-actinin-3 is a structural component of skeletal muscle that contributes to muscle contraction effectiveness. Aim: To investigate whether the presence of the ACTN3 R577X polymorphism is associated to Decathlon athletes’ performance in the different modalities of Decathlon. Methods: Thirty-one male athletes from the Brazilian national federation of Decathlon aged between 18 and 50 years were genotyped for the ACTN3 R577X polymorphism using polymerase chain reaction (PCR). The athletes' latest Decathlon performances were recorded over ten competitions. The Hardy-Weinberg equilibrium was verified. Pearson's correlation coefficient was utilized to assess the relationship between the obtained sports performance (score) by event and by sets of events (speed events, jumps, and throws) with significance considered at p < 0.05. Results: Strong and significant correlations were identified between the speed events, the jumping, and the launching performances. The frequencies of 51.6%, 48.4%, and 0% for R577R, R577X, and R577R genotypes, respectively, revealed a greater presence of the 577R allele (p < 0.01) among the athletes. However, no significant differences in sports performance (score) were observed between the genotype groups (R577R vs. R577X). Conclusion: Our results support the importance of the ACTN3 genotype, specifically, the presence of the 577R allele, as one of the contributive factors for athletes’ performance in modalities that involve muscle strength, power, and speed.
Keywords:
ACTN3 polymorphism
; decathlon
; α-actin
; sports performance
; muscle strength
1. Introduction
Alpha-actinin 3 is an actin-binding protein located in the Z-discs of skeletal muscles, where it forms a lattice-like structure and stabilizes the actin filaments within sarcomeres. It is expressed only in fast-twitch skeletal muscle fibers (type II), more specifically in 100% of type IIb and about 50% of type IIa muscle fibers [1], however, the absence of alpha-actinin-3does not confer a phenotypic change to the muscle tissue due to a compensatory role of alpha-actinin-2 [2,3].
A common nonsense polymorphism in the gene encoding alpha-actinin-3 (ACTN3) is found in 18 to 25% of the general population. This polymorphism involves a cytosine to a thymine (C→T) nucleotide substitution, resulting in the replacement of an arginine (R) with a stop codon (X) and the subsequent synthesis of a truncated, non-functional form of alpha-actinin [4,5]. This ACTN3 R577X polymorphism has been reported to influence athletes' performance in high-intensity and short-duration sports [6]. In addition, a high frequency of the RX and XX genotypes is present in endurance sports athletes [7]. It has been shown that the ACNT3 577R genotype is associated with better performances in speed, strength, and hypertrophic response adaptations to training, while the 577X genotype is related to better performance in endurance sports [6,8,9].
Decathlon consists of ten athletics events held over two competition days. It requires a mix of different athletic competencies and skills such as speed, agility, and endurance . The Decathlon modalities can be categorized into speed events (100m dash, 110m hurdles, and 400m dash), jumping events (long jump, high jump, and pole vault), throwing events (shot put, discus throw, and javelin throw), and endurance running events (middle-distance - 1500m run).
As previously mentioned, the ACTN3 R577X polymorphism has been extensively studied in the context of its influence on physical performance, with evidence suggesting its role in power, speed, and endurance activities. However, despite the physiological demands of decathlon—a sport requiring a unique combination of strength, speed, endurance, and agility—the relationship between this genetic variant and performance across its ten distinct events has not been previously explored. This study is particularly novel because it examines genotypic distributions and their potential impact on diverse athletic modalities within a single cohort of national-level decathlon athletes. Understanding these relationships could provide critical insights into how genetic factors contribute to multisport performance, guiding talent identification, training optimization, and athlete development strategies. Additionally, the findings may further elucidate the mechanistic role of ACTN3 in determining physical capabilities across different athletic contexts, contributing to the broader field of sports genomics. This study, therefore, addresses a significant knowledge gap in the scholarly literature, offering practical implications for sports science and genetic research. Our aim was to investigate the frequency distribution of the ACTN3 polymorphism in a sample of national Decathlon athletes, exploring whether the genotypes can influence performance in different modalities.
2. Materials and Methods
In this cross-sectional descriptive study design, 31 male Decathlon athletes aged 18 to 50 years were recruited for genotyping according to the following inclusion criteria: (i) having participated in the Brazilian Championship, South Americans, Ibero-Americans, World Championship, or the Olympics games, and (ii) reporting at least 10 years of competing experience. Data from the athlete's scores from 1996 to 2015 were collected from the Troféu Brasil competition in São Paulo, the Brazilian University’s Games (JUBS) in Aracaju City, and the website of the Brazilian Athletics Confederation (CBA). It is important to highlight that the decathlete's scores presented in this study represent 58.06% of the top ranks in the Olympic games.
The sample size was determined a priori using the G*Power software. For this purpose, we considered the following inputs: "T-tests family," point biserial correlation model, with an r2 of 0.35 (associative explanation of at least 35%) for two-tailed correlations. Accordingly, we adopted α of 0.05 and β of 0.95. As such, the minimum sample size indicated for the present study was 30 subjects.
Ethics
This study was designed according to the guidelines of the Declaration of Helsinki and approved by the Ethical Committee for research involving human subjects of the University Center of João Pessoa (UNIPÊ), following the National Health Council - Resolution 466/2012 of December 12, 2012, CAAE number #45665215.5.0000.5176. All participants were informed about the objectives, procedures, and potential risks involved in the study and provided their consent.
Sampling and Genotyping
All participants were asked to refrain from eating for two hours before collecting the buccal epithelium A sterile swab (Eppendorf) was used for this purpose. Immediately before taking the biomaterial, the subjects thoroughly rinsed their mouths with warm water. Samples were separated by centrifugation. The resulting aliquots were placed in a refrigerator and kept until DNA profiling at −18 °C. Genomic DNA was extracted by the Chelex method as described by Walsh et al. [10]. Allelic variants and genotypes of the SNP rs1815739 (C577T) of the ACTN3 gene on chromosome 11q13.2 were determined in all participants using real-time polymerase chain reaction (RT-PCR) (Applied Biosystems). The allelic discrimination was performed using a TaqMan® Pre-Designed SNP Genotyping Assay with fluorescent probes delivered by the validated Assay Id C_590093_1_ (Thermofisher Scientific), according to the manufacturer’s protocol. In addition, 20% of the DNA samples were tested twice with 100% agreement of the results to ensure the accuracy of PCR diagnostics. Primers were designed according to the Applied Biosystems protocol for the Context Sequence [VIC/FAM]: CAAGGCAACACTGCCCGAGGCTGAC[T/C]GAGAGCGAGGTGCCATCATGGGCAT. The following designations of the C (R) and T (X) alleles for the rs1815739 of the ACTN3 gene were used: homozygous fully functional CC genotype (cytosine/cytosine) or RR (arginine/arginine); heterozygous (intermediate) CT genotype (cytosine/thymine) or RX (arginine/stop codon); and homozygous low functional TT genotype (thymine/thymine) or XX (stop codon/stop codon).
Statistics
Descriptive statistics were performed, including measures of central tendency (mean, standard deviation, and variance), along with respective percentages. The normality of distributions was assessed using the Kolmogorov-Smirnov test, and the homogeneity of the variance was examined through the Levene test. Pearson's correlation coefficient was utilized to assess the relationship between the obtained sports performance (score) in each event and sets of events (speed events, jumps, and throws). The Hardy-Weinberg equilibrium was verified for describing and predicting the genotype and allele frequency in a given population. Allele frequencies were estimated by counting, and the allele observance was compared to its expected frequency based on the Chi-square statistical test. Differences in sports performance (in points) between genotypic groups (R577R vs. R577X) were analyzed using the non-parametric Mann-Whitney U test. For all inferential data analyses, a significance level of p < 0.05 was considered. The data were analyzed using the statistical software SPSS® for Windows, version 23.
3. Results
The data from the best scores of the study’s participants, according to the modality’s classification is shown in Table 3.
There was a strong correlation between the athlete's performance in the 100m dash and most of the other events, e.g. 400m dash (r = .759) and long jump (r = .727). The athlete's performance in the middle-distance run (1500m) had only a moderate association with the 100m dash (r = .463) (Table 3). Table 4 presents the associations among the athlete's performances in the Decathlon modalities.
There was a strong correlation between athletes’ performance in the speed events (100m, 400m dash, and 110m hurdles) with the set of jumping events (long jump, high jump, and pole vault) (r = .786) and of throwing events (shot put, discus throw, and javelin throw) (r = .747) (Table 4).
In Table 5, the genotypes and allelic frequencies for the entire sample are presented. Results show an equitable distribution of RR (51.6%) and RX (48.4%) genotypes, with the absence of XX genotype (0%) and a consequent high allelic frequency for the R allele amongst the athletes (0.76). The study’s population adhered to the Hardy-Weinberg equilibrium (p = .758; q = .242; X2 = 3.119, p > 0.05).
Figure 1-A describes the performance points per event according to the genotypic distribution of the ACTN3 R577X polymorphism for the entire athlete's sample. There were no significant differences (p > 0.05) between genotypes regarding the athlete's performance per event. In Figure 1-B, we present the performance points per set of events, according to the genotypic distribution of the ACTN3 R577X polymorphism for the entire sample. There were no significant differences (p > 0.05) in the scores obtained in each set of events between both genotypic groups.
4. Discussion
Our results provide a description of the genotypic profile of national competitors of Decathlon for the ACTN3 R577X polymorphism. Our results show that there is a moderate to strong association between performances in the different events that require speed and agility in Decathlon. The sole exception was noticed for the performances in the half-distance race (1500m), which is an endurance task, that was weakly associated with those of 100m dash.
Despite technical specificities that are peculiar to each event, these associations are expected as it is well established that speed and agility are strongly associated with motor skills that rely on muscular strength and power like jumping and throwing [11,12]. This is due to the rate of force development (RFD) required for performance in expressly explosive motor activities [12], and the RFD is partially determined by innate qualities of the neuromuscular system which include the biochemical and contractile properties of the muscle fibers [13].
Considering the scores by sets of events, the athletes achieved the highest scores in the speed events (100m, 400m, and 110m hurdles) (total of 2,418 points; average of 806.0 points), followed by the jumping events (long jump, high jump and pole vault) (total of 2,188 points; average of 729.0 points), and launchings (shot put, discus and javelin throw) (total of 1,932 points; average of 644 points). The average score in the medium-distance run (633.0 points) was the lowest of all events, indicating that the sample of athletes was primarily composed of sprinters.
Analysis of the distribution of the ACTN3 R577X polymorphism revealed that 51.6% (n: 16) of athletes were homozygotes for the arginine base (R577R genotype) and that 48.4% (n: 15) of athletes presented the stop codon in one of the alleles (R577X genotype). A significantly higher frequency of the 577R allele compared to the 577X allele in athletes specializing in activities requiring explosive muscle contractions and power suggests that individuals expressing the non-polymorphic ACTN3 isoform possess a morphophysiological advantage in developing muscle power. Our findings are in agreement with the reports of several studies that identified a higher frequency of the R allele in strength/power activities and of the X allele in resistance activities [8,14,15].
The available body of scholarly evidence indicates that the ACTN3 genotype influences muscle phenotypes and physical performance. Yang et al. [6] reported a lower frequency of the X577X genotype in speed athletes (8%) compared to endurance athletes (24%) and non-athletes (18%). In contrast to our findings, Niemi and Majamaa's [8], studying Finnish endurance and sprint athletes found that the frequency of the ACTN3 X577X genotype was higher while the frequency of the R577R genotype was lower among endurance athletes. Additionally, they pointed out that none of the top Finnish sprinters had the X577X genotype. Similarly, the study by Moran et al. [16] of 992 adolescent Greeks reported a strong association between the 577R allele and faster times in a 40-meter sprint. In addition, Cieszczyk et al. [17] data reported that sprint swimmers, sprint runners, and weightlifters showed a higher frequency of the 577R allele compared to non-athletes. Nevertheless, this has not been evidenced in all sports. For instance, in the sample of all-time-best Spanish judo male athletes that were compared with ethnically matched nonathletes in Rodríguez-Romo et al. [18] study, no differences were found in allele or genotype distributions, implying that the R577X polymorphism was not associated with the status of being an elite judo athlete in the Spanish population.
The ACTN3 R577X polymorphism appears associated with several aspects of athletes' performance, such as strength, speed, and the proportion of muscle fibers - type II (fast) and type I (slow) [3,19,20]. In the human muscle tissue, there are two sarcomeric (calcium-insensitive) alpha-actinin isoforms: α-actinin-2 and α-actinin-3, encoded by the genes ACTN2 and ACTN3, respectively. As previously mentioned, these a-actinins mainly compose the sarcomeric Z line, where they form a lattice structure that anchors together the actin-containing thin filaments to stabilize muscle contraction and interact with a variety of signaling and metabolic pathways. The expression of alpha-actinin-3 is likely restricted to fast glycolytic skeletal muscle fibers, while the alpha-actinin-2 isoform is predominantly present in cardiac and oxidative skeletal muscle fibers [21].
The experiment by Vincent et al. [22] compared the fiber type composition in the right vastus lateralis of X577X vs. R577R subjects. Their results showed that individuals R577R homozygotes show higher relative dynamic quadriceps torques than X577X homozygotes, and that their fiber type characteristics significantly differed. They found that the percentage surface and number of types IIx fibers were greater in the vastus lateralis of R577R compared to X577X homozygotes and that alpha-actinin-3 protein content was systematically higher in type IIx compared with type IIa fibers (1.17 IIx to IIa ratio). Later on, Norman, et al. [9] investigated the power and fatigability as well as the expression levels of alpha-actinin-2 and alpha-actinin-3 in the vastus lateralis of moderately trained individuals with different ACTN3 genotypes. Their findings showed no difference in the power output, fatigability, or force-velocity across individuals with different ACTN3 genotypes. However, training was able to promptly increase the peak torque in R577R but not with X577X homozygote individuals. Their data revealed that the expression of alpha-actinin-2 is affected by the muscle content of alpha-actinin-3 in a manner that alpha-actinin-2 might compensate for the lack of alpha-actinin-3 and potentially counteract phenotypic consequences. Together, these findings indicate that the ACTN3 577X polymorphism influences the mechanistic role of alpha-actinin-3 in generating muscle contraction and, consequently, the capability to develop strength and speed.
On another perspective, by comparing muscle damage (creatine kinase – CK, alpha-actin, and interleukin 6 (IL-6) in professional soccer players with different ACTN3 genotypes, Pimenta et al. [24] reported that players who were X577X homozygotes presented a higher level of exercise-induced CK and alpha-actin compared to R577R and R577X athletes. Meanwhile, R577R and R577X athletes had higher levels of testosterone and IL-6 compared to athletes X577X. Their results suggest that X577X homozygous athletes are more susceptible to eccentric muscle damage.
It is important to notice that sports performance is not solely supported by pure manifestations of strength and power. Also, homozygosity for the 577X allele might result in deficiency in the alpha-actinin-3 function but does not necessarily result in an abnormal muscular phenotype [25]. Alternatively, the 577X has been proposed as a metabolically thrifty allele [17]. All these findings may justify the absence of differences in performances between the genotypic groups R577R vs. R577X that were exhibited by the Decathlon athletes involved in the present study.
Our study is the first study presenting data on the relationship between the ACTN3 R577X polymorphism genotypic and allelic frequency and sporting performance in Decathlon. Despite the originality and relevance of our findings, the study has some limitations. Firstly, it does not include the genotype study of a non-athlete control group. Secondly, the broad age range of participants in our study necessitated the retrospective use of their best recorded competitive performances over their careers. Consequently, environmental factors such as location, timing within the season, and the age at which these scores were achieved were neither controlled nor adjusted for. Nevertheless, our study contains important merits such as access to high-level multisport athlete’s data.
Despite the limitations mentioned above, the findings of the present study provide valuable insights for coaches, sports scientists, and athletic trainers working with decathlon athletes. By identifying the presence of the ACTN3 R577X genotype, it may be possible to tailor training programs to better suit the genetic predispositions of athletes. For instance, athletes with the R577R genotype, associated with greater muscle strength and power, could benefit from a focus on explosive power and speed-oriented training to maximize their potential in events such as sprints and jumps. Conversely, athletes with the RX genotype, who may have a more balanced strength-endurance profile, could have their training adjusted to improve performance in endurance events like the 1500m run while maintaining their strength capabilities. Furthermore, the results could aid talent identification programs by providing genetic information that complements traditional performance testing, helping identify athletes with the potential for success in multisport disciplines. Lastly, these insights may also contribute to injury prevention strategies by understanding the role of genotype in muscle adaptation and recovery, optimizing long-term athlete health and performance.
5. Conclusions
Our findings revealed a strong association between speed, jumping, and throwing performances in decathlon athletes, reflecting the multifaceted physical demands of this sport. Decathletes demonstrated a superior specialization in speed tasks, which heavily rely on muscular strength and power, further emphasizing the physiological requirements for success in these events. The genotypic distribution in our sample showed a frequency of 51.6% for R577R homozygous and 48.4% for R577X heterozygous athletes, with the absence of the XX genotype, aligning with previous research suggesting a predisposition of the R allele in power and strength-oriented sports. Although no statistically significant differences were observed between genotypes in performance across decathlon events, our results support the contributive role of the 577R allele in the development of muscle strength and power, key attributes for high-level performance in speed and jumping events. The lack of performance disparity between genotypes highlights the multifactorial nature of athletic success in decathlon, where factors such as training, technical proficiency, and environmental influences likely interact with genetic predispositions. This study is the first to investigate the influence of the ACTN3 R577X polymorphism on performance in a cohort of national-level decathlon athletes, filling a critical gap in sports genomics research. These findings underscore the complexity of genetic contributions to multisport performance and provide a foundation for future research exploring gene-environment interactions in decathlon and other multisport disciplines. Practical applications of this work include personalized training strategies and talent identification programs that integrate genetic insights with traditional performance metrics, ultimately advancing athlete development and sports science.
Funding
This research received no external funding.
Institutional Review Board Statement
This study was approved by the Ethical Committee for research involving human subjects of the University Center of João Pessoa (UNIPÊ), Brazil, following the National Health Council - Resolution 466/2012 of December 12, 2012, CAAE number #45665215.5.0000.5176.
Informed Consent Statement
Written informed consent has been obtained from the participants for the publication of this paper.
Data Availability Statement
The data underlying this article cannot be shared publicly due to privacy or ethical restrictions. The data will be shared at reasonable request to the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Dias, R. G., Pereira, A. D. C., Negrão, C. E. & Krieger, J. E. Genetic polymorphisms determining of the physical performance in elite athletes. Rev. Bras. Med. do Esporte 2007. 13, 209–216.
- Blanchard, A., Ohanian, V. & Critchley, D. The structure and function of α-actinin. J. Muscle Res. Cell Motil 1989. 10, 280–289. [CrossRef]
- Pasqua, L. A., Artioli, G. G., Pires, F. de O. & Bertuzzi, R. ACTN 3 e desempenho esportivo: Um gene candidato ao sucesso em provas de curta e longa duração. Rev. Bras. Cineantropometria e Desempenho Hum 2011. 13, 477–483.
- Dias, R. G. Genetics, human physical performance and gene doping: The common sense versus the scientific reality. Rev. Bras. Med. do Esporte 2011. 17, 62–70.
- Mills, Michelle A., Nan Yang, Ron Weinberger, Douglas L. Vander Woude, Alan H. Beggs, Simon Easteal, and Kathryn N. North. Differential expression for the actin-binding proteins, α-actinin-2 and -3, in different species: Implications for the evolution of functional redundancy. Hum. Mol. Genet. 2001. 10, 1335–1346. [CrossRef]
- Yang, Nan, Daniel G. MacArthur, Jason P. Gulbin, Allan G. Hahn, Alan H. Beggs, Simon Easteal, and Kathryn North. ACTN3 genotype is associated with human elite athletic performance. Am. J. Hum. Genet 2003, 73, 627–631. [CrossRef]
- Chae, J. H., Eom, S.-H., Lee, S.-K., Jung, J.-H. & Kim, C.-H. Association between Complex ACTN3 and ACE Gene Polymorphisms and Elite Endurance Sports in Koreans: A Case–Control Study. Genes (Basel). 2024, 15, 1110. [CrossRef]
- Niemi, A. K. & Majamaa, K. Mitochondrial DNA and ACTN3 genotypes in Finnish elite endurance and sprint athletes. Eur. J. Hum. Genet 2005, 13, 965–969. [CrossRef]
- Norman, Barbara, Mona Esbjörnsson, Håkan Rundqvist, Ted Österlund, Ferdinand Von Walden, and Per A. Tesch. Strength, power, fiber types, and mRNA expression in trained men and women with different ACTN3 R577X genotypes. J. Appl. Physiol. 2009, 106, 959–965.
- Walsh, P. S., Metzger, D. A. & Higuchi, R. Chelex® 100 as a medium for simple extraction of DNA for PCR-based typing from forensic material. Biotechniques 1991, 10, 506–513.
- Habibi, Abdolhamid, Mehrzad Shabani, Esmaeil Rahimi, Rouhollah Fatemi, Abdolrahman Najafi, Hossein Analoei, and Morad Hosseini. Relationship between jump test results and acceleration phase of sprint performance in national and regional 100m sprinters. J. Hum. Kinet 2010, 23, 29–35. [CrossRef]
- Marques, M. C. & González-Badillo, J. J. Relationship between strength parameters and squat jump performance in trained athletes. Motricidade 2011, 7, 43–48.
- Vescovi, J. D. & McGuigan, M. R. Relationships between sprinting, agility, and jump ability in female athletes. J. Sports Sci. 2008, 26, 97–107. [CrossRef]
- Druzhevskaya, A. M., Ahmetov, I. I., Astratenkova, I. V. & Rogozkin, V. A. Association of the ACTN3 R577X polymorphism with power athlete status in Russians. Eur. J. Appl. Physiol. 2008, 103, 631–634. [CrossRef]
- Papadimitriou, D. T., Marakaki, C., Fretzayas, A., Nicolaidou, P. & Papadimitriou, A. Negativation of type 1 diabetes-associated autoantibodies to glutamic acid decarboxylase and insulin in children treated with oral calcitriol. J. Diabetes 2013, 5, 344–348.
- Moran, Colin N., Nan Yang, Mark E. S. Bailey, Athanasios Tsiokanos, Athanasios Jamurtas, Daniel G. MacArthur, Kathryn North, Yannis P. Pitsiladis, and Richard H. Wilson. Association analysis of the ACTN3 R577X polymorphism and complex quantitative body composition and performance phenotypes in adolescent Greeks. Eur. J. Hum. Genet. 2007, 15, 88–93. [CrossRef]
- Cieszczyk, Paweł, Jerzy Eider, Magdalena Ostanek, Aleksandra Arczewska, Agata Leońska-Duniec, Stanisław Sawczyn, Krzysztof Ficek, and Krzysztof Krupecki. Association of the ACTN3 R577X polymorphism in Polish power-orientated athletes. J. Hum. Kinet. 2011, 28, 55–61. [CrossRef]
- Rodríguez-Romo, Gabriel, Thomas Yvert, Alfonso de Diego, Catalina Santiago, Alfonso L. Díaz de Durana, Vicente Carratalá, Nuria Garatachea, and Alejandro Lucia. No Association Between ACTN3 R577X Polymorphism and Elite Judo Athletic Status. Int. J. Sports Physiol. Perform 2013, 8, 579–581. [CrossRef]
- Harrison, A. J., Keane, S. P. & Coglan, J. Force-velocity relationship and stretch-shortening cycle function in sprint and endurance athletes. J. strength Cond. Res. 2004, 18, 473–479. [CrossRef]
- Paulo, By, Jorge Paixão, Victor Manuel, Machado Reis, Paulo Jorge, and Paixão Miguel. 400M Performance. Methods 2004, 39–45.
- MacArthur, D. G. & North, K. N. A gene for speed? The evolution and function of α-actinin-3. BioEssays 2004, 26, 786–795.
- Vincent, Barbara, Katrien De Bock, Monique Ramaekers, Els Van Den Eede, Marc Van Leemputte, Peter Hespel, and Martine A. Thomis. ACTN3 (R577X) genotype is associated with fiber type distribution. Physiol. Genomics 2007. 32, 58–63. [CrossRef]
- Pimenta, Eduardo Mendonça, Daniel Barbosa Coelho, Izinara Rosse Cruz, Rodrigo Figueiredo Morandi, Christiano Eduardo Veneroso, Guilherme De Azambuja Pussieldi, Maria Raquel Santos Carvalho, Emerson Silami-Garcia, and José Antonio De Paz Fernández. 2012. The ACTN3 genotype in soccer players in response to acute eccentric training. Eur. J. Appl. Physiol 2012, 112, 1495–1503. [CrossRef]
- Saunders, C. J., A. V. September, S. L. Xenophontos, M. A. Cariolou, L. C. Anastassiades, T. D. Noakes, and M. Collins. No association of the ACTN3 gene R577X polymorphism with endurance performance in Ironman Triathlons. Ann. Hum. Genet. 2007, 71, 777–781. [CrossRef]
Figure 1.
A: Average athletes’ performance per event according to genotypic distribution. B: Average sports performance obtained in the sets of speed, jumps, and throws according to the genotypic distribution.
Figure 1.
A: Average athletes’ performance per event according to genotypic distribution. B: Average sports performance obtained in the sets of speed, jumps, and throws according to the genotypic distribution.
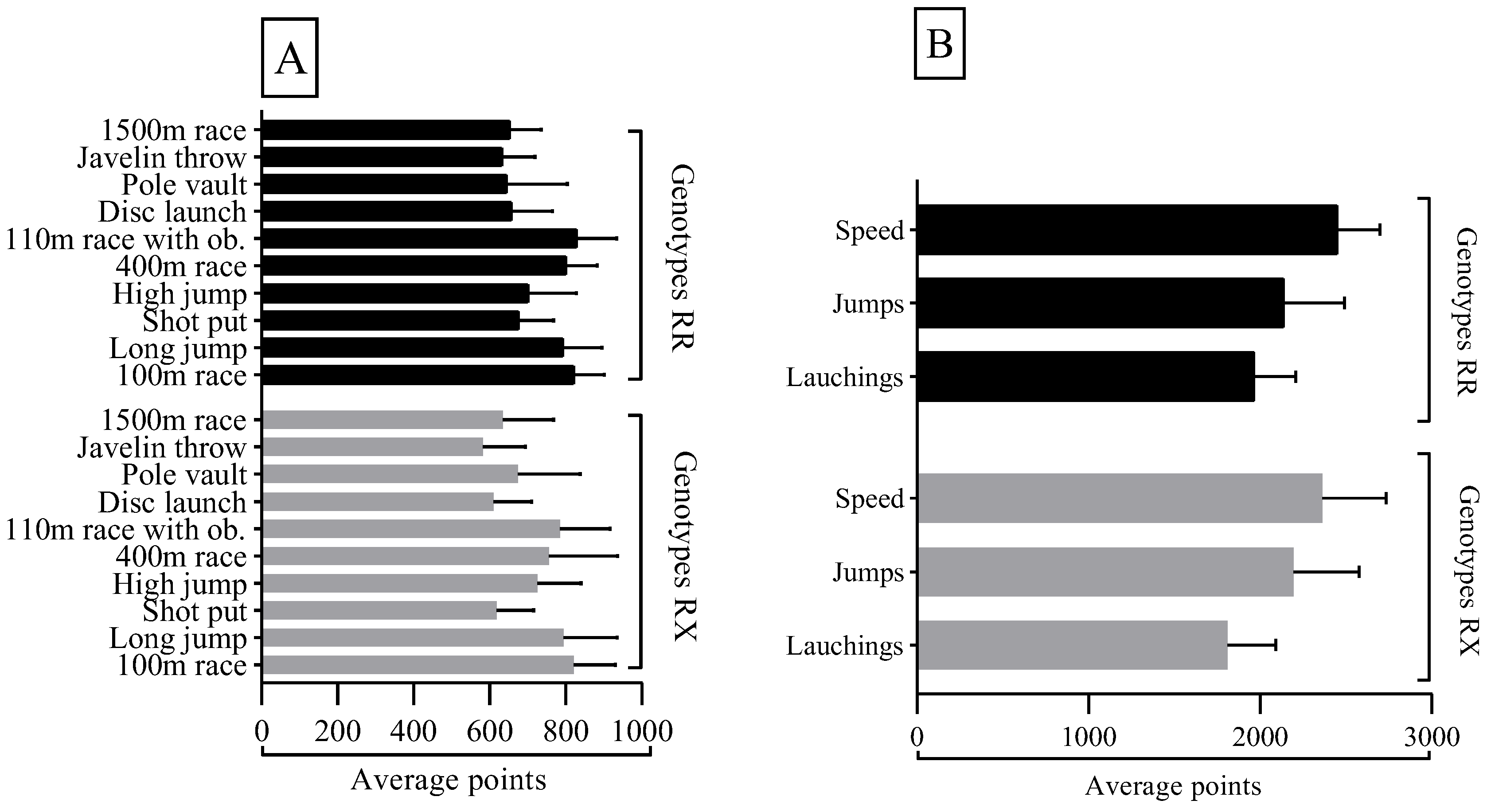

Table 1.
presents the points per result in day-one Decathlon events.
| 100m race | Long jump | Shot put | High jump | 400m race | |||||
| RES - sec | Points | RES - m | Points | RES – m | Points | RES. - | Points | RES. – sec | Points |
| 10.50 | 975 | 7.39 | 908 | 14.02 | 730 | 1.99 | 794 | 49.67 | 830 |
| 10.51 | 973 | 7.38 | 905 | 14.01 | 729 | 1.98 | 785 | 49.69 | 829 |
| 10.52 | 970 | 7.37 | 903 | 13.99 | 728 | 1.97 | 776 | 49.72 | 828 |
| 10.53 | 968 | 7.36 | 900 | 13.97 | 727 | 1.96 | 767 | 49.74 | 827 |
| 10.54 | 966 | 7.35 | 898 | 13.96 | 726 | 1.95 | 758 | 49.76 | 826 |
| 10.55 | 963 | 7.34 | 896 | 13.94 | 725 | 1.94 | 749 | 49.78 | 825 |
| 10.56 | 961 | 7.33 | 893 | 13.93 | 724 | 1.93 | 740 | 49.80 | 824 |
| 10.57 | 959 | 7.32 | 891 | 13.91 | 723 | 1.92 | 731 | 49.82 | 823 |
| 10.58 | 956 | 7.31 | 888 | 13.89 | 722 | 1.91 | 723 | 49.85 | 822 |
| 10.59 | 954 | 7.30 | 886 | 13.88 | 721 | 1.90 | 714 | 49.87 | 821 |
RES. = Resultados: S. = Salto: A. = Arremesso: RES.: Result. - sec: Seconds. – m: Meters.
Table 2.
presents the points per result in day-two Decathlon events.
| 110 w/obstacle | Disc launch | Pole jump | Javelin throw | 1500m race | |||||
| RES - sec | Points | RES - m | Points | RES - m | Points | RES - m | Points | RES - min | Points |
| 14.20 | 949 | 44.16 | 750 | 4.79 | 846 | 60.12 | 740 | 4:39.33 | 685 |
| 14.21 | 948 | 44.11 | 749 | 4.78 | 843 | 60.05 | 739 | 4:39.49 | 684 |
| 14.22 | 946 | 44.06 | 748 | 4.77 | 840 | 59.98 | 738 | 4:39.65 | 683 |
| 14.23 | 945 | 44.02 | 747 | 4.76 | 837 | 59.92 | 737 | 4:39.80 | 682 |
| 14.24 | 944 | 43.97 | 746 | 4.75 | 834 | 59.85 | 736 | 4:39.96 | 681 |
| 14.25 | 942 | 43.92 | 745 | 4.74 | 831 | 59.78 | 735 | 4:40.12 | 680 |
| 14.26 | 941 | 43.87 | 744 | 4.73 | 828 | 59.72 | 734 | 4:40.28 | 679 |
| 14.27 | 940 | 43.82 | 743 | 4.72 | 825 | 59.65 | 733 | 4:40.44 | 678 |
| 14.28 | 939 | 43.77 | 742 | 4.71 | 822 | 59.58 | 732 | 4:40.60 | 677 |
| 14.29 | 937 | 43.72 | 741 | 4.70 | 819 | 59.52 | 731 | 4:40.76 | 676 |
RES.: Result. – m: Meters. – min: Minutes.
Table 3.
The scores (in points) obtained by athletes in the study.
| Set of Events | Sum of points | Average points | Fitness skills |
| Speed | 2.418 | 806 | Explosive strength, displacement speed, agility |
| Jumps | 2.188 | 729 | Travel speed, explosive force |
| Throwing and Launching | 1.932 | 644 | Maximum strength, explosive strength |
| Medium Distance - 1500m | -- | 633 | Aerobic Power |
Table 4.
Pearson Correlation Coefficient among scores in the Decathlon events for the entire sample.
Table 4.
Pearson Correlation Coefficient among scores in the Decathlon events for the entire sample.
| Tests | Race. 100m | L.J | S. put | H.J | C.400m | R.110m w/H | T. of disc | Pole jump | T. of the dart | R. 1500m | |
| 100m race | r | 1 | .727** | .375* | .526** | .759** | .625** | .336 | .428* | .292 | .463** |
| p | 0 | .038 | .002 | 0 | 0 | .064 | .016 | .111 | .009 | ||
| 400m race | r | .759** | .651** | .581** | .583** | 1 | .640** | .489** | .612** | .500** | .285 |
| p | 0 | 0 | .001 | .001 | 0 | .005 | 0 | .004 | .12 | ||
| Long jump | r | .727** | 1 | .558** | .789** | .651** | .829** | .580** | .623** | .385* | .328 |
| p | 0 | .001 | 0 | 0 | 0 | .001 | 0 | .033 | .071 | ||
| Shot put | r | .375* | .558** | 1 | .583** | .581** | .651** | .736** | .699** | .640** | .035 |
| p | .038 | .001 | .001 | .001 | 0 | 0 | 0 | 0 | .853 | ||
| High jump | r | .526** | .789** | .583** | 1 | .583** | .745** | .712** | .790** | .397* | .04 |
| p | .002 | 0 | .001 | .001 | 0 | 0 | 0 | .027 | .832 | ||
| 110m race with ob. | r | .625** | .829** | .651** | .745** | .640** | 1 | .675** | .607** | .433* | .309 |
| p | 0 | 0 | 0 | 0 | 0 | 0 | 0 | .015 | .09 | ||
| Disc launch | r | .336 | .580** | .736** | .712** | .489** | .675** | 1 | .805** | .608** | -.155 |
| p | .064 | .001 | 0 | 0 | .005 | 0 | 0 | 0 | .406 | ||
| Pole jump | r | .428* | .623** | .699** | .790** | .612** | .607** | .805** | 1 | .546** | .037 |
| p | .016 | 0 | 0 | 0 | 0 | 0 | 0 | .001 | .842 | ||
| Javelin throw | r | .292 | .385* | .640** | .397* | .500** | .433* | .608** | .546** | 1 | -.129 |
| p | .111 | .033 | 0 | .027 | .004 | .015 | 0 | .001 | .49 | ||
| 1500m race | r | .463** | .328 | .035 | .04 | .285 | .309 | -.155 | .037 | -.129 | 1 |
| p | .009 | .071 | .853 | .832 | .12 | .09 | .406 | .842 | .49 | ||
r: Correlation coefficient. R.= Race – L.J. = Long Jump - S. put= Shot put – H.J.= High Jump – w/H = with Hurdle - T. = Throw - ** The correlation is significant at the 0.01 level (two-sided). * Correlation is significant at the 0.05 level (two-sided).
Table 4.
Pearson correlation coefficient and its significance between sets of events for the entire sample.
Table 4.
Pearson correlation coefficient and its significance between sets of events for the entire sample.
| Set of events | Variable | Speed | Jumps | Launches |
| Speed | R | 1 | .786 | .634 |
| p-value | <.01 | <.01 | ||
| N | 31 | 31 | 31 | |
| Jumps | R | .786 | 1 | .747 |
| p-value | <.01 | <.01 | ||
| N | 31 | 31 | 31 | |
| Launching and Throws |
R | .634 | .747 | 1 |
| p-value | <.01 | <.01 | ||
| N | 31 | 31 | 31 |
r: Correlation coefficient.
Table 5.
Genotype and allele frequencies observed for the ACTN3 locus for the entire sample are shown in the table below.
Table 5.
Genotype and allele frequencies observed for the ACTN3 locus for the entire sample are shown in the table below.
| Genotypes Frequency | Alleles Frequency | |||
| R577R(%) | R577X(%) | X577X(%) | 577R(%) | 577X(%) |
| 16 (51.6%) |
15 (48.4%) |
0 (0.0%) |
76 | 24 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



